Строение белков
Среди органических веществ белки, или протеины, — самые многочисленные, наиболее разнообразные и имеющие первостепенное значение биополимеры. На их долю приходится 50 — 80% сухой массы клетки.
Молекулы белков имеют большие размеры, поэтому их называют макромолекулами. Кроме углерода, кислорода, водорода и азота, в состав белков могут входить сера, фосфор и железо. Белки отличаются друг от друга числом (от ста до нескольких тысяч), составом и последовательностью мономеров. Мономерами белков являются аминокислоты (рис. 1)
Бесконечное разнообразие белков создается за счет различного сочетания всего 20 аминокислот. Каждая аминокислота имеет свое название, особое строение и свойства. Их общую формулу можно представить в следующем виде:
Молекула аминокислоты состоит из двух одинаковых для всех аминокислот частей, одна из которых является аминогруппой (—NH2) с основными свойствами, другая — карбоксильной группой (—COOH) с кислотными свойствами. Часть молекулы, называемая радикалом (R), у разных аминокислот имеет различное строение. Наличие в одной молекуле аминокислоты основной и кислотной групп обусловливает их высокую реакционную способность. через эти группы происходит соединение аминокислот при образовании белка. При этом возникает молекула воды, а освободившиеся электроны образуют пептидную связь. Поэтому белки называют полипептидами.
Молекулы белков могут иметь различные пространственные конфигурации, и в их строении различают четыре уровня структурной организации.
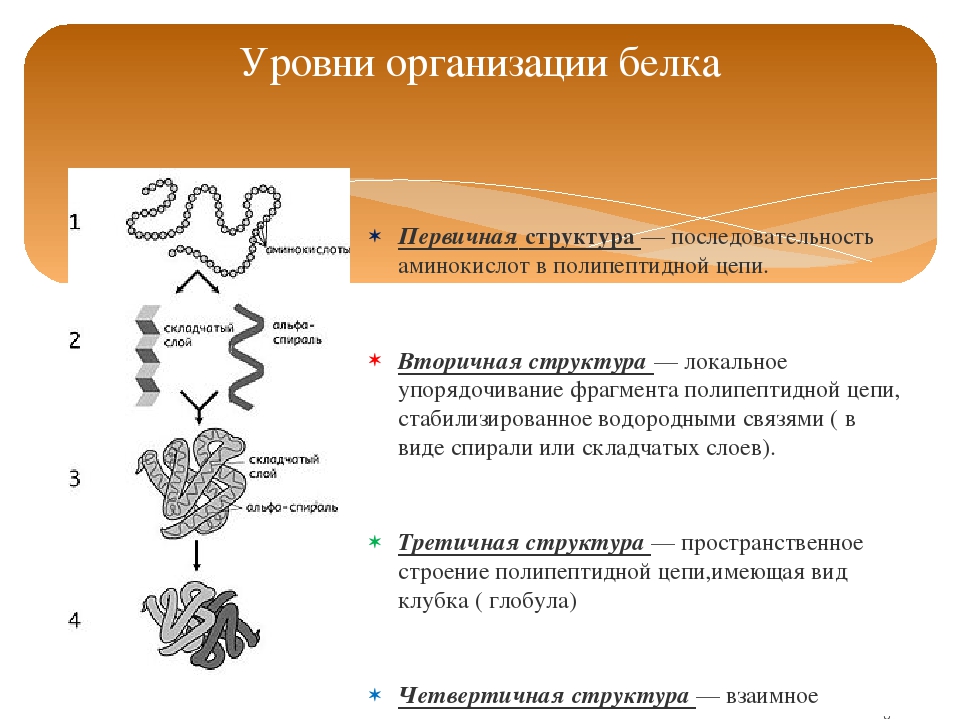
Последовательность аминокислот в составе полипептидной цепи представляет первичную структуру белка. Она уникальна для любого белка и определяет его форму, свойства и функции.
Большинство белков имеют вид спирали в результате образования водородных связей между —CO- и —NH- группами разных аминокислотных остатков полипептидной цепи. Водородные связи малопрочные, но в комплексе они обеспечивают довольно прочную структуру. Эта спираль — вторичная структура белка.
Водородные связи малопрочные, но в комплексе они обеспечивают довольно прочную структуру. Эта спираль — вторичная структура белка.
Третичная структура — трехмерная пространственная «упаковка» полипептидной цепи. В результате возникает причудливая, но для каждого белка специфическая конфигурация — глобула. Прочность третичной структуры обеспечивается разнообразными связями, возникающими между радикалами аминокислот.
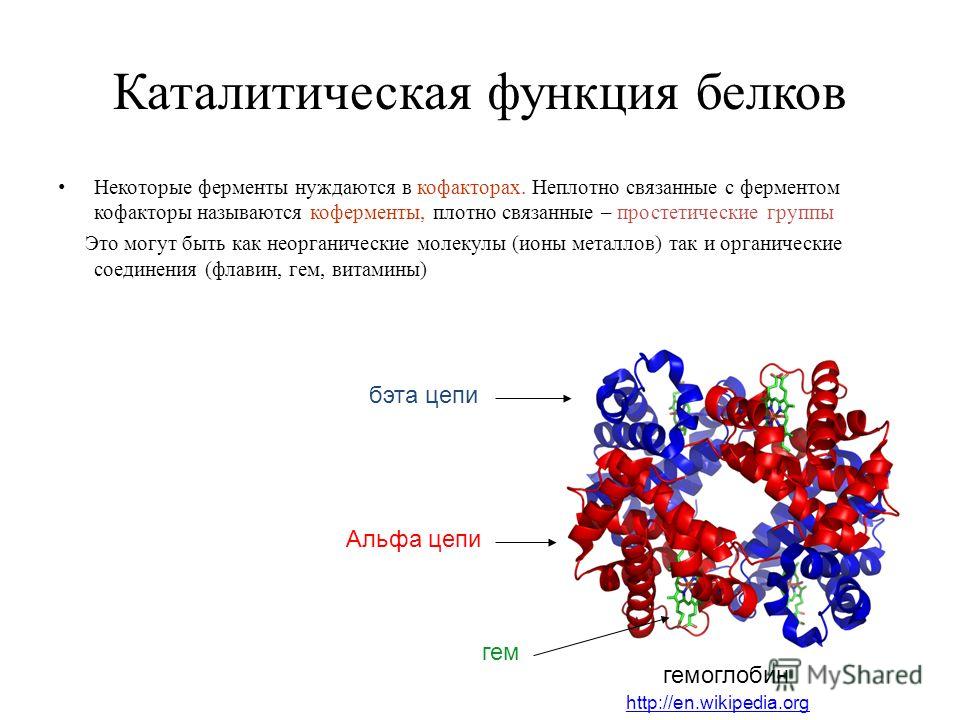
Четвертичная структура характерна не для всех белков. Она возникает в результате соединения нескольких макромолекул с третичной структурой в сложный комплекс. Например, гемоглобин крови человека представляет комплекс из четырех макромолекул белка.
Такая сложность структуры белковых молекул связана с разнообразием функций, свойственных этим биополимерам.
Нарушение природной структуры белка называют денатурацией. Она может происходить под воздействием температуры, химических веществ, лучистой энергии и других факторов. При слабом воздействии распадается только четвертичная структура, при более сильном — третичная, а затем — вторичная, и белок остается в виде полипептидной цепи.
Этот процесс частично обратим: если не нарушена первичная структура, то денатурированный белок способен восстанавливать свою структуру. Отсюда следует, что все особенность строение макромолекулы белка определяются его первичной структурой.
Кроме простых белков, состоящих только из аминокислот, есть еще и сложные белки
Другие заметки по биологии
Строение и функции белков
Доказано, что белки составляют, в среднем, до 50% от сухой массы практически всех живых существ. Основным структурным элементом белка является аминокислота. В составе каждой из аминокислот имеется карбоксильная (кислотная) и аминогруппа. При комбинации этих групп появляется пептидная связь, поэтому второе название белков – полипептиды.
Структуры белка
1. Первичная структура — цепь, состоящая из аминокислот, соединенных пептидной связью (мощной, ковалентной).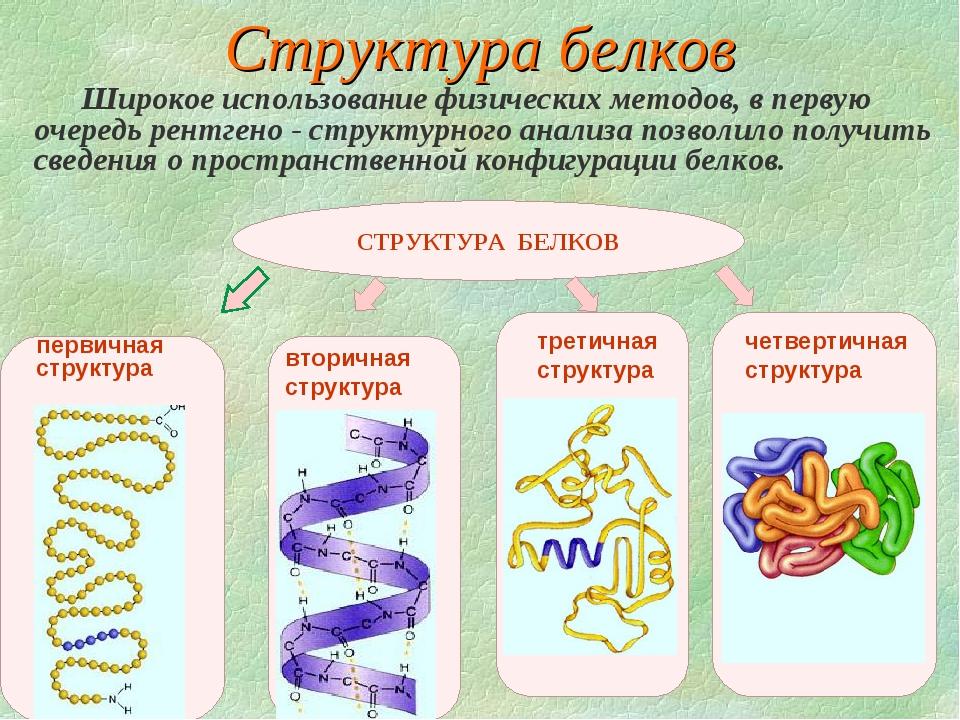 При чередовании 20 аминокислот в разном порядке образуется множество разнообразных белков. В случае замены в определенной белковой цепочке хотя бы одной аминокислоты полностью меняется структура и, соответственно, функции белка. Поэтому первичная структура по праву является основной в белке.
При чередовании 20 аминокислот в разном порядке образуется множество разнообразных белков. В случае замены в определенной белковой цепочке хотя бы одной аминокислоты полностью меняется структура и, соответственно, функции белка. Поэтому первичная структура по праву является основной в белке.
2. Вторичная. Это спираль, форма которой поддерживается водородными связями (относительно слабыми).
3. Третичная структура – это шар, или так называемая глобула. Она образована, благодаря четырем видам связей. Из них наиболее мощная дисульфидная (так называемый серный мостик). Более слабыми являются такие связи, как гидрофобные, ионные, водородные). Функции белка определяются формой глобулы, по которой белки сильно отличаются друг от друга. Вследствие денатурации происходит изменение формы белка, что влечет нарушение его функции.
4. Четвертичная структура характерна не для всех белков. Представлена несколькими глобулами, объединенными друг с другом теми же связями, что и в третичной структуре. Ярким примером является гемоглобин.
Денатурация
Это процесс внешнего влияния на белок, который приводит к изменению формы его глобулы. Воздействие на белок оказывает высокая температура, избыточная соленость, кислотность, реакции с другими веществами. В случае, когда воздействие на белок не сильное (например, повышение температуры на 1 градус), возникает обратимая денатурация. При значительных воздействиях белок подвергается необратимой денатурации. В этом случае повреждаются все структуры, за исключением первичной.
Функции белков
В любом живом организме белки выполняют множество разных функций. Самые важные из них:
1. Структурная (строительная) заключается в том, что любая живая клетка состоит, в большей степени, из белков и воды.
2. Ферментативная выражается в том, что существуют белки-ферменты, выполняющие роль ускорителей химических реакций. Это происходит, так как активная часть фермента подходит к определенному веществу как ключик к замку.
Это происходит, так как активная часть фермента подходит к определенному веществу как ключик к замку.
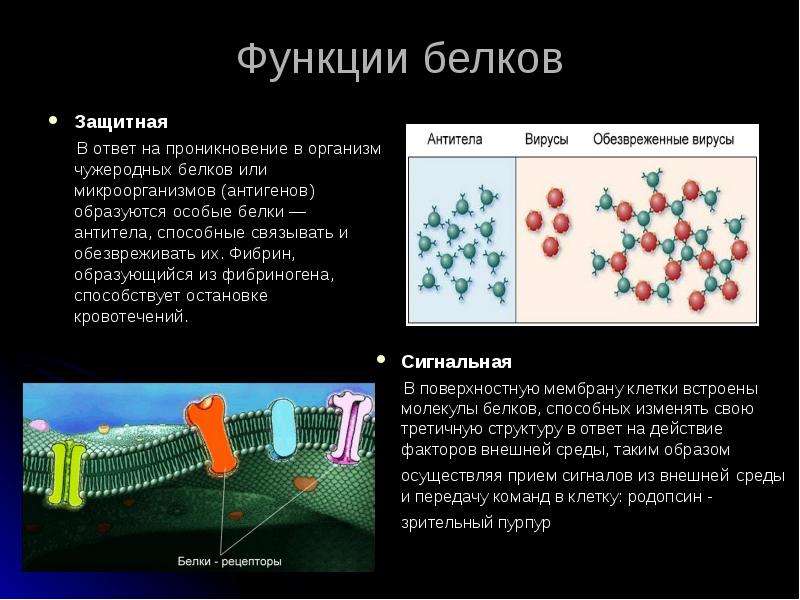
3. Защитная функция ярко прослеживается на примере организма животного, где белки-антитела иммунной системы уничтожают болезнетворные микроорганизмы.
Похожие материалы:
Биология 9 класс. Тема урока: Строение и функции белков. | Уроки по Биологии
Биология 9 класс. Тема урока: Строение и функции белков.
13.09.2014
20641
0
Цели
урока: 1. Продолжить
расширение и углубление знаний о важнейших
органических
веществах клетки на основе изучения
строения
и функции
белков.
2.Знать из чего
состоят белки, функции белков.
3.Продолжить
формирование умения выявлять связи между
строением и
функциями веществ.
Оборудование: карточки -информаторы, учебник 9 кл.,
мет. пособие.
Тип
урока: комбинированный
Методы
обучения: беседа
Ход урока
I. Проверка знаний.
Фронтальная беседа.
- На какие
группы делят все элементы, входящие в состав живого? - На какие
группы делятся углеводы в зависимости от количества молекул? - Почему
углеводы получили такое название? - Что такое
липиды?
II. Изучение
нового материала.
1.
Рассказ об особенностях строение молекул белков.
Белки- нерегулярные
биополимеры, мономерами которых являются аминокислоты. В состав белков входит
20 аминокислот. Все аминокислоты имеют
Все аминокислоты имеют
радикал (R), аминогруппу (Nh3),
карбоксильную группу (COOH ).
H — N — C = O H — N — C — С— N
/ \ + / \\ \
H OH
H -h3O O Н
ПЕПТИДНАЯ
СВЯЗЬ
- Самостоятельное
изучение учащимися текста учебника с. 36 о классификации белков. - Рассказ о
многообразии функций белков с краткой записью в тетрадях.
Строительная — белки входят в
состав клеточных мембран и органелл
клетки. Стенки
кровеносных сосудов, хрящи, сухожилия, волосы, ногти,
когти у высших
животных состоят из белков.
Ферментативная- белки-ферменты
катализируют протекание всех химических
реакций в организме. Они обеспечивают расщепление питательных веществ в
пищеварительном тракте, фиксацию углерода при
фотосинтезе.
Транспортная— белки способны
присоединять и переносить различные вещества. Белок гемоглобин переносит кислород, альбумины
крови переносят жирные кислоты.
Защитная- ее выполняют
иммуноглобулины (антитела) крови, обеспечивающие иммунную защиту
организма. Фибриноген и тромбин
участвуют в свертывании крови и предотвращают
кровотечение.
Сократительная- благодаря
движению относительно друг друга нитей
белков актина и
миозина осуществляется сокращения мышц.
Регуляторная- многие гормоны,
регулирующие транскрипцию, являются белками: инсулин, глюкагон.
Рецепторная- некоторые
белки, встроенные в клеточную мембрану,
способны изменить свою структуру на
действие внешней среды. Фитохром-
светочувствительный белок,
регулирующий фотопериодическую реакцию
растений, оксин- составная часть
родопсина, пигмента, находящегося в клетках
сетчатки глаза.
Пигментную— белки имеющие
цвет, меланин — кожи, волос, радужной
оболочки глаза.
4. Беседа об уровнях организации белковой молекулы.
Первичная структура — последовательность аминокислот в
полипептидной цепи. Первичная структура специфична для каждого белка, замена
одной аминокислоты влечет за собой изменение функции.
Вторичная структура— за
счет водородных связей, 2-спиральная
конфигурация,
структура белка волос, ногтей… .
Третичная- за счет ионных,
водородных , сульфидных связей.
Четвертичная— четырех
отдельных полипептидных
субъединиц и небелковой части —
гема. Гемоглобин .
Денатурация- процесс нарушения структуры белка.
III. Закрепление знаний.
В процессе беседы с помощью следующих вопросов6:
- Какие
органические вещества клетки можно назвать самыми важными? - Каким
образом создается бесконечное разнообразие белков? - Что собой
представляют мономеры биополимеры белка? - Как
формируется пептидная связь? - Что собой
представляет первичная структура белка? - Приведите
примеры белков, выполняющих самые разные функции.
При ответе можно
использовать следующую схему: №1
IV. Итог. На уроке мы повторили и расширили свои знания
по
теме «Белки». Узнали о
функциях белков их строении,
о роли белков как самых
необходимых химических
соединений для
жизнедеятельности всего живого на
Земле.
Урок биологии №6. Функции белков
Тест №1 «Строение и функции белков» — 1 вариант.
Зад. № 1 Ответьте на предложенные вопросы, выберите один правильные ответы.
№ 1 Ответьте на предложенные вопросы, выберите один правильные ответы.
I. Какие соединения являются мономерами молекул белка?
а) глюкоза б) глицерин в) жирные кислоты д) аминокислоты
2. Сколько из известных аминокислот участвуют в синтезе белка?
а) 20 б) 23 в) 100
3. Какая часть молекул аминокислот отличает их друг от друга?
а) радикал б) аминогруппа в) карбоксильная группа
4.Посредством какой химической связи соединены между собой аминокислоты в молекуле белка первичной структуры?
a) пептидная связь
б) водородная связь между группами -СО и -NH, расположенных в соседних завитках спирали
в) связи между радикалами аминокислот
5.О каком уровне структурной организации белка идёт речь :«Это трёхмерная, пространственная «упаковка» полипептидной цепи, в результате которой возникает — глобула»? а) первичная структура б) вторичная структура в) третичная структура
г) четвертичная структура
6.Простые белки состоят из:
а) только из аминокислот в)аминокислот и липидов
б) аминокислот и углеводов г) аминокислот и нуклеиновых кислот
7. О какой функции белка идёт речь: «Специальные белки способны ускорять биохимические реакции, идущие в клетке»?
а) пластическая г) защитная ж) рецепторная б) каталитическая д) сократительная з) энергетическая д) регуляторная
8.О какой функции белка идет речь: «Специальные белки поддерживают постоянную концентрацию веществ в крови и клетках, участвуют в росте, размножении и других жизненно важных процессах»?
а) пластическая б) каталитическая в) сократительная г) защитная д) рецепторная
е) энергетическая ж) регуляторная
9.Процесс потери белком своих свойств и нарушение структуры белковой молекулы – это:
а) ренатурация; б) репликация; в) денатурация; г) спирализация
10.Вторичная структура белка: а) глобула; б) конформация; в) свёрнута в спираль.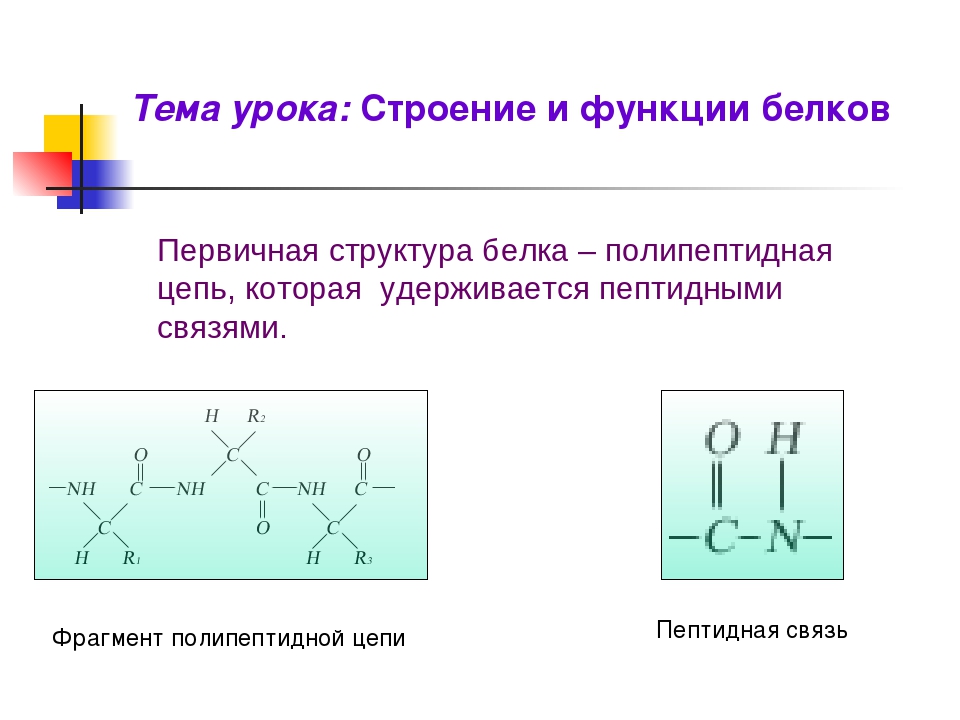
11.При обратимой денатурации нарушаются следующие структуры белка:
а) вторичная; б) первичная; в) третичная; г) четвертичная; д) все, кроме первичной.
Зад. № 2 Ответьте на предложенные вопросы, выберите два правильных ответа.
9. Что является общим для всех аминокислот?
а) радикал б) аминогруппа в) карбоксильная группа
Зад. № 3 Ответьте на предложенные вопросы, выберите три правильных ответа.
10.Какие структуры белка способны нарушаться, а затем восстанавливаться?
а) первичная структура б) вторичная структура в) третичная структура
г) четвертичная структура
Тест №1 «Строение и функции белков» — 2 вариант.
Зад. № 1 Ответьте на предложенные вопросы, выберите один правильные ответы.
1.Какие соединения являются мономерами молекул белков?
а) жирные кислоты б) глицерин в) аминокислоты г) глюкоза
2.Сколько из известных аминокислот участвуют в синтезе белка?
а) 150 6)5 в) 20
3.Какая часть молекул аминокислот отличает их друг
от друга? а) карбоксильная группа б) радикал в) аминогруппа
4.Посредством какой химической связи возникает третичная структура белка?
а) пептидная б) водородная связь между группами -СО и -NH, расположенными в соседних завитках спирали в) связи между радикалами аминокислот
5.О каком уровне структурной организации белка идёт речь: «Она возникает в результате соединения нескольких макромолекул с третичной структурой в сложный комплекс? а) первичная структура б) вторичная структура в) третичная структура
г) четвертичная структура
6.О какой функции белков идёт речь: «Специфические белки предохраняют организмы от вторжения чужеродных белков и организмов и от повреждения»?
а) пластическая б) каталитическая в) защитная г) транспортная д) сократительная ж) рецепторная е) регуляторная з) энергетическая
7. О какой функции белков идёт речь: «Специальные белки способны присоединять и переносить различные вещества».
О какой функции белков идёт речь: «Специальные белки способны присоединять и переносить различные вещества».
а) пластическая б) каталитическая в) транспортная г) защитная ж) рецепторная
д) сократительная з) энергетическая е) регуляторная.
8.Первичная структура молекулы белка удерживается:
а) водородными связями; б) пептидными связями; в) гидрофобными связями; г) дисульфидными связями.
9.Ферменты от других белков отличаются тем, что они:
а) синтезируются на рибосомах; б) включают в свой состав витамины, металлы;
в) являются катализаторами химических реакций
10.Ренатурация – это процесс: а) нарушение естественной структуры белка;
б) восстановления естественной структуры белка
Зад. № 2 Ответьте на предложенные вопросы, выберите два правильных ответа
8.Что является общим для всех аминокислот?
а) карбоксильная группа б) радикал в) аминогруппа
Зад. № 3. Ответьте на предложенные вопросы, выберите три правильных ответа.
9.Какие структуры молекул белка способны нарушаться при денатурации, а затем вновь восстанавливаться? а) четвертичная структура б) третичная структура в) вторичная структура г) первичная структура
10.Сложные белки состоят из: а) только аминокислот б) аминокислот и углеводов в) аминокислот и липидов г) аминокислот и нуклеиновых кислот
Тест по биологии на тему «Молекулярный уровень» (9 класс)
Тест. Молекулярный уровень. 1 вариант. 9 класс.
А1.Какой из химических элементов содержится в клетках в наибольшем количестве:
1.азот
2.кислород
3.углегод
4.водород
А2.Назовите химический элемент, который входит в состав АТФ, всех мономеров белков и нуклеиновых кислот.
1)N 2)P 3)S 4)Fe
А3.Укажите химическое соединение, которое углеводом НЕ является.
1)лактоза 2)хитин 3)кератин 4)крахмал
А4. Как называется структура белка, которая представляет собой спираль из цепочки аминокислот, свернутую в пространстве клубком?
Как называется структура белка, которая представляет собой спираль из цепочки аминокислот, свернутую в пространстве клубком?
1)первичная 2)вторичная 3)третичная 4)четвертичная
А5.В клетках животных запасным углеводом является:
1.крахмал
2.целлюлоза
3.глюкоза
4.гликоген
А6.Основным источником энергии для новорожденных млекопитающих является:
1.глюкоза
2.крахмал
3.гликоген
4.лактоза
А7.Что является мономером РНК?
1)азотистое основание 2)нуклеотид 3)рибоза 4)урацил
А8.Сколько видов азотистых оснований входит в состав молекулы РНК?
1)5 2)2 3)3 4)4
А9.Какое азотистое основание ДНК комплиментарно цитозину?
1)аденин 2)гуанин 3)урацил 4)тимин
А10. Универсальным биологическим аккумулятором энергии являются молекулы
1).белков 2).липидов 3).ДНК 4).АТФ
А11. В молекуле ДНК количество нуклеотидов с гуанином составляет 5% от общего числа. Сколько нуклеотидов с тимином содержится в этой молекуле
1).40% 2).45% 3).90% 4).95%
А12.Какова роль молекул АТФ в клетке?
1-обеспечивают транспортную функцию 2-передают наследственную информацию
3-обеспечивают процессы жизнедеятельности энергией 4-ускоряют биохимические
реакции
Часть В ( выберите три верных ответа из шести предложенных)
В1. Какие функции в клетке выполняют углеводы?
-
Каталитическую 4) структурную
-
Энергетическую 5) запасающую
-
Двигательную 6) сократительную
В2. Какие структурные компоненты входят в состав нуклеотидов молекулы ДНК?
-
Азотистые основания: А,Т,Г,Ц.
-
Разнообразные кислоты
-
Липопротеины
-
Углевод дезоксирибоза
-
Азотная кислота
-
Фосфорная кислота
В3. Установите соответствие между строением и функцией органического вещества и его видом:
СТРОЕНИЕ И ФУНКЦИИ ВЕЩЕСТВА
А. состоят из остатков молекул глицерина и жирных кислот 1. липиды
состоят из остатков молекул глицерина и жирных кислот 1. липиды
Б. состоят из остатков молекул аминокислот 2. Белки
В. Участвуют в терморегуляции
Г. Защищают организм от чужеродных веществ
Д. образуются за счет пептидных связей.
Е. Являются наиболее энергоемкими.
С1. Решите задачу.
В молекуле ДНК находится 1250 нуклеотидов с аденином (А), что составляет 20% от их общего числа. Определите, сколько нуклеотидов с тимином (Т), цитозином (Ц) и гуанином (Г) содержится в отдельности в молекуле ДНК. Ответ поясните.
Часть А – 1 балл (максимальное количество 12 баллов)
Часть В – 2 балла (максимальное количество 6 баллов
)
Часть С – 3 балла (максимальное количество 3 балла)
Итого : 21 балл
Критерии оценивания:
19 -21 балл – «5»
13 – 18 баллов – «4»
9 – 12 баллов – «3»
1 – 8 баллов – «2»
Тест. Молекулярный уровень. 2 вариант. 9 класс
А1.На долю четырех химических элементов приходится 98%всего содержимого клетки. Укажите химический элемент, НЕ относящийся к ним.
1)О 2)Р 3)С 4)N
А2.У детей развивается рахит при недостатке:
1.марганца и железа
2.кальция и фосфора
3.меди и цинка
4.серы и азота
А3.Назовите дисахарид.
1)лактоза 2)фруктоза 3)крахмал 4)гликоген
А4. Как называется структура белка, представляющая собой спираль , которую свернута цепочка из аминокислот?
1)первичная 2)вторичная 3)третичная 4)четвертичная
А5.В клетках растений запасным углеводом является:
1.крахмал
2.целлюлоза
3.глюкоза
4.гликоген
А6.Наибольшее количество энергии выделяется при разложении 1 грамма:
1.жира
2.белка
3.глюкоза
4.углеводов
А7.Что является мономером ДНК?
1)азотистое основание 2)нуклеотид 3)дезоксирибоза 4)урацил
А8.Сколько полинуклеотидных нитей входит в состав одной молекулы ДНК?
1)1 2)2 3)3 4)4
А9. Назовите химическое соединение, которое имеется в РНК, но отсутствует в ДНК.
Назовите химическое соединение, которое имеется в РНК, но отсутствует в ДНК.
1)тимин 2)дезоксмирибоза 3)рибоза 4)гуанин
А10. Источником энергии клетки являются молекулы
1).белков 2).липидов 3).ДНК 4).АТФ
А11. В молекуле ДНК количество нуклеотидов с цитозином составляет 5% от общего числа. Сколько нуклеотидов с тимином содержится в этой молекуле
1).40% 2).45% 3).90% 4).95%
А12.Какие соединения входят в состав АТФ?
1-азотистое основание аденин,углевод рибоза,3 молекулы фосфорной кислоты
2-азотистое основание гуанин, сахар фруктоза, остаток фосфорной кислоты.
3-рибоза,глицерин и какая-либо аминокислота
Часть В ( выберите три верных ответа из шести предложенных)
В1. Липиды выполняют функции:
-
Ферментативную 4) транспортную
-
Энергетическую 5) запасающую
-
Гормональную 6) передача наследственной информации
В2. Какие структурные компоненты входят в состав нуклеотидов молекулы РНК?
-
Азотистые основания: А,У,Г,Ц.
-
Разнообразные кислоты
-
Азотистые основания: А,Т,Г,Ц.
-
Углевод рибоза
-
Азотная кислота
-
Фосфорная кислота
В3. Установите соответствие между особенностями и молекулами для которых они характерны.
ОСОБЕННОСТИ МОЛЕКУЛЫ
А) хорошо растворяются в воде 1) моносахариды
Б) имеют сладкий вкус 2) полисахариды
В) сладкий вкус отсутствуют
Г) глюкоза, рибоза, фруктоза
Д) в воде нерастворимы
Е) крахмал, гликоген, хитин.
С1. В молекуле ДНК находится 1100 нуклеотидов с цитозином (Ц) что составляет 20% от их общего числа. Определите, сколько нуклеотидов с тимином (Т), гуанином(Г), аденином (А) содержится в отдельности в молекуле ДНК, объясните полученный результат.
Часть А – 1 балл (максимальное количество 12 баллов)
Часть В – 2 балла (максимальное количество 6 баллов)
Часть С – 3 балла (максимальное количество 3 балла)
Итого : 21 балл
Критерии оценивания:
19 — 21 балл – «5»
13 – 18 баллов – «4»
9 – 12 баллов – «3»
1 – 8 баллов – «2»
2.
 3.3. Белки, их строение и функции. Биология [Полный справочник для подготовки к ЕГЭ]
3.3. Белки, их строение и функции. Биология [Полный справочник для подготовки к ЕГЭ]
2.3.3. Белки, их строение и функции
Белки – это биологические гетерополимеры, мономерами которых являются аминокислоты. Белки синтезируются в живых организмах и выполняют в них определенные функции.
В состав белков входят атомы углерода, кислорода, водорода, азота и иногда серы. Мономерами белков являются аминокислоты – вещества, имеющие в своем составе неизменяемые части аминогруппу NH2 и карбоксильную группу СООН и изменяемую часть – радикал. Именно радикалами аминокислоты отличаются друг от друга. Аминокислоты обладают свойствами кислоты и основания (они амфотерны), поэтому могут соединяться друг с другом. Их количество в одной молекуле может достигать нескольких сотен. Чередование разных аминокислот в разной последовательности позволяет получать огромное количество различных по структуре и функциям белков.
В белках встречается 20 видов различных аминокислот, некоторые из которых животные синтезировать не могут. Они получают их от растений, которые могут синтезировать все аминокислоты. Именно до аминокислот расщепляются белки в пищеварительных трактах животных. Из этих аминокислот, поступающих в клетки организма, строятся его новые белки.
Структура белковой молекулы. Под структурой белковой молекулы понимают ее аминокислотный состав, последовательность мономеров и степень скрученности молекулы, которая должна умещаться в различных отделах и органоидах клетки, причем не одна, а вместе с огромным количеством других молекул.
Последовательность аминокислот в молекуле белка образует его первичную структуру. Она зависит от последовательности нуклеотидов в участке молекулы ДНК (гене), кодирующем данный белок. Соседние аминокислоты связаны пептидными связями, возникающими между углеродом карбоксильной группы одной аминокислоты и азотом аминогруппы другой аминокислоты.
Длинная молекула белка сворачивается и приобретает сначала вид спирали. Так возникает вторичная структура белковой молекулы. Между СО и NH – группами аминокислотных остатков, соседних витков спирали, возникают водородные связи, удерживающие цепь.
Так возникает вторичная структура белковой молекулы. Между СО и NH – группами аминокислотных остатков, соседних витков спирали, возникают водородные связи, удерживающие цепь.
Молекула белка сложной конфигурации в виде глобулы (шарика), приобретает третичную структуру. Прочность этой структуры обеспечивается гидрофобными, водородными, ионными и дисульфидными S-S связями.
Некоторые белки имеют четвертичную структуру, образованную несколькими полипептидными цепями (третичными структурами). Четвертичная структура так же удерживается слабыми нековалентными связями – ионными, водородными, гидрофобными. Однако прочность этих связей невелика и структура может быть легко нарушена. При нагревании или обработке некоторыми химическими веществами белок подвергается денатурации и теряет свою биологическую активность. Нарушение четвертичной, третичной и вторичной структур обратимо. Разрушение первичной структуры необратимо.
В любой клетке есть сотни белковых молекул, выполняющих различные функции. Кроме того, белки имеют видовую специфичность. Это означает, что каждый вид организмов обладает белками, не встречающимися у других видов. Это создает серьезные трудности при пересадке органов и тканей от одного человека к другому, при прививках одного вида растений на другой и т.д.
Функции белков. Каталитическая (ферментативная) – белки ускоряют все биохимические процессы, идущие в клетке: расщепление питательных веществ в пищеварительном тракте, участвуют в реакциях матричного синтеза. Каждый фермент ускоряет одну и только одну реакцию (как в прямом, так и в обратном направлении). Скорость ферментативных реакций зависит от температуры среды, уровня ее рН, а также от концентраций реагирующих веществ и концентрации фермента.
Транспортная – белки обеспечивают активный транспорт ионов через клеточные мембраны, транспорт кислорода и углекислого газа, транспорт жирных кислот.
Защитная – антитела обеспечивают иммунную защиту организма; фибриноген и фибрин защищают организм от кровопотерь.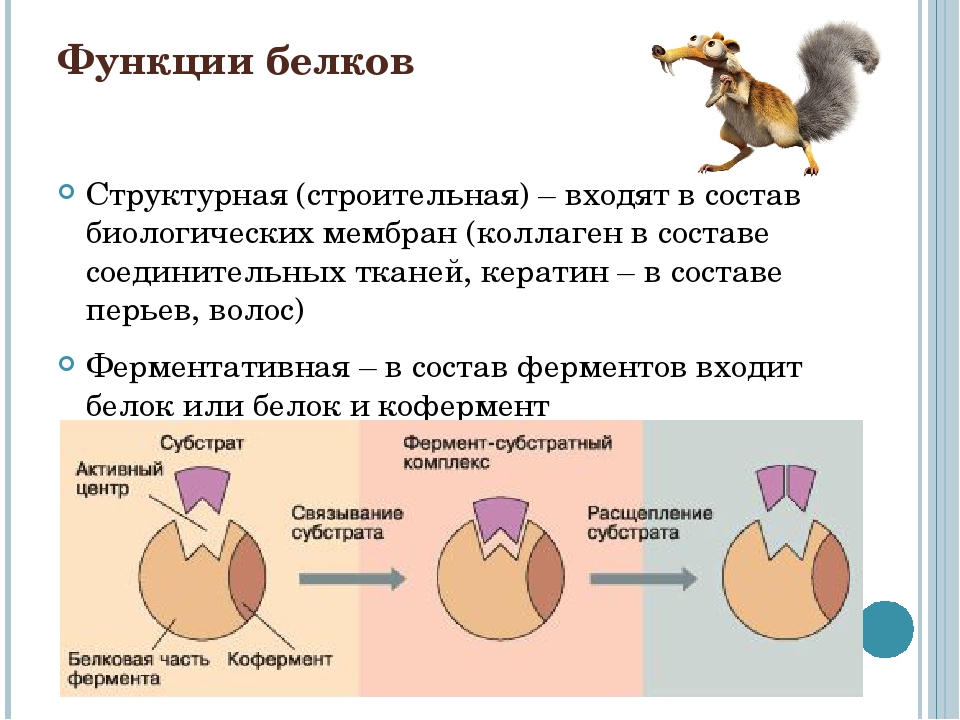
Структурная – одна из основных функций белков. Белки входят в состав клеточных мембран; белок кератин образует волосы и ногти; белки коллаген и эластин – хрящи и сухожилия.
Сократительная – обеспечивается сократительными белками – актином и миозином.
Сигнальная – белковые молекулы могут принимать сигналы и служить их переносчиками в организме (гормонами). Следует помнить, что не все гормоны являются белками.
Энергетическая – при длительном голодании белки могут использоваться в качестве дополнительного источника энергии после того, как израсходованы углеводы и жиры.
ПРИМЕРЫ ЗАДАНИЙ
Часть А
А1. Последовательность аминокислот в молекуле белка зависит от:
1) структуры гена 3) их случайного сочетания
2) внешней среды 4) их строения
А2. Человек получает незаменимые аминокислоты путем
1) их синтеза в клетках 3) приема лекарств
2) поступления с пищей 4) приема витаминов
А3. При понижении температуры активность ферментов
1) заметно повышается
2) заметно понижается
3) остается стабильной
4) периодически изменяется
А4. В защите организма от кровопотерь участвует
1) гемоглобин 3) фибрин
2) коллаген 4) миозин
А5. В каком из указанных процессов белки не участвуют?
обмен веществ
кодирование наследственной информации
ферментативный катализ
транспорт веществ
А6. Укажите пример пептидной связи:
Часть В
В1. Выберите функции, характерные для белков
1) каталитическая 4) транспортная
2) кроветворная 5) рефлекторная
3) защитная 6) фотосинтетическая
В2. Установите соответствие между структурой белковой молекулы и ее особенностями
Часть С
С1. Почему продукты хранят в холодильнике?
С2. Почему продукты, подвергшиеся тепловой обработке, хранятся дольше?
СЗ. Объясните понятие «специфичность» белка, и какое биологическое значение имеет специфичность?
Объясните понятие «специфичность» белка, и какое биологическое значение имеет специфичность?
С4. Прочитайте текст, укажите номера предложений, в которых допущены ошибки и объясните их 1) Большая часть химических реакций в организме катализируется ферментами. 2) Каждый фермент может катализировать множество типов реакций. 3) У фермента есть активный центр, геометрическая форма которого изменяется в зависимости от вещества, с которым фермент взаимодействует. 4) Примером действия фермента может быть разложение мочевины уреазой. 5) Мочевина разлагается на двуокись углерода и аммиак, которым пахнет кошачий лоток с песком. 6) За одну секунду уреаза расщепляет до 30 ООО молекул мочевины, в обычных условиях на это потребовалось бы около 3 млн лет.
Данный текст является ознакомительным фрагментом.
Продолжение на ЛитРес
|
ЮРИИ АНАТОЛЬЕВИЧ ОВЧИННИКОВ (родился в 1934 г.), академик, вице-президент АН СССР, председатель Секции химикотехнологических и биологических наук Президиума АН СССР, директор Института биоорганической химии им. М. М. Шемякина. Герой Социалистического Труда, лауреат Ленинской и Государственной премий.
|
 Автор более 150 научных трудов, в том числе 10 монографий.
Автор более 150 научных трудов, в том числе 10 монографий.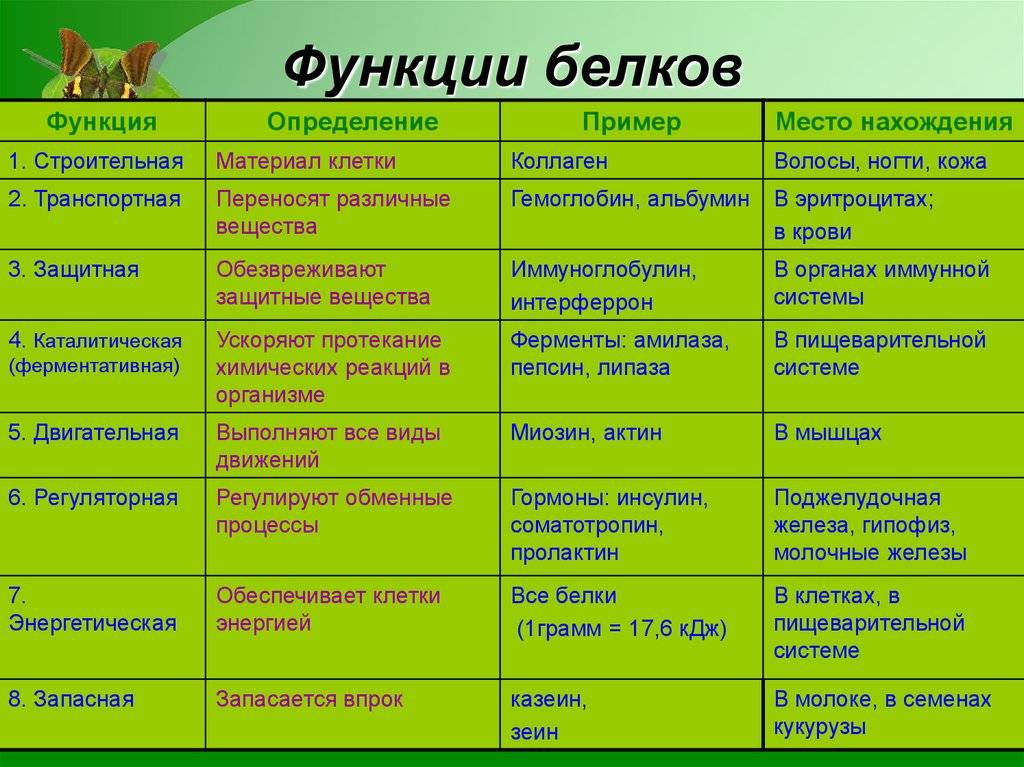
 Исследования оказались ориентированы на поиски специфических соединений, с которыми могли соединяться, притом строго специфично, пептидные гормоны. Естественно было предположить, что это тоже будут белки. Мысль была правильной, но ученые долго не могли выявить вещества, которые должны были играть роль «включателей — выключателей» для гормонов.
Исследования оказались ориентированы на поиски специфических соединений, с которыми могли соединяться, притом строго специфично, пептидные гормоны. Естественно было предположить, что это тоже будут белки. Мысль была правильной, но ученые долго не могли выявить вещества, которые должны были играть роль «включателей — выключателей» для гормонов. Оказалось, что они также могут быть объяснены с помощью пространственных взаимодействий с белковыми структурами.
Оказалось, что они также могут быть объяснены с помощью пространственных взаимодействий с белковыми структурами.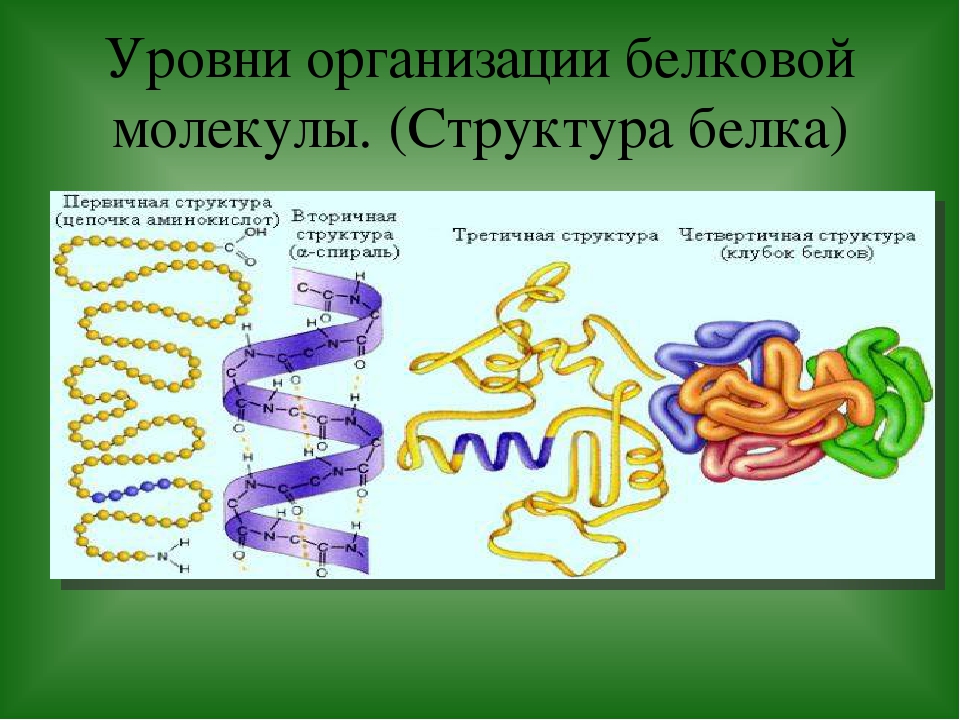 Поэтому, делая заключения о строении рецепторов, пользуются косвенными данными:- знанием конформации гормона, тех его участков, которые ответственны за связывание с рецептором.
Поэтому, делая заключения о строении рецепторов, пользуются косвенными данными:- знанием конформации гормона, тех его участков, которые ответственны за связывание с рецептором. Такими веществами могут быть и стероидные гормоны, и феромоны — вещества, влияющие на поведение насекомых и обмен веществ в их сообществах, и т. д.
Такими веществами могут быть и стероидные гормоны, и феромоны — вещества, влияющие на поведение насекомых и обмен веществ в их сообществах, и т. д.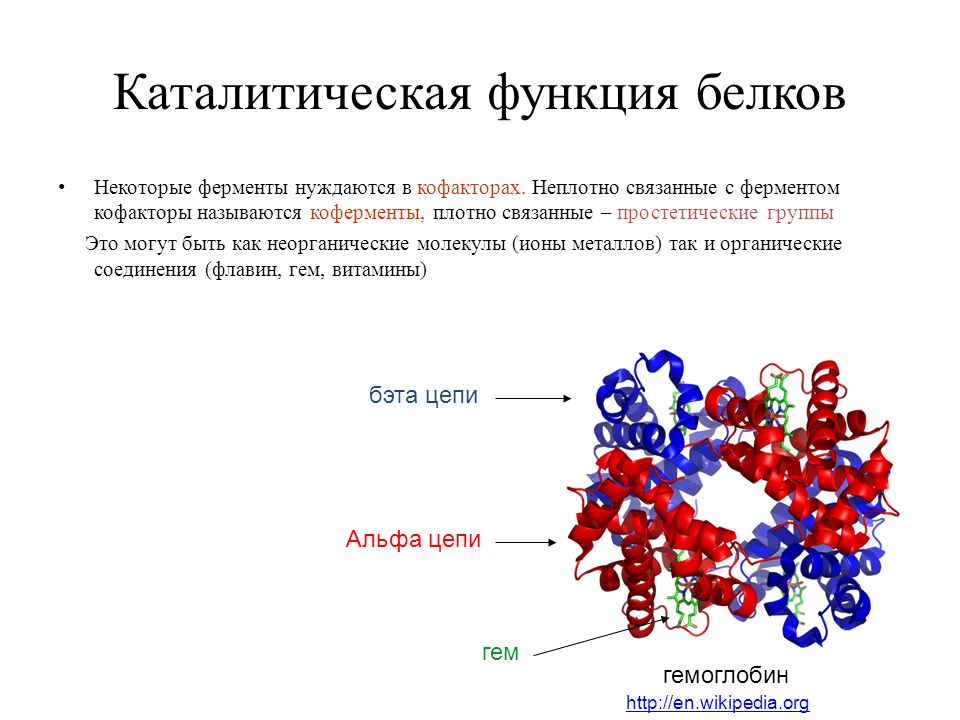
 Правда, ряд специалистов в области биохимии насекомых считает, что нельзя проводить параллель между органами обоняния животных и органами фиксации запахов насекомых. Они полагают, что органы распознавания присутствия органических веществ в среде у насекомых физиологически отличаются от органов обоняния насекомых и биологический смысл этих процессов различен. Говорить, что насекомые обладают органами обоняния, возможно, скорее приписывать им свойства, которыми обладаем мы с вами. Но возможно, что это и не так. Скорее всего, исследуя насекомых, удалось обнаружить явление, в той или иной мере присущее всем представителям мира животных, включая и человека. Во всяком случае, сейчас известно, что сходный механизм взаимного ориентирования имеется у многих морских животных, включая китов. Правда, нельзя говорить о «запахе», имея в виду присутствие ничтожных концентраций некоторых веществ в толще воды, но биологический смысл их распознавания близок тому, которым обладают насекомые. Последние отличаются лишь совершенно особым устройством своих органов восприятия запахов и их необычайной чувствительностью, но молекулярные основы их функционирования, безусловно, очень близки всему миру животных. Надо отметить также, что открытие химической коммуникации в мире животных считается одним из самых важных биологических открытий последних десятилетий и сейчас привлекает пристальное внимание.
Правда, ряд специалистов в области биохимии насекомых считает, что нельзя проводить параллель между органами обоняния животных и органами фиксации запахов насекомых. Они полагают, что органы распознавания присутствия органических веществ в среде у насекомых физиологически отличаются от органов обоняния насекомых и биологический смысл этих процессов различен. Говорить, что насекомые обладают органами обоняния, возможно, скорее приписывать им свойства, которыми обладаем мы с вами. Но возможно, что это и не так. Скорее всего, исследуя насекомых, удалось обнаружить явление, в той или иной мере присущее всем представителям мира животных, включая и человека. Во всяком случае, сейчас известно, что сходный механизм взаимного ориентирования имеется у многих морских животных, включая китов. Правда, нельзя говорить о «запахе», имея в виду присутствие ничтожных концентраций некоторых веществ в толще воды, но биологический смысл их распознавания близок тому, которым обладают насекомые. Последние отличаются лишь совершенно особым устройством своих органов восприятия запахов и их необычайной чувствительностью, но молекулярные основы их функционирования, безусловно, очень близки всему миру животных. Надо отметить также, что открытие химической коммуникации в мире животных считается одним из самых важных биологических открытий последних десятилетий и сейчас привлекает пристальное внимание. В 1971 г. В. Стокениус и Д. Остерхельт выделили интересный белок из суспензии пурпурных мембран, полученных из клеточной стенки галофильных (солеустойчивых) бактерий. Этот белок был назван бактериородопсином.
В 1971 г. В. Стокениус и Д. Остерхельт выделили интересный белок из суспензии пурпурных мембран, полученных из клеточной стенки галофильных (солеустойчивых) бактерий. Этот белок был назван бактериородопсином. Ученые с помощью химических методов смогли «разобрать» клетку на части и «вынуть» из нее ядро. Это сделал Ф. Мишер, швейцарский ученый. Его открытие противоречило всем догмам биологии того времени. Ведь и клетка, и протоплазма считались коллоидным образованием, этаким раствором, в котором плавали капельки: капелька-ядро, капельки-включения, вакуоли и т. д. Капельки изучали, либо окрашивая их различными красками, либо фиксируя, переводя в нерастворимое состояние. Но считали, что при этом природа клеточного содержимого, протоплазмы безвозвратно нарушалась. Никаких данных о ее функционировании получить не удавалось. Можно было лишь приблизительно узнать, какие вещества входят в ее состав.
Ученые с помощью химических методов смогли «разобрать» клетку на части и «вынуть» из нее ядро. Это сделал Ф. Мишер, швейцарский ученый. Его открытие противоречило всем догмам биологии того времени. Ведь и клетка, и протоплазма считались коллоидным образованием, этаким раствором, в котором плавали капельки: капелька-ядро, капельки-включения, вакуоли и т. д. Капельки изучали, либо окрашивая их различными красками, либо фиксируя, переводя в нерастворимое состояние. Но считали, что при этом природа клеточного содержимого, протоплазмы безвозвратно нарушалась. Никаких данных о ее функционировании получить не удавалось. Можно было лишь приблизительно узнать, какие вещества входят в ее состав. Все эти процессы еще не разгаданы до конца.
Все эти процессы еще не разгаданы до конца.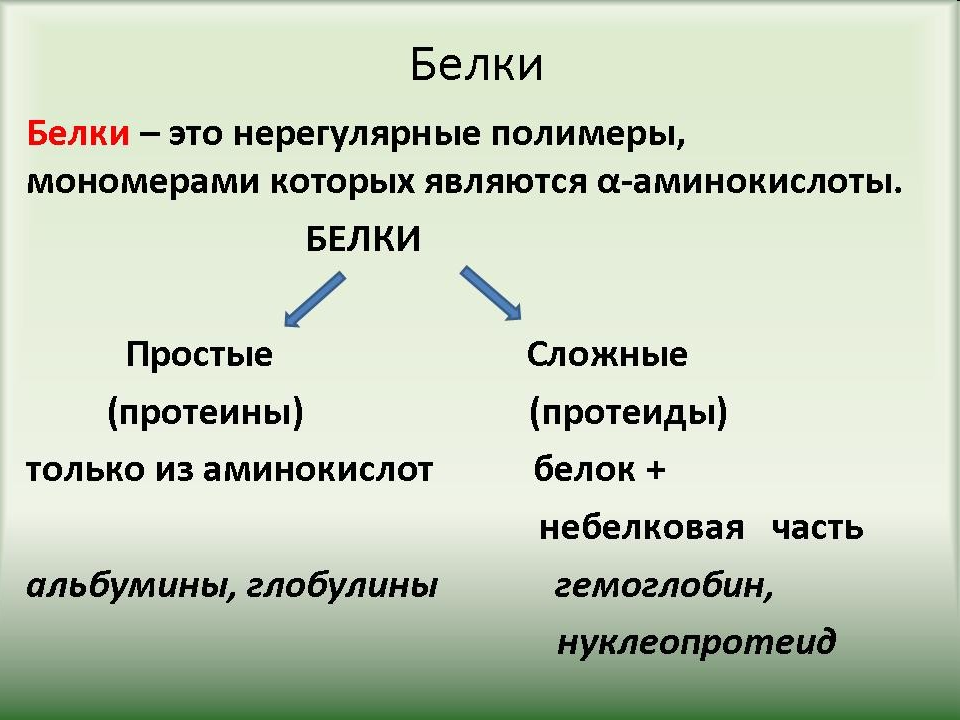 В коротких токсинах 4 внутримолекулярных дисуль-фидных мостика, а в длинных — 5. Эти токсины обладают одним важным свойством — они очень устойчивы. Их можно кипятить в течение 30 минум, и они не потеряют своей активности. Их можно обрабатывать различными, вызывающими денатурацию белков веществами — они также не потеряют активности. Их «ядовитость» обусловлена тем, что они образуют исключительно прочные связи с рецепторами. В результате их удалось использовать для выявления рецепторов практически в чистом виде — получали комплекс нейротоксин-рецептор, подобно тому, как комплекс фермент-ингибитор.
В коротких токсинах 4 внутримолекулярных дисуль-фидных мостика, а в длинных — 5. Эти токсины обладают одним важным свойством — они очень устойчивы. Их можно кипятить в течение 30 минум, и они не потеряют своей активности. Их можно обрабатывать различными, вызывающими денатурацию белков веществами — они также не потеряют активности. Их «ядовитость» обусловлена тем, что они образуют исключительно прочные связи с рецепторами. В результате их удалось использовать для выявления рецепторов практически в чистом виде — получали комплекс нейротоксин-рецептор, подобно тому, как комплекс фермент-ингибитор. Мало того, иногда они представляли собой фрагменты других, более сложно устроенных гормонов. Наибольший интерес вызвали пептиды — регуляторы поведения, так называемые нейропептиды. Нейропептиды связаны с проявлением многих поведенческих реакций. Такие ощущения, как пищевое насыщение, жажда, удовольствие, а также боль, обучаемость и память, двигательная активность, сон и зимняя спячка и многое другое (включая элементы симптомов психических заболеваний), вызываются различными пептидами, среди которых уже известные нам вазопрес-син, фрагменты кортикотропина, а также пептиды холецистокинин, ангиотензин П, пептид дельта-сна, люлиберин, а также очень интересная группа так называемых опиоидных пептидов (Met-энкефалин, Р-эндорфин, брадикинин, пептид Р, нейротензин и т. д.).
Мало того, иногда они представляли собой фрагменты других, более сложно устроенных гормонов. Наибольший интерес вызвали пептиды — регуляторы поведения, так называемые нейропептиды. Нейропептиды связаны с проявлением многих поведенческих реакций. Такие ощущения, как пищевое насыщение, жажда, удовольствие, а также боль, обучаемость и память, двигательная активность, сон и зимняя спячка и многое другое (включая элементы симптомов психических заболеваний), вызываются различными пептидами, среди которых уже известные нам вазопрес-син, фрагменты кортикотропина, а также пептиды холецистокинин, ангиотензин П, пептид дельта-сна, люлиберин, а также очень интересная группа так называемых опиоидных пептидов (Met-энкефалин, Р-эндорфин, брадикинин, пептид Р, нейротензин и т. д.). Но ведь клетка — очень точный механизм. Ее форма строго запрограммирована. Информация об этой программе заключена в генах — молекулах ДНК, реализуется она через синтез белка.
Но ведь клетка — очень точный механизм. Ее форма строго запрограммирована. Информация об этой программе заключена в генах — молекулах ДНК, реализуется она через синтез белка.
 Нуклеиновые кислоты выполняют функции хранения и передачи наследственной информации в клетке. А белки являются движителями всех процессов, протекающих в клетке.
Нуклеиновые кислоты выполняют функции хранения и передачи наследственной информации в клетке. А белки являются движителями всех процессов, протекающих в клетке.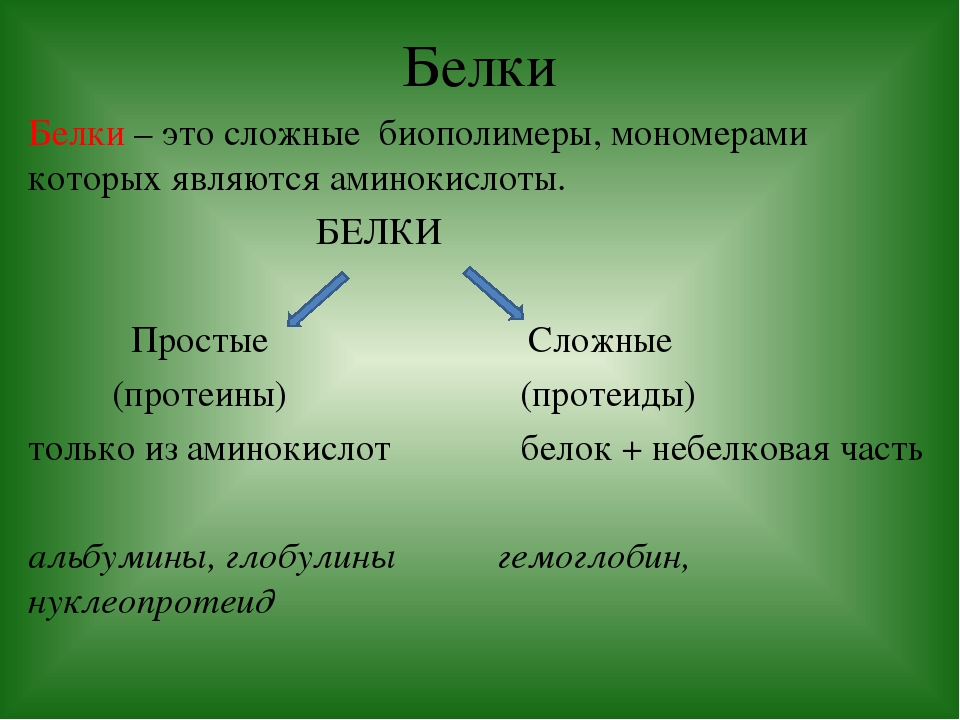 Вы уже знаете, что жизнь — это процесс поддержания и воспроизведения специфической структуры. Для воспроизведения специфической структуры может быть достаточен (сразу оговоримся, что не абсолютно) процесс воссоздания определенной специфической последовательности аминокислотных остатков. Так с молекулярной структурой белков оказывается связана форма биологических объектов.
Вы уже знаете, что жизнь — это процесс поддержания и воспроизведения специфической структуры. Для воспроизведения специфической структуры может быть достаточен (сразу оговоримся, что не абсолютно) процесс воссоздания определенной специфической последовательности аминокислотных остатков. Так с молекулярной структурой белков оказывается связана форма биологических объектов.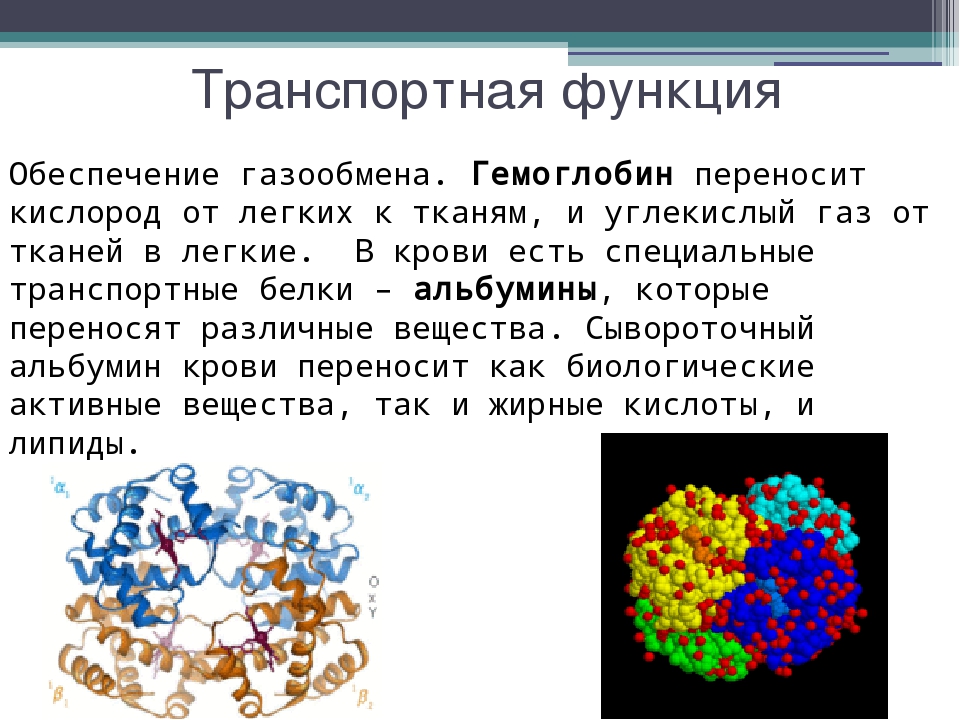 А окончательное соединение осуществляется за счет ван-дер-ваальсовых связей. Основой же взаимодействия является стерическая дополнительность молекул белков и нуклеиновых кислот — тоже одно из проявлений принципа «ключа и замка» или принципа «узнавания».
А окончательное соединение осуществляется за счет ван-дер-ваальсовых связей. Основой же взаимодействия является стерическая дополнительность молекул белков и нуклеиновых кислот — тоже одно из проявлений принципа «ключа и замка» или принципа «узнавания».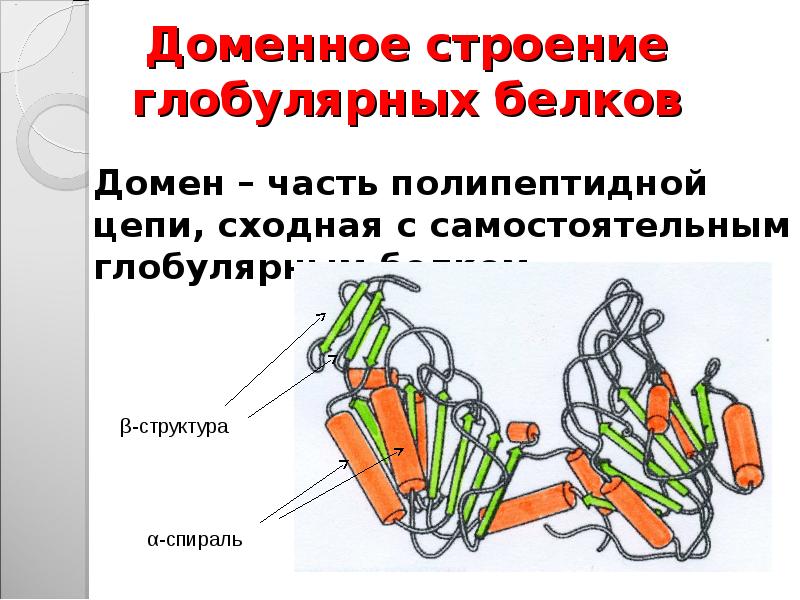
Строение и функции белков — презентация онлайн
1. Строение и функции белков
СТРОЕНИЕ И ФУНКЦИИ
БЕЛКОВ
2. Строение белков
СТРОЕНИЕ БЕЛКОВ
Белки — высокомолекулярные органические
соединения, состоящие из остатков αаминокислот.
В состав белков входят углерод, водород, азот,
кислород, сера. Часть белков образует комплексы
с другими молекулами, содержащими фосфор,
железо, цинк и медь.
Белки обладают большой молекулярной массой:
яичный альбумин — 36 000, гемоглобин —
152 000, миозин — 500 000. Для сравнения:
молекулярная масса спирта — 46, уксусной
кислоты — 60, бензола — 78.
3. Аминокислотный состав белков
АМИНОКИСЛОТНЫЙ СОСТАВ БЕЛКОВ
Белки — непериодические полимеры,
мономерами которых являются α-аминокислоты.
Обычно в качестве мономеров белков называют
20 видов α-аминокислот, хотя в клетках и тканях
их обнаружено свыше 170.
В зависимости от того, могут ли аминокислоты
синтезироваться в организме человека и других
животных, различают: заменимые аминокислоты
— могут синтезироваться; незаменимые
аминокислоты — не могут синтезироваться.
Незаменимые аминокислоты должны поступать в
организм вместе с пищей. Растения синтезируют
все виды аминокислот.
4. Аминокислотный состав белков
АМИНОКИСЛОТНЫЙ СОСТАВ БЕЛКОВ
В зависимости от аминокислотного состава,
белки бывают:
полноценными — содержат весь набор
аминокислот;
неполноценными — какие-то аминокислоты в их
составе отсутствуют. Если белки состоят только из
Если белки состоят только из
аминокислот, их называют простыми. Если белки
содержат помимо аминокислот еще и
неаминокислотный компонент (простетическую
группу), их называют сложными. Простетическая
группа может быть представлена металлами
(металлопротеины), углеводами (гликопротеины),
липидами (липопротеины), нуклеиновыми
кислотами (нуклеопротеины).
5. Аминокислотный состав белков
АМИНОКИСЛОТНЫЙ СОСТАВ БЕЛКОВ
Все аминокислоты содержат: 1) карбоксильную группу (–
СООН), 2) аминогруппу (–Nh3), 3) радикал или R-группу
(остальная часть молекулы). Строение радикала у разных
видов аминокислот — различное. В зависимости от
количества аминогрупп и карбоксильных групп, входящих в
состав аминокислот, различают: нейтральные
аминокислоты, имеющие одну карбоксильную группу и
одну аминогруппу; основные аминокислоты, имеющие
более одной аминогруппы; кислые аминокислоты,
имеющие более одной карбоксильной группы.
аминогруппа
карбоксильная
группа
6. Пептидная связь
ПЕПТИДНАЯ СВЯЗЬ
Пептиды — органические вещества, состоящие из остатков аминокислот,
соединенных пептидной связью.
Образование пептидов происходит в результате реакции конденсации
аминокислот.
При взаимодействии аминогруппы одной аминокислоты с карбоксильной
группой другой между ними возникает ковалентная азот-углеродная связь,
которую и называют пептидной. В зависимости от количества
аминокислотных остатков, входящих в состав пептида, различают
дипептиды, трипептиды, тетрапептиды и т.д. Образование пептидной связи
может повторяться многократно.
Это приводит к образованию полипептидов. На одном конце пептида
находится свободная аминогруппа (его называют N-концом), а на другом
— свободная карбоксильная группа (его называют С-концом).
7. Пространственная организация белковых молекул
ПРОСТРАНСТВЕННАЯ ОРГАНИЗАЦИЯ
БЕЛКОВЫХ МОЛЕКУЛ
Выполнение белками определенных
специфических функций зависит от
пространственной конфигурации их молекул,
кроме того, клетке энергетически невыгодно
держать белки в развернутой форме, в виде
цепочки, поэтому полипептидные цепи
подвергаются укладке, приобретая
определенную трехмерную структуру, или
конформацию. Выделяют 4 уровня
Выделяют 4 уровня
пространственной организации белков:
первичный, вторичный, третичный и
четвертичный.
9. Первичная структура белка
ПЕРВИЧНАЯ СТРУКТУРА БЕЛКА
Это последовательность расположения аминокислотных
остатков в полипептидной цепи, составляющей молекулу
белка. Связь между аминокислотами — пептидная.
Если молекула белка состоит всего из 10 аминокислотных остатков, то число
теоретически возможных вариантов белковых молекул, отличающихся порядком
чередования аминокислот, — 1020. Имея 20 аминокислот, можно составить из них
еще большее количество разнообразных комбинаций. В организме человека
обнаружено порядка десяти тысяч различных белков, которые отличаются как друг от
друга, так и от белков других организмов.
10. Вторичная структура
ВТОРИЧНАЯ СТРУКТУРА
Это упорядоченное свертывание полипептидной цепи в спираль
(имеет вид растянутой пружины). Витки спирали укрепляются
водородными связями, возникающими между карбоксильными
группами и аминогруппами.
Практически все СО- и NН-группы принимают участие в
образовании водородных связей. Они слабее пептидных, но,
повторяясь многократно, придают данной конфигурации
устойчивость и жесткость. На уровне вторичной структуры
существуют белки: фиброин (шелк, паутина), кератин (волосы,
ногти), коллаген (сухожилия).
11. Третичная структура
ТРЕТИЧНАЯ СТРУКТУРА
укладка полипептидных цепей в глобулы, возникающая в результате
возникновения химических связей (водородных, ионных, дисульфидных) и
установления гидрофобных взаимодействий между радикалами
аминокислотных остатков.
Основную роль в образовании третичной структуры играют гидрофильногидрофобные взаимодействия. В водных растворах гидрофобные
радикалы стремятся спрятаться от воды, группируясь внутри глобулы, в то
время как гидрофильные радикалы в результате гидратации
(взаимодействия с диполями воды) стремятся оказаться на поверхности
молекулы. У некоторых белков третичная структура стабилизируется
У некоторых белков третичная структура стабилизируется
дисульфидными ковалентными связями, возникающими между атомами
серы двух остатков цистеина.
На уровне третичной структуры существуют ферменты, антитела,
некоторые гормоны.
12. Четвертичная структура
ЧЕТВЕРТИЧНАЯ СТРУКТУРА
Она характерна для сложных белков, молекулы которых
образованы двумя и более глобулами. Субъединицы
удерживаются в молекуле благодаря ионным, гидрофобным и
электростатическим взаимодействиям. Иногда при
образовании четвертичной структуры между субъединицами
возникают дисульфидные связи.
Наиболее изученным белком, имеющим четвертичную
структуру, является гемоглобин. Он образован двумя αсубъединицами (141 аминокислотный остаток) и двумя βсубъединицами (146 аминокислотных остатков). С каждой
субъединицей связана молекула гема, содержащая железо.
13. Свойства белков
СВОЙСТВА БЕЛКОВ
Белки сочетают в себе основные и кислотные
свойства, определяемые радикалами аминокислот:
чем больше кислых аминокислот в белке, тем ярче
выражены его кислотные свойства.
Способность отдавать и присоединять Н+ определяют
буферные свойства белков; один из самых мощных
буферов — гемоглобин в эритроцитах,
поддерживающий рН крови на постоянном уровне.
Есть белки растворимые (фибриноген), есть
нерастворимые, выполняющие механические функции
(фиброин, кератин, коллаген).
Есть белки активные в химическом отношении
(ферменты), есть химически неактивные, устойчивые к
воздействию различных условий внешней среды и
крайне неустойчивые.
14. Свойства белков
СВОЙСТВА БЕЛКОВ
Внешние факторы (нагревание, ультрафиолетовое излучение, тяжелые
металлы и их соли, изменения рН, радиация, обезвоживание) могут
вызывать нарушение структурной организации молекулы белка.
Процесс утраты трехмерной конформации, присущей данной молекуле
белка, называют денатурацией. Причиной денатурации является
Причиной денатурации является
разрыв связей, стабилизирующих определенную структуру белка.
Первоначально рвутся наиболее слабые связи, а при ужесточении
условий и более сильные. Поэтому сначала утрачивается четвертичная,
затем третичная и вторичная структуры. Изменение пространственной
конфигурации приводит к изменению свойств белка и, как следствие,
делает невозможным выполнение белком свойственных ему
биологических функций.
Если денатурация не сопровождается разрушением первичной
структуры, то она может быть обратимой, в этом случае происходит
самовосстановление свойственной белку конформации. Такой
денатурации подвергаются, например, рецепторные белки мембраны.
Процесс восстановления структуры белка после денатурации
называется ренатурацией. Если восстановление пространственной
конфигурации белка невозможно, то денатурация называется
необратимой.
15. Функции белков
ФУНКЦИИ БЕЛКОВ
Функция
Примеры и пояснения
Строительная
Белки участвуют в образовании клеточных и внеклеточных структур: входят в состав клеточных мембран
(липопротеины, гликопротеины), волос (кератин), сухожилий (коллаген) и т.д.
Транспортная
Белок крови гемоглобин присоединяет кислород и транспортирует его от легких ко всем тканям и органам, а от них в
легкие переносит углекислый газ; в состав клеточных мембран входят особые белки, которые обеспечивают
активный и строго избирательный перенос некоторых веществ и ионов из клетки во внешнюю среду и обратно.
Регуляторная
Гормоны белковой природы принимают участие в регуляции процессов обмена веществ. Например, гормон инсулин
регулирует уровень глюкозы в крови, способствует синтезу гликогена, увеличивает образование жиров из углеводов.
Защитная
В ответ на проникновение в организм чужеродных белков или микроорганизмов (антигенов) образуются особые
белки — антитела, способные связывать и обезвреживать их.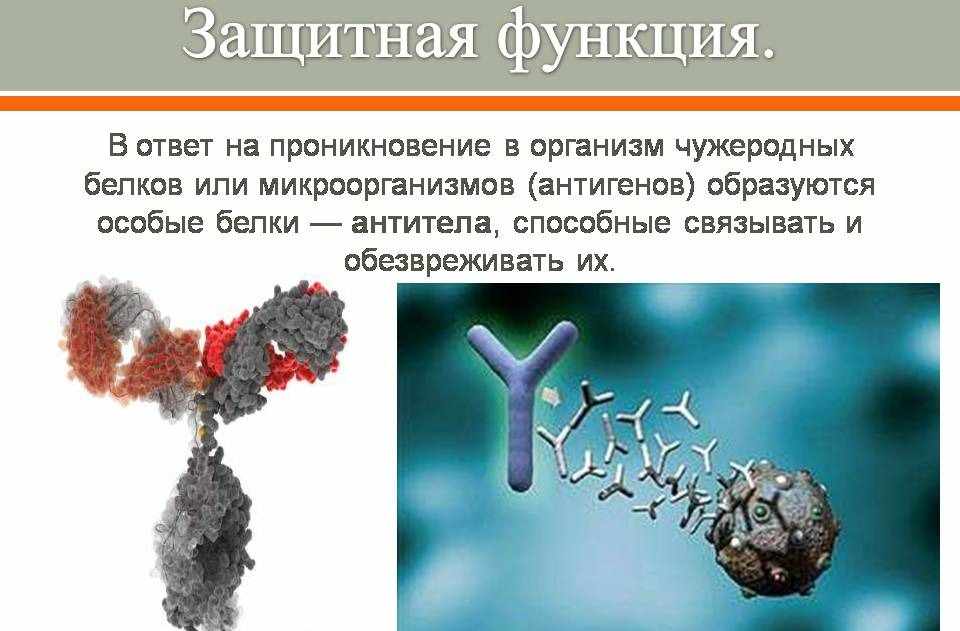 Фибрин, образующийся из фибриногена, способствует
Фибрин, образующийся из фибриногена, способствует
остановке кровотечений.
Двигательная
Сократительные белки актин и миозин обеспечивают сокращение мышц у многоклеточных животных.
Сигнальная
В поверхностную мембрану клетки встроены молекулы белков, способных изменять свою третичную структуру в
ответ на действие факторов внешней среды, таким образом осуществляя прием сигналов из внешней среды и
передачу команд в клетку.
Запасающая
В организме животных белки, как правило, не запасаются, исключение: альбумин яиц, казеин молока. Но благодаря
белкам в организме могут откладываться про запас некоторые вещества, например, при распаде гемоглобина железо
не выводится из организма, а сохраняется, образуя комплекс с белком ферритином.
При распаде 1 г белка до конечных продуктов выделяется 17,6 кДж. Сначала белки распадаются до аминокислот, а
Энергетическая затем до конечных продуктов — воды, углекислого газа и аммиака. Однако в качестве источника энергии белки
используются только тогда, когда другие источники (углеводы и жиры) израсходованы.
Одна из важнейших функций белков. Обеспечивается белками — ферментами, которые ускоряют биохимические
Каталитическая реакции, происходящие в клетках. Например, рибулезобифосфаткарбоксилаза катализирует фиксацию СО2 при
фотосинтезе.
Структура и функции белка
Структура белка закладывает основу для его взаимодействия с другими молекулами организма и, следовательно, определяет его функцию. В этой статье будут рассмотрены структурные принципы белков и их влияние на функцию белка.
Первичная структура белка
Белки состоят из длинной цепи аминокислот. Даже при ограниченном количестве аминокислотных мономеров — в организме человека обычно встречается всего 20 аминокислот — их можно расположить множеством способов, чтобы изменить трехмерную структуру и функцию белка.Простое секвенирование белка известно как его первичная структура.
Вторичная структура белка
Вторичная структура белка зависит от локальных взаимодействий между частями белковой цепи, которые могут влиять на укладку и трехмерную форму белка. Есть две основные вещи, которые могут изменить вторичную структуру:
- α-спираль: группы N-H в основной цепи образуют водородную связь с группой C = O четырех аминокислотных остатков ранее в спирали.
- β-складчатый лист: группы N-H в основной цепи одной нити образуют водородные связи с группами C = O в основной цепи полностью вытянутой цепи рядом с ней.
Также может быть несколько функциональных групп, таких как спирты, карбоксамины, карбоновые кислоты, тиоэфиры, тиолы и другие основные группы, связанные с каждым белком. Эти функциональные группы также влияют на складывание белков и, следовательно, на их функцию в организме.
Третичная структура
Третичная структура белков относится к общей трехмерной форме после вторичных взаимодействий.К ним относится влияние полярных, неполярных, кислотных и основных R-групп, которые существуют на белке.
Четвертичный белок
Четвертичная структура белка относится к ориентации и расположению субъединиц в белках с мульти-субъединицами. Это актуально только для белков с несколькими полипептидными цепями.
Белки складываются в определенные формы в соответствии с последовательностью аминокислот в полимере, и функция белка напрямую связана с полученной трехмерной структурой.
Белки могут также взаимодействовать друг с другом или с другими макромолекулами в организме, создавая сложные сборки. В этих сборках белки могут развивать функции, которые были невозможны в автономном белке, такие как выполнение репликации ДНК и передача клеточных сигналов.
Природа белков также очень разнообразна. Например, некоторые из них довольно жесткие, а другие несколько гибкие. Эти характеристики также соответствуют функции белка. Например, более жесткие белки могут играть роль в структуре цитоскелета или соединительных тканей. С другой стороны, те, у кого есть некоторая гибкость, могут действовать как шарниры, пружины или рычаги, помогая в работе других белков.
С другой стороны, те, у кого есть некоторая гибкость, могут действовать как шарниры, пружины или рычаги, помогая в работе других белков.
Функции белков
Белки играют важную роль во многих важных биологических процессах и функциях. Они очень универсальны и выполняют множество различных функций в организме, как указано ниже:
- Действовать как катализатор
- Транспортные другие молекулы
- Хранить другие молекулы
- Обеспечить механическую поддержку
- Обеспечивает иммунную защиту
- Создать движение
- Передавать нервные импульсы
- Контроль роста и дифференцировки клеток
Степень, в которой структура белков влияет на их функцию, демонстрируется влиянием изменений в структуре белка.Любое изменение белка на любом структурном уровне, включая небольшие изменения в укладке и форме белка, может сделать его нефункциональным.
Список литературы
- https://www.ncbi.nlm.nih.gov/books/NBK21177/
- http://genome.tugraz.at/MolecularBiology/WS11_Chapter03.pdf
- https://www.boundless.com/biology/textbooks/boundless-biology-textbook/biological-macromolecules-3/proteins-56/protein-structure-304-11437/
Что такое белки и для чего они нужны? — Биология муниципального колледжа Маунт-Худ 102
Как у прокариот, так и у эукариот, основная цель ДНК — предоставить информацию, необходимую для создания белков, необходимых для того, чтобы клетка могла выполнять все свои функции.Белки — это большие сложные молекулы, которые играют важную роль в организме. Они выполняют большую часть работы в клетках и необходимы для структуры, функции и регулирования тканей и органов тела.
Белки состоят из сотен или тысяч более мелких единиц, называемых аминокислотами, которые связаны друг с другом длинными цепями. Существует 20 различных типов аминокислот, которые можно комбинировать для получения белка. Последовательность аминокислот определяет уникальную трехмерную структуру каждого белка и его конкретную функцию.
Белки можно описать в соответствии с их широким спектром функций в организме, перечисленных в алфавитном порядке:
| Функция |
Описание |
| Антитело
|
Антитела связываются с определенными инородными частицами, такими как вирусы и бактерии, чтобы защитить организм. |
| Фермент
|
Ферменты осуществляют почти все тысячи химических реакций, протекающих в клетках.Они также помогают формированию новых молекул, считывая генетическую информацию, хранящуюся в ДНК. |
| Посланник
|
Белки-мессенджеры, такие как некоторые типы гормонов, передают сигналы для координации биологических процессов между различными клетками, тканями и органами. |
| Конструктивный элемент
|
Эти белки обеспечивают структуру и поддержку клеток. В большем масштабе они также позволяют телу двигаться. |
| Транспортировка / хранение
|
Эти белки связывают и переносят атомы и небольшие молекулы внутри клеток и по всему телу. |
Вы могли заметить, что «источник энергии» не был указан среди функций белков. Это связано с тем, что белки в нашем рационе обычно снова расщепляются на отдельные аминокислоты, которые наши клетки затем собирают в наши собственные белки. На самом деле люди не могут вырабатывать некоторые аминокислоты внутри наших собственных клеток — они необходимы нам в нашем рационе (это так называемые «незаменимые» аминокислоты).Наши клетки могут переваривать белки для высвобождения энергии, но обычно это происходит только тогда, когда углеводы или липиды недоступны.
Рис. 4: Примеры продуктов с высоким содержанием белка. («Белок» Национального института рака находится в открытом доступе)
Функции белков очень разнообразны, потому что они состоят из 20 различных химически различных аминокислот, которые образуют длинные цепи, и аминокислоты могут располагаться в любом порядке. Функция белка зависит от формы белка.Форма белка определяется порядком аминокислот. Белки часто состоят из сотен аминокислот и могут иметь очень сложную форму, потому что существует очень много различных возможных порядков для 20 аминокислот!
Рисунок 5: Основные уровни структуры белка. («Основные уровни структуры белка en» от LadyofHats находится в открытом доступе)
Уникальная форма каждого белка в конечном итоге определяется геном, кодирующим этот белок. Любое изменение в последовательности гена может привести к добавлению другой аминокислоты к полипептидной цепи, вызывая изменение структуры и функции белка.Люди, страдающие серповидно-клеточной анемией, могут иметь множество серьезных проблем со здоровьем, таких как одышка, головокружение, головные боли и боли в животе. При этом заболевании β-цепь гемоглобина имеет единственную аминокислотную замену, вызывающую изменение как структуры (формы), так и функции (работы) белка. Примечательно то, что молекула гемоглобина состоит примерно из 600 аминокислот. Структурная разница между нормальной молекулой гемоглобина и молекулой серповидно-клеточных клеток заключается в одной из 600 аминокислот.
Рисунок 6: Уникальная форма нормального белка гемоглобина. («Структура гемоглобина Gower 2» от Emw под лицензией CC BY-SA 3.0)
Если не указано иное, изображения на этой странице лицензированы OpenStax в соответствии с CC-BY 4.0.
OpenStax, Биология. OpenStax CNX. 27 мая 2016 г. http://cnx.org/contents/[email protected]:QhGQhr4x@6/Biological-Molecules
«Что такое белки и для чего они нужны?» Национальной медицинской библиотеки США находится в общественном достоянии
Комплексный анализ взаимосвязи структура-функция в белках на основе сходства локальной структуры
Аннотация
Фон
Сходство последовательностей с охарактеризованными белками обеспечивает проверяемые функциональные гипотезы для менее чем 50% белков, идентифицированных в проектах секвенирования генома.В структурной геномике считается, что структурное сходство может дать функциональные гипотезы для многих оставшихся белков.
Методология / основные выводы
Мы обеспечиваем систематический анализ взаимосвязи структура-функция в белках, используя новую концепцию локальных дескрипторов структуры белка. Локальный дескриптор — это небольшая субструктура белка, которая включает как короткие, так и дальние взаимодействия. Мы используем библиотеку часто встречающихся локальных дескрипторов, достаточно общих для сборки большинства существующих белковых структур.Затем мы моделируем отношения между этими локальными формами и генной онтологией, используя обучение на основе правил. Наша модель правил IF-THEN предлагает четкие описания с высоким разрешением, которые объединяют локальные подструктуры и способны различать функции даже для функционально универсальных складок, таких как часто встречающийся ствол TIM и складка Россманна. Оценивая прогностическую эффективность модели, мы обеспечиваем исчерпывающую количественную оценку взаимосвязи структура-функция, основанную только на локальном сходстве структуры.Наши результаты, среди прочего, заключаются в том, что консервативная структура является более сильным условием для ферментативной активности, чем для специфичности связывания, и что предсказания, основанные на структуре, дополняют предсказания, основанные на последовательностях. Модель способна генерировать правильные гипотезы, что подтверждается литературными исследованиями, даже когда не существует значительного сходства последовательностей с охарактеризованными белками.
Выводы / Значение
Наш подход предлагает новое и полное описание и количественную оценку взаимосвязи структура-функция в белках.Демонстрируя, как наши прогнозы обеспечивают более высокую чувствительность, чем использование глобальной структуры, и дополняют использование последовательности, мы показываем, что представленные идеи могут способствовать развитию мета-серверов в прогнозировании функций.
Образец цитирования: Hvidsten TR, Lægreid A, Kryshtafovych A, Andersson G, Fidelis K, Komorowski J (2009) Комплексный анализ взаимосвязи структура-функция в белках на основе сходства локальной структуры. PLoS ONE 4 (7):
e6266.https://doi.org/10.1371/journal.pone.0006266
Редактор: Джоэл Л. Суссман, Институт науки Вейцмана, Израиль
Поступила: 7 октября 2008 г .; Одобрена: 10 июня 2009 г .; Опубликовано: 15 июля 2009 г.
Это статья в открытом доступе, распространяемая в соответствии с условиями декларации Creative Commons Public Domain, которая предусматривает, что после размещения в общественном достоянии эта работа может свободно воспроизводиться, распространяться, передаваться, модифицированы, созданы на основе или иным образом использованы кем-либо в любых законных целях.
Финансирование: Эта работа была поддержана Фондом Кнута и Алисы Валленберг, Шведским фондом стратегических исследований, Шведским исследовательским советом, Шведским правительственным агентством инновационных систем (VINNOVA) и Национальными институтами здравоохранения США / Национальной библиотекой. медицины (LM007085 по KF). Эти спонсоры не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Выявление функций белков — одна из основных задач молекулярной биологии. Инструменты поиска сходства последовательностей, такие как BLAST [1], произвели революцию в биологических исследованиях, предоставив функциональные гипотезы, которые можно было проверить экспериментально. Однако идентификация функционально охарактеризованных гомологов с использованием сходства последовательностей возможна только для менее чем 50% белков, предсказанных в проектах секвенирования генома. Поскольку структура эволюционно более консервативна, чем последовательность, считается, что структурная информация обеспечивает решение для многих оставшихся белков [2], [3].Действительно, расширенная цель структурной геномики состоит в том, чтобы систематически определять структуры белков для новых семейств белков [4], использовать эти структуры в качестве шаблонов для методов предсказания структуры in silico [5], [6], а затем использовать решенные и предсказанные структуры для вывода функции [7], [8]. Однако это требует новых вычислительных методов, которые используют структуру для прогнозирования функций. Таким образом, понимание и предсказание взаимосвязей между структурой и функцией белков многие считают святым Граалем вычислительной биологии.
Подходы к анализу взаимосвязей структура-функция в белках основываются либо на глобальном сходстве (складка), либо на локальном сходстве (мотивы) [9] — [12]. Было показано, что сходство складок связано с функцией [13], [14], а также использовалось для вывода паттернов последовательностей, специфичных для функции [15]. Однако многие складки, такие как ствол TIM и складка Россмана, обнаруживаются в белках с несколькими различными функциями [2], и это привело к различным методам локальных структур-мотивов, основанных, например, на известных функциональных сайтах или функционально-специфической последовательности. узоры [16] — [21].Недавно мета-серверы получили функциональные прогнозы, позволяя большому количеству различных свидетельств (включая глобальные и локальные свойства) независимо голосовать за конкретную функцию [22] — [24].
Здесь мы предоставляем всесторонний анализ взаимосвязи структура-функция в белках, в котором библиотека повторяющихся мультифрагментных структурных мотивов, называемых локальными дескрипторами структуры белка [25], [26], используется для изучения IF-THEN правила [27], [28], которые связывают комбинации локальных субструктур со специфическими функциями белков.В отличие от предыдущих исследований, мы исследуем всех повторяющихся мотивов и всех аннотированных белков без предварительного знания функциональных сайтов или какой-либо информации о последовательностях. Таким образом, мы создаем модель-правило, которая представляет собой полное представление взаимосвязи структура-функция в белках, основанных только на структурном сходстве. Путем вычислительной оценки способности модели обобщать и предсказывать функцию невидимых белков мы предлагаем полную количественную оценку взаимосвязи структура-функция.Это позволяет нам сделать критические наблюдения о важности структуры в различных аспектах функции белка. Наши результаты можно резюмировать следующим образом: (а) почти две трети всех молекулярных функций предсказываются со статистически значимой точностью, (б) биологические процессы и клеточные компоненты значительно труднее предсказать по структуре, чем по молекулярным функциям, (в) объединение локальное сходство дает лучшую предсказательную силу, чем использование глобального сходства, в частности, для функционально универсальных складок, а также позволяет предсказывать функцию новых складок, (d) каталитическая активность предсказывается лучше, чем большинство функций, связанных со связыванием, и это связано с динамикой белка и беспорядок, и (e) предсказания на основе структуры дополняют предсказания, основанные на последовательностях, и показаны посредством литературной проверки, чтобы обеспечить множество правильных предсказаний, даже если не существует значительного сходства последовательностей.
Результаты
Библиотека аннотированных локальных субструктур белков
Локальный дескриптор структуры белка — это набор коротких непрерывных фрагментов (сегментов) основной цепи, центрированных в трех измерениях вокруг определенной аминокислоты (рис. 1A, B). Мы построили библиотеку из 4197 таких повторяющихся локальных субструктур [25] из репрезентативного набора всех экспериментально определенных доменов структуры белков в Protein Data Bank (PDB) с менее чем 40% идентичностью последовательностей друг другу [29], [30].Библиотека была использована для автоматического представления всех белковых структур с точки зрения соответствия или несоответствия каждой из локальных субструктур. Затем мы организовали аннотации генной онтологии (GO) [31], [32] всех охарактеризованных белков в 113 классов молекулярных функций, 139 классов биологических процессов и 30 классов клеточных компонентов (см. Таблицу 1 и Материалы и методы для получения подробной информации. ).
Рисунок 1. Локальная группа подструктур с центральным дескриптором 1qama_ # 37.
Называются дескрипторы: «доменное имя белка PDB» # «центральная аминокислота».А) Рисунок вторичной структуры центрального дескриптора и его структурное соответствие с десятью ближайшими дескрипторами в группе. B) Выравнивание последовательностей в результате структурного выравнивания в A. C) Расположение в онтологии генов значительно избыточно представленных (FDR контролируется на 0,05 [39]) молекулярных функций, аннотированных для 68 белков, соответствующих локальной субструктуре в A (отмечены красным) . Всего к 68 белкам были аннотированы 28 молекулярных функций. D) Правило IF (1qama_ # 37 И 1xvaa_ # 68) THEN (GO: 0008757 OR GO: 0000287), объединяющее субструктуру 1qama_ # 37 в A с субструктурой 1xvaa_ # 68 для однозначного описания 12 белков, аннотированных GO: 0008757 : Активность S-аденозилметионин-зависимой метилтрансферазы .Два из этих белков дополнительно аннотированы GO: 0000287: , связывающим ионы магния . Таким образом, правило эффективно объединяет локальные подструктуры для обращения только к одному из трех статистически значимых классов GO, связанных с 1qama_ # 37.
https://doi.org/10.1371/journal.pone.0006266.g001
Модель индукционная
Взаимосвязь между структурой и функцией была смоделирована с использованием правил IF-THEN [27], [28], где IF-часть каждого правила определяет минимальную комбинацию локальных подструктур, позволяющих отличить конкретную структуру белка от структур, аннотированных для других классов GO (рис. 1С, Г).Модель правил была индуцирована с использованием только субструктур, наблюдаемых в белковых структурах, статистически избыточно представленных по крайней мере в одном классе GO (Таблица S1). Классы GO не исключают друг друга. Например, каталитическая активность металлоэндопептидазы с участием иона цинка приведет к аннотациям молекулярных функций GO: GO: 0004222: , активность металлоэндопептидазы и GO: 0008270: , связывающая ион цинка . Кроме того, некоторые функции не полностью различимы с точки зрения структуры, потому что, например,g., функционально различающие свойства слишком редки, чтобы их можно было выделить по общим правилам. Следовательно, THEN-часть правил часто содержит несколько GO-классов с разными вероятностями (рисунок 1D). Наша модель молекулярной функции ГО включает около 20000 правил, описывающих различные перекрывающиеся структурно-функциональные отношения на разных уровнях специфичности (Таблица S2). В качестве отправной точки мы также создали правила, основанные на глобальном структурном сходстве, специфичном для предметной области, с точки зрения ориентации и связности основных вторичных структурных элементов (складка CATH, см. Материалы и методы) [33].
Количественная оценка взаимосвязи структура-функция
Мы утверждаем, что строгая оценка способности структурных моделей предсказывать функцию невидимых белков — лучший способ количественно оценить степень, в которой функция зависит от структуры. С этой целью мы оценили прогностическую эффективность моделей, используя перекрестную проверку и анализ рабочих характеристик приемника (ROC), и составили отчет о площади под кривой ROC (AUC) [34] для каждого класса молекулярных функций, биологических процессов и клеток. компонент (рисунок 2, таблица S3).
Рисунок 2. Прогнозирование характеристик модели с использованием перекрестной проверки и анализа ROC.
A) Список из десяти наиболее предсказуемых классов молекулярных функций GO, измеренных с помощью AUC и ее стандартной ошибки [34]. Мы также сообщаем о чувствительности (SENS), специфичности (SPEC) и количестве истинных положительных результатов (TP), ложных положительных результатов (FP), истинных отрицательных результатов (TN) и ложных отрицательных результатов (FN) при одном конкретном пороге принятия решения (THR). Подробности см. В разделе «Материалы и методы». (B, C и D) Производительность для всех классов GO и всех трех субонтологий GO с использованием локальных подструктур или складок CATH при разных порогах принятия решения (в результате варьирования затрат на ложные срабатывания, подробности см. В разделе «Материалы и методы»). Покрытие — это процент белков с хотя бы одним правильным предсказанием или процент аннотаций, правильно предсказанных, а точность — процент правильных предсказаний. Цифры, соответствующие порогам принятия решения в A, обведены кружком.
https://doi.org/10.1371/journal.pone.0006266.g002
Методы, основанные на локальных и глобальных структурах, лучше подходят для прогнозирования молекулярной функции, чем для прогнозирования биологического процесса и клеточного компонента (рис. 2).Это не является неожиданным, поскольку белки, разделяющие клеточное местоположение или являющиеся частью широкого биологического процесса, не должны быть структурно связаны. Это добавляет дополнительные доказательства к другим исследованиям, которые показали, что временные профили экспрессии генов необходимы для объяснения биологических процессов [35]. Следовательно, мы сосредоточим наш подробный анализ на молекулярной функции.
Для выбранного набора пороговых значений решения подход локальной субструктуры правильно предсказывает 51% аннотаций и, по крайней мере, одну аннотацию для 56% белков, при этом 37% прогнозов являются правильными (т.е., точность). Локальный подход неизменно превосходит глобальный подход (рис. 2B) из-за гибкости, связанной с объединением нескольких локальных подструктур для получения правил для конкретных функций. В частности, мы видим выраженную разницу для белков с одинаковой складкой, но с разной функцией. Например, 69% из 169 белков со складкой Россмана имели одну функцию, правильно предсказанную методом локальной субструктуры (точность = 27%), по сравнению с только 17% для CATH (точность = 9%), в то время как соответствующие числа для 50 TIM бочкообразные белки составляли 66% (точность = 21%) для локальных субструктур и 50% (точность = 12%) для CATH.Очевидно, что использование локальных субструктур увеличивает разрешение и позволяет нам функционально различать белки с одинаковой складкой.
Каталитическая активность зависит от консервативной структуры
Используя локальные субструктуры, мы получаем значительные значения AUC (т.е. AUC> 0,7) для 82 из 113 классов молекулярных функций GO. Однако не все аспекты молекулярной функции в равной степени зависят от структуры. Когда качество прогнозирования классов ГО было исследовано по отношению к группам более широких функциональных категорий, определяемых иерархической природой ГО, мы обнаружили, что 53 из 63 классов молекулярных функций ГО, расположенных под каталитической активностью ГО: 0003824: , были значительно прогнозируемыми ( Р <0.0020). С другой стороны, 15 из 37 классов под GO: 0005488: связывают (P <0,027) и все четыре класса находятся под GO: 0030528: активности регулятора транскрипции (P <0,0049), три из которых также находятся под связывание , были , а не , которые значительно предсказывались. Та же тенденция наблюдалась и в прогнозах на основе CATH. Таким образом, наши результаты показывают, что свойства, связанные со связыванием, трудно смоделировать на основе используемых представлений о структуре, в то время как каталитические механизмы, по-видимому, хорошо связаны с консервативным структурным сходством (Таблица S4).Это может быть связано с тем, что каталитическое действие ферментов не ограничивается каталитическим сайтом, но связано с внутренней динамикой белка [36]. CATH-складки и, в некоторой степени, локальные субструктуры в первую очередь описывают белковые ядра. Таким образом, они могут хорошо подходить для моделирования каталитической активности . Связывание , с другой стороны, в основном требует, чтобы белок имел поверхность с подходящими свойствами, определяемыми электростатическим, гидрофобным и ван-дер-ваальсовым взаимодействиями, и такая поверхность может быть создана с помощью альтернативных структур.Исключения из наблюдения, что связывание трудно предсказать, включают некоторые взаимодействия с ионами металлов (AUC 0,95, 0,92, 0,80, 0,75), которые часто участвуют в каталитическом механизме, и связывание GTP и ATP (AUC 0,89 и 0,77), которые играют очень важную роль в ферментативной активности.
Локальные дескрипторы, которые одновременно встречаются в правилах модели, выбираются, потому что они зависят от функции. Следовательно, интересно наблюдать, что такие сопутствующие субструктуры значительно чаще, чем случайно выбранные субструктуры (P <2.2 × 10 −16 ), образуют связанные комплексы, в которых один или несколько остатков каждой субструктуры находятся в пределах 5 Å друг от друга (подробности см. В разделе «Материалы и методы»). Недавно опубликованный контакт между участком петли и гидрофобным кластером, связанный с внутренней динамикой фермента циклофилина A (CypA), точно описывается одним из наших правил (Рисунок 3A) [36]. Мы ожидаем, что правила, которые объединяют локальные субструктуры, представляющие стабильные контактные поверхности, обнаруженные во многих белках, могут оказаться для описания общих механизмов, лежащих в основе функций белков (Figure 3B).Тот факт, что локальные субструктурные комплексы возникают из правил, которые могут предсказать функцию белка, указывает на то, что выбранный здесь подход способен обобщать и описывать функцию белка за пределами подходов, основанных на глобальном сходстве. Это также демонстрирует преимущество моделирования отношений структура-функция с использованием явных и понятных правил IF-THEN.
Рисунок 3. Правила объединения локальных подструктур в связанные комплексы.
A) Структура CypA (идентификатор PDB: 1aka).Область петли, представленная Phe 67, коррелирует с динамикой ядра, представленного гидрофобным кластером, включающим Leu 39, Phe 46, Phe 48 и Ile 78 [36]. Правило объединения локальных подструктур 1elva1 # 604 и 1bif_2 # 398 описывает именно этот механизм. 1elva1 # 604 (желтый) соответствует области цикла, включая Phe 67 (залитый желтым пространством), а 1bif_2 # 398 (синий) соответствует частям ядра, включая Leu 39 (залитый синим пространством). Перекрытие между двумя локальными подструктурами отмечено зеленым цветом.Хотя все остатки в гидрофобном кластере описаны в нашей локальной библиотеке субструктур, правило минимального IF-THEN требует только одного остатка в кластере, чтобы различать функцию. B) Две локальные субструктуры в правиле на Рисунке 1D, соответствующие ферменту Cytosine-N4-Specific (PDB id: 1boo): 1qama_ # 37 желтым, 1xvaa_ # 68 синим, перекрытие зеленым и остатки в контакте как заполненное пространство . Комбинированные локальные субструктуры имеют очень похожее количество контактирующих остатков в 12 совпадающих белках (средняя контактная поверхность включает 25% неперекрывающихся остатков в двух локальных субструктурах со стандартным отклонением 6.4).
https://doi.org/10.1371/journal.pone.0006266.g003
Белковые расстройства
Становится все более очевидным, что некоторые функции белков требуют внутреннего нарушения [37]. Используя пробелы из трех или более остатков в белках PDB, охарактеризованных с помощью рентгеновских лучей, как признак нарушения (http://www.disprot.org/), мы обнаружили значительную корреляцию между значением AUC каждого класса молекулярных функций и степень неупорядоченности белков этих классов (коэффициент корреляции −0.36, что отличается от отсутствия корреляции при P <9.9 × 10 −5 ). Кроме того, мы обнаружили, что для более широких функциональных категорий в GO, таких как каталитическая активность , и , связывание , классы GO, которые не прогнозируются в значительной степени, демонстрируют постоянно более высокую степень нарушения по сравнению с белками в классах GO, которые хорошо прогнозируются (Таблица S5) . Та же тенденция наблюдалась и при прогнозировании на основе CATH. Это указывает на то, что некоторые аспекты функции белка нарушают предположение, что последовательность определяет конкретную структуру как предварительное условие для функции, и согласуется с другими результатами, опубликованными недавно [38].Примеры включают активность димеризации GO: 0046983: (AUC = 0,69, нарушение = 9,8%), активность катионного канала GO: 0005261: (AUC = 0,42, нарушение = 8,1%) и активность коактиватора транскрипции GO: 0003713: (AUC = 0,58, беспорядок = 8,1%). Таким образом, такие функции можно правильно предсказать, только включив информацию в правила о беспорядке.
Взаимодополняемость последовательности и структуры при прогнозировании функций
Окончательная проверка прогнозов проводится экспериментально.Однако проверка in silico предлагает преимущества в том, что можно проверить гораздо большее количество гипотез и сделать статистически обоснованные выводы. Мы применили нашу модель к функционально охарактеризованным белкам, которые не были структурно решены во время индукции модели. Мы разделили этот тестовый набор на белки со слабым, но статистически значимым сходством последовательностей с обучающим набором и белки без статистически значимого сходства.
Мы предсказали молекулярную функцию 429 белковых структур (с 634 аннотациями) со слабым, но статистически значимым сходством последовательностей (идентичность последовательностей менее 40% и E-оценка менее 0.05) к обучающей выборке. Для этих белков мы смогли предсказать 45% всех аннотаций и по крайней мере одну правильную аннотацию для 53% белков с точностью 29%. Поскольку эта производительность сопоставима с оценками перекрестной проверки, полученными из обучающей выборки (рисунок 2), мы можем сделать вывод, что правила, основанные на библиотеке локальных подструктур, хорошо обобщаются на невидимые структуры по всему континууму сходства последовательностей. Комбинируя прогнозы на основе подхода локального дескриптора с прогнозами, полученными из аннотаций ближайшего соседа по последовательности в обучающем наборе (обнаруженного с помощью PSI-BLAST [1], подробности см. В материалах и методах), мы могли правильно предсказать 70% все аннотации и хотя бы одна правильная аннотация для 76% белков с точностью 30%.Из всех 444 правильных прогнозов 398 были сделаны с помощью PSI-BLAST (62%), а остальные 46 были сделаны исключительно методом на основе дескрипторов. Таким образом, подход, сочетающий PSI-BLAST и наш метод, основанный на структуре, правильно предсказывает больше аннотаций, чем использование только PSI-BLAST, даже если существует сходство последовательностей.
Мы, наконец, поставили перед системой задачу предсказать функцию для 167 невидимых белков (с 224 аннотациями) без существенного сходства последовательностей с обучающей выборкой (E-оценка больше 0.05). Для этих довольно требовательных целей метод локального дескриптора обеспечил охват и точность только около 10%, показывая, что модель не является независимой от последовательности, даже если она основана исключительно на структуре. Однако автоматические аннотации составляют 92,4% базы данных, и, как известно, эти аннотации являются неполными. Следовательно, мы вручную проверили все прогнозы, сделанные дескрипторным подходом для этих 167 целей. Этот анализ показал, что из 190 предсказаний, сделанных для 93 белков, 91 предсказание, сделанное для 57 белков, нашло некоторую поддержку в научной литературе (Таблица S6).Одним из примеров является протеин аланил-тРНК синтетаза (PDB id. 1riq) с четырьмя предсказаниями: GO: 0000049: связывание тРНК , GO: 0000287: связывание с ионом магния , GO: 0005524: связывание с АТФ и GO: 0004812 : Активность тРНК-лигазы . Были аннотированы только два последних прогноза. Однако все они были подтверждены литературным поиском как правильные. Более того, складка этого белка не была представлена в обучающей выборке, и, следовательно, этот белок нельзя было правильно предсказать, используя глобальное структурное сходство.Тот факт, что метод локальной субструктуры является полностью автоматическим, также является преимуществом по сравнению с методами, основанными на ручном назначении, поскольку можно делать прогнозы для недавно решенных целей структурной геномики. Только 58 из 167 недавно решенных структур, обсуждаемых здесь, до сих пор были отнесены к CATH. Наконец, хотя структурное сходство при отсутствии сходства последовательностей может быть результатом конвергентной эволюции, наши результаты ясно показывают, что локальные субструктуры могут обеспечивать полезные функциональные гипотезы даже для этих белков.
Обсуждение
Поскольку функция белка по своей природе зависит от глобальной архитектуры, внутренней динамики складок и тонких поверхностных свойств, которые придают специфичность связывания, мы ожидаем, что вычислительные методы, которые включают информацию на всех уровнях, будут превосходить методы, основанные исключительно на последовательностях. . Здесь мы представляем общий подход для представления функции белка с точки зрения локального и глобального структурного сходства и для количественной оценки взаимосвязи структура-функция.Это сильно отличается от любой ранее опубликованной работы с точки зрения полноты, использования многофрагментных локальных подструктур и того факта, что связь с функцией была исследована без использования информации о функциональных сайтах или паттернах последовательностей. Правила легко интерпретировать и позволяют включать в модель различные типы данных. В будущем мы хотели бы объединить этот подход в мета-метод, в который также будет включена информация о глобальном сходстве структуры и последовательности.
Были высказаны опасения, помогут ли предсказанные структуры в предсказании функций, поскольку эти методы in silico в основном правильно предсказывают ядро белка, в то время как функция часто зависит от свойств поверхности [3]. Однако наши результаты показывают, что локальные субструктуры, в основном связанные с ядром, сильно связаны с некоторыми аспектами молекулярных функций и, в частности, с каталитической активностью . Наши оценки показывают, что чисто предсказания на основе структуры служат дополнением к предсказаниям, полученным на основе последовательности, и что правильное предсказание также может быть обеспечено, когда сходства последовательностей не существует.Таким образом, мы оказали существенную поддержку жизнеспособности цели структурной геномики, то есть сокращения количества функционально не охарактеризованных белков посредством определения структуры и прогнозирования функций.
Материалы и методы
На рисунке 4 схематично показан наш метод прогнозирования функций. Подробности приведены в этом разделе.
Рисунок 4. Обзор метода прогнозирования функции.
Библиотека локальных дескрипторов структуры белка построена из репрезентативного подмножества PDB (т.е.е. Обучающий набор). Библиотека используется для представления белковых структур, а модель, которая различает классы аннотаций генной онтологии, индуцируется с использованием комбинаций локальных подструктур. Модель оценивается как внутри, так и на внешнем тестовом наборе.
https://doi.org/10.1371/journal.pone.0006266.g004
Библиотека локальных субструктур белков
Локальный дескриптор структуры белка — это набор коротких фрагментов основной цепи, центрированных в трех измерениях вокруг определенной аминокислоты [25], [26].Локальный дескриптор создается путем а) идентификации всех близких аминокислот в радиусе 6,5 Å (аминокислота представлена точкой на векторе [C α , C β ], который находится на расстоянии 2,5 Å от C . α ), б) для каждой близкой аминокислоты, добавление четырех соседей последовательности, по два с каждой стороны, чтобы получить непрерывные фрагменты основной цепи из пяти аминокислот, и в) объединение любых перекрывающихся фрагментов в сегменты. Мы вычислили локальные дескрипторы из всех аминокислот в репрезентативном наборе белковых доменов из PDB с менее чем 40% идентичностью последовательностей друг другу (ASTRAL версия 1.57 [30]). В результате получено 374 558 дескрипторов из 4006 доменов. Затем мы создали библиотеку часто встречающихся локальных дескрипторов: а) для каждого локального дескриптора, идентифицируя группу структурно подобных локальных дескрипторов, и б) выбирая набор из 4197 репрезентативных, частично перекрывающихся групп дескрипторов . Мы рассматривали только группы с минимум семью дескрипторами с минимум тремя неперекрывающимися фрагментами последовательности.
Белки в этом исследовании были представлены в виде цепочек из нулей и единиц, указывающих, соответствует ли структура белка соответствующим локальным субструктурам или нет.Это было сделано для белков с доменами в ASTRAL 1.57 (т.е. обучающего набора), а также для белков в ASTRAL 1.67 с менее чем 40% идентичностью последовательностей обучающему набору (т.е. внешнему набору тестов).
Аннотации генных онтологий
GO — это не зависящий от организма управляемый словарь для описания клеточной роли генов и генных продуктов с точки зрения молекулярных функций (т. Е. Задач, выполняемых отдельными генными продуктами), биологических процессов (т. Е. Широких биологических целей, достигаемых упорядоченной сборкой молекулярных продуктов). функции) и клеточные компоненты (т.е. места, где активны генные продукты).
Мы получили аннотации с домашней страницы GO (http://www.geneontology.org) [32] для белков, используемых для построения локальной библиотеки субструктур, описанной ранее (2878 белков с 4006 белковыми доменами в ASTRAL). Мы распределили эти аннотации (вверх) в графе GO (версия 1.419) и отбросили все термины (узлы) GO, используемые для аннотирования менее десяти белков. Затем мы выбрали среди оставшихся терминов наиболее конкретные термины в качестве наших учебных классов (Таблица 1, Таблица S3).При рассмотрении только терминов GO, используемых для аннотирования по крайней мере десяти белков, некоторые аннотации были потеряны. Однако большинство белков сохраняло по крайней мере одну аннотацию, что указывает на то, что существует набор больших классов, обеспечивающих по крайней мере одну аннотацию почти для каждого белка, и что дополнительные аннотации часто взяты из менее заполненных терминов GO. Кроме того, выбор конкретных классов в качестве учебных классов привел к потере некоторых общих аннотаций.
Значимые классы GO в группах дескрипторов
Мы использовали гипергеометрическое распределение для расчета p-значений, отражающих, в какой степени белки, аннотированные для определенного класса GO, были избыточно представлены в группе дескрипторов.Затем мы использовали коэффициент ложного обнаружения (FDR) [39], контролируемый на уровне 0,05, для определения статистически значимых локальных дескрипторов. FDR — это метод корректировки нескольких гипотез при статистической проверке гипотез.
В библиотеке 84% дескрипторных групп имели значительную избыточную представленность белков, аннотированных, по крайней мере, к одному из 113 классов молекулярных функций (FDR контролируется на уровне 0,05). Соответствующие числа для 139 биологических процессов и 30 клеточных компонентов составили 77% и 29% соответственно.Все классы GO для всех трех частей GO были значительно перепредставлены по крайней мере в одной группе дескрипторов (за исключением клеточного компонента GO: 0005938: клеточной коры ). См. Подробности в таблице S1.
Основополагающим принципом машинного обучения является то, что более высокое соотношение примеров к функциям создает модели, которые лучше работают в невидимых случаях (при той же разделимости классов). Чтобы справиться с большим количеством структурных особенностей (т.е. 4197 локальных субструктур) по сравнению с количеством белков (2815 для молекулярной функции) в этом исследовании, мы использовали только значимые группы дескрипторов FDR для создания моделей правил.
CATH
CATH [33] (версия 2.6.0) — это классификационное дерево, которое классифицирует доменные структуры с возрастающей специфичностью в соответствии с классом (C), архитектурой (A), топологией (T) и гомологичным суперсемейством (H). Класс присваивается в соответствии с составом вторичной структуры и упаковкой области структуры. В 90% случаев это делается автоматически. Архитектура относится к общей форме доменной структуры с точки зрения относительной ориентации элементов вторичной структуры.Архитектура назначается вручную. Топология относится к связности элементов вторичной структуры в аналогичных архитектурах, и назначение выполняется автоматически. Наконец, гомологичное суперсемейство относится к белкам, которые являются гомологами, как определено по сходству последовательностей. Эти назначения также выполняются вручную.
Локальные дескрипторы классифицируются по группам в соответствии с относительным расположением и ориентацией их сегментов. Следовательно, это соответствует архитектуре в CATH.Однако архитектура CATH слишком общая для прогнозирования функций (результаты не показаны). Более того, гомологичное суперсемейство CATH внесет в анализ сходство последовательностей и, следовательно, будет скрывать чистый сигнал структура-функция. Следовательно, в этой статье мы предпочитаем использовать топологию CATH (то есть свертку), а не гомологичное суперсемейство CATH или другие, более вручную выведенные базы данных [11].
Прогнозы на основе последовательностей
Программа сравнения последовательностей PSI-BLAST [1] использовалась для получения предсказаний на основе последовательностей.Каждый домен во внешнем тестовом наборе сравнивался с обучающим набором с профилем последовательности, полученным с использованием базы данных неизбыточных последовательностей NCBI (ftp://ftp.ncbi.nlm.nih.gov/blast/db/nr) (PSI -BLAST был запущен с тремя итерациями и порогом E-value 0,005 для включения последовательностей в модель). Аннотации для наиболее близкого совпадения в обучающем наборе (определяемого значением E) использовались в качестве прогнозов, а прогнозы для белка были приняты как прогнозы для всех его доменов.
Контакты между локальными подструктурами
Для каждого правила и каждого подходящего белка мы вычислили среднюю долю остатков в парах локальной субструктуры, которые были в контакте (остатки, общие для обеих локальных субструктур, не рассматривались). Мы определили, что два остатка должны контактировать, если кратчайшее расстояние между атомами в этих аминокислотах было менее 5 Å. Этот порог основан на расстоянии гидрофобного контакта между лигандами и белками [40]. Следовательно, для каждого правила мы получили среднее количество контактов между парами субструктур в совпадающих белках и стандартное отклонение, указывающее на стабильность этих контактных поверхностей по отношению к различным белкам.Средняя контактная поверхность пары локальных субструктур в правилах охватывала 11% неперекрывающихся остатков в этой паре со средним стандартным отклонением 9,3. Для сравнения мы случайным образом отобрали 1000 пар локальных субструктур, соответствующих по крайней мере двум белкам, и где субструктуры встречались по крайней мере в одном из правил. Контактные поверхности для правил, зависящих от функции, были значительно больше, чем для этих случайно выбранных пар (при P <2,2 × 10 −16 с использованием критерия Колмогорова-Смирнова), в то время как стандартные отклонения были значительно меньше (также при P <2.2 × 10 −16 ). Некоторые большие контактные конформации были особенно стабильными; 8,5% правил были связаны со средней контактной поверхностью, которая включала более 20% остатков и , где стандартное отклонение было менее 5%. Это было верно только для 2,8% случайно выбранных пар локальных субструктур.
Обучение правилам
Грубая теория множеств [27], [28] представляет собой математическую основу для вывода правил из примеров. Мы использовали этот фреймворк, реализованный в системе приблизительных наборов ROSETTA [28] (http: // rosetta.lcb.uu.se), для изучения правил IF-THEN, связывающих комбинации локальных подструктур белков с определенными классами GO. Эта структура ранее использовалась для изучения биологического процесса ГО по временным профилям экспрессии генов [35], [41] (см. Hvidsten et al. (2003) для более теоретической / математической обработки метода обучения правилам).
В принципе, метод находит минимальные наборы локальных субструктур, которые отличают конкретный белок от всех других белков, аннотированных для другого класса GO.Затем из каждого такого набора конструируется одно правило, так что IF-часть представляет собой комбинацию этих локальных подструктур, а THEN-часть — все классы GO, используемые для аннотирования белков, соответствующих IF-части. Если правило включает несколько классов GO, это означает, что соответствующий белок аннотирован классом GO, который не может быть однозначно определен из данных локальной субструктуры (то есть класс называется или ). В этом исследовании мы использовали генетический алгоритм, чтобы найти приблизительные минимальные наборы, которые отличают каждый белок от достаточно большой фракции (не менее 90%) белков из других классов GO.Правила из таких приближенных решений с меньшей вероятностью переоценивают данные и справляются с шумом лучше, чем точные решения.
Мы сравнили подход с использованием правил, основанных на минимальных, различающих подмножествах локальных подструктур, с подходом, использующим все правила на основе одной единственной локальной подструктуры. Такие очень простые правила принятия решений, называемые правилами 1R, были предложены Холте [42]. Используя этот подход, мы обнаружили, что комбинации важны для подхода локальных дескрипторов, но не помогают при использовании сверток CATH.
Прогнозирование и оценка
Мы проверили обобщающую способность нашего подхода к правилам, используя десятикратную перекрестную проверку. Набор белков был случайным образом разделен на десять подмножеств одинакового размера. Классификатор правил был вызван из девяти подмножеств (обучающий набор) и использовался для классификации белков в оставшемся подмножестве (тестовый набор). Эту процедуру повторяли десять раз, так что каждый белок был один раз в тестовой выборке, а в обучающей — девять раз.
Белок был классифицирован, позволив каждому правилу сопоставления отдать голоса классам GO, указанным этим правилом.Количество голосов, отданных каждым правилом каждому классу, соответствовало количеству белков в обучающем наборе из этого класса, которые соответствовали этому правилу (то есть поддерживаемому правилу). Затем для каждого класса рассчитывалось значение p на основе голосов с использованием гипергеометрического распределения. Эти p-значения были получены во время перекрестной проверки, и для каждого класса была рассчитана ROC-кривая, построенная на графике зависимости чувствительности от специфичности для всех возможных пороговых p-значений. Чувствительность — TP / (TP + FN), а специфичность — TN / (TN + FP), где TP — истинно положительные, FP — ложноположительные, TN — истинно отрицательные, а FN — ложно отрицательные.Кривая ROC оценивает работу классификатора, не зависящую от пороговых значений. Мы указали площадь под кривой ROC (AUC) как меру производительности. Это значение находится в диапазоне от 0 до 1, где 1 означает идеальную дискриминацию, а 0,5 означает отсутствие дискриминирующей способности. При прогнозировании фактических функций мы использовали пороговые значения p из кривых ROC, соответствующие точкам максимальной чувствительности плюс специфичность (специфичность всегда была больше 0,90, чтобы контролировать количество ложных срабатываний из-за большого количества классов).
Случайным образом перемешивая аннотации молекулярных функций, мы показали, что значения AUC перекрестной проверки, равные или превышающие 0,7, вряд ли будут получены случайно (P <0,01). Таким образом, в этом исследовании AUC ≥0,7 была обозначена как статистически значимая.
Дополнительная информация
Таблица S1.
Все пары значимых локальных дескрипторов FDR и классов GO. (a) Молекулярная функция: все пары классов значимый локальный дескриптор FDR-GO. (b) Биологический процесс: все пары классов значимый локальный дескриптор FDR-GO.(c) Сотовый компонент: все пары классов значимый локальный дескриптор FDR-GO. ПАРАМЕТРЫ относятся к параметрам в гипергеометрическом распределении, используемом для вычисления p-значений: N, n, k, x, где N — количество пар класса белок-GO в наборе данных, n — количество белков, совпадающих с локальный дескриптор, k — количество белков в классе GO, а x — количество белков, соответствующих локальному дескриптору и в классе GO.
https://doi.org/10.1371/journal.pone.0006266.s001
(1,28 МБ PDF)
Таблица S2.
Все индуцированные правила молекулярной функции. Для каждого класса молекулярных функций Gene Ontology в THEN-части значение p дается вместе с параметрами гипергеометрического распределения, используемыми для вычисления значений p: N, n, k, x, где N = 2725 — число пар белок-класс GO в наборе данных, n — количество белков, соответствующих IF-части правила, k — количество белков в классе GO, а x — количество белков, соответствующих правилу и в GO класс.
https://doi.org/10.1371/journal.pone.0006266.s002
(1,16 МБ PDF)
Таблица S3.
Прогнозирование производительности. (а) Прогнозирование производительности. 10-кратная перекрестная проверка оценок AUC для всех классов молекулярных функций (б) Прогнозирование. Оценки AUC с 10-кратной перекрестной проверкой для всех классов биологических процессов (c) Прогнозирование. 10-кратная перекрестная проверка оценок AUC для всех классов сотовых компонентов
https://doi.org/10.1371/journal.pone.0006266.s003
(0,04 МБ PDF)
Таблица S4.
Избыточное количество классов GO со значительными значениями AUC. (а) Местные подструктуры. Избыточное представительство классов GO со значительными значениями AUC (AUC> = 0,7) и незначительными значениями (AUC <0,7). P-значения рассчитываются на основе количества белков и количества классов GO в каждом из более общих терминов GO. (б) CATH складывается. Избыточное представительство классов GO со значительными значениями AUC (AUC> = 0.7) и незначимые значения (AUC <0,7). P-значения рассчитываются на основе количества белков и количества классов GO в каждом из более общих терминов GO.
https://doi.org/10.1371/journal.pone.0006266.s004
(0,01 МБ PDF)
Таблица S5.
Белковая болезнь. (а) Местные подструктуры. Белковое расстройство. Среднее нарушение на верхнем уровне онтологии генов и корреляция между прогностической эффективностью с точки зрения перекрестной проверки AUC и нарушения белка.(б) CATH складывается. Белковое расстройство. Среднее нарушение на верхнем уровне онтологии генов и корреляция между прогностической эффективностью с точки зрения перекрестной проверки AUC и нарушения белка.
https://doi.org/10.1371/journal.pone.0006266.s005
(0,03 МБ PDF)
Вклад авторов
Задумал и спроектировал эксперименты: TRH. Проведены эксперименты: TRH. Проанализированы данные: TRH AL AK GA KF. Предоставленные реагенты / материалы / инструменты анализа: TRH AK KF JK.Написал статью: TRH AL AK GA KF JK.
Ссылки
- 1.
Альтшул С.Ф., Мэдден Т.Л., Шаффер А.А., Чжан Дж., Чжан З. и др. (1997) Gapped BLAST и PSI-BLAST: новое поколение программ поиска в базе данных белков. Nucleic Acids Res 25: 3389–3402. - 2.
Киношита К., Накамура Х (2003) Белковая информатика для идентификации функций. Curr Opin Struct Biol 13: 396–400. - 3.
Сколник Дж., Фетроу Дж. С. (2000) От генов до структуры и функции белков: новые применения вычислительных подходов в эпоху генома.Trends Biotechnol 18: 34–39. - 4.
Чандония Дж. М., Бреннер С. Е. (2006) Влияние структурной геномики: ожидания и результаты. Наука 311: 347–351. - 5.
Бейкер Д., Сали А. (2001) Прогнозирование структуры белка и структурная геномика. Наука 294: 93–96. - 6.
Копп Дж., Бордоли Л., Бэтти Дж. Н., Кифер Ф., Шведе Т. (2007) Оценка прогнозов CASP7 для целей моделирования на основе шаблонов. Белки 69: Suppl 838–56. - 7.Чжан С., Ким С.Х. (2003) Обзор структурной геномики: от структуры к функции. Curr Opin Chem Biol 7: 28–32.
- 8.
Мурзин А.Г., Патти Л. (1999) Последовательности и топология: от последовательности к структуре к функции. Curr Opin Struct Biol 9: 359–362. - 9.
Orengo CA, Todd AE, Thornton JM (1999) От структуры белка к функции. Curr Opin Struct Biol 9: 374–382. - 10.
Thornton JM, Todd AE, Milburn D, Borkakoti N, Orengo CA (2000) От структуры к функции: подходы и ограничения.Nat Struct Biol 7: Suppl991–994. - 11.
Ouzounis CA, Coulson RM, Enright AJ, Kunin V, Pereira-Leal JB (2003) Схемы классификации структуры и функции белка. Нат Рев Женет 4: 508–519. - 12.
Ли Д., Редферн О., Оренго С. (2007) Предсказание функции белка по последовательности и структуре. Nat Rev Mol Cell Biol 8: 995–1005. - 13.
Шахнович Б.Е., Дохолян Н.В., ДеЛиси С., Шахнович Е.И. (2003) Функциональные отпечатки пальцев складок: свидетельство коррелированной структурно-функциональной эволюции.J Mol Biol 326: 1–9. - 14.
Hegyi H, Gerstein M (1999) Взаимосвязь между структурой и функцией белка: всесторонний обзор с применением к геному дрожжей. J Mol Biol 288: 147–164. - 15.
Пазос Ф., Штернберг М.Дж. (2004) Автоматизированное предсказание функции белка и обнаружение функциональных сайтов по структуре. Proc Natl Acad Sci U S A 101: 14754–14759. - 16.
Ди Дженнаро Дж. А., Сью Н., Хоффман Б. Т., Чжан Л., Сколник Дж. И др.(2001) Улучшенная функциональная аннотация белковых последовательностей за счет использования структурных дескрипторов. J Struct Biol 134: 232–245. - 17.
Kasuya A, Thornton JM (1999) Анализ трехмерной структуры паттернов PROSITE. J Mol Biol 286: 1673–1691. - 18.
Jonassen I, Eidhammer I, Taylor WR (1999) Открытие локальных мотивов упаковки в белковых структурах. Белки 34: 206–219. - 19.
Russell RB (1998) Обнаружение трехмерных паттернов боковых цепей белков: новые примеры конвергентной эволюции.J Mol Biol 279: 1211–1227. - 20.
Ferre F, Ausiello G, Zanzoni A, Helmer-Citterich M (2004) SURFACE: база данных участков поверхности белков для функциональной аннотации. Nucleic Acids Res 32: D240–244. - 21.
Polacco BJ, Babbitt PC (2006) Автоматическое обнаружение трехмерных мотивов для аннотации функции белка. Биоинформатика 22: 723–730. - 22.
Ласковски Р.А., Уотсон Дж. Д., Торнтон Дж. М. (2005) ProFunc: сервер для прогнозирования функции белка по трехмерной структуре.Nucleic Acids Res 33: W89–93. - 23.
Pal D, Eisenberg D (2005) Вывод функции белка из структуры белка. Структура 13: 121–130. - 24.
Уотсон Дж. Д., Сандерсон С., Езерский А., Савченко А., Эдвардс А. и др. (2007) К полностью автоматизированному предсказанию функций на основе структуры в структурной геномике: тематическое исследование. J Mol Biol 367: 1511–1522. - 25.
Hvidsten TR, Kryshtafovych A, Fidelis K (2009) Локальные дескрипторы структуры белка: систематический анализ взаимосвязи структура-последовательность в белках с использованием коротко- и дальнодействующих взаимодействий.Белки 75: 870–884. - 26.
Hvidsten TR, Kryshtafovych A, Komorowski J, Fidelis K (2003) Новый подход к распознаванию складок с использованием свойств, полученных из последовательностей из наборов структурно подобных локальных фрагментов белков. Биоинформатика 19: Дополнение 2II81 – II91. - 27.
Pawlak Z (1991) Грубые множества: теоретические аспекты рассуждений о данных. Библиотека теории и решений. Серия D, теория систем, инженерия знаний и решение проблем. Дордрехт; Бостон: Kluwer Academic Publishers. - 28.
Коморовски Дж., Эрн А., Сковрон А. (2002) Программная система ROSETTA Rough Set. В: Klösgen W, Zytkow J, редакторы. Справочник по интеллектуальному анализу данных и открытию знаний. Издательство Оксфордского университета. С. 554–559. - 29.
Берман Х.М., Вестбрук Дж., Фенг З., Гиллиланд Дж., Бхат Т.Н. и др. (2000) Банк данных о белках. Nucleic Acids Res 28: 235–242. - 30.
Brenner SE, Koehl P, Levitt M (2000) Сборник ASTRAL для анализа структуры и последовательности белка.Nucleic Acids Res 28: 254–256. - 31.
Эшбернер М., Болл К.А., Блейк Дж. А., Ботштейн Д., Батлер Н. и др. (2000) Генная онтология: инструмент для объединения биологии. Консорциум генных онтологий. Нат Генет 25: 25–29. - 32.
Камон Э., Магран М., Баррелл Д., Биннс Д., Флейшманн В. и др. (2003) Проект Gene Ontology Annotation (GOA): реализация GO в SWISS-PROT, TrEMBL и InterPro. Genome Res 13: 662–672. - 33.
Orengo CA, Michie AD, Jones S, Jones DT, Swindells MB, et al.(1997) CATH — иерархическая классификация структур белковых доменов. Структура 5: 1093–1108. - 34.
Hanley JA, McNeil BJ (1982) Значение и использование площади под кривой рабочей характеристики приемника (ROC). Радиология 143: 29–36. - 35.
Lægreid A, Hvidsten TR, Midelfart H, Komorowski J, Sandvik AK (2003) Прогнозирование биологического процесса онтологии генов на основе временных паттернов экспрессии генов. Genome Res 13: 965–979. - 36.
Eisenmesser EZ, Millet O, Labeikovsky W, Korzhnev DM, Wolf-Watz M, et al.(2005) Внутренняя динамика фермента лежит в основе катализа. Природа 438: 117–121. - 37.
Дункер А.К., Браун С.Дж., Лоусон Дж.Д., Якучева Л.М., Обрадович З. (2002) Внутреннее расстройство и функция белка. Биохимия 41: 6573–6582. - 38.
Lobley A, Swindells MB, Orengo CA, Jones DT (2007) Определение функции с использованием паттернов нативного нарушения в белках. PLoS Comput Biol 3: e162. - 39.
Бенджамини Ю., Хохберг Ю. (1995) Контроль над показателем ложного обнаружения: практический и мощный подход к множественному тестированию.Журнал Королевского статистического общества B 57: 289–300. - 40.
Рэри М., Крамер Б., Ленгауэр Т. (1999) Докинг гидрофобных лигандов с помощью алгоритмов сопоставления на основе взаимодействия. Биоинформатика 15: 243–250. - 41.
Hvidsten TR, Lægreid A, Komorowski J (2003) Изучение основанных на правилах моделей биологического процесса на основе временных профилей экспрессии генов с использованием генной онтологии. Биоинформатика 19: 1116–1123. - 42.
Holte RC (1993) Очень простые правила классификации хорошо работают с наиболее часто используемыми наборами данных.Машинное обучение 11: 63–91.
Proteins I: Структура и функции — Видео и стенограмма урока
Нам нужны наши белки не только как основная группа продуктов питания, но и для многих полезных ролей, которые они играют в нашем организме. В нашем вводном уроке, посвященном белкам, вы узнаете о многих функциях, для выполнения которых мы полагаемся.
Белки и пептидные связи
Мы слышали о белках, потому что они являются важной частью нашего рациона.Если мы посмотрим на этикетку с пищевой добавкой для макарон с сыром, то увидим, что они содержат не только несколько граммов углеводов и жиров, но и белки. Белки — это большие биологические молекулы, состоящие из аминокислот, соединенных пептидными связями.
Для образования белков многие аминокислоты объединяются путем дегидратации с образованием пептидных связей , которые представляют собой связь между карбонильной группой аминокислоты и азотом аминогруппы другой аминокислоты.Пептидные связи невероятно сильны и помогают белкам оставаться вместе, чтобы выполнять свою работу.
Белки как структура
Белки могут использоваться для создания структуры. Одним из примеров является белок под названием кератин . Кератин — это белок, который содержится в наших волосах, коже и ногтях.
Кератиновые белки объединяются и образуют нити, которые представляют собой длинные переплетенные нити. Если вы думаете о толстой веревке, которая состоит из множества разных меньших кусков веревки, она становится сильнее благодаря тому факту, что все эти разные части сплетены вместе, и это как раз случай с кератином.Кератин гидрофобен и не растворяется в воде; этот страх перед водой — одна из причин, почему кератин так силен. Различные нити кератиновых белков очень притягиваются друг к другу, а не к внешней среде, что помогает им оставаться вместе. Другой важный структурный белок — коллаген . Это важный компонент соединительной ткани, которая находится в сухожилиях и связках. Опять же, это очень важно для структуры, которую мы находим в наших телах.
Ферменты
Другая важная роль, которую могут играть белки, заключается в том, что они действуют как ферментов , которые могут вызывать химические реакции в нашем организме.
Способ, которым они вызывают химические реакции, состоит в том, что они уменьшают активационный барьер, чтобы реакция произошла. Итак, чтобы понять, что я имею в виду под активационным барьером, допустим, ваш будильник срабатывает, но вы действительно не хотите вставать с постели. За пределами кровати действительно холодно, и это увеличивает активационный барьер, мешающий вам встать с постели, или количество «умф», чтобы встать с постели.
В случае химических реакций мы говорим о количестве энергии, необходимом для того, чтобы эта химическая реакция произошла.Уменьшая активационный барьер, ферменты повышают вероятность возникновения химической реакции. Это было бы похоже на то, как если бы кто-то заварил вам хорошую чашку горячего кофе или, возможно, положил хороший теплый коврик, когда вы встаете с постели. Может быть, это уменьшит ваше сопротивление вставанию с постели.
Теперь ферменты могут вызывать химические реакции, которые действительно важны для многих областей нашего тела. Один пример вне нашего тела, о котором вы, возможно, слышали, называется лактаид. Эта таблетка содержит ферменты, которые помогают людям переваривать лактозный сахар.Это действительно важно для некоторых людей, которые не могут переваривать лактозу, поскольку это спасает их от несварения желудка. Расщепляя этот лактозный сахар, эти ферменты помогают желудку людей чувствовать себя лучше.
Другие функции белков
Другая важная функция белков заключается в том, что они позволяют двигаться. Двумя наиболее важными белками, которые участвуют в движении, являются актин и миозин .Актин играет важную роль в подвижности и подвижности клеток. Это также важно для сокращения наших мышц, чтобы мы могли двигаться или ходить. Миозины — я называю миозины, потому что существует множество различных версий этого белка — являются моторными белками и также очень важны для сокращения наших мышц.
Белки также могут перемещать вещи по нашему телу. Примером этого является гемоглобин , который представляет собой белок, который переносит кислород из наших легких в ткани по всему нашему телу.
Липопротеины — это белки, которые могут транспортировать множество молекул жира по нашей крови.
Белки также могут действовать как посланники, передавая сообщения от одной части нашего тела к другой. Одним из примеров этого является инсулин , который представляет собой белок, способный снизить уровень сахара в крови. Когда он высвобождается из нашей поджелудочной железы другими сигналами, он посылает сигналы нашим клеткам, чтобы они потребляли больше глюкозы, что снижает уровень сахара в крови. Гормон роста человека — еще один белок, который может действовать как посредник.После того, как он высвобождается из гипофиза, он может путешествовать по нашему телу, передавая сигналы для роста или воспроизведения другим клеткам.
Еще одна роль, которую могут играть белки, заключается в том, что они могут защищать нас. Антитела — это белки, которые могут распознавать чужеродные молекулы в нашем организме и прикрепляться к ним. Будь то вирус простуды, который у нас уже был раньше и который распознают антитела, или пыльца, на которую у нас аллергия, и антитела пытаются атаковать это, антитела — это белки, которые пытаются нас защитить.
Наконец, белки действительно важны, потому что они способны регулировать гены и другие белки. Иногда они делают это, разрезая другие белки и активируя их, или это могут быть факторы транскрипции , , которые связывают ДНК и решают, можно ли их включить или выключить.
Краткое содержание урока
Белки — это большие биологические молекулы, которые состоят из аминокислот и соединены пептидными связями.
Пептидные связи — это связи между карбонильным углеродом одной аминокислоты и аминным азотом другой аминокислоты.
Белки важны, потому что они могут накапливать энергию, обеспечивать структуру, действовать как ферменты, вызывая реакции, обеспечивать движение, транспортировать вещи, действовать как посланники, защищать нас от захватчиков и регулировать гены и другие белки.
Структурная функция белка | Протокол
4.15: Структурная функция белка
Структурные белки — это категория белков, отвечающих за различные функции, от формы и движения клеток до поддержки основных структур, таких как кости, хрящи, волосы и мышцы.В эту группу входят такие белки, как коллаген, актин, миозин и кератин.
Коллаген, самый распространенный белок у млекопитающих, встречается по всему телу. В соединительной ткани, такой как кожа, связки и сухожилия, он обеспечивает прочность на разрыв и эластичность. В костях и зубах он минерализуется с образованием твердых тканей и способствует их несущей способности. Помимо структурной поддержки, коллаген также может взаимодействовать с рецепторами клеточной поверхности и другими промежуточными молекулами для регулирования клеточных процессов, таких как рост и миграция, которые включают изменения формы клеток и тканей.
Структурные белки составляют основу цитоскелета клетки. Цитоскелет состоит из трех типов нитей, микрофиламентов, промежуточных нитей и микротрубочек, каждый из которых состоит из различных структурных белков. Микрофиламент образуется, когда актин самополимеризуется в длинные повторяющиеся структуры. Эти актиновые филаменты вносят вклад в форму и организацию клеток; Кроме того, микрофиламенты также могут способствовать перемещению и делению клеток, когда они действуют вместе с миозином.Состав промежуточных волокон варьируется в зависимости от типа клетки. Существует около 70 различных генов, кодирующих различные промежуточные филаменты. Промежуточные филаменты в эпителиальных клетках содержат кератин, периферические нейроны содержат периферин, а саркомеры в мышечных клетках содержат десмин. Основная структурная функция этих нитей — укреплять клетки и организовывать их в ткани. Микротрубочки состоят из структурных белков, называемых тубулинами. Тубулины самоорганизуются с образованием микротрубочек, которые способствуют организации цитоплазмы, включая расположение органелл.Микротрубочки также необходимы для митоза и деления клеток.
Поскольку структурные белки широко распространены, мутация в гене, кодирующем любой из этих белков, может иметь серьезные пагубные последствия. Например, мутация в гене, кодирующем коллаген, может привести к состоянию, известному как несовершенный остеогенез, которое характеризуется слабостью костей и деформациями соединительных тканей. Различные мутации в гене коллагена могут привести к синдрому Альпорта, который характеризуется проблемами в таких органах, как почки, глаза и уши.
Рекомендуемая литература
- Лодиш, Х., Берк, А., Зипурски, С. Л., Мацудаира, П., Балтимор, Д., и Дарнелл, Дж. (2000). Молекулярная клеточная биология 4-е издание. Национальный центр биотехнологической информации, книжная полка .
- Ricard-Blum, S. (2011). Семейство коллагена. Биологические перспективы Колд-Спринг-Харбор , 3 (1), a004978.
- Домингес, Р., и Холмс, К.С. (2011). Структура и функции актина. Годовой обзор биофизики , 40 , 169-186.
- Даунинг, К. Х., & Ногалес, Э. (1998). Строение тубулина и микротрубочек. Текущее мнение в области клеточной биологии , 10 (1), 16-22.
- Гейслер Ф. и Лейбе Р. Э. (2016). Эпителиальные промежуточные нити: защитники от микробной инфекции ?. Ячейки , 5 (3), 29. doi: 10.3390 / Cell5030029
Строение и функция белковых комплексов митохондриальной мембраны | BMC Biology
Риццуто Р., Де Стефани Д., Рафаэлло А., Маммукари С. Митохондрии как сенсоры и регуляторы передачи сигналов кальция. Nat Rev Mol Cell Biol. 2012; 13 (9): 566–78.
CAS
Статья
PubMed
Google Scholar
Пеллегрино М.В., Хейнс СМ. Митофагия и митохондриальный развернутый белковый ответ при нейродегенерации и бактериальной инфекции. BMC Biol. 2015; 13:22.
PubMed Central
Статья
PubMed
Google Scholar
Chandel NS. Митохондрии как сигнальные органеллы. BMC Biol. 2014; 12:34.
PubMed Central
Статья
PubMed
Google Scholar
Bratic A, Larsson NG. Роль митохондрий в старении. J Clin Invest. 2013. 123 (3): 951–7.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Gray MW, Lang BF, Cedergren R, Golding GB, Lemieux C, Sankoff D, et al.Структура генома и содержание генов в митохондриальной ДНК протистов. Nucleic Acids Res. 1998. 26 (4): 865–78.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Amunts A, Brown A, Bai X, Llacer JL, Hussain T., Emsley P, et al. Структура большой рибосомной субъединицы митохондрий дрожжей. Наука. 2014; 343 (6178): 1485–9.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Шмидт О., Пфаннер Н., Мейзингер С. Импорт митохондриального белка: от протеомики к функциональным механизмам. Nat Rev Mol Cell Biol. 2010. 11 (9): 655–67.
CAS
Статья
PubMed
Google Scholar
Lackner LL. Формирование динамической митохондриальной сети. BMC Biol. 2014; 12:35.
PubMed Central
Статья
PubMed
Google Scholar
Detmer SA, Chan DC.Функции и нарушения митохондриальной динамики. Nat Rev Mol Cell Biol. 2007. 8 (11): 870–9.
CAS
Статья
PubMed
Google Scholar
Хоппинс С., Лакнер Л., Нуннари Дж. Машины, которые делят и объединяют митохондрии. Анну Рев Биохим. 2007. 76: 751–80.
CAS
Статья
PubMed
Google Scholar
Alexandre A, Reynafarje B, Lehninger AL.Стехиометрия векторных движений H +, связанных с транспортом электронов и синтезом АТФ в митохондриях. Proc Natl Acad Sci U S. A. 1978; 75 (11): 5296–300.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Meeusen S, McCaffery JM, Nunnari J. Промежуточные звенья слияния митохондрий, обнаруженные in vitro. Наука. 2004; 305: 1747–52.
CAS
Статья
PubMed
Google Scholar
Гутман М., Котляр А.Б., Боровок Н., Нахлиэль Э. Реакция объемных протонов с препаратом внутренней митохондриальной мембраны: измерения с временным разрешением и их анализ. Биохимия. 1993. 32 (12): 2942–6.
CAS
Статья
PubMed
Google Scholar
Перкинс Г.А., Фрей Т.Г. Недавние исследования структуры митохондрий с помощью микроскопии. Микрон. 2000. 31 (1): 97–111.
CAS
Статья
PubMed
Google Scholar
Liao M, Cao E, Julius D, Cheng Y. Структура ионного канала TRPV1, определенная с помощью электронной криомикроскопии. Природа. 2013; 504 (7478): 107–12.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Кюльбрандт В. Революция в разрешении. Наука. 2014; 343 (6178): 1443–4.
Артикул
PubMed
Google Scholar
Дэвис К.М., Штраус М., Даум Б., Киф Дж. Х., Осевач HD, Рыцовска А. и др.Макромолекулярная организация АТФ-синтазы и комплекса I в целых митохондриях. Proc Natl Acad Sci U S. A. 2011; 108 (34): 14121–6.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Бхарат Т.А., Дэйви Н.Е., Ульбрих П., Ричес Дж. Д., де Марко А., Румлова М. и др. Структура незрелого ретровирусного капсида при разрешении 8 A с помощью криоэлектронной микроскопии. Природа. 2012. 487 (7407): 385–9.
CAS
Статья
PubMed
Google Scholar
Байрхубер М., Майнс Т., Хабек М., Беккер С., Гиллер К., Виллингер С. и др. Структура человеческого потенциалзависимого анионного канала. Proc Natl Acad Sci U S. A. 2008; 105 (40): 15370–5.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Лопис Дж., Маккаффери Дж. М., Мияваки А., Фаркуар М. Г., Цзянь Р. Я. Измерение цитозольного, митохондриального и рН Гольджи в отдельных живых клетках с зелеными флуоресцентными белками.Proc Natl Acad Sci U S. A. 1998; 95 (12): 6803–8.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Кукат С., Вурм К.А., Спар Х., Фалькенберг М., Ларссон Н.Г., Якобс С. Микроскопия со сверхвысоким разрешением показывает, что митохондриальные нуклеоиды млекопитающих имеют одинаковый размер и часто содержат единственную копию мтДНК. Proc Natl Acad Sci U S. A. 2011; 108 (33): 13534–9.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Kukat C, Davies K, Wurm CA, Spähr H, Bonekamp NA, Kühl I, et al. Перекрестное связывание TFAM с одной молекулой мтДНК образует митохондриальный нуклеоид. Proc Natl Acad Sci U S A. 2015. В печати.
Pfeffer S, Woellhaf MW, Herrmann JM, Forster F. Организация митохондриального механизма трансляции, изученная in situ с помощью криоэлектронной томографии. Nat Commun. 2015; 6: 6019.
CAS
Статья
PubMed
Google Scholar
Schleyer M, Neupert W. Транспорт белков в митохондрии: транслокационные промежуточные соединения, охватывающие места контакта между внешней и внутренней мембранами. Клетка. 1985. 43 (1): 339–50.
CAS
Статья
PubMed
Google Scholar
Перкинс Г., Ренкен С., Мартон М.Э., Янг С.Дж., Эллисман М., Фрей Т. Электронная томография митохондрий нейронов: трехмерная структура и организация крист и мембранных контактов.J. Struct Biol. 1997. 119 (3): 260–72.
CAS
Статья
PubMed
Google Scholar
Голд В.А., Иева Р., Вальтер А., Пфаннер Н., ван дер Лаан М., Кульбрандт В. Визуализация активных мембранных белковых комплексов с помощью электронной криотомографии. Nat Commun. 2014; 5: 4129.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Pebay-Peyroula E, Dahout-Gonzalez C, Kahn R, Trezeguet V, Lauquin GJ, Brandolin G.Структура митохондриального носителя АДФ / АТФ в комплексе с карбоксиатрактилоидом. Природа. 2003. 426 (6962): 39–44.
CAS
Статья
PubMed
Google Scholar
Sjostrand FS. Электронная микроскопия митохондрий и двойных мембран цитоплазмы. Природа. 1953, 171 (4340): 30–2.
CAS
Статья
PubMed
Google Scholar
Palade GE. Электронно-микроскопическое исследование структуры митохондрий.J Histochem Cytochem. 1953; 1 (4): 188–211.
CAS
Статья
PubMed
Google Scholar
Дэвис К.М., Ансельми С., Виттиг И., Фаральдо-Гомес Дж. Д., Кюльбрандт В. Структура димера дрожжевой F1Fo-АТФ-синтазы и его роль в формировании митохондриальных крист. Proc Natl Acad Sci U S. A. 2012; 109 (34): 13602–7.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Pfanner N, van der Laan M, Amati P, Capaldi RA, Caudy AA, Chacinska A, et al. Единая номенклатура сайта контакта митохондрий и системы организации крист. J Cell Biol. 2014. 204 (7): 1083–6.
PubMed Central
CAS
Статья
PubMed
Google Scholar
von der Malsburg K, Muller JM, Bohnert M, Oeljeklaus S, Kwiatkowska P, Becker T, et al. Двойная роль митофилина в организации митохондриальной мембраны и биогенезе белков.Dev Cell. 2011. 21 (4): 694–707.
Артикул
PubMed
Google Scholar
Vogel F, Bornhovd C, Neupert W, Reichert AS. Динамическая субкомпартментализация внутренней мембраны митохондрий. J Cell Biol. 2006. 175 (2): 237–47.
PubMed Central
CAS
Статья
PubMed
Google Scholar
фон Баллмос С., Виденманн А., Димрот П. Основы синтеза АТФ с помощью АТФ-синтаз F1F0.Анну Рев Биохим. 2009. 78: 649–72.
Артикул
Google Scholar
Ватт IN, Монтгомери MG, Рансуик MJ, Leslie AG, Walker JE. Биоэнергетическая стоимость создания молекулы аденозинтрифосфата в митохондриях животных. Proc Natl Acad Sci U S. A. 2010; 107 (39): 16823–7.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Stock D, Leslie AGW, Walker JE.Молекулярная архитектура вращательного двигателя в АТФ-синтазе. Наука. 1999; 286: 17701705.
Артикул
Google Scholar
Аллен Р.Д., Шредер С.С., Фок АК. Исследование внутренних мембран митохондрий методами быстрого замораживания и глубокого травления. J Cell Biol. 1989. 108 (6): 2233–40.
CAS
Статья
PubMed
Google Scholar
Strauss M, Hofhaus G, Schroder RR, Kühlbrandt W.Димерные ленты АТФ-синтазы формируют внутреннюю митохондриальную мембрану. EMBO J. 2008; 27 (7): 1154–60.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Аллегретти М., Клуш Н., Миллс Д. Д., Вонк Дж., Кюльбрандт В., Дэвис К. М.. Горизонтальные присущие мембране альфа-спирали в а-субъединице статора АТФ-синтазы F-типа. Природа. 2015; 521 (7551): 237–40.
CAS
Статья
PubMed
Google Scholar
Кюльбрандт В., Карен Д. АТФ-синтазы: новый поворот в древней машине. Trends Biochem Sci. 2015. В печати.
Paumard P, Vaillier J, Coulary B, Schaeffer J, Soubannier V, Mueller DM, et al. АТФ-синтаза участвует в формировании морфологии митохондриальных крист. EMBO J. 2002; 21 (3): 221–30.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Bornhövd C, Vogel F, Neupert W, Reichert AS.Потенциал митохондриальной мембраны зависит от олигомерного состояния супракомплексов F1F0-АТФ-синтазы. J Biol Chem. 2006. 281 (20): 13990–8.
Артикул
PubMed
Google Scholar
Джорджио В., фон Штокум С., Антониэль М., Фаббро А., Фоголари Ф, Форте М. и др. Димеры митохондриальной АТФ-синтазы образуют поры перехода проницаемости. Proc Natl Acad Sci U S. A. 2013; 110 (15): 5887–92.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Hunte C, Koepke J, Lange C, Roßmanith T, Michel H. Структура с разрешением 2,3 Å комплекса цитохрома bc1 из дрожжевых Saccharomyces cerevisiae, совместно кристаллизованных с Fv-фрагментом антитела. Состав. 2000. 8: 669–84.
CAS
Статья
PubMed
Google Scholar
Цукихара Т., Аояма Х., Ямасита Э., Томизаки Т., Ямагути Х., Синдзава-Ито К. и др. Вся структура 13-субъединицы окисла цитохром с оксидазы в 2.8 A. Наука. 1996. 272 (5265): 1136–44.
CAS
Статья
PubMed
Google Scholar
Цикерманн В., Вирт С., Насири Х., Зигмунд К., Швальбе Х., Хант С. и др. Механистическое понимание кристаллической структуры митохондриального комплекса I. Наука. 2015; 347 (6217): 44–9.
CAS
Статья
PubMed
Google Scholar
Ефремов Р.Г., Барадаран Р., Сазанов Л.А.Архитектура дыхательного комплекса I. Природа. 2010. 465 (7297): 441–5.
CAS
Статья
PubMed
Google Scholar
Виноткумар К.Р., Чжу Дж., Херст Дж. Архитектура респираторного комплекса млекопитающих I. Природа. 2014; 515 (7525): 80–4.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Schägger H, Pfeiffer K. Суперкомплексы в дыхательных цепях митохондрий дрожжей и млекопитающих.EMBO J. 2000; 19 (8): 1777–83.
PubMed Central
Статья
PubMed
Google Scholar
Schäfer E, Seelert H, Reifschneider NH, Krause F, Dencher NA, Vonck J. Архитектура активных суперкомплексов дыхательной цепи млекопитающих. J Biol Chem. 2006. 281 (22): 15370–5.
Артикул
PubMed
Google Scholar
Альтхофф Т., Миллс Д. Д., Попот Дж. Л., Кюльбрандт В.Расположение компонентов цепи переноса электронов в митохондриальном суперкомплексе крупного рогатого скота I1III2IV1. EMBO J. 2011; 30 (22): 4652–64.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Lapuente-Brun E, Moreno-Loshuertos R, Acin-Perez R, Latorre-Pellicer A, Colas C, Balsa E, et al. Сборка суперкомплекса определяет поток электронов в митохондриальной цепи переноса электронов. Наука. 2013; 340 (6140): 1567–70.
CAS
Статья
PubMed
Google Scholar
Милейковская Е., Пенчек П.А., Фанг Дж., Маллампалли В.К., Спарагна Г.К., Даухан В. Расположение комплексов дыхательной цепи в суперкомплексе Saccharomyces cerevisiae III2IV2, выявленное с помощью криоэлектронной микроскопии одиночных частиц. J Biol Chem. 2012. 287 (27): 23095–103.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Блаза Дж. Н., Серрели Р., Джонс А. Дж., Мохаммед К., Херст Дж. Кинетические доказательства против разделения пула убихинона и каталитической значимости суперкомплексов дыхательной цепи. Proc Natl Acad Sci U S. A. 2014; 111 (44): 15735–40.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Li P, Nijhawan D., Budihardjo I., Srinivasula SM, Ahmad M, Alnemri ES, et al. Цитохром c и dATP-зависимое образование комплекса Apaf-1 / каспаза-9 запускает каскад апоптотических протеаз.Клетка. 1997. 91 (4): 479–89.
CAS
Статья
PubMed
Google Scholar
Даум Б., Вальтер А., Хорст А., Осиевач HD, Кюльбрандт В. Возрастная диссоциация димеров АТФ-синтазы и потеря крист внутренней мембраны в митохондриях. Proc Natl Acad Sci U S. A. 2013; 110 (38): 15301–6.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Наварро А., Боверис А. Система митохондриальной передачи энергии и процесс старения. Am J Physiol Cell Physiol. 2007; 292 (2): C670–86.
CAS
Статья
PubMed
Google Scholar
Dröse S, Brandt U. Молекулярные механизмы производства супероксида дыхательной цепью митохондрий. Adv Exp Med Biol. 2012; 748: 145–69.
Артикул
PubMed
Google Scholar
Twig G, Shirihai OS. Взаимодействие митохондриальной динамики и митофагии. Антиоксидный окислительно-восстановительный сигнал. 2011; 14 (10): 1939–51.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Scheckhuber CQ, Erjavec N, Tinazli A, Hamann A, Nyström T, Osiewacz HD. Уменьшение деления митохондрий приводит к увеличению продолжительности жизни и пригодности двух моделей старения грибов. Nat Cell Biol. 2007. 9 (1): 99–105.
CAS
Статья
PubMed
Google Scholar
Calvo SE, Mootha VK. Митохондриальный протеом и болезнь человека. Анну Рев Геномикс Хум Генет. 2010; 11: 25–44.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Хендерсон Р. Возможности и ограничения нейтронов, электронов и рентгеновских лучей для микроскопии с атомным разрешением неокрашенных биологических молекул. Q Rev Biophys. 1995; 28: 171–93.
CAS
Статья
PubMed
Google Scholar
Рис DM, Leslie AG, Walker JE. Структура внешней части мембраны АТФ-синтазы крупного рогатого скота. Proc Natl Acad Sci U S. A. 2009; 106 (51): 21597–601.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Symersky J, Pagadala V, Osowski D, Krah A, Meier T., Faraldo-Gomez JD, et al. Структура c (10) кольца митохондриальной АТФ-синтазы дрожжей в открытой конформации. Nat Struct Mol Biol.2012. 19 (5): 485–91. S481.
PubMed Central
CAS
Статья
PubMed
Google Scholar
Растительная жизнь: белки и аминокислоты
Белки находятся во всех клетках и выполняют множество важных клеточных функций. Внутри любой клетки могут быть тысячи различных белков, имеющих различные размеры, структуры и функции. Белки также являются важными структурными компонентами клеточной стенки.
Белки являются наиболее сложными и многочисленными макромолекулами.Внутри клетки многие белки функционируют как ферменты в катализе метаболических реакций, в то время как другие служат транспортными молекулами, запасными белками, переносчиками электронов и структурными компонентами клетки.
Они особенно важны для семян, где они составляют до 40 процентов веса семян и служат для хранения аминокислот для развивающегося зародыша.
Белки также являются важными структурными компонентами клеточной стенки. Поскольку белки и их строительные блоки, аминокислоты, образуют такой крупный компонент жизни растений, растения служат важным пищевым источником восьми-десяти незаменимых аминокислот для людей и других животных.
Аминокислоты
Аминокислоты — это молекулярные строительные блоки белков. Все аминокислоты имеют общую структуру с центральным атомом углерода, альфа-углеродом, ковалентно связанным с атомом водорода, аминогруппой, группой карбоновой кислоты и группой, обозначенной как R-группа, структура которой варьируется от аминокислоты до аминокислота.
Именно разнообразная природа группы R обеспечивает белку многие его структурные и функциональные характеристики. Некоторые группы R являются полярными или электрически заряженными при физиологическом pH, что делает группы R гидрофильными (водолюбивыми).Другие группы R неполярны и гидрофобны (избегают воды).
Двадцать стандартных аминокислот, которые клетка использует для синтеза своих белков, это аланин, аргинин, аспартат (аспарагиновая кислота), аспарагин, цистеин, глутамат (глутаминовая кислота), глутамин, глицин, гистидин, изолейцин, лейцин, лизин, метионин, фенилаланин. , пролин, серин, треонин, триптофан, тирозин и валин.
Каждая из двадцати аминокислот отличается от других девятнадцати структурой своей R-группы.После включения в белок стандартная аминокислота может подвергаться модификации, чтобы создать нестандартные аминокислоты и еще большее разнообразие белковых структур.
Одной из наиболее распространенных нестандартных аминокислот, содержащихся в белках, является гидроксипролин, который обычно содержится в белках клеточной стенки растений. В дополнение к двадцати аминокислотам, из которых строятся белки, многие нестандартные аминокислоты свободно встречаются в клетке и не обнаруживаются в белках. Канаванин, например, содержится в семенах многих бобовых.
Основываясь на информации о клеточной дезоксирибонуклеиновой кислоте (ДНК), клетка соединяет двадцать стандартных аминокислот пептидными связями в определенных последовательностях, в результате чего образуются цепи длиной от двух до многих тысяч. Более короткие цепи аминокислот называются пептидами или олигопептидами, а более длинные цепи называются полипептидами.
Термин «белок» обычно зарезервирован для тех олигопептидов и полипептидов, которые выполняют биологические функции, поскольку отдельные полипептиды часто не выполняют биологических функций, если они не связаны с другими полипептидами.
Первичная структура
Белки отличаются друг от друга последовательностями своих аминокислот. Последовательность аминокислот белка называется его первичной структурой. Было показано, что мутации приводят к изменению всего лишь одной аминокислоты в белке. Поскольку ДНК определяет первичную структуру белка, информация о последовательности белка часто используется для изучения эволюционных взаимоотношений между организмами.
Белки часто образуют комплексы с другими соединениями в их биологически активном состоянии.Эти белки называются конъюгированными белками. Белки в комплексе с металлами, липидами, сахарами и рибофлавином называются металлопротеинами, липопротеинами, гликопротеинами и флавопротеинами соответственно.
Гликопротеины (буквально «сахарные белки») являются важными составляющими плазматической мембраны. Эти молекулы сахара могут встречаться по отдельности или в виде коротких простых разветвленных цепей.
| Превращение амоновых кислот в белки |
Белковая цепь может быть свернута в различные трехмерные формы.Трехмерная форма, которую принимает белок, называется его конформацией и определяется его аминокислотной последовательностью.
Чтобы белок был активным, он должен принять определенную конформацию. Любое изменение его строения может привести к снижению активности. Денатурирующие агенты изменяют структуру белка, так что он теряет свою конформацию, биологическую функцию и активность.
Вторичная структура
Вторичная структура относится к локальному складыванию или конформации полипептидной цепи на относительно коротких (около пятидесяти аминокислот) участках.
Две общие вторичные структуры, альфа-спираль и бета-лист, регулярно встречаются в белках. В среднем только около половины полипептидной цепи принимает альфа- или бета-конформацию, тогда как остальная часть существует в виде чередований и случайных структур.
Некоторые белки показывают только альфа-структуру, другие — только бета-структуру, в то время как третьи демонстрируют либо смесь двух структур, либо ни одну вторичную структуру. Как альфа-, так и бета-структуры повышают структурную стабильность белка.Аминокислотная последовательность определяет, будет ли конкретная последовательность аминокислот в белке иметь альфа- или бета-структуру.
Третичная структура
Общая пространственная ориентация всей полипептидной цепи в пространстве называется ее третичной структурой. Обычно различают две третичные структуры. Волокнистые или нитчатые белки расположены в виде волокон или листов, в то время как глобулярные белки расположены примерно в виде сферических или глобулярных структур.