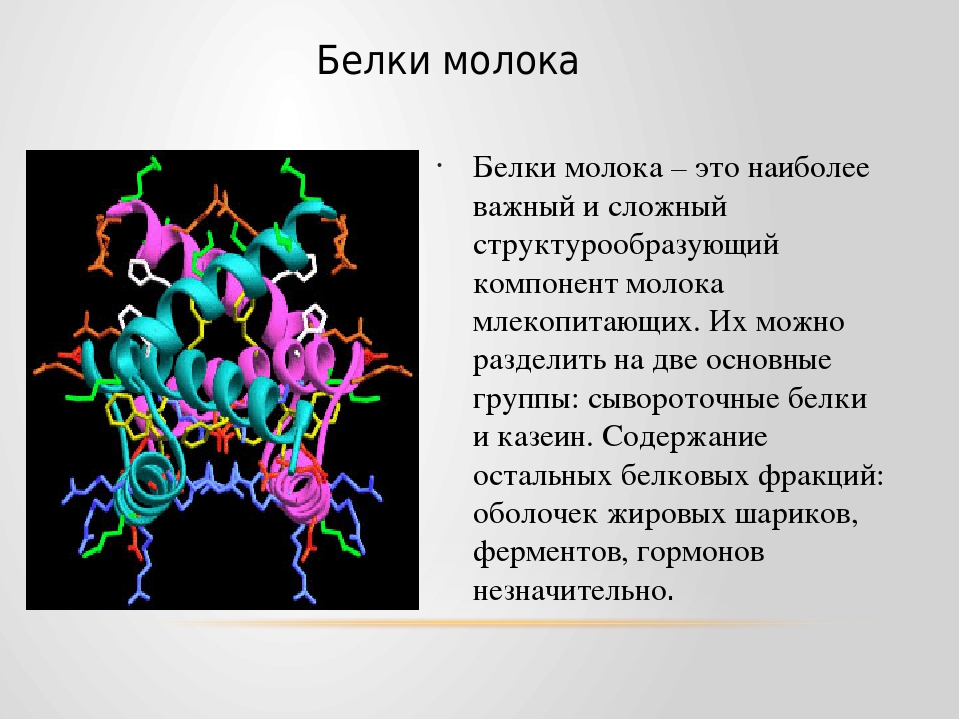
Белки основные — Справочник химика 21
Не следует забывать, что среди аминокислот, входящих в состав природных белков, имеются также моноаминодикарбоновые кислоты и диаминомонокарбоновые кислоты. Избыток первых в данном белке увеличивает его кислотный характер. В белках основного характера содержится некоторый избыток диаминокислот. [c.395]
Гистоны—белки основного характера, так как содержат значительное количество диаминокислот со свободными аминогруппами они растворимы в воде и в разбавленных кислотах, но не растворимы в разбавленных щелочах. Обычно представляют собой собственно белковые части сложных белков. Представитель глобин — белок, входящий в состав сложного белка крови — гемоглобина. [c.297]
Обе стадии невозможно отделить одну от другой. Сущность тепловой денатурации можно рассмотреть на примере глобулярных белков. Основная молекула глобулярного белка, как известно, состоит из одной или нескольких полипептидных цепей, сложенных складками и образующих клубки. Такая структура стабилизируется непрочными связями, среди которых большую роль играют водородные связи, образующие поперечные мостики между параллельными пептидными цепями или их складками. При нагревании белков происходит усиленное движение полипептидных цепей или их складок, что вызывает разрыв непрочных связей между ними. В результате этого наблюдается развертывание и перегруппировка складок, сопровождаемые перераспределением полярных и неполярных радикалов, причем неполярные радикалы концентрируются на поверхности глобул, понижая их гидрофильность, а следовательно, и растворимость. [c.370]
Такая структура стабилизируется непрочными связями, среди которых большую роль играют водородные связи, образующие поперечные мостики между параллельными пептидными цепями или их складками. При нагревании белков происходит усиленное движение полипептидных цепей или их складок, что вызывает разрыв непрочных связей между ними. В результате этого наблюдается развертывание и перегруппировка складок, сопровождаемые перераспределением полярных и неполярных радикалов, причем неполярные радикалы концентрируются на поверхности глобул, понижая их гидрофильность, а следовательно, и растворимость. [c.370]
Последние годы отмечены повышенным интересом к ферментам, взаимодействующим с ДНК. Большинство из них — белки основного характера, и их удобнее очищать на катионообменниках чаще всего для этой цели используют фосфоцеллюлозу. [c.304]
Подобно аминокислотам, белки сочетают в себе как кислотные, так и основные свойства. Являясь амфотерными полиэлектролитами, белки тем не менее существенно отличаются от свободных аминокислот, кислотно-основные свойства которых обусловлены а-амино- и а-карбоксильными группами. В белках основной вклад в формирование кислотно-основных свойств вносят заряженные радикалы аминокислотных остатков, расположенные на поверхности белковой глобулы. Основные свойства белков связаны с такими аминокислотами, как аргинин, лизин или гистидин, а кислые — с аспарагиновой и глутаминовой аминокислотами. Что касается а-аминных и а-кар-боксильных групп аминокислот, то их ионизация не имеет существенного значения, так как подавляющее их число участвует в образовании пептидных связей. Кривые титрования белков достаточно сложны для интерпретации. Это связано, во-первых, с наличием большого числа титруемых групп, а также с тем, что рА для каждой титруемой группы в белке может существенно отличаться от таковой в аминокислоте. Это связано с электростатическими взаимодействиями между ионизированными группами белка, наличием близко расположенных гидрофобных остатков, а также влиянием водородных связей. [c.52]
В белках основной вклад в формирование кислотно-основных свойств вносят заряженные радикалы аминокислотных остатков, расположенные на поверхности белковой глобулы. Основные свойства белков связаны с такими аминокислотами, как аргинин, лизин или гистидин, а кислые — с аспарагиновой и глутаминовой аминокислотами. Что касается а-аминных и а-кар-боксильных групп аминокислот, то их ионизация не имеет существенного значения, так как подавляющее их число участвует в образовании пептидных связей. Кривые титрования белков достаточно сложны для интерпретации. Это связано, во-первых, с наличием большого числа титруемых групп, а также с тем, что рА для каждой титруемой группы в белке может существенно отличаться от таковой в аминокислоте. Это связано с электростатическими взаимодействиями между ионизированными группами белка, наличием близко расположенных гидрофобных остатков, а также влиянием водородных связей. [c.52]
Гистоны. Белки основного характера. Находятся в виде нуклеопротеидов (см. стр. 392) в лейкоцитах и красных кровяных шариках. [c.390]
стр. 392) в лейкоцитах и красных кровяных шариках. [c.390]
Белки — это важнейшие для жизни вещества. Белки — основной структурный компонент живых тканей. Посмотрите на своего соседа. Все, что вы видите кожа, волосы, глаза, ногти, — это белки. Костные ткани, кровь, мозг — все содержит белки. Кроме того, все ферменты, контролирующие химические процессы в организме, представляют собой белки. В каждом чело- [c.258]
Физиологическое значение белков трудно переоценить не случайно их называют носителями жизни . Белки-основной материал, из которого построен живой организм, т.е. протоплазма каждой живой клетки. [c.230]
Огромное разнообразие белков есть следствие их эволюции. Эволюция явилась результатом многочисленных природных экспериментов (мутации и последующий отбор), которые могут быть использованы для изучения принципов строения белков. Основной мутационной ступенью эволюции белков является замена аминокислотного остатка на следующих по значимости этапах происходят вставки и делеции одного или большего числа остатков очень большие изменения являются результатом мультипликации и слияния генов. [c.241]
[c.241]
Японскую ассоциацию по пищевому растительному белку. Основные фирмы, фигурирующие в голубой книге Американской ассоциации по сое, представлены ниже. [c.660]
Исследования механизма свертывания, отвечающие второму подходу к установлению структурной организации белка, базируются на многочисленных физических, химических и биологических методах исследования, которые дают прямую или косвенную информацию о геометрических, термодинамических и кинетических аспектах процессов денатурации и ренатурации, механизме клеточного синтеза аминокислотной последовательности и взаимодействия белковых цепей с шаперона-ми. В исследованиях этого плана, как и предшествующего, надежда возлагается на то, что в результате анализа экспериментальных данных в конечном счете удастся разработать эмпирические правила, позволяющие предсказывать по известному химическому строению белка основные этапы свертывания, в первом случае, и нативную пространственную структуру, во втором. Далее, предполагается, если эти цели будут достигнуты, то станет ясно не только как возникает физиологически активная конформация, но и почему она возникает, т.е. бу- [c.77]
Далее, предполагается, если эти цели будут достигнуты, то станет ясно не только как возникает физиологически активная конформация, но и почему она возникает, т.е. бу- [c.77]
Представление о пространственной структуре пептидов и белков, якобы «предопределенной конформацией остова», не следует ни из экспериментальных фактов, ни из результатов расчета. Оно родственно а-спиральной концепции и является следствием стереотипности мышления, а также, по-видимому, магии слов. Появление таких терминов, как «остов», «основа», «скелет», обычно связано с необходимостью подчеркнуть фундаментальные, самые существенные свойства структуры или ее частей, В лексикон исследователей пространственного строения пептидов и белков слова «пептидный остов» и «пептидный скелет» пришли от исследователей химического строения этих соединений. Там они совершенно точно передавали суть химической структуры изучаемых объектов. Но эти слова потеряли свой первозданный смысл и приобрели ложный, иллюзорный, как только стали употребляться в описаниях пространственного строения пептидов и белков. Основные цепи пептидных и белковых молекул обретают лишь видимость остова или скелета в нативных конформациях, т.е. в состоянии, когда реализована полная схема межостаточных невалентных взаимодействий, прежде всего, взаимодействий типа «боковая цепь — боковая цепь» и «боковая цепь — основная цепь». Вне этих взаимодействий, т,е, в условиях денатурации, видимость пропадает, иллюзия рассеивается и химическая основа пептидов и белков превращается в гибкую цепь, которая не может самостоятельно удерживать свою форму, В предположении об особой конформационной роли пептидного остова авторы [22] делают одно исключение, В связи с этим они замечают «Сказанное не относится к ситуации, когда следует учитывать дающие весьма существенный вклад в конформационную энергию электростатические взаимодействия ионогенных групп в этом случае конформация боковой цепи, несущей ионогенную группу, должна быть «приравнена» к конформации пептидного остова» [22, С, 36], Таким образом, в структурной организации пептидов особая роль отводится также электростатическим взаимодействиям и, прежде всего, взаимодействиям между заряженными группами.
Основные цепи пептидных и белковых молекул обретают лишь видимость остова или скелета в нативных конформациях, т.е. в состоянии, когда реализована полная схема межостаточных невалентных взаимодействий, прежде всего, взаимодействий типа «боковая цепь — боковая цепь» и «боковая цепь — основная цепь». Вне этих взаимодействий, т,е, в условиях денатурации, видимость пропадает, иллюзия рассеивается и химическая основа пептидов и белков превращается в гибкую цепь, которая не может самостоятельно удерживать свою форму, В предположении об особой конформационной роли пептидного остова авторы [22] делают одно исключение, В связи с этим они замечают «Сказанное не относится к ситуации, когда следует учитывать дающие весьма существенный вклад в конформационную энергию электростатические взаимодействия ионогенных групп в этом случае конформация боковой цепи, несущей ионогенную группу, должна быть «приравнена» к конформации пептидного остова» [22, С, 36], Таким образом, в структурной организации пептидов особая роль отводится также электростатическим взаимодействиям и, прежде всего, взаимодействиям между заряженными группами. [c.399]
[c.399]
В процессе эволюции белков происходят замены отдельных остатков, вставки и делеции нескольких остатков, удвоение и слияние генов. Для белков основные этапы исторического процесса знаменуются заменами аминокислотных остатков в полипептидной цепи. С течением времени эти замены накапливаются, так что в конечном счете какое-либо сходство между исходной и окончательной аминокислотными последовательностями может исчезнуть. Однако, как правило, даже после того как исчезнет сходство аминокислотных последовательностей двух гомологичных белков, сохраняется соответствие в укладке их цепей. Тенденции к замещениям заметно отличаются у остатков в разных положениях в цепи (рис. 7.1, б). Различия гомологичных белков не исчерпываются за- [c.197]
Мембраны эритроцитов содержат около восьми основных полипептидов [6]. Пять из них являются внешними и составляют 40 % общего содержания белка. Основным внутренним белком является гликофорин, один из немногих внутренних белков с установленной аминокислотной последовательностью (рис. 25.3.7) . В его молекуле несколько аминокислотных остатков связано с олигосахаридными фрагментами, которые в основном определяют антигенные и рецепторные свойства эритроцитов эти олигосахариды локализованы исключительно в Л -концевой части аминокислотной последовательности и находятся на внешней поверхности мембраны. Примечательна также высокая концентрация остатков дикарбоновых аминокислот в С-концевой последовательности. Однако наибольший интерес представляет участок между М- и концевыми последовательностями, содержащий около двадцати [c.121]
25.3.7) . В его молекуле несколько аминокислотных остатков связано с олигосахаридными фрагментами, которые в основном определяют антигенные и рецепторные свойства эритроцитов эти олигосахариды локализованы исключительно в Л -концевой части аминокислотной последовательности и находятся на внешней поверхности мембраны. Примечательна также высокая концентрация остатков дикарбоновых аминокислот в С-концевой последовательности. Однако наибольший интерес представляет участок между М- и концевыми последовательностями, содержащий около двадцати [c.121]
При помощи метода фракционирования гомогенатов органов и тканей в центрифугах было показано, что ядерная фракция печени и почек содержит незначительное число ферментов, хотя известно, что в ядрах осуществляется синтез некоторых белков. Основное место синтеза белка, как теперь установлено,— фракция рибосом цитоплазмы. Показано, кроме [c.158]
Гистоны — белки основного характера, растворимые в разбавленных растворах кислот и щелочей осаждаются этанолом. [c.80]
[c.80]
Активацию рассматривают как суммарный результат физико-химических процессов реорганизации структуры и снижения стабильности макромолекул и мембран. Их термодинамические и кинетические характеристики сходны с показателями тепловой денатурации белков и плавления нуклеиновых кислот. Все химические реагенты и воздействия, вызывающие активацию, влияют на структурную организацию клеточных биополимеров и мембран. Активация спор при низких значениях pH происходит в той же зоне, где и денатурация белков. Основной итог активации — подготовка клетки к следующему этапу прорастания — инициации. [c.107]
Молекула ДНК состоит из 2 сплетенных комплементарных цепей нуклеотидов, которые удерживаются вместе за счет так называемого спаривания оснований. Особенности синтеза ДНК приводят к антипараллельности комплементарной цепи, 3 -конец одной цепи находится рядом с 5 -концом другой. Структурно цепи образуют двойную спираль, представляющую как бы винтовую лестницу, закрученную вправо вокруг воображаемой центральной оси. В свою очередь ДНК и РНК отвечают за синтез белка — основной структурной единицы любого организма. [c.440]
В свою очередь ДНК и РНК отвечают за синтез белка — основной структурной единицы любого организма. [c.440]
В процессе высушивания зерна не должны быть денатурированы белки, основное количество которых находится в его зародыше. Эти белки чрезвычайно термолабильны, и при неправильном режиме сушки зерно теряет свою всхожесть. Поэтому максимально допустимая температура нагрева зерна в основном определяется термоустойчивостью его белкового комплекса. С повышением влажности зерна его термоустойчивость падает. Сохранение семенных и продовольственных качеств зерна зависит не только от температуры, но и от скорости нагревания и времени выдержки при максимальной температуре [50]. Поэтому для сушки зерна применяют многокамерные сушилки с различным режимом по зонам и с охлаждением. По-видимому, целесообразно также применение цилиндро-конических аппаратов, в которых создается организованное движение материала. [c.212]
Физиологическое значение белков трудно переоценить не случайно их называют носителями жизни . Белки — основной материал, из которого построен живой организм, т. е. протоплазма каждой живой клетки. Особый класс белков образуют биологические катализаторы— ферменты (см. 40.5) белковую природу имеет также часть гормонов (см. 40.4). Склеропротеины служат живым организмам для создания органов защиты (рога) или теплоизоляции (перьевой или волосяной покров). [c.547]
Белки — основной материал, из которого построен живой организм, т. е. протоплазма каждой живой клетки. Особый класс белков образуют биологические катализаторы— ферменты (см. 40.5) белковую природу имеет также часть гормонов (см. 40.4). Склеропротеины служат живым организмам для создания органов защиты (рога) или теплоизоляции (перьевой или волосяной покров). [c.547]
Гистоны — белки основного характера, растворимые в разбавленных растворах кислот и щелочей осаждаются этанолом. Содержатся в клеточных ядрах, где они слабо связаны с нуклеиновыми кислотами. [c.550]
Мы не пытались углубляться во многие интересные применения систем с водородными связями. В гл. 11 дается лишь краткий обзор некоторых применений, главным образом для характеристики разнообразных возможностей. Даже в гл. 10 (где рассматриваются белки) основное внимание сосредоточено только на одном вопросе — роли водородной связи в структуре белков. [c.7]
Несмотря на всю сложность молекулы белка, основные принципы ее структуры делаются все яснее, и все более очевидной становится важная роль Н-связи в этой структуре. [c.254]
[c.254]
Гистоны, также обладающие основным характером, имеют более сложный состав и больший молекулярный вес, чем протамины, приближаясь тем самым к обычным белкам. И у этих белков основность обусловлена высоким содержанием аргинина. Они растворимы в воде и осаждаются аммиаком при нагревании они свертываются только в присутствии электролитов и то частично. Гистоны гидролизуются пепсином. Они находятся в ядрах клеток, связанные, как и протамины, с нуклеиновыми кислотами в виде нуклеопротеидов (они получаются легче всего из богатых ядрами органов, например щитовидной железы), [c.446]
Первыми генной инженерией всерьез заинтересовались фармацевтические фирмы. Для них возможность сравнительно дешево производить практически любые белки в больших количествах открывает совершенно новые горизонты. Ведь, помимо того, что белки — основные рабочие молекулы в клетке, они играют ещё ключевую [c.122]
Глинистые материалы, адсорбируя микробные клетки, образуют с ними крупные агрегаты размером 50—200 мкм. Подобные агрегаты образуются со всеми испытанными дрожжами и бактериями, а также при внесении белков, основных аминокислот и других веществ, подвергающихся адсорбции на частицах глин. Если адсорбция клеток не происходит или происходит очень слабо, то подобных агрегатов не образуется. Размеры агрегатов в определенной степени зависят от числа микробных клеток, адсорбированных минералом. [c.70]
Подобные агрегаты образуются со всеми испытанными дрожжами и бактериями, а также при внесении белков, основных аминокислот и других веществ, подвергающихся адсорбции на частицах глин. Если адсорбция клеток не происходит или происходит очень слабо, то подобных агрегатов не образуется. Размеры агрегатов в определенной степени зависят от числа микробных клеток, адсорбированных минералом. [c.70]
В водных растворах белки обладают свойствами слабых кислот или слабых оснований, в зависимости от преобладания в молекуле белка дикарбоновых кислот (глютаминовая, аспарагиновая) или диаминокислот (лизин, аргинин). Белки кислого характера (альбумины, глобулины) в водном растворе несут отрицательный заряд, белки основного характера (протамины, гистоны) — положительный. [c.25]
Заряд белковой молекулы в нейтральней среде определяется соотношением количества свободных групп —СООН и —ЫНд и степенью их диссоциации. Чем больше карбоксильных групп —СООН, тем выше окажется отрицательный заряд, и белок будет проявлять свойства слабой кислоты. Преобладание аминных 1 рупп —Nh3 сообшает белку основные свойства и положительный заряд. [c.188]
Преобладание аминных 1 рупп —Nh3 сообшает белку основные свойства и положительный заряд. [c.188]
Белки—основной материал, из ]соторою строится структура живой клетки, [c.3]
Наиболее интересной и важной группой из числа сложных белков являются нуклеопротеиды, в них белок соединен с нуклеиновой кислотой. В отличие от других сложных белков нуклеопротеиды состоят из сравнительно простого и низкомолекулярного белка основного характера — протамина или гистона, и высокомолекулярной простетической группы — нуклеиновой кислоты. Нуклеопротеиды входят в состав всех клеток живого организма, являются важной частью хромозом и т. д. К этому классу соединений относятся и молекулы ]-иганты — вирусы, инициаторы многих инфекционных заболеваний, например, полиомиелита. [c.533]
Все нуклеопротеиды можно разделить по меньшей мере на два типа. К первому типу относятся нуклеопротеиды, в которых нуклеиновая кислота связана солевой связью с простыми белками основного характера и низкого молекулярного веса. Такими белками могут быть протамины (сальмин, клупеин, сту-рин), встречающиеся в сперме рыб. К этому же типу относятся нуклеопротеиды, в которых нуклеиновая кислота связана с основными белками более высокого молекулярного веса — гистолами. Примером могут служить нуклеопротеиды, встречающиеся в тканях зобной и поджелудочной желез. Ко второму типу мы относим более сложные структуры — вирусы растений (например, вирус табачной мозаики) и бактериофаги. Содержание нуклеиновых кислот в вирусах колеблется от 5 до 50%. Природа связи между белками и нуклеиновыми кислотами в вирусных нуклеопротеидах изучена слабее, чем в нуклеопро-теидах первого типа. Известно, что в вирусном нуклеопротеиде связи между белком и нуклеиновыми кислотами более лабильны и что для белков вирусов характерно высокое содержание основных аминокислот. Даже сравнительно простые вирусы имеют весьма сложное строение. Еще более сложное строение у таких вирз сов, как вирусы гриппа и пситтакоза. Последние могут даже быть отнесены к микроорганизмам.
Такими белками могут быть протамины (сальмин, клупеин, сту-рин), встречающиеся в сперме рыб. К этому же типу относятся нуклеопротеиды, в которых нуклеиновая кислота связана с основными белками более высокого молекулярного веса — гистолами. Примером могут служить нуклеопротеиды, встречающиеся в тканях зобной и поджелудочной желез. Ко второму типу мы относим более сложные структуры — вирусы растений (например, вирус табачной мозаики) и бактериофаги. Содержание нуклеиновых кислот в вирусах колеблется от 5 до 50%. Природа связи между белками и нуклеиновыми кислотами в вирусных нуклеопротеидах изучена слабее, чем в нуклеопро-теидах первого типа. Известно, что в вирусном нуклеопротеиде связи между белком и нуклеиновыми кислотами более лабильны и что для белков вирусов характерно высокое содержание основных аминокислот. Даже сравнительно простые вирусы имеют весьма сложное строение. Еще более сложное строение у таких вирз сов, как вирусы гриппа и пситтакоза. Последние могут даже быть отнесены к микроорганизмам. Подробное строение вирусов этой группы здесь не рассматривается. [c.246]
Подробное строение вирусов этой группы здесь не рассматривается. [c.246]
Наряду с жирами и углеводами белки — основная составная часть пищи человека. В индустриальных странах главным источником пищевых белков являются продукты животного пронсхождення, в то время как в развивающихся странах в пище преобладают биологически неполноценные растительные белки. Для удовлетворения потребности постоянно растущего населения помимо увеличения производства животных и растительных продуктов, выведения сортов зерновых с повышенным содержанием недостающих аминокислот и повышения ценности биологически неполноценных растительных белков добавлением синтетических аминокислот все большее значение приобретает дальнейшее развитие микробиологических щюцессов получения белков одноклеточных микроорганизмов [10 — 15]. Микробиологические процессы основаны на способности определенных микроорганизмов использовать в обмене веществ в качестве источника углерода такие вешества, как углеводороды нефти, спирты или сырье, содержащее углеводы (крахмал, меласса, целлюлоза). Обзор важнейших процессов дан в табл. 3-1. [c.341]
Обзор важнейших процессов дан в табл. 3-1. [c.341]
Белки — основные вещества живой материи, их физиологическая и структурная роль чрезвычайно важна. В самом деле, в форме голопротеинов или гетеропротеинов, т. е. белков, связанных с другими молекулами, они содержатся во всех ком-партментах того биологического единства, которым является клетка. [c.125]
При филировании белков основным критерием служит молекулярная масса. Общепризнано, что в диапазоне 10—50 тыс. Да белки проявляют высокую склонность к филированию. Молекулы с очень малой массой дают прядильные растворы с чрезвычайно низкой вязкостью, образующие нестойкие белковые нити, которые быстро диспергируются в коагулирующем растворе. Наоборот, из белков с очень высокой молекулярной массой на этапе денатурации получаются прядильные растворы с очень сильной вязкостью, что делает невозможным прядение волокон. Некоторые белки также образуют очень густые гели в щелочной среде за счет появления межцепочечных ковалентных связей типа дисульфидный мостик , что вызывает необходимость подгонять, приспосабливать параметры филирования [97]. [c.537]
[c.537]
Исследование структурных и конформационных свойств индивидуальной полипептидной субъединицы обычно ведется по трем аспектам рассматриваются первичная, вторичная и третичная структуры, как это было предложено Линдерстрем-Лангом [И]. Организмы используют для синтеза белков основной набор из 20 аминокислот (см. гл. 23.2). Необычные аминокислоты, которые эпизодически встречаются в белковых структурах, часто являются результатом химической модификации простетической группы гема (остатки 70—80) они остались неизменными в процессе эволюции, тогда как другие части молекулы играют, по-видимому, меньшую роль для спецификаций, особенно остатки, не участвующие в агре- [c.222]
Гистоны также являются белками основного характера. В их состав входят лизин и аргинин, содержание которых, однако, не превышает 20—30%. Молекулярная масса гистонов намного больше нижнего предела молекулярной массы белков. Эти белки сосредоточены в основном в ядрах клеток в составе дезоксирибонуклеопротеинов и играют важную роль в регуляции экспрессии генов (см. главы 2 и 3). [c.73]
главы 2 и 3). [c.73]
В 1916 г. в опытах на животных было показано токсичное действие сырого яичного белка употребление печени или дрожжей снимало этот эффект. Фактор, предотвращающий развитие токсикоза, был назван витамином Н. Позже было установлено, что в дрожжевом экстракте печени и желтке куриного яйца содержится пищевой фактор, отличный от всех других известных к этому времени витаминов. Этот фактор стимулирует рост дрожжей и азотфиксирующих бактерий Rhizobium, в связи с чем он и получил название биотин (от греч. bios—жизнь), или коэнзим R. В 1940 г. было установлено, что все три названия (биотин, витамин Н и коэнзим R) относятся к одному и тому же химически индивидуальному соединению. Выделенное из сырого яичного белка вещество оказалось гликопротеином—белком основного характера, названным авидином этот белок обладает высоким сродством связывания с биотином с образованием нерастворимого в воде комплекса. Комплекс не подвергается расщеплению в пищеварительном тракте, поэтому биотин не всасывается, хотя и содержится в пищевых продуктах. [c.228]
[c.228]
От чего зависит заряд белка в водном растворе Назовите белкй основного и кислого характера. [c.44]
Свободные аминокислоты и входящие в состав белков основные и кислотные остатки являются электролитами. Функционируя в водной среде, они диссоциируют на ионы. Рационально пользоваться следующим определением Бренстеда кислота — молекула, от которой отщепляется протон основание — иолеку-ла, присоединяющая протон. Сам растворитель — вода — выступает как кислота в реакции НаО ОН и как основание в реакции НаО 4-Н+Н3О+ (ион оксония). [c.61]
Гистоны. Белки основного характера. Содержатся в нук-леопрртеидах лейкоцитов и красных кровяных шариков. [c.501]
Точное предсказание структуры белка даст прорыв в создании лекарств
Фантасты прошлого мечтали об искусственном сверхразуме, который решит все проблемы человечества. Но сейчас мы чаще видим такие заголовки: «Нейросеть подделала голоса знаменитостей», «нейросеть научилась заменять лица актеров в фильмах», «нейросеть записала рэп-альбом»…
Но сейчас мы чаще видим такие заголовки: «Нейросеть подделала голоса знаменитостей», «нейросеть научилась заменять лица актеров в фильмах», «нейросеть записала рэп-альбом»…
Впрочем, есть и по-настоящему серьезные достижения. Одно из них связано с предсказанием поведения живых систем. В конце 2020 года программисты DeepMind — исследовательского подразделения Google — создали программу, которая может построить структуру белка по последовательности его аминокислот. На основе этих данных ученые могут узнать о белке очень многое. В том числе и то, как он работает.
Олег Михайловский — сотрудник Лаборатории биомолекулярного ядерно-магнитного резонанса СПбГУ, один из тех, кто в России занимается изучением структуры белков. Он разрабатывает дополнение к программному пакету Amber — одному из двух крупнейших в мире инструментов для биомолекулярного моделирования. Разработка ученого позволяет получать высокоточные структуры белков на основе данных рентгеновской дифракции и компьютерной симуляции молекулярной динамики. Мы попросили его рассказать простыми словами об исследованиях, которые мало кто понимает, но которые могут изменить нашу жизнь уже очень скоро.
Мы попросили его рассказать простыми словами об исследованиях, которые мало кто понимает, но которые могут изменить нашу жизнь уже очень скоро.
О том, почему структура белка — это «загадка века»
Белки выполняют очень много функций в организме. Проще сказать, чего они не делают. Например, ферменты, которые помогают нам переваривать еду — это белки. Из фибриллярных белков состоят наши ткани, такие как мышцы или волосы. Белки участвуют в формировании иммунитета, обеспечивают саморегуляцию организма — например, свертываемость крови. Представьте длинную нить из связанных друг с другом шнурков разного цвета, которые сплетены в очень затейливую фигуру. Шнурки — это аминокислоты, строительные элементы белка. Основных аминокислот всего двадцать, но они расположены в разном порядке. Узнав их последовательность, можно попытаться теоретически предсказать структуру белка и то, как он ведет себя в организме. А это открывает огромные возможности.
Могу привести в пример болезнь Альцгеймера. Один из ее характерных признаков — накопление в мозге неправильно свернутых белков бета-амилоидов, которые вызывают гибель нейронов. По сути, из-за неправильной структуры белки начинают друг на друга наслаиваться и блокировать взаимодействие между клетками. Такие поломки в белках неплохо бы идентифицировать. А потом уже на основании модели внести мутацию туда, где она будет полезна. Увеличить активность какого-нибудь белка. Или, наоборот, снизить ее.
Один из ее характерных признаков — накопление в мозге неправильно свернутых белков бета-амилоидов, которые вызывают гибель нейронов. По сути, из-за неправильной структуры белки начинают друг на друга наслаиваться и блокировать взаимодействие между клетками. Такие поломки в белках неплохо бы идентифицировать. А потом уже на основании модели внести мутацию туда, где она будет полезна. Увеличить активность какого-нибудь белка. Или, наоборот, снизить ее.
О прорыве, которые обеспечили нейросети
Технически мы можем определить структуру белка вручную. Например, с помощью рентгеновской кристаллографии. Как это происходит? Мы выращиваем кристаллик с белком, светим на него лучом и получаем красивые картиночки с детектора. Дальше мы смотрим, где у нас на картинке точки интенсивности, и по ним можем рассчитать модель этого белка — то есть понять, где какой атом находится.
Но с нуля получить эту модель очень сложно и затратно. На одно только выращивание кристаллика хорошего качества может уйти несколько месяцев, в зависимости от белка, однако заранее предсказать, сколько займет процесс подбора «правильных» условий для его роста, практически невозможно. А есть белки, которые вообще никак не кристаллизуются. Есть и другие методы спектроскопии, такие как ЯМР, но они по своим причинам могут не сработать. И тут нам помогает компьютерное моделирование. Если мы не знаем структуру какого-то нового белка, ее можно попробовать предсказать по структуре его «родственника». За много лет на основе экспериментальных данных у нас накопилось порядка 170 тысяч уже известных структур. Если обучить машинный алгоритм на их основе, можно получить относительно точные предсказания. Например, нейросеть AlphaFold2 от Google. Но их модель ошибается примерно на размер атома, а это может быть важным для предсказания взаимодействий этого белка с другими молекулами.
А есть белки, которые вообще никак не кристаллизуются. Есть и другие методы спектроскопии, такие как ЯМР, но они по своим причинам могут не сработать. И тут нам помогает компьютерное моделирование. Если мы не знаем структуру какого-то нового белка, ее можно попробовать предсказать по структуре его «родственника». За много лет на основе экспериментальных данных у нас накопилось порядка 170 тысяч уже известных структур. Если обучить машинный алгоритм на их основе, можно получить относительно точные предсказания. Например, нейросеть AlphaFold2 от Google. Но их модель ошибается примерно на размер атома, а это может быть важным для предсказания взаимодействий этого белка с другими молекулами.
Но мы все равно не можем обойтись без экспериментов. Модель AlphaFold тренируется на определенном наборе, но что она делает помимо этого набора, сложно сказать. Для известных белков мы можем подтвердить или опровергнуть результат, полученный с помощью нейросети. А для новых мы проверить результаты никак не можем. Поэтому нужно получать экспериментальные данные, чтобы подтверждать предсказания. И как раз обработка этих данных с помощью нашей программной «надстройки» позволяет строить максимально точные модели белков, которые пока не под силу ни одной нейросети.
Поэтому нужно получать экспериментальные данные, чтобы подтверждать предсказания. И как раз обработка этих данных с помощью нашей программной «надстройки» позволяет строить максимально точные модели белков, которые пока не под силу ни одной нейросети.
О том, как провести эксперимент в виртуальной реальности
Эксперимент на компьютере — звучит странно. Но это именно эксперимент. Ты запускаешь симуляцию и смотришь на результат. У нас есть определенный базовый протокол, по которому идет симуляция. Ты настраиваешь условия, и нужно посмотреть, что будет происходить в зависимости от этих условий. А этих условий, например, сотня. Естественно, руками их настраивать никто не будет — для этого я пишу скрипт. Думаю, любой современный ученый должен уметь работать со скриптами, без этого уже невозможно. 100 условий руками щелкать — это не прикольно.
Как это происходит на практике? Допустим, нам нужно попытаться предсказать, как одна молекула соединяется с другой. Есть математическая функция, которая описывает, как себя ведут атомы друг по отношению к другу. Мы подставляем условия и просчитываем их для тысячи разных молекул — по отношению к одной целевой. Запускаем тысячу симуляций. Потом на основании результатов можно, например, понять, как будет работать молекула лекарства.
Мы подставляем условия и просчитываем их для тысячи разных молекул — по отношению к одной целевой. Запускаем тысячу симуляций. Потом на основании результатов можно, например, понять, как будет работать молекула лекарства.
Самое интересное — когда ты получил результаты и пытаешься вычленить какой-то паттерн в том, что ты получил. Какую-то закономерность. У тебя есть критерии оценки. Бывает, ты обнаруживаешь, что паттерна нету. Но ты все равно пытаешься найти объяснение, почему это так. Конечно, когда ты попробовал кучу разного и понял, что ничего не работает, — наступает разочарование. Но я стараюсь такие дни забывать. Утром просыпаешься свежим и пытаешься пробовать снова.
О пути из математиков в биологи
Хотя сейчас моя работа происходит в основном на компьютере, я могу и электрод к мозгу мышки подключить, и провести спектроскопический эксперимент. Хотя начинал я с математики. Я учился в обычной школе в Петербурге и параллельно ходил в Центр компьютерных технологий в Аничковом дворце, учился программированию.
Как-то в очередной год я пришел записываться дальше, и преподаватель мне говорит: попробуй поступить в Лабораторию непрерывного математического образования. Я пришел туда, пообщался с директором. Прошел какое-то тестирование — было очень сложно. Но начал ходить. Было очень непривычно, что там уже занятия назывались парами — с 9 класса. И нам уже тогда давали научных руководителей. От этой школы я стал ездить на российские и международные научные конференции для школьников, участвовать в конкурсах. Например, ездил на знаменитую Intel ISEF в США и получил там премию Американского математического общества.
Дальше был матмех СПбГУ по специализации дифференциальные уравнения и динамические системы. Я даже поступил в аспирантуру, но что-то все-таки не удовлетворяло. И в итоге я стал подавать документы в другие университеты на другие специальности. Я слушал курсы на Coursera и других площадках, не связанных с математикой. Стал интересоваться биохимией, нейронными сетями. И так сошлись звезды, что мою заявку одобрили и мне предложили поступить на междисциплинарную программу в Университете Пердью в США. В первый год на этой программе каждый студент проходит четыре так называемых ротации — у разных научных руководителей. Смотрит, что ему больше по душе. Причем ты выбираешь их сам. Я успел позаниматься программированием в области биоинформатики, был в лаборатории изучения нейронной деятельности. Даже мышек резал. Но никаких зверств, все было очень гуманно.
В первый год на этой программе каждый студент проходит четыре так называемых ротации — у разных научных руководителей. Смотрит, что ему больше по душе. Причем ты выбираешь их сам. Я успел позаниматься программированием в области биоинформатики, был в лаборатории изучения нейронной деятельности. Даже мышек резал. Но никаких зверств, все было очень гуманно.
Я знаю, как аккуратно подключить электрод к мозгу мышки. А это очень непросто! Руководитель говорил, что у меня классно все получается. Но меня отвернул от этой работы… запах. Виварий, где мышки растут — он просто ужасно пахнул. После двух месяцев работы там мне казалось, что от меня постоянно пахнет виварием. Даже когда я менял одежду.
По итогу ротаций мой выбор пал на лабораторию химического факультета, которая занималась ЯМР-спектроскопией белков. В диссертации тематика сменилась с одной спектроскопии на другую — рентгеновскую. Защищался я уже во время пандемии, но не дистанционно. В большой аудитории на 100 человек были 4-5 членов комитета, рассаженные по разным концам комнаты. А потом я вернулся в Россию. Так получилось, что глава диссертационного комитета в Пердью, Николай Скрынников, также работал и здесь, в Лаборатории биомолекулярного ЯМР Института трансляционной биомедицины СПбГУ. Мы продолжаем развивать начатый еще во время работы над моей диссертацией проект, и сейчас я тружусь в его лаборатории.
А потом я вернулся в Россию. Так получилось, что глава диссертационного комитета в Пердью, Николай Скрынников, также работал и здесь, в Лаборатории биомолекулярного ЯМР Института трансляционной биомедицины СПбГУ. Мы продолжаем развивать начатый еще во время работы над моей диссертацией проект, и сейчас я тружусь в его лаборатории.
О конкуренции и сотрудничестве
Сегодня наука — это коллективная работа. Конечно, мы конкурируем — например, за гранты. Тебе интересна какая-то тема, ты плюс-минус знаешь, какие люди занимаются ей по всему миру. И ищешь пробелы в работах других. Такие пробелы, которые бы принесли пользу. И хочется опубликовать свой результат раньше других.
Бывает так, что за полгода до твоей публикации кто-то другой выпускает статью по той же теме. Приходит разочарование, но ты пытаешься искать выходы из этой ситуации. Смотришь: ага, вот они сделали это, а я сделаю что-то другое. Похожее, но все-таки в другую сторону. Но нет такого, что наука — это джунгли, и тебя обязательно съедят. Конкуренция есть, но можно кому-то написать, посоветоваться. Можно даже договориться: «Я подожду, пока вы опубликуетесь, а я потом выпущу свой результат». В мировой практике вообще приняты коллаборации — между учеными из разных стран, лабораторий. У нас, например, есть коллаборации с университетами в Европе, Америке и Азии, например, с Ратгерским университетом в США и Университетом Цинхуа в Китае.
Конкуренция есть, но можно кому-то написать, посоветоваться. Можно даже договориться: «Я подожду, пока вы опубликуетесь, а я потом выпущу свой результат». В мировой практике вообще приняты коллаборации — между учеными из разных стран, лабораторий. У нас, например, есть коллаборации с университетами в Европе, Америке и Азии, например, с Ратгерским университетом в США и Университетом Цинхуа в Китае.
Знаете, я вспоминаю лекцию Джеймса Уотсона, первооткрывателя ДНК, которая проходила в СПбГУ, по-моему, в 2017 году. Его спросили, что для ученого важно в целом. А его область практически моя. И он сказал три вещи… Не ручаюсь за точность, но постараюсь передать смысл. Первое: stay curious. А второе: be kind. Третью я не запомнил, но первые две абсолютно, на мой взгляд, точны. Оставаться голодным до результатов и быть приятным по отношению к своим коллегам. Без этого сложно выстроить коммуникации и существовать в сообществе.
О вкладе в науку и планах
Проблема рентгеновских моделей в том, что они статичны, а белок — система динамическая. В некоторых местах модель структуры белка получается «замыленной», и непонятно, что там происходит. Моя задача как раз состоит в том, чтобы позволить программе «додумать», как белок ведет себя в местах, пропущенных на рентгеновской картинке.
В некоторых местах модель структуры белка получается «замыленной», и непонятно, что там происходит. Моя задача как раз состоит в том, чтобы позволить программе «додумать», как белок ведет себя в местах, пропущенных на рентгеновской картинке.
Если этот инструмент найдет широкого пользователя — будет классно. Но это задача, так сказать, фундаментальная, внутренняя, научная. А если говорить о глобальных планах… Мне было бы интересно заниматься чем-то более прикладным, например, разработкой продуктов, которые уже в ближайшей перспективе могут помочь в борьбе с экологическими проблемами. Например, дизайном энзимов — это специальный класс белков — которые перерабатывают пластик. И дают окружающему нас миру ощутимую пользу уже сейчас. Я все-таки не зря так часто менял свои области деятельности. Мне интересно то, что можно применить к природе, к миру, который нас окружает. Вообще, страшно идти в новую отрасль. Но как-то перебарываешь себя — и шагаешь в пропасть. А как иначе?
О превосходстве машин над человеком — и наоборот
Нейросеть не дает понимание того, как она приходит к выводу. Она выдает какой-то результат, но не объясняет, почему. Если нейросеть написала картину или музыку — она может мне нравиться, а может не нравиться. Мне интересно, когда барабанщик не попал в такт. Я буду думать: это сделано намеренно или случайно? Когда это сделала нейросеть — это абсолютно точно намеренно. Это алгоритм. Мы вроде как понимаем, что он делает, но не понимаем, что там внутри.
Она выдает какой-то результат, но не объясняет, почему. Если нейросеть написала картину или музыку — она может мне нравиться, а может не нравиться. Мне интересно, когда барабанщик не попал в такт. Я буду думать: это сделано намеренно или случайно? Когда это сделала нейросеть — это абсолютно точно намеренно. Это алгоритм. Мы вроде как понимаем, что он делает, но не понимаем, что там внутри.
Машина — черный ящик, но и человек в каком-то смысле тоже. Бывает, у меня долго в голове не складывается какое-то решение. Или я не понимаю, как применять ту или иную формулу — когда-то, еще на матмехе, так было. И вдруг в какой-то момент я просыпаюсь и осознаю: вот зачем это нужно! Кто скажет, почему именно в этот момент все случилось? Что соединилось в мозге, под влиянием чего? Так что — да, человек тоже черный ящик. Но гораздо более интересный.
Роль и место смесей на основе частично гидролизованных белков в питании доношенных детей: пищеварительный комфорт и предупреждение аллергии | Мигачёва
1. Szajewska H, Horvath A. Meta-analysis of the evidence for a partially hydrolyzed 100% whey formula for the prevention of allergic diseases. Curr Med Res Opin. 2010;26(2):423–437. doi: 10.1185/03007990903510317.
Szajewska H, Horvath A. Meta-analysis of the evidence for a partially hydrolyzed 100% whey formula for the prevention of allergic diseases. Curr Med Res Opin. 2010;26(2):423–437. doi: 10.1185/03007990903510317.
2. Vandenplas Y, Alarcon P, Fleischer D, et al. Should Partial Hydrolysates Be Used as Starter Infant Formula? A Working Group Consensus. J Pediatr Gastroenterol Nutr. 2016;62(1):22–35. doi: 10.1097/MPG.0000000000001014.
3. Hochwallner H, Schulmeister U, Swoboda I, et al. Cow’s milk allergy: from allergens to new forms of diagnosis, therapy and prevention. Methods. 2014;66(1):22–33. doi:10.1016/j.ymeth.2013.08.005.
4. Rueter K, Prescott SL, Palmer DJ. Nutritional approaches for the primary prevention of allergic disease: An update. J Paediatr Child Health. 2015;51(10):962–969; quiz 968–969. doi: 10.1111/jpc.12951.
5. Monaci L, Tregoat V, van Hengel AJ, Anklam E. Milk allergens, their characteristics and their detection in food: A review. Eur Food Res Technol. 2006;223(2):149–179. doi: 10.1007/S00217-005-0178-8.
6. Jost R, Fritsche R, Pahud JJ. Reduction of milk protein allergenicity through processing. Bibl Nutr Dieta. 1991;(48):127–137. doi: 10.1159/000420043.
7. Vandenplas Y, Latiff AHA, Fleischer DM, et al. Partially hydrolyzed formula in non-exclusively breastfed infants: A systematic review and expert consensus. Nutrition. 2019;57:268–274. doi: 10.1016/j.nut.2018.05.018.
8. Bernard H, Creminon C, Yvon M, et al. Specificity of the human IgE response to the different purified caseins in allergy to cow’s milk proteins. Int Arch Allergy Immunol. 1998;115(3):235–244. doi: 10.1159/000023906.
Int Arch Allergy Immunol. 1998;115(3):235–244. doi: 10.1159/000023906.
9. Ruiter B, Tregoat V, M’rabet L, et al. Characterization of T cell epitopes in alphas1-casein in cow’s milk allergic, atopic and non-atopic children. Clin Exp Allergy. 2006;36(3):303–310. doi: 10.1111/j.1365-2222.2006.02436.x.
10. Spuergin P, Mueller H, Walter M, et al. Allergenic epitopes of bovine a S1-casein recognized by human IgE and IgG. Allergy. 1996; 51(5):306–312. doi: 10.1111/j.1398-9995.1996.tb04614.x.
11. Restani P, Gaiaschi A, Plebani A, et al. Cross-reactivity between milk proteins from different animal species. Clin Exp Allergy. 1999; 29(7):997–1004. doi: 10.1046/j.1365-2222.1999.00563.x.
12. Rozenfeld P, Docena GH, Anon MC, Fossati CA. Detection and identification of a soy protein component that cross-reacts with caseins from cow’s milk. Clin Exp Immunol. 2002;130(1):49–58. doi: 10.1046/j.1365-2249.2002.t01-1-01935.x.
Rozenfeld P, Docena GH, Anon MC, Fossati CA. Detection and identification of a soy protein component that cross-reacts with caseins from cow’s milk. Clin Exp Immunol. 2002;130(1):49–58. doi: 10.1046/j.1365-2249.2002.t01-1-01935.x.
13. Han GD, Matsuno M, Ito S, et al. Meat allergy: investigation of potential allergenic proteins in beef. Biosci Biotechnol Biochem. 2000;64(9):1887–1895. doi: 10.1271/bbb.64.1887.
14. Vicente-Serrano J, Caballero ML, Rodriguez-Perez R, et al. Sensitization to serum albumins in children allergic to cow’s milk and epithelia. Pediatr Allergy Immunol. 2007;18(6):503–507. doi: 10.1111/j.1399-3038.2007.00548.x.
15. Anderson PJ, Brooks CL, Berliner LJ. Functional identification of calcium binding residues in bovine alpha-lactalbumin. Biochemistry. 1997;36(39):11648–11654. doi: 10.1021/bi9709598.
Biochemistry. 1997;36(39):11648–11654. doi: 10.1021/bi9709598.
16. Svensson M, Hakansson A, Mossberg AK, et al. Conversion of alpha-lactalbumin to a protein inducing apoptosis. Proc Natl Acad Sci U S A. 2000;97(8):4221–4226. doi: 10.1073/pnas.97.8.4221.
17. Permyakov EA, Berliner LJ. Alpha-Lactalbumin: structure and function. FEBS Lett. 2000;473(3):269–274. doi: 10.1016/s0014-5793(00)01546-5.
18. Maynard F, Jost R, Wal JM. Human IgE binding capacity of tryptic peptides from bovine alpha-lactalbumin. Int Arch Allergy Immunol. 1997;113(4):478–488. doi: 10.1159/000237625.
19. Suutari TJ, Valkonen KH, Karttunen TJ, et al. IgE cross reactivity between reindeer and bovine milk beta-lactoglobulins in cow’s milk allergic patients. J Investig Allergol Clin Immunol. 2006; 16(5):296–302.
J Investig Allergol Clin Immunol. 2006; 16(5):296–302.
20. Aymard P, Durand D, Nicolai T. The effect of temperature and ionic strength on the dimerisation of beta-lactoglobulin. Int J Biol Macromol. 1996;19(3):213–221. doi: 10.1016/0141-8130(96)01130-0.
21. Adler-Nissen J. Determination of degree of hydrolysis of food protein hydrolysates by trinitrobenzenesulfonic acid. J Agric Food Chem. 1979;27(6):1256–1279. doi: 10.1021/jf60226a042.
22. El-Agamy EI. The challenge of cow milk protein allergy. Small Rumin Res. 2007;68(1–2):64–72. doi: 10.1016/j.smallrumres.2006.09.016.
23. Schmidt DG, Meijer RJ, Slangen CJ, van Beresteijn EC. Raising the pH of the pepsin-catalysed hydrolysis of bovine whey proteins increases the antigenicity of the hydrolysates. Clin Exp Allergy. 1995; 25(10):1007–1017. doi: 10.1111/j.1365-2222.1995.tb00404.x.
24. Ehn B-M, Ekstrand B, Bengtsson U, Ahlstedt S. Modification of IgE-binding during heat processing of the cow’s milk allergen betalactoglobulin. J Agric Food Chem. 2004;52(5):1398–1403. doi: 10.1021/jf0304371.
25. Fritsche R. The role of immune tolerance in allergy prevention. Nestle Nutr Workshop Ser Pediatr Program. 2005;56:1–9; discussion 9–14. doi: 10.1159/000086231.
26. EFSA NDA Panel (EFSA Panel on Dietetic Products, Nutrition and Allergies). Scientific Opinion on the essential composition of infant and follow-on formulae. EFSA J. 2014;12(7):3760. doi: 10.2903/J.EFSA.2014.3760.
27. Salvatore S, Vandenplas Y. Hydrolyzed Proteins in Allergy. In: Bhatia J, Shamir R, Vandenplas Y, eds. Protein in Neonatal and Infant Nutrition: Recent Updates. Nestle Nutr Inst Workshop Ser. 2016; 86:11–27. doi: 10.1159/000442699.
28. Greer FR, Sicherer SH, Burks AW, et al. Effects of early nutritional interventions on the development of atopic disease in infants and children: the role of maternal dietary restriction, breastfeeding, timing of introduction of complementary foods, and hydrolyzed formulas. Pediatrics. 2008;121(1):183–191. doi: 10.1542/peds.2007-3022.
29. Vandenplas Y, Bathia J, Shamir R, et al. Hydrolyzed formulas for allergy prevention. J Pediatr Gastroenterol Nutr. 2014;58(5): 549–552. doi: 10.1097/MPG.0000000000000318.
30. Tsabouri S, Douros K, Priftis KN. Cow’s milk allergenicity. Endocr Metab Immune Disord Drug Targets. 2014;14(1):16–26. doi: 10.2174/1871530314666140121144224.
31. Terracciano L, Isoardi P, Arrigoni S, et al. Use of hydrolysates in the treatment of cow’s milk allergy. Ann Allergy Asthma Immunol. 2002;89(6 Suppl 1):86–90. doi: 10.1016/s1081-1206(10)62130-8.
32. Pecquet S, Bovetto L, Maynard F, Fritsche R. Peptides obtained by tryptic hydrolysis of bovine beta-lactoglobulin induce specific oral tolerance in mice. J Allergy Clin Immunol. 2000;105(3):514–521. doi: 10.1067/mai.2000.103049.
33. Lozano-Ojalvo D, Berin C, Tordesillas L. Immune basis of allergic reactions to food. J Investig Allergol Clin Immunol. 2019;29(1): 1–14. doi: 10.18176/jiaci.0355.
34. Kumfer AM, Commins SP. Primary prevention of food allergy. Curr Allergy Asthma Rep. 2019;19(1):7. doi: 10.1007/s11882-019-0838-6.
35. Nowak-Wegrzyn A, Chatchatee P. Mechanisms of tolerance induction. Ann Nutr Metab. 2017;70(Suppl 2):7–24. doi: 10.1159/000457915.
36. Rezende RM, Weiner HL. History and mechanisms of oral tolerance. Semin Immunol. 2017;30:3–11. doi: 10.1016/j.smim.2017.07.004.
37. Смолкин Ю.С., Грищенко Е.А. Современные представления о формировании оральной толерантности (Часть 1) // Аллергология и иммунология в педиатрии. — 2015. — Т. 43. — № 4. — С. 29–35.
38. Berin MC, Sampson HA. Mucosal immunology of food allergy. Curr Biol. 2013;23(9):R389–R400. doi: 10.1016/j.cub.2013.02.043.
39. Tanoue T, Atarashi K, Honda K. Development and maintenance of intestinal regulatory T cells. Nat Rev Immunol. 2016;16(5): 295–309. doi: 10.1038/nri.2016.36.
40. Hadis U, Wahl B, Schulz O, et al. Intestinal tolerance requires gut homing and expansion of FoxP3+ regulatory T cells in the lamina propria. Immunity. 2011;34(2):237–246. doi: 10.1016/j.immuni.2011.01.016.
41. Torgerson TR, Linane A, Moes N, et al. Severe food allergy as a variant of IPEX syndrome caused by a deletion in a noncoding region of the FOXP3 gene. Gastroenterology. 2007;132(5):1705–1717. doi: 10.1053/j.gastro.2007.02.044.
42. Jaensson E, Uronen-Hansson H, Pabst O, et al. Small intestinal CD103+ dendritic cells display unique functional properties that are conserved between mice and humans. J Exp Med. 2008; 205(9):2139–2149. doi: 10.1084/jem.20080414.
43. Klebanoff CA, Spencer SP, Torabi-Parizi P, et al. Retinoic acid controls the homeostasis of precDC-derived splenic and intestinal dendritic cells. J Exp Med. 2013;210(10):1961–1976. doi: 10.1084/jem.20122508.
44. Tsuji NM, Kosaka A. Oral tolerance: intestinal homeostasis and antigen-specific regulatory T cells. Trends Immunol. 2008;29(11): 532–540. doi: 10.1016/j.it.2008.09.002.
45. Украинцев С.Е. Профилактика аллергии: от иммунологии беременности до вскармливания детей первых месяцев жизни // Вопросы современной педиатрии. — 2016. — Т. 15. — № 6. — С. 604–609. doi.org/10.15690/vsp.v15i6.1658.
46. Wang Y, Allen KJ, Koplin JJ. Dietary intervention for preventing food allergy in children. Curr Opin Pediatr. 2017;29(6):704–710. doi: 10.1097/MOP.0000000000000552.
47. Grulee C, Sanford H. The influence of breast and artificial feeding on infantile eczema. J Pediatr. 1936;9(2):223–225. doi: 10.1016/S0022-3476(36)80058-4.
48. Lucas A, Brooke O, Morley R, et al. Early diet of preterm infants and development of allergic or atopic disease: randomized prospective study. BMJ. 1990;300(6728):837–840. doi: 10.1136/bmj.300.6728.837.
49. Gdalevich M, Mimouni D, David M. Breastfeeding and the onset of atopic dermatitis in childhood: a systematic review and metaanalysis of prospective studies. J Am Acad Dermatol. 2001;45(4): 520–527. doi: 10.1067/mjd.2001.114741.
50. Von Berg A, Koletzko S, Filipiak-Pittroff B, et al. Certain hydrolyzed formulas reduce the incidence of atopic dermatitis but not that of asthma: three-year results of the German Infant Nutritional Intervention Study. J Allergy Clin Immunol. 2007;119(3): 18–25. doi: 10.1016/j.jaci.2006.11.017.
51. Von Berg A, Filipiak-Pittroff B, Kramer U, et al. Allergies in highrisk schoolchildren after early intervention with cow’s milk protein hydrolysates: 10-year results from the German Infant Nutritional Intervention (GINI) study. J Allergy Clin Immunol. 2013;131(6): 1565–1573. doi: 10.1016/j.jaci.2013.01.006.
52. Osborn DA, Sinn J. Formulas containing hydrolysed protein for prevention of allergy and food intolerance in infants. Cochrane Database Syst Rev. 2006;18(4):CD003664. doi: 10.1002/14651858.CD003664.pub3.
53. Szajewska H, Horvath A. A partially hydrolyzed 100% whey formula and the risk of eczema and any allergy: an updated metaanalysis. World Allergy Organ J. 2017;26;10(1):27. doi: 10.1186/s40413-017-0158-z.
54. Von Berg A, Filipiak-Pittroff B, Schulz H, et al. Allergic manifestation 15 years after early intervention with hydrolyzed formulas — the GINI Study. Allergy. 2016;71(2):210–219. doi: 10.1111/all.12790.
55. Spieldenner J, Belli D, Dupont C, et al. Partially hydrolysed 100% whey-based infant formula and the prevention of atopic dermatitis: comparative pharmacoeconomic analyses. Ann Nutr Metab. 2011;59(Suppl 1):44–52. doi: 10.1159/000334232.
56. Botteman MF, Bhanegaonkar AJ, Horodniceanu EG, et al. Economic value of using partially hydrolysed infant formula for risk reduction of atopic dermatitis in high-risk, not exclusively breastfed infants in Singapore. Singapore Med J. 2018;59(8):439–448. doi: 10.11622/smedj.2017113.
57. Намазова–Баранова Л.С., Макарова С.Г., Новик Г.А., Вишнева Е.А. Национальные клинические рекомендации по оказанию медицинской помощи детям с аллергией к белкам коровьего молока. Краткий обзор документа // Российский аллергологический журнал. — 2017. — Т. 14. — № 2. — С. 55–65.
58. Первичная профилактика аллергии у детей. Согласительный документ Ассоциации детских аллергологов и иммунологов России. — М.; 2010.
59. Wood RA, Sicherer SH, Vickery BP, et al. The natural history of milk allergy in an observational cohort. J Allergy Clin Immunol. 2013; 131(3):805–812. doi: 10.1016/j.jaci.2012.10.060.
60. Elizur A, Rajuan N, Goldberg MR, et al. Natural course and risk factors for persistence of IgE-mediated cow’s milk allergy. J Pediatr. 2012;161(3):482–487.e1. doi: 10.1016/j.jpeds.2012.02.028.
61. Sackesen C, Altintas DU, Bingol A, et al. Current Trends in Tolerance Induction in Cow’s Milk Allergy: From Passive to Proactive Strategies. Front Pediatr. 2019;7:372. doi: 10.3389/fped.2019.00372.
62. Fiocchi A, Brozek J, Schunemann H, et al. World Allergy Organization (WAO) Diagnosis and Rationale for Action against Cow’s Milk Allergy (DRACMA) Guidelines. Pediatr Allergy Immunol. 2010; 21(Suppl 21):1–125. doi: 10.1111/j.1399-3038.2010.01068.x.
63. Vandenplas Y. Prevention and Management of Cow’s Milk Allergy in Non-Exclusively Breastfed Infants. Nutrients. 2017;9(7):731. doi: 10.3390/nu9070731.
64. Luyt D, Ball H, Makwana N, et al. BSACI guideline for the diagnosis and management of cow’s milk allergy. Clin Exp Allergy. 2014;44(5):642–672. doi: 10.1111/cea.12302.
65. Brozek JL, Terracciano L, Hsu J, et al. Oral immunotherapy for IgE-mediated cow’s milk allergy: a systematic review and meta-analysis. Clin Exp Allergy. 2012;42(3):363–374. doi: 10.1111/j.1365-2222.2011.03948.x.
66. D’Auria E, Salvatore S, Pozzi E, et al. Cow’s Milk Allergy: Immunomodulation by Dietary Intervention. Nutrients. 2019;11(6): 1399. doi: 10.3390/nu11061399.
67. Berni Canani R, Nocerino R, Terrin G, et al. Formula selection for management of children with cow’s milk allergy influences the rate of acquisition of tolerance: a prospective multicenter study. J Pediatr. 2013;163(3):771–777. doi: 10.1016/j.jpeds.2013.03.008.
68. Kim AR, Kim HS, Nam ST, et al. Mesenteric IL-10-producing CD5+ regulatory B cells suppress cow’s milk casein-induced allergic responses in mice. Sci Rep. 2016;6:19685. doi: 10.1038/srep19685.
69. Kiewiet MBG, Dekkers R, Gros M, et al. Toll-like receptor mediated activation is possibly involved in immunoregulating properties of cow’s milk hydrolysates. PLoS One. 2017;12(6): e0178191. doi: 10.1371/journal.pone.0178191.
70. Inuo C, Tanaka K, Suzuki S, et al. Oral Immunotherapy Using Partially Hydrolyzed Formula for Cow’s Milk Protein Allergy: A Randomized, Controlled Trial. Int Arch Allergy Immunol. 2018; 177(3):259–268. doi: 10.1159/000490804.
71. Inuo C, Tanaka K, Nakajima Y, et al. Tolerability of partially and extensively hydrolysed milk formulas in children with cow’s milk allergy. Asia Pac J Clin Nutr. 2019;28(1):49–56. doi: 10.6133/apjcn.201903_28(1).0008.
72. Astier C, Morisset M, Roitel O, et al. Predictive value of skin prick tests using recombinant allergens for diagnosis of peanut allergy. J Allergy Clin Immunol. 2006;118:250–256. doi: 10.1016/j.jaci.2006.04.053.
73. Vandenplas Y, Al-Hussaini B, Al-Mannaei K, et al. Prevention of Allergic Sensitization and Treatment of Cow’s Milk Protein Allergy in Early Life: The Middle-East Step-Down Consensus. Nutrients. 2019;11(7).1444. doi: 10.3390/nu11071444.
74. Vandenplas Y, Benninga M, Broekaert I, et al. Functional gastrointestinal disorder algorithms focus on early recognition, parental reassurance and nutritional strategies [published correction appears in Acta Paediatr. 2016;105(8):984]. Acta Paediatr. 2016; 105(3):244–252. doi:10.1111/apa.13270.
75. Rosen R, Vandenplas Y, Singendonk M, et al. Pediatric Gastroesophageal Reflux Clinical Practice Guidelines: Joint Recom mendations of the North American Society for Pediatric Gastroenterology, Hepatology, and Nutrition and the European Society for Pediatric Gastroenterology, Hepatology, and Nutrition. J Pediatr Gastroenterol Nutr. 2018;66(3):516–554. doi: 10.1097/MPG.0000000000001889.
76. Vandenplas Y, Hauser B, Salvatore S. Functional Gastrointestinal Disorders in Infancy: Impact on the Health of the Infant and Family. Pediatr Gastroenterol Hepatol Nutr. 2019;22(3):207–216. doi: 10.5223/pghn.2019.22.3.207.
77. Salvatore S, Abkari A, Cai W, et al. Review shows that parental reassurance and nutritional advice help to optimise the management of functional gastrointestinal disorders in infants. Acta Paediatr. 2018;107(9):1512–1520. doi: 10.1111/apa.14378.
78. Salvatore S, Savino F, Singendonk M, et al. Thickened infant formula: What to know. Nutrition. 2018; 49:51–56. doi: 10.1016/j.nut.2017.10.010.
79. Vandenplas Y, Leluyer B, Cazaubiel M, et al. Double-blind comparative trial with two antiregurgitation formulae. J Pediatr Gastroenterol Nutr. 2013;57(3):389–393. doi: 10.1097/MPG.0b013e318299993e.
80. Indrio F, Riezzo G, Giordano P, et al. Effect of a Partially Hydrolysed Whey Infant Formula Supplemented with Starch and Lactobacillus reuteri DSM 17938 on Regurgitation and Gastric Motility. Nutrients. 2017;9(11):1181. doi: 10.3390/nu9111181.
81. Gonzalez-Bermudez CA, Frontela-Saseta C, Lopez-Nicolas R, et al. Effect of adding different thickening agents on the viscosity properties and in vitro mineral availability of infant formula. Food Chem. 2014;159:5–11. doi: 10.1016/j.foodchem.2014.02.168.
82. Horvath A, Dziechciarz P, Szajewska H. The effect of thickened feed interventions on gastroesophageal reflux in infants: systematic review and meta-analysis of randomized, controlled trials. Pediatrics. 2008;122(6):e1268–1277. doi: 10.1542/peds.2008-1900.
83. Макарова Е.Г., Клепикова Т.В., Украинцев С.Е. Минимальные расстройства пищеварения: когда проблемы есть, а лечить еще рано // Вопросы современной педиатрии. — 2019. — Т. 18. — № 4. — С. 247–256. doi: 10.15690/vsp.v18i4.2041.
84. Gutierrez-Castrellon P, Indrio F, Bolio-Galvis A, et al. Efficacy of Lactobacillus reuteri DSM 17938 for infantile colic: Systematic review with network meta-analysis. Medicine (Baltimore). 2017; 96(51):e9375. doi: 10.1097/MD.0000000000009375.
85. Salanti G, Ades AE, Ioannidis JP. Graphical methods and numerical summaries for presenting results from multiple-treatment metaanalysis: an overview and tutorial. J Clin Epidemiol. 2011;64(2): 163–171. doi: 10.1016/j.jclinepi.2010.03.016.
86. Красавина Н.А., Кабанова Н.К., Перминова О.А. и др. Возможности диетологической коррекции сочетанных форм функциональных расстройств пищеварения у детей первого года жизни: результаты клинической апробации нового продукта в открытом сравнительном проспективном исследовании // Вопросы детской диетологии. — 2017. — Т. 15. — № 6. — С. 18–24. doi: 10.20953/1727-5784-2017-6-18-24.
87. Tabbers MM, Di Lorenzo C, Berger MY, et al. Evaluation and treatment of functional constipation in infants and children: evidence-based recommendations from ESPGHAN and NASPGHAN. J Pediatr Gastroenterol Nutr. 2014;58(2):265–281. doi: 10.1097/MPG.0000000000000266.
88. Czerkies LA, Kineman BD, Cohen SS, et al. A Pooled Analysis of Growth and Tolerance of Infants Exclusively Fed Partially Hydrolyzed Whey or Intact Protein-Based Infant Formulas. Int J Pediatr. 2018; 2018:4969576. doi: 10.1155/2018/4969576.
89. Vandenplas Y, Munasir Z, Hegar B, et al. A perspective on partially hydrolyzed protein infant formula in nonexclusively breastfed infants. Korean J Pediatr. 2019;62(5):149–154. doi: 10.3345/kjp.2018.07276.
Группа термодинамики белка | Институт белка РАН
Основные научные достижения
Одним из первых крупных достижений группы в начале 70-х годов была разработка принципиально новых высокочувствительных приборов для исследования разбавленных растворов биологических макромолекул. По мере усложнения объектов исследования возникла необходимость в разработке и компьютерной реализации алгоритмов анализа калориметрических данных, в частности полученных для мультидоменных белков и нуклеиновых кислот. Были созданы программы, которые до сих пор широко используются для сбора и анализа калориметрических данных. Эти методические прорывы позволили нам провести многочисленные исследования термодинамики разворачивания малых глобулярных белков, ДНК, мультидоменных и фибриллярных белков, РНК и мембран, то есть практически всех классов биологических макромолекул. Не случайно результаты наших исследований были не только опубликованы в оригинальных работах, но и легли в основу многочисленных обзорных статей. Не так давно нами был разработан уникальный сканирующий микрокалориметр способный работать при давлениях до 5000 атм. Это позволило нам начать новое направление исследований – изучение физических основ объемных изменений при конформационных превращениях макромолекул.
К основным принципиальным результатам, полученным группой за последние десятилетия, надо отнести:
- Создание теории обобщенных переходов между двумя состояниями. Этот аппарат устанавливает взаимно однозначное соответствие между любым комплексным числом и определенным пиком теплопоглощения, что во многих случаях упрощает структурную интерпретацию термодинамических данных.
- Изучение белков, структура которых неравновесна и сохраняется только в силу кинетических причин. Доказательство существования “кора” (части структуры, определяющей стабильность) у таких белков.
- Изучение структуры δ-эндотоксина в составе липидной мембраны. Создание модели взаимодействия токсина с мембраной.
- Прояснение общих принципов влияния органических растворителей на стабильность белков.
- Выяснение некоторых принципов, определяющих структуры многозаходных α-суперспиральных белков и конструирование суперспиральных пептидов с заданными свойствами.
- Создание уникального сканирующего микрокалориметра высокого давления и разработка методов анализа получаемых с его помощью данных.
- Изучение объемных изменений фосфолипидов и модельных полимеров при термотропных конформационных переходах.
Перспективы исследований
Работы, которые будут проводиться в группе в ближайшие годы, объединены общей фундаментальной целью – изучить механизмы самоорганизации структур основных биологически важных макромолекул и их комплексов, оценить роль различных физико-химических факторов в этих процессах, природу и относительный вклад различных сил. Особый интерес будет представлять изучение роли растворителя в процессах самоорганизации. Исследования будут проводиться с использованием широкого набора физико-химических методов. Особая роль будет отведена термодинамическим методам, в том числе методу сканирующей микрокалориметрии при высоком давлении.
Общий белок в сыворотке крови, total protein со скидкой до 50% в Lab4U
Интерпретация результатов исследования
«Общий белок»
Интерпретация результатов анализов носит информационный характер, не является диагнозом и не
заменяет консультации врача. Референсные значения могут отличаться от указанных в
зависимости от используемого оборудования, актуальные значения будут указаны на бланке
результатов.
Изменения концентрации общего белка в сыворотке крови могут быть как абсолютными, так и относительными (нарушение водного баланса).
Единица измерения: г/л
Референсные значения:
|
Возраст |
Общий белок, г/л |
|
1 — 14 дней |
52 — 79 |
|
15 дн -1 год |
43 — 68 |
|
2 — 6 лет |
59 — 72 |
|
7 — 9 лет |
62 — 74 |
|
10 — 15 лет |
63 — 77 |
|
>15 лет |
66 — 83 |
|
Беременные женщины |
|
|
I триместр |
62 — 76 |
|
II триместр |
57 — 69 |
|
III триместр |
56 — 67 |
Повышение:
Гиперпротеинемия абсолютная:
- Острые и хронические инфекции.
- Миеломная болезнь (плазмоцитома).
- Аутоиммунные заболевания.
- Парапротеинемические гемобластозы.
- Саркоидоз.
- Цирроз печени.
- Болезнь Вальденстрема.
- Болезни тяжёлых цепей.
- Лимфогранулематоз.
- Хронический полиартрит.
Гиперпротеинемия относительная:
- Перитонит.
- Тяжёлые ожоги.
- Кишечная непроходимость.
- Рвота, понос.
- Несахарный диабет.
- Хронический нефрит.
- Усиленное потоотделение.
Снижение:
Гипопротеинемия абсолютная:
- Снижение синтеза белка (цирроз печени, гепатиты, интоксикации).
- Врождённые нарушения синтеза отдельных белков (болезнь Вильсона-Коновалова, анальбуминемия, диспротеинемии).
- Повышенный распад белка в организме (гипертиреоз, обширные ожоги, послеоперационные состояния, травмы, длительные лихорадочные состояния, опухоли).
- Кровотечения.
- Повышенное выделение белка с мочой (нефротический синдром, заболевания почек, сахарный диабет).
- Недостаточное поступление белка с пищей (голодание, недоедание).
Гипопротеинемия относительная:
- Гипергидратация.
Белка обыкновенная | Барнаульский зоопарк «Лесная сказка»
Белки — очень шустрые и безмерно любопытные зверьки. Каждый посетитель, задержавшийся у их вольере, становится объектом беличьего внимания. Наши белочки дружелюбны и вполне хорошо уживаются с морскими свинками, с которыми делят вольер в летний период. Любят орехи, фрукты, ягоды. Запасов никогда не делают, так как корм получают ежедневно.
О виде животного
Белки — род некрупных грызунов семейства беличьих. Род включает в себя порядка 30 различных видов, которые населяют практически всю планету от Азии до Северной Америки.
Характерные особенности белок:
- Обыкновенная белка — единственный представитель семейства, обитающий на просторах России.
- У белки круглая голова и большие черные глаза-бусины.
- Окрас беличьей шубки меняется ежесезонно от рыжего до серого цветов.
- Пальцы белки оканчиваются острыми когтями.
- Задние лапы белки намного длиннее передних.
Вес этих мелких грызунов редко превышает 350 граммов, при этом белки обладают длинным стройным телом до 30 сантиметров длиной и роскошным пушистым хвостом. Для лучшего осязания природа наградила белок сверхчувствительными вибриссами на мордочке, животе и передних лапках. Цвет шерсти белок и размер их тел различается в зависимости от места обитания: обитатели равнин существенно мельче своих горных собратьев.
Белки населяют леса, преимущественно хвойные, поскольку именно там легче всего добывать любимую пищу — семена. В южных регионах России белки приспособились к жизни в плодовых садах и даже виноградниках. Живя на деревьях, белки обладают невероятной юркостью и подвижностью. Для них не составляет никакого труда совершить прыжок длиной 4 метра, при этом «рулем» им служит хвост. Большую часть дня белки посвящают поискам пропитания, происходит это утром и вечером. Белки — любители орехов и желудей, а также ягод, клубней и даже грибов. Бывает, не гнушаются перекусить личинками, насекомыми и птенцами.
В период размножения самку белки окружает стайка самцов, которые всячески демонстрируют друг другу и возлюбленной свое превосходство: издают урчащие звуки, бьют лапами ветки, гоняются друг за другом. Беременность у самки продолжается до 38 дней, на свет появляются до 10 слепых крохотных бельчат. Мать-белка кормит их молоком до 50 дней. Половозрелый возраст белки — 9 месяцев. В условиях дикой природы белка уже в 4 года считается отжившей свое, однако в неволе срок ее жизни увеличивается до 12 лет. Численность этих зверьков высока, поэтому вымирание в ближайшие годы им точно не грозит.
Интересные факты о белках:
- Белки — самые чистоплотные среди грызунов: самцы зачастую тратят больше времени на уход за собой, чем самки.
- Белки обладают великолепным обонянием: самец чует запах самки на расстоянии 1,5 километров.
- Новорожденные беличьи детеныши весят всего 30 граммов.
- Несмотря на стройность, за 7 дней белка способна съесть количество еды, равное собственному весу.
- Взрослые белки живут поодиночке.
Избыточное потребление белка приводит к подагре, определить суточную норму в Москве
Сколько белка нужно человеку?
На этот вопрос нет ответа, одинакового для всех. Потребность в белке возрастает при интенсивном росте (у детей), во время беременности, при некоторых патологических состояниях (ожогах, заболеваниях легких). При отдельных заболеваниях белок следует ограничивать, так как у организма снижены возможности полной утилизации продуктов его распада, в первую очередь — аммиака (мочевины).
Зачем нужен белок?
Любой белок — это комбинация двадцати аминокислот. Мы употребляем в пищу растительные или животные белки только для того, чтобы наш организм смог построить из них собственные. При пищеварении белки расщепляются на аминокислоты. И эти аминокислоты используются для построения клеток, синтеза гормонов и многих других важных веществ, без которых жизнь человека становится невозможной.
К чему приводит избыточное потребление белка?
В организме белок разлагается на воду, аммиак и глюкозу. Глюкоза — источник энергии, вода полезна, а аммиак — яд. Поэтому в печени связывается особыми ферментами и превращается в мочевину. Мочевина выводится почками. Если поступление белка значительно превышает норму, железы внутренней секреции работают с большим напряжением, повышается нервная возбудимость, страдают печень, почки, суставы, нарушается обмен веществ.
Обычно с избыточным потреблением белка связывают два заболевания: мочекаменную болезнь и подагру. Однако они вызваны нарушением другого вещества — мочевой кислоты, которая накапливается в организме в результате нарушения пуринового обмена. Ошибка простительна. Она объясняется тем, что и белок, и пурины в большом количестве содержатся в мясе и рыбе. То есть, употребляя без меры мясо (особенно мясные деликатесы!), человек получает не только белки, но и пурины.
Мочекаменная болезнь почек
Соли мочевой кислоты кристаллизуются в канальцах почек. В результате образуются камни, которые затрудняют отток мочи. Пока конкременты маленькие, человек ощущает незначительный дискомфорт в области поясницы. По мере их роста дискомфорт усиливается. В случае, когда камень перекрывает проток возникает почечная колика. Её симптомы резко выражены:
- резкая боль в пояснице;
- человек мечется, меняя положение тела в поисках позы, в которой боль станет хоть чуть меньше.
Это состояние требует неотложных мер. Нужно вызвать бригаду скорой помощи и следовать указаниям доктора.
Подагра
Хроническое обменное заболевание, которое развивается при поражении почек или чрезмерной выработке пуринов. Мочевая кислота накапливается — появляется подагра.
Характерные признаки:
- артрит, чаще поражаются мелкие суставы;
- асимметрия поражений;
- острые приступы сменяются ремиссиями.
Лечение подагры проводится в стационаре. Проводится медикаментозная терапия, подбирается диета.
Диета при подагре, мочекаменной болезни
Обычно рекомендуется шестой диетический стол, ограничивающий употребление мяса и соли. Исключаются мясные деликатесы, субпродукты, бобовые, щавель, шпинат, крепкие бульоны.
Рекомендуется увеличить в рационе долю растительного белка, усилить питьевой режим.
Определение суточной нормы белка в Москве
В клинике MAJOR CLINIC ведет прием опытный диетолог высокой квалификации. Вы можете сделать своё питание правильным и полезным, записавшись на консультацию к доктору. После изучения ваших особенностей обмена и двигательной активности, врач составит для вас индивидуальный рацион и примерное меню. Придерживаясь рационального питания, вы очень скоро почувствуете себя значительно лучше. Ощутите прилив энергии, станете более активным и жизнерадостным человеком.
Basic Protein — обзор
I СТРУКТУРА ОСНОВНОГО БЕЛКА
Основные белки бычьего (Eylar et al ., 1971; Brostoff et al ., 1974b) и человеческого (Carnegie, 1971) миелина ЦНС имеют был полностью упорядочен. Определение частичной последовательности и аминокислотный анализ триптических пептидов других основных белков млекопитающих (Shapira et al ., 1971; Martenson et al ., 1971a; Kornguth et al ., 1972; Brostoff & Eylar, 1972; Martenson и др. ., 1972) показали, что, за одним заметным исключением (см. Ниже), первичных структур этих белков очень похожи на структуру бычьего или человеческого белка. На рис. 1 последовательность бычьего основного белка проиллюстрирована с точки зрения положений (1) остатков, заряженных при нейтральном pH, (2) остатков, которые являются гидрофобными или слабо гидрофильными (отрицательно гидрофобными), (3) гистидиловых остатков, и (4) пролильных остатков. Все заряженные остатки сильно гидрофильны и неизменно находятся на поверхности белка.Шкала гидрофобности показывает относительную тенденцию каждого остатка избегать водной среды и располагаться в неполярной области (внутренней части) белка. Слабогидрофильные остатки (Gln, Asn, Ser), слабогидрофобные остатки Thr и His и сильно гидрофобные остатки Tyr и Trp могут участвовать в водородном связывании. Гистидиловые остатки могут быть либо сильно гидрофильными (заряженными), либо слабогидрофобными (незаряженными) в зависимости от их точного расположения в белке и, как следствие, значений имидазолила pK a . Остатки пролила завершают α-спиральные сегменты полипептидной цепи (если такие сегменты присутствуют) и меняют ее направление. Четыре остатка в основном белке подвергаются модификации in vivo : Ala-1 ацетилирован, Arg-106 частично метилирован, а Thr-97 и Ser-164 частично фосфорилированы. Особый интерес представляет последовательность трипролина (остатки 98–100), которая может вызвать разворот пептидной цепи (Hagopian et al ., 1971), частое появление заряженных остатков, а также высокогидрофобных остатков. в тандеме низкая гидрофобность области между остатками 43 и 85 и высокая концентрация лизила и низкая концентрация аргинильных остатков в С-концевой области между Trp-115 и Gly-156.В меньшем из двух основных белков крысы (S-белок ) отсутствует область между Trp-115 и Gly-156. Ряд отличительных особенностей этой последовательности будет подробно рассмотрен ниже.
Рис. 1. Аминокислотная последовательность основного белка бычьего миелина, иллюстрирующая распределение остатков в соответствии с их гидрофобным (положительное направление) или гидрофильным (отрицательное направление) характером. Незаштрихованные столбики обозначают гистидильные остатки, которые могут быть либо слабо гидрофобными (незаряженными), либо сильно гидрофильными (заряженными).Заряженным остаткам даются стандартные однобуквенные обозначения: Arg, R; Lys, К; Клей; Asp, D. Пролильные остатки показаны сплошными кружками; остатки глицила, не имеющие боковой цепи, выглядят как пробелы. Показаны положения метильной (Me) и фосфатной (P) групп, а также области, удаленной в S-белке крысы. Относительная гидрофобность согласно Nozaki & amp; Танфорд (1971) следующие: Джин и Сер, −3; Asn, −2; Его (незаряженный) и Ала, +5; Met, +13; Вал, +15; Лей и Иль, +18; Тюр, +23; Phe, +25; Трп, +34.В соответствии с этим представлением можно идентифицировать все остатки, за исключением пар Ser, Gin и Leu, Ile.
За исключением относительно коротких последовательностей, кажется, нет очевидной регулярности в расположении остатков какого-либо конкретного типа. Фактически, нерегулярная, но явно специфическая линейная последовательность аминокислот сама по себе, по-видимому, не имеет никакого смысла с точки зрения функции in vivo белка, если линейная последовательность не была преобразована в какую-то трехмерную структуру. .Ряд исследований показал, что изолированный основной белок в водном растворе при нейтральном или близком к нейтральному pH имеет мало уникальной вторичной или третичной структуры , если вообще имеет ее, и, по-видимому, сильно разупорядочен и расширен (Palmer & Dawson, 1969; Eylar & Томпсон, 1969; Оширо и Эйлар, 1970; Чао и Эйнштейн, 1970; Энтони и Москарелло, 1971; Кригбаум и Хсу, 1975; Либес и др. ., 1975; Джонс и Рамсби, 1975). В этом отношении выделенный основной белок миелина очень похож на другой основной белок, цитохром с, после того, как его гемовая группа была химически удалена.Апо-цитохром c имеет сильно дезорганизованную конформацию «случайной спирали» (Stellwagen et al. ., 1972; Fisher et al. ., 1973). Нативный цитохром c, напротив, представляет собой компактный глобулярный белок с очень высокой степенью вторичной и третичной структуры (Dickerson et al ., 1971). Какие бы конформационные детерминанты ни существовали в коротких участках полипептидной цепи апо-цитохрома с, их потенциал, по-видимому, не может быть выражен до тех пор, пока дальнодействующие взаимодействия между различными последовательностями внутри белка не будут вызваны специфическим ковалентным связыванием гема (Fisher et al ., 1973). По аналогии, похоже, не было бы оснований для a priori предполагать, что конформация основного белка после специфического взаимодействия с другими составляющими миелина in vivo не будет компактной и высокоструктурированной, хотя и не обязательно спиральной. Фактически, некоторая структура, очевидно спиральная, вводится в основной белок, когда он связывается с додецилсульфатом натрия (SDS) или липидами, которые являются кислотными или цвиттерионными (Palmer & Dawson, 1969; Anthony & Moscarello, 1971; Liebes et al. ., 1976; Moore и др. ., 1976). Epand et al. (1974) исследовали основные белковые молекулы с помощью электронной микроскопии. Образцы, которые были растворены в водном растворе при pH 7,4 перед подготовкой к микроскопии, выглядели как компактные скрученные частицы в форме стержня с размерами примерно 13 × 130 Å.
Основы белков | Бостонская детская больница
Белок — это большая органическая молекула, состоящая из более мелких молекул аминокислот.Хотя небольшие белки могут содержать всего 50 аминокислот, они, как правило, намного больше и состоят из сотен или даже тысяч аминокислот.
Иногда белки намеренно разрезают на более мелкие сегменты. Эти сегменты называются пептидами. Пептид может состоять от двух до 50 аминокислот.
В белке аминокислоты связаны одна за другой, образуя длинную цепь. Они удерживаются вместе пептидными связями, которые представляют собой химические связи между атомами углерода и азота аминокислот.Помимо углерода и азота, аминокислоты содержат водород, кислород и иногда серу.
Белки содержат 20 видов аминокислот. За исключением лейцина и изолейцина, которые имеют одинаковую атомную массу 113 дальтон, каждая аминокислота имеет уникальную массу. (Дальтон — единица измерения атомной массы. Один Дальтон равен 1/12 массы одного атома углерода 12.)
Белки, составляющие от 12 до 18 процентов стройного взрослого тела, играют важную роль в физиологии человека.Вот несколько:
- Белки образуют структурный каркас тканей и других структур организма. Например, коллаген способствует образованию сухожилий, связок, хрящей и других соединительных тканей.
- Белки переносят вещества по всему телу. Например, гемоглобин — переносчик кислорода и углекислого газа.
- Белки работают как ферменты, облегчая химические реакции. В организме человека насчитывается более 1000 известных типов ферментов.
- Белки миозин и актин позволяют мышцам сокращаться.
- Белки регулируют физиологические процессы и контролируют рост.
- Белки являются неотъемлемой частью иммунной системы. Например, антитела состоят из типа белка, используемого иммунной системой.
Как гены кодируют белки?
Вы можете представить себе геном человека как одну длинную цепочку ДНК, разделенную на 46 частей, называемых хромосомами. В каждой хромосоме тысячи генов. Последовательность ДНК, из которой состоит ген, похожа на программный код, который инструктирует клетку, как производить белки.
Четыре типа оснований составляют цепь ДНК — аденин (A), цитозин (C), гуанин (G) и тимин (T). У каждой базы есть дополнение — другая база, с которой она обычно сочетается. Пара А с Т, а пара С с Дж. В целом геном человека состоит из 3 миллиардов пар оснований.
Три последовательных основания в цепи ДНК, называемые кодоном, могут действовать как инструкция для клетки по выбору определенного типа аминокислоты. Серия кодонов инструктирует клетку собрать последовательность аминокислот для образования белка.Клетка знает, когда начинать и прекращать кодирование белков, распознавая специальные кодоны запуска и остановки, которые находятся в цепи ДНК.
Генетические инструкции выполняются различными молекулами и клеточными структурами, включая транспортную РНК (тРНК), информационную РНК (мРНК) и рибосомы. Другими словами, клетка способна считывать фрагменты кода ДНК и использовать информацию для соединения белка.
Аминокислоты бывают разных форм. Клетка, руководимая генами, использует это свойство и строит белки, которые имеют совершенно разные размеры и формы.
Что такое белок? Биолог объясняет
Примечание редактора: Натан Альгрен — доцент кафедры биологии Университета Кларка. В этом интервью он подробно объясняет, что такое белки, как они производятся, а также широкий спектр функций, которые они выполняют в организме человека.
Натан Альгрен объясняет, что делают белки в нашем организме.
Что такое белок?
Белок — это основная структура, которая встречается во всем живом.Это молекула. И главное в белке — это то, что он состоит из более мелких компонентов, называемых аминокислотами. Мне нравится думать о них как о бусинах разного цвета. Каждая бусинка представляет собой аминокислоту, представляющую собой более мелкие молекулы, содержащие атомы углерода, кислорода, водорода и иногда атомы серы. Итак, белок — это, по сути, цепочка, состоящая из этих маленьких отдельных аминокислот. Есть 22 различных аминокислоты, которые вы можете комбинировать по-разному.
Белок обычно не существует в виде цепочки, но фактически сворачивается в определенную форму, в зависимости от порядка и того, как эти разные аминокислоты взаимодействуют друг с другом.Эта форма влияет на то, что белок делает в нашем организме.
Откуда берутся аминокислоты?
Аминокислоты в нашем организме поступают из пищи, которую мы едим. Мы также производим их в нашем теле. Например, другие животные производят белки, а мы их едим. Наш организм берет эту цепочку и разбивает ее на отдельные аминокислоты. Затем он может преобразовать их в любой белок, который нам нужен.
После того, как белки расщепляются на аминокислоты в пищеварительной системе, они попадают в наши клетки и как бы плавают внутри клетки, как эти маленькие отдельные шарики в нашей аналогии.А внутри клетки ваше тело в основном связывает их вместе, чтобы вырабатывать белки, необходимые вашему организму.
Мы можем производить около половины необходимых нам аминокислот самостоятельно, а остальные мы должны получать из пищи.
Что делают белки в нашем организме?
Ученые точно не уверены, но большинство из них согласны с тем, что в нашем теле около 20 000 различных белков. Некоторые исследования предполагают, что их может быть даже больше. Они выполняют множество функций — от некоторых метаболических преобразований до удержания ваших клеток вместе и заставляя ваши мышцы работать.
Их функции делятся на несколько широких категорий. Один структурный. Ваше тело состоит из множества различных структур — представьте себе струнные структуры, шарики, якоря и т. Д. Они образуют вещество, которое скрепляет ваше тело. Коллаген — это белок, который придает структуру вашей коже, костям и даже зубам. Интегрин — это белок, который обеспечивает гибкие связи между вашими клетками. Ваши волосы и ногти состоят из протеина, называемого кератином.
Еще одна важная роль, которую они берут на себя, — это биохимия — то, как ваше тело выполняет определенные реакции в вашей клетке, такие как расщепление жиров или аминокислот.Помните, я сказал, что наше тело расщепляет белок из пищи, которую мы едим? Даже эту функцию выполняют такие белки, как пепсин. Другой пример — гемоглобин — белок, переносящий кислород в крови. Итак, они проводят эти особые химические реакции внутри вас.
Белки также могут обрабатывать сигналы и информацию, например белки циркадных часов, которые отслеживают время в наших клетках, но это несколько основных категорий функций, которые белки выполняют в клетке.
Почему белок часто ассоциируется с мышцами и мясом?
В разных продуктах питания разное содержание белка. В таких растениях, как пшеница и рис, много углеводов, но они менее богаты белком. Но в мясе вообще больше белка. Для создания мышц вашего тела требуется много белка. Вот почему белок часто ассоциируется с употреблением мяса и наращиванием мышечной массы, но на самом деле белки задействованы гораздо больше.
[ Получайте удовольствие от разговора каждые выходные. Подпишитесь на нашу еженедельную рассылку.]
Взаимосвязь между изоэлектрической точкой и: длиной белков, таксономией и экологией организмов | BMC Genomics
Общие свойства распределений изоэлектрических точек в протеомах
Некоторые примеры распределений изоэлектрических точек белков, кодируемых выбранными протеомами, показаны на рис. 1 (дополнительные протеомы см. В дополнительных файлах данных 2, 3 и 4).Все проанализированные протеомы обычно показывают бимодальное распределение с меньшим третьим пиком между двумя основными пиками, что согласуется с результатами других авторов [7, 8] и предположениями о том, что мультимодальное распределение pI соответствует значениям pK аминокислотных фрагментов [ 8, 14].
Рисунок 1
Гистограммы значений pI с интервалами 0,1 единицы (левая панель) и отношения между длиной белков (log L) и их pI (правая панель) для выбранных прокариотических протеомов с различным смещением pI (b): (A ) Natronomonas pharaonis DSM 2160, b = -87%.(B) Silicibacter pomeroyi DSS-3, b = -44%; (C) Ehrlichia ruminantium str. Гардель, b = 0%; (D) Mycoplasma pneumoniae M129, b = 43%; (E) Wigglesworthia glossinidia , b = 86%. Черные точки представляют собой набор кислых белков, а серые — набор основных белков. Диаграммы для всех проанализированных протеомов доступны в дополнительных файлах данных 2, 3 и 4.
Чтобы упростить сравнительный анализ распределения pI различных протеомов, мы разделили все наборы белков на два набора, названные позже кислотным и основным наборами.Точка деления соответствует значению pI, для которого распределение pI достигает минимума между кислотным и основным наборами (подробности см. В разделе «Методы»). Для большинства протеомов точка деления находилась между 7,4 и 7,5. Распределение значений пунктов деления очень узкое — диапазон 7,22–7,54 при среднем и медиане 7,41. Мы не обнаружили никакой корреляции между pH в этих точках и таксономической или экологической классификацией организмов и размерами генома.Универсальность значения pH минимума в распределении pI подтверждает вывод других авторов о том, что бимодальное распределение pI является результатом внутренних химических свойств аминокислот [8].
Для каждого протеома мы рассчитали средний pI отдельно для основного и кислого наборов белков, среднюю длину белков и смещение pI (см. Раздел «Методы»). Смещение pI просто описывает асимметрию бимодального распределения pI (рис. 1). Диапазон составляет от -100% до 100%.Эти два крайних значения указывают на то, что все белки в данном протеоме являются кислотными или основными, соответственно. Значение, близкое к нулю, означает, что данный протеом имеет одинаковую долю кислотных и основных белков. Мы обнаружили, что если протеом имеет более кислые белки, средний pI белков кислотного набора ниже, а взаимосвязь статистически значима (коэффициент корреляции, r = 0,76, p <0,001) - дополнительный файл данных 5A. С другой стороны, если в протеоме преобладают основные белки, это не связано со сдвигом среднего значения pI основных белков, за исключением некоторых внутриклеточных микроорганизмов (дополнительный файл данных 5B).Мы также обнаружили, что базовые наборы имеют большую дисперсию их pI, чем кислые наборы, что указывает на большее разнообразие основных белков.
Взаимосвязь между значением pI и размером белков
Виртуальное двумерное распределение белков показывает характерный образец, который может напоминать «бабочку» [7] или «легкие», и никакой корреляции между pI и молекулярной массой не наблюдалось. белки. Nandi et al. [15] обнаружили, что молекулярная масса белков является гораздо более консервативной характеристикой, чем их pI, что дополнительно предполагает отсутствие корреляции между pI и молекулярной массой.Однако после разделения протеомов на кислотные и основные наборы можно увидеть корреляцию между pI и размером белков для многих протеомов — положительную для кислых белков и отрицательную для основных белков (рис. 1). Хотя коэффициент корреляции не очень высок, для более чем 92% кислотных наборов и почти 99% основных наборов кодируемых хромосомами протеомов архей, бактерий и эукариот он является статистически значимым с поправкой Бенджамини-Хохберга на p <0,05. Кроме того, корреляция между размером и pI белков является общей чертой с очень узким распределением коэффициентов корреляции (показано на рис.2 отдельно для кислых и основных белков).
Рисунок 2
Распределение коэффициентов корреляции между значением pI и длиной белков, рассчитанных отдельно для кислотного и основного наборов протеомов.
Чтобы показать, что эти корреляции являются внутренними свойствами протеомов, мы создали три искусственных протеома, состоящих из случайно сгенерированных белков, характеризующихся: (i) средним аминокислотным составом белков E. coli и длиной настоящего белка E. .белки coli ; (ii) равные частоты аминокислот и длина реальных белков E. coli ; (iii) средний аминокислотный состав белков E. coli и равномерное распределение длины в диапазоне белков E. coli . (См. Дополнительный файл данных 6 для распределения pI и отношений между длиной белков и их pI для этих протеомов.) Создание искусственных протеомов повторяли 100 раз для каждого набора. В каждом случае мы получили значительную корреляцию между длиной и значениями pI (табл.1). Каждая версия искусственных протеомов давала бимодальное распределение pI и более сильные корреляции между pI и длиной белков, чем в реальном протеоме E. coli . Смещение pI реального протеома не попадает в диапазон значений pI сгенерированных протеомов и является ближайшим к сгенерированным протеомам, исходя из среднего аминокислотного состава и длины реальных белков E. coli . Смещение pI было более сбалансированным, когда белки были получены из однородного распределения аминокислот, но все еще без белков с pI, близким к 7.4. Кроме того, протеом становился более кислым, когда длина белков соответствовала равномерному распределению. Это предполагает, что отбор по длине белков может до некоторой степени влиять на распределение pI.
Таблица 1. Свойства искусственных и реальных протеомов E. coli .
Результаты означают, что более длинные белки могут поддерживать нейтральный или почти нейтральный pI, тогда как самые экстремальные значения pI специфичны для более коротких белков. Вероятно, это связано с чисто статистическими причинами.Более короткие белки демонстрируют более высокие колебания аминокислотного состава, что сильно влияет на их pI — включение даже одного заряженного остатка значительно сдвигает pI белка в сторону более низкого или более высокого значения. С другой стороны, длинные белки, обычно состоящие из более заряженных аминокислот, намного лучше амортизируют эффект колебаний в их составе и могут поддерживать свой pI даже близким к 7,4. Это «статистическое» объяснение взаимосвязи между размером белков и pI не исключает возможности того, что эта взаимосвязь может иметь некоторые биологические последствия в диверсификации структуры и функции белков.Например, если существует ограничение выбора для получения очень кислого или очень основного белка, самый простой способ добиться этого — «сделать» его коротким. На самом деле очень простые белки, взаимодействующие с нуклеиновыми кислотами — рибосомные белки — обычно очень короткие. Мы считаем, что необходимо найти больше белков, подвергнутых такому отбору. Эти белки могут принадлежать к определенным регуляторным белкам, факторам транскрипции, модуляторам, сигнальным белкам, небольшим белкам, взаимодействующим с другими белками и т. Д.
Мы также заметили разницу в длине кислых и основных белков. Сравнительный анализ показал, что кислотные белки значительно длиннее основных (t-критерий Стьюдента, скорректированный по Бенджамини-Хохбергу p <0,05) более чем в 95% протеомов архей, бактерий и эукариот. Кислые белки в среднем на 73 и 107 аминокислотных остатков длиннее основных у Prokaryota и Eukaryota соответственно. Вероятно, это связано с наличием в основном наборе очень коротких рибосомных белков и длинных аминоацил-тРНК-синтетаз в кислом [15].
Nandi et al. [15] наблюдали, что размер ортологичных белков, обнаруженных у близкородственных организмов, намного более консервативен, чем их pI. Мы сравнили длину ортологичных белков многих протеомов, чьи значения pI были изменены с кислых на основные или наоборот, но мы также не наблюдали каких-либо статистически значимых различий в длине этих белков. Мы обнаружили, что изменение pI не зависело от длины сравниваемых ортологов.
Связь между значениями pI протеомов и таксономией
При анализе потенциальных взаимосвязей между pI протеомов и таксономией мы использовали смещение pI, которое является довольно кратким, но очень информативным параметром, подчеркивающим наиболее четкие и основные различия в распределениях pI. .На рис. 3 показан результат статистического анализа смещения pI различных групп протеомов и дендрограмма UPGMA, классифицирующая различные группы протеомов согласно медиане смещения pI. Анализ позволяет разделить протеомы как минимум на две группы: (1) более основные протеомы, кодируемые пластидами, вирусами и плазмидами, и (2) более кислые протеомы архей, бактерий, эукариот, митохондрий и фагов. Это разделение поддерживается 95% (метод подвыборки), WLS-LRT и F-тестом (оба p <0.001). Более того, каждое сравнение между членами, отнесенными к этим различным группам, проведенное с помощью теста Краскела-Уоллиса, является статистически значимым (p <0,002). Внутри кислой группы можно выделить некоторые подгруппы в соответствии с высоким процентом поддержки: эукариоты с митохондриями и бактерии с фагами. Клады, содержащие прокариот и фаги, довольно слабо подтверждены анализом подвыборки, но очень значимы согласно WLS-LRT и F-тесту (для обоих p <0,001). Наблюдаемая основность протеомов пластид, вирусов и плазмид, вероятно, связана с преобладанием основных белков (напр.г. рибосомный в случае пластид и основных белков, взаимодействующих с нуклеиновыми кислотами в случае вирусов), а также может быть результатом более высокой скорости накопления мутаций, особенно в вирусах и плазмидах (см. раздел «Связь между значением pI протеомов, их размеры и содержание GC в геномах »для аналогичного объяснения внутриклеточных бактерий).
Рис. 3
Статистический анализ смещения pI различных групп протеомов и их кластеризации на основе UPGMA согласно медиане смещения pI.Числа в узлах означают процентную поддержку, основанную на методе подвыборки, а звездочки обозначают результаты тестов WLS-LRT / F (оба с p <0,001).
Наш анализ протеомов прокариот, выполненный на нижнем таксономическом уровне, не выявил какой-либо связи между смещением pI и филогенией, что согласуется с результатами Knight et al. [7]. Многие монофилетические или близкородственные группы (например, таксоны архей или протеобактерий) не были сгруппированы, когда смещение pI использовалось в качестве критерия.Это указывает на то, что распределение pI не сохранилось во время эволюции прокариот. Мы не наблюдали такой взаимосвязи и для протеомов эукариот.
С другой стороны, мы обнаружили интересную группировку, связанную с филогенетическим сигналом, при анализе митохондриальных протеомов, которые намного меньше и более консервативны в соответствии с их содержанием белка, чем большие протеомы, кодируемые хромосомами (рис.4, см. Также Табл. 2). Анализируемые группы протеомов четко делятся на две подгруппы: Viridiplantae + Protista и Fungi + Metazoa.Это деление строго поддерживается в 100%, WLS-LRT и F-тесте (для обоих p <0,001). Это согласуется с мнением о том, что клады Fungi + Metazoa, называемые Opisthokonta, являются хорошо известной группой, что подтверждается многими анализами молекулярных данных [16, 17]. Кроме того, две подгруппы Metazoa: Chordata и non-Chordata (включая большинство типов беспозвоночных) образуют хорошо поддерживаемую кладу (97%; WLS-LRT и F-тест: для обоих p <0,001). Группа Viridiplantae + Protista в среднем имеет гораздо больше основных протеомов, чем Fungi + Metazoa.В группе Opisthokonta у грибов есть умеренно основные протеомы, в то время как у Metazoa есть нейтральные (нехордовые) или слабокислые (хордовые).
Рисунок 4
Статистический анализ смещения pI митохондриальных протеомов и их кластеризации на основе UPGMA согласно медиане смещения pI. Числа в узлах означают процентную поддержку, основанную на методе подвыборки, а звездочки обозначают результаты тестов WLS-LRT / F (оба с p <0,001).
Таблица 2 Сравнение смещения pI для подмножеств органеллярных протеомов.
Наблюдаемый филогенетический сигнал в смещении pI, вероятно, является результатом различного состава протеомов. Митохондриальные протеомы Viridiplantae, Protista и Fungi обычно в несколько раз больше, чем протеомы Metazoa, которые обычно содержат 12–13 белков. Фактически, большинство белков (например, рибосомных белков), которые отсутствуют у Metazoa, но присутствуют в Viridiplantae, Protista и Fungi, являются очень простыми (дополнительный файл данных 7). Поэтому, когда мы провели такой же анализ на основе протеомов, состоящих только из 12 белков, присутствующих в большинстве митохондриальных протеомов, филогенетический сигнал исчез, а смещение pI основных протеомов стало нейтральным или даже кислым (для грибов).Мы не обнаружили никакой связи между смещением pI и филогенезом на более низких таксономических уровнях: между типами многоклеточных животных и подгруппами Craniata при анализе как полных протеомов, так и протеомов, содержащих только 12 общих белков.
Аналогично Knight et al. [7], но анализируя более широкий набор протеомов, мы обнаружили значительную корреляцию между смещением pI протеомов, кодируемых прокариотическими геномами и их плазмидами (N = 63, r = 0,74, p <0,001). Нет такой корреляции между смещением pI протеомов с ядерным кодированием эукариот и протеомами их органелл (N = 36, r = 0.04, p = 0,81). Мы получили аналогичные результаты при анализе средних значений pI для кислотных и основных наборов по отдельности, и никакой корреляции не было обнаружено в этих двух случаях, когда изучалась средняя длина белков в этих двух наборах. Кажется, что геномы органелл более независимы, чем плазмиды, чтобы следовать геномным тенденциям в pI, что, по-видимому, не верно для плазмид. Например, органеллы разделены двумя мембранами и обладают собственными репликационными, транскрипционными и трансляционными механизмами.Плазмиды обычно кодируют больше основных белков, чем хромосомы, но мутационное давление или некоторые ограничения отбора влияют на эти два протеома одновременно, и перенос генов между плазмидами и хромосомами происходит, вероятно, чаще, чем между органеллярным и ядерным геномами. Если будет доступно больше данных, было бы интересно проанализировать такие отношения между вирусными или фагальными протеомами и протеомами инфицированных организмов.
Взаимосвязь между значениями pI и субклеточной локализацией белков
Анализируемые митохондриальные и пластидные протеомы включают только белки, кодируемые органеллярными геномами.Однако существует множество белков, кодируемых ядерными генами, которые нацелены на органеллы. Кодируемые ядром органеллярные белки обычно снабжены N-концевыми сигналами нацеливания (транзитными пептидами), ответственными за их импорт в органеллы. Эти пептиды отщепляются органеллярными пептидазами после транспортировки белков. Поскольку эти пре-последовательности богаты основными аминокислотными остатками, pI преждевременных необработанных белков (то есть целых последовательностей) должна быть более основной, чем pI зрелых белков (т.е.е. без транзитного пептида). В закладке. 2 мы сравнили смещение pI преждевременных и зрелых белков с протеомами, кодируемыми органеллярными геномами. Поскольку митохондриальные протеомы различаются в разных таксономических группах, мы проанализировали их отдельно. (См. Дополнительный файл данных 8 для распределения pI и взаимосвязей между длиной белков и их pI для этих протеомов.) Как и ожидалось, присутствие очень основных транзитных пептидов (pI смещение 93 — 100%) в преждевременных белках сдвигает pI распределение этих белков по основности (т.е.е. более высокие значения pI bias), тогда как большинство зрелых белков являются кислыми. Интересно, что только в пластидных и митохондриальных протеомах растений все еще существует слабый избыток кислых преждевременных белков. Кодируемые ядром органелларные протеомы обычно отличаются от протеомов, кодируемых органеллами. Значения смещения pI только митохондриальных белков преждевременного созревания Viridiplantae и Fungi и митохондриальных зрелых белков Chordata попадают в пределы квартильного диапазона смещения pI соответствующих протеомов, кодируемых органеллами.
Schwartz et al. [6] проанализировали три субклеточных локализации белков и обнаружили, что цитоплазматические и интегральные мембранные протеомы содержат больше кислых и основных белков, соответственно, тогда как ядерный протеом имеет довольно равные пропорции кислотных и основных белков. В этой статье мы рассмотрели 12 субклеточных локализаций в анализах (табл. 3). (См. Также дополнительный файл данных 9 для распределения pI и отношений между длиной белков и их pI для этих протеомов.Результаты показали, что наиболее кислые протеомы присутствуют не только в цитозоле, но также в вакуолях и лизосомах, а многие кислые белки строят цитоскелет. Самым основным из них является митохондриальный протеом, и распределение pI белков, интегрированных в мембрану, немного смещено в сторону основности. Остальные субклеточные локализации содержат более сбалансированное соотношение кислотных и основных белков. Основной характер интегральных мембранных белков можно объяснить присутствием основных остатков по обе стороны от области, охватывающей мембрану, которые играют роль в стабилизации белков в мембране [6].Основность митохондриальных белков обусловлена наличием положительно заряженных транзитных пептидов. Другие взаимосвязи между pI и субклеточной локализацией, вероятно, связаны с присутствием некоторых специфических групп белков в определенных локализациях (например, кислотных гидролаз в лизосомах, деградирующих ферментов в вакуолях и кислых белков, таких как актины, динеины, кератины, кинезины и т. ламины, миозины, тубулины в цитоскелете). Интересно сопоставить распределение pI конкретных протеомов с pH компартментов, в которых эти протеомы расположены.Однако мы не наблюдали какой-либо статистически значимой корреляции (r = 0,38, p = 0,27, n = 10).
Таблица 3 Сравнение смещения pI для протеомов различной субклеточной локализации.
Взаимосвязь между значениями pI протеомов и экологией их организмов
Для детального анализа взаимосвязей между pI протеомов и экологией организмов все проанализированные прокариотические протеомы были классифицированы по различным экологическим подгруппам (см. Подробности в разделе «Методы») и затем сравнивают в отношении смещения pI (Табл.4). Тесты Краскела-Уоллиса показали, что различия не были статистически значимыми, когда организмы были сгруппированы в соответствии с их потребностями в кислороде и температуре (p> 0,1), но они были очень значимыми при классификации в соответствии с их средой обитания, связью с клетками-хозяевами и требованиями к солености (p <0,001). К такому же выводу мы пришли (на том же уровне значимости) при выполнении теста χ 2 . На рис. 5 представлены отношения наблюдаемого к ожидаемому количеству протеомов в данном классе смещения pI для различных экологических классификаций.
Таблица 4 Статистический анализ смещения pI для различных экологических групп. Рисунок 5
Отношения наблюдаемого и ожидаемого количества протеомов в данном классе смещения pI для различных экологических классификаций: (A) кислород, (B) температура, (C) соленость, (D) среда обитания и (E) отношение к клетке-хозяину.
Анализы показали, что соленость положительно коррелирует с кислотностью протеомов — чем более галофильные организмы имеют более кислые протеомы. Это согласуется с результатами других авторов, наблюдавших у галофилов преобладание кислотных остатков над основными [18–22] и низкую изоэлектрическую точку их белков [10–12].Чрезвычайно галофильные и умеренно галофильные бактерии присутствуют только в «кислом» классе, а мезогалофилы исчезают в «основном» классе (рис. 5C). Эта связь обычно объясняется более высокой стабильностью и растворимостью белков, богатых кислотными остатками, в гиперсоленой среде [19, 23–28].
Принимая во внимание предпочтения среды обитания, организмы, ассоциированные с хозяином, имеют наименее кислые протеомы по сравнению с другими группами, а водные бактерии обладают наиболее кислыми протеомами. В « кислом » классе водные бактерии представляют собой наиболее представленную группу, а виды, связанные с хозяевами, — наиболее недопредставленную (рис.5D). С другой стороны, «базовый» класс содержит только микроорганизмы, ассоциированные с хозяином. Хотя протеомы видов, ассоциированных с хозяином, смещены в сторону более основных протеомов, в среднем они все еще являются кислыми (Табл. 4). Однако более подробная классификация организмов с учетом их отношения к клетке-хозяину показала, что протеомы внутриклеточных бактерий в среднем являются основными, а внеклеточные и свободноживущие / внутриклеточные виды имеют слегка кислые протеомы (рис. 5E). Более кислые протеомы характерны для свободноживущих / внеклеточных и свободноживущих видов.Фактически, эти две группы чрезмерно представлены в «кислотном» классе, тогда как внутриклеточные бактерии сильно представлены в «основном» классе. Результаты показывают, что чем больше организм связан с клеткой-хозяином, тем больше у него основного протеома. Объяснение этого результата будет обсуждаться в следующем разделе, где рассматривается взаимосвязь между смещением pI, размером протеома и содержанием GC в геноме. Мы также заметили, что все внутриклеточные организмы, которые имеют слабокислые протеомы (Anaplasma, Brucella, Chlamydiae, Ehrlichia) и большинство из них (за одним исключением), имеют слабоосновные протеомы со смещением pI ≤ 20% (Bartonella, Coxiella, Parachlamydia, Rickettsia conorii , Tropheryma, Wolbachia) находятся и обычно размножаются в вакуолях или фагосомах.Что интересно, R. conorii имеет наименее основной протеом в пределах рода Rickettsia и как единственный представитель этого рода наблюдается в вакуоли [29]. Это означало бы, что среда вакуолей изменяет протеомы внутриклеточных организмов в сторону кислотности.
Экологические изменения довольно сильно и быстро влияют на смещение pI в процессе эволюции протеомов, поскольку изменения смещения pI наблюдаются даже среди близкородственных организмов. Мы собрали все проанализированные виды, принадлежащие к одному роду и имеющие разное экологическое назначение (см. Дополнительный файл данных 10).В каждом случае смещение pI видов, ассоциированных с хозяином, больше смещено в сторону более высоких значений (то есть основности), чем смещение видов, живущих в нескольких средах. Наиболее яркими примерами являются два вида Burkholderia, из которых один живет в наземной среде обитания и обладает кислотным протеомом, а другой связан с хозяином и имеет основной протеом. Протеомы других видов, ассоциированных с хозяином, хотя и смещены в сторону основности, все же остаются кислыми, вероятно, потому, что эти виды все еще являются факультативными и внеклеточными паразитами.Более того, явный сдвиг смещения pI виден при сравнении видов, живущих с разными требованиями к солености. Протеомы галофильных и мезогалофильных видов содержат более кислые белки, чем негалофильные.
Взаимосвязь между значениями pI протеомов, их размерами и GC-содержанием геномов
Взаимосвязь между pI-значением протеомов, их размерами и GC-содержанием геномов была проанализирована Knight et al. [7]. Они пришли к выводу, что нет корреляции между медианной pI и содержанием GC в небольших базовых протеомах, и открыли дискуссию о причинах основности крошечных протеомов, дав два непроверенных возможных объяснения: отбор белков в протеоме и отбор внутри определенных белков.Чтобы увидеть, насколько общими являются эти отношения, мы выполнили анализ более широкого набора протеомов, используя смещение pI и более подробно рассматривая связь организмов с клеткой-хозяином.
На фиг. 6А представлена взаимосвязь между смещением pI и размером протеома (выраженным как логарифм количества белков) для прокариотических организмов. График в дополнительном файле 11 данных включает также эукариотические организмы. Для прокариотических протеомов видна четкая тенденция — меньшие протеомы содержат больше основных белков, тогда как большинство протеомов Eukaryota смещены в сторону больших значений размера протеома.Коэффициент корреляции для набора, содержащего прокариотические протеомы, составляет r = -0,64 (p <0,001). Отрицательная корреляция была обнаружена также для смещения pI и содержания GC геномов (для всего набора: r = -0,49, для прокариотических протеомов r = -0,50; для обоих p <0,001). Как правило, геномы с более низким содержанием GC кодируют более простые протеомы. Детальный анализ прокариотических протеомов с учетом взаимосвязи между организмами и клеткой-хозяином показал, что чем больше штамм бактерий полагается на своего хозяина, тем выше фракция AT в его геноме и тем меньше и более базовый протеом он имеет (рис.6). Большинство этих организмов являются внутриклеточными паразитами или эндосимбионтами, или, по крайней мере, они каким-то образом связаны со своим хозяином (внеклеточными). Кажется, что правила являются общими, потому что протеом, кодируемый нуклеоморфом эукариота Guillardia theta [30], также является AT-богатым, редуцированным и самым основным во всем наборе. G. theta — одноклеточная водоросль-криптофит, которая является хозяином для другой клетки с уменьшенным остаточным ядром, называемой нуклеоморфом. Клетка также может считаться эндосимбионтом.
Рисунок 6
Связь между смещением pI и: (A) логарифмом размера протеома и (B) содержанием геномных GC для различных экологических групп прокариот.
Наблюдаемые корреляции становятся более ясными, если рассматривать только виды с протеомами менее 1000 белков. Коэффициенты корреляции изменяются от -0,64 до -0,82 для взаимосвязи между смещением pI и размером протеома и от -0,49 до -0,58 для взаимосвязи между смещением pI и содержанием GC и являются статистически значимыми.Таким образом, нельзя полностью исключить гипотезу о том, что неэффективность механизмов репарации ДНК у внутриклеточных микроорганизмов вызывает большое смещение AT их малых геномов и, как следствие, большее содержание основного лизина [31]. Возможно, что существуют и другие причины наблюдаемой взаимосвязи между основностью протеома и уменьшением его размера, например: увеличение или потеря некоторых групп белков, вызывающих сдвиг pI всего протеома или отбор на изменение pI протеома. определенные белки, напримерг. участвует в адаптации к новой среде или хозяину [7]. Первая гипотеза объясняет, по крайней мере, различия между митохондриальными протеомами (см. Выше).
Мы рассмотрели, объясняет ли эта гипотеза основность малых прокариотических протеомов. Мы можем исключить приобретение основных белков как объяснение, потому что трудно найти какие-либо новые гены, полученные внутриклеточными бактериями, которые отсутствовали бы у близкородственных свободноживущих родственников [32–34]. Обычно у внутриклеточных бактерий многие гены разрушаются или, скорее, уничтожаются, оставляя консервативный набор генов (например,г. кодирующие основные рибосомальные белки). Утрата незаменимых генов, кодирующих кислые белки, может привести к сдвигу всего протеома в сторону основности. Можно предположить, что консервативные ортологичные белки, присутствующие как во внутриклеточных бактериях, так и в их свободноживущих аналогах, не изменили своих функций в ходе эволюции, и эти наборы имеют одинаковые пропорции различных классов белков. Тогда мы должны ожидать, что эти протеомы имеют схожее распределение pI и являются более основными, чем белки, которые все еще присутствуют в свободноживущих бактериях, но были потеряны во внутриклеточном организме.Чтобы проверить это, мы проанализировали распределение pI для трех наборов белков: присутствующих только в свободноживущих бактериях, но не во внутриклеточных, и ортологичные белки, общие для внутриклеточных бактерий и их свободноживущие аналоги, проанализированные отдельно для этих двух групп. бактерий. Результаты сравнения Escherichia coli K12 с: Buchnera aphidicola str. Bp, Candidatus Blochmannia floridanus и Wigglesworthia glossinidia представлены на рис.7. Набор ортологичных белков этих эндосимбионтов очень базовый. Однако ортологичные белки E. coli даже более кислые, чем набор белков, присутствующих только в E. coli и потерянных у эндосимбионтов. Мы получили аналогичные результаты для семи других пар бактерий. Предполагая, что протеом E. coli похож на свободно живущего предка эндосимбионтов, мы можем оценить, что от 300 до 400 его белков (то есть 55-65 процентов общей части протеомов) изменили свою pI с кислой на основную во время переход к внутриклеточному образу жизни.Следовательно, в отличие от митохондриальных протеомов, наблюдаемый сдвиг малых бактериальных протеомов в сторону основности не может быть объяснен увеличением или потерей некоторых белков в протеоме, а сдвигом pI многих белков.
Рисунок 7
Распределение pI для трех наборов белков: присутствует в E . coli только K12, а не во внутриклеточном эндосимбионте (только в Ec), E. coli белков, ортологичных внутриклеточным бактериям [(common (Ec)], и эндосимбионтным белкам, ортологичным E.coli [(общий (инициалы эндосимбионта)]. Сравнения сделаны для E . coli K12 с: Buchnera aphidicola str. Bp (A), Candidatus Blochmannia floridanus (B) и gloss (C).
Чтобы найти объяснение этому сдвигу, мы создали для каждого организма протеом с таким же распределением белков по длине, что и в его реальном протеоме, но с аминокислотным составом, рассчитанным для базового состава, характерного для данного генома. это в некоторой степени отражает состав, вызванный мутационным давлением [35].Мы создали 100 таких протеомов для каждого организма и усреднили их смещение pI. Эти протеомы демонстрируют равновесное устойчивое состояние протеомов, развивающихся только под давлением мутаций без каких-либо ограничений отбора. Все сгенерированные протеомы показали чрезвычайно высокое (более 85%) смещение pI со средним значением = 94% (фиг. 8A, см. Дополнительный файл данных 6 для примера E. coli ). На самом деле, согласно таблице генетического кода, при равных частотах кодонов представительство кислотных остатков ниже, чем основных.Отношение между долей основной или кислой аминокислоты и содержанием GC является нелинейным, но любой нуклеотидный состав благоприятствует основным остаткам, особенно для экстремального содержания GC (рис. 8B). Наименьшая разница между долей кодируемой основной и кислой аминокислот составляет 37% ГХ. Это согласуется с соотношением, полученным для сгенерированных протеомов. Наименьшее смещение pI находится между 30% и 40% от содержания GC. Кроме того, точки раздваиваются в сторону более высокого смещения pI для крайних значений содержания GC, показывая отрицательную корреляцию ниже и положительную корреляцию выше 40% содержания GC.Отрицательная связь, вероятно, связана с увеличением основного лизина, кодируемого кодонами, богатыми AT, а положительная — с ростом основного аргинина, кодируемого кодонами, богатыми GC. Поскольку любая композиция ДНК (особенно та, которая находится в равновесии с мутационным давлением) сдвигает протеомы в сторону основности, мы можем предположить, что кислые протеомы подвергаются сильному давлению отбора, противоположному мутационному давлению. Чем более кислый протеом, тем более сильный отбор должен производиться на него.На данный момент наиболее кислые протеомы принадлежат к экстремальным галофилам, поэтому следует ожидать очень сильных ограничений отбора на эту группу со стороны гиперсоленой среды, в которой они живут.
Рисунок 8
Изоэлектрическая точка и содержание GC. (A) Связь между компьютерной смещением pI и содержанием геномного GC прокариотических организмов. Сгенерированное смещение pI представляет собой среднее значение, рассчитанное для 100 виртуальных протеомов, сгенерированных для каждого организма, при том же распределении белков по длине, что и в реальных протеомах, и аминокислотном составе, рассчитанном на основе базового состава, характерного для данного генома.(B) Соотношение между долей основных и кислых аминокислот и содержанием GC.
Результаты показывают, что смещение AT само по себе не может объяснить наблюдаемую основность крошечных протеомов. Мы не наблюдали никакой корреляции для этих виртуальных протеомов между смещением pI и размером протеома. Похоже, что ускорение накопления мутаций само по себе может быть ответственно за основность крошечных протеомов. Это согласуется с хорошо задокументированными более высокими темпами эволюции внутриклеточных бактерий, чем их свободноживущие родственники.Было высказано предположение, что более высокая скорость эволюции является результатом повышенной скорости мутаций [36] и / или эффекта храповика Мюллера или более легкой фиксации мутаций за счет генетического дрейфа в небольших бесполых популяциях [37–46]. Соответственно, увеличение основности протеомов и увеличение смещения мутаций AT будут параллельными явлениями, возникающими в результате более высокой скорости эволюции геномов внутриклеточных бактерий. Повышенное смещение AT, вероятно, является результатом устранения генов, кодирующих ферменты, связанные с репарацией ДНК и рекомбинацией, или, по крайней мере, из-за снижения их эффективности во внутриклеточных организмах [31, 32, 37, 47–49].Эти ферменты обычно корректируют связанную с ошибкой тенденцию к обогащению AT, например дезаминирование цитозина до урацила, который затем заменяется тимином.
Чтобы подтвердить наше объяснение, мы проанализировали 39 наборов консервативных ортологичных белков, присутствующих в каждом из 100 отобранных прокариотических организмов, что довольно равномерно представляет значения смещения pI. Мы предположили, что если увеличение скорости замены влияет на распределение pI, мы должны наблюдать положительную корреляцию между pI этих белков и смещением pI всех протеомов.Собственно, для 36 из них мы обнаружили такую статистически значимую корреляцию (см. Дополнительный файл данных 12). Три набора, для которых корреляция не была значимой, представляют самые основные рибосомные белки, и, возможно, их дальнейшее изменение было маловероятным. Результаты показывают, что даже очень консервативные белки, подвергающиеся строгим ограничениям отбора, следуют глобальной тенденции в распределении pI. Такая взаимосвязь, вероятно, касается многих других белков, поскольку анализ pI кластеров ортологичных групп (COG) показал, что белки только нескольких кластеров консервативны и остаются в том же кислотном или основном наборе, в то время как большинство из них прыгают между двумя наборами [15 ].Многие из этих беспорядочных белков являются мембранными белками, которые имеют прямой контакт с внешней средой и могут считаться адаптивными белками. Следовательно, это было бы в пользу гипотезы о выборе для изменения pI определенных белков. Однако это не полностью объясняет основность небольших прокариотических протеомов, потому что изменения pI затрагивают также многие немембранные и очень консервативные белки. (Связь между pI и другими явлениями, которые мы обсуждали в дополнительном файле данных 13.)
Было бы интересно исследовать взаимосвязь между изменением распределения pI протеомов и переходом организмов от свободного к внутриклеточному образу жизни на разных стадиях редукции генома. Очень проницательные результаты, касающиеся этого предмета, дадут секвенирование и анализ сокращенных геномов бактериальных эндосимбионтов, идентифицированных в эукариотических хозяевах, например цианобактерия в амебе Paulinella chromatophora , цианобактерия Cyanothece в диатоме Rhopalodia gibba , грамотрицательная бактерия в диатоме Pinnularia nobilis и многих различных бактериальных эндосимбионах.Дополнительные интересные результаты могут быть получены при анализе нуклеоморфов — небольших редуцированных эукариотических ядер, обнаруженных в определенных пластидах, присутствующих в некоторых группах водорослей, таких как криптомонады и хлорарахниофиты. Более того, интересно оценить, сколько мутаций требуется или должно быть принято для ремоделирования одного протеома в другой в аспекте их pI. Однако следует принимать во внимание многие факторы, такие как длина, аминокислотный состав, pI исходных белков, скорость мутаций и паттерны нуклеотидных замен, а также результирующие паттерны аминокислотных замен.
Белки: строительные блоки тела
Без воды и жира человеческое тело почти полностью состоит из белков. Белок является основным компонентом мышц, костей, органов, кожи и ногтей. Без воды мышцы состоят примерно на 80% из белка, что делает это питательное вещество особенно важным для спортсменов.
Как используется белок?
Организм расщепляет потребленный белок на аминокислоты и поглощает его. Он используется для наращивания мышц и органов, для выработки гормонов и антител, для хранения в виде жира и для сжигания в качестве энергии.
Виды протеиновых добавок
Существует много различных видов протеиновых добавок. Сывороточный протеин богат BCAA * , что делает его легко усваиваемым и высокоэффективным.
- BCAA (аминокислоты с разветвленной цепью) — это общее название незаменимых аминокислот валина, лейцина и изолейцина. BCAA — незаменимые аминокислоты, которые в мышцах превращаются в энергию.
Белок является неотъемлемой частью организма.
Сколько белка нужно организму?
Максимальное суточное количество, которое организм может использовать для синтеза белка, составляет около 2 граммов на 1 килограмм веса тела. Потребление большего количества белка не увеличит синтез, но увеличит количество потребляемой энергии и приведет к увеличению жировых отложений. Слишком много белка также может вызвать нагрузку на печень и почки.
Максимальное дневное количество, используемое для синтеза белка, составляет около 2 г на 1 кг массы тела
Поэтому важно регулировать количество потребляемого белка.
Если вы едите три хорошо сбалансированных приема пищи в день, вы легко сможете получить количество белка, необходимое вашему организму. Важно не только то, сколько вы едите, но и когда вы это едите.
Потребляйте белки и углеводы вместе
Когда белок потребляется вместе с углеводами (сахаром или глюкозой), повышается уровень сахара в крови и вырабатывается инсулин, способствуя синтезу аминокислот.
Углеводы также используются в качестве первого источника энергии, предотвращая использование аминокислот.После тренировки рекомендуется выпить что-нибудь вроде желеобразного напитка, который позволит вам одновременно потреблять углеводы и белок.
Когда употреблять белок
Важно употреблять белок в правильное время. Идеальное время для его употребления — сразу после тренировки.
Организм быстрее расщепляет белок сразу после тренировки, а также активнее его синтезирует, в результате чего в мышцы всасывается больше аминокислот.
Белок, потребленный сразу после тренировки, синтезируется лучше, чем белок, потребленный через 2 часа после тренировки.
Другие направления деятельности
Глава 3. Белки и аминокислоты
Глава 3. Белки и аминокислоты
1. БЕЛКИ
2. ПИЩЕВАРЕНИЕ БЕЛКОВ
И МЕТАБОЛИЗМ
3. ОБЩИЕ ТРЕБОВАНИЯ К БЕЛКАМ
4. АМИНОКИСЛОТЫ
5.КОЛИЧЕСТВЕННЫЙ
ТРЕБОВАНИЯ К АМИНОКИСЛОТЕ
6. ДОБАВКА
ДИЕТЫ С АМИНОКИСЛОТАМИ
7. ССЫЛКИ
Дж. Э. Халвер
Вашингтонский университет
Сиэтл, Вашингтон
1.1 Классификация
1.2 Структура
1.3 Свойства
1.4 Химическое определение
Белки представляют собой сложные органические соединения, состоящие из многих аминокислот, связанных вместе пептидными связями и поперечно связанных цепей сульфгидрильными связями, водородными связями и силами Ван-дер-Ваальса.Химический состав белков больше, чем у любой другой группы биологически активных соединений. Белки в различных клетках животных и растений придают этим тканям их биологическую специфичность.
1.1 Классификация
Белки можно классифицировать как:
(а) Простые белки. При гидролизе они дают только аминокислоты и иногда небольшие углеводные соединения. Примеры: альбумины, глобулины, глютелины, альбуминоиды, гистоны и протамины.
(б) Конъюгированные белки. Это простые белки в сочетании с некоторыми небелковыми веществами в организме. Примеры: нуклеопротеины, гликопротеины, фосфопротеины, гемоглобины и лецитопротеины.
(c) Производные белки. Это белки, полученные из простых или конъюгированных белков физическими или химическими способами. Примеры: денатурированные белки и пептиды.
1,2 Структура
Потенциальная конфигурация белковых молекул настолько сложна, что многие типы белковых молекул могут быть сконструированы и обнаружены в биологических материалах с различными физическими характеристиками.Глобулярные белки обнаруживаются в крови и тканевых жидкостях в аморфной глобулярной форме с очень тонкими или отсутствующими мембранами. Коллагеновые белки находятся в соединительной ткани, такой как кожа или клеточные мембраны. Волокнистые белки содержатся в волосах, мышцах и соединительной ткани. Кристаллические белки представлены хрусталиком глаза и подобными тканями. Ферменты — это белки с определенными химическими функциями, которые опосредуют большинство физиологических процессов жизни. Несколько небольших полипептидов действуют как гормоны в тканевых системах, контролирующих различные химические или физиологические процессы.Мышечный белок состоит из нескольких форм полипептидов, которые позволяют мышцам сокращаться и расслабляться при физических движениях.
1.3 Недвижимость
Белки также можно охарактеризовать по их химическим реакциям. Большинство белков растворимы в воде, спирте, разбавленной основе или в различных концентрациях солевых растворов. Белки имеют характерную спиральную структуру, которая определяется последовательностью аминокислот в первичной полипептидной цепи и стереоконфигурацией радикальных групп, присоединенных к альфа-углероду каждой аминокислоты.Белки термолабильны, проявляя различную степень лабильности в зависимости от типа белка, раствора и температурного профиля. Белки могут быть обратимыми или необратимыми, денатурированными при нагревании, концентрации соли, замораживании, ультразвуковой нагрузке или старении. Белки подвергаются характерному связыванию с другими белками в так называемой пластеиновой реакции и соединяются со свободными альдегидными и гидроксильными группами углеводов с образованием соединений типа Майяра.
1.4 Химическое определение
Содержание азота в большинстве белков, обнаруженных в тканях животных, орехов и зерна, составляет около 16 процентов; поэтому содержание белка обычно выражается как содержание азота × 6.25.
Проглоченные белки сначала расщепляются на более мелкие фрагменты пепсином в желудке или трипсином или химотрипсином из поджелудочной железы. Эти пептиды затем дополнительно восстанавливаются под действием карбоксипептидазы, которая гидролизует одну аминокислоту за раз, начиная со свободного карбоксильного конца молекулы, или с помощью аминопептидазы, которая отщепляет одну аминокислоту за раз, начиная со свободного амино-конца полипептида. цепь. Свободные аминокислоты, высвобождаемые в пищеварительную систему, затем всасываются через стенки желудочно-кишечного тракта в кровоток, где они затем повторно синтезируются в новые тканевые белки или катаболизируются для получения энергии или фрагментов для дальнейшего тканевого метаболизма.
Валовая потребность в белке была определена для нескольких видов рыб (см. Таблицу 1). Имитация цельного яичного протеина в тестовых диетах содержит избыток незаменимых аминокислот. Эти диеты поддерживались приблизительно изокалорийными за счет корректировки общего белка плюс усвояемые углеводные компоненты до фиксированного количества, поскольку лечение белковыми диетами варьировалось в испытанных диапазонах. Испытания на кормлении мальков, сеголетков и годовалых рыб показали, что общие потребности в белке наиболее высоки у начальных кормовых мальков и что они уменьшаются по мере увеличения размера рыбы.Чтобы расти с максимальной скоростью, мальки должны иметь диету, в которой почти половина легкоусвояемых ингредиентов состоит из сбалансированного белка; через 6-8 недель это требование снижается примерно до 40 процентов рациона лосося и форели и примерно до 35 процентов рациона годовалых лососевых, выращенных при стандартной температуре окружающей среды (SET). См. Рисунки 1 и 2. Общие потребности в белке молоди сома, по-видимому, меньше, чем у лососевых. Первоначально кормление мальков требует, чтобы около 50 процентов усвояемых компонентов рациона составлял белок, и потребность в них уменьшается с увеличением размера.Некоторые испытания кормления лососем показали прямую связь между изменениями потребности в белке молоди рыбы и изменениями температуры воды. Лосось чавычи в воде с температурой 7 ° C требует около 40 процентов цельного яичного белка для максимального роста; той же рыбе в воде с температурой 15 ° C требуется около 50% белка. Лосось, форель и сом могут использовать больше белка, чем требуется для максимального роста, благодаря эффективному удалению азотистых отходов в виде растворимых соединений аммиака через жаберные ткани непосредственно в водную среду.Эта система удаления азота более эффективна, чем система, доступная для птиц и млекопитающих. Птица и млекопитающие потребляют энергию для синтеза мочевины, мочевой кислоты или других соединений азота, которые выводятся через ткань почек и выводятся с мочой. Перевариваемые углеводы и жиры сохранят избыток белка в рационе до тех пор, пока удовлетворяются потребности в белке для максимального роста (рисунки 1 и 2).
Таблица 1 — Расчетная потребность в белке с пищей для некоторых видов рыб 1/
|
Виды |
Уровень сырого протеина в рационе для оптимального роста (г / кг) |
|
Радужная форель ( Salmo gairdneri ) |
400-460 |
|
Карп ( Cyprinus carpio ) |
380 |
|
Чавыча ( Oncorhynchus tshawytscha ) |
400 |
|
Угорь ( Ангилья японский ) |
445 |
|
Камбала ( Pleuronectes platessa ) |
500 |
|
Золотистый лещ ( Chrysophrys aurata ) |
400 |
|
Белый амур ( Ctenopharyngodon idella ) |
410-430 |
|
Brycon sp. |
356 |
|
Морской лещ ( Chrysophrys major ) |
550 |
|
Желтохвост ( Seriola quinqueradiata ) |
550 |
1/ По материалам C.B. Cowey, 1978
Рис. 1. Потребность в белке чавычи при 47 ° F. Верхняя кривая: начальный индивидуальный средний вес рыбы, 1.5г. Нижняя кривая: исходная индивидуальная средняя масса рыбы 5,6 г.
Рис. 2. Потребность в белке чавычи при температуре 58 ° F. Верхняя кривая: исходный индивидуальный средний вес рыбы 2,6 г. Нижняя кривая: исходная индивидуальная средняя масса рыбы 5,8 г.
(Оба рисунка адаптированы из: DeLong, D.C., J.E. Halver and E.T. Mertz, 1958, J.Nutr ., 65: 589-99)
По сути, рыбе необходимо давать диету, содержащую дифференцированный уровень высококачественного белка и энергии, а также адекватный баланс незаменимых жирных кислот, витаминов и минералов в течение длительного периода времени.Из полученной кривой доза / ответ потребность в белке обычно получают по графику Альмквиста. Считается, что эти различия в очевидной потребности в белке связаны с различиями в методах культивирования и составе рациона.
Относительно высокий уровень пищевого белка, необходимый для максимального роста некоторых рыб, таких как амазон, Ctenopharyngodon idella, и Brycon spp. Удивительны тем, что эти рыбы всеядны. Brycon spp.выращиваются на нежелательных фруктах и другом растительном материале с низким содержанием белка, и в этих условиях, по-видимому, существенный вклад в потребление ими белка вносит естественная пищевая цепь.
Потребность в белке эвриталинных рыб, таких как радужная форель, Salmo gairdneri, и кижуч, Oncorhynchus кисуч, , выращенных в воде с соленостью 20 ppt, примерно такая же, как потребность в пресной воде. Нет данных о потребности этих видов в белке в морской воде с полной концентрацией.(35 п.
4.1 Essential
и заменимые аминокислоты
4.2 Незаменимые
Аминокислоты и качество протеина
Аминокислоты являются строительными блоками белков; около 23 аминокислот были выделены из природных белков. Десять из них незаменимы для рыб. Животное не способно синтезировать незаменимые аминокислоты и поэтому должно получать их с пищей.
4.1 Незаменимые и заменимые аминокислоты
Корм для лосося, форели и канального сома без содержания аргинина, гистидина, изолейцина, лейцина, лизина, метионина, фенилаланина, треонина, триптофана или валина не рос (рис.3). Те же самые рыбы, которых кормили рационами, лишенными других L-аминокислот, росли так же, как и рыбы, получавшие все 18 протестированных аминокислот (рис. 4). Азотный компонент в тестируемых диетах состоял из 18 L-аминокислот по образцу, обнаруженному в цельном яичном протеине. Вся тестируемая рыба быстро выздоравливала, когда в рационе была заменена недостающая аминокислота. Наклон кривой роста в группе восстановления был идентичен таковому у рыб, получавших полный тестовый рацион с аминокислотами.
Заменяемые аминокислоты были протестированы: аланин, аспарагиновая кислота, цистин, глутаминовая кислота, глицин, пролин, серин и тирозин.Было обнаружено, что эти аминокислоты не являются необходимыми для роста лосося, форели и канального сома.
Для количественных исследований требований к 10 незаменимым аминокислотам использовалась смесь казеина и желатина с добавлением кристаллических L-аминокислот. Тестовая диета содержала 40 процентов цельного яичного белка для азотного компонента. Эксперименты, проведенные с карпом и угрем, показали аналогичное отсутствие роста, когда в рационе отсутствовала незаменимая аминокислота.
Рис. 3. Рост рыб с дефицитом аргинина. Группа с дефицитом была разделена через шесть недель на диете с дефицитом, и недостающая аминокислота была заменена в одной из двух частей.
Рис. 4. Рост рыб с дефицитом цистина.
(Оба рисунка взяты из: DeLong, D.C., J.E. Halver and E.T. Mertz, 1958, J.Nutr., 65: 589-99)
4.2 Основные аминокислоты и качество белка
Если известны потребности рыбы в незаменимых аминокислотах, должно быть возможно удовлетворить эти потребности в системах культивирования различными способами за счет различных пищевых белков или комбинаций пищевых белков.
Фенилаланин избавлен от тирозина. Неизвестно, что он химически модифицирован или становится недоступным из-за суровых условий, которым обычно подвергаются кормовые белки во время обработки. Измерение фенилаланина в белках несложно, поэтому обеспечение и оценка фенилаланина в белках в практических диетах не представляет особых трудностей.
Лизин — основная аминокислота. В дополнение к -аминокислотной группе, обычно связанной пептидной связью, он также содержит вторую -аминогруппу.Эта альфа-аминогруппа должна быть свободной и реактивной, иначе лизин, хотя и поддается химическому измерению, не будет доступен биологически. Во время обработки белков корма α-аминогруппа лизина может реагировать с небелковыми молекулами, присутствующими в корме, с образованием дополнительных соединений, которые делают лизин биологически недоступным.
Метионин избавлен от цистина. Однако измерить содержание метионина в кормовых белках непросто, поскольку аминокислота подвержена окислению во время обработки.После обработки метионин может присутствовать как таковой, или в виде сульфоксида, или в виде сульфона. Сульфоксид может образовываться из метионина во время кислотного гидролиза кормового белка перед измерением его кислотного состава, не содержащего кислоты. Кислотный гидролиз белков перед анализом нарушает исходное равновесие между двумя соединениями, так что состав гидролизата больше не отражает состав белка. При определении содержания метионина в чистых белках окисление аминокислоты до метионинсульфона обычно является количественным.Однако в случае кормовых белков это не покажет, сколько метионина или сульфоксида метионина присутствовало в белке до его окисления и гидролиза.
Сульфоксид метионина может иметь некоторую биологическую ценность для рыб, которые могут иметь некоторую способность обратного преобразования его в метионин и, таким образом, частично восполнять часть метионина, окисленного во время обработки.
Недавно появились сообщения о методах измерения метионина в белках с использованием йодоплатинатного реагента до и после восстановления трихлоридом титана, чтобы получить значения как для метионина, так и для сульфоксида в исходном белке.Также был описан способ измерения метионина конкретно по расщеплению цианогенбромида. Оба метода еще предстоит оценить независимо. Микробиологический анализ метионина в белках кормов является ценным инструментом, хотя существует опасность того, что оксиды метионина могут различаться по своей активности в отношении микроорганизмов и искажать значения.
Количественные потребности лососевых в десяти незаменимых аминокислотах определялись путем кормления линейными приращениями одной аминокислоты за раз в тестируемой диете, содержащей аминокислотный профиль, идентичный цельному яичному белку, за исключением тестируемой аминокислоты.Повторяющиеся группы рыб подвергались диетическому лечению до тех пор, пока не появились большие различия в росте опытных партий. График реакции роста Альмквиста показывает уровень аминокислот, необходимый для максимального роста в этих конкретных условиях испытания. Рационы были разработаны таким образом, чтобы содержать белок на уровне или немного ниже оптимальной потребности в белке для данного вида и условий тестирования, чтобы гарантировать максимальное использование ограничивающей аминокислоты. Сравнение требований к десяти незаменимым аминокислотам между видами показано в таблице 2.
Недавним нововведением стало использование в тестовых диетах белков, относительно дефицитных по данной незаменимой аминокислоте. Таким образом, комбинации рыбной муки и зеина использовались в тестовых диетах для определения потребности радужной форели в аргинине. Рационы, содержащие различные относительные количества казеина и желатина, показали, что увеличение уровня связанного с белками аргинина с 11 до 17 г / кг привело к значительному увеличению роста канального сома.
Таблица 2 Потребность семи животных в аминокислотах 1/
|
Аминокислота |
Молодь угря |
Мальки карпа |
Канальный сом |
Молодь чавычи |
Цыпленок |
Молодой поросенок |
Крыса |
|
Аргинин |
3.9 (1,7 / 42) |
4,3 (1,65 / 38,5) |
6,0 (2,4 / 40) |
6,1 (1,1 / 18) |
1,5 (0,2 / 13) |
1,0 (0,2 / 19) |
|
|
Гистидин |
1,9 (0,8 / 42) |
1,8 (0,7 / 40) |
1,7 (0,3 / 18) |
1.5 (0,2 / 13) |
2,1 (0,4 / 19) |
||
|
Изолейцин |
3,6 (1,5 / 42) |
2,6 (1,0 / 38,5) |
2,2 (0,9 / 41) |
4,4 (0,8 / 18) |
4,6 (0,6 / 13) |
3,9 (0,5 / 13) |
|
|
лейцин |
4.1 (1,7 / 42) |
3,9 (1,5 / 38,5) |
3,9 (1,6 / 41) |
6,7 (1,2 / 18) |
4,6 (0,6 / 13) |
4,5 (0,9 / 19) |
|
|
Лизин |
4,8 (2,0 / 42) |
5,1 (1,23 / 24,0) |
5,0 (2,0 / 40) |
6.1 (1.1 / 18) |
4,7 (0,65 / 13) |
5,4 (1,0 / 19) |
|
|
метионин 2/ |
4,5 (2,1 / 42) 3/ |
3,1 (1,2 / 38,5) |
2,3 (0,56 / 24,0) |
4,0 (1,6 / 40) 3/ |
4.4 (0,8 / 18) |
3,0 (0,6 / 20) |
3,0 (0,6 / 20) |
|
Фенилаланин 4/ |
5,1 (2,1 / 41) 5/ |
7,2 (1,3 / 18) |
3.6 (0,45 / 13) |
5,3 (0,9 / 17) |
|||
|
Треонин |
3,6 (1,5 / 42) |
2,2 (0,9 / 40) |
3,3 (0,6 / 18) |
3,0 (0,4 / 13) |
3,1 (0,2 / 19) |
||
|
Триптофан |
1,0 (0,4 / 42) |
0.5 (0,2 / 40) |
1,1 (0,2 / 18) |
0,8 (0,2 / 25) |
1,0 (0,2 / 19) |
||
|
Валин |
3,6 (1,5 / 42) |
3,2 (1,3 / 40) |
4,4 (0,8 / 18) |
3,1 (0,4 / 13) |
3,1 (0,4 / 13) |
1/ Выражается в процентах от диетического белка.В скобках числители — это потребности в процентах от сухого рациона, а знаменатели — это процент общего содержания белка в рационе.
2/ При отсутствии цистина
3/ Метионин плюс цистин
4/ При отсутствии синусоидального синуса
5/ Фенилаланин плюс тирозин
(по материалам: Национальный исследовательский совет, 1977 г.)
Потребность радужной форели в аргинине была определена по стандартной кривой доза / реакция (рост), а также путем измерения уровней свободного аргинина в тканях (крови и мышцах) в группах форели, получавших возрастающее количество аргинина в рационе.После того, как диетическая потребность форели в аргинине была удовлетворена, любое дальнейшее увеличение потребления аргинина привело к увеличению концентрации свободного аргинина в крови и мышцах. Было получено хорошее согласие между двумя методами.
Данные, представленные в таблице 2, предполагают, что между видами рыб существуют реальные различия в их потребностях в определенных аминокислотах. Это приводит к трудностям при составлении белкового компонента практического рациона для тех видов, потребности которых в аминокислотах еще не известны.Возможное решение — использовать для каждой аминокислоты наивысший уровень, необходимый для любого из тех видов, по которым имеются данные. Необходимость дополнительных количественных данных о потребностях рыб в аминокислотах, особенно тех, которые действительно или потенциально могут использоваться в качестве сельскохозяйственных животных, очевидна.
Одним из решений использования белков, относительно дефицитных по одной или нескольким аминокислотам, является добавление к белку соответствующих количеств аминокислоты, необходимых в практических диетах. Рыба, по-видимому, использует свободные аминокислоты с разной степенью эффективности.
Молодой карп, Cyprinus carpio, , как было показано, неспособен расти на диетах, в которых белковый компонент (казеин, желатин) был заменен смесью аминокислот, аналогичных по общему составу. Гидролизат трипсина казеина также оказался неэффективным. Однако, если диета, содержащая свободные аминокислоты в качестве белкового компонента, тщательно нейтрализуется NaOH до pH 6,5-6,7, то некоторый рост молоди карпа действительно происходит. Этот рост был заметно ниже, чем при сопоставимой казеиновой диете в тех же условиях.
Канальный сом также не может использовать свободные аминокислоты в качестве добавок к дефицитным белкам. При изонитрогенной замене соевого шрота на муку менхадена рост и эффективность корма канального сома существенно снизились. Добавление свободного метионина, цистина или лизина, наиболее ограничивающих аминокислот, к этим заменителям сои не привело к увеличению веса.
Повышение уровня аргинина в рационах сома с 11 до 17 г / кг путем изонитрогенной замены желатина на казеин значительно увеличивало набор веса, но добавление свободного аргинина, цистина, триптофана или метионина к казеину мало влияло на рост или преобразование пищи.
Лососевые могут использовать свободные аминокислоты для роста. Было показано, что зеин-желатиновая диета с добавлением лизина и тритофана заметно превосходит зеин-желатиновую диету для радужной форели, когда в качестве критериев использовались прибавка в весе и использование белка.
Несколько исследователей продемонстрировали возможность дополнения белков с дефицитом аминокислот ограничивающими аминокислотами в рационах лососевых. Казеин с добавкой шести аминокислот давал коэффициенты конверсии корма для атлантического лосося, аналогичные тем, которые были получены при использовании изолированного рыбного белка в качестве источника пищевого белка.Соевый шрот с добавлением пяти или более аминокислот (включая метионин и лизин) был лучшим источником белка для радужной форели по сравнению с соевым шротом. Однако однократное добавление метионина и лизина не привело к повышению ценности соевого шрота. Эти результаты позволяют предположить, что аминокислотный спектр выделенного рыбьего белка, который они использовали, может приблизительно соответствовать потребности в аминокислотах радужной форели. Пищевая ценность изолята соевого белка может быть увеличена путем добавления в него первой ограничивающей аминокислоты; я.е., метионин.
Рационы, содержащие в качестве белкового компонента рыбную муку, мясокостную муку, а также дрожжевую и соевую муку, можно улучшить, добавляя цистин (10 г / кг) и триптофан (5 г / кг) вместе. Рыбную муку можно полностью заменить без снижения конверсии корма в рационах для радужной форели смесью из субпродуктов домашней птицы и перьевой муки вместе с 17 г лизина HCL / кг, 4,8 г DL-метионина / кг и 1,44 г DL. -триптофан / кг.
Коуи, К.Б. и Дж. Р. Сардженты, 1972 Кормление рыб. Adv. Mar.Biol., 10: 383-492
Cowey, C.B., 1979 Потребности рыб в белках и аминокислотах. В Технология кормления и кормления рыб, под редакцией Дж. Э. Халвера и К. Тьюса. Материалы Всемирного симпозиума, спонсируемого EIFAC / FAO, ICES и IUNS, Гамбург, 20-23 июня 1978 г. Schr . Bundesforschungsanst . Fisch ., Hamb ., (14/15) vol. 1: 3-16
Мерц, Э.Т., 1972 г. Потребности в белке и аминокислотах. В Питание рыб, под редакцией Дж. Э. Халвера. Нью-Йорк, Academic Press, стр. 106-43.
Национальный исследовательский совет, Подкомитет по тепловодным рыбам 1977 года, Потребность теплопроводных рыб в питательных веществах. Вашингтон, округ Колумбия, Национальная академия наук (потребности домашних животных в питательных веществах) 78 стр.
Новые исследования в области секвенирования белков готовы изменить медицину
7 июня 2021 г.
В то время как ДНК представляет собой книгу генетических рецептов для биологических форм и функций, работа белков организма — выполнять сложные команды, продиктованные генетическим кодом ДНК.
Стюарт Линдсей, исследователь из Института биодизайна при Университете штата Аризона, был в авангарде усилий по совершенствованию быстрого секвенирования ДНК и недавно применил свои таланты для исследования гораздо более сложной проблемы секвенирования белковых молекул, одна молекула за молекулой. время.
Белки состоят из 20 аминокислот, которые складываются в сложные трехмерные формы. Здесь представлены три возможных представления трехмерной структуры протеинтриозофосфат-изомеразы.Изображение предоставлено Opabinia regalis.
Скачать полное изображение
В новой обзорной статье описываются усилия Линдси, наряду с усилиями международных коллег, по применению множества инновационных стратегий для секвенирования белков на одноклеточном и даже одномолекулярном уровне.
«Одним из сюрпризов проекта« Геном человека »было то, что люди могут быть построены всего из 25 000 генов», — сказал Линдси. «Настоящая сложность заключается в почти бесконечном количестве способов, которыми белки могут быть собраны вместе из этого. ограниченный набор генов и способы, которыми отдельные аминокислоты — строительные блоки белков — могут быть химически модифицированы. По этим причинам нам нужны инструменты для анализа белков на уровне одной молекулы ».
Линдси — директор Центра биодизайна биофизики одномолекул, президентского кафедры физики им. Эдварда и Надин Карсон в АГУ и профессора Риджентс Колледжа свободных искусств и наук в области химии и биохимии.
Стюарт Линдсей — директор Центра биодизайна биофизики одномолекул, президентского кафедры физики им. Эдварда и Надин Карсон в АГУ и профессора Риджентс в Колледже свободных искусств и наук.
Обзор проводился совместно Хавьером Альфаро, Гданьский университет, Польша; Амит Меллер, Технион – Израильский технологический институт, Хайфа, Израиль; Чирлмин Джу, Делфтский технологический университет, Нидерланды; и Адам Поморски, Вроцлавский университет, Польша.
Исследование опубликовано в текущем номере журнала Nature Methods.
Хотя клетки могут внешне казаться идентичными, на самом деле они представляют собой уникальные сущности с индивидуальными свойствами, которые могут резко отличаться, что может иметь важные последствия для здоровья и болезней. Поскольку белки выполняют большую часть работы внутри клетки, исследование межклеточных вариаций в органах, тканях и культурах клеток может дать критически важную информацию, особенно для изучения рака, когда отдельные клеточные мутации могут усиливаться, что способствует развитию болезнь.
В области исследования белков (известных как протеомика) традиционно доминировали масс-спектрометрические технологии, которые являются дорогостоящими и, как правило, только последовательностями белков партии, содержащимися в сложных смесях. Выявление отдельных белков в отдельных клетках потребует преобразования технологии секвенирования, аналогичной революции в секвенировании ДНК. По словам Альфаро, «проблема в том, что, в отличие от ДНК, нет способа амплифицировать белки, и это представляет собой серьезную проблему для чувствительности современного оборудования.
Подход Линдси к этой проблеме включает метод, известный как туннельное распознавание, основанный на технологии, известной как сканирующая туннельная микроскопия (СТМ). В более ранних исследованиях Линдсей применил СТМ для идентификации оснований ДНК, последовательно пропускаемых через крошечное отверстие, известное как нанопора, как будто нить можно пропустить через игольное ушко.
Когда нуклеотиды ДНК проходят через нанопору, они на мгновение удерживаются в плену между парой электродов. Ток, проходящий через каждое основание, известный как туннельный ток, дает сигнал. Сказать электрические шипы, которые похожи на отпечатки пальцев каждого из четырех оснований ДНК: аденина, тимина, цитозина и гуанина.
Последние достижения позволили применить ту же основную идею к идентификации белков путем последовательной идентификации 20 аминокислот, из которых построены все белки. В то время как распознавание туннелирования белков следует тому же базовому плану игры, который используется для секвенирования оснований ДНК, задача для секвенирования белков гораздо сложнее, требуя связывания аминокислот со специализированными молекулами адаптера для прохождения туннельного тока, а также алгоритмов машинного обучения чтобы помочь выбрать точную последовательность аминокислот.