Белки катаболизм — Справочник химика 21
Метаболизм включает в себя катаболизм, или расщепление пищевых веществ, богатых энергией, и анаболизм, или биосинтез новых клеточных компонентов. В катаболических и анаболических процессах различают три главные стадии. На первой стадии катаболизма полисахариды, жиры и белки расщеп- [c.398]
Все процессы, протекающие в бактериальной клетке, — образование аминокислот, нуклеотидов и других важных метаболитов, репликация, транскрипция, трансляция, катаболизм, высвобождение энергии, реакции на внешние воздействия — требуют участия белков. Однако энергетических ресурсов клетки не хватает для одновременного осуществления транскрипции и трансляции (экспрессии) всех структурных генов. Поэтому постоянно экспрессируются толь- [c.41]
Таким образом, взаимопревращение метаболитов, образующихся при катаболизме веществ разных классов, тесно связано с энергетическим обменом. Известно, что одним из энергоемких процессов в организме является биосинтез белка, и становится понятна в этом отношении интеграция этого процесса с катаболическими реакциями превращения глюкозы и триацилглицерола — основными источниками синтеза АТФ в процессе окислительного фосфорилирования. В свою очередь, все реакции углеводного и липидного обмена катализируются ферментами, являющимися белками. Следует отметить, что единство метаболических процессов находится под воздействием условий внешней среды и способность живых организмов сохранять постоянство внутренней среды — биохимический гомеостаз — при помощи механизмов саморегуляции является одним из важнейших свойств всех живых систем. [c.449]
Известно, что одним из энергоемких процессов в организме является биосинтез белка, и становится понятна в этом отношении интеграция этого процесса с катаболическими реакциями превращения глюкозы и триацилглицерола — основными источниками синтеза АТФ в процессе окислительного фосфорилирования. В свою очередь, все реакции углеводного и липидного обмена катализируются ферментами, являющимися белками. Следует отметить, что единство метаболических процессов находится под воздействием условий внешней среды и способность живых организмов сохранять постоянство внутренней среды — биохимический гомеостаз — при помощи механизмов саморегуляции является одним из важнейших свойств всех живых систем. [c.449]
Стабильность белкового состава организма — следствие устойчивого динамического равновесия, при котором количество постоянно распадающихся белков практически равно синтезируемому их количеству. Поддержание этого равновесия между анаболизмом и катаболизмом — явление тонкое, так как организм располагает лишь очень малым резервом аминокислот. В организме человека соотношение свободных аминокислот и возобновимых белков составляет приблизительно 1 5000 [65]. [c.568]
В организме человека соотношение свободных аминокислот и возобновимых белков составляет приблизительно 1 5000 [65]. [c.568]
Живая природа характеризуется рядом свойств, отличающих ее от неживой природы, и почти все эти свойства связаны с белками. Прежде всего для живых организмов характерны широкое разнообразие белковых структур и их высокая упорядоченность последняя существует во времени и пространстве. Удивительная способность живых организмов к воспроизведению себе подобных также связана с белками. Сократимость, движение — непременные атрибуты живых систем —имеют прямое отношение к белковым структурам мышечного аппарата. Наконец, жизнь немыслима без обмена веществ, постоянного обновления составных частей живого организма, т.е. без процессов анаболизма и катаболизма (этого удивительного единства противоположностей живого), в основе которых лежит деятельность каталитически активных белков—ферментов. [c.19]
Характерной особенностью белкового обмена является его чрезвычайная разветвленность. Достаточно указать, что в обмене 20 аминокислот, входящих в состав белковых молекул, в организме животных участвуют сотни промежуточных метаболитов, тесно связанных с обменом углеводов и липидов. Число ферментов, катализирующих химические реакции азотистого обмена, также исчисляется сотнями. Следует добавить, что блокирование одного какого-либо специфического пути обмена даже одной аминокислоты, обычно наблюдаемое при врожденных пороках обмена, может привести к образованию совершенно неизвестных продуктов обмена, так как возникают условия для неспецифических превращений всех предшествующих компонентов в данной цепи реакций. Отсюда становятся понятными трудности интерпретации данных о регуляции процессов азотистого обмена в норме и особенно при патологии. Этими обстоятельствами можно объяснить исключительную перспективность изучения обмена белков с целью выяснения особенностей их катаболизма и синтеза, овладение тонкими молекулярными механизмами которых, несомненно, даст в руки исследователя ключ к пониманию развития и течения патологических процессов и соответственно к целенаправленному воздействию на многие процессы жизни.
Достаточно указать, что в обмене 20 аминокислот, входящих в состав белковых молекул, в организме животных участвуют сотни промежуточных метаболитов, тесно связанных с обменом углеводов и липидов. Число ферментов, катализирующих химические реакции азотистого обмена, также исчисляется сотнями. Следует добавить, что блокирование одного какого-либо специфического пути обмена даже одной аминокислоты, обычно наблюдаемое при врожденных пороках обмена, может привести к образованию совершенно неизвестных продуктов обмена, так как возникают условия для неспецифических превращений всех предшествующих компонентов в данной цепи реакций. Отсюда становятся понятными трудности интерпретации данных о регуляции процессов азотистого обмена в норме и особенно при патологии. Этими обстоятельствами можно объяснить исключительную перспективность изучения обмена белков с целью выяснения особенностей их катаболизма и синтеза, овладение тонкими молекулярными механизмами которых, несомненно, даст в руки исследователя ключ к пониманию развития и течения патологических процессов и соответственно к целенаправленному воздействию на многие процессы жизни. [c.410]
[c.410]
Кажущаяся стабильность химического состава целостного организма является результатом существования определенного равновесия между скоростями синтеза и распада его составляющих. Внедрение в биохимическую и клиническую практику метода меченых атомов позволило доказать, что белки нужны не только растущему, но и сформировавшемуся организму, когда его рост прекратился, т.е. имеются доказательства существования в организме механизма постоянного обновления химических составных частей тела. При нормальных физиологических условиях, как и при патологических состояниях, скорости синтеза и распада специфических веществ определяются, помимо нервно-гормонального влияния, химической природой веществ и внутриклеточной их локализацией. В растущем организме скорость синтеза многих компонентов органов и тканей преобладает над скоростью их распада. Тяжелые изнуряющие болезни, а также голодание, напротив, характеризуются преобладанием скорости катаболизма над скоростью синтеза. Почти все белки тела, включая структурные белки, гемоглобин, белки плазмы и других биологических жидкостей организма, также подвергаются постепенному распаду и синтезу. Например, более половины белков печени, сыворотки крови и слизистой оболочки кишечника подвергается распаду и ресинтезу в течение 10 дней. Медленнее обновляются белки мышц, кожи и мозга. [c.410]
Почти все белки тела, включая структурные белки, гемоглобин, белки плазмы и других биологических жидкостей организма, также подвергаются постепенному распаду и синтезу. Например, более половины белков печени, сыворотки крови и слизистой оболочки кишечника подвергается распаду и ресинтезу в течение 10 дней. Медленнее обновляются белки мышц, кожи и мозга. [c.410]
Сериновые протеазы широко распространены в природе и вместе с протеолитическими ферментами других классов (аспартильными, цистеиновыми и металлопротеиназами) обеспечивают расщепление белков (катаболизм) и целый ряд реакций ограниченного протеолиза, имеющих регуляторное значение для жизни клетки. [c.36]
Как было указано, обмен веществ в организме человека протекает не хаотично он интегрирован и тонко настроен. Все превращения органических веществ, процессы анаболизма и катаболизма тесно связаны друг с другом. В частности, процессы синтеза и распада взаимосвязаны, координированы и регулируются нейрогормональными механизмами, придающими химическим процессам нужное направление. В организме человека, как и в живой природе вообще, не существует самостоятельного обмена белков, жиров, углеводов и нуклеиновых кислот. Все превращения объединены в целостный процесс метаболизма, подчиняющийся диалектическим закономерностям взаимозависимости и взаимообусловленности, допускающий также взаимопревращения между отдельными классами органических веществ. Подобные взаимопревращения диктуются физиологическими потребностями организма, а также целесообразностью замены одних классов органических веществ другими в условиях блокирования какого-либо процесса при патологии. [c.545]
В организме человека, как и в живой природе вообще, не существует самостоятельного обмена белков, жиров, углеводов и нуклеиновых кислот. Все превращения объединены в целостный процесс метаболизма, подчиняющийся диалектическим закономерностям взаимозависимости и взаимообусловленности, допускающий также взаимопревращения между отдельными классами органических веществ. Подобные взаимопревращения диктуются физиологическими потребностями организма, а также целесообразностью замены одних классов органических веществ другими в условиях блокирования какого-либо процесса при патологии. [c.545]
Транскрипция, контролируемая /ас-промотором, регулируется также с помощью белка — активатора катаболизма (САР) (рис. 6.2). При связывании САР с промотором повышается сродство последнего к РНК-полимеразе и усиливается транскрипция примыкающих к нему генов. В свою очередь сродство САР к промотору повышается при его связывании с цикличе- [c.107]
Желчные пигменты млекопитающих — это продукты окислительного расщепления гема (красящего вещества крови, см. далее в этом разд.). При этом первичным продуктом гемового катаболизма является тетрапиррол 6.69. Однако его в организме немного, так как он быстро восстанавливается специальным ферментом до билирубина. Ни первый, ни второй в воде нерастворимы. В комплексе с белками крови билирубин попадает в печень, где происходит его конъюгация, т.е. перевод в водорастворимую форму путем реакции с глюкуроновой кислотой. Водорастворимый глюкуронид 6.71 выделяется в желчь и с нею через кишечник выводится из организма. Если этот процесс нарушается на какой-либо стадии, происходит накопление билирубина в крови. От этого кожные покровы у человека окрашиваются в желтый цвет. Развивается заболевание, именуемое желтухой. На самом деле это несколько заболеваний, так как причины накопления билирубина могут быть разными. Чаще всего нарушается функция печени, и этот орган в той или иной степени теряет способность конъюгировать желчный пигмент. [c.444]
далее в этом разд.). При этом первичным продуктом гемового катаболизма является тетрапиррол 6.69. Однако его в организме немного, так как он быстро восстанавливается специальным ферментом до билирубина. Ни первый, ни второй в воде нерастворимы. В комплексе с белками крови билирубин попадает в печень, где происходит его конъюгация, т.е. перевод в водорастворимую форму путем реакции с глюкуроновой кислотой. Водорастворимый глюкуронид 6.71 выделяется в желчь и с нею через кишечник выводится из организма. Если этот процесс нарушается на какой-либо стадии, происходит накопление билирубина в крови. От этого кожные покровы у человека окрашиваются в желтый цвет. Развивается заболевание, именуемое желтухой. На самом деле это несколько заболеваний, так как причины накопления билирубина могут быть разными. Чаще всего нарушается функция печени, и этот орган в той или иной степени теряет способность конъюгировать желчный пигмент. [c.444]
В организме синтез и распад белков тесно взаимосвязаны. Постоянство массы и белкового состава взрослого организма является результатом тонко настроенного равновесия между анаболизмом и катаболизмом. Аминокислоты белков пищи и аминокислоты, возникающие в результате распада белков тканей в процессе самообновления, составляют общий фонд аминокислот организма, равный приблизительно 500 г. [c.167]
Постоянство массы и белкового состава взрослого организма является результатом тонко настроенного равновесия между анаболизмом и катаболизмом. Аминокислоты белков пищи и аминокислоты, возникающие в результате распада белков тканей в процессе самообновления, составляют общий фонд аминокислот организма, равный приблизительно 500 г. [c.167]
Рассмотрим теперь катаболизм более подробно. Ферментативное расшепление тех главных питательных веществ, которые служат клетке источником энергии, а именно углеводов, жиров и белков совершается постепенно-через ряд последовательных ферментативных реакций. В аэробном катаболизме различают три главные стадии (рис. 13-6). На стадии [c.380]
Если в рационе недостает одной из незаменимых аминокислот, то белковый синтез будет продолжаться до тех пор, пока запас этой аминокислоты не истощится. Чтобы пополнить запас, необходимо дополнительное потребление несбалансированной смеси аминокислот или разрушение белков организма. При этом неизбежно возникнет избыток других аминокислот, которые должны подвергнуться катаболизму, что приведет к вьщелению азота и в результате к отрицательному азотному балансу. [c.1001]
При этом неизбежно возникнет избыток других аминокислот, которые должны подвергнуться катаболизму, что приведет к вьщелению азота и в результате к отрицательному азотному балансу. [c.1001]
Благодаря использованию большого набора мутаций по промоторам и генам активирующих белков дрожжей удалось выяснить некоторые особенности взаимодействия белков-активаторов с АП, а также характерные свойства этих белков. Белок GAL4 активирует гены, необходимые для катаболизма галактозы. GAL4 связывается с АП, представленной повторяющимися элементами по 17 п. н-Степень активирующего действия пропорциональна числу этих элементов в промоторе. Функция связывания ДНК и активации транскрипции принадлежит разным участкам белка GAL4, который содержит 881 аминокислоту. 73 остатка с N-конца молекулы белка достаточны для обеспечения специфического связывания с ДНК. Эгот участок связывает ионы цинка и содержит характерную структуру — цинковые пальцы , обнаруженные в целом ряде белков, активирующих транскрипцию (см. раздел 4 этой главы). Два других дискретных участка белка, включающих аминокислоты 149—196 и 768—881, достаточны для обеспечения активации транскрипции. Эти участки содержат кислые аминокислотные остатки. По-видимому, в разных активаторных белках эти районы обладают [c.196]
раздел 4 этой главы). Два других дискретных участка белка, включающих аминокислоты 149—196 и 768—881, достаточны для обеспечения активации транскрипции. Эти участки содержат кислые аминокислотные остатки. По-видимому, в разных активаторных белках эти районы обладают [c.196]
В обоих случаях регуляция осуществляется с помощью специальных белков, нацеленных на регуляцию определенного оперона. Существуют аналогичные регуляторные белки с более широким спектром действия. Важнейшим их представителем у бактерий является белок активатор катаболизма (САР, atabolite. [c.429]
КАТЕПСИНЫ, ферменты класса гидролаз внутриклеточные протеиназы. Играют важную роль в катаболизме белков катализируют гидролиз белков и пептидов в слабокислой среде. Для формы А из мышц карпа (мол. м. 34 ООО) оптнм. каталитич. активность при pH 4,6—5,8 обладает пептидазной активностью и гидролизует эфиры и амиды [c.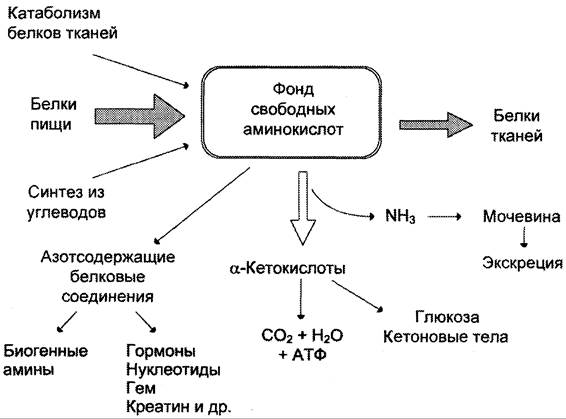 249]
249]
Адреналин и глюкагон осуществляют регуляцию метаболизма гликогена путем изменения активности гликогенфосфорилазы и гликогенсинтазы (через цАМФ) таким образом, что торможение гликогеногенеза и стимуляция гликогенолиза осуществляются одновременно, т. е. реципропно. Глюкокортикоиды (11-гидроксистероиды) усиливают глюконеогенез за счет интенсификации катаболизма белков и аминокислот в тканях и вовлечения промежуточных метаболитов в процесс глюконеогенеза. Таким образом, в рассмотренных случаях адреналин, глюкагон, глюкокортикоиды действуют как антагонисты инсулина. На содержание сахара в крови влияет также гормон щитовидной железы тироксин (подобно инсулину). Гормоны передней доли гипофиза — гормон роста (соматотропин), АКТГ и, вероятно, другие факторы повышают уровень сахара в крови, однако механизмы действия этих гормонов в значительной степени являются опосредованными, поскольку они стимулируют мобилизацию из жировой ткани свободных жирньгх кислот, которые являются ингибиторами потребления глюкозы. [c.283]
[c.283]
Катаболизм белков у всех организмов начинается с их расщепления по пептидным связям протеолитич. ферментами. В желудочно-кишечном тракте животных белки гидролизуются трипсином, химотрипсином, пепсином и др. ментами до своб. аминокислот, к-рые всасываются стенками кишечника и попадают в кровоток. Часть аминокислот подвергается дезаминированию до оксокислот, претерпевающих дальнейшее расщепление, др. часть используется печенью или тканями организма для биосинтеза белков. У млекопитающих отщепляющийся от аминокислот аммиак превращ. в орнитиновом х икле в мочевину. Этот процесс осуществляется в печени. Образующаяся мочевина вместе с др. р-римыми продуктами О.в. выводится из кровотока почками. [c.315]
Кроме того, ТДФ принимает участие в окислит, декарбоксилировании кетокислот с разветвленным углеродным скелетом — 2-оксоизовалериановой, З-метил-2-оксовалериано-вой и 4-метил-2-оксопентановой, являющихся продуктами дезаминирования аминокислот валина, изолейцина и лейцина. Эти р-ции играют важную роль в катаболизме белков. [c.564]
Эти р-ции играют важную роль в катаболизме белков. [c.564]
Несмотря на большое число исследований, чисто химический аспект действия инсулина остается неясным — . Обычно считается, что гормон действует на плазматические мембраны всех тканей, вызывая заметные изменения проницаемости, что поиводит к возрастанию поглощения глюкозы, различных ионов и других веществ. Такого рода изменения проницаемости могут обусловить сильное влияние инсулина на важнейшие процессы биосинтеза имеет место, в частности, повышение синтеза гликогена, липидов и белков. В то же время процессы катаболизма подавляются и активность катаболических ферментов, например глюкозо-6-фосфатазы, снижается. Ключом к пониманию действия инсулина может явиться выяснение вопроса о природе его вторичного посредника , аналогичного по своему действию сАМР. Высказывались предположения, что вторичным посредником для инсулина является сАМР, однако более вероятно, что эту роль выполняет какой-то ион, возможно К+ . [c.505]
[c.505]
Промежуточный метаболизм складывается из двух фаз-катаболизма и анаболизма. Катаболизм-это фаза, в которой происходит расщепление сложных органических молекул до более простых конечных продуктов. Углеводы, жиры и белки, поступившие извне с пищей или присутствующие в самой клетке в качестве запасных веществ, распадаются в серии последовательных реакций до таких соединений, как молочная кислота, СО 2 и аммиак. Катаболические процессы сопровождаются высвобождением свободной энергии, заключенной в сложной структуре больших органических молекул. На определенных этапах соответствующих катаболических путей значительная часть свободной энергии запасается благодаря сопряженным ферментативным реакциям в форме высокоэнергетического соединения — аденозинтрифосфата (АТР). Часть ее запасается также в богатых энергией водородных атомах кофермента никотинамид адениндинуклеотидфосфата, находящегося в [c.379]
Продукты катаболизма пиримидинов либо выводятся из организма, либо повторно утилизируются в других метаболических процессах. Так, р-аланин используется при биосинтезе витамина В3 (пантотеновая кислота), который, в свою очередь, необходим для синтеза коэнзима А и ацилпереносящего белка — компонента, участвующего в синтезе жирных кислот. [c.428]
Так, р-аланин используется при биосинтезе витамина В3 (пантотеновая кислота), который, в свою очередь, необходим для синтеза коэнзима А и ацилпереносящего белка — компонента, участвующего в синтезе жирных кислот. [c.428]
Катаболизм — фаза метаболизма, включающая ферментивное расщепление белков, липидов, углеводов и других веществ и сопрово)цдаю-щаяся выделением энергии. [c.141]
Клеточный метаболизм основан на принципе максимальной экономии. Общая скорость катаболизма, обеспечивающего клетку энергией, определяется не просто наличием или концентрацией клеточного топлива она обусловлена потребностью клетки в энергии в форме АТР и NADPH. Клетка потребляет в каждый данный момент как раз такое количество питательных веществ, какое позволяет ей удовлетворять свои энергетические нужды. Точно так же обусловлена потребностями данного момента скорость синтеза строительных блоков и макромолекул клетки. В растущих клетках, например, все 20 видов аминокислот синтезируются как раз с такой скоростью и в таких соотношениях, какие необходимы для того, чтобы обеспечить сборку новых белков, требующихся в данный момент. Таким образом, ни одна из 20 аминокислот не вырабатывается в избытке и не остается без использования. У многих животных и растений в организме откладываются запасные питательные вещества, способные служить источником энергии и углерода. Такими запасными питательными веществами являются, в частности, жиры и углеводы. [c.388]
Таким образом, ни одна из 20 аминокислот не вырабатывается в избытке и не остается без использования. У многих животных и растений в организме откладываются запасные питательные вещества, способные служить источником энергии и углерода. Такими запасными питательными веществами являются, в частности, жиры и углеводы. [c.388]
Синтез белка у прокариот регулируется главным образом на уровне транскрипции ДНК, т. е. на уровне образования мРНК. Транскрипция группы метаболически связанных между собой генов регулируется путем присоединения (или отделения) особого белка-репрессора к операторному участку ДНК. Оператор и группа связанных друг с другом генов вместе составляют оперон. Транскрипция такой группы генов может индуцироваться специфическим питательным субстратом, например лактозой. Лактоза может связывать репрессор и вызывать тем самым его отделение от оператора. Благодаря этому разрешается транскрипция генов, кодирующих белки, необходимые клетке для использования лактозы в качестве источника углерода и энергии. Некоторые опероны имеют также промоторный участок, содержащий регуляторную частъ-так называемый САР-участок последний предназначен для связывания комплекса, образованного белком, активирующим катаболитный ген (САР), и сАМР. Этот комплекс, формирующийся при отсутствии в среде глюкозы, дает возможность РНК-полимеразе присоединиться к месту инициации транскрипции генов, ответственных за катаболизм лактозы. [c.961]
Некоторые опероны имеют также промоторный участок, содержащий регуляторную частъ-так называемый САР-участок последний предназначен для связывания комплекса, образованного белком, активирующим катаболитный ген (САР), и сАМР. Этот комплекс, формирующийся при отсутствии в среде глюкозы, дает возможность РНК-полимеразе присоединиться к месту инициации транскрипции генов, ответственных за катаболизм лактозы. [c.961]
Превращения веществ в клетке (обмен веществ, или метаболизм), в результате которых из сравнительно простых предшественников, например глюкозы, жирных кислот с длинной цепью или ароматических соединений, образуется новое клеточное вещество, можно ради простоты подразделить на три основные группы. Сначала питательные вещества расщепляются на небольшие фрагменты (распад, или катаболизм), а затем в ходе реакций промежуточного обмена, или амфиболизма, они превращаются в ряд органических кислот и фосфорных эфиров. Эти два пути переходят незаметно один в другой. Многообразные низкомолекулярные соединения-это тот субстрат, из которого синтезируются основные строительные блоки клетки. Строительными блоками мы называем аминокислоты, пуриновые и пиримидиновые основания, фос-форилированные сахара, органические кислоты и другие метаболиты — конечные продукты цепей биосинтеза, иногда длинных. Из них строятся полимерные макромолекулы (нуклеиновые кислоты, белки, резервные вещества, компоненты клеточной стенки и т.п.), из которых состоит клетка. Эти два этапа биосинтеза клеточных веществ-синтез строительных блоков и синтез полимеров-составляют синтетическую ветвь метаболизма, или анаболизм (рис. 7.1). [c.214]
Многообразные низкомолекулярные соединения-это тот субстрат, из которого синтезируются основные строительные блоки клетки. Строительными блоками мы называем аминокислоты, пуриновые и пиримидиновые основания, фос-форилированные сахара, органические кислоты и другие метаболиты — конечные продукты цепей биосинтеза, иногда длинных. Из них строятся полимерные макромолекулы (нуклеиновые кислоты, белки, резервные вещества, компоненты клеточной стенки и т.п.), из которых состоит клетка. Эти два этапа биосинтеза клеточных веществ-синтез строительных блоков и синтез полимеров-составляют синтетическую ветвь метаболизма, или анаболизм (рис. 7.1). [c.214]
Обмен веществ
Что такое метаболизм, он же обмен веществ? Это процесс химических реакций в организме человека, который состоит из катаболизма и анаболизма. В первом случае происходит расщепление сложных веществ (белки, жиры и углеводы) до простых, а также протекает окисление различных молекул — все это приводит к высвобождению энергии, которая необходима для существования, из калорий еды.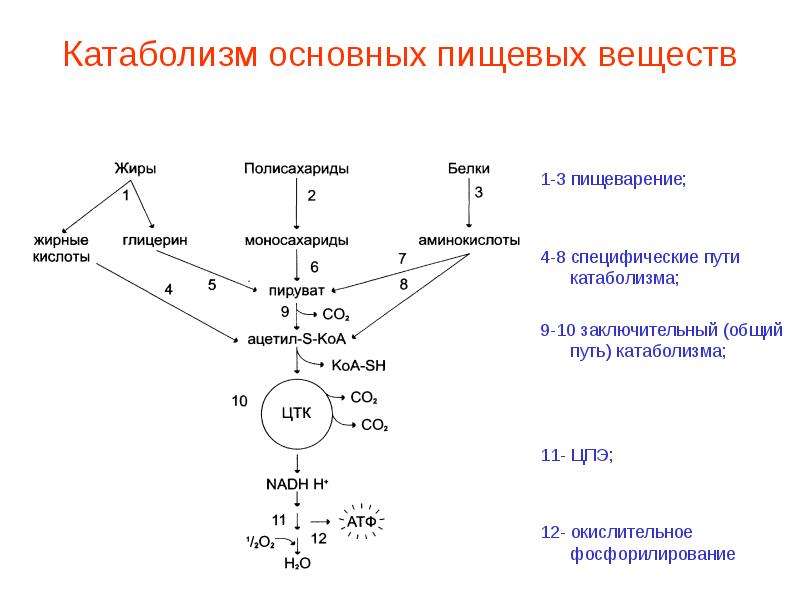 Анаболизм же можно охарактеризовать, как синтез тканевых, клеточных, соединительных структур.
Анаболизм же можно охарактеризовать, как синтез тканевых, клеточных, соединительных структур.
Что приводит к нарушению обмена веществ?
Для начала разберемся, что, с медицинской точки зрения, называется нарушением метаболизма. Это неправильная работа связей между биохимическими цепочками — например, замедление, ускорение или полное отсутствие процесса катаболизма или же анаболизма.
К основным причинам, приводящим к сбоям в обмене веществ, относят следующие:
- Генетические мутацию и врожденные патологии;
- Неправильное питание: переедание, частые диеты;
- Перенесенные вирусные болезни;
- Гормональные нарушения;
- Паразиты;
- Стрессы, депрессия;
- Сидячий образ жизни;
- Послеродовые нарушения — у женщины сбивается гормональный фон;
- Возраст;
- Гормональные контрацептивы;
- Курение, злоупотребление алкоголем.
Все вышеперечисленные факторы могут спровоцировать серьезные болезни.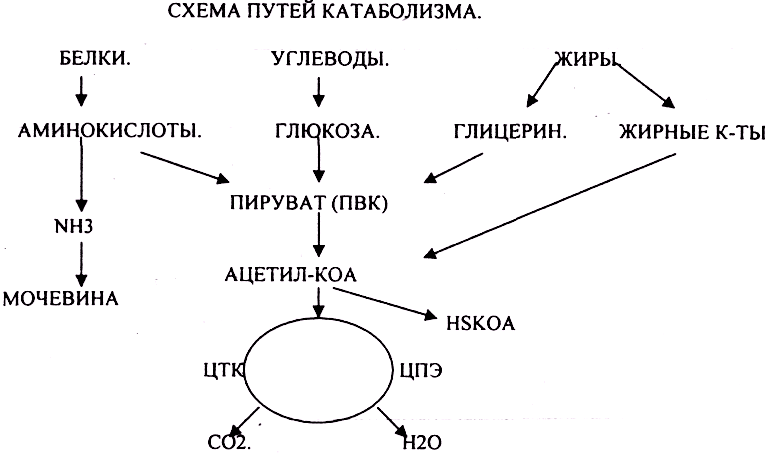 Среди них сахарный диабет, так как организм человека утрачивает способность усваивать глюкозу. Нередко встречаются поражения сердца и суставов, развитие атеросклероза, избыточная масса тела, проблемы с костями и мышцами, анемия.
Среди них сахарный диабет, так как организм человека утрачивает способность усваивать глюкозу. Нередко встречаются поражения сердца и суставов, развитие атеросклероза, избыточная масса тела, проблемы с костями и мышцами, анемия.
Для того чтобы опровергнуть или подтвердить проблемы с обменом веществ, лечащий врач, как правило, назначает целый комплекс обследований. Это общий и биохимический анализы крови и мочи, тест на глюкозу, измерение холестерина в организме человека, КТ, ЭКГ, липидный профиль. Также при необходимости добавляют ультразвуковые исследования органов эндокринной системы.
После того как пациент прошел обязательные обследования, доктор составляет для больного индивидуальный план лечения. Обычно в него входят активный образ жизни, соблюдение режима дня, сбалансированное и полезное питание, витаминные и лекарственные препараты.
Кроме того, любой человек может заняться профилактикой сбоев метаболизма. Для этого необходимо не сидеть на «голодных» диетах, избегать переедания, сладкой и жирной пищи, употреблять продукты, насыщенные витаминами и минералами, тренировать стрессоустойчивость, избавиться от пагубных привычек.
Помните, что профилактикой является и своевременное обращение к специалисту. Если вы обнаружили у себя несколько симптомов нарушения обмена веществ, то не откладывайте визит к врачу в дальний ящик.
Катаболизм
Катаболизм
Катаболизм (от греч. katabole — сбрасывание, разрушение), совокупность химических процессов, составляющих противоположную анаболизму сторону обмена веществ.Катаболизм это первый враг спортсмена на пути сохранения приобретенных путем силовых тренировок мышц. В результате катаболизма происходит разрушение мышечной ткани: белки распадаются на исходные аминокислоты, которые частично идут на биосинтез, а частично просто выводятся из организма. Такие гормоны, как кортизол, глюкагон и адреналин сдвигают баланс обмена веществ в сторону преобладания катаболизма, и в том числе распада мышечной ткани. Стрессы, ограничительные диеты, чрезмерные нагрузки и недосыпания создают неблагоприятный гормональный фон, что негативно сказывается на состоянии мышечной ткани. На мышечную массу влияют два различных фактора: скорость анаболизма, увеличивающего мышцу, и скорость катаболизма, уменьшающего её. Два этих процесса происходят постоянно в мышцах. Если скорость анаболизма выше скорости катаболизма, то вы наращиваете массу. Если скорость катаболизма выше, вы теряете мышечную массу. По окончании тренировки повышенный фон кортизола так и остается на высоком уровне, продолжая во время отдыха «сжигать» ткани нашего организма, особенно мышцы, превращая аминокислоты мышечной ткани в глюкозу, хотя в это время нам нужно совсем другое, нам нужно восстанавливать потраченные во время тренировки ресурсы и восстанавливать, добиваясь прогресса в увеличении выносливости, мышечной массы и силы.Чем сильнее тренировочный стресс, тем больше мы расходуем энергии в единицу времени, тем ниже уровень глюкозы в крови и тем сильнее сигнал о необходимости секретировать как можно большее количество кортизола, что бы этот уровень глюкозы повысить, превратив аминокислоты мышечной ткани (преимущественно ВСАА, глютамин) в глюкозу.
На мышечную массу влияют два различных фактора: скорость анаболизма, увеличивающего мышцу, и скорость катаболизма, уменьшающего её. Два этих процесса происходят постоянно в мышцах. Если скорость анаболизма выше скорости катаболизма, то вы наращиваете массу. Если скорость катаболизма выше, вы теряете мышечную массу. По окончании тренировки повышенный фон кортизола так и остается на высоком уровне, продолжая во время отдыха «сжигать» ткани нашего организма, особенно мышцы, превращая аминокислоты мышечной ткани в глюкозу, хотя в это время нам нужно совсем другое, нам нужно восстанавливать потраченные во время тренировки ресурсы и восстанавливать, добиваясь прогресса в увеличении выносливости, мышечной массы и силы.Чем сильнее тренировочный стресс, тем больше мы расходуем энергии в единицу времени, тем ниже уровень глюкозы в крови и тем сильнее сигнал о необходимости секретировать как можно большее количество кортизола, что бы этот уровень глюкозы повысить, превратив аминокислоты мышечной ткани (преимущественно ВСАА, глютамин) в глюкозу. Этот процесс называется глюконеогенез – образование глюкозы из не углеводных источников.Но катаболизм возникает не только после тренировки. Утро — вот настоящий ужас для спортсмена. Как только мы просыпаемся организму в огромных количествах нужны аминокислоты и глюкоза. И именно с этой повышенной секрецией кортизола нам и следует бороться принимая Протеин, либо аминокислотные комплексы, либо BCAA они отлично восполнят запас аминокислот в организме. Можно ещё во время тренеровки добавлять глюкозу (чем ниже уровень глюкозы в крови и тем сильнее сигнал о необходимости секретировать как можно большее количество кортизола) в аминокислоты или ВСАА, в этом случае происходит выброс инсулина, который так же является антикатаболиком. Не следует добавлять глюкозу если вы находитесь на сбросе жира так как инсулин остановит движение жирных кислот на переработку в энергию, тут следует увеличить дозировку аминокислот,ВСАА что бы катаболизм не добрался до ваших мышц, по этой причине как ни странно бы это звучало белка нам нужно больше на сбросе жира, а не на наборе массы.
Этот процесс называется глюконеогенез – образование глюкозы из не углеводных источников.Но катаболизм возникает не только после тренировки. Утро — вот настоящий ужас для спортсмена. Как только мы просыпаемся организму в огромных количествах нужны аминокислоты и глюкоза. И именно с этой повышенной секрецией кортизола нам и следует бороться принимая Протеин, либо аминокислотные комплексы, либо BCAA они отлично восполнят запас аминокислот в организме. Можно ещё во время тренеровки добавлять глюкозу (чем ниже уровень глюкозы в крови и тем сильнее сигнал о необходимости секретировать как можно большее количество кортизола) в аминокислоты или ВСАА, в этом случае происходит выброс инсулина, который так же является антикатаболиком. Не следует добавлять глюкозу если вы находитесь на сбросе жира так как инсулин остановит движение жирных кислот на переработку в энергию, тут следует увеличить дозировку аминокислот,ВСАА что бы катаболизм не добрался до ваших мышц, по этой причине как ни странно бы это звучало белка нам нужно больше на сбросе жира, а не на наборе массы.
| Обычно катаболизм высвобождает энергию, а анаболизм потребляет энергию. | |
| Катаболизм аминокислот приводит к отходам аммиака. | |
| Она включает в себя прием пищи, всасывание, ассимиляцию, биосинтез, катаболизм и экскрецию. | |
| Однако добавки с витамином А будут полезны только в исключительных случаях, потому что обычно местный катаболизм витамина А будет регулирующим фактором. | |
| Катаболизм расщепляет молекулы, а анаболизм собирает их вместе. | |
| Катаболизм-это совокупность метаболических процессов, которые разрушают крупные молекулы. | |
| Катаболизм белка включает цикл мочевины, который расположен в печени и может легко подавить способность уже поврежденного органа. | |
Во-вторых, было высказано предположение, что витамин С также играет определенную роль в регуляции липидов, контролируя катаболизм холестерина и желчных кислот. |
|
| Второе — это катаболизм, распад молекул. | |
| Начальный катаболизм фруктозы иногда называют фруктолизом, по аналогии с гликолизом, катаболизмом глюкозы. | |
| Катаболизм-это метаболическая реакция, которую клетки претерпевают, чтобы расщепить более крупные молекулы, извлекая энергию. | |
| Моносахаридные единицы могут затем вступать в катаболизм моносахаридов. | |
| Кроме того, курение может еще больше ускорить катаболизм биотина у женщин. | |
| Другие результаты | |
| В биологии ферменты являются белковыми катализаторами метаболизма и катаболизма. | |
| Метаболизм клетки достигает этого путем соединения самопроизвольных процессов катаболизма с не-самопроизвольными процессами анаболизма. | |
| Метаболизм-это совокупность анаболизма и катаболизма. | |
Фосфорилирование сахаров часто является первой стадией их катаболизма. |
|
| Дикарбоновые кислоты могут быть получены путем ω-окисления жирных кислот в процессе их катаболизма. | |
| К ним относятся азотистые отходы мочевины, образующиеся в результате катаболизма белков, и мочевая кислота, образующаяся в результате метаболизма нуклеиновых кислот. | |
| Типичный коричневый цвет отходов жизнедеятельности млекопитающих обусловлен билирубином-продуктом распада нормального катаболизма гема. | |
| Другие возможные механизмы включают антагонизм серотонина и гистамина, а также снижение катаболизма тканевых мукополисахаридов. | |
| Только лакз и Лейси, по-видимому, необходимы для катаболизма лактозы. | |
| Этот механизм преобладает во время катаболизма, усиливаясь по мере нарастания тяжести голодания и голодания. | |
| Синтез жирных кислот требует ацетилкоа, который происходит, в частности, от катаболизма глюкозы. | |
Эти соединения образуются при катаболизме углеводов и липидов в реакциях конденсации, а также в некоторых других метаболических реакциях аминокислот. |
|
| Чтобы быть использованным в клеточном катаболизме, он должен сначала быть преобразован в глюкозу 6-фосфат ферментом фосфоглюкомутазой. | |
| Биотин играет важную роль в синтезе жирных кислот, катаболизме аминокислот с разветвленной цепью и глюконеогенезе. | |
| Анаболизм-это совокупность конструктивных метаболических процессов, при которых высвобождаемая катаболизмом энергия используется для синтеза сложных молекул. | |
| АТФ действует как мост между катаболизмом и анаболизмом. | |
| Это проявляется усилением клеточного дыхания, катаболизмом белков и метаболическим ацидозом с компенсаторным дыхательным алкалозом. | |
≡ Восстановление обмена веществ и нормализация веса в г. Днепр
Для чего нужен метаболизм? Он позволяет нашему телу расти, размножатся, заживлять повреждения и реагировать на окружающую среду. Но более распространённое выражение метаболизма — называют процесс переработки употребляемой человеком пищи. Этот процесс делится на анаболизм и катаболизм.
Этот процесс делится на анаболизм и катаболизм.
Анаболизм — процесс биосинтеза в результате которого, из простых веществ (молекул) образуются более сложные вещества (например, мышечные волокна). Очень энергетически затратный процесс.
Катаболизм — это процесс, наоборот, когда происходит распад сложных веществ на простые элементы, а также выделяется необходимая энергия.
Анаболизм и Катаболизм не могут существовать друг без друга. Если мы тратим такое же количество энергии, что получаем из пищи, у нас не появится проблем ни с весом, ни с общим здоровьем. Причем нарушение этого баланса в какую-то из сторон одинаково вредно. Когда количество съеденных калорий больше, чем количество затраченных, мы постепенно оплываем жирком в неположенных местах. Когда же калорий слишком мало, начинает разрушаться организм, пытаясь найти энергию для своей жизнедеятельности. И хорошо если он берёт энергию из жировых отложений. Хуже будет тогда, когда начинает разрушаться мышечная ткань. Чтобы вы знали. Небольшая часть полученной и нерастраченной энергии складывается в виде углевода гликогена – источника энергии для активной работы мышц. Он запасается в самих мышцах и печени. Остальное накапливается в жировых клетках. Причём для их образования и жизни требуется гораздо меньше энергии, чем для постройки мышц или костей. Даже если вы двигаетесь очень много, нужно, так или иначе, следить за потребляемыми калориями, чтобы их не оказалось больше положенного.
Чтобы вы знали. Небольшая часть полученной и нерастраченной энергии складывается в виде углевода гликогена – источника энергии для активной работы мышц. Он запасается в самих мышцах и печени. Остальное накапливается в жировых клетках. Причём для их образования и жизни требуется гораздо меньше энергии, чем для постройки мышц или костей. Даже если вы двигаетесь очень много, нужно, так или иначе, следить за потребляемыми калориями, чтобы их не оказалось больше положенного.
Как метаболизм связан с массой тела? Можно сказать, что вес тела — это катаболизм минус анаболизм. Другими словами, разница между количеством поступившей в организм энергии и использованной им.
Несложный пример: съедаете бутерброд (колбаса с хлебом, самый простой) и ложитесь на диван. Из хлеба и колбасы организм получил жиры, белки, углеводы и 140 ккал. При этом лежащее тело потратит полученные калории только на расщепление съеденной пищи и немного на поддержание функций дыхания и кровообращения – около 50 ккал в час. Остальные 90 ккал превратятся в 10 г жира и отложатся в жировое депо. Если же любитель бутербродов выйдет на спокойную прогулку, полученные калории организм потратит примерно за час.
Остальные 90 ккал превратятся в 10 г жира и отложатся в жировое депо. Если же любитель бутербродов выйдет на спокойную прогулку, полученные калории организм потратит примерно за час.
Нормализация веса в клинике Health Partner
«Хороший» и «плохой» метаболизм? Исследования показали, что замедленный метаболизм наблюдается при ряде заболеваний, например, гипотиреозе — недостатке гормона щитовидной железы. Тогда как у большинства людей с лишним весом наблюдается энергетический дисбаланс. То есть энергии в организм поступает гораздо больше, чем её нужно на самом деле. И именно она складируется про запас Поэтому и стоит задуматься о восстановление обмена веществ и нормализации веса.
Статьи расхода калорий
Чтобы расход и получение калорий держать под контролем, стоит помнить основные направления дополнительных энергозатрат.
- Чем выше масса тела, тем больше калорий ему требуется. Но, как мы знаем, жировой ткани надо совсем мало энергии для жизни, а вот мышечная потребляет достаточно.
 Поэтому 100-килограммовый культурист потратит как минимум в два раза больше калорий на ту же работу, что и его 100 — килограммовый ровесник с неразвитыми мышцами и высоким процентом жира.
Поэтому 100-килограммовый культурист потратит как минимум в два раза больше калорий на ту же работу, что и его 100 — килограммовый ровесник с неразвитыми мышцами и высоким процентом жира. - Чем старше становится человек, тем выше у него разница между поступлением энергии и её тратами за счёт гормонального дисбаланса и резкого снижения физической активности.
- В метаболизме мужского организма активно участвует гормон тестостерон. Это настоящий естественный анаболик, заставляющий организм тратить энергию и ресурсы на выращивание дополнительных мышц. Именно поэтому мышечная масса у мужчин обычно гораздо выше, чем у женщин. А поскольку на поддержание жизнедеятельности мышц требуется гораздо больше энергии, чем для сохранения жира, то мужчина и женщина одного роста и веса тратят неодинаковое количество калорий на одни и те же действия. Проще говоря: мужчины больше тратят энергии, им требуется больше еды, а при желании они гораздо быстрее худеют.
Простая формула расчёта калорий Для мужчины: (9,99 умножить на вес в кг) + (6,25 умножить на рост в см) — (4. 92 умножить на возраст в годах) + 5 = ;
92 умножить на возраст в годах) + 5 = ;
Для женщины: (9,99 умножить на вес в кг) + (6,25 умножить на рост в см) — (4,92 умножить на возраст в годах) — 161 = .
Чтобы было понятно, как считать, возьмём женщину 30 лет весом 70 кг и ростом 160 сантиметров: (9,99×70) + (6,25×160)— (4,92×30) – 161 = (699,3 + 1000 – 196,8— 161) = 1390,7 ккал— это то количество калорий, меньше которого давать организму нельзя, иначе метаболизм начнёт замедляться в целях сохранить энергию, так как для него наступают голодные дни, а значит, пора делать запасы.
Восстановление обмена веществ в частной клинике
Как влияют физические нагрузки и спорт на нормализацию метаболизм и восстановление обмена веществ? Нередко те, кому нужна нормализация веса, идут в тренажерный зал, начинают активно заниматься спортом, чтобы увеличить энергозатраты и таким путём избавиться от лишних килограммов. Чтобы вес и объёмы успешно покидали ваше тело, важно владеть информацией, как различные виды физической нагрузки воздействуют на организм. Клиника в г. Днепр знает как помочь Вам для нормализация веса и восстановления обмена веществ.
Клиника в г. Днепр знает как помочь Вам для нормализация веса и восстановления обмена веществ.
Физические нагрузки состоят из двух основных видов: аэробные и анаэробные. К первым относят бег, интенсивную ходьбу, различные виды танцев, аэробику, велосипед, ко вторым относят тренировки с различными отягощениями. С их помощью тратится много калорий, так как обычно они проводятся интенсивно, расход энергии происходит, даже когда вы уже не тренируетесь, во сне, в том числе. Однако чтобы такие чудеса происходили, нужно иметь мышцы, привычные к нагрузкам, чтобы в течение определённого времени проводить подобную тренировку. Занимаясь, например, бегом, хороших результатов вряд ли можно добиться, если мышечный корсет развит слабо. Чтобы исправить ситуацию и развить мышечный корсет, стоит сначала заняться анаэробными, силовыми упражнениями, а уже потом подключить аэробные. У кого же мышечный корсет уже достаточно развит, чтобы создать красивую фигуру и укрепить здоровье, хорошо подойдёт сочетание силовых и аэробных нагрузок.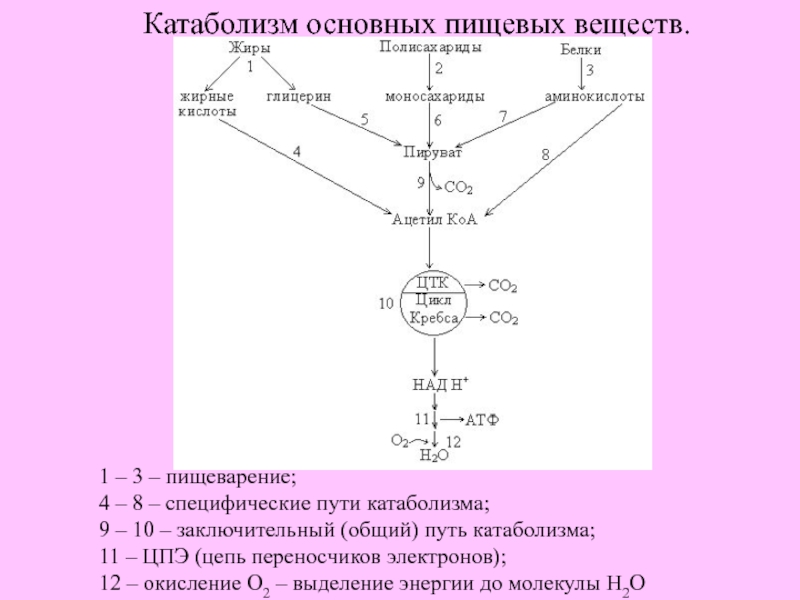 Они обе приводят к расходу калорий, вот только происходит это разными способами.
Они обе приводят к расходу калорий, вот только происходит это разными способами.
Немного о питании и диетах Для того чтобы ускорить свой метаболизм необходимо изменить подход к своему питанию. А это значит, что необходимо убрать определённые продукты из своего рациона, которые просто делают «помойку» из вашего организма и начать есть больше. Не нужно искать какую-то особую, строгую и убийственную диету. Достаточно начать питаться дробно (небольшими порциями) и часто (5-6 приёмов пищи в день). Не забываем при этом подсчитывать калории. Ни в коем случае нельзя допускать голода для организма.
Важно! Для восстановление обмена веществ, ограничьте сладкие и жирные продукты, полуфабрикаты в первую очередь. Более подробно о том, отчего следует по возможности избавиться в своём рационе, вы можете прочитать в нашей статье или.
Несколько советов по ускорению метаболизма.
- Чистая вода. Пейте больше простой и чистой воды. Благодаря этому вы сможете контролировать свой аппетит и включать накопленные жировые отложения в обменные процессы.

- Стабильный сон. Очень важно для отличного обмена веществ спать 7-8 часов в сутки. Желательно, чтобы сон был в период с 22:00-23:00 до 05:00-06:00. Самое оптимальное время, для выработки, например, мелатонина (гормон жизни и долголетия).
- Стрессы. Постарайтесь максимально избавиться от стрессов. Стресс, влияет на наш организм не очень положительно изменяя наш гормональный фон. При сильном стрессе вырабатывается кортизол, который разрушающие действует на вашу мышечную ткань.
- Контрастный душ — очень хороший помощник в ускорении вашего метаболизма.
- Спорт. Займитесь своим телом. Измените свой образ жизни. Просто начните заниматься ходьбой, бегом, делать базовые упражнения (отжимания, приседания, подтягивания) обязательно начните делать растяжку или займитесь йогой. Если вас это все бесит, начните хотя бы ежедневно танцевать.
- Ешьте фрукты богатые витамином С и овощи богатые клетчаткой. Все фрукты и цитрусовые, содержащие в больших количествах витамин С, в частности, это касается ананасов, кислых ягод, киви, зелёных яблок, способствуют улучшению пищеварительного процесса, помогают сжигать жиры и ускорять метаболизм.
 А богатые клетчаткой овощи будут помогать лучше работать кишечнику и желудку. Быстро наполняя желудок, они на продолжительное время дают ощущение сытости.
А богатые клетчаткой овощи будут помогать лучше работать кишечнику и желудку. Быстро наполняя желудок, они на продолжительное время дают ощущение сытости. - И наконец для ленивых , Health partner предлагает программы : Health, Детокс, Антистресс, Релакс, которые не только нормализуют метаболизм , но и помогают адекватно привести в порядок физическое и психическое здоровье.
Impact Oral | Nestlé Health Science
Вода, сахароза, мальтодектрин, молочный белок, L- аргинин, частично гидролизованная гуаровая камедь, рыбий жир, минеральные вещества (цитрат калия, цитрат натрия, натрия хлорид, хлорид магния, хлорид калия, магния фосфат, кальция цитрат, железа пирофосфат, сульфат цинка, меди глюконат, сульфат марганца, натрия фторид, йодид калия, молибдат натрия, натрия селенат, хрома хлорид), эмульгаторы (Е471, соевый лецитин), эйкозапентаеновая кислота, битартрат холина, ароматизаторы, витамины (С, ниацин, пантотеновая кислота, Е, В6, А, тиамин, рибофлавин, D, фолиевая кислота, К, биотин, В12), стабилизаторы (Е460, каррагинан, Е466), антиоксидант (Е306), краситель (бета-каротин).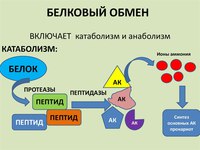 Не содержит глютен и лактозу.
Не содержит глютен и лактозу.
Пищевая ценность на 100 мл готовой смеси
Калорийность / Энергетическая ценность
144 ккал / 606 кДж
Основные компоненты
Пищевые волокна
1,4 г
Расчетное количество соли
0,38 г
Минералы
Марганец
0,3 мкг
Молибден
22,5 мкг
Фториды
0,21 мкг
Витамины
Фолиевая кислота
28 мкг
Пантотеновая кислота
1,14 мг
Другие питательные вещества
Эйкозапентаеновая кислота (EPA) +
Докозагексаеновая
кислота (DHA)
0,5 г
Среднецепочечные
триглицериды
1,1 г
Нуклеотиды
0,18 г
Лабораторная диагностика сахарного диабета в Санкт-Петербурге
Диабет — эндокринное заболевание, комплекс патологических состояний, в основе которых лежит патология синтеза и функции инсулина, а также происходит нарушение обмена глюкозы. В результате у пациента возникает стойкая гипергликемия — высокая концентрация глюкозы в крови. Патология относится к хроническим заболеваниям. Постепенно, кроме углеводного патология затрагивает остальные виды обменных процессов — жировой, белковый, минеральный. Клиническая картина диабета разнообразна, ведь он затрагивает разные органы и системы. Последствия диабета сказываются на состоянии кожи, периферических нервов, головного мозга, почек, сердца и сосудов. Каждое из нарушений влечет за собой соответствующие последствия и в результате диабет представляет собой сочетание нескольких патологий.
Лабораторная диагностика диабета важна для постановки диагноза, контроля лечения, определения степени и стадии процесса. Различные особенности гипергликемии являются показателями для той или иной лечебной тактики.
Как распознать заболевание?
Симптомы диабета развиваются в зависимости от стадии процесса. Заболевание может постепенно развиваться или дебютировать в виде острого состояния — комы. Чтобы упорядочить признаки диабета, их делят на две категории: основные и дополнительные.
Рассмотрим основные признаки диабета:
Частое мочеиспускание, увеличение объема выделяемой жидкости. Это связано с тем, что высокий уровень глюкозы повышает осмотическое давление, что сказывается на работе почечных клубочков и канальцев. Развивается глюкозурия — в моче становится много глюкозы, хотя в норме её там нет вовсе. Пациенты отмечают учащение ночных позывов к мочеиспусканию.
При диабете наблюдается сильная жажда, которая не связана с физическими нагрузками или погодными условиями. Если в привычном режиме жизни пациент отмечает сильную жажду и частое желание употребления воды — стоит задуматься о том, чтобы сдать анализ крови на глюкозу. Такой клинический признак связан с повышенной потерей жидкости.
Человеку постоянно хочется есть. Этот голод так же как и жажда не связан с окружающими условиями или повышенными нагрузками. Причина заключается в том, что больше расходуются питательные вещества и нарушается их обмен в тканях. Глюкоза находится в крови, но не попадает в клетки. Это воздействует на рецепторы, организм воспринимает ситуацию, как недостаток глюкозы и повышает аппетит, чтобы восполнить запас.
Так как обмен глюкозы нарушается, усиливается катаболизм жиров и белков. Это приводит к уменьшению объема и массы тела. Похудение происходит при повышенном аппетите, чем еще больше его усиливает.
Это характерная картина для 1 типа диабета. В данном случае патология может начинаться остро, симптомы развиваются быстро и пациент помнит примерное время начала заболевания.
Менее специфические, дополнительные симптомы характерны для всех видов диабета. Они возникают постепенно, пациент не может точно вспомнить, как давно они его беспокоят. Могут наблюдаться такие изменения:
- кожа и слизистые оболочки постоянно чешутся и зудят;
- появляется неприятная сухость слизистой оболочки рта;
- чувствуется слабость мышц;
- постоянная головная боль;
- частые и длительные воспаления кожи, в том числе гнойного характера, которые не заживают даже на фоне лечения;
- зрительные нарушения;
- появление ацетона в моче.
Могут быть и другие общие симптомы, такие как слабость и повышенная утомляемость. Человеку становится трудно работать или учиться, снижается работоспособность, нарушается сон.
Перейти к анализам
Современная диагностическая программа при патологии
Различают следующие методы диагностики при нарушениях обмена глюкозы:
Рассмотрим более детально данные методики и их особенности.
Методика показана при патологии печени, поджелудочной железы, ожирении, для подтверждения или исключения диабета, а также для контроля его лечения.
Кровь для анализа собирается утром, до еды и не ранее чем через 8 часов после последнего приема пищи. Пациент не должен подвергаться физическим или эмоциональным нагрузкам с вечера до исследования.
2) Тест на толерантность к глюкозе
Исследование показано тогда, когда концентрация глюкозы в крови не выходит за критические рамки. Если измерение уровня глюкозы показывает результат от 3,88 до 7 ммоль/л, рано говорить о гипер- или гипогликемии и необходимо уточнить состояние пациента. Тест применяется также для тех, у кого есть факторы риска развития диабета.
Исследование проводится утром, до еды и не раньше, чем через 8 часов после последнего приема пищи. Пациент должен находится в спокойном состоянии, не подвергаться физическим нагрузкам и стрессам. Тест не проводится, если уровень глюкозы в крови превышает 7 ммоль/л.
Как проходит тест? Вначале глюкометром измеряется уровень гликемии, после чего происходит забор крови. Далее пациента просят выпить 300 мл воды с растворенными в ней 75 г глюкозы. Через 2 часа происходит повторный забор крови. Вс это время нельзя пить и есть, курить и выполнять физические нагрузки.
Расшифровка теста происходит следующим образом: позитивный результат — это концентрация глюкозы в крови менее чем 7,8 ммоль/л, от 7,8 до 11,0 — толерантность считается нарушенной, если гликемия выше 11,0 — ставится диагноз “Сахарный диабет”.
3) Гликированный гемоглобин — показатель, который позволяет отличить кратковременную гипергликемию от длительного течения патологического обмена глюкозы. Принцип исследования построен на обнаружении и измерении того количества гемоглобина, который связался с молекулами глюкозы необратимо. Методика позволяет оценить картину за последние 3 месяца, так как именно столько времени в крови сохраняется эритроцит с измененным гемоглобином. Оценка важна для прогнозирования осложнений диабета и постановки диагноза. Также, анализ показывает, насколько качественным было лечение болезни за прошедший период и нужна ли коррекция.
Нормальная концентрация гликированного гемоглобина от 4 до 5,9%. Если значение превышает 8% — срочно требуется коррекция лечебной тактики.
Особенной подготовки перед анализом не требуется. Результаты не зависят от приема пищи, нагрузок и времени суток.
Исследование показано при диагностике диабета, для контроля динамики процесса.
4) Исследование мочи при диабете включает в себя анализ на уровень ацетона и глюкозы. Применяется оценка суточного количества мочи, чтобы определить общую глюкозурию за сутки. В норме — глюкозы в моче быть не должно. Иногда анализ разбивают на временные промежутки так, чтобы получилось четыре порции мочи. Это позволяет определить суточные колебания и выяснить их причину. Однократная сдача мочи — стандартная процедура для определения глюкозурии или её исключения.
Оценка уровня ацетона происходит аналогично и выполняется одновременно с определением глюкозы. Ацетон в норме также отсутствует в моче, его появление говорит о патологии.
Методика применяется для контроля качества лечения. Применяется также при плохом самочувствии, общих расстройствах здоровья, во время болезни.
5) Определение уровня инсулина необходимо при длительной гипергликемии. Гормон повышается в ответ на высокую концентрацию глюкозы — это естественно и физиологично. А врачу удается оценить особенности глюкозо-инсулинового обмена, благодаря данной методике. Для анализа используется венозная кровь. Желательно сочетать данный тест вместе с тестом на толерантность глюкозы.
Исследование проводится натощак и после 8-часового перерыва после приема пищи. Следует избегать интенсивных нагрузок. За сутки следует прекратить прием еды с высоким содержанием сахара. Нельзя курить в течении 2 часов до анализа.
6) Анализ на С-пептид входит в диагностику диабета, так как это продукт углеводного обмена и показывает его качество в организме.Как правило, уровень данного вещества соответствует концентрации инсулина в крови. Устойчивость к инсулину и патология островков поджелудочной железы сопровождается снижением С-пептида. Нормальный показатель — 0,9-7,9. Повышенная концентрация говорит о наличии сахарного диабета типа 2, реакции на препараты, снижающие уровень глюкозы, могут говорить о наличии инсулиномы, антител к инсулину, нейроэндокринных процессах и проблемах с почками.
Подготовка к анализу подразумевает отказ от интенсивных нагрузок, физических упражнений, стрессов. Необходимо сдавать кровь натощак, отказаться перед сдачей не только от еды, но и от курения.
7) Анализ на лептин также входит в комплексную программу оценки здоровья при диабете. Это гормон, который регулирует аппетит и массу тела. Его вырабатывают адипоциты — жировые клетки. Норма — 1,1 — 27,6 нг/мл для мужчин и 0,5-13,5 нг/мл для женщин.
Подготовка к анализу включает в себя отказ от пищи за 8 часов до сдачи крови, разрешается небольшое количество воды. Пациенту следует избегать нагрузок и переживаний.
Исследование показано, если у пациента подозревают дефицит данного гормона, нарушение репродуктивной функции, если необходимо дифференцировать диабет или ожирение, а также при частых тромбозах. Значение может повышаться при повышенном потреблении пищи, сахарном диабете, который не зависит от уровня инсулина, а также при ожирении. Сниженный результат может сопровождать стремительное похудение, длительное голодание и генетический дефицит гормона.
8) Анализ антител к клеткам поджелудочной железы применяется для диагностики аутоиммунных нарушений углеводного обмена. Это показатель деструкции клеток железы, которая имеет аутоиммунный механизм. Применяется для коррекции терапии, дифференциальной диагностики, определения типа диабета в сложных случаях и при непонятной клинике.
В норме антител к клеткам поджелудочной а организме быть не должно. Их наличие говорит о неисправности иммунной системы.
Анализ крови сдается натощак, необходимо избегать физических и эмоциональных нагрузок перед обследованием. Помимо еды стоит отказаться от курения, можно пить небольшое количество воды без газа.
Все анализы должны проводиться в соответствующих условиях, с соблюдением асептики и антисептики. Собранный биологический материал необходимо правильно хранить и транспортировать в лабораторию.
Назначение тех или иных методов исследования зависит от стадии процесса, рекомендаций врача. Желательно комбинировать различные анализы, чтобы диагностика была полноценной и охватывала разные аспекты углеводного обмена. Качественная диагностика диабета позволяет вовремя начать лечение, которое соответствует состоянию организма. Если стабилизировать уровень глюкозы в крови и придерживаться её нормальной концентрации, можно избежать целого ряда осложнений. В этом и заключается ценность диагностических процедур при нарушениях глюкозного обмена.
Биохимия, катаболизм белков — StatPearls
Введение
Катаболизм белков — это расщепление белков на абсорбируемые мономеры для дальнейшей деградации или повторной сборки. Катаболизм белков в просвете кишечника важен по нескольким причинам, одна из которых — мобилизация незаменимых аминокислот для всасывания. Незаменимые аминокислоты не могут быть синтезированы в организме человека, но они необходимы для биосинтеза жизненно важных белков, поэтому их единственный источник — распад полипептидов с помощью пищеварительных ферментов.Этот процесс начинается в желудке и продолжается в тонком кишечнике. Большие белковые цепи разбираются, чтобы в конечном итоге оставить свободные аминокислоты, которые могут попасть в кровь и транспортироваться к различным клеткам по всему телу для дальнейшего расщепления.
Эндопептидазы в форме зимогена высвобождаются слизистой оболочкой желудка и экзокринной поджелудочной железой для расщепления полипептидной цепи между определенными аминокислотными остатками. Оказавшись в меньшей форме, экзопептидазы по одной удаляют последние аминокислоты с C- или N-конца дипептида или трипептида, способствуя абсорбции на микроворсинках.Клетки могут использовать эти аминокислоты для создания жизненно важных белков или в качестве субстратов для создания энергии. Белки, созданные внутриклеточно, также могут катаболизироваться по тем же причинам. Внутриклеточные белки, которые были неправильно свернуты или больше не функционируют в клетке, также подвергаются внутриклеточному катаболизму белков в лизосомах с помощью образования убиквитина и протеасом. Если клетка находится в низкоэнергетическом состоянии, свободные аминокислоты в цитозоле далее разлагаются с образованием промежуточных продуктов цикла лимонной кислоты и направляются туда для производства АТФ.В то время как углеродный каркас вступает в энергетические пути, азотный каркас модифицируется и выводится в основном через почки.
Основы
rКатаболический процесс начинается, когда протеолитические ферменты высвобождаются в форме зимогена из слизистой оболочки желудочно-кишечного тракта. После секреции зимогены расщепляются на активные протеазы. Пепсин из слизистой оболочки желудка гидролизует более крупный полипептид на более мелкие полипептиды, которые позже могут быть расщеплены протеазами поджелудочной железы, высвобождаемыми в двенадцатиперстной кишке.Трипсин, химотрипсин, эластаза и карбоксипептидазы, синтезируемые экзокринной поджелудочной железой, могут расщеплять полипептид в определенных точках расщепления на аминокислоты и олигопептиды. Аминопептидазы, расположенные на щеточной кайме эпителиальных клеток кишечника, гидролизуют оставшиеся олигопептиды до аминокислот, которые затем попадают в кровоток через трансмембранные транспортные системы. Существуют как облегченные, так и активные транспортные механизмы для транспортировки аминокислот в клетки.
Катаболизм белков — важная часть клеточного обновления.Когда цитозольные белки, такие как сигнальные или структурные пептиды, больше не нужны, они должны расщепляться в лизосомах для создания новых белков, которые могут выполнять необходимые метаболические функции. Если полученные аминокислоты не используются для синтеза новых белков для жизненно важных внутриклеточных функций, они могут войти в цикл лимонной кислоты для выработки энергии. Когда запрограммированная смерть — это судьба клетки, катепсины и другие катаболические ферменты работают вместе в процессе апоптоза.
Пептидазы желудка: главные клетки слизистой оболочки желудка секретируют пепсиноген.[1] С помощью соляной кислоты, высвобождаемой париетальными клетками желудка, пепсиноген изменяет конформацию в сильнокислой среде желудка и расщепляется на активную протеазу, пепсин. Кислота также денатурирует пищевые белки и частично разворачивает их для облегчения протеолиза. Пепсин обычно расщепляет пептидные связи, содержащие карбоксильную группу, от кислой или ароматической аминокислоты; однако его широкая специфичность позволяет ему расщеплять пептидные связи и в других местах. [2] Меньшие пептиды, а иногда и свободные аминокислоты, затем попадают в двенадцатиперстную кишку, чтобы встретить пептидазы, секретируемые поджелудочной железой.
Пептидазы поджелудочной железы и кишечника: экзокринная поджелудочная железа секретирует различные пищеварительные ферменты, включая амилазу, липазу, колипазу, бикарбонат и неактивные протеазы. Бикарбонат, высвобождаемый в просвет кишечника, повышает pH и позволяет протеазам поджелудочной железы активироваться из их зимогенной формы. Одна конкретная протеаза, трипсин, является наиболее важной для процесса пищеварения, потому что она сама расщепляет белки, а также активирует другие протеазы из их неактивных форм.Трипсин отщепляется от его проферментной формы трипсиногена энтеропептидазой. Затем трипсин продолжает активировать химотрипсиноген, проэластазу и прокарбоксипептидазы в химотрипсин, эластазу и карбоксипептидазы соответственно [3]. Трипсин гидролизует пептидные связи с карбоксильной группой аргинина или лизина. Химотрипсин, с другой стороны, имеет тенденцию воздействовать на пептидные связи между гидрофобными аминокислотами. [4] Трипсин и химотрипсин относятся к сериновым протеазам и имеют аналогичный метод протеолиза.Они образуют каталитический тетраэдрический промежуточный продукт, располагая каталитическую триаду таким образом, что протоны, образующие водородные связи, могут разорвать пептидную связь между двумя аминокислотами белка-субстрата. [4] Эти два фермента также очень похожи в своем S1-связывающем кармане, который содержит основную аминокислоту в трипсине и гидрофобную аминокислоту в химотрипсине. [4] Тем не менее, особенности и механизмы намного сложнее с их динамическими свойствами, а структурная гибкость является одним из наиболее важных.
Эластаза, также сериновая протеаза, расщепляет эластин, а также белки с небольшими гидрофобными боковыми цепями, такие как глицин или аланин. Ацинарные клетки поджелудочной железы выделяют две формы эластазы, причем эластаза II имеет более широкую специфичность, чем эластаза I. [5] Нуклеофильная аминокислота в активном центре эндопептидазы гидролизует пептидную связь до более мелких пептидных цепей, которые нуждаются в дальнейшей деградации; это может происходить через экзопептидазы, которые удаляют одну аминокислоту с конца белковой цепи.Примером экзопептидазы является активируемая трипсином протеаза поджелудочной железы карбоксипептидаза. Существует две формы этого протеолитического фермента, A и B. Карбоксипептидаза A имеет тенденцию отщеплять гидрофобные аминокислоты от пептидной цепи, а карбоксипептидаза B отщепляет основные аминокислоты от пептида. Эпителиальные клетки кишечника также содержат экзопептидазы на щеточной кайме, называемые аминопептидазами, которые выделяют отдельные аминокислоты, которые переносятся в клетки [6].
Внутриклеточный обмен белков: белки внутри клетки также подвергаются катаболизму, чтобы пополнить пул внутриклеточных аминокислот.Хотя у всех белков есть период полураспада, некоторые из них разлагаются в течение нескольких минут, а некоторые — в течение часов или дней. Некоторыми примерами сильно синтезируемых и деградированных белков являются белки мышечных клеток, гемоглобин, эпителиальные клетки желудочно-кишечного тракта и сами протеолитические ферменты. Многие из этих белков расщепляются лизосомными протеазами, так что их аминокислотные составляющие могут использоваться для восстановления белков, потребности которых зависят от текущего физиологического статуса клетки. Например, в периоды голодания аминокислоты, высвобождаемые в результате катаболизма мышечных белков, могут попасть в глюконеогенез и действовать как источник энергии.В сытом состоянии или во время метаболического ацидоза глутамин может быть топливом для почечной системы. Скелетные мышцы могут абсорбировать аланин, аспартат и глутамат и посредством трансаминирования образовывать полезные промежуточные соединения цикла Кребса.
Клеточный
Трансмембранный транспорт: Работа эндопептидаз, химотрипсина, эластазы и карбоксипептидазы и экзопептидаз, карбоксипептидазы и аминопептидазы в конечном итоге дает свободные аминокислоты. Существуют механизмы облегченной диффузии и активного транспорта для поглощения этих соединений из просвета кишечника посредством вторичного активного транспорта [7].Аминокислоты пересекают щеточную границу кишечных клеток через натрий-зависимую транспортную систему, в которой низкая внутриклеточная концентрация натрия способствует совместному поступлению натрия и аминокислоты. Натрий-калиевая АТФаза на базолатеральной мембране поддерживает низкую концентрацию натрия внутри клетки. Как только аминокислоты попадают в клетку на апикальной мембране, они транспортируются вниз по своему градиенту на базальной мембране в кровоток, обычно в портальную циркуляцию. Шесть различных натрийзависимых аминокислотных переносчиков частично совпадают по своей специфичности в отношении аминокислот, которые они транспортируют через апикальную мембрану щеточной каймы; однако носители отдают предпочтение типам транспортируемых аминокислот.[7] Предпочтения включают нейтральные аминокислоты, кислые аминокислоты, основные аминокислоты, цистин, пролин и гидроксипролин. Натрий-зависимые аминокислотные переносчики на клетках просвета кишечника также существуют на почечном эпителии в различных формах изоферментов [8]. С другой стороны, облегченные транспортные системы на базальной стороне сопоставимы с системами, обнаруженными в других типах тканей. Аминокислоты также могут абсорбироваться из крови через эти облегченные переносчики и использоваться в качестве субстратов для цикла лимонной кислоты для выработки энергии — двунаправленность облегченных переносчиков помогает выживанию кишечного эпителия во время голодания.
Транспортеры на апикальной поверхности мышц, печени и других тканей поглощают аминокислоты из крови и концентрируют их. Хотя эти переносчики сходны с люминальными натрий-зависимыми переносчиками аминокислот на кишечном эпителии, они различаются по своим генетическим основам, белковому составу и специфичности [8]. Многие ткани экспрессируют A-систему, ASC-систему и L-систему для транспорта аминокислот в клетку; однако из них L-система — единственная без натрий-зависимого механизма.Клетки кишечника и почек экспрессируют B-систему, X-систему и иминосистему. [9] Конкретный тип ткани может содержать конкретный тип переносчика аминокислот, не обнаруживаемый в других тканях, например, экспрессия N-системы в печени и ее специфичность в отношении абсорбции глутамина. Изоформа N-системы для поглощения глутамина может экспрессироваться в другом типе ткани. Однако его свойства и состав будут отличаться от того, что содержится в печени. Каждая система отличается своей специфичностью в отношении аминокислот и имеет значение для множества различных патологий.
Лизосомная деградация: если белки больше не нужны или синтезируются неправильно, внутриклеточные белки расщепляются с использованием лизосом и протеасомного комплекса. Белки попадают в лизосомы посредством аутофагии, строго регулируемого процесса, при котором везикулы сливаются с лизосомной мембраной. [10] Аутофагии могут способствовать состояния низкой энергии, такие как низкий уровень АТФ или высокий уровень АМФ в клетке. Поскольку АМФ-активированная протеинкиназа активируется при высоких уровнях АМФ, она может фосфорилировать комплекс 1 и 2 туберозного склероза, что приводит к активации Rheb-GTPase.[11] Эта активация важна, потому что она инактивирует Rheb и, таким образом, также инактивирует mTOR, главный ингибитор аутофагии; вот как низкоэнергетические состояния способствуют катаболизму белков. Напротив, активация комплекса 1 и 2 туберозного склероза инсулином через киназу Akt приводит к активному mTOR. [11] Активированный mTOR способствует синтезу белка, а не деградации.
Убиквитинирование — это процесс, при котором молекула убиквитина становится ковалентно прикрепленной к расщепляемым белкам и взаимодействует с протеасомой в лизосоме для расщепления указанного белка с использованием АТФ-зависимого процесса.[12] Убиквитин сначала должен активироваться через E1, E2 и E3, прежде чем он ковалентно присоединится к эпсилон-аминогруппе боковых цепей лизина через ферментный комплекс. [13] Обычно белок получает больше молекул убиквитина на конце лизина, образуя хвост. После формирования хвоста убиквитинированный белок отделяется от комплекса лигирующих ферментов и перемещается в протеасому. Протеасома представляет собой цилиндрический комплекс протеазы 26S с несколькими каталитическими центрами внутри. Протеасомный комплекс 26S имеет два альфа- и два бета-кольца, содержащих три внутренних каталитических сайта, подобных трипсину, химотрипсину и постглутамилпептидилгидролазе.Конструкция четырехкольцевой структуры такова, что альфа-кольца играют стабилизирующую роль, тогда как бета-субъединицы являются активными каталитическими частями комплекса. [13] Он сохраняет молекулы убиквитина, но использует гидролиз АТФ, чтобы развернуть и продвинуть белок глубже в комплекс и высвободить свободные аминокислоты. [10] Белки, гидролизуемые этим процессом, известны как последовательности PEST, названные так в честь общих аминокислот, включенных в цепь. Аминокислоты пролин, глутамат, серин и треонин находятся в высоких концентрациях в белках последовательности PEST.[14]
Попав внутрь лизосомы, лизосомальные протеазы, называемые катепсинами, также могут разрушать цитозольные белки, помеченные как катаболизм. Катепсины — это еще один тип цистеиновой протеазы, присутствующий в перицеллюлярной среде, ядрах цитозоля и митохондриях. [15] Передача сигналов фактора некроза опухоли, клеточный стресс и каспаза могут запускать лизосомное высвобождение катепсина, расщепляя внутриклеточные белки, такие как Bid, что приводит к последующей активации каскада апоптотических каспаз. [16]
Молекулярный
Судьба пула внутриклеточных аминокислот сильно зависит от метаболического состояния отдельных тканей и организма в целом.В физиологическом состоянии сытости аминокислоты могут разлагаться дальше до их углеродного и азотного скелетов. В то время как углеродная основа может подвергаться окислению для получения энергии, азотный компонент выводится из организма через азотсодержащие отходы. Первоначально азот присутствует в форме аммиака, токсичного для человеческого организма вещества. Печень превращает токсичный аммиак в мочевину через цикл мочевины в нетоксичную водорастворимую форму азота, которая легко выводится через почки.
По мере того, как азотный скелет выбрасывается, углеродный скелет аминокислот может использоваться для производства энергии путем окисления в пируват (триптофан, глицин, аланин, серин, цистеин) и ацетил-КоА. Пируват и ацетилкофермент А являются субстратами цикла лимонной кислоты, в конечном итоге производящим АТФ и диоксид углерода. Ацетил-КоА может также образовывать кетоновые тела, попадающие в кровоток для использования в качестве источника энергии в определенных тканях. Аминокислоты, которые становятся ацетил-КоА или ацетоацетатом, называются кетогенными из-за их склонности превращаться в кетоновые тела.[17] Например, лизин и лейцин являются кетогенными, а также незаменимыми аминокислотами.
Другая возможная судьба — превращение углеродного скелета в промежуточные продукты цикла лимонной кислоты альфа-кетоглутарат (аргинин, гистидин, глутамин, пролин, глутамат), сукцинил-КоА (валин, треонин, изолейцин, метионин), фумарат (аспартат, тирозин, фенилаланин) и оксалоацетат (аспартат, аспарагин). [18] Стоит отметить, что малат, промежуточный продукт цикла лимонной кислоты, может перемещаться по кровообращению в печень и участвовать в глюконеогенезе.[19] Если аминокислота может стать предшественником глюкозы, она называется глюкогенной. Из-за различных промежуточных продуктов и точек входа в цикл Кребса аминокислоты могут быть как глюкогенными, так и кетогенными.
Тестирование
Убиквитинирование — начало внутриклеточного катаболизма белков. Тестирование функции этого процесса для понимания конкретных патологий включает комбинирование культивируемых клеток с плазмидами, содержащими белок с убиквитином, меченным эпитопом. Следующие шаги включают добавление стабилизирующих буферов, лизирование и обработку клеток ультразвуком с последующими циклами инкубации.Поскольку окончательная форма анализа включает иммуноблоттинг с гелем SDS-PAGE, должны быть сформированы конъюгированные антитела против белка [20]. Существуют также способы анализа процесса убиквитинирования in vitro, а не in vivo. [20]
Существуют способы проверки ферментов, выделяемых экзокринной поджелудочной железой, трипсина, химотрипсина и эластазы. Одним из тестов является тест на эластазу кала, при котором стул анализируется на концентрацию эластазы, что подтверждает проходимость протока поджелудочной железы, секреторную функцию экзокринной функции поджелудочной железы и наличие активирующего трипсина.[21] Этот тест является наиболее широко используемым из-за его высокой специфичности и неинвазивности. Более инвазивный тест — это эндоскопический тест функции поджелудочной железы, который доступен только в некоторых медицинских учреждениях. В этом тесте пациенту вводят успокоительное и перед эндоскопической процедурой вводят секретин [22]. Эндоскоп отсасывает другую жидкость из верхних отделов желудочно-кишечного тракта по пути к двенадцатиперстной кишке. Затем образцы секрета поджелудочной железы берутся на большом сосочке и анализируются на бикарбонат в лаборатории.[22] Как правило, образцы собираются с интервалами в 15, 30, 45 и 60 минут. Оценка секреции и активности этих ферментов важна для оценки патологий мальабсорбции белка.
Патофизиология
Муковисцидоз — аутосомно-рецессивная мутация в гене CFTR, который кодирует белки, составляющие поры хлоридного канала. [23] Тяжесть заболевания варьируется, но некоторые особенности являются общими для всех форм, например, образование толстых слизистых пробок в протоке поджелудочной железы, легких и мужской мочеполовой системе.Поскольку белки должны разрушаться для всасывания, слизистая пробка, блокирующая высвобождение зимогена из экзокринной поджелудочной железы, приведет к отсутствию катаболизма жизненно важных белков в просвете кишечника. [23] Пациенты с муковисцидозом страдают серьезным дефицитом белка и должны получать добавку экзогенных ферментов поджелудочной железы, хотя следует упомянуть, что побочные эффекты длительного приема добавок ферментов поджелудочной железы в настоящее время неизвестны. [24]
Квашиоркор — еще один серьезный дефицит белка; однако это связано с недостаточным потреблением белка, а не с генетическим заболеванием.Пищеварительные ферменты присутствуют в квашиоркоре; однако из-за того, что потребляется мало белка, присутствуют симптомы дефицита белка. У людей с истощением, принимающих квашиоркор, жизненные реакции в организме поддерживаются за счет внутриклеточного катаболизма белков. Потребность в доступных незаменимых аминокислотах приводит к значительному снижению периферической мышечной массы из-за разрушения мышц. [10] Дефицит белка также приводит к снижению уровня альбумина в сыворотке крови, что снижает внутрисосудистое коллоидное давление, что приводит к отеку и вздутию живота.Тяжелая форма квашиоркора быстро ухудшается, поскольку пищеварительные ферменты больше не вырабатываются, а эпителий тонкого кишечника не регенерируется.
Нарушение механизмов облегченного и активного транспорта может привести к патологическим состояниям мальабсорбции. Цистинурия и болезнь Хартнупа являются генетическими нарушениями, связанными с мембранными переносчиками аминокислот, но различаются группами транспортируемых аминокислот и поэтому проявляются по-разному клинически [9]. Цистинурия — это нарушение транспорта основных аминокислот через мембраны почечной и желудочно-кишечной систем.Отличительным признаком этого заболевания является образование почечных камней из-за неспособности резорбировать основную аминокислоту цистин из клубочкового фильтрата. Поскольку переносчик также существует в эпителии тонкого кишечника, цистин и другие основные аминокислоты плохо всасываются из просвета кишечника. Болезнь Хартнупа — заболевание, похожее по механизму, представляет собой дефект в транспортировке нейтральных аминокислот через почечную и кишечную систему. Поскольку триптофан является одной из нейтральных аминокислот, плохо усваиваемых, при физикальном осмотре выявляются сыпь, диарея и психические расстройства, похожие на пеллагру.В отличие от пеллагры, добавление ниацина мало помогает избавиться от симптомов и должно выявить необходимость генетического тестирования.
Клиническая значимость
Правильное функционирование катаболизма белков имеет первостепенное значение для поддержания метаболических потребностей человеческого организма. Разрушение больших полипептидных цепей с высвобождением свободных незаменимых и заменимых аминокислот обеспечивает клетки необходимыми субстратами для синтеза белка или выработки энергии. Распад внутриклеточного белка важен для нормального обмена веществ, а также при недоедании и истощенных физиологических состояниях, чтобы мобилизовать аминокислоты на экспорт для поддержки энергетических потребностей жизненно важных органов.Нарушения катаболизма белков могут привести к разнообразным клиническим проявлениям, затрагивающим почти все системы органов.
Ссылки
- 1.
- Magee DF. Пепсин. Med Clin North Am. 1974 ноя; 58 (6): 1277-87. [PubMed: 4610293]
- 2.
- Fruton JS. Специфика и механизм действия пепсина. Adv Enzymol Relat Areas Mol Biol. 1970; 33: 401-43. [PubMed: 4916858]
- 3.
- Джи Б, Logsdon CD. Усвоение новой информации о роли трипсина при панкреатите.Гастроэнтерология. 2011 декабрь; 141 (6): 1972-5. [Бесплатная статья PMC: PMC4327863] [PubMed: 22033179]
- 4.
- Ma W, Tang C, Lai L. Специфичность трипсина и химотрипсина: определяемая динамическая корреляция, контролируемая движением петли. Biophys J. 2005 Aug; 89 (2): 1183-93. [Бесплатная статья PMC: PMC1366603] [PubMed: 15923233]
- 5.
- Del Mar EG, Largman C, Brodrick JW, Fassett M, Geokas MC. Субстратная специфичность эластазы поджелудочной железы человека 2. Биохимия. 1980, 5 февраля; 19 (3): 468-72.[PubMed: 6898442]
- 6.
- Tobey N, Heizer W., Yeh R, Huang T.I, Hoffner C. Пептидазы щеточной каймы кишечника человека. Гастроэнтерология. 1985 Апрель; 88 (4): 913-26. [PubMed: 3882510]
- 7.
- Холмс Р., Лобли Р. В.. Повторный визит к краю кишечной щетки. Кишечник. 1989 декабрь; 30 (12): 1667-78. [Бесплатная статья PMC: PMC1434444] [PubMed: 2693228]
- 8.
- Bröer S. Транспорт аминокислот через эпителий кишечника и почек млекопитающих. Physiol Rev.2008 Янв; 88 (1): 249-86.[PubMed: 18195088]
- 9.
- Bröer S, Fairweather SJ. Транспорт аминокислот через кишечник млекопитающих. Compr Physiol. 2018 13 декабря; 9 (1): 343-373. [PubMed: 30549024]
- 10.
- Сандри М. Распад белка при истощении мышц: роль аутофагии-лизосомы и убиквитин-протеасомы. Int J Biochem Cell Biol. 2013 Октябрь; 45 (10): 2121-9. [Бесплатная статья PMC: PMC3775123] [PubMed: 23665154]
- 11.
- Пуэртоллано Р. mTOR и регуляция лизосом. F1000Prime Rep.2014; 6: 52. [Бесплатная статья PMC: PMC4108950] [PubMed: 25184042]
- 12.
- Доэрти Ф.Дж., Доусон С., Майер Р.Дж.. Убиквитин-протеасомный путь внутриклеточного протеолиза. Очерки Биохимии. 2002; 38: 51-63. [PubMed: 12463161]
- 13.
- Цехановер А. Путь убиквитин-протеасома: гибель белка и жизнь клетки. EMBO J. 15 декабря 1998; 17 (24): 7151-60. [Бесплатная статья PMC: PMC1171061] [PubMed: 9857172]
- 14.
- Rechsteiner M, Rogers SW. Последовательности PEST и регуляция протеолизом.Trends Biochem Sci. 1996 июл; 21 (7): 267-71. [PubMed: 8755249]
- 15.
- Брикс К., Дункхорст А., Майер К., Джорданс С. Катепсины цистеина: клеточная дорожная карта для различных функций. Биохимия. 2008 Февраль; 90 (2): 194-207. [PubMed: 17825974]
- 16.
- Лейст М., Яаттеля М. Запуск апоптоза катепсинами. Смерть клетки отличается. 2001 апр; 8 (4): 324-6. [PubMed: 11550083]
- 17.
- Харрис Р.А., Джоши М., Чжунг Н.Х., Обаяши М. Обзор молекулярных и биохимических основ катаболизма аминокислот с разветвленной цепью.J Nutr. 2005 июн; 135 (6 доп.): 1527S-30S. [PubMed: 15930464]
- 18.
- БЕЛЛАМИ Д. Эндогенные промежуточные продукты цикла лимонной кислоты и аминокислоты митохондрий. Biochem J. 1962, январь; 82: 218-24. [Бесплатная статья PMC: PMC1243435] [PubMed: 13866863]
- 19.
- Айкава Т., Мацутака Х., Ямамото Х., Окуда Т., Исикава Э. Глюконеогенез и метаболизм аминокислот. II. Межорганические отношения и роль глутамина и аланина в метаболизме аминокислот голодных крыс. J Biochem.1973 Ноябрь; 74 (5): 1003-17. [PubMed: 4770365]
- 20.
- Чу Ю.С., Чжан З. Обнаружение убиквитинирования белков. J Vis Exp. 2009 августа 19; (30) [Бесплатная статья PMC: PMC3149903] [PubMed: 19692941]
- 21.
- Löser C, Möllgaard A, Fölsch UR. Фекальная эластаза 1: новый, высокочувствительный и специфический бесконтактный тест на функцию поджелудочной железы. Кишечник. 1996 Октябрь; 39 (4): 580-6. [Бесплатная статья PMC: PMC1383273] [PubMed: 8944569]
- 22.
- Forsmark C, Adams PC. Функциональное тестирование поджелудочной железы — полезно, но недостаточно.Можно J Гастроэнтерол. 2009 августа; 23 (8): 529-30. [Бесплатная статья PMC: PMC2732171] [PubMed: 19668794]
- 23.
- Ooi CY, Durie PR. Мутации гена трансмембранного регулятора проводимости муковисцидоза (CFTR) при панкреатите. J Cyst Fibros. 2012 сентябрь; 11 (5): 355-62. [PubMed: 22658665]
- 24.
- Сомараджу У. Р., Солис-Моя А. Заместительная терапия ферментами поджелудочной железы для людей с муковисцидозом. Кокрановская база данных Syst Rev. 23 ноября 2016 г .; 11: CD008227. [Бесплатная статья PMC: PMC6734127] [PubMed: 27878805]
Биохимия, катаболизм белков — StatPearls
Введение
Катаболизм белков — это расщепление белков на всасываемые мономеры для дальнейшей деградации или повторной сборки.Катаболизм белков в просвете кишечника важен по нескольким причинам, одна из которых — мобилизация незаменимых аминокислот для всасывания. Незаменимые аминокислоты не могут быть синтезированы в организме человека, но они необходимы для биосинтеза жизненно важных белков, поэтому их единственный источник — распад полипептидов с помощью пищеварительных ферментов. Этот процесс начинается в желудке и продолжается в тонком кишечнике. Большие белковые цепи разбираются, чтобы в конечном итоге оставить свободные аминокислоты, которые могут попасть в кровь и транспортироваться к различным клеткам по всему телу для дальнейшего расщепления.
Эндопептидазы в форме зимогена высвобождаются слизистой оболочкой желудка и экзокринной поджелудочной железой для расщепления полипептидной цепи между определенными аминокислотными остатками. Оказавшись в меньшей форме, экзопептидазы по одной удаляют последние аминокислоты с C- или N-конца дипептида или трипептида, способствуя абсорбции на микроворсинках. Клетки могут использовать эти аминокислоты для создания жизненно важных белков или в качестве субстратов для создания энергии. Белки, созданные внутриклеточно, также могут катаболизироваться по тем же причинам.Внутриклеточные белки, которые были неправильно свернуты или больше не функционируют в клетке, также подвергаются внутриклеточному катаболизму белков в лизосомах с помощью образования убиквитина и протеасом. Если клетка находится в низкоэнергетическом состоянии, свободные аминокислоты в цитозоле далее разлагаются с образованием промежуточных продуктов цикла лимонной кислоты и направляются туда для производства АТФ. В то время как углеродный каркас вступает в энергетические пути, азотный каркас модифицируется и выводится в основном через почки.
Основы
rКатаболический процесс начинается, когда протеолитические ферменты высвобождаются в форме зимогена из слизистой оболочки желудочно-кишечного тракта. После секреции зимогены расщепляются на активные протеазы. Пепсин из слизистой оболочки желудка гидролизует более крупный полипептид на более мелкие полипептиды, которые позже могут быть расщеплены протеазами поджелудочной железы, высвобождаемыми в двенадцатиперстной кишке. Трипсин, химотрипсин, эластаза и карбоксипептидазы, синтезируемые экзокринной поджелудочной железой, могут расщеплять полипептид в определенных точках расщепления на аминокислоты и олигопептиды.Аминопептидазы, расположенные на щеточной кайме эпителиальных клеток кишечника, гидролизуют оставшиеся олигопептиды до аминокислот, которые затем попадают в кровоток через трансмембранные транспортные системы. Существуют как облегченные, так и активные транспортные механизмы для транспортировки аминокислот в клетки.
Катаболизм белков — важная часть клеточного обновления. Когда цитозольные белки, такие как сигнальные или структурные пептиды, больше не нужны, они должны расщепляться в лизосомах для создания новых белков, которые могут выполнять необходимые метаболические функции.Если полученные аминокислоты не используются для синтеза новых белков для жизненно важных внутриклеточных функций, они могут войти в цикл лимонной кислоты для выработки энергии. Когда запрограммированная смерть — это судьба клетки, катепсины и другие катаболические ферменты работают вместе в процессе апоптоза.
Пептидазы желудка: главные клетки слизистой оболочки желудка секретируют пепсиноген. [1] С помощью соляной кислоты, высвобождаемой париетальными клетками желудка, пепсиноген изменяет конформацию в сильнокислой среде желудка и расщепляется на активную протеазу, пепсин.Кислота также денатурирует пищевые белки и частично разворачивает их для облегчения протеолиза. Пепсин обычно расщепляет пептидные связи, содержащие карбоксильную группу, от кислой или ароматической аминокислоты; однако его широкая специфичность позволяет ему расщеплять пептидные связи и в других местах. [2] Меньшие пептиды, а иногда и свободные аминокислоты, затем попадают в двенадцатиперстную кишку, чтобы встретить пептидазы, секретируемые поджелудочной железой.
Пептидазы поджелудочной железы и кишечника: экзокринная поджелудочная железа секретирует различные пищеварительные ферменты, включая амилазу, липазу, колипазу, бикарбонат и неактивные протеазы.Бикарбонат, высвобождаемый в просвет кишечника, повышает pH и позволяет протеазам поджелудочной железы активироваться из их зимогенной формы. Одна конкретная протеаза, трипсин, является наиболее важной для процесса пищеварения, потому что она сама расщепляет белки, а также активирует другие протеазы из их неактивных форм. Трипсин отщепляется от его проферментной формы трипсиногена энтеропептидазой. Затем трипсин продолжает активировать химотрипсиноген, проэластазу и прокарбоксипептидазы в химотрипсин, эластазу и карбоксипептидазы соответственно.[3] Трипсин гидролизует пептидные связи с карбоксильной группой аргинина или лизина. Химотрипсин, с другой стороны, имеет тенденцию воздействовать на пептидные связи между гидрофобными аминокислотами. [4] Трипсин и химотрипсин относятся к сериновым протеазам и имеют аналогичный метод протеолиза. Они образуют каталитический тетраэдрический промежуточный продукт, располагая каталитическую триаду таким образом, что протоны, образующие водородные связи, могут разорвать пептидную связь между двумя аминокислотами белка-субстрата.[4] Эти два фермента также очень похожи по своему S1-связывающему карману, который содержит основную аминокислоту в трипсине и гидрофобную аминокислоту в химотрипсине. [4] Тем не менее, особенности и механизмы намного сложнее с их динамическими свойствами, а структурная гибкость является одним из наиболее важных.
Эластаза, также сериновая протеаза, расщепляет эластин, а также белки с небольшими гидрофобными боковыми цепями, такие как глицин или аланин. Ацинарные клетки поджелудочной железы выделяют две формы эластазы, причем эластаза II имеет более широкую специфичность, чем эластаза I.[5] Нуклеофильная аминокислота в активном центре эндопептидазы гидролизует пептидную связь на более мелкие пептидные цепи, которые нуждаются в дальнейшей деградации; это может происходить через экзопептидазы, которые удаляют одну аминокислоту с конца белковой цепи. Примером экзопептидазы является активируемая трипсином протеаза поджелудочной железы карбоксипептидаза. Существует две формы этого протеолитического фермента, A и B. Карбоксипептидаза A имеет тенденцию отщеплять гидрофобные аминокислоты от пептидной цепи, а карбоксипептидаза B отщепляет основные аминокислоты от пептида.Эпителиальные клетки кишечника также содержат экзопептидазы на щеточной кайме, называемые аминопептидазами, которые выделяют отдельные аминокислоты, которые переносятся в клетки [6].
Внутриклеточный обмен белков: белки внутри клетки также подвергаются катаболизму, чтобы пополнить пул внутриклеточных аминокислот. Хотя у всех белков есть период полураспада, некоторые из них разлагаются в течение нескольких минут, а некоторые — в течение часов или дней. Некоторыми примерами сильно синтезируемых и деградированных белков являются белки мышечных клеток, гемоглобин, эпителиальные клетки желудочно-кишечного тракта и сами протеолитические ферменты.Многие из этих белков расщепляются лизосомными протеазами, так что их аминокислотные составляющие могут использоваться для восстановления белков, потребности которых зависят от текущего физиологического статуса клетки. Например, в периоды голодания аминокислоты, высвобождаемые в результате катаболизма мышечных белков, могут попасть в глюконеогенез и действовать как источник энергии. В сытом состоянии или во время метаболического ацидоза глутамин может быть топливом для почечной системы. Скелетные мышцы могут абсорбировать аланин, аспартат и глутамат и посредством трансаминирования образовывать полезные промежуточные соединения цикла Кребса.
Клеточный
Трансмембранный транспорт: Работа эндопептидаз, химотрипсина, эластазы и карбоксипептидазы и экзопептидаз, карбоксипептидазы и аминопептидазы в конечном итоге дает свободные аминокислоты. Существуют механизмы облегченной диффузии и активного транспорта для поглощения этих соединений из просвета кишечника посредством вторичного активного транспорта [7]. Аминокислоты пересекают щеточную границу кишечных клеток через натрий-зависимую транспортную систему, в которой низкая внутриклеточная концентрация натрия способствует совместному поступлению натрия и аминокислоты.Натрий-калиевая АТФаза на базолатеральной мембране поддерживает низкую концентрацию натрия внутри клетки. Как только аминокислоты попадают в клетку на апикальной мембране, они транспортируются вниз по своему градиенту на базальной мембране в кровоток, обычно в портальную циркуляцию. Шесть различных натрийзависимых аминокислотных переносчиков частично совпадают по своей специфичности в отношении аминокислот, которые они транспортируют через апикальную мембрану щеточной каймы; однако носители отдают предпочтение типам транспортируемых аминокислот.[7] Предпочтения включают нейтральные аминокислоты, кислые аминокислоты, основные аминокислоты, цистин, пролин и гидроксипролин. Натрий-зависимые аминокислотные переносчики на клетках просвета кишечника также существуют на почечном эпителии в различных формах изоферментов [8]. С другой стороны, облегченные транспортные системы на базальной стороне сопоставимы с системами, обнаруженными в других типах тканей. Аминокислоты также могут абсорбироваться из крови через эти облегченные переносчики и использоваться в качестве субстратов для цикла лимонной кислоты для выработки энергии — двунаправленность облегченных переносчиков помогает выживанию кишечного эпителия во время голодания.
Транспортеры на апикальной поверхности мышц, печени и других тканей поглощают аминокислоты из крови и концентрируют их. Хотя эти переносчики сходны с люминальными натрий-зависимыми переносчиками аминокислот на кишечном эпителии, они различаются по своим генетическим основам, белковому составу и специфичности [8]. Многие ткани экспрессируют A-систему, ASC-систему и L-систему для транспорта аминокислот в клетку; однако из них L-система — единственная без натрий-зависимого механизма.Клетки кишечника и почек экспрессируют B-систему, X-систему и иминосистему. [9] Конкретный тип ткани может содержать конкретный тип переносчика аминокислот, не обнаруживаемый в других тканях, например, экспрессия N-системы в печени и ее специфичность в отношении абсорбции глутамина. Изоформа N-системы для поглощения глутамина может экспрессироваться в другом типе ткани. Однако его свойства и состав будут отличаться от того, что содержится в печени. Каждая система отличается своей специфичностью в отношении аминокислот и имеет значение для множества различных патологий.
Лизосомная деградация: если белки больше не нужны или синтезируются неправильно, внутриклеточные белки расщепляются с использованием лизосом и протеасомного комплекса. Белки попадают в лизосомы посредством аутофагии, строго регулируемого процесса, при котором везикулы сливаются с лизосомной мембраной. [10] Аутофагии могут способствовать состояния низкой энергии, такие как низкий уровень АТФ или высокий уровень АМФ в клетке. Поскольку АМФ-активированная протеинкиназа активируется при высоких уровнях АМФ, она может фосфорилировать комплекс 1 и 2 туберозного склероза, что приводит к активации Rheb-GTPase.[11] Эта активация важна, потому что она инактивирует Rheb и, таким образом, также инактивирует mTOR, главный ингибитор аутофагии; вот как низкоэнергетические состояния способствуют катаболизму белков. Напротив, активация комплекса 1 и 2 туберозного склероза инсулином через киназу Akt приводит к активному mTOR. [11] Активированный mTOR способствует синтезу белка, а не деградации.
Убиквитинирование — это процесс, при котором молекула убиквитина становится ковалентно прикрепленной к расщепляемым белкам и взаимодействует с протеасомой в лизосоме для расщепления указанного белка с использованием АТФ-зависимого процесса.[12] Убиквитин сначала должен активироваться через E1, E2 и E3, прежде чем он ковалентно присоединится к эпсилон-аминогруппе боковых цепей лизина через ферментный комплекс. [13] Обычно белок получает больше молекул убиквитина на конце лизина, образуя хвост. После формирования хвоста убиквитинированный белок отделяется от комплекса лигирующих ферментов и перемещается в протеасому. Протеасома представляет собой цилиндрический комплекс протеазы 26S с несколькими каталитическими центрами внутри. Протеасомный комплекс 26S имеет два альфа- и два бета-кольца, содержащих три внутренних каталитических сайта, подобных трипсину, химотрипсину и постглутамилпептидилгидролазе.Конструкция четырехкольцевой структуры такова, что альфа-кольца играют стабилизирующую роль, тогда как бета-субъединицы являются активными каталитическими частями комплекса. [13] Он сохраняет молекулы убиквитина, но использует гидролиз АТФ, чтобы развернуть и продвинуть белок глубже в комплекс и высвободить свободные аминокислоты. [10] Белки, гидролизуемые этим процессом, известны как последовательности PEST, названные так в честь общих аминокислот, включенных в цепь. Аминокислоты пролин, глутамат, серин и треонин находятся в высоких концентрациях в белках последовательности PEST.[14]
Попав внутрь лизосомы, лизосомальные протеазы, называемые катепсинами, также могут разрушать цитозольные белки, помеченные как катаболизм. Катепсины — это еще один тип цистеиновой протеазы, присутствующий в перицеллюлярной среде, ядрах цитозоля и митохондриях. [15] Передача сигналов фактора некроза опухоли, клеточный стресс и каспаза могут запускать лизосомное высвобождение катепсина, расщепляя внутриклеточные белки, такие как Bid, что приводит к последующей активации каскада апоптотических каспаз. [16]
Молекулярный
Судьба пула внутриклеточных аминокислот сильно зависит от метаболического состояния отдельных тканей и организма в целом.В физиологическом состоянии сытости аминокислоты могут разлагаться дальше до их углеродного и азотного скелетов. В то время как углеродная основа может подвергаться окислению для получения энергии, азотный компонент выводится из организма через азотсодержащие отходы. Первоначально азот присутствует в форме аммиака, токсичного для человеческого организма вещества. Печень превращает токсичный аммиак в мочевину через цикл мочевины в нетоксичную водорастворимую форму азота, которая легко выводится через почки.
По мере того, как азотный скелет выбрасывается, углеродный скелет аминокислот может использоваться для производства энергии путем окисления в пируват (триптофан, глицин, аланин, серин, цистеин) и ацетил-КоА. Пируват и ацетилкофермент А являются субстратами цикла лимонной кислоты, в конечном итоге производящим АТФ и диоксид углерода. Ацетил-КоА может также образовывать кетоновые тела, попадающие в кровоток для использования в качестве источника энергии в определенных тканях. Аминокислоты, которые становятся ацетил-КоА или ацетоацетатом, называются кетогенными из-за их склонности превращаться в кетоновые тела.[17] Например, лизин и лейцин являются кетогенными, а также незаменимыми аминокислотами.
Другая возможная судьба — превращение углеродного скелета в промежуточные продукты цикла лимонной кислоты альфа-кетоглутарат (аргинин, гистидин, глутамин, пролин, глутамат), сукцинил-КоА (валин, треонин, изолейцин, метионин), фумарат (аспартат, тирозин, фенилаланин) и оксалоацетат (аспартат, аспарагин). [18] Стоит отметить, что малат, промежуточный продукт цикла лимонной кислоты, может перемещаться по кровообращению в печень и участвовать в глюконеогенезе.[19] Если аминокислота может стать предшественником глюкозы, она называется глюкогенной. Из-за различных промежуточных продуктов и точек входа в цикл Кребса аминокислоты могут быть как глюкогенными, так и кетогенными.
Тестирование
Убиквитинирование — начало внутриклеточного катаболизма белков. Тестирование функции этого процесса для понимания конкретных патологий включает комбинирование культивируемых клеток с плазмидами, содержащими белок с убиквитином, меченным эпитопом. Следующие шаги включают добавление стабилизирующих буферов, лизирование и обработку клеток ультразвуком с последующими циклами инкубации.Поскольку окончательная форма анализа включает иммуноблоттинг с гелем SDS-PAGE, должны быть сформированы конъюгированные антитела против белка [20]. Существуют также способы анализа процесса убиквитинирования in vitro, а не in vivo. [20]
Существуют способы проверки ферментов, выделяемых экзокринной поджелудочной железой, трипсина, химотрипсина и эластазы. Одним из тестов является тест на эластазу кала, при котором стул анализируется на концентрацию эластазы, что подтверждает проходимость протока поджелудочной железы, секреторную функцию экзокринной функции поджелудочной железы и наличие активирующего трипсина.[21] Этот тест является наиболее широко используемым из-за его высокой специфичности и неинвазивности. Более инвазивный тест — это эндоскопический тест функции поджелудочной железы, который доступен только в некоторых медицинских учреждениях. В этом тесте пациенту вводят успокоительное и перед эндоскопической процедурой вводят секретин [22]. Эндоскоп отсасывает другую жидкость из верхних отделов желудочно-кишечного тракта по пути к двенадцатиперстной кишке. Затем образцы секрета поджелудочной железы берутся на большом сосочке и анализируются на бикарбонат в лаборатории.[22] Как правило, образцы собираются с интервалами в 15, 30, 45 и 60 минут. Оценка секреции и активности этих ферментов важна для оценки патологий мальабсорбции белка.
Патофизиология
Муковисцидоз — аутосомно-рецессивная мутация в гене CFTR, который кодирует белки, составляющие поры хлоридного канала. [23] Тяжесть заболевания варьируется, но некоторые особенности являются общими для всех форм, например, образование толстых слизистых пробок в протоке поджелудочной железы, легких и мужской мочеполовой системе.Поскольку белки должны разрушаться для всасывания, слизистая пробка, блокирующая высвобождение зимогена из экзокринной поджелудочной железы, приведет к отсутствию катаболизма жизненно важных белков в просвете кишечника. [23] Пациенты с муковисцидозом страдают серьезным дефицитом белка и должны получать добавку экзогенных ферментов поджелудочной железы, хотя следует упомянуть, что побочные эффекты длительного приема добавок ферментов поджелудочной железы в настоящее время неизвестны. [24]
Квашиоркор — еще один серьезный дефицит белка; однако это связано с недостаточным потреблением белка, а не с генетическим заболеванием.Пищеварительные ферменты присутствуют в квашиоркоре; однако из-за того, что потребляется мало белка, присутствуют симптомы дефицита белка. У людей с истощением, принимающих квашиоркор, жизненные реакции в организме поддерживаются за счет внутриклеточного катаболизма белков. Потребность в доступных незаменимых аминокислотах приводит к значительному снижению периферической мышечной массы из-за разрушения мышц. [10] Дефицит белка также приводит к снижению уровня альбумина в сыворотке крови, что снижает внутрисосудистое коллоидное давление, что приводит к отеку и вздутию живота.Тяжелая форма квашиоркора быстро ухудшается, поскольку пищеварительные ферменты больше не вырабатываются, а эпителий тонкого кишечника не регенерируется.
Нарушение механизмов облегченного и активного транспорта может привести к патологическим состояниям мальабсорбции. Цистинурия и болезнь Хартнупа являются генетическими нарушениями, связанными с мембранными переносчиками аминокислот, но различаются группами транспортируемых аминокислот и поэтому проявляются по-разному клинически [9]. Цистинурия — это нарушение транспорта основных аминокислот через мембраны почечной и желудочно-кишечной систем.Отличительным признаком этого заболевания является образование почечных камней из-за неспособности резорбировать основную аминокислоту цистин из клубочкового фильтрата. Поскольку переносчик также существует в эпителии тонкого кишечника, цистин и другие основные аминокислоты плохо всасываются из просвета кишечника. Болезнь Хартнупа — заболевание, похожее по механизму, представляет собой дефект в транспортировке нейтральных аминокислот через почечную и кишечную систему. Поскольку триптофан является одной из нейтральных аминокислот, плохо усваиваемых, при физикальном осмотре выявляются сыпь, диарея и психические расстройства, похожие на пеллагру.В отличие от пеллагры, добавление ниацина мало помогает избавиться от симптомов и должно выявить необходимость генетического тестирования.
Клиническая значимость
Правильное функционирование катаболизма белков имеет первостепенное значение для поддержания метаболических потребностей человеческого организма. Разрушение больших полипептидных цепей с высвобождением свободных незаменимых и заменимых аминокислот обеспечивает клетки необходимыми субстратами для синтеза белка или выработки энергии. Распад внутриклеточного белка важен для нормального обмена веществ, а также при недоедании и истощенных физиологических состояниях, чтобы мобилизовать аминокислоты на экспорт для поддержки энергетических потребностей жизненно важных органов.Нарушения катаболизма белков могут привести к разнообразным клиническим проявлениям, затрагивающим почти все системы органов.
Ссылки
- 1.
- Magee DF. Пепсин. Med Clin North Am. 1974 ноя; 58 (6): 1277-87. [PubMed: 4610293]
- 2.
- Fruton JS. Специфика и механизм действия пепсина. Adv Enzymol Relat Areas Mol Biol. 1970; 33: 401-43. [PubMed: 4916858]
- 3.
- Джи Б, Logsdon CD. Усвоение новой информации о роли трипсина при панкреатите.Гастроэнтерология. 2011 декабрь; 141 (6): 1972-5. [Бесплатная статья PMC: PMC4327863] [PubMed: 22033179]
- 4.
- Ma W, Tang C, Lai L. Специфичность трипсина и химотрипсина: определяемая динамическая корреляция, контролируемая движением петли. Biophys J. 2005 Aug; 89 (2): 1183-93. [Бесплатная статья PMC: PMC1366603] [PubMed: 15923233]
- 5.
- Del Mar EG, Largman C, Brodrick JW, Fassett M, Geokas MC. Субстратная специфичность эластазы поджелудочной железы человека 2. Биохимия. 1980, 5 февраля; 19 (3): 468-72.[PubMed: 6898442]
- 6.
- Tobey N, Heizer W., Yeh R, Huang T.I, Hoffner C. Пептидазы щеточной каймы кишечника человека. Гастроэнтерология. 1985 Апрель; 88 (4): 913-26. [PubMed: 3882510]
- 7.
- Холмс Р., Лобли Р. В.. Повторный визит к краю кишечной щетки. Кишечник. 1989 декабрь; 30 (12): 1667-78. [Бесплатная статья PMC: PMC1434444] [PubMed: 2693228]
- 8.
- Bröer S. Транспорт аминокислот через эпителий кишечника и почек млекопитающих. Physiol Rev.2008 Янв; 88 (1): 249-86.[PubMed: 18195088]
- 9.
- Bröer S, Fairweather SJ. Транспорт аминокислот через кишечник млекопитающих. Compr Physiol. 2018 13 декабря; 9 (1): 343-373. [PubMed: 30549024]
- 10.
- Сандри М. Распад белка при истощении мышц: роль аутофагии-лизосомы и убиквитин-протеасомы. Int J Biochem Cell Biol. 2013 Октябрь; 45 (10): 2121-9. [Бесплатная статья PMC: PMC3775123] [PubMed: 23665154]
- 11.
- Пуэртоллано Р. mTOR и регуляция лизосом. F1000Prime Rep.2014; 6: 52. [Бесплатная статья PMC: PMC4108950] [PubMed: 25184042]
- 12.
- Доэрти Ф.Дж., Доусон С., Майер Р.Дж.. Убиквитин-протеасомный путь внутриклеточного протеолиза. Очерки Биохимии. 2002; 38: 51-63. [PubMed: 12463161]
- 13.
- Цехановер А. Путь убиквитин-протеасома: гибель белка и жизнь клетки. EMBO J. 15 декабря 1998; 17 (24): 7151-60. [Бесплатная статья PMC: PMC1171061] [PubMed: 9857172]
- 14.
- Rechsteiner M, Rogers SW. Последовательности PEST и регуляция протеолизом.Trends Biochem Sci. 1996 июл; 21 (7): 267-71. [PubMed: 8755249]
- 15.
- Брикс К., Дункхорст А., Майер К., Джорданс С. Катепсины цистеина: клеточная дорожная карта для различных функций. Биохимия. 2008 Февраль; 90 (2): 194-207. [PubMed: 17825974]
- 16.
- Лейст М., Яаттеля М. Запуск апоптоза катепсинами. Смерть клетки отличается. 2001 апр; 8 (4): 324-6. [PubMed: 11550083]
- 17.
- Харрис Р.А., Джоши М., Чжунг Н.Х., Обаяши М. Обзор молекулярных и биохимических основ катаболизма аминокислот с разветвленной цепью.J Nutr. 2005 июн; 135 (6 доп.): 1527S-30S. [PubMed: 15930464]
- 18.
- БЕЛЛАМИ Д. Эндогенные промежуточные продукты цикла лимонной кислоты и аминокислоты митохондрий. Biochem J. 1962, январь; 82: 218-24. [Бесплатная статья PMC: PMC1243435] [PubMed: 13866863]
- 19.
- Айкава Т., Мацутака Х., Ямамото Х., Окуда Т., Исикава Э. Глюконеогенез и метаболизм аминокислот. II. Межорганические отношения и роль глутамина и аланина в метаболизме аминокислот голодных крыс. J Biochem.1973 Ноябрь; 74 (5): 1003-17. [PubMed: 4770365]
- 20.
- Чу Ю.С., Чжан З. Обнаружение убиквитинирования белков. J Vis Exp. 2009 августа 19; (30) [Бесплатная статья PMC: PMC3149903] [PubMed: 19692941]
- 21.
- Löser C, Möllgaard A, Fölsch UR. Фекальная эластаза 1: новый, высокочувствительный и специфический бесконтактный тест на функцию поджелудочной железы. Кишечник. 1996 Октябрь; 39 (4): 580-6. [Бесплатная статья PMC: PMC1383273] [PubMed: 8944569]
- 22.
- Forsmark C, Adams PC. Функциональное тестирование поджелудочной железы — полезно, но недостаточно.Можно J Гастроэнтерол. 2009 августа; 23 (8): 529-30. [Бесплатная статья PMC: PMC2732171] [PubMed: 19668794]
- 23.
- Ooi CY, Durie PR. Мутации гена трансмембранного регулятора проводимости муковисцидоза (CFTR) при панкреатите. J Cyst Fibros. 2012 сентябрь; 11 (5): 355-62. [PubMed: 22658665]
- 24.
- Сомараджу У. Р., Солис-Моя А. Заместительная терапия ферментами поджелудочной железы для людей с муковисцидозом. Кокрановская база данных Syst Rev. 23 ноября 2016 г .; 11: CD008227. [Бесплатная статья PMC: PMC6734127] [PubMed: 27878805]
Биохимия, катаболизм белков — StatPearls
Введение
Катаболизм белков — это расщепление белков на всасываемые мономеры для дальнейшей деградации или повторной сборки.Катаболизм белков в просвете кишечника важен по нескольким причинам, одна из которых — мобилизация незаменимых аминокислот для всасывания. Незаменимые аминокислоты не могут быть синтезированы в организме человека, но они необходимы для биосинтеза жизненно важных белков, поэтому их единственный источник — распад полипептидов с помощью пищеварительных ферментов. Этот процесс начинается в желудке и продолжается в тонком кишечнике. Большие белковые цепи разбираются, чтобы в конечном итоге оставить свободные аминокислоты, которые могут попасть в кровь и транспортироваться к различным клеткам по всему телу для дальнейшего расщепления.
Эндопептидазы в форме зимогена высвобождаются слизистой оболочкой желудка и экзокринной поджелудочной железой для расщепления полипептидной цепи между определенными аминокислотными остатками. Оказавшись в меньшей форме, экзопептидазы по одной удаляют последние аминокислоты с C- или N-конца дипептида или трипептида, способствуя абсорбции на микроворсинках. Клетки могут использовать эти аминокислоты для создания жизненно важных белков или в качестве субстратов для создания энергии. Белки, созданные внутриклеточно, также могут катаболизироваться по тем же причинам.Внутриклеточные белки, которые были неправильно свернуты или больше не функционируют в клетке, также подвергаются внутриклеточному катаболизму белков в лизосомах с помощью образования убиквитина и протеасом. Если клетка находится в низкоэнергетическом состоянии, свободные аминокислоты в цитозоле далее разлагаются с образованием промежуточных продуктов цикла лимонной кислоты и направляются туда для производства АТФ. В то время как углеродный каркас вступает в энергетические пути, азотный каркас модифицируется и выводится в основном через почки.
Основы
rКатаболический процесс начинается, когда протеолитические ферменты высвобождаются в форме зимогена из слизистой оболочки желудочно-кишечного тракта. После секреции зимогены расщепляются на активные протеазы. Пепсин из слизистой оболочки желудка гидролизует более крупный полипептид на более мелкие полипептиды, которые позже могут быть расщеплены протеазами поджелудочной железы, высвобождаемыми в двенадцатиперстной кишке. Трипсин, химотрипсин, эластаза и карбоксипептидазы, синтезируемые экзокринной поджелудочной железой, могут расщеплять полипептид в определенных точках расщепления на аминокислоты и олигопептиды.Аминопептидазы, расположенные на щеточной кайме эпителиальных клеток кишечника, гидролизуют оставшиеся олигопептиды до аминокислот, которые затем попадают в кровоток через трансмембранные транспортные системы. Существуют как облегченные, так и активные транспортные механизмы для транспортировки аминокислот в клетки.
Катаболизм белков — важная часть клеточного обновления. Когда цитозольные белки, такие как сигнальные или структурные пептиды, больше не нужны, они должны расщепляться в лизосомах для создания новых белков, которые могут выполнять необходимые метаболические функции.Если полученные аминокислоты не используются для синтеза новых белков для жизненно важных внутриклеточных функций, они могут войти в цикл лимонной кислоты для выработки энергии. Когда запрограммированная смерть — это судьба клетки, катепсины и другие катаболические ферменты работают вместе в процессе апоптоза.
Пептидазы желудка: главные клетки слизистой оболочки желудка секретируют пепсиноген. [1] С помощью соляной кислоты, высвобождаемой париетальными клетками желудка, пепсиноген изменяет конформацию в сильнокислой среде желудка и расщепляется на активную протеазу, пепсин.Кислота также денатурирует пищевые белки и частично разворачивает их для облегчения протеолиза. Пепсин обычно расщепляет пептидные связи, содержащие карбоксильную группу, от кислой или ароматической аминокислоты; однако его широкая специфичность позволяет ему расщеплять пептидные связи и в других местах. [2] Меньшие пептиды, а иногда и свободные аминокислоты, затем попадают в двенадцатиперстную кишку, чтобы встретить пептидазы, секретируемые поджелудочной железой.
Пептидазы поджелудочной железы и кишечника: экзокринная поджелудочная железа секретирует различные пищеварительные ферменты, включая амилазу, липазу, колипазу, бикарбонат и неактивные протеазы.Бикарбонат, высвобождаемый в просвет кишечника, повышает pH и позволяет протеазам поджелудочной железы активироваться из их зимогенной формы. Одна конкретная протеаза, трипсин, является наиболее важной для процесса пищеварения, потому что она сама расщепляет белки, а также активирует другие протеазы из их неактивных форм. Трипсин отщепляется от его проферментной формы трипсиногена энтеропептидазой. Затем трипсин продолжает активировать химотрипсиноген, проэластазу и прокарбоксипептидазы в химотрипсин, эластазу и карбоксипептидазы соответственно.[3] Трипсин гидролизует пептидные связи с карбоксильной группой аргинина или лизина. Химотрипсин, с другой стороны, имеет тенденцию воздействовать на пептидные связи между гидрофобными аминокислотами. [4] Трипсин и химотрипсин относятся к сериновым протеазам и имеют аналогичный метод протеолиза. Они образуют каталитический тетраэдрический промежуточный продукт, располагая каталитическую триаду таким образом, что протоны, образующие водородные связи, могут разорвать пептидную связь между двумя аминокислотами белка-субстрата.[4] Эти два фермента также очень похожи по своему S1-связывающему карману, который содержит основную аминокислоту в трипсине и гидрофобную аминокислоту в химотрипсине. [4] Тем не менее, особенности и механизмы намного сложнее с их динамическими свойствами, а структурная гибкость является одним из наиболее важных.
Эластаза, также сериновая протеаза, расщепляет эластин, а также белки с небольшими гидрофобными боковыми цепями, такие как глицин или аланин. Ацинарные клетки поджелудочной железы выделяют две формы эластазы, причем эластаза II имеет более широкую специфичность, чем эластаза I.[5] Нуклеофильная аминокислота в активном центре эндопептидазы гидролизует пептидную связь на более мелкие пептидные цепи, которые нуждаются в дальнейшей деградации; это может происходить через экзопептидазы, которые удаляют одну аминокислоту с конца белковой цепи. Примером экзопептидазы является активируемая трипсином протеаза поджелудочной железы карбоксипептидаза. Существует две формы этого протеолитического фермента, A и B. Карбоксипептидаза A имеет тенденцию отщеплять гидрофобные аминокислоты от пептидной цепи, а карбоксипептидаза B отщепляет основные аминокислоты от пептида.Эпителиальные клетки кишечника также содержат экзопептидазы на щеточной кайме, называемые аминопептидазами, которые выделяют отдельные аминокислоты, которые переносятся в клетки [6].
Внутриклеточный обмен белков: белки внутри клетки также подвергаются катаболизму, чтобы пополнить пул внутриклеточных аминокислот. Хотя у всех белков есть период полураспада, некоторые из них разлагаются в течение нескольких минут, а некоторые — в течение часов или дней. Некоторыми примерами сильно синтезируемых и деградированных белков являются белки мышечных клеток, гемоглобин, эпителиальные клетки желудочно-кишечного тракта и сами протеолитические ферменты.Многие из этих белков расщепляются лизосомными протеазами, так что их аминокислотные составляющие могут использоваться для восстановления белков, потребности которых зависят от текущего физиологического статуса клетки. Например, в периоды голодания аминокислоты, высвобождаемые в результате катаболизма мышечных белков, могут попасть в глюконеогенез и действовать как источник энергии. В сытом состоянии или во время метаболического ацидоза глутамин может быть топливом для почечной системы. Скелетные мышцы могут абсорбировать аланин, аспартат и глутамат и посредством трансаминирования образовывать полезные промежуточные соединения цикла Кребса.
Клеточный
Трансмембранный транспорт: Работа эндопептидаз, химотрипсина, эластазы и карбоксипептидазы и экзопептидаз, карбоксипептидазы и аминопептидазы в конечном итоге дает свободные аминокислоты. Существуют механизмы облегченной диффузии и активного транспорта для поглощения этих соединений из просвета кишечника посредством вторичного активного транспорта [7]. Аминокислоты пересекают щеточную границу кишечных клеток через натрий-зависимую транспортную систему, в которой низкая внутриклеточная концентрация натрия способствует совместному поступлению натрия и аминокислоты.Натрий-калиевая АТФаза на базолатеральной мембране поддерживает низкую концентрацию натрия внутри клетки. Как только аминокислоты попадают в клетку на апикальной мембране, они транспортируются вниз по своему градиенту на базальной мембране в кровоток, обычно в портальную циркуляцию. Шесть различных натрийзависимых аминокислотных переносчиков частично совпадают по своей специфичности в отношении аминокислот, которые они транспортируют через апикальную мембрану щеточной каймы; однако носители отдают предпочтение типам транспортируемых аминокислот.[7] Предпочтения включают нейтральные аминокислоты, кислые аминокислоты, основные аминокислоты, цистин, пролин и гидроксипролин. Натрий-зависимые аминокислотные переносчики на клетках просвета кишечника также существуют на почечном эпителии в различных формах изоферментов [8]. С другой стороны, облегченные транспортные системы на базальной стороне сопоставимы с системами, обнаруженными в других типах тканей. Аминокислоты также могут абсорбироваться из крови через эти облегченные переносчики и использоваться в качестве субстратов для цикла лимонной кислоты для выработки энергии — двунаправленность облегченных переносчиков помогает выживанию кишечного эпителия во время голодания.
Транспортеры на апикальной поверхности мышц, печени и других тканей поглощают аминокислоты из крови и концентрируют их. Хотя эти переносчики сходны с люминальными натрий-зависимыми переносчиками аминокислот на кишечном эпителии, они различаются по своим генетическим основам, белковому составу и специфичности [8]. Многие ткани экспрессируют A-систему, ASC-систему и L-систему для транспорта аминокислот в клетку; однако из них L-система — единственная без натрий-зависимого механизма.Клетки кишечника и почек экспрессируют B-систему, X-систему и иминосистему. [9] Конкретный тип ткани может содержать конкретный тип переносчика аминокислот, не обнаруживаемый в других тканях, например, экспрессия N-системы в печени и ее специфичность в отношении абсорбции глутамина. Изоформа N-системы для поглощения глутамина может экспрессироваться в другом типе ткани. Однако его свойства и состав будут отличаться от того, что содержится в печени. Каждая система отличается своей специфичностью в отношении аминокислот и имеет значение для множества различных патологий.
Лизосомная деградация: если белки больше не нужны или синтезируются неправильно, внутриклеточные белки расщепляются с использованием лизосом и протеасомного комплекса. Белки попадают в лизосомы посредством аутофагии, строго регулируемого процесса, при котором везикулы сливаются с лизосомной мембраной. [10] Аутофагии могут способствовать состояния низкой энергии, такие как низкий уровень АТФ или высокий уровень АМФ в клетке. Поскольку АМФ-активированная протеинкиназа активируется при высоких уровнях АМФ, она может фосфорилировать комплекс 1 и 2 туберозного склероза, что приводит к активации Rheb-GTPase.[11] Эта активация важна, потому что она инактивирует Rheb и, таким образом, также инактивирует mTOR, главный ингибитор аутофагии; вот как низкоэнергетические состояния способствуют катаболизму белков. Напротив, активация комплекса 1 и 2 туберозного склероза инсулином через киназу Akt приводит к активному mTOR. [11] Активированный mTOR способствует синтезу белка, а не деградации.
Убиквитинирование — это процесс, при котором молекула убиквитина становится ковалентно прикрепленной к расщепляемым белкам и взаимодействует с протеасомой в лизосоме для расщепления указанного белка с использованием АТФ-зависимого процесса.[12] Убиквитин сначала должен активироваться через E1, E2 и E3, прежде чем он ковалентно присоединится к эпсилон-аминогруппе боковых цепей лизина через ферментный комплекс. [13] Обычно белок получает больше молекул убиквитина на конце лизина, образуя хвост. После формирования хвоста убиквитинированный белок отделяется от комплекса лигирующих ферментов и перемещается в протеасому. Протеасома представляет собой цилиндрический комплекс протеазы 26S с несколькими каталитическими центрами внутри. Протеасомный комплекс 26S имеет два альфа- и два бета-кольца, содержащих три внутренних каталитических сайта, подобных трипсину, химотрипсину и постглутамилпептидилгидролазе.Конструкция четырехкольцевой структуры такова, что альфа-кольца играют стабилизирующую роль, тогда как бета-субъединицы являются активными каталитическими частями комплекса. [13] Он сохраняет молекулы убиквитина, но использует гидролиз АТФ, чтобы развернуть и продвинуть белок глубже в комплекс и высвободить свободные аминокислоты. [10] Белки, гидролизуемые этим процессом, известны как последовательности PEST, названные так в честь общих аминокислот, включенных в цепь. Аминокислоты пролин, глутамат, серин и треонин находятся в высоких концентрациях в белках последовательности PEST.[14]
Попав внутрь лизосомы, лизосомальные протеазы, называемые катепсинами, также могут разрушать цитозольные белки, помеченные как катаболизм. Катепсины — это еще один тип цистеиновой протеазы, присутствующий в перицеллюлярной среде, ядрах цитозоля и митохондриях. [15] Передача сигналов фактора некроза опухоли, клеточный стресс и каспаза могут запускать лизосомное высвобождение катепсина, расщепляя внутриклеточные белки, такие как Bid, что приводит к последующей активации каскада апоптотических каспаз. [16]
Молекулярный
Судьба пула внутриклеточных аминокислот сильно зависит от метаболического состояния отдельных тканей и организма в целом.В физиологическом состоянии сытости аминокислоты могут разлагаться дальше до их углеродного и азотного скелетов. В то время как углеродная основа может подвергаться окислению для получения энергии, азотный компонент выводится из организма через азотсодержащие отходы. Первоначально азот присутствует в форме аммиака, токсичного для человеческого организма вещества. Печень превращает токсичный аммиак в мочевину через цикл мочевины в нетоксичную водорастворимую форму азота, которая легко выводится через почки.
По мере того, как азотный скелет выбрасывается, углеродный скелет аминокислот может использоваться для производства энергии путем окисления в пируват (триптофан, глицин, аланин, серин, цистеин) и ацетил-КоА. Пируват и ацетилкофермент А являются субстратами цикла лимонной кислоты, в конечном итоге производящим АТФ и диоксид углерода. Ацетил-КоА может также образовывать кетоновые тела, попадающие в кровоток для использования в качестве источника энергии в определенных тканях. Аминокислоты, которые становятся ацетил-КоА или ацетоацетатом, называются кетогенными из-за их склонности превращаться в кетоновые тела.[17] Например, лизин и лейцин являются кетогенными, а также незаменимыми аминокислотами.
Другая возможная судьба — превращение углеродного скелета в промежуточные продукты цикла лимонной кислоты альфа-кетоглутарат (аргинин, гистидин, глутамин, пролин, глутамат), сукцинил-КоА (валин, треонин, изолейцин, метионин), фумарат (аспартат, тирозин, фенилаланин) и оксалоацетат (аспартат, аспарагин). [18] Стоит отметить, что малат, промежуточный продукт цикла лимонной кислоты, может перемещаться по кровообращению в печень и участвовать в глюконеогенезе.[19] Если аминокислота может стать предшественником глюкозы, она называется глюкогенной. Из-за различных промежуточных продуктов и точек входа в цикл Кребса аминокислоты могут быть как глюкогенными, так и кетогенными.
Тестирование
Убиквитинирование — начало внутриклеточного катаболизма белков. Тестирование функции этого процесса для понимания конкретных патологий включает комбинирование культивируемых клеток с плазмидами, содержащими белок с убиквитином, меченным эпитопом. Следующие шаги включают добавление стабилизирующих буферов, лизирование и обработку клеток ультразвуком с последующими циклами инкубации.Поскольку окончательная форма анализа включает иммуноблоттинг с гелем SDS-PAGE, должны быть сформированы конъюгированные антитела против белка [20]. Существуют также способы анализа процесса убиквитинирования in vitro, а не in vivo. [20]
Существуют способы проверки ферментов, выделяемых экзокринной поджелудочной железой, трипсина, химотрипсина и эластазы. Одним из тестов является тест на эластазу кала, при котором стул анализируется на концентрацию эластазы, что подтверждает проходимость протока поджелудочной железы, секреторную функцию экзокринной функции поджелудочной железы и наличие активирующего трипсина.[21] Этот тест является наиболее широко используемым из-за его высокой специфичности и неинвазивности. Более инвазивный тест — это эндоскопический тест функции поджелудочной железы, который доступен только в некоторых медицинских учреждениях. В этом тесте пациенту вводят успокоительное и перед эндоскопической процедурой вводят секретин [22]. Эндоскоп отсасывает другую жидкость из верхних отделов желудочно-кишечного тракта по пути к двенадцатиперстной кишке. Затем образцы секрета поджелудочной железы берутся на большом сосочке и анализируются на бикарбонат в лаборатории.[22] Как правило, образцы собираются с интервалами в 15, 30, 45 и 60 минут. Оценка секреции и активности этих ферментов важна для оценки патологий мальабсорбции белка.
Патофизиология
Муковисцидоз — аутосомно-рецессивная мутация в гене CFTR, который кодирует белки, составляющие поры хлоридного канала. [23] Тяжесть заболевания варьируется, но некоторые особенности являются общими для всех форм, например, образование толстых слизистых пробок в протоке поджелудочной железы, легких и мужской мочеполовой системе.Поскольку белки должны разрушаться для всасывания, слизистая пробка, блокирующая высвобождение зимогена из экзокринной поджелудочной железы, приведет к отсутствию катаболизма жизненно важных белков в просвете кишечника. [23] Пациенты с муковисцидозом страдают серьезным дефицитом белка и должны получать добавку экзогенных ферментов поджелудочной железы, хотя следует упомянуть, что побочные эффекты длительного приема добавок ферментов поджелудочной железы в настоящее время неизвестны. [24]
Квашиоркор — еще один серьезный дефицит белка; однако это связано с недостаточным потреблением белка, а не с генетическим заболеванием.Пищеварительные ферменты присутствуют в квашиоркоре; однако из-за того, что потребляется мало белка, присутствуют симптомы дефицита белка. У людей с истощением, принимающих квашиоркор, жизненные реакции в организме поддерживаются за счет внутриклеточного катаболизма белков. Потребность в доступных незаменимых аминокислотах приводит к значительному снижению периферической мышечной массы из-за разрушения мышц. [10] Дефицит белка также приводит к снижению уровня альбумина в сыворотке крови, что снижает внутрисосудистое коллоидное давление, что приводит к отеку и вздутию живота.Тяжелая форма квашиоркора быстро ухудшается, поскольку пищеварительные ферменты больше не вырабатываются, а эпителий тонкого кишечника не регенерируется.
Нарушение механизмов облегченного и активного транспорта может привести к патологическим состояниям мальабсорбции. Цистинурия и болезнь Хартнупа являются генетическими нарушениями, связанными с мембранными переносчиками аминокислот, но различаются группами транспортируемых аминокислот и поэтому проявляются по-разному клинически [9]. Цистинурия — это нарушение транспорта основных аминокислот через мембраны почечной и желудочно-кишечной систем.Отличительным признаком этого заболевания является образование почечных камней из-за неспособности резорбировать основную аминокислоту цистин из клубочкового фильтрата. Поскольку переносчик также существует в эпителии тонкого кишечника, цистин и другие основные аминокислоты плохо всасываются из просвета кишечника. Болезнь Хартнупа — заболевание, похожее по механизму, представляет собой дефект в транспортировке нейтральных аминокислот через почечную и кишечную систему. Поскольку триптофан является одной из нейтральных аминокислот, плохо усваиваемых, при физикальном осмотре выявляются сыпь, диарея и психические расстройства, похожие на пеллагру.В отличие от пеллагры, добавление ниацина мало помогает избавиться от симптомов и должно выявить необходимость генетического тестирования.
Клиническая значимость
Правильное функционирование катаболизма белков имеет первостепенное значение для поддержания метаболических потребностей человеческого организма. Разрушение больших полипептидных цепей с высвобождением свободных незаменимых и заменимых аминокислот обеспечивает клетки необходимыми субстратами для синтеза белка или выработки энергии. Распад внутриклеточного белка важен для нормального обмена веществ, а также при недоедании и истощенных физиологических состояниях, чтобы мобилизовать аминокислоты на экспорт для поддержки энергетических потребностей жизненно важных органов.Нарушения катаболизма белков могут привести к разнообразным клиническим проявлениям, затрагивающим почти все системы органов.
Ссылки
- 1.
- Magee DF. Пепсин. Med Clin North Am. 1974 ноя; 58 (6): 1277-87. [PubMed: 4610293]
- 2.
- Fruton JS. Специфика и механизм действия пепсина. Adv Enzymol Relat Areas Mol Biol. 1970; 33: 401-43. [PubMed: 4916858]
- 3.
- Джи Б, Logsdon CD. Усвоение новой информации о роли трипсина при панкреатите.Гастроэнтерология. 2011 декабрь; 141 (6): 1972-5. [Бесплатная статья PMC: PMC4327863] [PubMed: 22033179]
- 4.
- Ma W, Tang C, Lai L. Специфичность трипсина и химотрипсина: определяемая динамическая корреляция, контролируемая движением петли. Biophys J. 2005 Aug; 89 (2): 1183-93. [Бесплатная статья PMC: PMC1366603] [PubMed: 15923233]
- 5.
- Del Mar EG, Largman C, Brodrick JW, Fassett M, Geokas MC. Субстратная специфичность эластазы поджелудочной железы человека 2. Биохимия. 1980, 5 февраля; 19 (3): 468-72.[PubMed: 6898442]
- 6.
- Tobey N, Heizer W., Yeh R, Huang T.I, Hoffner C. Пептидазы щеточной каймы кишечника человека. Гастроэнтерология. 1985 Апрель; 88 (4): 913-26. [PubMed: 3882510]
- 7.
- Холмс Р., Лобли Р. В.. Повторный визит к краю кишечной щетки. Кишечник. 1989 декабрь; 30 (12): 1667-78. [Бесплатная статья PMC: PMC1434444] [PubMed: 2693228]
- 8.
- Bröer S. Транспорт аминокислот через эпителий кишечника и почек млекопитающих. Physiol Rev.2008 Янв; 88 (1): 249-86.[PubMed: 18195088]
- 9.
- Bröer S, Fairweather SJ. Транспорт аминокислот через кишечник млекопитающих. Compr Physiol. 2018 13 декабря; 9 (1): 343-373. [PubMed: 30549024]
- 10.
- Сандри М. Распад белка при истощении мышц: роль аутофагии-лизосомы и убиквитин-протеасомы. Int J Biochem Cell Biol. 2013 Октябрь; 45 (10): 2121-9. [Бесплатная статья PMC: PMC3775123] [PubMed: 23665154]
- 11.
- Пуэртоллано Р. mTOR и регуляция лизосом. F1000Prime Rep.2014; 6: 52. [Бесплатная статья PMC: PMC4108950] [PubMed: 25184042]
- 12.
- Доэрти Ф.Дж., Доусон С., Майер Р.Дж.. Убиквитин-протеасомный путь внутриклеточного протеолиза. Очерки Биохимии. 2002; 38: 51-63. [PubMed: 12463161]
- 13.
- Цехановер А. Путь убиквитин-протеасома: гибель белка и жизнь клетки. EMBO J. 15 декабря 1998; 17 (24): 7151-60. [Бесплатная статья PMC: PMC1171061] [PubMed: 9857172]
- 14.
- Rechsteiner M, Rogers SW. Последовательности PEST и регуляция протеолизом.Trends Biochem Sci. 1996 июл; 21 (7): 267-71. [PubMed: 8755249]
- 15.
- Брикс К., Дункхорст А., Майер К., Джорданс С. Катепсины цистеина: клеточная дорожная карта для различных функций. Биохимия. 2008 Февраль; 90 (2): 194-207. [PubMed: 17825974]
- 16.
- Лейст М., Яаттеля М. Запуск апоптоза катепсинами. Смерть клетки отличается. 2001 апр; 8 (4): 324-6. [PubMed: 11550083]
- 17.
- Харрис Р.А., Джоши М., Чжунг Н.Х., Обаяши М. Обзор молекулярных и биохимических основ катаболизма аминокислот с разветвленной цепью.J Nutr. 2005 июн; 135 (6 доп.): 1527S-30S. [PubMed: 15930464]
- 18.
- БЕЛЛАМИ Д. Эндогенные промежуточные продукты цикла лимонной кислоты и аминокислоты митохондрий. Biochem J. 1962, январь; 82: 218-24. [Бесплатная статья PMC: PMC1243435] [PubMed: 13866863]
- 19.
- Айкава Т., Мацутака Х., Ямамото Х., Окуда Т., Исикава Э. Глюконеогенез и метаболизм аминокислот. II. Межорганические отношения и роль глутамина и аланина в метаболизме аминокислот голодных крыс. J Biochem.1973 Ноябрь; 74 (5): 1003-17. [PubMed: 4770365]
- 20.
- Чу Ю.С., Чжан З. Обнаружение убиквитинирования белков. J Vis Exp. 2009 августа 19; (30) [Бесплатная статья PMC: PMC3149903] [PubMed: 19692941]
- 21.
- Löser C, Möllgaard A, Fölsch UR. Фекальная эластаза 1: новый, высокочувствительный и специфический бесконтактный тест на функцию поджелудочной железы. Кишечник. 1996 Октябрь; 39 (4): 580-6. [Бесплатная статья PMC: PMC1383273] [PubMed: 8944569]
- 22.
- Forsmark C, Adams PC. Функциональное тестирование поджелудочной железы — полезно, но недостаточно.Можно J Гастроэнтерол. 2009 августа; 23 (8): 529-30. [Бесплатная статья PMC: PMC2732171] [PubMed: 19668794]
- 23.
- Ooi CY, Durie PR. Мутации гена трансмембранного регулятора проводимости муковисцидоза (CFTR) при панкреатите. J Cyst Fibros. 2012 сентябрь; 11 (5): 355-62. [PubMed: 22658665]
- 24.
- Сомараджу У. Р., Солис-Моя А. Заместительная терапия ферментами поджелудочной железы для людей с муковисцидозом. Кокрановская база данных Syst Rev. 23 ноября 2016 г .; 11: CD008227. [Бесплатная статья PMC: PMC6734127] [PubMed: 27878805]
Биохимия, катаболизм белков — StatPearls
Введение
Катаболизм белков — это расщепление белков на всасываемые мономеры для дальнейшей деградации или повторной сборки.Катаболизм белков в просвете кишечника важен по нескольким причинам, одна из которых — мобилизация незаменимых аминокислот для всасывания. Незаменимые аминокислоты не могут быть синтезированы в организме человека, но они необходимы для биосинтеза жизненно важных белков, поэтому их единственный источник — распад полипептидов с помощью пищеварительных ферментов. Этот процесс начинается в желудке и продолжается в тонком кишечнике. Большие белковые цепи разбираются, чтобы в конечном итоге оставить свободные аминокислоты, которые могут попасть в кровь и транспортироваться к различным клеткам по всему телу для дальнейшего расщепления.
Эндопептидазы в форме зимогена высвобождаются слизистой оболочкой желудка и экзокринной поджелудочной железой для расщепления полипептидной цепи между определенными аминокислотными остатками. Оказавшись в меньшей форме, экзопептидазы по одной удаляют последние аминокислоты с C- или N-конца дипептида или трипептида, способствуя абсорбции на микроворсинках. Клетки могут использовать эти аминокислоты для создания жизненно важных белков или в качестве субстратов для создания энергии. Белки, созданные внутриклеточно, также могут катаболизироваться по тем же причинам.Внутриклеточные белки, которые были неправильно свернуты или больше не функционируют в клетке, также подвергаются внутриклеточному катаболизму белков в лизосомах с помощью образования убиквитина и протеасом. Если клетка находится в низкоэнергетическом состоянии, свободные аминокислоты в цитозоле далее разлагаются с образованием промежуточных продуктов цикла лимонной кислоты и направляются туда для производства АТФ. В то время как углеродный каркас вступает в энергетические пути, азотный каркас модифицируется и выводится в основном через почки.
Основы
rКатаболический процесс начинается, когда протеолитические ферменты высвобождаются в форме зимогена из слизистой оболочки желудочно-кишечного тракта. После секреции зимогены расщепляются на активные протеазы. Пепсин из слизистой оболочки желудка гидролизует более крупный полипептид на более мелкие полипептиды, которые позже могут быть расщеплены протеазами поджелудочной железы, высвобождаемыми в двенадцатиперстной кишке. Трипсин, химотрипсин, эластаза и карбоксипептидазы, синтезируемые экзокринной поджелудочной железой, могут расщеплять полипептид в определенных точках расщепления на аминокислоты и олигопептиды.Аминопептидазы, расположенные на щеточной кайме эпителиальных клеток кишечника, гидролизуют оставшиеся олигопептиды до аминокислот, которые затем попадают в кровоток через трансмембранные транспортные системы. Существуют как облегченные, так и активные транспортные механизмы для транспортировки аминокислот в клетки.
Катаболизм белков — важная часть клеточного обновления. Когда цитозольные белки, такие как сигнальные или структурные пептиды, больше не нужны, они должны расщепляться в лизосомах для создания новых белков, которые могут выполнять необходимые метаболические функции.Если полученные аминокислоты не используются для синтеза новых белков для жизненно важных внутриклеточных функций, они могут войти в цикл лимонной кислоты для выработки энергии. Когда запрограммированная смерть — это судьба клетки, катепсины и другие катаболические ферменты работают вместе в процессе апоптоза.
Пептидазы желудка: главные клетки слизистой оболочки желудка секретируют пепсиноген. [1] С помощью соляной кислоты, высвобождаемой париетальными клетками желудка, пепсиноген изменяет конформацию в сильнокислой среде желудка и расщепляется на активную протеазу, пепсин.Кислота также денатурирует пищевые белки и частично разворачивает их для облегчения протеолиза. Пепсин обычно расщепляет пептидные связи, содержащие карбоксильную группу, от кислой или ароматической аминокислоты; однако его широкая специфичность позволяет ему расщеплять пептидные связи и в других местах. [2] Меньшие пептиды, а иногда и свободные аминокислоты, затем попадают в двенадцатиперстную кишку, чтобы встретить пептидазы, секретируемые поджелудочной железой.
Пептидазы поджелудочной железы и кишечника: экзокринная поджелудочная железа секретирует различные пищеварительные ферменты, включая амилазу, липазу, колипазу, бикарбонат и неактивные протеазы.Бикарбонат, высвобождаемый в просвет кишечника, повышает pH и позволяет протеазам поджелудочной железы активироваться из их зимогенной формы. Одна конкретная протеаза, трипсин, является наиболее важной для процесса пищеварения, потому что она сама расщепляет белки, а также активирует другие протеазы из их неактивных форм. Трипсин отщепляется от его проферментной формы трипсиногена энтеропептидазой. Затем трипсин продолжает активировать химотрипсиноген, проэластазу и прокарбоксипептидазы в химотрипсин, эластазу и карбоксипептидазы соответственно.[3] Трипсин гидролизует пептидные связи с карбоксильной группой аргинина или лизина. Химотрипсин, с другой стороны, имеет тенденцию воздействовать на пептидные связи между гидрофобными аминокислотами. [4] Трипсин и химотрипсин относятся к сериновым протеазам и имеют аналогичный метод протеолиза. Они образуют каталитический тетраэдрический промежуточный продукт, располагая каталитическую триаду таким образом, что протоны, образующие водородные связи, могут разорвать пептидную связь между двумя аминокислотами белка-субстрата.[4] Эти два фермента также очень похожи по своему S1-связывающему карману, который содержит основную аминокислоту в трипсине и гидрофобную аминокислоту в химотрипсине. [4] Тем не менее, особенности и механизмы намного сложнее с их динамическими свойствами, а структурная гибкость является одним из наиболее важных.
Эластаза, также сериновая протеаза, расщепляет эластин, а также белки с небольшими гидрофобными боковыми цепями, такие как глицин или аланин. Ацинарные клетки поджелудочной железы выделяют две формы эластазы, причем эластаза II имеет более широкую специфичность, чем эластаза I.[5] Нуклеофильная аминокислота в активном центре эндопептидазы гидролизует пептидную связь на более мелкие пептидные цепи, которые нуждаются в дальнейшей деградации; это может происходить через экзопептидазы, которые удаляют одну аминокислоту с конца белковой цепи. Примером экзопептидазы является активируемая трипсином протеаза поджелудочной железы карбоксипептидаза. Существует две формы этого протеолитического фермента, A и B. Карбоксипептидаза A имеет тенденцию отщеплять гидрофобные аминокислоты от пептидной цепи, а карбоксипептидаза B отщепляет основные аминокислоты от пептида.Эпителиальные клетки кишечника также содержат экзопептидазы на щеточной кайме, называемые аминопептидазами, которые выделяют отдельные аминокислоты, которые переносятся в клетки [6].
Внутриклеточный обмен белков: белки внутри клетки также подвергаются катаболизму, чтобы пополнить пул внутриклеточных аминокислот. Хотя у всех белков есть период полураспада, некоторые из них разлагаются в течение нескольких минут, а некоторые — в течение часов или дней. Некоторыми примерами сильно синтезируемых и деградированных белков являются белки мышечных клеток, гемоглобин, эпителиальные клетки желудочно-кишечного тракта и сами протеолитические ферменты.Многие из этих белков расщепляются лизосомными протеазами, так что их аминокислотные составляющие могут использоваться для восстановления белков, потребности которых зависят от текущего физиологического статуса клетки. Например, в периоды голодания аминокислоты, высвобождаемые в результате катаболизма мышечных белков, могут попасть в глюконеогенез и действовать как источник энергии. В сытом состоянии или во время метаболического ацидоза глутамин может быть топливом для почечной системы. Скелетные мышцы могут абсорбировать аланин, аспартат и глутамат и посредством трансаминирования образовывать полезные промежуточные соединения цикла Кребса.
Клеточный
Трансмембранный транспорт: Работа эндопептидаз, химотрипсина, эластазы и карбоксипептидазы и экзопептидаз, карбоксипептидазы и аминопептидазы в конечном итоге дает свободные аминокислоты. Существуют механизмы облегченной диффузии и активного транспорта для поглощения этих соединений из просвета кишечника посредством вторичного активного транспорта [7]. Аминокислоты пересекают щеточную границу кишечных клеток через натрий-зависимую транспортную систему, в которой низкая внутриклеточная концентрация натрия способствует совместному поступлению натрия и аминокислоты.Натрий-калиевая АТФаза на базолатеральной мембране поддерживает низкую концентрацию натрия внутри клетки. Как только аминокислоты попадают в клетку на апикальной мембране, они транспортируются вниз по своему градиенту на базальной мембране в кровоток, обычно в портальную циркуляцию. Шесть различных натрийзависимых аминокислотных переносчиков частично совпадают по своей специфичности в отношении аминокислот, которые они транспортируют через апикальную мембрану щеточной каймы; однако носители отдают предпочтение типам транспортируемых аминокислот.[7] Предпочтения включают нейтральные аминокислоты, кислые аминокислоты, основные аминокислоты, цистин, пролин и гидроксипролин. Натрий-зависимые аминокислотные переносчики на клетках просвета кишечника также существуют на почечном эпителии в различных формах изоферментов [8]. С другой стороны, облегченные транспортные системы на базальной стороне сопоставимы с системами, обнаруженными в других типах тканей. Аминокислоты также могут абсорбироваться из крови через эти облегченные переносчики и использоваться в качестве субстратов для цикла лимонной кислоты для выработки энергии — двунаправленность облегченных переносчиков помогает выживанию кишечного эпителия во время голодания.
Транспортеры на апикальной поверхности мышц, печени и других тканей поглощают аминокислоты из крови и концентрируют их. Хотя эти переносчики сходны с люминальными натрий-зависимыми переносчиками аминокислот на кишечном эпителии, они различаются по своим генетическим основам, белковому составу и специфичности [8]. Многие ткани экспрессируют A-систему, ASC-систему и L-систему для транспорта аминокислот в клетку; однако из них L-система — единственная без натрий-зависимого механизма.Клетки кишечника и почек экспрессируют B-систему, X-систему и иминосистему. [9] Конкретный тип ткани может содержать конкретный тип переносчика аминокислот, не обнаруживаемый в других тканях, например, экспрессия N-системы в печени и ее специфичность в отношении абсорбции глутамина. Изоформа N-системы для поглощения глутамина может экспрессироваться в другом типе ткани. Однако его свойства и состав будут отличаться от того, что содержится в печени. Каждая система отличается своей специфичностью в отношении аминокислот и имеет значение для множества различных патологий.
Лизосомная деградация: если белки больше не нужны или синтезируются неправильно, внутриклеточные белки расщепляются с использованием лизосом и протеасомного комплекса. Белки попадают в лизосомы посредством аутофагии, строго регулируемого процесса, при котором везикулы сливаются с лизосомной мембраной. [10] Аутофагии могут способствовать состояния низкой энергии, такие как низкий уровень АТФ или высокий уровень АМФ в клетке. Поскольку АМФ-активированная протеинкиназа активируется при высоких уровнях АМФ, она может фосфорилировать комплекс 1 и 2 туберозного склероза, что приводит к активации Rheb-GTPase.[11] Эта активация важна, потому что она инактивирует Rheb и, таким образом, также инактивирует mTOR, главный ингибитор аутофагии; вот как низкоэнергетические состояния способствуют катаболизму белков. Напротив, активация комплекса 1 и 2 туберозного склероза инсулином через киназу Akt приводит к активному mTOR. [11] Активированный mTOR способствует синтезу белка, а не деградации.
Убиквитинирование — это процесс, при котором молекула убиквитина становится ковалентно прикрепленной к расщепляемым белкам и взаимодействует с протеасомой в лизосоме для расщепления указанного белка с использованием АТФ-зависимого процесса.[12] Убиквитин сначала должен активироваться через E1, E2 и E3, прежде чем он ковалентно присоединится к эпсилон-аминогруппе боковых цепей лизина через ферментный комплекс. [13] Обычно белок получает больше молекул убиквитина на конце лизина, образуя хвост. После формирования хвоста убиквитинированный белок отделяется от комплекса лигирующих ферментов и перемещается в протеасому. Протеасома представляет собой цилиндрический комплекс протеазы 26S с несколькими каталитическими центрами внутри. Протеасомный комплекс 26S имеет два альфа- и два бета-кольца, содержащих три внутренних каталитических сайта, подобных трипсину, химотрипсину и постглутамилпептидилгидролазе.Конструкция четырехкольцевой структуры такова, что альфа-кольца играют стабилизирующую роль, тогда как бета-субъединицы являются активными каталитическими частями комплекса. [13] Он сохраняет молекулы убиквитина, но использует гидролиз АТФ, чтобы развернуть и продвинуть белок глубже в комплекс и высвободить свободные аминокислоты. [10] Белки, гидролизуемые этим процессом, известны как последовательности PEST, названные так в честь общих аминокислот, включенных в цепь. Аминокислоты пролин, глутамат, серин и треонин находятся в высоких концентрациях в белках последовательности PEST.[14]
Попав внутрь лизосомы, лизосомальные протеазы, называемые катепсинами, также могут разрушать цитозольные белки, помеченные как катаболизм. Катепсины — это еще один тип цистеиновой протеазы, присутствующий в перицеллюлярной среде, ядрах цитозоля и митохондриях. [15] Передача сигналов фактора некроза опухоли, клеточный стресс и каспаза могут запускать лизосомное высвобождение катепсина, расщепляя внутриклеточные белки, такие как Bid, что приводит к последующей активации каскада апоптотических каспаз. [16]
Молекулярный
Судьба пула внутриклеточных аминокислот сильно зависит от метаболического состояния отдельных тканей и организма в целом.В физиологическом состоянии сытости аминокислоты могут разлагаться дальше до их углеродного и азотного скелетов. В то время как углеродная основа может подвергаться окислению для получения энергии, азотный компонент выводится из организма через азотсодержащие отходы. Первоначально азот присутствует в форме аммиака, токсичного для человеческого организма вещества. Печень превращает токсичный аммиак в мочевину через цикл мочевины в нетоксичную водорастворимую форму азота, которая легко выводится через почки.
По мере того, как азотный скелет выбрасывается, углеродный скелет аминокислот может использоваться для производства энергии путем окисления в пируват (триптофан, глицин, аланин, серин, цистеин) и ацетил-КоА. Пируват и ацетилкофермент А являются субстратами цикла лимонной кислоты, в конечном итоге производящим АТФ и диоксид углерода. Ацетил-КоА может также образовывать кетоновые тела, попадающие в кровоток для использования в качестве источника энергии в определенных тканях. Аминокислоты, которые становятся ацетил-КоА или ацетоацетатом, называются кетогенными из-за их склонности превращаться в кетоновые тела.[17] Например, лизин и лейцин являются кетогенными, а также незаменимыми аминокислотами.
Другая возможная судьба — превращение углеродного скелета в промежуточные продукты цикла лимонной кислоты альфа-кетоглутарат (аргинин, гистидин, глутамин, пролин, глутамат), сукцинил-КоА (валин, треонин, изолейцин, метионин), фумарат (аспартат, тирозин, фенилаланин) и оксалоацетат (аспартат, аспарагин). [18] Стоит отметить, что малат, промежуточный продукт цикла лимонной кислоты, может перемещаться по кровообращению в печень и участвовать в глюконеогенезе.[19] Если аминокислота может стать предшественником глюкозы, она называется глюкогенной. Из-за различных промежуточных продуктов и точек входа в цикл Кребса аминокислоты могут быть как глюкогенными, так и кетогенными.
Тестирование
Убиквитинирование — начало внутриклеточного катаболизма белков. Тестирование функции этого процесса для понимания конкретных патологий включает комбинирование культивируемых клеток с плазмидами, содержащими белок с убиквитином, меченным эпитопом. Следующие шаги включают добавление стабилизирующих буферов, лизирование и обработку клеток ультразвуком с последующими циклами инкубации.Поскольку окончательная форма анализа включает иммуноблоттинг с гелем SDS-PAGE, должны быть сформированы конъюгированные антитела против белка [20]. Существуют также способы анализа процесса убиквитинирования in vitro, а не in vivo. [20]
Существуют способы проверки ферментов, выделяемых экзокринной поджелудочной железой, трипсина, химотрипсина и эластазы. Одним из тестов является тест на эластазу кала, при котором стул анализируется на концентрацию эластазы, что подтверждает проходимость протока поджелудочной железы, секреторную функцию экзокринной функции поджелудочной железы и наличие активирующего трипсина.[21] Этот тест является наиболее широко используемым из-за его высокой специфичности и неинвазивности. Более инвазивный тест — это эндоскопический тест функции поджелудочной железы, который доступен только в некоторых медицинских учреждениях. В этом тесте пациенту вводят успокоительное и перед эндоскопической процедурой вводят секретин [22]. Эндоскоп отсасывает другую жидкость из верхних отделов желудочно-кишечного тракта по пути к двенадцатиперстной кишке. Затем образцы секрета поджелудочной железы берутся на большом сосочке и анализируются на бикарбонат в лаборатории.[22] Как правило, образцы собираются с интервалами в 15, 30, 45 и 60 минут. Оценка секреции и активности этих ферментов важна для оценки патологий мальабсорбции белка.
Патофизиология
Муковисцидоз — аутосомно-рецессивная мутация в гене CFTR, который кодирует белки, составляющие поры хлоридного канала. [23] Тяжесть заболевания варьируется, но некоторые особенности являются общими для всех форм, например, образование толстых слизистых пробок в протоке поджелудочной железы, легких и мужской мочеполовой системе.Поскольку белки должны разрушаться для всасывания, слизистая пробка, блокирующая высвобождение зимогена из экзокринной поджелудочной железы, приведет к отсутствию катаболизма жизненно важных белков в просвете кишечника. [23] Пациенты с муковисцидозом страдают серьезным дефицитом белка и должны получать добавку экзогенных ферментов поджелудочной железы, хотя следует упомянуть, что побочные эффекты длительного приема добавок ферментов поджелудочной железы в настоящее время неизвестны. [24]
Квашиоркор — еще один серьезный дефицит белка; однако это связано с недостаточным потреблением белка, а не с генетическим заболеванием.Пищеварительные ферменты присутствуют в квашиоркоре; однако из-за того, что потребляется мало белка, присутствуют симптомы дефицита белка. У людей с истощением, принимающих квашиоркор, жизненные реакции в организме поддерживаются за счет внутриклеточного катаболизма белков. Потребность в доступных незаменимых аминокислотах приводит к значительному снижению периферической мышечной массы из-за разрушения мышц. [10] Дефицит белка также приводит к снижению уровня альбумина в сыворотке крови, что снижает внутрисосудистое коллоидное давление, что приводит к отеку и вздутию живота.Тяжелая форма квашиоркора быстро ухудшается, поскольку пищеварительные ферменты больше не вырабатываются, а эпителий тонкого кишечника не регенерируется.
Нарушение механизмов облегченного и активного транспорта может привести к патологическим состояниям мальабсорбции. Цистинурия и болезнь Хартнупа являются генетическими нарушениями, связанными с мембранными переносчиками аминокислот, но различаются группами транспортируемых аминокислот и поэтому проявляются по-разному клинически [9]. Цистинурия — это нарушение транспорта основных аминокислот через мембраны почечной и желудочно-кишечной систем.Отличительным признаком этого заболевания является образование почечных камней из-за неспособности резорбировать основную аминокислоту цистин из клубочкового фильтрата. Поскольку переносчик также существует в эпителии тонкого кишечника, цистин и другие основные аминокислоты плохо всасываются из просвета кишечника. Болезнь Хартнупа — заболевание, похожее по механизму, представляет собой дефект в транспортировке нейтральных аминокислот через почечную и кишечную систему. Поскольку триптофан является одной из нейтральных аминокислот, плохо усваиваемых, при физикальном осмотре выявляются сыпь, диарея и психические расстройства, похожие на пеллагру.В отличие от пеллагры, добавление ниацина мало помогает избавиться от симптомов и должно выявить необходимость генетического тестирования.
Клиническая значимость
Правильное функционирование катаболизма белков имеет первостепенное значение для поддержания метаболических потребностей человеческого организма. Разрушение больших полипептидных цепей с высвобождением свободных незаменимых и заменимых аминокислот обеспечивает клетки необходимыми субстратами для синтеза белка или выработки энергии. Распад внутриклеточного белка важен для нормального обмена веществ, а также при недоедании и истощенных физиологических состояниях, чтобы мобилизовать аминокислоты на экспорт для поддержки энергетических потребностей жизненно важных органов.Нарушения катаболизма белков могут привести к разнообразным клиническим проявлениям, затрагивающим почти все системы органов.
Ссылки
- 1.
- Magee DF. Пепсин. Med Clin North Am. 1974 ноя; 58 (6): 1277-87. [PubMed: 4610293]
- 2.
- Fruton JS. Специфика и механизм действия пепсина. Adv Enzymol Relat Areas Mol Biol. 1970; 33: 401-43. [PubMed: 4916858]
- 3.
- Джи Б, Logsdon CD. Усвоение новой информации о роли трипсина при панкреатите.Гастроэнтерология. 2011 декабрь; 141 (6): 1972-5. [Бесплатная статья PMC: PMC4327863] [PubMed: 22033179]
- 4.
- Ma W, Tang C, Lai L. Специфичность трипсина и химотрипсина: определяемая динамическая корреляция, контролируемая движением петли. Biophys J. 2005 Aug; 89 (2): 1183-93. [Бесплатная статья PMC: PMC1366603] [PubMed: 15923233]
- 5.
- Del Mar EG, Largman C, Brodrick JW, Fassett M, Geokas MC. Субстратная специфичность эластазы поджелудочной железы человека 2. Биохимия. 1980, 5 февраля; 19 (3): 468-72.[PubMed: 6898442]
- 6.
- Tobey N, Heizer W., Yeh R, Huang T.I, Hoffner C. Пептидазы щеточной каймы кишечника человека. Гастроэнтерология. 1985 Апрель; 88 (4): 913-26. [PubMed: 3882510]
- 7.
- Холмс Р., Лобли Р. В.. Повторный визит к краю кишечной щетки. Кишечник. 1989 декабрь; 30 (12): 1667-78. [Бесплатная статья PMC: PMC1434444] [PubMed: 2693228]
- 8.
- Bröer S. Транспорт аминокислот через эпителий кишечника и почек млекопитающих. Physiol Rev.2008 Янв; 88 (1): 249-86.[PubMed: 18195088]
- 9.
- Bröer S, Fairweather SJ. Транспорт аминокислот через кишечник млекопитающих. Compr Physiol. 2018 13 декабря; 9 (1): 343-373. [PubMed: 30549024]
- 10.
- Сандри М. Распад белка при истощении мышц: роль аутофагии-лизосомы и убиквитин-протеасомы. Int J Biochem Cell Biol. 2013 Октябрь; 45 (10): 2121-9. [Бесплатная статья PMC: PMC3775123] [PubMed: 23665154]
- 11.
- Пуэртоллано Р. mTOR и регуляция лизосом. F1000Prime Rep.2014; 6: 52. [Бесплатная статья PMC: PMC4108950] [PubMed: 25184042]
- 12.
- Доэрти Ф.Дж., Доусон С., Майер Р.Дж.. Убиквитин-протеасомный путь внутриклеточного протеолиза. Очерки Биохимии. 2002; 38: 51-63. [PubMed: 12463161]
- 13.
- Цехановер А. Путь убиквитин-протеасома: гибель белка и жизнь клетки. EMBO J. 15 декабря 1998; 17 (24): 7151-60. [Бесплатная статья PMC: PMC1171061] [PubMed: 9857172]
- 14.
- Rechsteiner M, Rogers SW. Последовательности PEST и регуляция протеолизом.Trends Biochem Sci. 1996 июл; 21 (7): 267-71. [PubMed: 8755249]
- 15.
- Брикс К., Дункхорст А., Майер К., Джорданс С. Катепсины цистеина: клеточная дорожная карта для различных функций. Биохимия. 2008 Февраль; 90 (2): 194-207. [PubMed: 17825974]
- 16.
- Лейст М., Яаттеля М. Запуск апоптоза катепсинами. Смерть клетки отличается. 2001 апр; 8 (4): 324-6. [PubMed: 11550083]
- 17.
- Харрис Р.А., Джоши М., Чжунг Н.Х., Обаяши М. Обзор молекулярных и биохимических основ катаболизма аминокислот с разветвленной цепью.J Nutr. 2005 июн; 135 (6 доп.): 1527S-30S. [PubMed: 15930464]
- 18.
- БЕЛЛАМИ Д. Эндогенные промежуточные продукты цикла лимонной кислоты и аминокислоты митохондрий. Biochem J. 1962, январь; 82: 218-24. [Бесплатная статья PMC: PMC1243435] [PubMed: 13866863]
- 19.
- Айкава Т., Мацутака Х., Ямамото Х., Окуда Т., Исикава Э. Глюконеогенез и метаболизм аминокислот. II. Межорганические отношения и роль глутамина и аланина в метаболизме аминокислот голодных крыс. J Biochem.1973 Ноябрь; 74 (5): 1003-17. [PubMed: 4770365]
- 20.
- Чу Ю.С., Чжан З. Обнаружение убиквитинирования белков. J Vis Exp. 2009 августа 19; (30) [Бесплатная статья PMC: PMC3149903] [PubMed: 19692941]
- 21.
- Löser C, Möllgaard A, Fölsch UR. Фекальная эластаза 1: новый, высокочувствительный и специфический бесконтактный тест на функцию поджелудочной железы. Кишечник. 1996 Октябрь; 39 (4): 580-6. [Бесплатная статья PMC: PMC1383273] [PubMed: 8944569]
- 22.
- Forsmark C, Adams PC. Функциональное тестирование поджелудочной железы — полезно, но недостаточно.Можно J Гастроэнтерол. 2009 августа; 23 (8): 529-30. [Бесплатная статья PMC: PMC2732171] [PubMed: 19668794]
- 23.
- Ooi CY, Durie PR. Мутации гена трансмембранного регулятора проводимости муковисцидоза (CFTR) при панкреатите. J Cyst Fibros. 2012 сентябрь; 11 (5): 355-62. [PubMed: 22658665]
- 24.
- Сомараджу У. Р., Солис-Моя А. Заместительная терапия ферментами поджелудочной железы для людей с муковисцидозом. Кокрановская база данных Syst Rev. 23 ноября 2016 г .; 11: CD008227. [Бесплатная статья PMC: PMC6734127] [PubMed: 27878805]
26.9. Катаболизм белков
Цели обучения
- Чтобы описать, как разлагаются избыточные аминокислоты.
Печень является основным местом метаболизма аминокислот, но другие ткани, такие как почки, тонкий кишечник, мышцы и жировая ткань, принимают в нем участие. Как правило, первым шагом в расщеплении аминокислот является отделение аминогруппы от углеродного скелета, обычно в результате реакции трансаминирования. Углеродные скелеты, образовавшиеся в результате дезаминированных аминокислот, используются для образования либо глюкозы, либо жиров, либо они превращаются в промежуточный продукт метаболизма, который может окисляться циклом лимонной кислоты.Последняя альтернатива, катаболизм аминокислот, более вероятна при низком уровне глюкозы, например, когда человек голодает или голодает.
Трансаминирование
Трансаминирование — это обмен функциональными группами между любой аминокислотой (кроме лизина, пролина и треонина) и α-кетокислотой. Аминогруппа обычно переносится на кетоуглеродный атом пирувата, оксалоацетата или α-кетоглутарата, превращая α-кетокислоту в аланин, аспартат или глутамат соответственно.Реакции трансаминирования катализируются специфическими трансаминазами (также называемыми аминотрансферазами), которым в качестве кофермента требуется пиридоксальфосфат.
Рисунок \ (\ PageIndex {1} \)).
В α-кетокислоте карбонильная или кетогруппа расположена на атоме углерода, смежном с карбоксильной группой кислоты.
Рисунок \ (\ PageIndex {1} \): две реакции трансаминирования. В обеих реакциях конечным акцептором аминогруппы является α-кетоглутарат, а конечным продуктом является глутамат.
Окислительное дезаминирование
При расщеплении аминокислот для получения энергии конечным акцептором α-аминогруппы является α-кетоглутарат, образуя глутамат. Затем глутамат может подвергаться окислительному дезаминированию, при котором он теряет свою аминогруппу в виде иона аммония (NH 4 + ) и снова окисляется до α-кетоглутарата (готового принять другую аминогруппу):
Катаболизм белков — биохимия
Если вы считаете, что контент, доступный через Веб-сайт (как определено в наших Условиях обслуживания), нарушает
или несколько ваших авторских прав, сообщите нам, отправив письменное уведомление («Уведомление о нарушении»), содержащее
в
информацию, описанную ниже, назначенному ниже агенту.Если репетиторы университета предпримут действия в ответ на
ан
Уведомление о нарушении, оно предпримет добросовестную попытку связаться со стороной, которая предоставила такой контент
средствами самого последнего адреса электронной почты, если таковой имеется, предоставленного такой стороной Varsity Tutors.
Ваше Уведомление о нарушении прав может быть отправлено стороне, предоставившей доступ к контенту, или третьим лицам, таким как
в качестве
ChillingEffects.org.
Обратите внимание, что вы будете нести ответственность за ущерб (включая расходы и гонорары адвокатам), если вы существенно
искажать информацию о том, что продукт или действие нарушает ваши авторские права.Таким образом, если вы не уверены, что контент находится
на Веб-сайте или по ссылке с него нарушает ваши авторские права, вам следует сначала обратиться к юристу.
Чтобы отправить уведомление, выполните следующие действия:
Вы должны включить следующее:
Физическая или электронная подпись правообладателя или лица, уполномоченного действовать от их имени;
Идентификация авторских прав, которые, как утверждается, были нарушены;
Описание характера и точного местонахождения контента, который, по вашему мнению, нарушает ваши авторские права, в \
достаточно подробностей, чтобы позволить репетиторам университетских школ найти и точно идентифицировать этот контент; например нам требуется
а
ссылка на конкретный вопрос (а не только на название вопроса), который содержит содержание и описание
к какой конкретной части вопроса — изображению, ссылке, тексту и т. д. — относится ваша жалоба;
Ваше имя, адрес, номер телефона и адрес электронной почты; а также
Ваше заявление: (а) вы добросовестно полагаете, что использование контента, который, по вашему мнению, нарушает
ваши авторские права не разрешены законом, владельцем авторских прав или его агентом; (б) что все
информация, содержащаяся в вашем Уведомлении о нарушении, является точной, и (c) под страхом наказания за лжесвидетельство, что вы
либо владелец авторских прав, либо лицо, уполномоченное действовать от их имени.
Отправьте жалобу нашему уполномоченному агенту по адресу:
Чарльз Кон
Varsity Tutors LLC
101 S. Hanley Rd, Suite 300
St. Louis, MO 63105
Или заполните форму ниже:
Катаболизм белка — обзор
5.23 Деструкторы белка BET
Химеры, нацеленные на протеолиз (PROTAC), включают нацеленные белки для разложения убиквитинлигазами E3. 190 PROTAC dBET6, специфичный для BET-BRD, показал большую эффективность, чем (+) — JQ1, на панели из 39 клеточных линий, происходящих из ряда злокачественных новообразований, и снизил лейкемическую нагрузку на мышиных моделях. Кроме того, было продемонстрировано, что в линиях Т-клеточных лимфобластных лейкозных клеток деградация BRD4 происходит после обработки dBET6, тогда как более низкие концентрации, чем (+) — JQ1, приводят к расщеплению PARP, расщеплению каспазы 3 и остановке клеточного цикла. 189 Эффекты dBET6 были также установлены в клетках глиобластомы, что привело к потере экспрессии белка BET и жизнеспособности клеток, остановке клеточного цикла G2 / M, подавлению набора генов E2F1 и снижению экспрессии MYC. 98 Интересно, что длительное воздействие возрастающих доз (+) — JQ1, приводящее к резистентности, не предотвращало антипролиферативные эффекты dBET6. Наконец, в моделях ксенотрансплантатов мышей dBET6 превзошел ингибиторы BET BRD в снижении опухолевой нагрузки и увеличении продолжительности жизни мышей. 98 Эти результаты демонстрируют, что PROTAC могут быть эффективны в моделях in vitro, и in vivo, , происходящих из разнообразного набора злокачественных тканей, и поэтому представляют собой многообещающую потенциальную терапевтическую стратегию, особенно в случаях, когда возникла устойчивость к ингибиторам BET BRD. .
PROTACS также показал свою эффективность при лимфоме Беркитта и раке простаты. 190, 191 В исследовании с использованием лимфомы Беркитта в качестве модели лечение PROTAC привело к деградации BRD4 и потере cMYC в непрерывных клеточных линиях. Фенотипически клетки, обработанные PROTAC, показали большую потерю жизнеспособности, активацию каспаз и расщепление PARP по сравнению с ингибиторами BET BRD (+) — JQ1 или OXT015. 190 Хотя это обычно так, исследование с использованием dBET1 показало минимальное влияние на развитие клеточного цикла в линиях рака толстой кишки, груди и яичников.Кроме того, несмотря на то, что была повышена способность индуцировать активацию каспазы 3 по сравнению с обработкой (+) — JQ1 в линиях клеток груди и яичников, при сравнении PROTAC (dBET1) с контролем было небольшое различие в окрашивании аннексина V, что указывает на то, что, возможно, существует также был минимальный эффект на индукцию апоптоза. 103 Исследование непрерывных клеточных линий рака простаты также показало успешную индукцию E3-лигазозависимого расщепления белка BET под действием PROTAC и последующее подавление MYC. 191 Обработка приводила к активации путей апоптоза в форме активации каспазы 3/7 и расщепления PARP в течение 24 часов, тогда как лечение ингибитором BET BRD OTX015 не помогло. Интересно, что деградация BET приводит к снижению экспрессии полноразмерной и сплайсированной AR, а также нижестоящей мишени, ERG. Мыши с ксенотрансплантатом, которым вводили деструктор, показали потерю экспрессии BRD4, подавление экспрессии белка MYC и потерю экспрессии AR. 191 PROTAC превзошел контрольные обработки, стандартные химиотерапевтические схемы и OTX015, а некастрированные самцы мышей Nu / Nu переносили лечение ингибиторами BET PROTAC без потери массы тела. Однако в местах инъекций произошло обратимое изменение цвета кожи. 191 Мыши CB17 и SCID не переносили одинаковые схемы лечения. При периодическом лечении одним и тем же PROTAC у мышей CB17 и SCID проявлялись признаки токсичности, включая летаргию, снижение подвижности и сутулость позвоночника, хотя значительной потери веса не наблюдалось. 191 Эти неоднородные данные о токсичности разных линий мышей предполагают, что токсичность может быть проблематичной в будущих клинических исследованиях и требует дальнейшего изучения.
Подобно фармакологическим ингибиторам функции белка BET, механизмы потенциальной резистентности к разрушителям BET также должны быть приняты во внимание и изучены соответствующим образом. В недавнем исследовании устойчивые клетки как к VHL-, так и к цереблон-направленным PROTACS генерировались после воздействия возрастающих доз в течение 4 месяцев.Появились устойчивые клоны в линии клеток рака яичников, которые оставались устойчивыми даже при удалении PROTAC из среды в течение 2 месяцев, что позволяет предположить, что механизм устойчивости включает стабильную генетическую мутацию. Устойчивые клетки показали усиленный рост клеток, потерю расщепления PARP и активацию каспаз после обработки PROTAC. В устойчивых клетках наблюдались минимальные изменения уровней экспрессии белков BRD2, BRD3 и BRD4 после обработки PROTAC, даже когда время и доза увеличивались.Полное секвенирование экзома и секвенирование РНК показали, что мутации, нарушающие функцию компонентов механизма убиквитинирования, CUL2 и CRBN, лежат в основе этой устойчивости. Эти мутации привели к значительному подавлению CUL2 и CRBN, что было подтверждено с помощью количественной ПЦР и вестерн-блоттинга. Эктопическая сверхэкспрессия этих генов привела к восстановлению чувствительности к PROTAC и восстановлению деградации BRD2, BRD3 и BRD4 после лечения. Это исследование показывает, что чувствительность к разрушителям BET зависит от неизменной функции аппарата E3-лигазы, и выявляет потенциальный механизм ускользания, который может угрожать долгосрочной эффективности BET PROTAC при использовании в клиническом контексте.