Метаболическая функция — Большая Энциклопедия Нефти и Газа, статья, страница 1
Метаболическая функция
Cтраница 1
Метаболическая функция даже — простейших клеток включает несколько тысяч взаимосвязанных химических реакций и, следовательно, требует тонкого механизма для их координации и управления ими. Иначе говоря, необходима необычайно тонкая и сложная функциональная организация. Кроме того, метаболические реакции требуют специфических катализаторов — энзимов. Это — гигантские молекулы, обладающие пространственной организацией, и организм должен уметь синтезировать их. Напомню, что катализатором мы называем вещество, которое ускоряет некую химическую реакцию, но не расходуется в ходе ее. Каждый энзим, или катализатор, выполняет одну специфическую задачу.
[2]
Важными метаболическими функциями аскорбиновой кислоты являются расщепление тирозина и лизина и гидроксилирование пролина и допамина.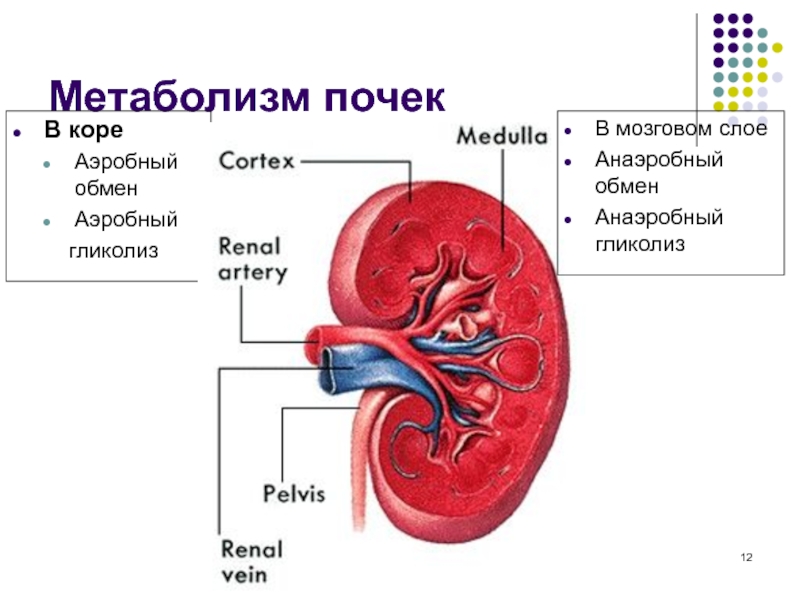 Она участвует также в обмене липидов.
Она участвует также в обмене липидов.
[3]
Какие метаболические функции выполняют почки.
[4]
Известно, что фермент может выполнять метаболическую функцию после приобретения соответствующей структуры.
[5]
Известно, что токоферолы выполняют в организме две главные метаболические функции. Во-первых, они являются наиболее активными и, возможно, главными природными жирорастворимыми антиоксидантами: разрушают наиболее реактивные формы кислорода и соответственно предохраняют от окисления полиненасыщенные жирные кислоты. Во-вторых, токоферолы играют специфическую, пока еще не полностью раскрытую роль в обмене селена. Селен, как известно, является интегральной частью глутатионпероксидазы — фермента, обеспечивающего защиту мембран от разрушающего действия пероксидных радикалов. Биологическая роль витамина Е сводится, таким образом, к предотвращению аутоокисления липидов биомембран и возможному снижению потребности в глутатионпероксидазе, необходимой для разрушения образующихся в клетке перекисей. Участие токоферолов в механизме транспорта электронов и протонов, как и в регуляции процесса транскрипции генов, и их роль в метаболизме убихинонов пока недостаточны выяснены.
Участие токоферолов в механизме транспорта электронов и протонов, как и в регуляции процесса транскрипции генов, и их роль в метаболизме убихинонов пока недостаточны выяснены.
[6]
Мп, В, Со, Си, Si — необходимые для других метаболических функций.
[7]
Однако новые данные о роли ДНК не только в репродуктивных, но и в метаболических функциях клетки выдвигают, как нам кажется, некоторые новые аспекты в проблеме действия радиации на нуклеиновые кислоты и их обмен.
[8]
В основе действия этих соединений на микробную клетку лежит, по-видимому, нарушение энзиматических или метаболических функций, связанных с репродукцией клеток, или торможение окислительных процессов, связанных, по данным Колдуэлла и Хиншел-вуда [74], с синтезом производных рибозы, например дезоксирибозы.
[9]
В настоящей работе предлагается познакомиться с основными закономерностями активного транспорта Са2 и связью этого процесса с метаболической функцией митохондрий печени крысы.
[10]
Кость — это основной материал, из которого построен скелет большинства позвоночных животных; она несет опорные, защитные и некоторые метаболические функции. Клетки кости погружены в плотный кальцифицированный матрикс, состоящий примерно на 30 % из органических соединений, главным образом из коллагеновых волокон и гликопротеинов, и на 70 % — из неорганических солей. Основной неорганический компонент кости составляют игловидные кристаллы гидроксиапатита, Саю ( РО4) е ( ОН) 2 — одной из форм фосфата кальция. В ней содержатся также в различных количествах натрий, магний, калий, хлор, фтор, гидрокарбонат — и цитрат-ионы.
[11]
Биотип ( или витамин Н, когда речь идет о человеческом организме) является необходимым кофактором ряда ферментов, которые несут различные метаболические функции. Более десятка различных ферментов используют биотин. К наиболее известным относятся ацетил — СоА — карбоксилаза, пируватдекарбоксилаза, про-пионил — СоА — карбоксилаза, карбоксилаза мочевины, метилмало-нил — СоА — декарбоксилаза и Оксалоацетатдекарбоксилаза. Биотин служит ковалентным переносчиком СО2 в реакциях, в которых С02 фиксируется на акцепторе с помощью карбоксилаз. Затем карбоксильная группа в независимой реакции переносится транс-карбоксилазами от одного акцепторного субстрата к другому или же может удаляться в виде СО2 под действием декарбоксилазы.
Более десятка различных ферментов используют биотин. К наиболее известным относятся ацетил — СоА — карбоксилаза, пируватдекарбоксилаза, про-пионил — СоА — карбоксилаза, карбоксилаза мочевины, метилмало-нил — СоА — декарбоксилаза и Оксалоацетатдекарбоксилаза. Биотин служит ковалентным переносчиком СО2 в реакциях, в которых С02 фиксируется на акцепторе с помощью карбоксилаз. Затем карбоксильная группа в независимой реакции переносится транс-карбоксилазами от одного акцепторного субстрата к другому или же может удаляться в виде СО2 под действием декарбоксилазы.
[12]
Одной из основных проблем современной биохимии является выяснение механизма превращения энергии, выделяющейся в результате взаимодействия связей С — Н с кислородом с образованием двуокиси углерода и воды в энергию фосфоангидридной связи АТФ — единой платежной единицы в процессах переноса химической энергии, используемой для большого числа синтетических и метаболических функций. Если энергетическое сопряжение имеет химический механизм ( хотя это еще не очевидно [185]), то оно может происходить либо непосредственно через окисление некоторых легко образующихся низкоэнергетических фосфатных производных до высокоэнергетических форм, которые могут затем переносить фосфат на АДФ, давая АТФ, либо через окисление некоторых других низкоэнергетических молекул до высокоэнергетических форм, которые могут дать макроэргический фосфат через серию реакций переноса. В последнее время стало известно несколько примеров такого актива-ционного процесса, в котором происходит образование высокоэнергетического тиолового эфира при окислении альдегида. Тиоловый эфир может реагировать дальше, давая ацилфосфат и при известных обстоятельствах АТФ. Этот тип активации является ответственным за образование макро-эргических фосфатных связей на субстратном уровне фосфорилирования, в котором метаболит, подвергающийся окислению, превращается в активированный продукт. В настоящее время, однако, еще нет уверенности, что аналогичный процесс происходит при многоступенчатом переносе электронов между субстратом и кислородом, который является ответственным за освобождение большей части энергии в аэробном метаболизме.
Если энергетическое сопряжение имеет химический механизм ( хотя это еще не очевидно [185]), то оно может происходить либо непосредственно через окисление некоторых легко образующихся низкоэнергетических фосфатных производных до высокоэнергетических форм, которые могут затем переносить фосфат на АДФ, давая АТФ, либо через окисление некоторых других низкоэнергетических молекул до высокоэнергетических форм, которые могут дать макроэргический фосфат через серию реакций переноса. В последнее время стало известно несколько примеров такого актива-ционного процесса, в котором происходит образование высокоэнергетического тиолового эфира при окислении альдегида. Тиоловый эфир может реагировать дальше, давая ацилфосфат и при известных обстоятельствах АТФ. Этот тип активации является ответственным за образование макро-эргических фосфатных связей на субстратном уровне фосфорилирования, в котором метаболит, подвергающийся окислению, превращается в активированный продукт. В настоящее время, однако, еще нет уверенности, что аналогичный процесс происходит при многоступенчатом переносе электронов между субстратом и кислородом, который является ответственным за освобождение большей части энергии в аэробном метаболизме. Интерес к этой проблеме стимулировал поиски реакций, в которых фосфатная группа превращается в энергетически богатую форму посредством окислительного процесса, что может служить моделью реакций с природным коферментом. Хотя в настоящее время еще нет доказательств, что какой-либо процесс такого рода ответствен за окислительное фосфорилирование, эти исследования интересны с химической точки зрения и в качестве источника некоторых потенциально полезных синтетических методов.
Интерес к этой проблеме стимулировал поиски реакций, в которых фосфатная группа превращается в энергетически богатую форму посредством окислительного процесса, что может служить моделью реакций с природным коферментом. Хотя в настоящее время еще нет доказательств, что какой-либо процесс такого рода ответствен за окислительное фосфорилирование, эти исследования интересны с химической точки зрения и в качестве источника некоторых потенциально полезных синтетических методов.
[13]
Автокатализ ( присутствие вещества X ускоряет процесс образования его в результате реакции), автоингибиция ( присутствие вещества X блокирует катализ, необходимый для производства X) и кросс-катализ ( каждое из двух веществ, принадлежащих различным цепям реакций, является катализатором для синтеза другого) лежат в основе классического механизма регуляции, обеспечивающего согласованность метаболической функции.
[14]
НАД ( Ф) и НАД ( Ф) Н в печени, например, превышает 1 мкмоль / г сырого веса ткани. Пиридиннуклеотиды выполняют важную метаболическую функцию, являясь кофакторами большого числа дегидрогеназ.
Пиридиннуклеотиды выполняют важную метаболическую функцию, являясь кофакторами большого числа дегидрогеназ.
[15]
Страницы:
1
2
3
4
МЕТАБОЛИЧЕСКАЯ ФУНКЦИЯ
|
biokhimija.ru |
Тимин О.А. Лекции по биологической химии |
347 |
|
|
|
|
ФУНКЦИИ ПЕЧЕНИ
1.Пищеварительная – печень является крупнейшей пищеварительной железой. Она образует желчь, включающую воду (82%), желчные кислоты (12%), фосфатидилхолин (4%), холестерол (0,7%), прямой билирубин, белки, продукты распада стероидных гормонов, электролиты, лекарственные средства и их метаболиты.
Желчь обеспечивает эмульгирование и переваривание жиров пищи, стимулирует пери-
стальтику кишечника.
Из крови воротной вены желчные кислоты поглощаются симпортом с ионами Na+. В желчный капилляр синтезированные de novo и используемые вторично желчные кислоты секретируются АТФ-зависимым транспортом.
2.Экскреторная функция, близка к пищеварительной – с помощью желчи выводятся билирубин, немного креатинина и мочевины, ксенобиотики и продукты их обезвреживания, холестерол. Последний выводится из организма только в составе желчи.
3.Секреторная – печень осуществляет биосинтез и секрецию в кровь альбумина и некоторых белков других фракций, белков свертывающей системы, липопротеинов, глюкозы, кетоновых тел, 25-оксикальциферола, креатина.
4.Депонирующая – здесь находится место депонирования энергетических резервов
гликогена, накапливаются минеральные вещества, особенно железо, витамины A, D, K, B12 и фолиевая кислота.
5.Метаболическая функция (подробно см ниже)
6.Обезвреживающая функция (подробно см ниже)
Печень является центральным органом метаболизма.
Углеводный обмен
В гепатоцитах активно протекают процессы углеводного обмена. Благодаря синтезу и распаду гликогена печень поддерживает концентрацию глюкозы в крови. Активный синтез
|
Биохимия печени |
348 |
|
|
|
гликогена происходит после приема пищи, когда концентрация глюкозы в крови воротной вены достигает 20 ммоль/л. Запасы гликогена в печени составляют от 30 до 100 г. При кратковременном голодании происходит гликогенолиз, в случае длительного голодания основным источником глюкозы крови является глюконеогенез из аминокислот и глицерина.
Печень осуществляет взаимопревращение сахаров, т.е. превращение гексоз (фруктозы, галактозы) в глюкозу.
Активные реакции пентозофосфатного пути обеспечивают наработку НАДФН, необходимого для микросомального окисления и синтеза жирных кислот из глюкозы.
Липидный обмен
Если во время приема пищи в печень поступает избыток глюкозы, который не используется для синтеза гликогена и других синтезов, то она превращается в липиды – холестерол и триацилглицеролы. Поскольку запасать ТАГ печень не может, то их удаление происходит при помощи липопротеинов очень низкой плотности (ЛПОНП). Холестерол используется, в первую очередь, для синтеза желчных кислот, также он включается в состав липопротеинов низкой плотности (ЛПНП) и ЛПОНП.
Поскольку запасать ТАГ печень не может, то их удаление происходит при помощи липопротеинов очень низкой плотности (ЛПОНП). Холестерол используется, в первую очередь, для синтеза желчных кислот, также он включается в состав липопротеинов низкой плотности (ЛПНП) и ЛПОНП.
При определенных условиях – голодание, длительная мышечная нагрузка, сахарный диабет I типа, богатая жирами диета – в печени активируется синтез кетоновых тел, используемых большинством тканей как альтернативный источник энергии.
Белковый обмен
Больше половины синтезируемого за сутки в организме белка приходится на печень. Скорость обновления всех белков печени составляет 7 суток, тогда как в других органах эта величина соответствует 17 суткам и более. К ним относятся не только белки собственно гепатоцитов, но и идущие на «экспорт» – альбумины, многие глобулины, ферменты крови, а
также фибриноген и факторы свертывания крови.
Аминокислоты подвергаются трансаминированию, дезаминированию, декарбоксилированию с образованием биогенных аминов. Происходят реакции синтеза холина и креатина благодаря переносу метильной группы от аденозилметионина. В печени идет утилизация избыточного азота и включение его в состав мочевины.
Происходят реакции синтеза холина и креатина благодаря переносу метильной группы от аденозилметионина. В печени идет утилизация избыточного азота и включение его в состав мочевины.
Мышцы метаболическая функция — Справочник химика 21
Участие компонентов биомембран в осуществлении и регулировании метаболических процессов в клетке. Общая характеристика процессов передачи информации в клетке. Понятие о первичных и вторичных мессенджерах. Классификация, особенности структурно-функциональной организации мембранных белков-рецепторов. Характеристика аденилатциклазного и фосфо-инозитидного пути передачи сигнала в клетку. Роль ионов в осуществлении метаболических процессов с участием мембран. Адсорбционный тип регуляции метаболизма. Понятие о метаболоне, физиологическое значение его образования. Пространствен-но-структурная организация ферментных систем клетки (на примере гликолитического комплекса и цикла Кребса), Экспериментальные исследования взаимодействия ферментов гликолиза с различными структурными компонентами клетки. Модели структуры гликолитического комплекса в скелетных мышцах и на внутренней поверхности мембран эритроцитов. Эстафетный механизм работы ферментов в клетке. Механизмы регулирования функциональной активности векторных ферментов биомембран. Пути нейрогуморальной регуляции функций клеток. [c.284]
Модели структуры гликолитического комплекса в скелетных мышцах и на внутренней поверхности мембран эритроцитов. Эстафетный механизм работы ферментов в клетке. Механизмы регулирования функциональной активности векторных ферментов биомембран. Пути нейрогуморальной регуляции функций клеток. [c.284]
Железо функционирует как основной переносчик электронов в биологических реакциях окисления — восстановления [231]. Ионы железа, и Fe +, и Fe +, присутствуют в человеческом организме и, действуя как переносчики электронов, постоянно переходят из одного состояния окисления в другое. Это можно проиллюстрировать на примере цитохромов . Ионы железа также служат для транспорта и хранения молекулярного кислорода — функция, необходимая для жизнедеятельности всех позвоночных животных. В этой системе работает только Ре(П) [Fe(111)-гемоглобин не участвует в переносе кислорода]. Чтобы удовлетворить потребности метаболических процессов в кислороде, большинство животных имеет жидкость, циркулирующую по телу эта жидкость и переносит кислород, поглощая его из внешнего источника, в митохондрии тканей.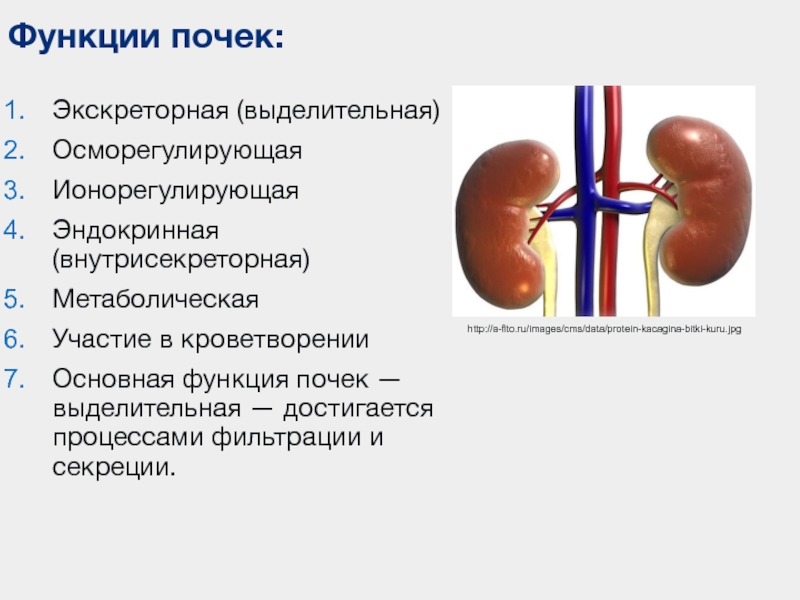 Здесь он необходим для дыхательной цепи, чтобы обеспечивать окислительное фосфорилирование и производство АТР. Одиако растворимость кислорода в воде слишком низка для поддержания дыхания у живых существ. Поэтому в состав крови обычно входят белки, которые обратимо связывают кислород. Эти белковые молекулы способствуют проникновению кислорода в мышцы (ткани), а также могут служить хранилищем кислорода. [c.359]
Здесь он необходим для дыхательной цепи, чтобы обеспечивать окислительное фосфорилирование и производство АТР. Одиако растворимость кислорода в воде слишком низка для поддержания дыхания у живых существ. Поэтому в состав крови обычно входят белки, которые обратимо связывают кислород. Эти белковые молекулы способствуют проникновению кислорода в мышцы (ткани), а также могут служить хранилищем кислорода. [c.359]
Белки выполняют в организме различные функции и в соответствии с этим обладают разными свойствами. Так, белки, из которых построены ткани (кожа, сухожилия, мышцы и т. д.), ногти и волосы, не растворяются в воде, в то время как белки, участвующие в метаболических процессах (например, белки плазмы крови), растворяются в воде или в водных растворах солей. [c.192]
Биохимические функции. Глюкагон является гормоном-антагонистом инсулина. Он стимулирует гликогенолиз и липолиз, а также активирует процесс глюконеогенеза. Глюкагон взаимодействует с клетками-мишенями по мембрано-опосредованному механизму (гл. 11). Через вторичный посредник — цАМФ он активирует протеинкиназу, киназу фосфорилазу и фосфорилазу Ь, что приводит к мобилизации глюкозы из гликогена. Как и инсулин, глюкагон регулирует метаболические процессы преимущественно в печени, мышцах и жировой ткани. [c.167]
11). Через вторичный посредник — цАМФ он активирует протеинкиназу, киназу фосфорилазу и фосфорилазу Ь, что приводит к мобилизации глюкозы из гликогена. Как и инсулин, глюкагон регулирует метаболические процессы преимущественно в печени, мышцах и жировой ткани. [c.167]
В большинстве рассмотренных нами примеров эндотермии тепло, используемое для регуляции температуры тела, было побочным продуктом той или иной рабочей функции, например мышечного сокращения. Очевидно, что основное назначение работы таких органов, как мышцы, — это не образование тепла для целей терморегуляции. Эндотермия (или необходимый для нее энергетический потенциал) является добавочным продуктом метаболических реакций, лежащих в основе таких функций, как сокращение или перенос ионов. [c.239]
Все ЭТИ функции имеют тенденцию удалить глюкозу из кровяного русла. Согласно последним данным, инсулин функционирует как привратник клеточной стенки. В отсутствие инсулина молекула глюкозы не может проходить сквозь клеточную стенку. Она, однако, легко проходит через нее в присутствии инсулина. Если инсулин почему-либо отсутствует (например, при сахарном диабете, который разбирается более подробно в гл. 20), то использование глюкозы серьезно нарушается и избыточные ее количества остаются в крови (гипергликемия). В этом случае обычно уровень сахара в крови повышается до уровня почечного порога и глюкоза появляется в моче (глюкозурия). Для усвоения глюкозы клетками мозга или сердечной мышцы инсулин не нужен. Поэтому его отсутствие непосредственно на этих тканях не сказывается. Потеря глюкозы (обусловленная отсутствием инсулина) клетками скелетных мышц и печени также вначале большой опасности не представляет, так как эти клетки в случае крайней необходимости люгут получать энергию за счет других источников. Однако продолжительное отсутствие инсулина (как, например, при сахарном диабете) приводит уже к кризису, т. е. к резкому нарушению вышеуказанного метаболического баланса организма, и в конце концов наступает смерть, если только не ввести в организм инсулин.
В отсутствие инсулина молекула глюкозы не может проходить сквозь клеточную стенку. Она, однако, легко проходит через нее в присутствии инсулина. Если инсулин почему-либо отсутствует (например, при сахарном диабете, который разбирается более подробно в гл. 20), то использование глюкозы серьезно нарушается и избыточные ее количества остаются в крови (гипергликемия). В этом случае обычно уровень сахара в крови повышается до уровня почечного порога и глюкоза появляется в моче (глюкозурия). Для усвоения глюкозы клетками мозга или сердечной мышцы инсулин не нужен. Поэтому его отсутствие непосредственно на этих тканях не сказывается. Потеря глюкозы (обусловленная отсутствием инсулина) клетками скелетных мышц и печени также вначале большой опасности не представляет, так как эти клетки в случае крайней необходимости люгут получать энергию за счет других источников. Однако продолжительное отсутствие инсулина (как, например, при сахарном диабете) приводит уже к кризису, т. е. к резкому нарушению вышеуказанного метаболического баланса организма, и в конце концов наступает смерть, если только не ввести в организм инсулин. [c.385]
[c.385]
Клетка регулирует функции митохондрий и более обычными способами. У млекопитающих главным метаболическим путем переработки азотсодержащих продуктов обмена служит цикл мочевины. Образующаяся при этом мочевина выводится с мочой. Ферменты, кодируемые ядерным геномом, катализируют несколько этапов этого цикла в митохондриальном матриксе. Мочевина образуется лишь в некоторых органах, таких как печень, и ферменты цикла мочевины синтезируются и переходят в митохондрии только в этих органах. Кроме того, дыхательные ферментные комплексы, входящие в состав внутренней митохондриальной мембраны, у млекопитающих содержат несколько тканеспецифических субъединиц, которые кодируются ядром и, вероятно, действуют как регуляторы переноса электронов. Например, > некоторых людей с наследственным заболеванием мышц одна из субъединиц цитохромоксидазы дефектна поскольку эта субъединица специфична для скелетных мышц, волокна сердечной мышцы у этих людей функционируют нормально, что позволяет таким больным выживать Как и следовало ожидать, тканеспецифические различия свойственны и хлоропластным белкам, кодируемым ядерными генами [c. 497]
497]
Установление факта клеточного строения нервной системы еще не означало раскрытия механизмов ее функции. Этот факт мог служить лишь отправной точкой. Как следует из второй половины данного нами определения нейробиологии, задача заключается в том, чтобы понять, как нервные клетки организуются в функциональные системы. Обратимся снова к рис. 1.2 и сравним организацию клеток в других тканях организма. Для железистых органов, например печени, основными функциями должны быть метаболическая и секреторная активность отдельных клеток пространственное расположение клеток таких органов важно только с точки зрения транспорта веществ между клетками и кровью. В других же тканях, например коже, мышцах и кости, на первый план выступают механические факторы, а в таких органах, как легкие или почка, комбинируются метаболические и механические функции. [c.29]
Характерная функция ионов Са + у живых существ состоит в способности активировать различные метаболические процессы. Это происходит при резких -изменениях проницаемости плазматических мембран или мембран эндоплазматического ретикулума, в результате которых становится возможной диффузия ионов Са + в цитоплазму. Так, например, при сокращении мышцы в результате освобождения ионов Са + из эндоплазматич0окого ретикулума его концентрация увеличивается приблизительно от 0,1 до 10 мкМ . Связывание ионов Са + с тропонином С инициирует сокращение (гл. 4, разд. Е.1) . Мембраны эндоплазматического ретикулума мышечного волокна содержат большое количество белка кальциевого пасоса, а также ряд белков, связывающих кальций (гл. 4, разд. В.8.в) . Один из Са +нсвязывающих белков мышцы кролика, кальсеквестрин (мол. вес 46 500), способен связывать до 43 молей Са + на моль белка» [c.373]
Это происходит при резких -изменениях проницаемости плазматических мембран или мембран эндоплазматического ретикулума, в результате которых становится возможной диффузия ионов Са + в цитоплазму. Так, например, при сокращении мышцы в результате освобождения ионов Са + из эндоплазматич0окого ретикулума его концентрация увеличивается приблизительно от 0,1 до 10 мкМ . Связывание ионов Са + с тропонином С инициирует сокращение (гл. 4, разд. Е.1) . Мембраны эндоплазматического ретикулума мышечного волокна содержат большое количество белка кальциевого пасоса, а также ряд белков, связывающих кальций (гл. 4, разд. В.8.в) . Один из Са +нсвязывающих белков мышцы кролика, кальсеквестрин (мол. вес 46 500), способен связывать до 43 молей Са + на моль белка» [c.373]
Активный транспорт в таких клетках, как мышцы или эритроциты, в действительности не связан с механизмом, рассмотренным выше в рамках модели Хирона — Пригожина. Обычно считается, что сопряжение является свойством самой мембраны, связанным с ферментами, которые составляют ее неотъемлемую часть. В случае такой мембраны, как кожа лягушки, где обе стороны эпителия легко доступны, метаболическая реакция протекает, несомненно, внутри мембраны. Простая модель такой системы, основанная на стационарном сопряжении, исследовалась экспериментально и теоретически Блюмента-лем, Кепленом и Кедем [1]. Эта модель состоит из последовательной комбинации двух мембран, одна из которых обладает катионообменными, а другая — анионообменными свойствами, с заключенным между ними тонким слоем водного раствора фермента. Эта система при наличии снаружи соответствующего субстрата способна создавать электрический ток между двумя идентичными растворами. В стационарном состоянии ее диссипативная функция имеет сокращенную форму [c.36]
В случае такой мембраны, как кожа лягушки, где обе стороны эпителия легко доступны, метаболическая реакция протекает, несомненно, внутри мембраны. Простая модель такой системы, основанная на стационарном сопряжении, исследовалась экспериментально и теоретически Блюмента-лем, Кепленом и Кедем [1]. Эта модель состоит из последовательной комбинации двух мембран, одна из которых обладает катионообменными, а другая — анионообменными свойствами, с заключенным между ними тонким слоем водного раствора фермента. Эта система при наличии снаружи соответствующего субстрата способна создавать электрический ток между двумя идентичными растворами. В стационарном состоянии ее диссипативная функция имеет сокращенную форму [c.36]
Железо играет ключевую роль во многих метаболических процессах, начиная от переноса кислорода гемоглобином в крови позвоночных животных и кончая центральной ролью в цитохромных структурах. Соответственно минералы железа служат для осуществления столь же широкого ряда функций, включающего перенос и запасание железа, удаление железосодержащих отходов жизнедеятельности, укрепление зубов и ориентацию на местности. Перенос и запасание железа представляют собой, по-видимому, основную функцию минерала ферри-гидрита, образующего, как отмечалось выше, сердцевинную мицеллу сохраняющего железо белка-ферритина. Механизм запасания железа необходим даже организмам, не имеющим в качестве переносчика кислорода гемоглобина. Например, хотя наутилусы и используют для переноса кислорода медьсодержащий белок гемоцианин, их ротовые мышцы содержат миоглобин и, следовательно, железо. Минералы железа служат также для укрепления поверхностного слоя зубцов, покрывающих радулу у хитонов и блюдечек. Животные этих двух групп, живущие в тропических и субтропических водах, используют свои минерализованные зубцы при питании обитающими в каменистом субстрате бактериями и водорослями. Как теперь принято считать, именно они вызывают сильное разрушение береговых утесов на [c.23]
Перенос и запасание железа представляют собой, по-видимому, основную функцию минерала ферри-гидрита, образующего, как отмечалось выше, сердцевинную мицеллу сохраняющего железо белка-ферритина. Механизм запасания железа необходим даже организмам, не имеющим в качестве переносчика кислорода гемоглобина. Например, хотя наутилусы и используют для переноса кислорода медьсодержащий белок гемоцианин, их ротовые мышцы содержат миоглобин и, следовательно, железо. Минералы железа служат также для укрепления поверхностного слоя зубцов, покрывающих радулу у хитонов и блюдечек. Животные этих двух групп, живущие в тропических и субтропических водах, используют свои минерализованные зубцы при питании обитающими в каменистом субстрате бактериями и водорослями. Как теперь принято считать, именно они вызывают сильное разрушение береговых утесов на [c.23]
МЕТАБОЛИЧЕСКАЯ ФУНКЦИЯ ПОЧЕК
1. Образование активной формы витамина D3. В почках в результате микросомального окисления происходит заключительный этап созревания активной формы витамина D3 – 1,25-диоксихолекальциферола, который синтезируется в коже под действием ультрафиолетовых лучей из холестерина, и затем гидроксилируется: сначала в печени (в положении 25), а затем в почках (в положении 1). Таким образом, участвуя в образовании активной формы витамина D3 , почки оказывают влияние на фосфорно-кальциевый обмен в организме. Поэтому при заболеваниях почек, когда нарушаются процессы гидроксилирования витамина D3 , может развиться остеодистрофия.
Таким образом, участвуя в образовании активной формы витамина D3 , почки оказывают влияние на фосфорно-кальциевый обмен в организме. Поэтому при заболеваниях почек, когда нарушаются процессы гидроксилирования витамина D3 , может развиться остеодистрофия.
2. Регуляция эритропоэза. В почках вырабатывается гликопротеин, названный почечным эритропоэтическим фактором (ПЭФ или эритропоэтин). Это гормон, который способен оказывать воздействие на стволовые клетки красного костного мозга, являющиеся клетками-мишенями для ПЭФ. ПЭФ направляет развитие этих клеток по пути эритропоэза, т.е. стимулирует образование эритроцитов. Скорость выделения ПЭФ зависит от обеспечения почек кислородом. Если количество поступающего кислорода снижается, то увеличивается выработка ПЭФ – это ведет к увеличению количества эритроцитов в крови и улучшению снабжения кислородом. Поэтому при заболеваниях почек иногда наблюдается почечная анемия.
3. Биосинтез белков. В почках активно идут процессы биосинтеза белков, которые необходимы другим тканям. Здесь синтезируются также компоненты системы свертывания крови, системы комплемента и системы фибринолиза.
В почках синтезируются фермент ренин и белок кининоген, участвующие в регуляции сосудистого тонуса и артериального давления.
4. Катаболизм белков. Почки участвуют в катаболизме некоторых белков, имеющих низкую молекулярную массу (5-6 кДа), и пептидов, которые фильтруются в первичную мочу. Среди них гормоны и некоторые другие биологически активные вещества. В клетках канальцев под действием лизосомальных протеолитических ферментов эти белки и пептиды гидролизуются до аминокислот, которые затем поступают в кровь и реутилизируются клетками других тканей.
Большие затраты АТФ почками связаны с процессами активного транспорта при реабсорбции, секреции, а также с биосинтезом белков. Основной путь получения АТФ – это окислительное фосфорилирование. Поэтому ткань почки нуждается в значительных количествах кислорода. Масса почек составляет 0,5% от общей массы тела, а потребление кислорода почками — 10% от всего поступающего кислорода.
Поэтому ткань почки нуждается в значительных количествах кислорода. Масса почек составляет 0,5% от общей массы тела, а потребление кислорода почками — 10% от всего поступающего кислорода.
7.4. РЕГУЛЯЦИЯ ВОДНО-СОЛЕВОГО ОБМЕНА
И МОЧЕОБРАЗОВАНИЯ
Объем мочи и содержание ионов в ней регулируется благодаря сочетанному действию гормонов и особенностям строения почки.
Ренин-ангиотензин-альдостероновая система. В почках, в клетках юкстагломерулярного аппарата (ЮГА), синтезируется ренин – протеолитический фермент, который участвует в регуляции сосудистого тонуса, превращая ангиотензиноген в декапептид ангиотензин I путем частичного протеолиза. Из ангиотензина I под действием фермента карбоксикатепсина образуется (тоже путем частичного протеолиза) октапептид ангиотензин II. Он обладает сосудосуживающим эффектом, а также стимулирует выработку гормона коры надпочечников — альдостерона.
Альдостерон – это стероидный гормон коры надпочечников из группы минералкортикоидов, который обеспечивает усиление реабсорбции натрия из дистальной части почечного канальца благодаря активному транспорту. Он начинает активно секретироваться при значительном снижении концентрации натрия в плазме крови. В случае очень низких концентраций натрия в плазме крови под действием альдостерона может происходить практически полное удаление натрия из мочи. Альдостерон усиливает реабсорбцию натрия и воды в почечных канальцах – это приводит к увеличению объёма крови, циркулирующей в сосудах. В результате повышается артериальное давление (АД) (рис. 19).
Он начинает активно секретироваться при значительном снижении концентрации натрия в плазме крови. В случае очень низких концентраций натрия в плазме крови под действием альдостерона может происходить практически полное удаление натрия из мочи. Альдостерон усиливает реабсорбцию натрия и воды в почечных канальцах – это приводит к увеличению объёма крови, циркулирующей в сосудах. В результате повышается артериальное давление (АД) (рис. 19).
Рис. 19. Ренин-ангиотензин-альдостероновая система
Когда молекула ангиотензина-II выполнит свою функцию, она подвергается тотальному протеолизу под действием группы специальных протез – ангиотензиназ.
Выработка ренина зависит от кровоснабжения почек. Поэтому при снижении АД выработка ренина увеличивается, а при повышении – снижается. При патологии почек иногда наблюдается повышенная выработка ренина и может развиваться стойкая гипертензия (повышение АД).
Гиперсекреция альдостерона приводит к задержке натрия и воды – затем развивается отек и гипертония, вплоть до сердечной недостаточности. Недостаточность альдостерона приводит к значительной потере натрия, хлоридов и воды и уменьшению объема плазмы крови. В почках одновременно нарушаются процессы секреции H+ и NH4+, что может приводить к ацидозу.
Недостаточность альдостерона приводит к значительной потере натрия, хлоридов и воды и уменьшению объема плазмы крови. В почках одновременно нарушаются процессы секреции H+ и NH4+, что может приводить к ацидозу.
Ренин-ангиотензин-альдостероновая система работает в тесном контакте с другой системой регуляции сосудистого тонуса калликреин-кининовой системой, действие которой приводит к понижению АД (рис. 20).
Рис. 20. Калликреин-кининовая система
В почках синтезируется белок кининоген. Попадая в кровь, кининоген под действием сериновых протеиназ — калликреинов превращается в вазоактиные пептиды — кинины: брадикинин и каллидин. Брадикинин и каллидин обладают сосудорасширяющим эффектом – понижают АД.
Инактивация кининов происходит при участии карбоксикатепсина – этот фермент одновременно влияет на обе системы регуляции сосудистого тонуса, что приводит к повышению АД (рис.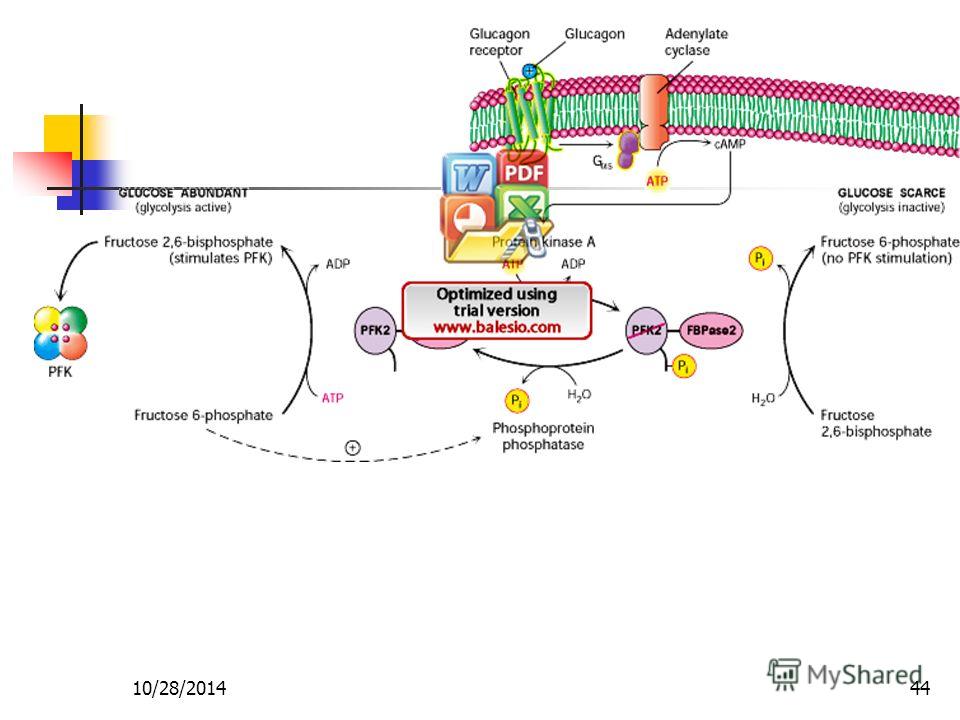 21). Ингибиторы карбоксикатепсина применяются в лечебных целях при лечении некоторых форм артериальной гипертензии. Участие почек в регуляции АД связано также с выработкой простагландинов, которые обладают гипотензивным эффектом.
21). Ингибиторы карбоксикатепсина применяются в лечебных целях при лечении некоторых форм артериальной гипертензии. Участие почек в регуляции АД связано также с выработкой простагландинов, которые обладают гипотензивным эффектом.
Рис. 21. Взаимосвязь ренин-ангиотензин-альдостероновой
и калликреин-кининовой систем
Вазопрессин – пептидный гормон, синтезируемый в гипоталамусе и секретируемый из нейрогипофиза, имеет мембранный механизм действия. Этот механизм в клетках-мишенях реализуется через аденилатциклазную систему. Вазопрессин вызывает сужение периферических сосудов (артериол), в результате повышается АД. В почках вазопрессин повышает скорость реабсорбции воды из начальной части дистальных извитых канальцев и собирательных трубочек. В результате увеличивается относительная концентрация Na, C1, P и общего N. Секреция вазопрессина увеличивается при повышении осмотического давления плазмы крови, например, при повышении потребления соли или обезвоживании организма. Считается, что действие вазопрессина связано с фосфорилированием белков апикальной мембраны почки, в результате чего увеличивается ее проницаемость. При поражении гипофиза, в случае нарушения секреции вазопрессина наблюдается несахарный диабет – резкое увеличение объема мочи (до 4-5 л) с низким удельным весом.
Считается, что действие вазопрессина связано с фосфорилированием белков апикальной мембраны почки, в результате чего увеличивается ее проницаемость. При поражении гипофиза, в случае нарушения секреции вазопрессина наблюдается несахарный диабет – резкое увеличение объема мочи (до 4-5 л) с низким удельным весом.
Натрийуретический фактор (НУФ) – пептид, который образуется в клетках предсердия в гипоталамусе. Это гормоноподобное вещество. Его мишени – клетки дистального отдела почечных канальцев. НУФ действует через гуанилатциклазную систему, т.е. внутриклеточным посредником его является цГМФ. Результатом влияния НУФ на клетки канальцев является снижение реабсорбции Na+, т.е. развивается натрийурия.
Паратгормон – гормон паращитовидной железы белково-пептидной природы. Он имеет мембранный механизм действия через цАМФ. Влияет на удаление солей из организма. В почках паратгормон усиливает канальцевую реабсорбцию Ca2+ и Mg2+, увеличивает экскрецию К+, фосфата, НСО3— и уменьшает экскрецию H+ и NH4+. В основном это происходит благодаря снижению канальцевой реабсорбции фосфата. Одновременно в плазме увеличивается концентрация кальция. Гипосекреция паратгормона приводит к обратным явлениям – увеличению содержания фосфатов в плазме крови и к снижению содержания Ca2+ в плазме.
В основном это происходит благодаря снижению канальцевой реабсорбции фосфата. Одновременно в плазме увеличивается концентрация кальция. Гипосекреция паратгормона приводит к обратным явлениям – увеличению содержания фосфатов в плазме крови и к снижению содержания Ca2+ в плазме.
Эстрадиол – женский половой гормон. Стимулирует синтез
1,25-диоксикальциферола, усиливает реабсорбцию кальция и фосфора в почечных канальцах.
На задержку некоторого количества воды в организме оказывает гормон надпочечников кортизон. В этом случае происходит задержка выделения из организма ионов Na и как следствие – задержка воды. Гормон тироксин приводит к падению массы тела за счет усиленного выделения воды, главным образом через кожу.
Эти механизмы находятся под контролем ЦНС. В регуляции водного обмена участвуют промежуточный мозг и серый бугор мозга. Возбуждение коры головного мозга приводит к изменению работы почек в результате либо прямой передачи соответствующих импульсов по нервным путям, либо путем возбуждения некоторых эндокринных желез, в частности, гипофиза.
Нарушения водного баланса при различных патологических состояниях могут приводить либо к задержке воды в организме, либо к частичному обезвоживанию тканей. Если задержка воды в тканях имеет хронический характер, обычно развиваются различные формы отёков (воспалительные, солевые, голодные).
Патологическое обезвоживание тканей обычно является следствием выделения через почки повышенного количества воды (до 15-20 л мочи в сутки). Такое усиленное мочеотделение, сопровождающееся сильнейшей жаждой, наблюдается при несахарном диабете (diabetes insipidus). У больных, страдающих несахарным диабетом на почве недостатка гормона вазопрессина, почки утрачивают способность концентрировать первичную мочу; моча становится очень разбавленной и имеет низкий удельный вес. Однако ограничение питья при этом заболевании может провести к несовместимому с жизнью обезвоживанию тканей.
Контрольные вопросы
1. Охарактеризуйте экскреторную функцию почек.
2. В чем заключается гомеостатическая функция почек?
3. Какую метаболическую функцию выполняют почки?
4. Какие гормоны участвуют в регуляции осмотического давления и объема внеклеточной жидкости?
5. Опишите механизм действия ренин-ангиотензиновой системы.
6. В чем заключается взаимосвязь ренин-альдостерон-ангиотензиновой и калликреин-кининовой систем?
7. Какие нарушения гормональной регуляции могут быть причиной гипертензии?
8. Укажите причины задержки воды в организме.
9. Что является причиной несахарного диабета?
БИОХИМИЯ НЕРВНОЙ ТКАНИ
Метаболические коррекция — лечение и диагностика нарушений метаболизма в Клинике Expert Clinics
Классическая медицина, как правило, лечит симптомы, в то время как антивозрастная сфокусирована на выявлении и устранении глубинных причин заболеваний.
При интегративном антивозрастном подходе к здоровью организм человека рассматривается как единая система, способная к саморегуляции, особенно если устранить корень нарушений.
Метаболическая коррекция — это тонкая настройка клеточной биохимии, которая способна выровнять состояние здоровья в целом и затормозить развитие многих недугов.
Благодаря персонализированному подходу врачам антивозрастной медицины удается восстановить функции организма на клеточном уровне, во всех тканях и органах.
Перезвоните мне
Основы метаболической коррекции
Научные исследования последних лет доказывают: многие хронические заболевания возникают из-за несбалансированного питания, некачественного сна, токсинов, стресса и определенных дефицитов, мешающих полноценному функционированию организма и истощающих его.
К примеру, нехватка различных нутриентов негативно влияет на метаболические функции организма, которые контролируют все физиологические процессы. В итоге клетки перестают нормально функционировать и повреждаются.
В результате процессы детоксикации становятся неэффективными и замедленными. А это, в свою очередь, приводит к накоплению токсинов в организме, снижению активности иммунной системы, дисбалансу микробиоты, изменению гормонального баланса и снижению функции митохондрий (“энергетических станций” наших клеток).
Ученые также доказали, что с возрастом организм человека производит большое количество токсинов — так называемые эндотоксины. При нарушении метаболических путей они перестают выводится.
Подобные изменения ведут к хроническим дегенеративным состояниям, которые сильно подрывают здоровье человека и снижают качество его жизни. “Плохая” наследственность только усиливает эти разрушительные эффекты.
Метаболическая коррекция заключается в использовании методик и персонализированных препаратов в оптимальных формах и дозах. Они регулируют функцию метаболических ферментов, а также устраняют дефициты полезных веществ.
Цели метаболической коррекции
Любая болезнь — результат нарушения нормальной физиологии, которое происходит, когда организм подвергается воздействию внешних или внутренних неблагоприятных факторов.
Суть метаболической коррекции — в поддержании здоровья и улучшении работы всех систем организма. Так, например, нормализуются белковый, углеводный и жировой обмены.
Коррекция метаболических нарушений может быть весьма эффективной для улучшения состояния людей с сахарным диабетом, тяжелыми формами аллергии и аутоиммунными заболеваниями.
Врачи, восстанавливающие метаболические процессы, добиваются сразу нескольких целей:
-
Предупреждение хронических заболеваний;
-
Коррекция уже имеющихся болезней;
-
Активное поддержание здоровья для компенсации генетической предрасположенности к различным недугам.
-
Повышение качества жизни пациентов.
Работа с пациентами на метаболическом уровне предполагает определенный рацион и применение различных лечебных методик (инфузионные, пероральные, диетотерапия). При грамотном применении они могут стать “первой линией обороны” нашего здоровья.
Записаться на консультацию
Метаболическая коррекция в Expert Clinics
В штате клиники Expert Clinics в Москве — опытные врачи антивозрастной медицины, прошедшие многочисленные обучения как в России, так и за рубежом. Они успешно помогают пациентам выравнивать метаболические процессы организма даже в запущенных случаях.
Они успешно помогают пациентам выравнивать метаболические процессы организма даже в запущенных случаях.
Лечение и диагностика дисфункций метаболизма должна быть комплексной. Для начала необходима полная клиническая оценка, выявляющая проблемы конкретного пациента. Это потребует подробного анамнеза, функциональной диагностики и всестороннего лабораторного тестирования.
Необходимо обследование может выглядеть так:
-
Генетическое тестирование;
-
Комплексный анализ крови;
-
Выявление маркеров воспаления;
-
Иммунные тесты;
-
Содержание тяжелых металлов;
-
Анализ мочи ;
-
УЗИ внутренних органов;
-
МРТ, КТ.
После пациенту будет назначена персонализированная схема лечения.
Как показывает обширная практика врачей Expert Clinics, благодаря метаболической коррекции многие пациенты обретают отличное самочувствие, становятся энергичнее и наблюдают ремиссию своих давних болезней.
Записаться
— функции почек — Биохимия
В организме почки выполняют три функции:
- Экскреция вредных веществ и соединений, находящихся в избытке.
- Поддержание объема и состава внеклеточной жидкости.
- Участие в обмене углеводов, органических кислот, минеральных солей и воды.
Экскреторная функция
Благодаря почкам из организма выводятся вещества, которые можно условно сгруппировать по их происхождению:
- конечные продукты азотистого обмена – мочевина, креатинин, мочевая кислота,
- продукты обезвреживания эндогенных токсичных веществ – гиппуровая кислота, билирубинглюкуронид, индикан,
- избыточные или не нужные вещества – витамины, гормоны, органические кислоты, вода,
- ксенобиотики и продукты их детоксикации – лекарственные препараты, никотин и т.п.
Регулирующая функция
Почки обеспечивают водно-солевой и кислотно-основной гомеостаз. Поддержание гомеостаза внеклеточных жидкостей осуществляется как непосредственно клетками почек, так и при помощи образования определенных активных веществ:
Поддержание гомеостаза внеклеточных жидкостей осуществляется как непосредственно клетками почек, так и при помощи образования определенных активных веществ:
- ацидогенез и аммониегенез обеспечивают удаление ионов Н+ из организма,
- реабсорбция бикарбонатов повышает буферную емкость крови (щелочной резерв),
- секреция ренина через альдостероновый механизм стимулирует реабсорбцию ионов Na+ и удаление ионов К+,
- почки являются объектом воздействия антидиуретического гормона, который усиливает реабсорбцию воды,
- паратгормон, воздействуя на почки, стимулирует реабсорбцию ионов Ca2+,
- синтезируемые здесь же простагландины участвуют в регуляции почечного кровотока, расширяя афферентные и эфферентные артериолы, они также уменьшают чувствительность клеток канальцев к АДГ,
- в почке вырабатывается сильный вазодилататор брадикинин.

Метаболическая функция
Почки осуществляют ряд биохимических синтезов:
- синтез эритропоэтина, стимулирующего продукцию эритроцитов,
- синтезируется активатор плазминогена урокиназа (u-PA).
- гидроксилирование 25-оксикальциферола и превращение его в 1,25-диоксикальциферол,
- в почках происходит синтез глюкозы (глюконеогенез) из реабсорбируемых органических кислот (лактата, пирувата и др). Основное значение глюконеогенез в почках имеет при полном голодании – при этом образуется до 50% всей глюкозы, находящейся в крови.
Функции микрофлоры кишечника | Энтерожермина
Также она выполняет роль защитного барьера, препятствуя проникновению в организм вредных микробов; регулирует функции иммунной системы, принимает участие в переваривании поступающей пищи, синтезирует различные витамины и другие полезные вещества. Эта сложная экосистема оказывает влияние на здоровье всего организма человека. Таким образом, можно выделить основные функции, которые выполняет микрофлора кишечника человека: метаболическую (обмен веществ), трофическую (пищевую), защитную и иммунную.
Эта сложная экосистема оказывает влияние на здоровье всего организма человека. Таким образом, можно выделить основные функции, которые выполняет микрофлора кишечника человека: метаболическую (обмен веществ), трофическую (пищевую), защитную и иммунную.
Метаболическая функция (функция обмена веществ) микрофлоры кишечника
Микрофлора кишечника человека обеспечивает поглощение кальция, магния, железа и других полезных веществ. Синтезирует витамины (фолиевую кислоту, витамин К, витамины группы В и др.). Ферментирует (расщепляет) углеводы (сахар). Последнее приводит к образованию «топлива» для клеток толстой кишки — короткоцепочечных жирных кислот, в частности масляной кислоты.
Постоянная активность микроорганизмов, населяющих микрофлору кишечника, обеспечивает человека различными витаминами и микроэлементами. Витамины группы B поддерживают мышцы в тонусе, обеспечивают лучшую производительность и большую выносливость во время физической нагрузки.
Трофическая (пищевая) функция микрофлоры кишечника
Важным эффектом, который создает микрофлора кишечника человека, является производство короткоцепочечных жирных кислот, таких как масляная кислота, поскольку они обеспечивают питание (трофику), являются источником энергии для клеток кишечника и тем самым способствуют физиологической активности толстой кишки.
Иммунная функция микрофлоры кишечника
Одним из важнейших видов деятельности, которую осуществляет бактериальная микрофлора кишечника человека, является обмен информацией (посредством сигналов) между микроорганизмами и лимфатической системой. Именно лимфатическая система вырабатывает различные вещества и специальные клетки, которые защищают наш организм от инфекций и других вредных воздействий.
Защитная функция микрофлоры кишечника
Защита от патогенных (вредных) микроорганизмов осуществляется множеством способов. Слюна, желудочный сок, сокращения кишечника (перистальтика), мембраны эпителиальных клеток, которые выстилают внутреннюю поверхность кишечника — все это препятствует проникновению инфекций в наш организм. Но первой линией обороны против внешних бактерий является нормальная микрофлора желудочно-кишечного тракта. Флора желудка повышает кислотность среды (снижает уровень pH), что убивает многие вредные микробы. Микрофлора кишечника человека действует путем конкуренции, занимая места на эпителии кишечника и отбирая питательные вещества у патогенных микроорганизмов, а также синтезирует специфические белки (например, бактериоцины), которые непосредственно разрушают вредные бактерии.
Слюна, желудочный сок, сокращения кишечника (перистальтика), мембраны эпителиальных клеток, которые выстилают внутреннюю поверхность кишечника — все это препятствует проникновению инфекций в наш организм. Но первой линией обороны против внешних бактерий является нормальная микрофлора желудочно-кишечного тракта. Флора желудка повышает кислотность среды (снижает уровень pH), что убивает многие вредные микробы. Микрофлора кишечника человека действует путем конкуренции, занимая места на эпителии кишечника и отбирая питательные вещества у патогенных микроорганизмов, а также синтезирует специфические белки (например, бактериоцины), которые непосредственно разрушают вредные бактерии.
Что такое метаболизм?
Метаболизм — это термин, который используется для описания всех химических реакций, участвующих в поддержании жизненного состояния клеток и организма. Обмен веществ можно условно разделить на две категории:
- Катаболизм — распад молекул для получения энергии
- Анаболизм — синтез всех соединений, необходимых клеткам
Метаболизм тесно связан с питанием и доступностью питательных веществ.Биоэнергетика — это термин, описывающий биохимические или метаболические пути, с помощью которых клетка в конечном итоге получает энергию. Образование энергии — один из жизненно важных компонентов обмена веществ.
Изображение предоставлено: VectorMine / Shutterstock.com
Питание, обмен веществ и энергия
Питание — это ключ к обмену веществ. Пути метаболизма зависят от питательных веществ, которые они расщепляют, чтобы произвести энергию. Эта энергия, в свою очередь, требуется организму для синтеза таких молекул, как новые белки и нуклеиновые кислоты (ДНК, РНК).
Питательные вещества, связанные с метаболизмом, включают такие факторы, как потребности организма в различных веществах, индивидуальные функции в организме, необходимое количество и уровень, ниже которого ухудшается состояние здоровья.
Основные питательные вещества поставляют энергию (калории) и поставляют необходимые химические вещества, которые сам организм не может синтезировать. Пища содержит множество веществ, которые необходимы для построения, содержания и восстановления тканей тела, а также для его эффективного функционирования.
Диета нуждается в основных питательных веществах, таких как углерод, водород, кислород, азот, фосфор, сера и около 20 других неорганических элементов. Основные элементы представлены углеводами, липидами и белком. Кроме того, необходимы витамины, минералы и вода.
Углеводы в обмене веществ
Продукты питания содержат углеводы в трех формах: крахмал, сахар и целлюлозу (клетчатку). Крахмал и сахар являются основными и необходимыми источниками энергии для человека. Волокна увеличивают объем рациона.
Ткани организма зависят от глюкозы во всех сферах деятельности. Углеводы и сахара производят глюкозу в результате пищеварения или метаболизма.
Общая реакция горения глюкозы записывается как:
C 6 H 12 O 6 + 6 O 2 ——> 6 CO 2 + 6 H 2 O + энергия
Большинство людей потребляют около половины своего рациона в виде углеводов. Это происходит из таких продуктов, как рис, пшеница, хлеб, картофель и макаронные изделия.
Белки в обмене веществ
Белки являются основными строителями тканей в организме. Они являются частью каждой клетки тела. Белки помогают в структуре клеток, функциях, формировании гемоглобина для переноса кислорода, ферментах для выполнения жизненно важных реакций и множестве других функций в организме. Белки также жизненно важны для снабжения азотом генетического материала ДНК и РНК и производства энергии.
Белки необходимы для питания, поскольку содержат аминокислоты. Из 20 или более аминокислот человеческий организм не может синтезировать 8, и они называются незаменимыми аминокислотами.
К незаменимым аминокислотам относятся:
- Лизин
- Триптофан
- метионин
- Лейцин
- Изолейцин
- Фенилаланин
- Валин
- Треонин
Продукты с высоким содержанием белка — яйца, молоко, соевые бобы, мясо, овощи и зерновые.
Жир в обмене веществ
Жиры — это концентрированные источники энергии. Они производят в два раза больше энергии, чем углеводы или белки, в пересчете на вес.
Функции жиров включают:
- Помогает формировать клеточную структуру;
- Образует защитную подушку и изоляцию вокруг жизненно важных органов;
- Способствует усвоению жирорастворимых витаминов,
- Обеспечение резервного хранилища энергии
Незаменимые жирные кислоты включают ненасыщенные жирные кислоты, такие как линолевая, линоленовая и арахидоновая кислоты. Их нужно принимать с пищей. Насыщенные жиры, наряду с холестерином, участвуют в артериосклерозе и сердечных заболеваниях.
Минералы и витамины в обмене веществ
Минералы, содержащиеся в пищевых продуктах, не вносят непосредственного вклада в энергетические потребности, но важны как регуляторы организма и играют роль в метаболических путях организма. В организме человека содержится более 50 элементов. Было обнаружено, что незаменимыми являются около 25 элементов, а это означает, что их дефицит вызывает определенные симптомы дефицита.
Важные минералы включают:
- Кальций
- фосфор
- Утюг
- Натрий
- Калий
- Хлорид-ионы
- Медь
- Кобальт
- Марганец
- Цинк
- Магний
- Фтор
- Йод
Витамины — это важные органические соединения, которые человеческий организм не может синтезировать сам по себе, и поэтому они должны присутствовать в рационе.Витамины, особенно важные для обмена веществ, включают:
- Витамин А
- B2 (рибофлавин)
- Ниацин или никотиновая кислота
- Пантотеновая кислота
Изображение предоставлено: Siberian Art / Shutterstock.com
Метаболические пути
Химические реакции метаболизма организованы в метаболические пути. Они позволяют преобразовать основные химические вещества из пищи с помощью последовательности ферментов через ряд этапов в другое химическое вещество.
Ферменты имеют решающее значение для метаболизма, потому что они позволяют организмам вызывать желательные реакции, требующие энергии. Эти реакции также связаны с реакциями, высвобождающими энергию. Поскольку ферменты действуют как катализаторы, они позволяют этим реакциям протекать быстро и эффективно. Ферменты также позволяют регулировать метаболические пути в ответ на изменения в клеточной среде или сигналы от других клеток.
Список литературы
Дополнительная литература
Метаболические функции печени
Метаболические функции печени
Гепатоциты — это метаболические сверхспособности в организме.Они играют решающую роль в синтезе молекул, которые используются где-то еще для поддержания гомеостаза, в преобразовании молекул одного типа в другой и в регулировании энергетического баланса. Если вы прослушали курс биохимии, вы, вероятно, потратили большую часть этого курса на изучение метаболических путей в печени. Рискуя быть осужденным слабой похвалой, основные метаболические функции печени можно разделить на несколько основных категорий:
Углеводный метаболизм
Для всех животных критически важно поддерживать концентрацию глюкозы в крови в узком нормальном диапазоне.Поддержание нормального уровня глюкозы в крови как в течение короткого (часы), так и длительного (от дней до недель) периодов времени — одна из особенно важных функций печени.
Гепатоциты содержат множество различных метаболических путей и используют десятки ферментов, которые попеременно включаются или выключаются в зависимости от того, повышается ли уровень глюкозы в крови или выходит за пределы нормального диапазона. Два важных примера этих способностей:
- Избыточная глюкоза, попадающая в кровь после еды, быстро поглощается печенью и секвестрируется в виде большого полимера, гликогена (процесс, называемый гликогенез ).Позже, когда концентрация глюкозы в крови начинает снижаться, печень активирует другие пути, которые приводят к деполимеризации гликогена (гликогенолиз , ) и экспорту глюкозы обратно в кровь для транспортировки во все другие ткани.
- Когда запасы гликогена в печени истощаются, как это происходит, когда животное не ест в течение нескольких часов, сдаются ли гепатоциты? Нет! Они распознают проблему и активируют дополнительные группы ферментов, которые начинают синтезировать глюкозу из таких веществ, как аминокислоты и негексозные углеводы ( глюконеогенез ).Способность печени синтезировать эту «новую» глюкозу имеет огромное значение для плотоядных животных, у которых, по крайней мере в дикой природе, диета практически не содержит крахмала.
Жировой обмен
Некоторые аспекты метаболизма липидов уникальны для печени, но многие из них выполняются преимущественно печенью. Основные примеры роли печени в метаболизме жиров включают:
- Печень чрезвычайно активна в окислении триглицеридов для выработки энергии. Печень расщепляет намного больше жирных кислот, в которых нуждаются гепатоциты, и экспортирует большие количества ацетоацетата в кровь, где он может быть захвачен и легко метаболизирован другими тканями.
- Большая часть липопротеинов синтезируется в печени.
- Печень является основным местом преобразования избыточных углеводов и белков в жирные кислоты и триглицериды, которые затем экспортируются и хранятся в жировой ткани.
- Печень синтезирует большое количество холестерина и фосфолипидов. Некоторые из них содержат липопротеины и становятся доступными для остального тела. Остальная часть выводится с желчью в виде холестерина или после преобразования в желчные кислоты.
Белковый метаболизм
Наиболее важные аспекты метаболизма белков, происходящие в печени:
- Дезаминирование и трансаминирование аминокислот с последующим превращением неазотистой части этих молекул в глюкозу или липиды. Некоторые из ферментов, используемых в этих путях (например, аланин и аспартатаминотрансферазы), обычно анализируются в сыворотке для оценки повреждения печени.
- Удаление аммиака из организма путем синтеза мочевины.Аммиак очень токсичен, и, если его быстро и эффективно не удалить из кровотока, он приведет к заболеванию центральной нервной системы. Частой причиной такой печеночной энцефалопатии у собак и кошек являются пороки кровоснабжения печени, называемые портосистемными шунтами.
- Синтез заменимых аминокислот.
- Гепатоциты отвечают за синтез большинства белков плазмы. Альбумин, основной белок плазмы, синтезируется почти исключительно в печени.Также печень синтезирует многие факторы свертывания, необходимые для свертывания крови.
Отправляйте комментарии [email protected]
Метаболическая функция резкой волновой ряби гиппокампа
Каноски, С. Э. и Гриль, Х. Дж. Вклад гиппокампа в контроль потребления пищи: мнемонические, нейроанатомические и эндокринные механизмы. Biol. Психиатрия 81 , 748–756 (2017).
PubMed
Статья
PubMed Central
Google ученый
Лавенекс, П. и Амарал, Д. Г. Взаимодействие гиппокампа и неокортекса: иерархия ассоциативности. Гиппокамп 10 , 420–430 (2000).
CAS
PubMed
Статья
PubMed Central
Google ученый
Сето К., Сайто Х., Оцука К. и Каваками М. Влияние электростимуляции лимбической структуры на уровень инсулина в плазме кролика. Exp. Clin. Эндокринол . 81, , 347–349 (1983).
CAS
PubMed
Статья
PubMed Central
Google ученый
Endroczi, E., Lissak, K., Bohus, B. & Kovacs, S. Тормозящее влияние архикортикальных структур на гипофизарно-надпочечниковую функцию. Acta Physiol. Акад. Sci. Повесил . 16 , 17–22 (1959).
Google ученый
Рубин Р. Т., Манделл А. Дж. И Крэндалл П.H. Ответы кортикостероидов на лимбическую стимуляцию у человека: локализация раздражителей. Наука 153 , 767–768 (1966).
ADS
CAS
PubMed
Статья
PubMed Central
Google ученый
Макгоу, Дж. Л. и Голд, П. Э. Гормональная модуляция памяти. Психоэндокринология (Academic, 1989).
Голд, П. Э. и ван Бускерк, Р. Влияние посттриальных инъекций гормонов на процессы памяти. Horm. Поведение . 7 , 509–517 (1976).
CAS
PubMed
Статья
PubMed Central
Google ученый
МакИвен, Б.С., Вайс, Дж. М. и Шварц, Л.С. Поглощение кортикостерона мозгом крысы и его концентрация в определенных лимбических структурах. Мозг Рес . 16 , 227–241 (1969).
CAS
PubMed
Статья
PubMed Central
Google ученый
Токарный Р. Гормоны и гиппокамп. Дж. Эндокринол . 169 , 205–231 (2001).
CAS
PubMed
Статья
PubMed Central
Google ученый
Сковилл, В. Б. и Милнер, Б. Потеря недавней памяти после двусторонних поражений гиппокампа. J. Neurol. Нейрохирург. Психиатрия 20 , 11–21 (1957).
CAS
PubMed
PubMed Central
Статья
Google ученый
Бохус, Б. в Гиппокамп: Том 1: Структура и развитие (ред. Исааксон, Р. Л. и Прибрам, К. Х.) 323–353 (Спрингер, 1975).
Якобсон, Л. и Сапольски, Р. Роль гиппокампа в регуляции обратной связи гипоталамо-гипофизарно-надпочечниковой оси. Endocr. Ред. . 12, , 118–134 (1991).
CAS
PubMed
Статья
PubMed Central
Google ученый
Hsu, T. M. et al. Передача сигналов грелина в гиппокампе опосредует аппетит через латеральные пути орексина в гипоталамусе. eLife 4 , e11190 (2015).
PubMed
PubMed Central
Статья
Google ученый
Сафьер, Д. и Фельдман, С. Влияние стимулов перегородки и гиппокампа на нейроны паравентрикулярного ядра. Neuroscience 20 , 749–755 (1987).
CAS
PubMed
Статья
PubMed Central
Google ученый
Пруцкова Н.П., Петров Ю. A. Электрофизиологическое исследование проекций гиппокампа на нейросекреторные клетки супраоптического ядра гипоталамуса крысы. Neurosci. Behav. Физиол . 20 , 194–200 (1990).
CAS
PubMed
Статья
PubMed Central
Google ученый
Тасали Э., Лепроулт Р., Эрманн Д. А. и Ван Каутер Э. Медленноволновой сон и риск диабета 2 типа у людей. Proc. Natl Acad. Sci. США 105 , 1044–1049 (2008).
ADS
CAS
PubMed
PubMed Central
Статья
Google ученый
Капуччио, Ф. П., Д’Элия, Л., Страцзулло, П., Миллер, М. А. Количество и качество сна и заболеваемость диабетом 2 типа: систематический обзор и метаанализ. Уход за диабетом 33 , 414–420 (2010).
PubMed
Статья
PubMed Central
Google ученый
Шпигель К., Лепроулт Р. и Ван Каутер Э. Влияние недосыпания на метаболические и эндокринные функции. Ланцет 354 , 1435–1439 (1999).
CAS
PubMed
Статья
PubMed Central
Google ученый
Амарал Д. и Лавенекс П. в Книга Гиппокампа (ред. Андерсон П. и др.) 45–66 (Oxford Univ. Press, 2006).
Cenquizca, L. A. & Swanson, L.W. Анализ прямых проекций аксонов СА1 поля коры гиппокампа на промежуточный мозг крысы. J. Comp. Neurol . 497 , 101–114 (2006).
PubMed
PubMed Central
Статья
Google ученый
Симадзу Т. Нейрональная регуляция метаболизма глюкозы в печени у млекопитающих. Diabetes Metab. Ред. . 3 , 185–206 (1987).
CAS
PubMed
Статья
PubMed Central
Google ученый
Buzsáki, G. Резкая волновая рябь в гиппокампе: когнитивный биомаркер эпизодической памяти и планирования. Гиппокамп 25 , 1073–1188 (2015).
PubMed
PubMed Central
Статья
Google ученый
Хансен, К. Колебания уровня сахара в крови у нормальных людей натощак. Acta Med. Сканд . 57 , 27–32 (1923).
Google ученый
Risold, P. Y. и Swanson, L. W. Соединения латерального перегородочного комплекса крысы. Brain Res. Brain Res. Ред. . 24 , 115–195 (1997).
CAS
PubMed
Статья
PubMed Central
Google ученый
Sosa, M., Joo, H.R. и Frank, L.M. Резкие волны на спине и вентральной части гиппокампа активируют отдельные сети прилежащего ядра. Нейрон 105 , 725–741 (2020).
CAS
PubMed
Статья
PubMed Central
Google ученый
Tingley, D. & Buzsáki, G. Направление ряби гиппокампа на подкорковые структуры через боковую перегородку. Нейрон 105 , 138–149 (2020).
CAS
PubMed
Статья
PubMed Central
Google ученый
Ниидзима, А. Нейронный контроль уровня глюкозы в крови. Jpn. J. Physiol . 36, , 827–841 (1986).
CAS
PubMed
Статья
PubMed Central
Google ученый
Бриттон, С. В. Исследования условий активности желез внутренней секреции: XVII. Нервный контроль секреции инсулина. Am. J. Physiol . 74 , 291–308 (1925).
CAS
Статья
Google ученый
Коркилл, А. Б., Маркс, Х. П. и Уайт, У. Э. Связь гипофиза с действием инсулина и адреналина. J. Physiol . 612 , 193–205 (1933).
Артикул
Google ученый
Куо, Т., Маккуин, А., Чен, Т.-К. И Ван, Ж.-К. Регулирование гомеостаза глюкозы глюкокортикоидами. Adv. Exp. Med. Биол . 872 , 99–126 (2015).
CAS
PubMed
PubMed Central
Статья
Google ученый
Ким, С. Х. и Парк, М.-Дж. Влияние гормона роста на метаболизм глюкозы и инсулинорезистентность у человека. Ann. Педиатр. Эндокринол. Метаб . 22 , 145–152 (2017).
PubMed
PubMed Central
Статья
Google ученый
Güemes, A. & Georgiou, P. Обзор роли нервной системы в гомеостазе глюкозы и будущих перспектив лечения диабета. Bioelectron.Мед . 4 , 9 (2018).
PubMed
PubMed Central
Статья
Google ученый
Бэнкс В. А. и Кастин А. Дж. Дифференциальная проницаемость гематоэнцефалического барьера для двух пептидов поджелудочной железы: инсулина и амилина. Пептиды 19 , 883–889 (1998).
CAS
PubMed
Статья
PubMed Central
Google ученый
Шулингкамп Р. Дж., Пагано Т. К., Хунг Д. и Раффа Р. Б. Рецепторы инсулина и действие инсулина в головном мозге: обзор и клинические последствия. Neurosci. Biobehav. Ред. . 24 , 855–872 (2000).
CAS
PubMed
Статья
PubMed Central
Google ученый
O’Malley, D., Shanley, LJ & Harvey, J. Инсулин ингибирует нейроны гиппокампа крысы посредством активации АТФ-чувствительных K + и высокой проводимости Ca 2+ -активированных K + каналы. Нейрофармакология 44 , 855–863 (2003).
PubMed
Статья
CAS
PubMed Central
Google ученый
Harasta, A. E. et al. Экспрессия рецептора глюкагоноподобного пептида 1 перегородки определяет подавление кокаин-индуцированного поведения. Нейропсихофармакология 40 , 1969–1978 (2015).
CAS
PubMed
PubMed Central
Статья
Google ученый
Сомерс, В. К., Дайкен, М. Е., Марк, А. Л. и Аббуд, Ф. М. Активность симпатического нерва во время сна у нормальных субъектов. N. Engl. J. Med . 328 , 303–307 (1993).
CAS
PubMed
Статья
PubMed Central
Google ученый
Alford, F. P. et al. Временные паттерны интегрированных уровней гормонов плазмы во время сна и бодрствования. I. Тиреотропный гормон, гормон роста и кортизол. J. Clin. Эндокринол. Метаб . 37 , 841–847 (1973).
CAS
PubMed
Статья
PubMed Central
Google ученый
Logothetis, N. K. et al. Гиппокампально-корковое взаимодействие в периоды подкоркового молчания. Природа 491 , 547–553 (2012).
ADS
CAS
PubMed
Статья
PubMed Central
Google ученый
Swanson, R.A., Levenstein, D., McClain, K., Tingley, D. & Buzsáki, G. Различная специфичность реактивации следов памяти во время резкой волновой ряби в гиппокампе. Curr. Opin. Behav. Sci . 32 , 126–135 (2020).
Артикул
Google ученый
Csicsvari, J., Hirase, H., Czurkó, A., Mamiya, A. & Buzsáki, G. Быстрые сетевые колебания в области CA1 гиппокампа крысы, ведущей себя. Дж.Neurosci . 19 , RC20 (1999).
CAS
PubMed
PubMed Central
Статья
Google ученый
Schomburg, E. W. et al. Тета-фазовая сегрегация входных гамма-паттернов в энторинально-гиппокампальных сетях. Нейрон 84 , 470–485 (2014).
CAS
PubMed
PubMed Central
Статья
Google ученый
Уотсон, Б.О., Левенштейн, Д., Грин, Дж. П., Гелинас, Дж. Н., Бузаки, Г. Сетевой гомеостаз и динамика состояния неокортикального сна. Нейрон 90 , 839–852 (2016).
CAS
PubMed
PubMed Central
Статья
Google ученый
Левенштейн, Д., Уотсон, Б. О., Ринзель, Дж. И Бужаки, Г. Регулирование во сне распределения скоростей возбуждения коры головного мозга. Curr. Opin. Нейробиол . 44 , 34–42 (2017).
CAS
PubMed
PubMed Central
Статья
Google ученый
Chen, C. et al. Последние достижения в области электрохимических биосенсоров глюкозы: обзор. RSC Advances 3 , 4473–4491 (2013).
ADS
CAS
Статья
Google ученый
Basu, A. et al. Время задержки глюкозы от внутрисосудистого до интерстициального компартмента у людей. Диабет 62 , 4083–4087 (2013).
CAS
PubMed
PubMed Central
Статья
Google ученый
Ковачев Б.П., Шилдс Д. и Бретон М. Графическая и числовая оценка временной задержки непрерывного измерения глюкозы. Diabetes Technol. Ther . 11 , 139–143 (2009).
PubMed
PubMed Central
Статья
Google ученый
Бейкер Д. А. и Гоф Д. А. Динамическая задержка и максимальная динамическая ошибка в непрерывных биосенсорах. Анал. Chem . 68, , 1292–1297 (1996).
CAS
PubMed
Статья
PubMed Central
Google ученый
Дэви, Р. Дж., Лоу, К., Джонс, Т. В. и Фурнье, П. А. Вклад внутреннего запаздывания систем непрерывного мониторинга глюкозы в различия в измеренных и фактических концентрациях глюкозы, изменяющихся с переменной скоростью in vitro. J. Diabetes Sci. Технол . 4 , 1393–1399 (2010).
PubMed
PubMed Central
Статья
Google ученый
Кинан, Д. Б., Мастрототаро, Дж. Дж., Восканян, Г. и Стейл, Г. М. Задержки в минимально инвазивных устройствах для непрерывного мониторинга глюкозы: обзор современных технологий. J. Diabetes Sci. Технол . 3 , 1207–1214 (2009).
PubMed
PubMed Central
Статья
Google ученый
Donoghue, T. et al. Параметризация нейронных спектров мощности на периодические и апериодические составляющие. Nat. Neurosci . 23 , 1655–1665 (2020).
PubMed
PubMed Central
Статья
CAS
Google ученый
Тингли Д. и Бужаки Г. Преобразование пространственной карты через гиппокамп-латеральную перегородку. Нейрон 98 , 1229–1242 (2018).
CAS
PubMed
PubMed Central
Статья
Google ученый
Stark, E. et al. Взаимодействия пирамидная клетка-интернейрон лежат в основе колебаний пульсации гиппокампа. Нейрон 83 , 467–480 (2014).
CAS
PubMed
PubMed Central
Статья
Google ученый
Fernández-Ruiz, A. et al. Длительная резкая волновая рябь в гиппокампе улучшает память. Наука 364 , 1082–1086 (2019).
ADS
PubMed
PubMed Central
Статья
CAS
Google ученый
Сон и метаболическая функция
Одним из путей, который может связать короткий сон с избыточным весом, является увеличение потребления калорий у короткоспящих.В следующих разделах будут представлены результаты лабораторных исследований, в которых использовалось полное или частичное лишение сна для изучения влияния недосыпания на субъективное чувство голода, гормоны, участвующие в контроле аппетита, калорийности и набора веса.
Исследования частичного лишения сна
Все большее количество исследований пытались определить влияние частичного лишения сна на регулирование приема пищи. резюмирует результаты этих исследований. Первое исследование, которое одновременно оценивало влияние ограничения сна на субъективное чувство голода и гормоны, участвующие в регуляции аппетита, было опубликовано в 2004 году [162].В этом рандомизированном исследовании с перекрестным дизайном дневные профили лептина и грелина оценивались у молодых мужчин с нормальным весом после двух ночей по 4 часа в постели по сравнению с двумя ночами по 10 часов в постели, в то время как испытуемые заполняли утвержденные шкалы для оценки голода. и аппетит к различным категориям продуктов питания [162]. Лептин снизился на 18%, а уровень грелина увеличился на 28%, а соотношение грелин / лептин увеличилось более чем на 70% при ограничении сна (). Недосыпание также привело к увеличению чувства голода на 23% и увеличению аппетита к питательным веществам с высоким содержанием углеводов на 33%.Семьдесят процентов разницы в увеличении голода объясняется увеличением соотношения грелин / лептин [162] (). Важно отметить, что эти различия в регуляции аппетита имели место, несмотря на одинаковое количество потребляемых калорий, одинаковый сидячий образ жизни и стабильный вес. Для сравнения, 3 дня недокармливания примерно на 900 кал / день у здоровых поджарых добровольцев привели к снижению уровня лептина на 22% [25]; Эти результаты подчеркивают функциональное значение снижения лептина, наблюдаемого при недосыпании.
a Среднее (± SEM) дневное соотношение грелина / лептина и оценки голода, полученные у девяти молодых здоровых людей при непрерывной инфузии глюкозы через 2 дня с 4-часовым отходом ко сну или через 2 дня с 10-часовым отходом ко сну. b Связь между повышением рейтинга голода и увеличением соотношения грелина / лептина при ограничении сна по сравнению с продолжительным
Таблица 2
Влияние частичного недосыпания на регулирование веса
| Показатель результата | Эффект сообщено | Не сообщалось об эффекте | Обратный эффект сообщил |
|---|---|---|---|
| Повышенный голод | |||
| Общее количество исследований | 5 [14, 99, 11782] 907, 144, 162 6 [11, 122, 145, 146, 170, 194] |
||
| Общее количество пациентов | 53 (50 M, 3 F) |
95 (53 M, 42 F) |
|
| Средняя продолжительность TIB (диапазон) | 4.6 ч (от 4 ч до 5,5 ч) |
4,3 ч (от 3 ч до 5,75 ч) |
|
| Средняя продолжительность ограничения сна (диапазон) | 4 ночи (1–14 ночей) |
3 ночей (1–5 ночей) |
|
| Количество исследований, в которых контролировалось потребление пищи до оценки голода | 5 [14, 99, 117, 144, 162] | 4 [11, 122, 146, 170 ] | |
| Повышенное потребление калорий | |||
| Общее количество исследований | 6 [11, 14, 104, 116, 170, 179]. | 1 [146] | |
| Общее количество субъектов | 85 (49 M, 36 F) |
15 (15 M) |
|
| Средняя продолжительность TIB (диапазон) | 4,9 ч (4 ч – 5,75 ч) |
4,25 ч | |
| Средняя продолжительность ограничения сна (диапазон) | 5,3 ночи (1–14) |
2 ночи | |
| Увеличение веса или ожирение | |||
| Общее количество исследований | 2 [11, 117] | 1 [116] | |
| Общее количество пациентов | 24 (17 F, 7 F, 7 F, 7 F, |
11 (6 месяцев, 5 жен) |
|
| Средняя продолжительность TIB (диапазон) | 5.6 ч (5,5 ч – 5,75 ч) |
5,5 ч | |
| Средняя продолжительность ограничения сна (диапазон) | 9 ночей (от 4 до 14 ночей) |
14 ночей | |
| Уменьшено уровни лептина | |||
| Общее количество исследований | 3 [62, 161, 162] | 5 [99, 116, 117, 144, 146] |
4 [11, 122, 154 , 194] |
| Общее количество пациентов | 31 31 M |
55 (47 M, 8 F) |
180 (84 M, 96 F) |
| Средняя продолжительность TIB (диапазон) | 4 ч. (4–4) |
4.95 ч (4,25–5,5) |
4 ч (3–5) |
| Средняя продолжительность ограничения сна (диапазон) | 5 ночей (2–7) |
6,6 ночи (1–14) |
3,5 ночи (1–5) |
| Количество исследований, в которых контролировалось питание до или во время взятия крови | 3 [62, 161, 162] | 3 [99, 117, 144] | 2 [122 , 194] |
| Средний диапазон ИМТ | 19,5–26,5 | 23.2–28,6 | 18,6–40,3 |
| Повышенный уровень грелина | |||
| Общее количество исследований | 3 [117, 144, 162] | 4 [11, 99, 116, 116 ] | |
| Общее количество пациентов | 29 (26 M, 3 F) |
50 (31 M, 19 F) |
|
| Средняя продолжительность TIB | 4,6 ч (4– 5.5) |
5.1 ч (4.25–5,75) |
|
| Средняя продолжительность ограничения сна | 5,6 ночи (1–14) |
5,5 ночи (2–14) |
|
| Количество исследований, в которых контролировалось питание до или во время забор крови | 3 [117, 144, 162] | 1 [99] | |
| Средний диапазон ИМТ | 22,2–27,6 | 22–32,8 |
Увеличение голода Наблюдаемое во время ограничения сна, что приводит к соразмерному увеличению потребления пищи, со временем будет происходить увеличение веса.В соответствии с этой гипотезой из семи исследований, в которых оценивалось потребление пищи во время ограничения сна, в шести наблюдалось повышенное потребление калорий (). Одна ночь на 4 часа в постели по сравнению с одной ночью на 8 часов в постели привела к увеличению потребления калорий на 22% в течение следующего дня у здоровых молодых людей [14]. Когда молодые женщины подвергались диете ad libitum в течение двух условий сна, т. Е. Четыре ночи по ~ 5,5 часов сна против двух ночей по ~ 9 часов сна, наблюдалось 20% -ное увеличение потребления пищи во время сна. ограничение [11].У взрослых среднего возраста с избыточной массой тела повышенное потребление калорий из закусок с более высоким содержанием углеводов наблюдалось после 2 недель ограничения сна (-1,5 ч / ночь) по сравнению с 2 неделями продления сна (+1,5 ч / ночь), в то время как потребление энергии из еды и общее потребление энергии не зависели от условий отхода ко сну [116]. Следует отметить, что каждые 2 недели проводились в условиях неограниченного доступа к вкусной пище. Поскольку потребление пищи в значительной степени зависит от количества и вкусовых качеств пищи, предоставленной субъекту [132], субъекты переедают (> 3400 калорий в день) в каждом условии перед сном, что могло замаскировать влияние продолжительности сна на общее потребление энергии.Недавно в перекрестном исследовании было измерено потребление пищи худыми и полными мужчинами и женщинами среднего возраста после пяти ночей с 4-часовым отходом ко сну и после пяти ночей с 9-часовым отходом ко сну. Добровольцы ели на ~ 300 ккал / день больше, в основном из жиров, особенно из насыщенных жиров, во время короткого сна [170]. Важно отметить, что в последних двух исследованиях ЭЭ, измеренная с помощью золотого стандарта, дважды помеченного методом воды, не различалась между условиями исследования, что позволяет предположить, что увеличение потребления пищи, чаще всего связанное с недосыпанием, не связано со сравнимым ростом потребностей в энергии и в конечном итоге приведет к положительному энергетическому балансу.Подобные предварительные данные были получены в двух других исследованиях, в которых оценивалось потребление калорий во время шведского стола ad libitum [104, 178]. Молодые мужчины и женщины с нормальным весом увеличили потребление калорий на 14%, особенно богатых углеводами питательных веществ, после четырех ночей по 4,5 часа в постели по сравнению с 4 ночами по 8,5 часов в постели [178], и субъекты среднего возраста с ожирением представили до четырех-пяти ночей ограничения или продления привычного графика сна на 2-3 часа в сутки съедали на 15% больше калорий в условиях ограничения времени отхода ко сну [104].Единственное исследование, в котором не сообщалось о каких-либо изменениях в потреблении калорий после недосыпания, предлагало вкусную пищу ad libitum через 1 час после пробуждения после двух ночей по 4 часа и после двух ночей по 8 часов сна. Опять же, эта неограниченная доступность аппетитной пищи привела к перееданию (+ 60%, независимо от условий перед сном) и, возможно, замаскировала влияние потери сна на потребление пищи [146]. Другое методологическое различие может способствовать тому, что недосыпание не влияет на потребление пищи в этом исследовании.Чтобы минимизировать влияние манипуляции продолжительностью сна на циркадную систему, в большинстве исследований центр периода сна оставался постоянным, поэтому ограничение сна связывали с более ранним пробуждением. Напротив, Schmidt et al. разбудил субъектов одновременно в обоих условиях перед сном. В этом последнем случае субъекты могут быть менее склонны есть больше, чем в других исследованиях, в которых ограничение сна было связано с увеличением времени бодрствования до проверки калорийности. Взятые вместе, эти результаты показывают, что потеря сна может стимулировать потребление пищи с избытком> 250 ккал / день.Такое увеличение имеет большое значение, поскольку количество накопленной положительной энергии, необходимое для объяснения эпидемии ожирения, составляет ~ 50 ккал / день [73]. Результаты, касающиеся субъективного чувства голода, кажутся менее последовательными, поскольку только пять исследований [14, 99, 117, 144, 162] из 11 [11, 14, 99, 117, 122, 144–146, 162, 170, 194] изучали влияние сокращения продолжительности сна на голод сообщил о положительных эффектах. Однако, если мы исключим исследования, которые позволяли утолить голод за счет увеличения потребления неконтролируемой доступности пищи, пять из девяти исследований сообщили о положительных эффектах.Следует отметить, что исследование St-Onge et al. сообщили об увеличении примерно на 300 ккал / день после 5 дней 4-часового отхода ко сну, хотя явного влияния продолжительности сна на субъективные чувства голода и аппетита в предыдущий день не было [11, 170], что указывает на ненадежность субъективных оценок.
Влияние потери сна на массу тела изучалось в двух исследованиях, которые позволяли неограниченный доступ к пище во время периодического ограничения сна [11, 116]. Были получены противоречивые результаты. Bosy-Westphal et al.сообщили о прибавке в весе на 0,4 кг в группе свободно живущих худощавых, полных и страдающих ожирением женщин после четырех ночей продолжительностью ~ 5,5 часов по сравнению с двумя ночами продолжительностью ~ 9 часов [11]. Напротив, Недельчева и др. наблюдали одинаковый вес в конце каждого 14-дневного периода продления сна или ограничения сна у мужчин и женщин с избыточным весом, получавших неограниченное количество вкусной пищи в лабораторных условиях. В соответствии с тем фактом, что переедание обычно происходит, когда доступно неограниченное количество съедобной пищи [132], субъекты переедают и набирают ~ 2 кг в каждом состоянии сна [116].Наконец, недавнее исследование изучало, может ли потеря сна поставить под угрозу эффективность низкокалорийной диеты при избыточном ожирении [117]. Мужчины и женщины среднего возраста с избыточной массой тела и ожирением были случайным образом распределялись по 8,5 или 5,5 часов для сна в течение 14 ночей в сочетании с умеренным ограничением калорийности. По истечении 2 недель ограничения сна доля потерянного веса за счет жира снизилась на 55%, а потеря массы тела без жира увеличилась на 60%. Эти результаты показывают, что недостаток сна может снизить эффективность диетических вмешательств для похудания [117].
Двенадцать исследований оценивали влияние частичного лишения сна на лептин и грелин [11, 62, 99, 116, 117, 122, 144, 146, 154, 161, 162, 194]. В трех исследованиях уровень лептина оценивался более чем в 2 временных точках в состоянии покоя и в состоянии ограничения сна у мужчин с нормальным весом после того, как в обоих состояниях потребовалось одинаковое количество пищи. Все обнаружили снижение уровня анорексигенного лептина после недосыпания [62, 161, 162]. Четыре исследования, которые обеспечивали неограниченный доступ к пище до или во время оценки лептина, обнаружили неизменные или повышенные уровни лептина после ограничения сна [11, 116, 146, 154].Поскольку уровни лептина повышаются при приеме пищи и коррелируют со степенью ожирения [25], эти последние результаты могут быть ожидаемым следствием положительного энергетического баланса, достигаемого за счет увеличения потребления пищи [11, 116, 146] и увеличения веса [11, 116]. Точно так же эти исследования не наблюдали изменений в уровнях грелина. В остальных пяти исследованиях наблюдались неизменные или повышенные, а не пониженные уровни лептина после ограничения сна, хотя потребление пищи было одинаковым в обоих состояниях сна. Этому расхождению могут способствовать методологические различия.Например, в исследовании, в котором испытуемые подвергались ограничению калорийности, уровень лептина снижался параллельно с потерей веса и ожирением без значительного независимого эффекта потери сна: отрицательный энергетический баланс, скорее всего, перевесил влияние продолжительности сна [117] . Однако в условиях ограничения сна наблюдались повышенные уровни грелина и повышенные показатели голода, что свидетельствует о том, что, в отличие от того, что наблюдалось с лептином, потеря сна способна усугубить активацию грелина и голода, вызванные отрицательным энергетическим балансом.В исследованиях Leeuwen et al. Однократная оценка лептина проводилась в 07:30 после пяти ночей по 4 часа в постели (время отхода ко сну с 03:00 до 07:00) и после пяти ночей обычного сна (время отхода ко сну с 23:00 до 07:00). [194]. Поскольку на ночное повышение уровня лептина влияет как сон, так и циркадная ритмичность [36], повышение уровня лептина могло произойти позже из-за смещения короткого времени отхода ко сну, что привело к более высокому уровню лептина по утрам. В исследовании Omisade et al.[122], время отхода ко сну ограничивалось 3 часами только в течение 1 дня, а уровни лептина измеряли только в 2 временных точках в дневное время. В исследовании Schmid et al. [144], которые получили однократный анализ крови после одной ночи в течение 4,5 часов в постели, на лептин не повлиял, тогда как голод и грелин были увеличены. Результаты этих двух последних исследований можно сравнить с результатами популяционного исследования, в котором оценивали привычный сон по дневникам и получали утренний забор крови после ночной полисомнографической записи (ПСГ) [174].Авторы обнаружили повышенный уровень грелина, который коррелировал в основном с острой потерей сна, измеренной с помощью ПСГ непосредственно перед забором крови, в то время как пониженный уровень лептина коррелировал с хроническим ограничением сна, о чем свидетельствуют показатели сна. Таким образом, одной ночи лишения сна может быть недостаточно, чтобы вызвать изменения лептина. Эти данные согласуются с ролью лептина в передаче сигналов о долгосрочном нутритивном статусе, в то время как грелин больше участвует в лечении острого голода [174]. Наконец, в пятом исследовании не наблюдалось изменений уровней лептина и грелина после двух ночей по 5 часов в постели по сравнению с.после полностью отдохнувшей ночи (8–10 часов в постели), но ограничение сна привело к снижению чувства насыщения и снижению уровня PYY, анорексигенного пептида, продуцируемого желудочно-кишечным трактом в ответ на прием пищи [99]. Хотя сон не отслеживался, а гормональные уровни оценивались в один момент времени после пробуждения, это первое сообщение о снижении уровней PYY после ограничения сна у людей, что может представлять собой еще один механизм, лежащий в основе снижения чувства сытости, о котором постоянно сообщают недосыпающие. частные лица.
Несогласованные результаты относительно лептина и грелина также могут частично объясняться различиями в размерах выборок, тяжести ограничения сна и исследуемой популяции (). В частности, различия в составе тела и распространенность синдрома обструктивного апноэ во сне (OSA) могут иметь особое значение, поскольку они оба изменяют регуляцию лептина и грелина, а также действие лептина. Ожирение связано с гипогрелинемией, гиперлептинемией и резистентностью к лептину, тогда как OSA связано с гипергрелинемией, гиперлептинемией и резистентностью к лептину.Поэтому неуместно предполагать, что влияние продолжительности сна на лептин и грелин, наблюдаемое у людей с нормальным весом, будет распространяться на людей с ожирением, особенно если они также страдают ОАС. В пользу этой гипотезы показано, что потеря сна приводила к увеличению лептина и отсутствию изменений в грелине в исследованиях, в которых участвовали субъекты с более высоким ИМТ, чем в исследованиях, в которых сообщалось о снижении лептина и повышении грелина (средний верхний диапазон ИМТ 40,3 и 32,8 соответственно по сравнению с исследованиями). 26.5 и 27,6 соответственно). Эти результаты предполагают, что короткий сон может усугубить гиперлептинемию и резистентность к лептину, обычно связанную с избыточным весом у субъектов с ожирением. Несколько наблюдательных, эпидемиологических исследований, в которых изучалась перекрестная связь между продолжительностью сна, лептином и грелином, также дали противоречивые результаты. В соответствии с большинством экспериментальных исследований, проведенных на людях с нормальным весом, когортное исследование сна в Висконсине и семейное исследование в Квебеке показали снижение уровня лептина после контроля ИМТ или ожирения у людей, которые обычно не спят [22, 174].Также было обнаружено, что более высокий уровень грелина связан с коротким сном [174]. В отличие от этих двух исследований, в которых участвовали мужчины и женщины с различным телосложением (худощавые, полные и полные), три последующих исследования не подтвердили связь между продолжительностью сна, уровнем лептина и грелина: в одном участвовали только женщины с избыточным весом в постменопаузе [94]. , еще одна — только женщины, в основном страдающие избыточным весом или ожирением, с диабетом 2 типа [209], а последняя — только страдающие ожирением [83]. Наконец, в недавнем отчете Кливлендского семейного исследования была обнаружена связь между продолжительностью сна и лептином, которая была в противоположном ожидаемом направлении: каждый час сокращения общего времени сна, оцениваемого PSG в течение 7 часов 55 минут принудительного пребывания в постели, было связано с увеличением на 6%. в утреннем лептине после корректировки с учетом различных факторов, включая ожирение и тяжесть апноэ во сне.Поскольку эта когорта была создана для изучения генетической основы апноэ во сне, распространенность ОАС и ожирения была очень высокой. Хотя эти обсервационные исследования скорректированы на ИМТ или ожирение, а иногда и на индексы тяжести ОАС в их анализах, они не оценивали связь между сном и лептином для различного состава тела и статуса апноэ во сне (например, худощавость, избыточный вес или ожирение, с или без OSA). Необходимы исследования, чтобы определить, вызваны ли несоответствия результатов лептина и грелина между исследованиями, как экспериментальными, так и наблюдательными, методологическими различиями или они представляют собой физиологические различия между худыми, полными и тучными людьми с или без ОАС.
Продолжительность сна, по-видимому, дозозависимо модулирует нейроэндокринную регуляцию аппетита. В исследовании, в котором оценивались 24-часовые профили лептина после 6 дней 4-, 8- и 12-часового укладывания спать у здоровых худых молодых мужчин в условиях стабильного потребления калорий и уровней активности, все характеристики 24-часового лептинового профиля (общее среднее, ночной максимум и амплитуда) постепенно увеличивались от 4-х до 12-ти часового отхода ко сну [161]. Доза-реакция между количеством сна, оценкой голода и уровнями грелина также сообщалось в исследовании с участием TSD, 4.5 и 7 ночей [144]. Эти результаты согласуются с метаанализом эпидемиологических исследований, в которых сообщается о тенденции зависимости доза-ответ между сокращением продолжительности сна и повышенным риском ожирения [23].
Границы | Метаболическая функция и дисфункция жировой ткани: последствия ожоговой травмы
Введение
Жировая ткань, в основном состоящая из адипоцитов, преадипоцитов, фибробластов, лейкоцитов и эндотелиальных клеток, широко признана метаболическим органом, играющим центральную роль в регулировании энергетического гомеостаза всего тела (Ahima and Flier, 2000; Coelho et al., 2013). Жировая ткань накапливает энергию в виде липидов и регулирует их системную мобилизацию и распределение в крови (Sethi and Vidal-Puig, 2007). Поступающая с пищей избыточная энергия сохраняется в адипоцитах в форме липидов или триацилглицеридов (ТАГ) посредством пути липогенеза (Tan and Vidal-Puig, 2008). Эволюционно жировая ткань функционирует, чтобы накапливать поступающий жир в существующих адипоцитах, увеличивая размер адипоцитов (гипертрофия) или увеличивая количество новых адипоцитов за счет адипогенеза (гиперплазия) (Ghaben and Scherer, 2019).Однако длительное снижение расхода энергии приводит к увеличению количества адипоцитов и, следовательно, к ожирению и связанному с ним метаболическому синдрому (Lafontan and Langin, 2009). Следовательно, когда приток липидов превышает емкость жировой ткани, липиды начинают накапливаться в эктопических тканях, что приводит к метаболической дисфункции и инсулинорезистентности из-за липотоксичности (Virtue and Vidal-Puig, 2010). В нескольких недавних обзорах подробно описана роль жировой ткани в предотвращении липотоксичности и метаболической дисфункции у здоровых людей по сравнению с тучными (Boura-Halfon and Zick, 2009; Virtue и Vidal-Puig, 2010; Nielsen et al., 2014; Фассауэр и Блюхер, 2015; Чжу и Шерер, 2018; Габен и Шерер, 2019). Кроме того, теперь ясно, что жировая ткань не только служит резервуаром энергии, но также действует как эндокринный орган и выделяет метаболиты и другие сигнальные компоненты, такие как адипонектин, лептин и определенные факторы роста, которые играют важную роль во множестве сигнальных путей ( Fasshauer and Bluher, 2015). Метаболиты, секретируемые жировой тканью, обычно называют «адипокинами». Другие примеры адипокинов включают висфатин, резистин, хемерин, апелин, интерлейкин 6 (IL6), интерлейкин 1β (IL1β), фактор некроза опухоли α (TNFα) и фактор роста фибробластов 21 (FGF21) (Fasshauer and Bluher, 2015).Некоторые ключевые примеры, иллюстрирующие роль адипокинов в координации метаболической функции, включают (но не ограничиваются ими): выход глюкозы в печени (адипонектин, FGF21), чувствительность к инсулину (адипонектин, лептин), воспаление (IL6, IL1β, TNFα) и метаболизм липидов. (фактор кластерной дифференциации 36) (Fasshauer, Bluher, 2015; Zhu, Scherer, 2018).
Жировая ткань является одним из крупнейших эндокринных органов человека и в основном бывает двух типов: белого и коричневого. Оба эти типа жировой ткани имеют различную морфологию, характеристики и функциональные аспекты.Коричневая жировая ткань (BAT) специализируется на производстве тепла (или термогенезе) и обычно присутствует у новорожденных и взрослых (Ahima and Flier, 2000; Coelho et al., 2013; Peirce et al., 2014). Первоначально считалось, что BAT не имеет большого физиологического значения для взрослых людей. Однако недавние исследования с использованием позитронно-эмиссионной томографии — компьютерной томографии (ПЭТ-КТ) показали, что повышенная активность BAT положительно коррелирует с расходом энергии в состоянии покоя. Кроме того, исследования ПЭТ-КТ с использованием аналога глюкозы 2-фтор-2-дезокси-D- [ 18 F] глюкозы ( 18 FDG) выявили наличие очаговых областей повышенного накопления индикаторов в надключичной области, периадреналовая область, подмышечная впадина, межреберье и вокруг магистральных сосудов у взрослых людей (Carter et al., 2011а, б). BAT, по сравнению с белой жировой тканью (WAT), имеет повышенное содержание митохондрий, повышенную экспрессию разобщающего белка 1 (UCP1) и более васкуляризован, что позволяет выделять тепло за счет окисления жирных кислот, что приводит к образованию мультилокулярных адипоцитов меньшего размера (Peirce et al. др., 2014). Хотя BAT также хранит липиды, их основная функция — выработка тепла, а не снабжение питательными веществами.
Белая жировая ткань, с другой стороны, имеет более крупные однокамерные адипоциты, пониженное содержание митохондрий и отсутствие экспрессии UCP1.ЖАТ рассредоточен по всему телу и в основном бывает двух типов: подкожный (паховый у грызунов) и висцеральный (эпидидимальный у грызунов) жировая ткань. WAT действует как метаболический орган и выполняет различные метаболические функции, поддерживая энергетический гомеостаз у людей (Bartelt and Heeren, 2014; Peirce et al., 2014), что делает его прибыльной терапевтической мишенью против метаболических нарушений, таких как диабет 2 типа и др. ожирение. В настоящее время физиологические изменения жировой ткани представляют собой серьезную клиническую проблему при лечении гиперметаболизма, хронического метаболического нарушения, встречающегося у пациентов с тяжелыми ожогами.Действительно, катаболизм сухих мышц и жировой ткани, связанный с гиперметаболической реакцией, является разрушительным и является основным фактором заболеваемости и смертности пациентов (Jeschke et al., 2011). Ожоговое повреждение и связанный с ним гиперметаболический ответ вызывают у пациентов последствия метаболического стресса, характеризующиеся повышенными уровнями глюкозы, инсулина, свободных жирных кислот, аминокислот и гормонов стресса в сыворотке (Jeschke et al., 2011), что вызывает жизненно важные органы, такие как печень, поджелудочная железа, мышцы, жировая ткань, сердце и мозг, должны функционировать с ускорением.После активации гиперметаболизм приводит к порочному кругу петель прямой связи у ожоговых пациентов, что в конечном итоге приводит к увеличению расхода энергии в состоянии покоя (REE), повышенному риску сепсиса, инфекций и, в конечном итоге, смерти (Jeschke et al., 2011; Auger et al. ., 2017). Распространенность гиперметаболизма среди пациентов с тяжелыми ожогами вызвала большой интерес к пониманию патофизиологических механизмов, лежащих в основе этой реакции, чтобы минимизировать неблагоприятные метаболические последствия гиперметаболизма, такие как инсулинорезистентность, дислипидемия, гепатостеатоз и сердечно-сосудистые осложнения (Рисунок 1).Хотя роль жировой ткани в эффективном хранении калорий и защитном экранировании кажется очевидной, мы и другие недавно начали понимать глубокое влияние метаболической дисфункции адипоцитов на усиление гиперметаболической реакции после ожоговой травмы. Действительно, вызванный ожогом переход от накопления калорий к катаболическому состоянию нарушает энергетический гомеостаз и приводит в действие системную липотоксичность и связанное с этим накопление липидов в жизненно важных органах, таких как сердце, печень и почки, что способствует дисфункции органов.Остается интригующий вопрос: какие факторы вызывают дисфункцию жировой ткани после ожоговой травмы и как они регулируются? Этот обзор направлен на критический анализ нашего понимания и научных достижений, достигнутых к настоящему времени в отношении механистической роли жировой ткани и полезных аспектов терапевтического воздействия на липолиз жировой ткани и потемнение, чтобы смягчить гиперметаболический ответ у ожоговых пациентов.
Рисунок 1. Дисфункция жировой ткани после ожоговой травмы.WAT подвергается липолизу и потемнению, опосредованным несколькими сигнальными медиаторами и адипокинами, которые способствуют увеличению системного потока FFA и расходу энергии в состоянии покоя.
Липолиз жировой ткани
Жировая ткань активирует липолитические пути при активации внешним раздражителем от мозга и надпочечников в режиме голодания, во время физических упражнений и при увеличении потребности в энергии (Nielsen et al., 2014). После активации липолиза запасы ТАГ в адипоцитах расщепляются на глицерин и свободные жирные кислоты (СЖК), которые могут использоваться, когда в системном кровотоке истощаются запасы глюкозы (Lafontan and Langin, 2009).Высвободившийся глицерин и СЖК попадают в кровоток и впоследствии помогают питать жизненно важные органы и поддерживать системный энергетический гомеостаз. Эти высвобожденные липиды могут метаболизироваться важнейшими органами в качестве субстрата для β-окисления и производства аденозинтрифосфата (АТФ) (Kolditz and Langin, 2010). Эта уникальная способность жировой ткани балансировать хранение и использование липидов позволяет людям иметь повышенную буферную способность FFA для удовлетворения потребности в энергии (Frayn, 2002). Однако метаболические последствия чрезмерного увеличения запасов жировой ткани приводят к ожирению, нежелательной секреции адипокина и связанной с этим метаболической дисфункции (Alberti et al., 2009).
В адипоцитах разрушение липидных капель достигается за счет последовательного действия трех липаз, которые расщепляют ТАГ на три СЖК. При липолизе млекопитающих TAG превращается в диацилглицерид (DAG) с помощью триглицерид липазы (ATGL), что является ферментативной стадией с ограниченной скоростью (Zimmermann et al., 2004). Далее DAG превращается в моноацилглицерид (MAG) за счет последовательного действия гормоночувствительной липазы (HSL), которая обладает субстратной специфичностью по отношению к DAG (Haemmerle et al., 2002).MAG дополнительно расщепляется под действием моноглицерид липазы (MGL), которая превращает его в глицерин и FFA (Fredrikson et al., 1986). Активация и дезактивация липолитических путей в жировой ткани строго регулируется гормонами и изобилием питательных веществ в кровообращении. Положительным регулятором и активаторами липолиза жировой ткани являются катехоламины, натрийуретические пептиды и гормон стресса, действующие на β-адренорецепторы. Напротив, антилиполитическое действие опосредуется инсулином (Jensen and Nielsen, 2007), катехоламинами и гормональным действием на α-адренорецепторы (Laugwitz et al., 1996; Чо и др., 2005; Эндо и Кобаяши, 2012; Родригес и др., 2013). Исследования у ожоговых пациентов показали повышенный уровень системных липидов и заметное повышение уровней катехоламинов в плазме и гормонов стресса, что свидетельствует о взаимосвязанной роли гормонов стресса в запуске липолиза WAT после ожоговой травмы (Wolfe et al., 1987a, b; Jeschke et al. ., 2008, 2011). Более того, исследование, проведенное на педиатрических ожоговых пациентах с использованием пропранолола (β-адреноблокатор) или плацебо, показало снижение экспрессии генов жировой ткани, связанных с метаболизмом липидов, у пациентов, получавших пропранолол (Barrow et al., 2006).
В физиологических условиях липолитический путь активируется связыванием катехоламинов с β-адренергическими (преимущественно β1 и β2 и в некоторой степени β3 у человека) рецепторами (Barbe et al., 1996; Jocken and Blaak, 2008). Напротив, антилиполитическое действие опосредуется связыванием с α2-адренорецепторами (Берлан и Лафонтан, 1982; Лафонтан и Берлан, 1993). Обе группы α- и β-адренорецепторов принадлежат к семейству рецепторов, сопряженных с G-белками, где α2 связывается с белками, содержащими субъединицу ингибитора G и , а β-рецептор связывается с белками, содержащими стимулирующую субъединицу G s , для взаимодействия с аденилилом. циклаза (Barrow et al., 2006). После активации G i ингибирует аденилатциклазу, тогда как G s активирует аденилатциклазу. В свою очередь, аденилатциклаза превращает АТФ в цитозольный аденозинмонофосфат (цАМФ), что приводит к быстрому накоплению внутриклеточных уровней цАМФ, что активирует передачу сигналов протеинкиназы А (ПКА) (также известную как цАМФ-зависимая передача сигналов). Весь этот процесс способствует активации ATGL и фосфорилированию HSL, что в конечном итоге запускает каскад липолиза жировой ткани.Более подробно об активации и передаче сигналов PKA-сигналов подробно рассказано в другом месте (Nielsen et al., 2014).
Повышенные уровни FFA и системных липидов обычно связаны с тяжелой ожоговой травмой и гиперметаболическим ответом (Jeschke et al., 2011). Фактически, усиленный липолиз может сохраняться у пациентов с тяжелой ожоговой травмой в течение периода времени от нескольких дней до лет после первоначальной травмы, что приводит к истощению жировой ткани и системной липотоксичности (Jeschke et al., 2011).Липолиз жировой ткани после ожоговой травмы стимулируется сигнализацией адренергического стресса, которая включает катехоламины, глюкокортикоиды и цитокины, опосредованные гиперметаболическим ответом. Катехоламины, такие как адреналин и норэпинефрин, а также гормоны стресса являются основными медиаторами метаболической реакции в жировой ткани через адренергические рецепторы (Robidoux et al., 2004). У ожоговых пациентов уровни адреналина, норэпинефрина и кортизола повышаются, что вызывает различные катаболические события, такие как липолиз, мышечное истощение и усиление РЗЭ (Jeschke et al., 2011, 2015). Например, одно исследование у детей с ожогами продемонстрировало повышенный уровень катехоламинов в течение 2 лет по сравнению со здоровыми людьми (Kulp et al., 2010). В другом исследовании уровень норадреналина был в 10 раз выше в образцах мочи и оставался повышенным более года (Jeschke et al., 2011).
Устойчивость к инсулину в жировой ткани
Адипоциты чувствительны к действию инсулина и останавливают липолиз для поддержания системного энергетического гомеостаза (Ghaben and Scherer, 2019).Инсулин связывается с α-субъединицей инсулинового рецептора (IRS) на внешней поверхности клетки, что приводит к конформационным изменениям β-субъединицы для связывания АТФ. Связывание АТФ запускает аутофосфорилирование β-субъединицы IRS внутри клетки, что активирует активность тирозинкиназы и обеспечивает дальнейшую клеточную активность инсулина (Wolever, 1990; Burks and White, 2001; Kido et al., 2001). Существует четыре известных типа IRS (1–4) (Burks and White, 2001). IRS-3 присутствует только в жировой ткани. Фосфорилирование белка IRS инсулином активирует множество сигнальных путей.Например, активация пути фосфоинозитол-3-киназа (PI3K) / протеинкиназа B (AKT) способствует липогенезу de novo , ингибирует липолиз WAT и подавляет глюконеогенез в печени (Burks and White, 2001). Внутриклеточный транспорт глюкозы опосредуется действием инсулина через транспортер глюкозы 4 (GLUT4), способствуя захвату глюкозы, липогенезу и подавлению липолиза, что снижает приток FFA в кровоток (Smith, 2002). Следовательно, в состоянии дефицита инсулина жировая ткань высвобождает FFA для поддержания энергетического гомеостаза за счет окисления жирных кислот и для прямого использования такими органами, как печень, сердце и мышцы.Кроме того, печень превращает FFA в кетоновые тела, которые являются альтернативным субстратом для мозга во время голодания (Wilcox, 2005).
Тяжелая ожоговая травма часто связана с системной инсулинорезистентностью, гипергликемией и гиперлипидемией, что значительно влияет на заболеваемость и смертность ожоговых пациентов (Jeschke et al., 2011, 2020). Таким образом, инсулинотерапия и подобные аналоги считаются важной терапевтической мерой против гиперметаболического ответа и показали многообещающие результаты у пациентов с тяжелыми ожогами (Jeschke et al., 2010, 2016). В нормальных условиях повышение уровня глюкозы в крови после приема пищи стимулирует высвобождение инсулина из β-клеток поджелудочной железы. Таким образом, высвобожденный инсулин способствует захвату глюкозы в скелетных мышцах и жировой ткани и ингибирует гликолиз, гликогенолиз и липолиз для поддержания эугликемии (Belfort et al., 2005; Gormsen et al., 2007; Kolka et al., 2015). В критических обстоятельствах, таких как тяжелые ожоговые травмы, высвобождение медиаторов стресса вызывает метаболические нарушения и значительно влияет на энергетический гомеостаз.Чтобы удовлетворить повышенную потребность в энергии в условиях стресса, медиаторы стресса противодействуют анаболическому действию инсулина (ингибирование выработки глюкозы), вызывая состояние инсулинорезистентности. Катехоламины подавляют действие инсулина во время стресса или физических упражнений через α2-адренорецепторы (Wilcox, 2005), предполагая роль адренергических сигналов стресса в развитии состояния инсулинорезистентности. Эти гиперметаболические условия активируют альтернативные источники энергии, такие как липолиз жировой ткани, высвобождение глюкозы в печени и протеолиз скелетных мышц (Jeschke et al., 2020). Таким образом, гипергликемия и инсулин не подавляют липолиз и производство глюкозы. Усиление системной циркуляции энергии приводит к липотоксичности, инсулинорезистентности и связанному с этим эктопическому отложению жира в жизненно важных органах, что влияет на их функцию. Следовательно, липолиз жировой ткани и инсулинорезистентность играют интригующую роль в развитии гиперметаболического состояния, вызванного ожогом.
Термогенез коричневой жировой ткани
Активация коричневой жировой ткани подразумевает запуск BAT для выработки тепла за счет «не дрожащего термогенеза».«Изначально считалось, что НИМ в первую очередь связаны с выделением тепла у новорожденных и животных, находящихся в спячке, с небольшим вкладом в метаболизм (Abdullahi and Jeschke, 2017). Недавние исследования с использованием ПЭТ-КТ показали метаболическую активацию BAT в ответ на различные стимуляторы стресса, такие как воздействие холода, ожоговые травмы и кожные раны (Carter et al., 2011a, b). Интересно, что у мышей, подвергшихся ожоговому повреждению, было пониженное содержание липидов и повышенное содержание UCP1 в BAT (Carter et al., 2011a, b).Кроме того, Yo et al. (2013) также показали, что активация BAT положительно коррелирует с повышенным расходом энергии, увеличением митохондриального биогенеза и усилением экспрессии Ucp1 . После нацеливания на митохондриальную активность с использованием SS31 Yo et al. (2013) показали, что пептид SS31, нацеленный на митохондрии, приводит к снижению расхода энергии у ожоговых мышей и подавляет экспрессию Ucp1 в изолированных BAT от ожоговых мышей. Совсем недавно Bhattarai et al. (2020) показали зависящее от времени пополнение BAT после ожоговой травмы, когда UCP1-зависимое дыхание усиливается в первые 24 часа после ожога и затем уменьшается.В целом, исследования, оценивающие роль BAT при ожоговом поражении, показали, что BAT активируется впоследствии, что свидетельствует об усилении экспрессии Ucp1 . Однако понимание роли BAT при ожоговой травме очень ограничено. Необходимы дальнейшие исследования, чтобы определить, способствует ли это депо системной дисфункции за счет усиленного липолиза или медиаторов воспаления.
Белый Браунинг жировой ткани
Концепция потемнения WAT подразумевает запуск WAT для перехода в коричневую жировую ткань, известную как бежевая жировая ткань.Последний характерен для промежуточного состояния между WAT и BAT, имея смесь одногокулярных и мультилокулярных адипоцитов, повышенное содержание митохондрий и проявляя экспрессию UCP1 (Bartelt and Heeren, 2014; Peirce et al., 2014). Исследования показывают, что WAT и BAT происходят из разных клонов мезенхимальных стволовых клеток и имеют разные миогенные сигнатуры (Timmons et al., 2007; Seale et al., 2008). Однако бежевые и белые адипоциты имеют одну и ту же клеточную линию, а белые адипоциты могут трансдифференцироваться в бежевые адипоциты при стимуляции (Park et al., 2014; Икеда и др., 2018). Теоретически бежевые адипоциты могут происходить из клеток-предшественников, находящихся в WAT, в ответ на внешние стимулы (Wang et al., 2013). Альтернативно, они также могут трансдифференцироваться от WAT или BAT, что включает прямое превращение WAT или BAT в бежевые адипоциты и наоборот (Barbatelli et al., 2010; Rosenwald et al., 2013). И WAT, и BAT сильно иннервируются и чувствительны к потребности в энергии, передаваемой симпатической нервной системой, которая регулирует транскрипционный контур процесса потемнения.
В ответ на воздействие холода или катехоламинов потемнение WAT инициируется рецептором γ, активируемым пролифератором пероксисом ( Ppar γ), который дополнительно активирует экспрессию термогенного гена ( Pgc1 α, Prdm16 и Ucp1 ). Кроме того, потемнение WAT может быть активировано несколькими гормонами через перекрестные помехи между тканями. Например, FGF21 и костный морфогенетический белок 4 (BMP4), которые продуцируются в ответ на передачу адренергических сигналов, могут способствовать потемнению WAT (Fisher et al., 2012; Грефхорст и др., 2015). Другие выделяемые WAT гормоны, такие как лептин и инсулин, также способствуют потемнению WAT через нейроны проопиомеланокортина (POMC) и симпатическую нервную систему (Dodd et al., 2015). Кроме того, хотя катехоламины необходимы для активации потемнения WAT, макрофаги M2, находящиеся в WAT, также являются источником катехоламинов (Nguyen et al., 2011; Abdullahi et al., 2019a), что предполагает наличие прямой петли, инициирующей потемнение WAT. и связанный с этим вклад в усиление липолиза REE и WAT.В целом потемнение WAT может быть вызвано ощущением энергии, метаболической потребностью, гормональной активацией и адренергической сигнализацией, все из которых предполагают решающую метаболическую роль WAT в энергетическом гомеостазе.
В эволюционном плане WAT действует как отличный теплоизолятор, который поддерживает температуру тела и энергетический баланс человека. Кроме того, активация потемнения WAT потенциально может помочь людям с ожирением или избыточным весом, стремящимся снизить массу тела и стать метаболически здоровыми (Bartelt and Heeren, 2014).Однако стрессовые условия (например, тяжелая ожоговая травма) также вызывают потемнение WAT, что приводит к увеличению REE, увеличению содержания митохондрий, экспрессии UCP1 и обильному поступлению питательных веществ, запуская липолиз WAT для окисления жирных кислот. Потемнение WAT, по-видимому, является обычным признаком у пациентов с тяжелыми ожоговыми травмами и является основным фактором, подпитывающим гиперметаболический ответ у пациентов с тяжелыми травмами (Abdullahi and Jeschke, 2017). Следовательно, потемнение WAT может рассматриваться как дисфункциональная черта жировой ткани и новая проблема, с которой врачи должны бороться при лечении пациентов с тяжелыми ожоговыми травмами.Хотя значительный успех был достигнут в выявлении критических предшественников, вызывающих потемнение WAT, продолжаются исследования по определению мер по ингибированию липолиза WAT и потемнения у пациентов с тяжелыми ожоговыми травмами (Abdullahi and Jeschke, 2017). Фактически, исследование, проведенное у детей и взрослых ожоговых пациентов, продемонстрировало, что потемнение WAT является основным фактором гиперметаболической реакции и связанной с ней метаболической дисфункции (Patsouris et al., 2015; Sidossis et al., 2015). Кроме того, исследования на грызунах показали критическую роль цитокинов, таких как IL6 и макрофаги, в активации потемнения WAT и возникновении стеатоза печени после ожогового повреждения (Abdullahi et al., 2017, 2019а).
Воспаление жировой ткани — порочный круг
Воспаление в клетках или тканях человека — это многоуровневый ответный механизм для коммуникации и защиты от любых вредных веществ, а также для подготовки защитного щита для борьбы с ними. В большинстве случаев воспалительные триггеры исчезают, как только проблема решена. Однако в некоторых случаях, когда триггер является постоянным, продолжающееся острое воспаление может превратиться в хроническое воспаление (Barrett et al., 2019). В стрессовых условиях адипоциты могут активно продуцировать и рекрутировать другие воспалительные клетки и медиаторы, которые способны активировать и рекрутировать иммунные клетки, такие как макрофаги и Т-лимфоциты (Т-клетки) (Abdullahi et al., 2019a). WAT действует как резервуар, из которого может исходить множество метаболических сигналов посредством секреции большого количества цитокинов и гормонов. Например, TNFα может напрямую нарушать передачу сигналов инсулина (Hotamisligil, 1999), а IL6 может активировать потемнение WAT (Abdullahi et al., 2017), что привело к развитию инсулинорезистентности и увеличению РЗЭ соответственно. Пациенты с тяжелыми ожоговыми травмами часто испытывают хроническое воспаление и связанную с ним метаболическую дисфункцию (Jeschke et al., 2011). Такое хроническое гипервоспаление часто влияет на заживление ран, вызывает потемнение ВАТ, липолиз, липотоксичность, сепсис и связанные с ним осложнения полиорганной недостаточности.
Исследования, проведенные за последние два десятилетия, показали, что воспаление в WAT является основным фактором гиперметаболической реакции, наблюдаемой у ожоговых пациентов.WAT действует как эндокринный орган и играет активную роль в секреции воспалительных компонентов, таких как цитокины, гормоны и другие факторы роста. При критическом стрессе адипоциты привлекают медиаторы воспаления, хемоаттрактанты (такие как хемоаттрактантный белок-1 моноцитов или MCP-1) и цитокины, которые активируют поляризацию макрофагов, а также множественные метаболические сигнальные пути. Установлено, что ожоговая травма приводит к структурным, функциональным и морфологическим изменениям WAT с повышенным уровнем циркулирующих адипокинов, полученных из WAT, медиаторов воспаления и гормонов, которые, как известно, регулируют воспаление и метаболизм WAT (Jeschke et al., 2011). Исследования на людях и грызунах показали, что нейтрализация TNFα ускоряет заживление ран (Ashcroft et al., 2012). Оценка WAT, полученная от ожоговых пациентов, выявила усиление инфильтрации лейкоцитов, макрофагов и активации белка Nod-подобного рецептора инфламмасомы-3 (NLRP3), который играет решающую роль во множественных сигнальных путях (Stanojcic et al., 2014). Кроме того, исследования, выясняющие роль NLRP3 в WAT после ожогового повреждения, показывают, что NLRP3 обладает эффектом против потемнения и что генетическая делеция этой инфламмасомы увеличивает потемнение WAT и гиперметаболический ответ (Vinaik et al., 2019). С другой стороны, рекрутирование макрофагов в WAT претерпевает попеременную поляризацию и активацию, что приводит к секреции катехоламинов и цитокинов, которые индуцируют множественные сигнальные каскады (Abdullahi et al., 2019a). Напр., Альтернативно активированные макрофаги секретируют IL6, который играет решающую роль в активации потемнения WAT и связанной с этим дисфункции после ожогового повреждения (Ashcroft et al., 2012). Более того, ингибирование альтернативно активированных макрофагов ухудшает метаболическую адаптацию и термогенный ответ жировой ткани.Кроме того, введение интерлейкина-4 восстанавливает экспрессию термогенных генов, системную мобилизацию жира и энергетический гомеостаз в ответ на холод (Nguyen et al., 2011), что свидетельствует о важной роли альтернативно активируемых макрофагов в организации термогенного ответа и метаболической адаптации жировой ткани. в ответ на стрессовые раздражители, такие как холод или ожоги.
Терапевтические достижения, направленные на липолиз WAT и браунинг
Терапевтические вмешательства, направленные на β-адренорецепторы, цитокины, липолиз WAT и медиаторы потемнения после ожоговой травмы, показали многообещающие результаты в улучшении REE, стеатоза печени, уменьшении гипергликемии, гиперлипидемии и хронического воспаления.Пропранолол, неселективный блокатор передачи β-адренергических сигналов, показал многообещающие преимущества в снижении REE и снижении экспрессии маркеров потемнения ( Ucp1 , Cox-iv ) в WAT, что свидетельствует о важности регулирования катехоламинов и гормонов стресса. для смягчения WAT-ассоциированной дисфункции (Sidossis et al., 2015). Пропранолол имеет более высокое сродство к рецепторам β1 и β2 (Barbe et al., 1996; Finnerty and Herndon, 2013). Однако исследования, проведенные независимой исследовательской группой, показали, что хронический адренергический стресс после ожоговой травмы усиливает экспрессию рецептора β3 в WAT, хотя его роль в потемнении WAT остается неуловимой (Saraf et al., 2015). Кроме того, исследования у детей с ожогами, получавших пропранолол в независимой группе, показали снижение сердечной нагрузки, сопровождающееся снижением липолиза, мышечного катаболизма, гепатостеатоза и, в конечном итоге, РЗЭ (Finnerty and Herndon, 2013). Однако неселективный характер пропранолола также подвергает пациентов значительному риску сердечной недостаточности (неопубликованные клинические данные). Кроме того, было обнаружено, что системные уровни IL6 повышаются в образцах пациентов с ожогами вскоре после ожоговой травмы и сохраняются более месяца (Patsouris et al., 2015). Последующие исследования по оценке роли IL6 в WAT с использованием модели мышей с глобальным нокаутом IL6 показали, что делеция IL6 предотвращает потемнение WAT после ожоговой травмы (Abdullahi et al., 2017). Кроме того, делеция IL6 снижает инфильтрацию макрофагов и подавляет альтернативную активацию и поляризацию макрофагов (Abdullahi et al., 2019a). Дальнейшие исследования, оценивающие роль блокады рецептора IL6 с использованием тоцилизумаба, показали положительные результаты в подавлении передачи сигналов IL6 и ослаблении потемнения WAT и стеатоза печени на мышиной модели C57BL / 6 после ожоговой травмы (Abdullahi et al., 2019b), что свидетельствует о терапевтическом потенциале тоцилизумаба в снижении гиперметаболического ответа, связанного с WAT, у ожоговых пациентов с повышенным уровнем IL6.
Метформин, успешный клинический препарат для лечения диабета, показал многообещающие результаты в улучшении инсулинорезистентности, не вызывая гипогликемии, в рандомизированных клинических испытаниях фазы II, проведенных у ожоговых пациентов (Jeschke et al., 2016). Механистические исследования на мышиной модели термического повреждения, оценивающие действие метформина, продемонстрировали, что метформин индуцирует активность протеинфосфатазы 2A (PP2A), дефосфорилируя, таким образом, ключевые ферменты липолитического пути WAT [такие как ацетил-CoA-карбоксилаза (ACC) и HSL] и способствует накоплению жира в адипоцитах (Auger et al., 2019). Фактически, лечение метформином также снижает митохондриальное дыхание и усиливает контроль митохондриального сцепления в WAT, что предполагает косвенный защитный эффект метформина в снижении потемнения WAT и REE (Auger et al., 2019). Хотя эти изменения не зависят от активации аденозинмонофосфаткиназы (AMPK), канонического механизма действия метформина, авторы постулируют, что для активации этого сигнального пути в высокоэнергетической бежевой жировой ткани необходимы более высокие концентрации этого бигуанида (Auger et al., 2019). Действительно, после поглощения печенью на животных моделях было продемонстрировано, что только 10-40 мкМ метформина является системным биодоступным, что может быть недостаточно для запуска прокатаболических эффектов AMPK, а также увеличения митохондриального биогенеза через PGC-1α (He и Вондисфорд, 2015). Кроме того, другое клиническое исследование метформина показало многообещающие результаты в отношении катаболизма скелетных мышц и резистентности к инсулину после тяжелой ожоговой травмы (Gore et al., 2005). Кроме того, исследования, оценивающие влияние метформина на воспаление в адипоцитах, показали, что введение метформина подавляет провоспалительные цитокины, такие как TNFα и IL-1β, а также косвенно усиливает противовоспалительный эффект метформина (Qi et al., 2017). Более того, у ожоговых пациентов есть доказательства того, что этот бигуанид может снижать содержание медиаторов воспаления в сыворотке, таких как IL-1β и MCP-1 (Jeschke et al., 2016). На сегодняшний день потенциальные преимущества других бигуанидов или агонистов PPARγ, таких как тиазолидиндионы, в отношении контроля уровня глюкозы и системной дисфункции после ожогов изучены недостаточно.
Аципимокс, производное ниацина, которое нацелено на липолиз WAT, показал эффективные результаты на мышиной модели при серьезной ожоговой травме.Аципимокс не только снижал системные уровни липидов, но и ослаблял потемнение WAT и инфильтрацию печеночного жира после ожоговой травмы (Barayan et al., 2019), предполагая, что терапевтический потенциал аципимокса в борьбе с гиперметаболическим ответом после ожоговой травмы заключается в смягчении WAT. дисфункция и сопутствующие осложнения. Кроме того, аципимокс показал многообещающие результаты в 3-месячных клинических испытаниях на ВИЧ-инфицированных пациентах (23) с гиперлипидемией и аномальным распределением жира. Лечение аципимоксом привело к снижению системных уровней липидов, снижению липолиза WAT и повышению чувствительности к инсулину у ВИЧ-инфицированных пациентов (Hadigan et al., 2006). Однако влияние аципимокса на воспаление WAT и системный метаболизм глюкозы при этих побочных эффектах еще предстоит выяснить.
Резюме и заключение
Жировая ткань обладает огромной буферной способностью для высвобождения, хранения и рассеивания энергии в случае необходимости. Исследования последних лет показали, что функция и дисфункция жировой ткани играют важную роль в ожоговой травме и связанной с ней гиперметаболической реакции, которая часто прогрессирует до мультиорганной дисфункции (рис. 2).Жировая ткань, являясь эндокринным органом, выделяет множество адипокинов и поддерживает энергетический гомеостаз у человека. Хотя был достигнут значительный успех в понимании факторов, запускающих дисфункцию адипоцитов, наши знания по-прежнему ограничены верхушкой айсберга. Большая часть исследований в этой области сосредоточена на выявлении ключевых маркеров, которые изменяются при ожоговой травме. Хотя роль инсулина была тщательно изучена, существует множество цитокинов, адипокинов (лептин, адипонектин) и гормонов стресса, которые до сих пор полностью не изучены, и то, как они влияют на действие инсулина и морфологию WAT, все еще остается предметом дискуссий.
Рисунок 2. Дисфункция жировой ткани и связанное с ней полиорганное повреждение после ожоговой травмы. Повышенные уровни системного потока FFA, медиаторов воспаления и адипокинов в совокупности способствуют возникновению петли прямой связи, гиперметаболизма и полиорганных повреждений (Jeschke et al., 2020).
Инфильтрация и поляризация макрофагов в жировой ткани после ожоговой травмы известна, однако точная роль медиаторов воспаления все еще не ясна. Исследования, выясняющие роль IL6 и его ингибирование, выявили пагубную роль этого цитокина в процессах потемнения WAT и стеатоза печени (Abdullahi et al., 2017, 2019b). Однако его роль в рекрутировании и поляризации макрофагов неясна (Abdullahi et al., 2019a). Точно так же исследования, проведенные для понимания роли TNFα у ожоговых пациентов, показали, что он активируется после ожогового повреждения (Yeh et al., 1997), однако его точная роль все еще не ясна. Исследования, проведенные на мышиной модели NLRP3, показали, что удаление NLRP3 усиливает потемнение, липолиз, стеатоз печени и ухудшает заживление ран (Stanojcic et al., 2014; Vinaik et al., 2018, 2019), что позволяет предположить, что медиаторы воспаления оказывают благотворное влияние в острой фазе после ожога. Однако необходимы дальнейшие исследования, чтобы понять защитную роль и механистическое действие NLRP3 при ожоговой травме.
Исследования последнего десятилетия и достижения в области клинической помощи при ожогах значительно расширили наши знания и значительно повысили выживаемость ожоговых пациентов (рис. 3). Такие препараты, как метформин, показали многообещающую безопасность и эффективность в клинических испытаниях фазы II.Кроме того, метформин защищает от липолиза WAT, потемнения и помогает поддерживать эугликемию. Молекулярные исследования механизма действия метформина продемонстрировали, что метаболические пути активированы для смягчения дисфункции WAT в исследованиях на грызунах. Однако необходимы дальнейшие подробные клинические исследования, чтобы выяснить его влияние на функцию жировой ткани после тяжелой ожоговой травмы. Недавно ингибитор липолиза WAT аципимокс показал многообещающие результаты в исследованиях на грызунах при ожоговом поражении, что свидетельствует о возможных преимуществах борьбы с дисфункцией WAT в будущем.Однако необходимы дальнейшие механистические исследования, чтобы выяснить действие аципимокса и его возможное влияние на чувствительность к инсулину и воспаление WAT. Сильная корреляция этих препаратов, направленных на дисфункцию WAT, предполагает, что снижение липолиза WAT и потемнения является важной терапевтической стратегией для лечения гиперметаболической реакции у ожоговых пациентов. Другими потенциально подходящими стратегиями могут быть понимание роли рекрутирования макрофагов в WAT и механизмов, участвующих в активации и поляризации макрофагов.С этой целью еще многое предстоит выяснить в отношении взаимодействия макрофагов с самими собой и взаимодействия макрофагов с адипоцитами при ожоговом поражении. Наконец, понимание роли адипокинов и их влияния на сигнальные пути в жизненно важных органах-мишенях, таких как мозг, центральная нервная система, сердце, печень и скелетные мышцы, возможно, может раскрыть новые терапевтические стратегии для снижения гиперметаболического ответа, связанного с WAT, при ожоге. пациенты.
Рисунок 3. Резюме терапевтических достижений, направленных на липолиз адипоцитов и потемнение после ожоговых повреждений. Метаболическое влияние медикаментозного лечения (A) Пропранолол (B) Тоцилизумаб (C) Метформин (D) Аципимокс послеожоговая травма.
Авторские взносы
Все перечисленные авторы внесли существенный, прямой и интеллектуальный вклад в работу и одобрили ее к публикации.
Финансирование
Особого финансирования на эту работу авторы не получали.Гранты, поддерживающие исследовательскую работу, предоставлены Канадским институтом исследований в области здравоохранения (123336), Национальным институтом здравоохранения (№ R01GM087285 и R01GM133961) и Институтом регенеративной медицины Онтарио.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Список литературы
Абдуллахи, А., Аугер, К., Станойчич, М., Пацурис, Д., Парусис, А., Epelman, S., et al. (2019a). Альтернативно активированные макрофаги вызывают потемнение белой жировой ткани при ожогах. Ann. Surg. 269, 554–563. DOI: 10.1097 / SLA.0000000000002465
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Абдуллахи А., Чен П., Станойчич М., Садри А. Р., Кобурн Н. и Йешке М. Г. (2017). Сигнал IL-6 из костного мозга необходим для потемнения белой жировой ткани после ожога. Амортизатор 47, 33–39. DOI: 10.1097 / SHK.0000000000000749
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Абдуллахи А. и Йешке М. Г. (2017). Укрощение пламени: борьба с потемнением белой жировой ткани в условиях гиперметаболизма. Endocr. Ред. 38, 538–549. DOI: 10.1210 / er.2017-00163
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Абдуллахи А., Самади О., Аугер К., Канагалингам Т., Бёнинг Д., Би, С. и др. (2019b). Побурение белой жировой ткани после ожоговой травмы способствует стеатозу и дисфункции печени. Cell Death Dis. 10: 870. DOI: 10.1038 / s41419-019-2103-2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ахима, Р. С., и Флиер, Дж. С. (2000). Жировая ткань как эндокринный орган. Trends Endocrinol. Метаб. 11, 327–332. DOI: 10.1016 / s1043-2760 (00) 00301-5
CrossRef Полный текст | Google Scholar
Альберти, К. Г., Экель, Р. Х., Гранди, С. М., Зиммет, П. З., Климан, Дж. И., Донато, К. А. и др. (2009). Гармонизация метаболического синдрома: совместное промежуточное заявление целевой группы международной федерации диабета по эпидемиологии и профилактике; национальный институт сердца, легких и крови; Американская Ассоциация Сердца; всемирная федерация сердца; международное общество атеросклероза; и международная ассоциация по изучению ожирения. Тираж 120, 1640–1645. DOI: 10.1161 / CIRCULATIONAHA.109.192644
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эшкрофт, Г. С., Джеонг, М. Дж., Эшворт, Дж. Дж., Хардман, М., Джин, В., Мутсопулос, Н. и др. (2012). Фактор некроза опухоли-альфа (TNF-альфа) является терапевтической мишенью для нарушенного заживления кожных ран. Регенерация для восстановления ран. 20, 38–49. DOI: 10.1111 / j.1524-475X.2011.00748.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Оже, К., Кнут, К. М., Абдуллахи, А., Самади, О., Парусис, А., и Йешке, М. Г. (2019). Метформин предотвращает патологическое потемнение подкожной белой жировой ткани. Mol. Метаб. 29, 12–23.
Google Scholar
Аугер К., Самади О. и Йешке М. Г. (2017). Биохимические изменения, лежащие в основе послеожогового гиперметаболизма. Biochim. Биофиз. Acta Mol. Основы дис. 1863 (10 Pt B), 2633–2644. DOI: 10.1016 / j.bbadis.2017.02.019
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бараян, Д., Винаик, Р., Аугер, К., Кнут, К. М., Абдуллахи, А., и Йешке, М. Г. (2019). Ингибирование липолиза с помощью аципимокса снижает потемнение белой жировой ткани после ожога и инфильтрацию жира в печени. Ударная волна 53, 137–145. DOI: 10.1097 / SHK.0000000000001439
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Барбателли, Г., Мурано, И., Мадсен, Л., Хао, К., Хименес, М., Кристиансен, К. и др. (2010). Появление индуцированных холодом коричневых адипоцитов в депо белого жира мышей определяется преимущественно трансдифференцировкой адипоцитов от белого к коричневому. Am. J. Physiol. Эндокринол. Метаб. 298, E1244 – E1253. DOI: 10.1152 / ajpendo.00600.2009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Барбе П., Милле Л., Галицки Дж., Лафонтан М. и Берлан М. (1996). Оценка in situ роли бета-1-, бета-2- и бета-3-адренорецепторов в контроле липолиза и питательного кровотока в подкожной жировой ткани человека. Br. J. Pharmacol. 117, 907–913. DOI: 10.1111 / j.1476-5381.1996.tb15279.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Барретт Л. В., Страх В. С., Уэйтман Дж. К., Вуд Ф. М. и Страх М. В. (2019). Понимание острой ожоговой травмы как хронического заболевания. Ожоговая травма 7:23. DOI: 10.1186 / s41038-019-0163-2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Барроу, Р. Э., Вулф, Р. Р., Дасу, М. Р., Барроу, Л. Н., и Херндон, Д. Н. (2006). Использование бета-адренергической блокады для предотвращения гепатомегалии, вызванной травмой. Ann. Surg. 243, 115–120. DOI: 10.1097 / 01.sla.0000193834.07413.91
CrossRef Полный текст | Google Scholar
Белфорт Р., Мандарино Л., Кашьяп С., Вирфель К., Пратипанаватр Т., Берриа Р. и др. (2005). Дозозависимый эффект повышенного содержания свободных жирных кислот в плазме на передачу сигналов инсулина. Диабет 54, 1640–1648. DOI: 10.2337 / диабет.54.6.1640
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Берлан М. и Лафонтан М.(1982). Альфа-2-адренорецептор жировых клеток человека: сравнительное исследование связывания альфа-2-адренергического радиолиганда и биологического ответа. J. Physiol. (Париж) 78, 279–287.
Google Scholar
Бхаттарай Н., Ронтоянни В. Г., Росс, Э., Огунбиледже, Дж. О., Муртон, А. Дж., И Портер, К. (2020). Рекрутмент коричневой жировой ткани в модели тяжелых ожогов на грызунах. Burns (в печати). DOI: 10.1016 / j.burns.2020.04.034
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бура-Халфон, С.и Зик Ю. (2009). Фосфорилирование белков IRS, действие инсулина и инсулинорезистентность. Am. J. Physiol. Эндокринол. Метаб. 296, E581 – E591. DOI: 10.1152 / ajpendo..2008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Картер, Э. А., Бонаб, А. А., Хамрахи, В., Питман, Дж., Винтер, Д., Макинтош, Л. Дж. И др. (2011a). Влияние ожоговой травмы, холодового стресса и повреждения кожной раны на морфологию и энергетический метаболизм коричневой жировой ткани мыши (BAT) in vivo. Life Sci. 89, 78–85. DOI: 10.1016 / j.lfs.2011.04.014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Картер, Э. А., Бонаб, А. А., Пол, К., Йеркса, Дж., Томпкинс, Р. Г., и Фишман, А. Дж. (2011b). Связь производства тепла с накоплением 18F-FDG в коричневой жировой ткани мыши после стресса. J. Nucl. Med. 52, 1616–1620. DOI: 10.2967 / jnumed.111.0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чо, К.J., Shim, J.H., Cho, M.C., Choe, Y.K., Hong, J.T., Moon, D.C., et al. (2005). Сигнальные пути, участвующие в индуцированном альфа-меланоцитами липолизе в адипоцитах 3T3-L1, стимулирующем гормон. J. Cell Biochem. 96, 869–878. DOI: 10.1002 / jcb.20561
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Додд, Г. Т., Декерф, С., Ло, К., Саймондс, С. Е., Виде, Ф., Балланд, Э. и др. (2015). Лептин и инсулин действуют на нейроны POMC, способствуя потемнению белого жира. Ячейка 160, 88–104. DOI: 10.1016 / j.cell.2014.12.022
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эндо Т., Кобаяши Т. (2012). Экспрессия функционального рецептора ТТГ в белых жировых тканях мышей hyt / hyt индуцирует липолиз in vivo. Am. J. Physiol. Эндокринол. Метаб. 302, E1569 – E1575. DOI: 10.1152 / ajpendo.00572.2011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фишер, Ф. М., Кляйнер, С., Douris, N., Fox, E.C., Mepani, R.J., Verdeguer, F., et al. (2012). FGF21 регулирует PGC-1альфа и потемнение белой жировой ткани в адаптивном термогенезе. Genes Dev. 26, 271–281. DOI: 10.1101 / gad.177857.111
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фредриксон, Г., Торнквист, Х., и Белфраге, П. (1986). И гормоночувствительная липаза, и моноацилглицеринлипаза необходимы для полной деградации триацилглицерина адипоцитов. Biochim.Биофиз. Acta 876, 288–293. DOI: 10.1016 / 0005-2760 (86)
-9
CrossRef Полный текст | Google Scholar
Гор, Д. К., Вольф, С. Е., Сэнфорд, А., Херндон, Д. Н., и Вулф, Р. Р. (2005). Влияние метформина на непереносимость глюкозы и катаболизм мышц после тяжелой ожоговой травмы. Ann. Surg. 241, 334–342. DOI: 10.1097 / 01.sla.0000152013.23032.d1
CrossRef Полный текст | Google Scholar
Гормсен, Л. К., Йессен, Н., Джедстед, Дж., Джедде, С., Norrelund, H., Lund, S., et al. (2007). Дозозависимые эффекты свободных жирных кислот на метаболизм глюкозы и липидов во время соматостатиновой блокады гормона роста и инсулина у людей. J. Clin. Эндокринол. Метаб. 92, 1834–1842. DOI: 10.1210 / jc.2006-2659
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Грефхорст, А., ван ден Бекель, Дж. К., ван Хаутен, Э. Л., Стинберген, Дж., Виссер, Дж. А. и Теммен, А. П. (2015). Эстрогены увеличивают экспрессию костного морфогенетического белка 8b в коричневой жировой ткани мышей. Biol. Половые различия. 6: 7. DOI: 10.1186 / s13293-015-0025-y
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хадиган, К., Либау, Дж., Торриани, М., Андерсен, Р., и Гринспун, С. (2006). Повышение чувствительности к триглицеридам и инсулину при применении аципимокса в течение 3 месяцев у инфицированных вирусом иммунодефицита человека пациентов с гипертриглицеридемией. J. Clin. Эндокринол. Метаб. 91, 4438–4444. DOI: 10.1210 / jc.2006-1174
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хеммерле, Г., Zimmermann, R., Hayn, M., Theussl, C., Waeg, G., Wagner, E., et al. (2002). Дефицит гормоночувствительной липазы у мышей вызывает накопление диглицеридов в жировой ткани, мышцах и семенниках. J. Biol. Chem. 277, 4806–4815. DOI: 10.1074 / jbc.M110355200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Икеда, К., Маретич, П., и Каджимура, С. (2018). Общие и отличительные черты коричневых и бежевых адипоцитов. Trends Endocrinol. Метаб. 29, 191–200.DOI: 10.1016 / j.tem.2018.01.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Йешке, М. Г., Абдуллахи, А., Бернетт, М., Реху, С., и Станойчич, М. (2016). Контроль уровня глюкозы у пациентов с тяжелыми ожогами, использующих метформин: промежуточный анализ безопасности и эффективности рандомизированного контролируемого исследования фазы II. Ann. Surg. 264, 518–527. DOI: 10.1097 / SLA.0000000000001845
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Йешке, М.Г., Чинкес, Д. Л., Финнерти, К. К., Кулп, Г., Суман, О. Е., Норбери, В. Б. и др. (2008). Патофизиологический ответ на тяжелую ожоговую травму. Ann. Surg. 248, 387–401. DOI: 10.1097 / SLA.0b013e3181856241
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Йешке, М. Г., Гауглиц, Г. Г., Кулп, Г. А., Финнерти, К. К., Уильямс, Ф. Н., Крафт, Р. и др. (2011). Длительная стойкость патофизиологического ответа на тяжелую ожоговую травму. PLoS One 6: e21245.DOI: 10.1371 / journal.pone.0021245
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Йешке, М. Г., Кулп, Г. А., Крафт, Р., Финнерти, К. К., Млчак, Р., Ли, Дж. О. и др. (2010). Интенсивная инсулинотерапия у педиатрических пациентов с тяжелыми ожогами: проспективное рандомизированное исследование. Am. J. Respir Crit. Care Med. 182, 351–359. DOI: 10.1164 / rccm.201002-0190OC
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Йешке, М.Г., Пацурис, Д., Станойчич, М., Абдуллахи, А., Реху, С., Пинто, Р., и др. (2015). Патофизиологический ответ на ожоги у пожилых людей. EBioMedicine 2, 1536–1548. DOI: 10.1016 / j.ebiom.2015.07.040
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Йешке, М. Г., ван Баар, М. Э., Чоудри, М. А., Чанг, К. К., Джебран, Н. С., и Логсетти, С. (2020). Ожоговая травма. Nat. Преподобный Дис. Праймеры 6:11. DOI: 10.1038 / s41572-020-0145-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джокен, Дж.W., и Blaak, E. E. (2008). Катехоламин-индуцированный липолиз в жировой ткани и скелетных мышцах при ожирении. Physiol. Behav. 94, 219–230. DOI: 10.1016 / j.physbeh.2008.01.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кидо Ю., Накаэ Дж. И Аччили Д. (2001). Клинический обзор 125: рецептор инсулина и его клеточные мишени. J. Clin. Эндокринол. Метаб. 86, 972–979. DOI: 10.1210 / jcem.86.3.7306
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Колка, с.М., Ричи, Дж. М., Кастро, А. В., Бруссар, Дж. Л., Ионут, В., и Бергман, Р. Н. (2015). Инсулинорезистентность, индуцированная липидами, не ухудшает доступ инсулина к скелетным мышцам. Am. J. Physiol. Эндокринол. Метаб. 308, E1001 – E1009. DOI: 10.1152 / ajpendo.00015.2015
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кулп, Г. А., Херндон, Д. Н., Ли, Дж. О., Суман, О. Е., и Йешке, М. Г. (2010). Степень и величина выброса катехоламинов у детей с ожогами. Амортизатор 33, 369–374. DOI: 10.1097 / SHK.0b013e3181b92340
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лафонтан М. и Берлан М. (1993). Адренергические рецепторы жировых клеток и контроль функции белых и коричневых жировых клеток. J. Lipid Res. 34, 1057–1091.
Google Scholar
Лаугвиц, К. Л., Аллгейер, А., Офферманнс, С., Спичер, К., Ван Санде, Дж., Дюмон, Дж. Э. и др. (1996). Рецептор тиреотропина человека: гепталический рецептор, способный стимулировать членов всех четырех семейств G-белков. Proc. Natl. Акад. Sci. США 93, 116–120. DOI: 10.1073 / pnas.93.1.116
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Nguyen, K. D., Qiu, Y., Cui, X., Goh, Y. P., Mwangi, J., David, T., et al. (2011). Альтернативно активированные макрофаги продуцируют катехоламины для поддержания адаптивного термогенеза. Природа 480, 104–108. DOI: 10.1038 / nature10653
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Нильсен, Т.С., Джессен, Н., Йоргенсен, Дж. О., Моллер, Н., и Лунд, С. (2014). Рассечение липолиза жировой ткани: молекулярная регуляция и последствия для метаболических заболеваний. J. Mol. Эндокринол. 52, R199 – R222. DOI: 10.1530 / JME-13-0277
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Парк, А., Ким, В. К., и Бэ, К. Х. (2014). Различие белых, бежевых и коричневых адипоцитов, полученных из мезенхимальных стволовых клеток. World J. Stem Cells 6, 33–42. DOI: 10.4252 / wjsc.v6.i1.33
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Patsouris, D., Qi, P., Abdullahi, A., Stanojcic, M., Chen, P., Parousis, A., et al. (2015). Ожог вызывает потемнение подкожной белой жировой ткани у мышей и людей. Cell Rep. 13, 1538–1544. DOI: 10.1016 / j.celrep.2015.10.028
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Qi, T., Chen, Y., Li, H., Pei, Y., Woo, S.L., Guo, X., et al. (2017).Роль PFKFB3 / iPFK2 в подавлении метформином воспалительных реакций адипоцитов. J. Mol. Эндокринол. 59, 49–59. DOI: 10.1530 / JME-17-0066
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Робиду, Дж., Мартин, Т. Л., и Коллинз, С. (2004). Бета-адренорецепторы и регуляция расхода энергии: дело семейное. Annu. Rev. Pharmacol. Toxicol. 44, 297–323. DOI: 10.1146 / annurev.pharmtox.44.101802.121659
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Родригес, А.Р., Алмейда, Х., Гувейя, А. М. (2013). Передача сигналов альфа-МСГ через рецептор меланокортина 5 способствует липолизу и ухудшает повторную этерификацию в адипоцитах. Biochim. Биофиз. Acta 1831, 1267–1275. DOI: 10.1016 / j.bbalip.2013.04.008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сараф М., Херндон Д., Портер К., Хондроникола М., Чао Т., Толивер Кински Т. и др. (2015). Хронический адренергический стресс вызывает активацию адренергических рецепторов β-3 в белой жировой ткани ожоговых пациентов. FASEB J. 29 (1 приложение): 995.916. DOI: 10.1096 / fasebj.29.1_supplement.995.16
CrossRef Полный текст | Google Scholar
Сил П., Бьорк Б., Янг В., Каджимура С., Чин С., Куанг С. и др. (2008). PRDM16 управляет переключением между коричневым жиром и скелетными мышцами. Nature 454, 961–967. DOI: 10.1038 / nature07182
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сетхи, Дж. К., и Видаль-Пуч, А. Дж. (2007). Серия тематических обзоров: биология адипоцитов.Адаптация к питанию зависит от функции и пластичности жировой ткани. J. Lipid Res. 48, 1253–1262. DOI: 10.1194 / мл. R700005-JLR200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сидосис, Л. С., Портер, К., Сараф, М. К., Борсхайм, Э., Радхакришнан, Р. С., Чао, Т. и др. (2015). Побурение подкожной белой жировой ткани у человека после тяжелого адренергического стресса. Cell Metab. 22, 219–227. DOI: 10.1016 / j.cmet.2015.06.022
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Смит, У.(2002). Нарушение («диабетический») передачи сигналов и действия инсулина происходит в жировых клетках задолго до непереносимости глюкозы — возникает ли инсулинорезистентность в жировой ткани? Внутр. J. Obes. Relat. Метаб. Disord. 26, 897–904. DOI: 10.1038 / sj.ijo.0802028
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Станойчич, М., Чен, П., Харрисон, Р.А., Ван, В., Антонишин, Дж., Зунига-Пфлюкер, Дж. К. и др. (2014). Инфильтрация лейкоцитов и активация инфламмасомы NLRP3 в белой жировой ткани после термического повреждения. Crit. Care Med. 42, 1357–1364. DOI: 10.1097 / CCM.0000000000000209
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тан, К. Ю., и Видаль-Пуч, А. (2008). Расширяемость жировой ткани: метаболические проблемы ожирения могут возникать из-за неспособности стать более тучным. Biochem. Soc. Пер. 36 (Pt 5), 935–940. DOI: 10.1042 / BST0360935
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тиммонс, Дж. А., Веннмальм, К., Ларссон, О., Уолден, Т. Б., Лассманн, Т., Петрович, Н. и др. (2007). Сигнатура экспрессии миогенного гена устанавливает, что коричневые и белые адипоциты происходят из разных клеточных линий. Proc. Natl. Акад. Sci. США 104, 4401–4406. DOI: 10.1073 / pnas.0610615104
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Винаик Р., Бараян Д., Абдуллахи А. и Йешке М. Г. (2019). Инфламмасома NLRP3 опосредует потемнение белой жировой ткани после ожога. Am.J. Physiol. Эндокринол. Метаб. 317, E751 – E759. DOI: 10.1152 / ajpendo.00180.2019
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Винаик Р., Станойчич М. и Йешке М. Г. (2018). Инфламмасома NLRP3 модулирует липолиз после ожога и инфильтрацию жира в печени через синтазу жирных кислот. Sci. Rep. 8: 15197. DOI: 10.1038 / s41598-018-33486-9
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Virtue, S., и Vidal-Puig, A.(2010). Расширяемость жировой ткани, липотоксичность и метаболический синдром — аллостатическая перспектива. Biochim. Биофиз. Acta 1801, 338–349. DOI: 10.1016 / j.bbalip.2009.12.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ван, К. А., Тао, К., Гупта, Р. К., и Шерер, П. Э. (2013). Отслеживание адипогенеза во время развития, расширения и регенерации белой жировой ткани. Nat. Med. 19, 1338–1344. DOI: 10,1038 / нм.3324
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Уилкокс, Г.(2005). Инсулино-инсулинорезистентность. Clin. Biochem. Rev. 26, 19–39.
Google Scholar
Волевер Т. М. (1990). Гликемический индекс. World Rev. Nutr. Диета 62, 120–185.
Google Scholar
Вулф Р. Р., Херндон Д. Н., Джахур Ф., Миёси Х. и Вулф М. (1987a). Влияние тяжелой ожоговой травмы на круговорот субстрата глюкозой и жирными кислотами. N. Engl. J. Med. 317, 403–408. DOI: 10.1056 / NEJM198708133170702
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вулф, Р.Р., Херндон, Д. Н., Питерс, Э. Дж., Джахур, Ф., Десаи, М. Х., и Холланд, О. Б. (1987b). Регуляция липолиза у детей с тяжелыми ожогами. Ann. Surg. 206, 214–221. DOI: 10.1097 / 00000658-198708000-00016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Йе, Ф. Л., Лин, В. Л., Шен, Х. Д. и Фанг, Р. Х. (1997). Изменения сывороточного фактора некроза опухоли альфа у пациентов с ожогами. Бернс 23, 6–10. DOI: 10.1016 / s0305-4179 (96) 00071-x
CrossRef Полный текст | Google Scholar
Йо, К., Ю., Ю. М., Чжао, Г., Бонаб, А. А., Айкава, Н., Томпкинс, Р. Г. и др. (2013). Коричневая жировая ткань и ее модуляция с помощью пептида, нацеленного на митохондрии, при гиперметаболизме у крыс, вызванном ожоговым повреждением. Am. J. Physiol. Эндокринол. Метаб. 304, E331 – E341. DOI: 10.1152 / ajpendo.00098.2012
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжу, К., и Шерер, П. Э. (2018). Иммунологические и эндокринные функции жировой ткани: последствия для заболевания почек. Nat. Преподобный Нефрол. 14, 105–120. DOI: 10.1038 / nrneph.2017.157
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Циммерманн Р., Штраус Дж. Г., Хеммерле Г., Шойсволь Г., Бирнер-Грюнбергер Р., Ридерер М. и др. (2004). Мобилизации жира в жировой ткани способствует липаза триглицеридов жиров. Science 306, 1383–1386. DOI: 10.1126 / science.1100747
PubMed Аннотация | CrossRef Полный текст | Google Scholar
5.4D: Метаболические функции — Медицина LibreTexts
Одна из метаболических функций кожи — выработка витамина D3, когда ультрафиолетовый свет вступает в реакцию с 7-дегидрохолестерином.
ЦЕЛЬ ОБУЧЕНИЯ
Опишите роль покровной системы в производстве витамина D
Основные выводы
Ключевые точки
- Витамин D относится к группе жирорастворимых стероидов, ответственных за увеличение абсорбции в кишечнике кальция, железа, магния, фосфата и цинка.
- Пищевых продуктов, богатых витамином D, относительно мало, поэтому организм сам синтезирует большую часть витамина D в коже.
- Дефицит витамина D связан с плохим развитием костей у детей и их размягчением у взрослых.
- Витамин D 3 образуется в коже, когда 7-дегидрохолестерин реагирует с ультрафиолетом. Витамин D вырабатывается в двух самых внутренних слоях эпидермиса. Холекальциферол (D 3 ) образуется в коже фотохимическим способом из 7-дегидрохолестерина.
- Витамин D вырабатывается в двух самых внутренних слоях эпидермиса, базальном слое и шиповидном слое.
Ключевой термин
- 7-дегидрохолестерин : 7-дегидрохолестерин является предшественником холестерина, который превращается в коже в витамин D3 и, следовательно, действует как провитамин-D3.
Покровная система — самая большая из систем органов тела, состоящая из кожи и связанных с ней придатков. Покровная система отличает, отделяет и защищает организм от окружающей среды, но также играет ключевую метаболическую функцию, являясь основной областью производства витамина D.
Что такое витамин D?
Витамин D относится к группе жирорастворимых стероидов, ответственных за увеличение кишечного всасывания кальция, железа, магния, фосфата и цинка. Для человека наиболее важными соединениями этой группы являются витамин D 3 (также известный как холекальциферол) и витамин D 2 (эргокальциферол). Холекальциферол и эргокальциферол могут поступать с пищей и с добавками, однако очень немногие продукты богаты витамином D; и поэтому синтез в коже является ключевым источником.
Дефицит витамина D связан с нарушением развития костей у детей, что приводит к развитию рахита и размягчению костей у взрослых. Дефицит витамина D был назван современным заболеванием, связанным как с плохим питанием, так и с сокращением времени, проводимого на улице.
Синтез витамина D
Витамин D: Химическая структура витамина D.
Кожа человека состоит из трех основных слоев: эпидермиса, дермы и гиподермы.Эпидермис образует самый внешний слой, обеспечивая первоначальный барьер для внешней среды. Под ним дерма состоит из двух частей, сосочкового и ретикулярного слоев, и содержит соединительные ткани, сосуды, железы, фолликулы, корни волос, сенсорные нервные окончания и мышечную ткань. Самый глубокий слой — это гиподерма, которая в основном состоит из жировой ткани.
Витамин D вырабатывается в двух самых внутренних слоях эпидермиса, базальном слое и шиповидном слое.
Витамин D 3 образуется в коже, когда 7-дегидрохолестерин реагирует с ультрафиолетовым светом типа UVB на длинах волн от 280 до 315 нм, с пиковым синтезом между 295 и 297 нм.
В зависимости от интенсивности UVB-лучей и минут воздействия в коже может развиться равновесие, и витамин D разлагается так же быстро, как и вырабатывается.
Витамин D из рациона или синтезированный организмом биологически неактивен; активация требует ферментативного преобразования в печени и почках.
Карта метаболизма и путей для витамина D: Путь синтеза витамина D
Костный метаболизм и модуляторы | Феликс Броннер
Недавнее исследование, которое Bone-Metabolic Function and Modulators расширяет, добавило новую поддержку идее о том, что кость не только служит опорной системой, но также функционирует как интегрирующий орган, играющий важную регулирующую роль для липидов и энергии. метаболизм.Связь между физической активностью и скелетом также становится все более очевидной. Этот полностью иллюстрированный том содержит самую свежую информацию о метаболической роли скелета и о том, что это может значить для лечения метаболических, а также скелетных и слуховых заболеваний.
Костная метаболическая функция и модуляторы представляет особый интерес для ученых-клиницистов, клинических и фундаментальных исследователей костей, ортопедов, эндокринологов, терапевтов, стоматологов, практикующих медсестер, медицинских и стоматологических ординаторов и физиотерапевтов, а также студентов, изучающих костно-мышечную систему.
Костная метаболическая функция и модуляторы — седьмой том из серии «Темы биологии костей» под редакцией Феликса Броннера и Мэри К. Фарач-Карсон.
Другие названия в этой серии: —
Формирование кости
Резорбция кости
Инженерия функциональных скелетных тканей
Кость и остеоартрит
Кость и рак
Кость и развитие
9000- 2 модуляторы метаболизма представляет особый интерес для ученых-клиницистов, клинических и фундаментальных исследователей костей, ортопедов, эндокринологов, терапевтов, стоматологов, практикующих медсестер, медицинских и стоматологических ординаторов и физиотерапевтов, а также студентов, изучающих костно-мышечную систему.
Костная метаболическая функция и модуляторы — седьмой том из серии «Темы биологии костей» под редакцией Феликса Броннера и Мэри К. Фарач-Карсон.
Другие названия в этой серии: —
Формирование кости
Резорбция кости
Инженерия функциональных скелетных тканей
Кость и остеоартрит
Кость и рак
Кость и развитие
9000- 2 модуляторы метаболизма представляет особый интерес для ученых-клиницистов, клинических и фундаментальных исследователей костей, ортопедов, эндокринологов, терапевтов, стоматологов, практикующих медсестер, медицинских и стоматологических ординаторов и физиотерапевтов, а также студентов, изучающих костно-мышечную систему.