как принимать, виды и типы аминокислот, полезные свойства
Активный рост мышц спортсмена нельзя представить без дополнительных аминокислот. Они являются строительными частицами белков в организме, что и делает тело спортсмена рельефным и очень привлекательным. Кроме этого немаловажного достоинства, аминокислоты укрепляют мышечную ткань, восстанавливают организм после тяжелых нагрузок и активно положительно влияют на похудение.
Виды аминокислот и их предназначение
По своей сути аминокислоты являются белками, расщепленными на частицы. Когда белок распадается (например, в процессе пищеварения), он образовывает эти питательные вещества.
Все аминокислоты можно разделить на три вида:
- заменимые (которые могут вырабатываться в организме человека) – это аланин, аспаргин, глицин, пролин, глютамин, серин;
- условно заменимые (вырабатываются только в благоприятных условиях или только у взрослых людей) – аргинин, цистеин, тирозин;
- все остальные аминокислоты относятся к категории незаменимых (они не вырабатываются самостоятельно и поступают в организм только с продуктами рациона).
В организме эти нутриенты выполняют множество функций:
- отвечают за создание новых клеток, а также регенерацию мышечных волокон;
- обеспечивают организм дополнительной энергией;
- способствуют нормальному обмену веществ, поддержанию здорового гормонального фона;
- улучшают память, повышают концентрацию внимания, влияют на состояние нервной системы;
- поддерживают иммунную систему;
- подавляют аппетит и способствуют избавлению от лишнего веса;
- играют важную роль при формировании мышечного рельефа;
- улучшают состояние волос, ногтей и кожи.
Если вы занимаетесь спортом, отдельное внимание следует обратить на комплекс ВСАА. Он состоит из трех незаменимых аминокислот – лейцина, изолейцина и валина, препятствующих распаду мышечных волокон (катаболизму). Принимайте этот вид спортпита, чтобы защитить мышцы во время интенсивной тренировки или сразу после нее.
Аминокислоты одинаково важны и для мужчин, и для женщин. Но если представители сильного пола используют их для прироста мышечной массы, то женщины таким образом избавляются от избытков жировой ткани. Таким образом, аминокислотные комплексы практически не требуют времени для усвоения организмом, и быстрее проникают в мышечные клетки, питая и восстанавливая их.
Но если представители сильного пола используют их для прироста мышечной массы, то женщины таким образом избавляются от избытков жировой ткани. Таким образом, аминокислотные комплексы практически не требуют времени для усвоения организмом, и быстрее проникают в мышечные клетки, питая и восстанавливая их.
Типы аминокислотных комплексов
- Свободная форма – это такие аминокислоты, которые моментально всасываются в кровеносные сосуды и не требуют дополнительное переваривание пищеварительной системой. За счет этого они способны очень быстро проникнуть в мышцы и предвидеть мышечный катаболизм.
- Гидролизованная форма – самые быстрые в усвоении организмом, активно питают мышечную ткань и являются основной всех анаболических реакцией.
- BCAA, которые еще называют «мышечными аминокислотами», ведь именно они наиболее положительно влияют на рост мышечной ткани.
- Ди- и трипептидные аминокислоты питают мышцы и активируют анаболические реакции.
Любые аминокислоты отлично комбинируются с другими продуктами спортпита, но далеко не все можно вместе употреблять и тем более смешивать. Для дополнительной консультации лучше обратиться к специалистам нашего магазина спортивного питания Bcaa.
Для дополнительной консультации лучше обратиться к специалистам нашего магазина спортивного питания Bcaa.
Польза аминокислот
- они отлично повышают силы и выносливость на тренировке, что ускоряет набор мышечной массы;
- быстро восстанавливают организм, устраняют боли после тренировки;
- отлично обогащают рацион питания полезными компонентами;
- активно устраняют чувство голода;
- сжигают лишнюю жировую прослойку, ускоряя метаболизм.
Как принимать аминокислотные комплексы?
Правильный прием зависит от вашей цели. Если вы хотите нарастать мышечную массу и выглядеть лучше, идеальное время для употребления аминокислот – это до и после тренировки, после пробуждения утром. Если вам нужно быстро похудеть, продукт стоить принимать чаще. Универсальная дозировка – не менее 5 грамм.
Самые популярные продукты
Ассортимент аминокислот на рынке спортивного питания поражает, но вот процесс выбора качественного и полезного комплекса часто оказывается большой проблемой.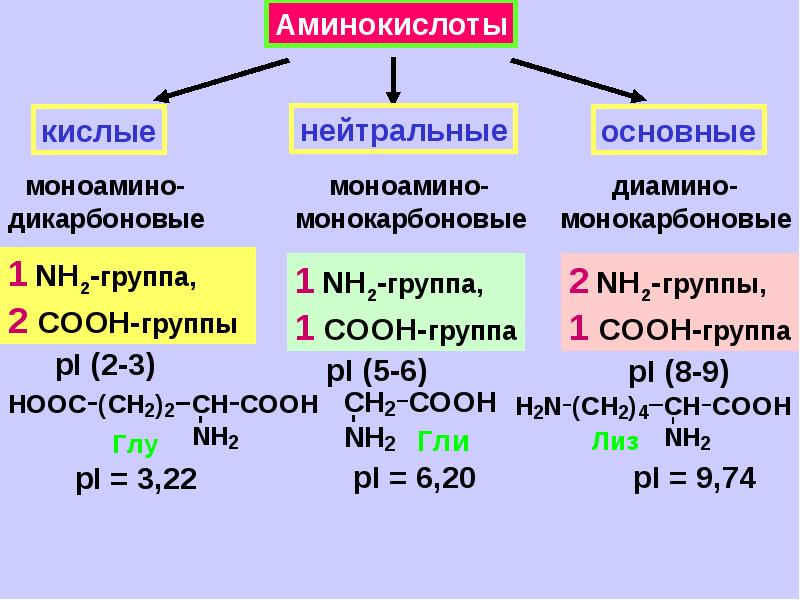 Какие же продукты считаются самыми популярными и завоевали доверие профессионалов? Это такие аминокислоты, как Whey Amino Tabs 2000 и Amino Max Hydrolysate от Maxler, Mega Amino 3200 от бренда BioTech и таблетки Amino 5600 от известного производителя Scitec Nutrition.
Какие же продукты считаются самыми популярными и завоевали доверие профессионалов? Это такие аминокислоты, как Whey Amino Tabs 2000 и Amino Max Hydrolysate от Maxler, Mega Amino 3200 от бренда BioTech и таблетки Amino 5600 от известного производителя Scitec Nutrition.
Другие полезные статьи:
что это такое, полезные свойства и как их правильно принимать
© Yulia Furman — stock.adobe.com
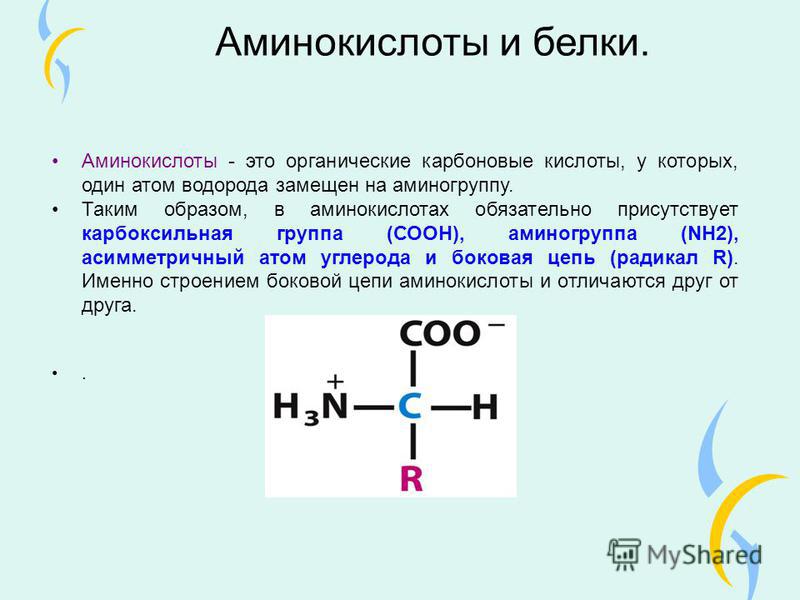
Аминокислоты – органические вещества, состоящие из углеводородного скелета и двух дополнительных групп: аминной и карбоксильной. Последние два радикала обусловливают уникальные свойства аминокислот – они могут проявлять свойства как кислот, так и щелочей: первые – за счет карбоксильной группы, вторые – за счет аминогруппы.
Итак, мы выяснили, что такое аминокислоты с точки зрения биохимии. Теперь рассмотрим их влияние на организм и применение в спорте. Для спортсменов аминокислоты важны своим участием в протеиновом обмене. Именно из отдельных аминокислот строятся протеины для роста мышечной массы нашего тела – мышечная, скелетная, печеночная, соединительная ткани. Помимо этого, некоторые аминокислоты напрямую участвуют в обмене веществ. К примеру, аргинин участвует в орнитиновом цикле мочевины – уникальном механизме обезвреживания аммиака, образующегося в печени в процессе переваривания белков.
Помимо этого, некоторые аминокислоты напрямую участвуют в обмене веществ. К примеру, аргинин участвует в орнитиновом цикле мочевины – уникальном механизме обезвреживания аммиака, образующегося в печени в процессе переваривания белков.
- Из тирозина в коре надпочечников синтезируются катехоламины – адреналин и норадреналин – гормоны, функция которых – поддержание тонуса сердечно сосудистой системы, мгновенная реакция на стрессовую ситуацию.
- Триптофан – предшественник гормона сна – мелатонина, вырабатывающегося в шишковидном теле головного мозга – эпифизе. При недостатке этой аминокислоты в рационе процесс засыпания усложняется, развивается бессонница и ряд других заболеваний, ею обусловленных.
Перечислять можно долго, однако остановимся на аминокислоте, значение которой особенно велико для спортсменов и людей, умеренно занимающихся спортом.
Для чего нужен глютамин
Глютамин – аминокислота, лимитирующая синтез протеина, из которого состоит наша иммунная ткань – лимфатические узлы и отдельные образования лимфоидной ткани. Значение этой системы переоценить трудно: без должного сопротивления инфекциям ни о каком тренировочном процессе говорить не приходится. Тем более, что каждая тренировка – не важно, профессиональная или любительская – это дозированный стресс для организма.
Значение этой системы переоценить трудно: без должного сопротивления инфекциям ни о каком тренировочном процессе говорить не приходится. Тем более, что каждая тренировка – не важно, профессиональная или любительская – это дозированный стресс для организма.
Стресс – необходимое условие, чтобы сдвинуть с места нашу “точку равновесия”, то есть вызвать определенные биохимические и физиологические изменения в организме. Любой стресс – это цепь реакций, мобилизующих тело. В промежуток, характеризующий регресс каскада реакций симпатоадреналовой системы (а именно они и представляют собой стресс), происходит снижение синтеза лимфоидной ткани. По этой причине процесс распада превышает скорость синтеза, а значит, иммунитет ослабевает. Так вот, дополнительный прием глютамина сводит к минимуму этот крайне нежелательный, но неизбежный эффект физической нагрузки
Незаменимые и заменимые аминокислоты
Чтобы понять, для чего нужны незаменимые аминокислоты в спорте, необходимо иметь общие представления о белковом обмене. Потребленные человеком белки на уровне желудочно-кишечного тракта обрабатываются ферментами – веществами, расщепляющими пищу, которую мы употребили.
Потребленные человеком белки на уровне желудочно-кишечного тракта обрабатываются ферментами – веществами, расщепляющими пищу, которую мы употребили.
В частности, белки распадаются сперва до пептидов – отдельных цепочек аминокислот, не имеющих четвертичной пространственной структуры. И уже пептиды распадутся на отдельные аминокислоты. Те, в свою очередь, усваиваются организмом человека. Это значит, что аминокислоты всасываются в кровь и только с этого этапа могут быть использованы в качестве продуктов для синтеза белка тела.
Забегая вперед скажем, что прием отдельных аминокислот в спорте сокращает этот этап – отдельные аминокислоты будут сразу же всасываться в кровь и процессы синтеза, а также биологический эффект аминокислот наступят быстрее.
Всего существует двадцать аминокислот. Чтобы процесс синтеза белка в теле человека стал возможным в принципе, в рационе человека должен присутствовать полный спектр – все 20 соединений.
Незаменимые
Вот с этого момента и появляется понятие незаменимости.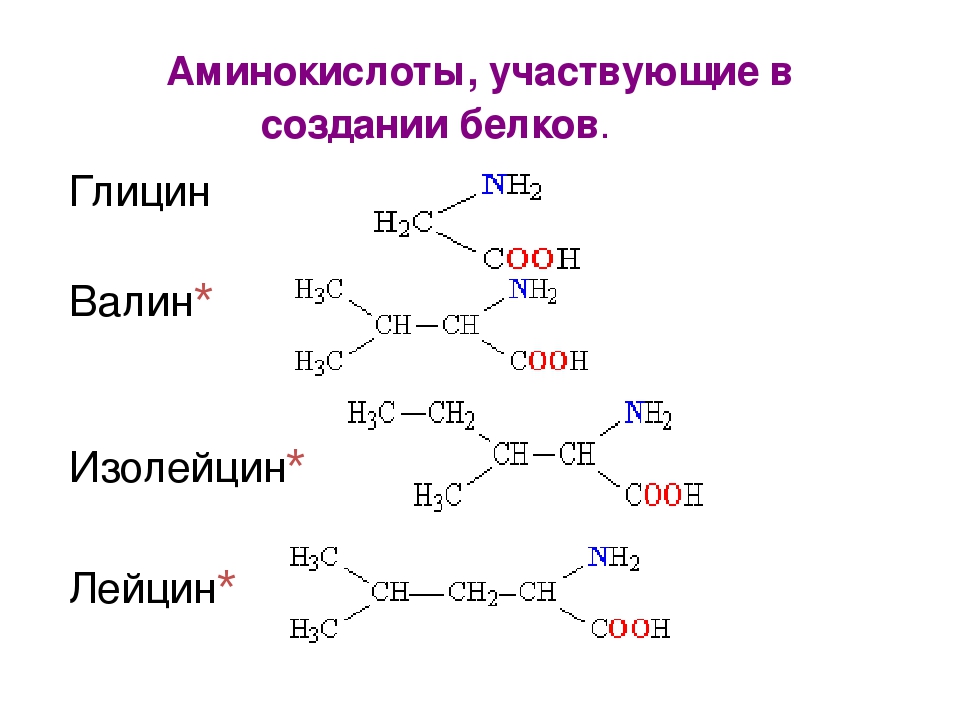 К незаменимым аминокислотам относятся те, которые наше тело не способно синтезировать самостоятельно из других аминокислот. А это значит, что появится им, кроме как из продуктов питания, неоткуда. Таких аминокислот насчитывается 8 плюс 2 частично-заменимые.
К незаменимым аминокислотам относятся те, которые наше тело не способно синтезировать самостоятельно из других аминокислот. А это значит, что появится им, кроме как из продуктов питания, неоткуда. Таких аминокислот насчитывается 8 плюс 2 частично-заменимые.
Рассмотрим в таблице, в каких продуктах содержится каждая незаменимая аминокислота и какова ее роль в организме человека:
| Название | В каких продуктах содержится | Роль в организме |
| Лейцин | Орехи, овес, рыба, яйца, курица, чечевица | Снижает содержание сахара в крови |
| Изолейцин | Нут, чечевица, кешью, мясо, соя, рыба, яйца, печень, миндаль, мясо | Восстанавливает мышечную ткань |
| Лизин | Амарант, пшеница, рыба, мясо, большинство молочных продуктов | Принимает участие в усвоении кальция |
| Валин | Арахис, грибы, мясо, бобовые, молочные продукты, многие зерновые | Принимает участие в обменных процессах азота |
| Фенилаланин | Говядина, орехи, творог, молоко, рыба, яйца, разные бобовые | Улучшение памяти |
| Треонин | Яйца, орехи, бобы, молочные продукты | Синтезирует коллаген |
| Метионин | Фасоль, соя, яйца, мясо, рыба, бобовые, чечевица | Принимает участие в защите от радиации |
| Триптофан | Кунжут, овес, бобовые, арахис, кедровые орехи, большинство молочных продуктов, курица, индейка, мясо, рыба, сушенные финики | Улучшает и делает сон глубже |
| Гистидин (частично-заменимая) | Чечевица, соевые бобы, арахис, тунец, лосось, говяжье и куриное филе, свиная вырезка | Принимает участие в противовоспалительных реакциях |
| Аргинин (частично-заменимая) | Йогурт, кунжут, семена тыквы, швейцарский сыр, говядина, свинина, арахис | Способствует росту и восстановлению тканей организма |
В достаточном количестве аминокислоты содержатся в животных источниках белка – рыбе, мясе, птице.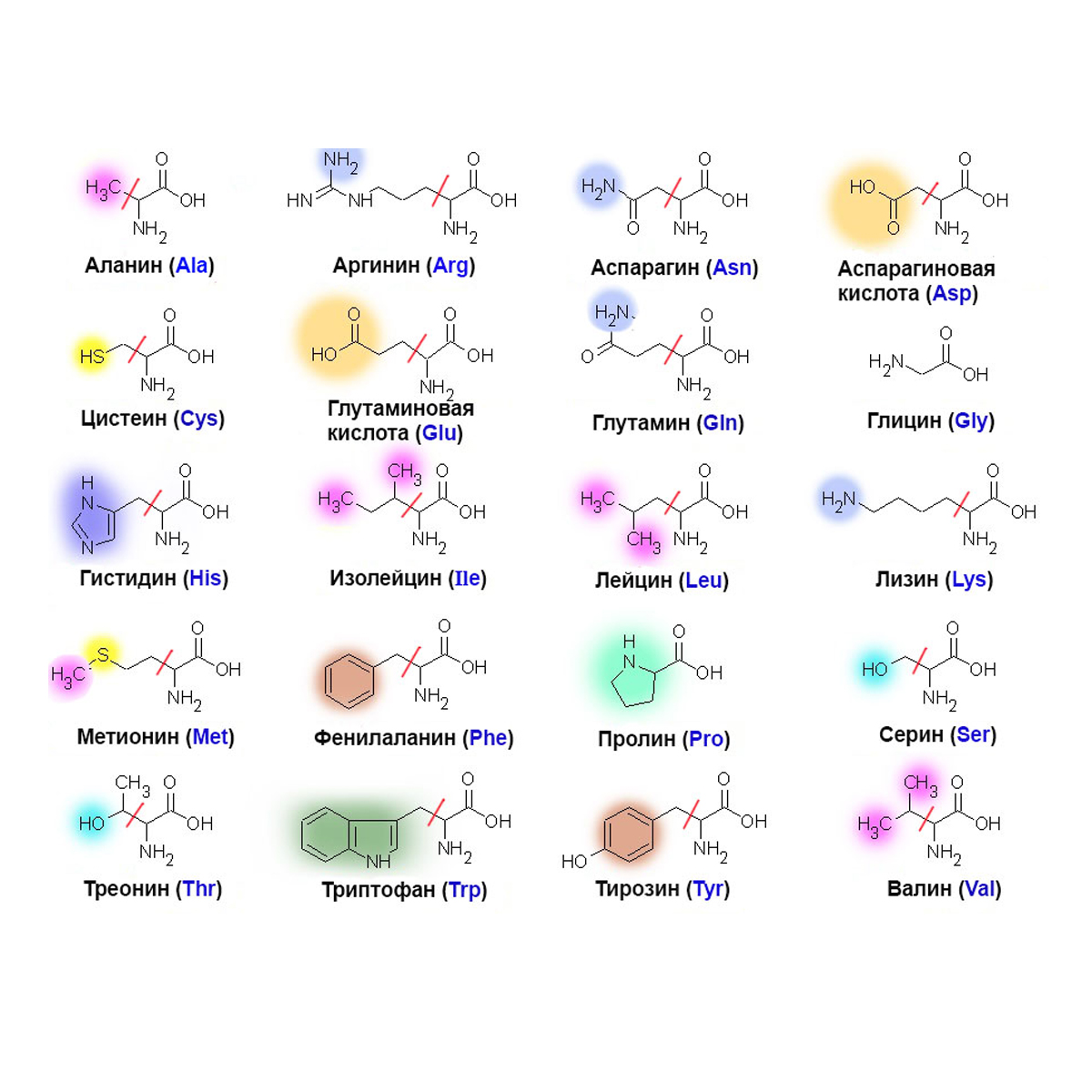 При отсутствии таковых в рационе весьма целесообразен прием недостающих аминокислот в качестве добавок спортивного питания, что особенно актуально для спортсменов-вегетарианцев.
При отсутствии таковых в рационе весьма целесообразен прием недостающих аминокислот в качестве добавок спортивного питания, что особенно актуально для спортсменов-вегетарианцев.
Основное внимание последним стоит обратить на такие добавки, как ВСАА – смесь лейцина, валина и изолейцина. Именно по этим аминокислотам возможна “просадка” в рационе, не содержащем животных источников белка. Для спортсмена (как профессионала, так и любителя) это абсолютно не допустимо, так как в долгосрочной перспективе приведет к катаболизму со стороны внутренних органов и к заболеваниям последних. В первую очередь страдает от недостатка аминокислот печень.
© conejota — stock.adobe.com
Заменимые
Заменимые аминокислоты и их роль рассмотрим в таблице ниже:
| Название | Роль в организме |
| Аланин | Принимает участие в глюконеогенезе печени |
| Пролин | Отвечает за составление прочной структуры коллагена |
| Левокарнитин | Поддерживает кофермент А |
| Тирозин | Отвечает за ферментативную активность |
| Серин | Отвечает за построение природных белков |
| Глютамин | Синтезирует протеины мышц |
| Глицин | Снижает напряжение т уменьшает агрессивность |
| Цистеин | Положительно влияет на текстуру и состояние кожи |
| Таурин | Оказывает метаболическое действие |
| Орнитин | Принимает участие в биосинтезе мочевины |
Вред, побочные эффекты, противопоказания
Если у вас есть заболевания, характеризующиеся непереносимостью одной из аминокислот, вы об этом знаете с рождения, так же, как и ваши родители. Этой аминокислоты нужно избегать и дальше. Если же этого нет, говорить о вреде и противопоказаниях добавок нет смысла, поскольку это полностью натуральные вещества.
Этой аминокислоты нужно избегать и дальше. Если же этого нет, говорить о вреде и противопоказаниях добавок нет смысла, поскольку это полностью натуральные вещества.
Аминокислоты – составляющая часть белка, белок – привычная часть рациона человека. Все то, что продается в магазинах спортивного питания – не является фармакологическими препаратами! Только дилетанты могут говорить о каком-то вреде и противопоказаниях. По той же причине нет смысла рассматривать такое понятие, как побочные эффекты аминокислот – при умеренному потреблении никаких негативных реакций быть не может.
Трезво подходите к своему рациону и спортивным тренировкам! Будьте здоровы!
Оцените материал
Научный консультант проекта.
Физиолог (биологический факультет СПБГУ, бакалавриат).
Биохимик (биологический факультет СПБГУ, магистратура).
Инструктор по хатха-йоге (Институт управления развитием человеческих ресурсов, проект GENERATION YOGA).
Научный сотрудник (2013-2015 НИИ акушерства, гинекологии и репродуктологии им. Отта, работа с маркерами женского бесплодия, анализ биологических образцов; 2015-2017 НИИ особо чистых биопрепаратов, разработка лекарственных средств)
Отта, работа с маркерами женского бесплодия, анализ биологических образцов; 2015-2017 НИИ особо чистых биопрепаратов, разработка лекарственных средств)
Автор и научный консультант сайтов по тематике ЗОЖ и науке (в области продления жизни)
C 2019 года научный консультант проекта Cross.Expert.
Редакция cross.expert
Цены на аминокислоты выросли почти в два раза
Аминокислоты – это кормовые добавки, напрямую влияющие на образование тканей в организме животного. При недостатке этих веществ останавливается синтез белка, в результате чего животное может потерять мышечную массу. Аминокислоты (треонин, валин, метионин, триптофан, лизин) применяют в кормлении сельскохозяйственных животных, большая часть этой продукции – импортная.
- «Например, цена на лизин из Китая на условиях поставки DAF (поставка до российской границы без последующей таможенной очистки. – Прим. ред.) выросла с 1,1 доллара за килограмм в октябре – ноябре прошлого года до 1,7–1,75 доллара за килограмм в январе 2021», – рассказал Сергей Михнюк.

Цена на треонин возросла с 0,98 до 1,54 доллара, на триптофан – с 7,5 до 9,8 доллара, на валин – с 2,35 до 4,1.
Основное сырье для производства аминокислот – кукуруза. Цены на нее еще в январе 2020 года достигли трехлетнего максимума. Связано это с неурожаем в США, Аргентине, Бразилии и на Украине – это основные поставщики сырья в Китай. К тому же в КНР выросло потребление кукурузы, прежде всего со стороны предприятий свиноводческого комплекса. Причем большая часть собственного урожая этой культуры в самом Китае была потеряна из-за тайфуна.
Вторая причина роста цен на аминокислоты – новая волна коронавируса в Китае, которая хотя и слабее предыдущей, но заставляет власти даже при единичных случаях выявления инфекции закрывать границы городов и провинций, приостанавливать отгрузки в портах.
- «Новые вспышки коронавирусной инфекции сейчас сильнее всего затронули северо-восточные провинции Китая, где сосредоточено наибольшее количество заводов по глубокой переработке кукурузы и производству аминокислот из этого сырья», – отметил Михнюк, добавив, что приостановка деятельности этих предприятий приводит к возникновению дефицита на рынке и увеличению цен.

Дополнительные сложности – логистические. Из-за рисков распространения коронавируса железная дорога Китая сократила число разрешений на отправку экспортных грузов, в особенности тех, где нужна ручная перегрузка, что привело к срыву ранее оговоренных поставок, в том числе аминокислот. К тому же в конце 2020 года в Китае сложился дефицит контейнеров для отправки грузов на экспорт. Отправленные ранее контейнеры застряли в Европе из-за новых ограничений, связанных с пандемией. Это привело к резкому росту цен на транспортные услуги, что отразилось и на стоимости перевозимого товара.
В Национальном кормовом союзе прогнозируют, что цены на аминокислоты стабилизируются только к лету.
- «Цена должна стабилизироваться в связи с новым урожаем, а также потому, что заработает большее количество предприятий в Китае, возрастет конкуренция, будет создан рынок покупателя, а не продавца», – добавил Михнюк.
Подпишитесь на нас в Telegram, чтобы читать новости сразу, как только они появляются на сайте.
Почему после сытного обеда хочется спать?
- Клаудиа Хаммонд
- BBC Future
Автор фото, Thinkstock
Существует мнение, что некоторые продукты питания вызывают сонливость. Обозреватель BBC Future выяснила, имеет ли эта теория научную основу.
Считается, что наибольшую сонливость у человека вызывает жареная индейка. Если вы позволяете себе лишнего за праздничным столом, эти ощущения вам должны быть знакомы. Однако справедливы ли подобные предположения?
Дело в том, что мясо индейки содержит вещество L-триптофан. В составе некоторых продуктов, однако, этой аминокислоты может быть значительно больше. В почках морского льва, например, или в других более повседневных продуктах питания: в яичном белке, треске или свином стейке.
Сложно придумать рецепт блюда, который соединил бы все эти составляещие. Однако, даже если вам это удастся, эффект от этого яства вряд ли был бы усыпляющим.
Однако, даже если вам это удастся, эффект от этого яства вряд ли был бы усыпляющим.
Мысль о том, что некоторые блюда могут вызвать сонливость, очевидно, не совсем верна. Чтобы понять почему, нужно разобраться в том, как тело и мозг поглощают питательные вещества.
L-триптофан является незаменимой амино-кислотой, которую наш организм не умеет производить самостоятельно. Именно поэтому вещество добывается из питания, а в дальнейшем служит для выработки серотонина.
Серотонин, как известно, является «гормоном счастья», но мало кто знает, что он также способствует выработке дрозофила — «гормона сна», который и приводит к повышенной сонливости у людей.
Это позволяет предположить, что прием триптофана поможет в борьбе с бессонницей, и такие его фармакологические качества, на самом деле, подтверждают результаты исследований, проведенных до 1986 года. В последующие годы свойства триптофана почти не изучались, однако один эксперимент 2002 года показал, что снижение уровня триптофана ухудшает сон.
Впрочем, свойства триптофана в фармакологических исследованиях, безусловно, отличаются от эффекта, который он производит, попадая в организм вместе с пищей, ведь в продуктах питания он взаимодействует с другими веществами.
Автор фото, Thinkstock
Підпис до фото,
Считается, что наибольшую сонливость у человека вызывает жареная индейка
В состав белка, содержащегося в продуктах питания, входят большие нейтральные аминокислоты (LNAA), которые «конкурируют» с триптофаном за прохождение через гематоэнцефалический барьер и фактически препятствуют попаданию в мозг большей части триптофана.
Чтобы триптофан, содержащийся в пище, мог превратиться в серотонин, вы не только должны употребить его на голодный желудок, где он не будет «конкурировать» с другими питательными элементами, но и само блюдо также не должно содержать другие аминокислоты. Однако это невозможно.
Впрочем, есть одно условие, при котором триптофан может проникнуть в мозг. В одном канадском исследовании участникам, страдающим от бессонницы, предлагали батончики с семенами мускатной тыквы с высоким содержанием триптофана.
В одном канадском исследовании участникам, страдающим от бессонницы, предлагали батончики с семенами мускатной тыквы с высоким содержанием триптофана.
Когда в батончики добавляли сахар глюкозы, сон у участников исследования улучшался (в эксперименте участвовала пара однояйцевых близнецов).
Причина, по которой глюкоза изменила результаты исследования, заключается в том, что сладкие углеводы вызывают выделение инсулина. Он стимулирует всасывание других аминокислот в тканях, тогда как путь к мозгу остается свободным для триптофана.
Впрочем, даже если вы подберете правильную комбинацию продуктов питания, вы совсем не обязательно достигните нужного эффекта. В 40-ка контролируемых исследованиях, проведенных до 1982 года, добавка L-триптофана действительно повышала у участников чувство сонливости и способствовала быстрому засыпанию.
Автор фото, Thinkstock
Підпис до фото,
Триптофан в виде лекарственных препаратов может способствовать хорошему сну, однако в составе продуктов питания он действует иначе
При этом препарат не влиял на количество часов сна, особенно если человек не страдал от бессонницы. А если нарушения сна были значительными, триптофан не помогал.
А если нарушения сна были значительными, триптофан не помогал.
Итак, возможно, дело не в празничной индейке, а в сочетании продуктов.
Хотя причин для ощущения усталости может быть множество: аврал на работе перед праздниками, приготовление ужина для гостей и чересчур активный отдых.
Если вы хорошо поели, да еще и запили алкоголем, нет ничего удивительного в том, что после обеда возникает неудержимое желание вздремнуть, даже если L-триптофан так и не добрался до вашего мозга.
Прочитать оригинал этой статьи на английском языке вы можете на сайтеBBC Future.
Аминокислоты, без которых нам не жить
Аминокислоты называют «строительным материалом» при синтезе в организме человека целого ряда белков. И любой белок – это цепочка из аминокислот, которые в определённой последовательности соединены между собой. При отсутствии хотя бы одной аминокислоты происходит сбой.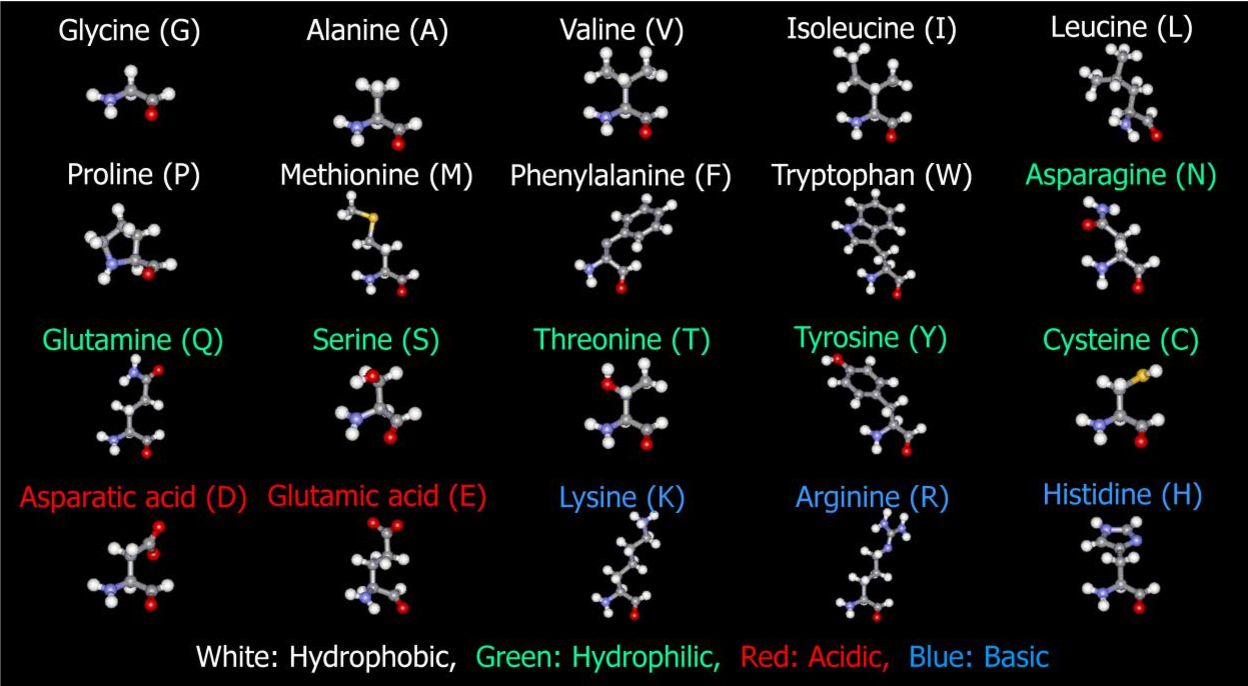
Из двадцати известных аминокислот, восемь являются незаменимыми. То есть сам организм синтезировать их не может, поэтому должен получать их вместе с пищей. Если же он их не получает, то нарушается работа нервной системы, водно-солевой обмен и многие другие функции в организме.
К незаменимым кислотам относятся:
Валин, который с лейцином и изолейцином участвует в синтезе тканей тела и стимулирует их рост, все трое они служат источником энергии в мышечных клетках.
Валин отвечает за мышечную координацию, понижает чувствительность организма к жаре, холоду и боли, поддерживает уровень гормона «счастья» – серотонина.
Содержится: в мясе, грибах, бобовых, зерновых, арахисе и молочных продуктах.
Лейцин необходим для активизации умственной деятельности и хорошей памяти, он защищает мышечные волокна от повреждений, восстанавливает кожные покровы, мышцы и кости, стимулирует гормон роста и снижает уровень сахара в крови. Содержится в нежирном мясе, печени, рыбе, твороге, молоке, натуральном йогурте, кефире, гречихе, чечевице, овсе, неочищенном рисе, люцерне.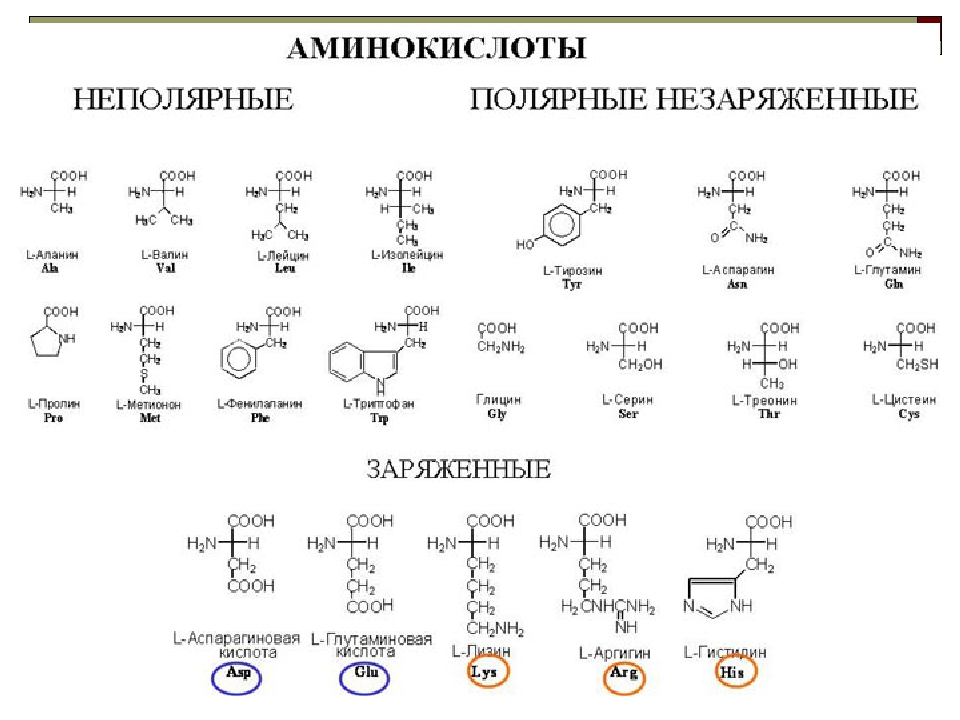
Изолейцин так же отвечает за уровень сахара в крови, обеспечивает энергией все к летки и повышает выносливость. Содержится: в мясе птицы, печени, рыбе, яйцах, бобовых, чечевице, во ржи, миндале, кешью, сое, семечках.
Лизин отвечает за работу мозга и ясное мышление до глубокой старости, поддерживает энергию и следит за здоровьем сердца, оказывает сопротивление вирусам, способствует усвоению кальция, восстанавливает ткани, формируя коллаген. Содержится в мясе птицы, рыбе, молочных продуктах, бобовых, кукурузе, орехах, семечках, какао, в горьком шоколаде.
Метионин снижает содержание холестерина и улучшает работу печени, препятствует развитию депрессии.
Содержится в рыбе, желтке яиц, бобовых, зелёном горошке, гречихе, капусте, моркови, в апельсинах, арбузах и дыне.
Треонин – препятствует ожирению печени, участвует жировом и белковом обмене, повышает иммунитет. Содержится: в яйцах, молочных продуктах, бобовых, орехах.
Триптофан нормализует психическое состояние, отвечает за нормальное функционирование мозга и замедляет общее старение организма. Кроме того снижает аппетит и способствует повышению выработки гормона роста. Содержится в мясе птицы, рыбе, молоке, твороге, бобовых, орехах, кунжуте, бананах, в винограде и таких сухофруктах, как курага, инжир, финики.
Кроме того снижает аппетит и способствует повышению выработки гормона роста. Содержится в мясе птицы, рыбе, молоке, твороге, бобовых, орехах, кунжуте, бананах, в винограде и таких сухофруктах, как курага, инжир, финики.
Фенилаланин снижает аппетит и повышает настроение, а так же отвечает за быстроту реакций и уменьшает чувствительность организма к боли. Содержится он в говядине, курином мясе, рыбе, яйцах, твороге, молоке, сметане.
Получается, что для того, чтобы обеспечить организм незаменимыми аминокислотами, нужно потратить не так уж много денег.
Бобовые, злаки, семечки, овощи стоят недорого, мясо птицы, молочные продукты, яйца тоже доступны, есть недорогие сорта рыбы. Дорогими можно назвать только шоколад, орехи и некоторые сухофрукты. Но и их в небольшом количестве можно себе позволить – те же финики и курагу в виде перекуса на работе вместо тоже недешёвых конфет.
Для того чтобы сохранить здоровье, врачи советуют не забывать о крестоцветных – всех видах капусты, о цитрусовых и листовых огородных травах. А так же об оливковом и подсолнечном масле.
А так же об оливковом и подсолнечном масле.
Всё это вместе снизит риск развития сердечно-сосудистых заболеваний, инсультов, помешает образованию склеротических бляшек и развитию слабоумия в старости.
границ | Связи между метаболизмом аминокислот в растениях: лизин в качестве примера
Введение
Аминокислоты играют ряд жизненно важных ролей в центральном метаболизме растений. Незаменимые аминокислоты (EAA), особенно лизин и метионин, не могут быть синтезированы людьми или животными, и поэтому их нужно получать через источников пищи. Однако дисбаланс в питании растений часто вызван отсутствием определенных EAA (Galili et al., 2016). Аминокислоты также действуют как промежуточные продукты конечных метаболитов в определенных метаболических путях, а также участвуют в регуляции множества метаболических путей и других физиологических и биохимических путей, тем самым влияя на многочисленные физиологические процессы в растениях (Amir et al., 2018; Ян и др. , 2018).
, 2018).
EAA в растениях широко изучены (Galili et al., 2016). Генная инженерия обеспечивает эффективный способ повышения уровня EAA в растениях, с многочисленными успешными результатами, хотя и с дополнительными неожиданными эффектами (Galili et al., 2016). Считается, что из всех живых организмов высшие растения обладают наивысшей сложностью с точки зрения метаболических сетей, что делает метаболическую инженерию композиционных характеристик особенно сложной задачей (Yang et al., 2018). В этом обзоре представлен обзор связей между метаболизмом аминокислот в растениях, с особым вниманием к новым открытиям, сделанным в последние годы. Углубленный анализ метаболических связей между аминокислотами углубит наше понимание генетической основы геномов и метаболомов растений, предоставив новую информацию для селекции культур с высоким содержанием EAA.
Метаболический поток аминокислот в растениях
Исследования показали, что регулирование метаболизма определенной аминокислоты влияет на уровень других аминокислот. Это в основном связано с биосинтезом и катаболизмом аминокислот, происходящих из одного и того же метаболического ствола и / или тесно связанных с другими метаболическими путями, действующих как синтетический субстрат или промежуточный продукт (Long et al., 2013; Song et al., 2013) .
Это в основном связано с биосинтезом и катаболизмом аминокислот, происходящих из одного и того же метаболического ствола и / или тесно связанных с другими метаболическими путями, действующих как синтетический субстрат или промежуточный продукт (Long et al., 2013; Song et al., 2013) .
Путь семейства аспартатов приводит к четырем ключевым EAA, лизину, метионину, треонину и изолейцину, но также прочно связан с гомосерином, глутаматом, глицином и пролином (рис. 1A). Разработка метаболизма лизина имеет тенденцию изменять другие аминокислоты в семействе аспартатов, даже те, которые получены другими путями, что свидетельствует о тесной регуляторной сети в биосинтезе свободных аминокислот (Yang et al., 2018). Подобные результаты были обнаружены при регуляции метаболизма метионина (Song et al., 2013). Интересно, что высокие уровни триптофана были связаны с растениями с высоким содержанием лизина (Yang et al., 2018; Wang et al., 2019), в то время как Yang et al. (2018) показали, что накопление лизина индуцирует синтез и метаболизм триптофана в трансгенном рисе с высоким содержанием свободного лизина ( Oryza sativa L. ). Однако в нескольких отчетах задокументирована взаимосвязь между метаболизмом различных семейств аминокислот, что подчеркивает необходимость дальнейших исследований.
). Однако в нескольких отчетах задокументирована взаимосвязь между метаболизмом различных семейств аминокислот, что подчеркивает необходимость дальнейших исследований.
Рисунок 1 Метаболические связи между аминокислотами, производными от аспартата в растениях. (A) Аминокислоты, полученные из аспартата (выделены оранжевым), метаболизм в растениях. (B) Краткое описание метаболизма лизина и связанных метаболических путей (зеленая рамка). (C) Аминокислоты, полученные из аспартата, связаны метаболическими путями (зеленый прямоугольник).
Метаболические связи между лизином и другими метаболитами в растениях
Метаболические пути лизина хорошо изучены, и усиленное накопление лизина было достигнуто с помощью метаболической инженерии .Тем не менее, потенциальные связи с фортификацией лизина с помощью метаболической инженерии еще предстоит изучить.
Эффекты обогащения лизином в растениях
Были предприняты попытки увеличить содержание лизина путем изменения метаболизма лизина или экспрессии белка с высоким содержанием лизина в растениях (Sun and Liu, 2004). Однако было обнаружено, что прорастание семян Arabidopsis с высоким содержанием лизина [ Arabidopsis thaliana (L.) Heynh.] Значительно замедлено и снижено (Zhu and Galili, 2004).Более того, у сои [ Glycine max (Linn.) Merr.] Было обнаружено, что высокая специфическая для семян экспрессия нечувствительных к обратной связи генов, кодирующих дигидродипиколинатсинтазу (DHPS) и аспартаткиназу (AK), приводит к морщинистым семенам и низкой скорости прорастания. (Falco et al., 1995), тогда как снижение урожайности зерна наблюдалось на культурах с высоким содержанием лизина (Liu et al., 2016). Исследования также показывают, что лизин влияет на синтез крахмала и / или развитие эндосперма у кукурузы ( Zea mays Linn.) И риса посредством биофортификации (Jia et al., 2013; Лю и др., 2016). Фенотип обострения меловости также был обнаружен в эндосперме семян трансгенных риса и кукурузы (Wong et al., 2015). Недавно мы создали трансгенный рис с повышенным содержанием свободного лизина, обнаружив небольшое снижение содержания изолейцина и повышение других EAA, со значительным увеличением содержания триптофана (Yang et al.
Однако было обнаружено, что прорастание семян Arabidopsis с высоким содержанием лизина [ Arabidopsis thaliana (L.) Heynh.] Значительно замедлено и снижено (Zhu and Galili, 2004).Более того, у сои [ Glycine max (Linn.) Merr.] Было обнаружено, что высокая специфическая для семян экспрессия нечувствительных к обратной связи генов, кодирующих дигидродипиколинатсинтазу (DHPS) и аспартаткиназу (AK), приводит к морщинистым семенам и низкой скорости прорастания. (Falco et al., 1995), тогда как снижение урожайности зерна наблюдалось на культурах с высоким содержанием лизина (Liu et al., 2016). Исследования также показывают, что лизин влияет на синтез крахмала и / или развитие эндосперма у кукурузы ( Zea mays Linn.) И риса посредством биофортификации (Jia et al., 2013; Лю и др., 2016). Фенотип обострения меловости также был обнаружен в эндосперме семян трансгенных риса и кукурузы (Wong et al., 2015). Недавно мы создали трансгенный рис с повышенным содержанием свободного лизина, обнаружив небольшое снижение содержания изолейцина и повышение других EAA, со значительным увеличением содержания триптофана (Yang et al. , 2016). Тем не менее, темно-коричневый эндосперм, связанный с фенотипом с высоким содержанием свободного лизина, также был обнаружен в сконструированных зрелых семенах (Yang et al., 2018). Приведенные выше данные свидетельствуют о том, что чрезмерное увеличение содержания лизина также влияет на другие метаболические пути, тем самым влияя на рост и развитие целевого растения. Однако эти эффекты также различаются у разных видов, что позволяет предположить, что различия в метаболическом потоке и связи с метаболизмом лизина также существуют.
, 2016). Тем не менее, темно-коричневый эндосперм, связанный с фенотипом с высоким содержанием свободного лизина, также был обнаружен в сконструированных зрелых семенах (Yang et al., 2018). Приведенные выше данные свидетельствуют о том, что чрезмерное увеличение содержания лизина также влияет на другие метаболические пути, тем самым влияя на рост и развитие целевого растения. Однако эти эффекты также различаются у разных видов, что позволяет предположить, что различия в метаболическом потоке и связи с метаболизмом лизина также существуют.
Связи между лизином и энергетическим метаболизмом
В дополнение к классической роли аспартатных кислот в синтезе внутриклеточных белков, в случае дефицита энергии, катаболизм лизина и изолейцина перемещается непосредственно в цикл трикарбоновой кислоты (TCA), в то время как треонин и метионин превращаются в изолейцин (рис. 1A) (Wang et al., 2018). Негативный эффект на прорастание семян наблюдался также у трансгенного Arabidopsis с высоким содержанием свободного лизина, экспрессирующего ген Escherichia coli dhps в мутанте lkr (Angelovici et al. , 2011). DHPS и редуктаза лизинкетоглутаровой кислоты (LKR) представляют собой два ключевых фермента, участвующих в биосинтезе и катаболизме лизина, соответственно. Между тем, результаты анализа метаболома и транскриптома показали значительное увеличение метаболитов, таких как цитрат, сукцинат, малат и фумарат, которые участвуют в цикле TCA, и транскрипта гена, кодирующего 2-оксоизовалератдегидрогеназу, который связан с изолейцином. катаболизм в цикле TCA в проросших семенах с высоким уровнем свободного лизина, потенциально влияющий на энергетический метаболизм в проросших семенах и последующее укоренение проростков (Angelovici et al., 2011). Кроме того, уровень глицина также увеличивался у растений с высоким содержанием лизина, и было обнаружено, что он является ключевым промежуточным метаболитом во время фотодыхания (Schulze et al., 2016). Более того, анализ транскриптома также выявил, что экспрессия генов, связанных с фотосинтезом и фотодыханием, ослаблялась во время прорастания семян Arabidopsis с высоким содержанием лизина (Angelovici et al.
, 2011). DHPS и редуктаза лизинкетоглутаровой кислоты (LKR) представляют собой два ключевых фермента, участвующих в биосинтезе и катаболизме лизина, соответственно. Между тем, результаты анализа метаболома и транскриптома показали значительное увеличение метаболитов, таких как цитрат, сукцинат, малат и фумарат, которые участвуют в цикле TCA, и транскрипта гена, кодирующего 2-оксоизовалератдегидрогеназу, который связан с изолейцином. катаболизм в цикле TCA в проросших семенах с высоким уровнем свободного лизина, потенциально влияющий на энергетический метаболизм в проросших семенах и последующее укоренение проростков (Angelovici et al., 2011). Кроме того, уровень глицина также увеличивался у растений с высоким содержанием лизина, и было обнаружено, что он является ключевым промежуточным метаболитом во время фотодыхания (Schulze et al., 2016). Более того, анализ транскриптома также выявил, что экспрессия генов, связанных с фотосинтезом и фотодыханием, ослаблялась во время прорастания семян Arabidopsis с высоким содержанием лизина (Angelovici et al. , 2009; Angelovici et al., 2011).
, 2009; Angelovici et al., 2011).
Связь между метаболизмом лизина и реакцией растений на стресс
Метаболизм лизина участвует в реакции растений на стресс в различных формах.Он в основном катаболизируется посредством сахаропинового пути, который, как было показано, играет роль в реакциях на абиотический и биотический стресс (рисунки 1B, C) (Kiyota et al., 2015; Bernsdorff et al., 2016; Arruda and Barreto, 2020) . Более того, при солевом и осмотическом стрессе было обнаружено усиление экспрессии LKR / SDH , в то время как последующий метаболит пипеколат усиливался (Kiyota et al., 2015). Кроме того, было обнаружено, что пипеколат значительно увеличивается у Arabidopsis, инфицированного патогенными бактериями, играя роль в защитных реакциях растений (Bernsdorff et al., 2016).
Увеличение пролина также индуцируется глутаматом сахаропинового пути и в ответ на осмотический и солевой стресс (Batista-Silva et al., 2019). Когда ткани или клетки подвергаются осмотическому стрессу, α-аминоадипическая полуальдегиддегидрогеназа (AASADH), третий фермент в пути сахаропина, значительно активируется, что позволяет предположить, что AASADH также связан с реакцией на осмотический стресс (Brocker et al. , 2010 ). Однако у кукурузы сахаропиновый путь индуцируется экзогенным лизином и подавляется солевым стрессом, тогда как синтез пролина и пипеколата значительно подавляется лизином.В то время как AASADH накапливается в тканях при солевом, осмотическом и окислительном стрессе, фермент LKR / SDH не продуцируется (Kiyota et al., 2015). Между тем, в развитии семян пролин, как полагают, играет важную роль в ответах на абиотический стресс у кукурузы, подобно роли пипеколата у Arabidopsis и канолы ( Brassica napus L.) (Kiyota et al., 2015). Помимо этого, пролин выполняет множество функций при других биотических и абиотических стрессах, передаче сигналов, экспрессии генов, регуляции ROS, окислительно-восстановительной адаптации и развитии растений (Trovato et al., 2019) (Рисунок 1С). Недавно мы предположили, что метаболизм лизина индуцирует сигнальный путь жасмоната и метаболизм триптофана во время стрессовых реакций, в отличие от основной роли пипеколата, пролина и AASADH у других видов растений (Yang et al.
, 2010 ). Однако у кукурузы сахаропиновый путь индуцируется экзогенным лизином и подавляется солевым стрессом, тогда как синтез пролина и пипеколата значительно подавляется лизином.В то время как AASADH накапливается в тканях при солевом, осмотическом и окислительном стрессе, фермент LKR / SDH не продуцируется (Kiyota et al., 2015). Между тем, в развитии семян пролин, как полагают, играет важную роль в ответах на абиотический стресс у кукурузы, подобно роли пипеколата у Arabidopsis и канолы ( Brassica napus L.) (Kiyota et al., 2015). Помимо этого, пролин выполняет множество функций при других биотических и абиотических стрессах, передаче сигналов, экспрессии генов, регуляции ROS, окислительно-восстановительной адаптации и развитии растений (Trovato et al., 2019) (Рисунок 1С). Недавно мы предположили, что метаболизм лизина индуцирует сигнальный путь жасмоната и метаболизм триптофана во время стрессовых реакций, в отличие от основной роли пипеколата, пролина и AASADH у других видов растений (Yang et al. , 2018). Вкратце, хотя катаболизм лизина участвует в реакции растений на абиотический и биотический стресс, существуют очевидные различия между видами растений в соответствии с различными реакциями на стресс однодольных и двудольных растений.
, 2018). Вкратце, хотя катаболизм лизина участвует в реакции растений на абиотический и биотический стресс, существуют очевидные различия между видами растений в соответствии с различными реакциями на стресс однодольных и двудольных растений.
Между тем, изолейцин, аминокислота с разветвленной цепью (BCAA), которая также может индуцироваться аспартатным путем, играет ключевую роль в устойчивости растений к стрессу в качестве фактора осмо-регуляции (Batista-Silva et al., 2019). Гомосерин и треонин также происходят из аспартатного пути, и было обнаружено, что дисбаланс аминокислот, связанный с накоплением гомосерина и треонина, повышает иммунитет растений к патогенам оомицетов (Zeier, 2013).
Метаболические связи между лизином и другими путями
На крахмал, как важный источник углеводов, также влияет накопление лизина (Jia et al., 2013). У мутантной кукурузы opaque2 наблюдалось увеличение экспрессии нескольких ферментов биосинтеза крахмала (GBSSI, Zpu1, SSIIa, BEI, BEIIb), что приводило к образованию высококристаллического крахмала (Jia et al. , 2013). Кроме того, Zhang et al. (2016) показали, что как Opaque — 2 , так и фактор связывания проламин-бокса регулируют синтез белка и крахмала.
, 2013). Кроме того, Zhang et al. (2016) показали, что как Opaque — 2 , так и фактор связывания проламин-бокса регулируют синтез белка и крахмала.
Ответы на развернутые белки (UPR) характеризуются активацией шаперонов и протеаз, облегчением сворачивания белка и разложением развернутого белка (Ruberti and Brandizzi, 2018). У риса с высоким содержанием лизина и сверхэкспрессией экзогенных белков BiP и PDI индуцировались в эндосперме, а также были тесно связаны с фенотипом зерна мучнистой / меловой, что позволяет предположить, что накопление лизина также связано с UPR (Wong et al., 2015).
У канолы и сои сверхэкспрессия генов Corynebacterium dapA и E. coli lysC привела к значительному увеличению свободного лизина, но снижению продукции масла (Falco et al., 1995). Аналогичные результаты были получены и для кукурузы (Medici et al., 2009). Однако кукуруза с высоким содержанием лизина PQ15 / CordapA, которая была разработана за счет уменьшения запаса белков семян с низким содержанием лизина, зеинов и сверхэкспрессии CordapA , показала увеличение содержания масла в зернах (Huang et al. , 2005). Кроме того, анализ метаболома и транскриптома показал снижение содержания липидов в трансгенном рисе с высоким содержанием лизина и повышение уровня мРНК липидных метаболических генов (Angelovici et al., 2009; Angelovici et al., 2011; Yang et al., 2018). ). Эти данные свидетельствуют о том, что регуляция метаболизма лизина влияет на метаболизм липидов, но с различиями между видами растений.
, 2005). Кроме того, анализ метаболома и транскриптома показал снижение содержания липидов в трансгенном рисе с высоким содержанием лизина и повышение уровня мРНК липидных метаболических генов (Angelovici et al., 2009; Angelovici et al., 2011; Yang et al., 2018). ). Эти данные свидетельствуют о том, что регуляция метаболизма лизина влияет на метаболизм липидов, но с различиями между видами растений.
Во время выработки лизина в микроорганизмах клетки вырабатывают ряд органических осмотических регуляторов, тем самым поддерживая нормальное осмотическое давление (Ying et al., 2014). Между тем, производство лизина отрицательно влияет на метаболизм, связанный с ацетатом, во время ферментации лизина (Anastassiadis, 2007). Однако у риса и арабидопсиса выявлена положительная корреляция между накоплением лизина и метаболизмом гликолиза (Angelovici et al., 2011; Yang et al., 2018).
Комплексный анализ метаболома и транскриптома показал, что 12 генов, кодирующих ферменты, связанные с метаболизмом нуклеотидов, были стимулированы, в то время как уровни аденина, псевдоуридина и урацила, которые связаны с метаболизмом нуклеотидов, также были улучшены у растений с высоким содержанием лизина (Angelovici et al. , 2009; Ян и др., 2018). Таким образом, метаболизм лизина влияет на путь семейства аспартата, который, в свою очередь, может влиять на другие пути метаболизма, подтверждая сложность пути семейства аспартата у растений.
, 2009; Ян и др., 2018). Таким образом, метаболизм лизина влияет на путь семейства аспартата, который, в свою очередь, может влиять на другие пути метаболизма, подтверждая сложность пути семейства аспартата у растений.
Связи между другими аминокислотами и связанными с ними метаболическими путями
В целом метаболизм аминокислот тесно связан с энергетическим и углеводным обменом, углеродно-азотным балансом и потребностями в синтезе белка и вторичном метаболизме (Pratelli and Pilot, 2014) .Метионин является основным ЕАА, ограничивающим серу, у растений, поскольку он может превращаться в цистеин у животных, таким образом удовлетворяя потребности обеих аминокислот (Song et al., 2013). И метионин, и S-аденозилметионин участвуют в путях биосинтеза этилена, никотианамина и полиамина (Sauter et al., 2013). Более того, было обнаружено, что арабидопсис с высоким содержанием метионина стимулирует метаболические и транскриптомные реакции, связанные со стрессом высыхания и энергетическим метаболизмом митохондрий (Cohen et al. , 2014). Предыдущее исследование также показало, что чем выше уровень метионина в растениях, тем выше их устойчивость к абиотическому стрессу (Ma et al., 2017). Цистеин — первый продукт ассимиляции сульфата, синтезируемый на последней стадии фотосинтетической ассимиляции сульфата (Gotor et al., 2015). Это не только белковый компонент, но и источник биосинтеза метионина, и многие другие серосодержащие метаболиты также участвуют в росте растений, передаче сигналов, стрессовых ответах и развитии противогрибковых средств (Roblin et al., 2018; Коприва и др., 2019). Кроме того, синтез цистеина является точкой слияния трех основных путей ассимиляции углерода, азота и серы (Jobe et al., 2019).
, 2014). Предыдущее исследование также показало, что чем выше уровень метионина в растениях, тем выше их устойчивость к абиотическому стрессу (Ma et al., 2017). Цистеин — первый продукт ассимиляции сульфата, синтезируемый на последней стадии фотосинтетической ассимиляции сульфата (Gotor et al., 2015). Это не только белковый компонент, но и источник биосинтеза метионина, и многие другие серосодержащие метаболиты также участвуют в росте растений, передаче сигналов, стрессовых ответах и развитии противогрибковых средств (Roblin et al., 2018; Коприва и др., 2019). Кроме того, синтез цистеина является точкой слияния трех основных путей ассимиляции углерода, азота и серы (Jobe et al., 2019).
BCAA изолейцин, валин и лейцин являются важными питательными веществами для людей и животных (Chen et al., 2010). BCAA и их производные также способствуют росту растений, реакции на стресс и выработке компонентов пищевого вкуса (Xing and Last, 2017). Пути катаболизма BCAA уже были определены как важные во время толерантности к дегидратации у Arabidopsis (Pires et al./molecular-3d-composition-of-amino-acid-arginine-157693924-58fdf61e3df78ca159b2591b.jpg) , 2016). Они также, по-видимому, влияют на устойчивость растений к различным классам патогенов, модулируя перекрестные помехи между защитными путями, регулируемыми салициловой кислотой и жасмоновой кислотой (Zeier, 2013). Более того, катаболизм BCAA обеспечивает альтернативный источник энергии при длительной обработке растений в темноте (Peng et al., 2015). Недавно Cao et al. (2019) также показали, что чрезмерное накопление BCAA приводит к усилению активности мишени рапамицина, что вызывает реорганизацию актинового цитоскелета и актин-ассоциированных эндомембран у мутантов Arabidopsis.Кроме того, было обнаружено, что изолейцин служит предшественником для синтеза β-аланина в растениях (Rouhier et al., 2019), в то время как у Arabidopsis была обнаружена метаболическая взаимосвязь между катаболизмом BCAA и кластерами железо-сера через . митохондриальный гомолог GRXS15 (Moseler et al., 2020).
, 2016). Они также, по-видимому, влияют на устойчивость растений к различным классам патогенов, модулируя перекрестные помехи между защитными путями, регулируемыми салициловой кислотой и жасмоновой кислотой (Zeier, 2013). Более того, катаболизм BCAA обеспечивает альтернативный источник энергии при длительной обработке растений в темноте (Peng et al., 2015). Недавно Cao et al. (2019) также показали, что чрезмерное накопление BCAA приводит к усилению активности мишени рапамицина, что вызывает реорганизацию актинового цитоскелета и актин-ассоциированных эндомембран у мутантов Arabidopsis.Кроме того, было обнаружено, что изолейцин служит предшественником для синтеза β-аланина в растениях (Rouhier et al., 2019), в то время как у Arabidopsis была обнаружена метаболическая взаимосвязь между катаболизмом BCAA и кластерами железо-сера через . митохондриальный гомолог GRXS15 (Moseler et al., 2020).
Аргинин и орнитин синтезируют полиамины и участвуют в реакции растений на стрессы. Кроме того, аргинин составляет высокий процент пула аминокислот в запасных белках хвойных пород, в то время как профилирование транскриптома и метаболома также выявило важные гены и метаболиты, участвующие в метаболизме аргинина на позднем эмбриогенезе (Businge et al., 2012; Canales et al., 2014). Поэтому считается, что метаболизм аргинина играет ключевую роль в хранении азота во время эмбриогенеза и мобилизации азота во время прорастания (Llebrés et al., 2018). Более того, декарбоксилированный саденозилметионин из метаболизма метионина служит донором аминопропила во время образования полиаминов (Gong et al., 2014). Данные также свидетельствуют о том, что растения синтезируют β-аланин из спермина, урацила и пропионата (Parthasarathy et al., 2019). Точно так же исследования метаболического отслеживания показывают, что пшеница синтезирует β-аланин как из изолейцина, так и из пропионата, как в Arabidopsis (Reinhart and Rouhier, 2019).У растений β-аланин важен для синтеза пантотената, а затем кофермента А, который является важным коферментом в метаболизме липидов и углеводов (Parthasarathy et al.
Кроме того, аргинин составляет высокий процент пула аминокислот в запасных белках хвойных пород, в то время как профилирование транскриптома и метаболома также выявило важные гены и метаболиты, участвующие в метаболизме аргинина на позднем эмбриогенезе (Businge et al., 2012; Canales et al., 2014). Поэтому считается, что метаболизм аргинина играет ключевую роль в хранении азота во время эмбриогенеза и мобилизации азота во время прорастания (Llebrés et al., 2018). Более того, декарбоксилированный саденозилметионин из метаболизма метионина служит донором аминопропила во время образования полиаминов (Gong et al., 2014). Данные также свидетельствуют о том, что растения синтезируют β-аланин из спермина, урацила и пропионата (Parthasarathy et al., 2019). Точно так же исследования метаболического отслеживания показывают, что пшеница синтезирует β-аланин как из изолейцина, так и из пропионата, как в Arabidopsis (Reinhart and Rouhier, 2019).У растений β-аланин важен для синтеза пантотената, а затем кофермента А, который является важным коферментом в метаболизме липидов и углеводов (Parthasarathy et al. , 2019). Гликолиз и цикл TCA связаны с помощью аланинаминотрансферазы во время гипоксии, вызванной переувлажнением (Rocha et al., 2010). Более того, у риса было обнаружено, что аланинаминотрансфераза 1, кодируемая геном Flo12 , одновременно регулирует углеродный и азотный метаболизм, в то время как мутант flo12 представляет собой мучнистый эндосперм с белым ядром (Zhong et al., 2019).
, 2019). Гликолиз и цикл TCA связаны с помощью аланинаминотрансферазы во время гипоксии, вызванной переувлажнением (Rocha et al., 2010). Более того, у риса было обнаружено, что аланинаминотрансфераза 1, кодируемая геном Flo12 , одновременно регулирует углеродный и азотный метаболизм, в то время как мутант flo12 представляет собой мучнистый эндосперм с белым ядром (Zhong et al., 2019).
В растениях ароматические аминокислоты фенилаланин, тирозин и триптофан являются не только важными компонентами синтеза белка, но также расположены выше ряда гормонов роста и вторичных метаболитов с множеством биологических функций и полезных для здоровья свойств, таких как защита от абиотического и биотического стресса (Цин, Галили, 2010). Фенилаланин необходим для биосинтеза белка и выживания клеток (Tzin and Galili, 2010), а в растениях он также действует как предшественник большого числа многофункциональных вторичных метаболитов.Среди них лигнин является основным структурным компонентом опорных тканей сосудистых растений и некоторых водорослей (Vanholme et al. , 2019). Тирозин является центральным звеном множества специализированных метаболических путей, в то время как витамин E и пластохинон являются важными метаболитами питания растений, фотосинтеза и синтеза антиоксидантов (Schenck and Maeda, 2018). Тирозин также является предшественником множества специализированных метаболитов с различными физиологическими функциями, таких как небелковые аминокислоты, аттрактанты и защитные соединения (Schenck and Maeda, 2018).Между тем, триптофан является важным EAA в синтезе большого количества биоактивных молекул, таких как ауксин, производные триптамина, фитоалексины, индолглюкозинолаты и терпеноидные индольные алкалоиды, а также играет ключевую роль в регуляции роста и развития растений. и стрессовые реакции. Соответственно, в последнее время все эти результаты широко обсуждались (Tzin and Galili, 2010; Hildebrandt et al., 2015; Datta et al., 2016).
, 2019). Тирозин является центральным звеном множества специализированных метаболических путей, в то время как витамин E и пластохинон являются важными метаболитами питания растений, фотосинтеза и синтеза антиоксидантов (Schenck and Maeda, 2018). Тирозин также является предшественником множества специализированных метаболитов с различными физиологическими функциями, таких как небелковые аминокислоты, аттрактанты и защитные соединения (Schenck and Maeda, 2018).Между тем, триптофан является важным EAA в синтезе большого количества биоактивных молекул, таких как ауксин, производные триптамина, фитоалексины, индолглюкозинолаты и терпеноидные индольные алкалоиды, а также играет ключевую роль в регуляции роста и развития растений. и стрессовые реакции. Соответственно, в последнее время все эти результаты широко обсуждались (Tzin and Galili, 2010; Hildebrandt et al., 2015; Datta et al., 2016).
Мало что известно о метаболизме гистидина и его связи с другими аминокислотами в растениях.Тесная корреляция между толерантностью к никелю, концентрацией корневого гистидина и обилием транскриптов ATP-PRT была обнаружена у растений с гипераккумуляторами, которые демонстрируют постоянно высокую экспрессию пути биосинтеза гистидина (Ingle et al. , 2005). Более того, исследования мутантов hisn1a показали, что гистидин регулирует отложение масла семян и накопление белка посредством биосинтеза абсцизовой кислоты и β-окисления у Arabidopsis (Ma and Wang, 2016). Биосинтез гистидина тесно связан с метаболизмом нуклеотидов через 5′-фосфорибозил-1-пирофосфат, который является промежуточным метаболитом антранилата.Таким образом, эти данные предполагают метаболическую связь между гистидином и триптофаном, метаболизм нуклеотидов (Koslowsky et al., 2008).
, 2005). Более того, исследования мутантов hisn1a показали, что гистидин регулирует отложение масла семян и накопление белка посредством биосинтеза абсцизовой кислоты и β-окисления у Arabidopsis (Ma and Wang, 2016). Биосинтез гистидина тесно связан с метаболизмом нуклеотидов через 5′-фосфорибозил-1-пирофосфат, который является промежуточным метаболитом антранилата.Таким образом, эти данные предполагают метаболическую связь между гистидином и триптофаном, метаболизм нуклеотидов (Koslowsky et al., 2008).
Перспективы на будущее
Наши знания о метаболизме аминокислот значительно выросли за последние три десятилетия. Аминокислоты и их производные выполняют различные важные функции в растениях, такие как синтез белка, рост и развитие, питание и стрессовые реакции (Hildebrandt et al., 2015). Между тем, метаболизм — одна из самых важных и сложных сетей в биологических системах, но наше понимание регуляции метаболизма остается ограниченным с точки зрения модульной работы этих сетей. Поэтому очень важна точная и подробная информация о биологических и молекулярных механизмах и метаболических связях. Недавние разработки омических подходов широко применялись для изучения метаболизма аминокислот и их связей (Gu et al., 2010; Angelovici et al., 2017; Xing and Last, 2017; Yang et al., 2018). Комбинации биохимии, молекулярной генетики, геномики и системной биологии будут и дальше способствовать фундаментальным исследованиям, позволяя нам разрабатывать идеи и стратегии, направленные на изучение новых свойств регуляторных сетей ген-белок-метаболит (рис. 2).Более того, исследования эпитранскриптомики могут предоставить новую стратегию для анализа метаболических связей в растениях, дающую представление о том, как различные маркеры регулируют множество биологических процессов, от биосинтеза до катаболизма и транспорта для функционирования (Vandivier and Gregory, 2018). Кроме того, постоянно оптимизируемая технология редактирования генов CRISPR-Cas позволила изменить экспрессию или активность одного или нескольких ключевых регуляторных ферментов, поддерживая исследования, направленные на улучшение качества питания растений (Chen et al.
Поэтому очень важна точная и подробная информация о биологических и молекулярных механизмах и метаболических связях. Недавние разработки омических подходов широко применялись для изучения метаболизма аминокислот и их связей (Gu et al., 2010; Angelovici et al., 2017; Xing and Last, 2017; Yang et al., 2018). Комбинации биохимии, молекулярной генетики, геномики и системной биологии будут и дальше способствовать фундаментальным исследованиям, позволяя нам разрабатывать идеи и стратегии, направленные на изучение новых свойств регуляторных сетей ген-белок-метаболит (рис. 2).Более того, исследования эпитранскриптомики могут предоставить новую стратегию для анализа метаболических связей в растениях, дающую представление о том, как различные маркеры регулируют множество биологических процессов, от биосинтеза до катаболизма и транспорта для функционирования (Vandivier and Gregory, 2018). Кроме того, постоянно оптимизируемая технология редактирования генов CRISPR-Cas позволила изменить экспрессию или активность одного или нескольких ключевых регуляторных ферментов, поддерживая исследования, направленные на улучшение качества питания растений (Chen et al. , 2019).
, 2019).
Рис. 2 Схематическое изображение рабочего процесса, изображающего применение различных подходов для получения системного понимания метаболических связей аминокислот в растениях и их применения для получения биообогащенных аминокислотами сельскохозяйственных культур.
Помимо технологических вопросов, в наших знаниях в определенных областях есть существенные пробелы. Учитывая важность питательной ценности аминокислот, влияние накопления аминокислот (особенно ЕАА) на другие метаболические пути во время роста и развития растений требует дальнейшего внимания.Для этого необходим анализ связей между метаболизмом аминокислот, факторами регуляции транскрипции и посттрансляционными модификациями. Таким образом, несмотря на наши растущие знания о метаболизме растительных аминокислот и их метаболических связях, ясно, что многие важные открытия еще предстоит сделать.
Вклад авторов
QY и QL организовали и написали рукопись. DZ дал критическую оценку и отредактировал текст. Все авторы внесли свой вклад в статью и одобрили представленную версию.
Все авторы внесли свой вклад в статью и одобрили представленную версию.
Финансирование
Эта работа была поддержана Национальным фондом естественных наук Китая (31801322 и 31701393), Министерством сельского хозяйства (2016ZX08001006-005) и правительством провинции Цзянсу (BE2018357 и PAPD) Китая.
Конфликт интересов
Авторы заявляют, что это исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Источники
Ангеловичи, Р., Fait, A., Zhu, X., Szymanski, J., Feldmesser, E., Fernie, A. R., et al. (2009). Расшифровка транскрипционных и метаболических сетей, связанных с метаболизмом лизина во время развития семян арабидопсиса. Plant Physiol. 151, 2058–2072. doi: 10.1104 / pp.109.145631
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ангеловичи Р., Фейт А., Ферни А. Р., Галили Г. (2011). Признак высокого содержания лизина в семенах отрицательно связан с циклом TCA и замедляет прорастание семян Arabidopsis. New Phytol. 189, 148–159. doi: 10.1111 / j.1469-8137.2010.03478.x
New Phytol. 189, 148–159. doi: 10.1111 / j.1469-8137.2010.03478.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ангеловичи Р., Батушанский А., Дисон Н., Гонсалес-Хорхе С., Гор М. А., Фейт А. и др. (2017). GWAS с сетевым управлением улучшает идентификацию генов, влияющих на свободные аминокислоты. Plant Physiol. 173, 872–886. doi: 10.1104 / pp.16.01287
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Arruda, P., Баррето, П. (2020). Катаболизм лизина через сахаропиновый путь: ферменты и промежуточные соединения, участвующие в ответах растений на абиотический и биотический стресс. Фронт. Plant Sci. 11, 587. doi: 10.3389 / fpls.2020.00587
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Батиста-Сильва, В., Хайнеманн, Б., Руген, Н., Нунес-Неси, А., Араужо, В. Л., Браун, Х. П. и др. (2019). Роль метаболизма аминокислот при снятии абиотического стресса. Среда растительных клеток. 42, 1630–1644. doi: 10.1111 / pce.13518
doi: 10.1111 / pce.13518
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Bernsdorff, F., Döring, A.C., Gruner, K., Schuck, S., Bräutigam, A., Zeier, J. (2016). Пипеколиновая кислота регулирует приобретенную системную устойчивость растений и прайминг защиты через зависимые и независимые от салициловой кислоты пути. Растительная клетка 28, 102–129. doi: 10.1105 / tpc.15.00496
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Brocker, C., Лассен, Н., Эстей, Т., Паппа, А., Канторе, М., Орлова, В.В. и др. (2010). Альдегиддегидрогеназа 7A1 (ALDH7A1) — новый фермент, участвующий в защите клеток от гиперосмотического стресса. J. Biol. Chem. 285, 18452–18463. doi: 10.1074 / jbc.M109.077925
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Businge, E., Brackmann, K., Moritz, T., Egertsdotter, U. (2012). Профилирование метаболитов выявляет четкие метаболические изменения во время соматического развития зародыша ели европейской ( Picea abies ).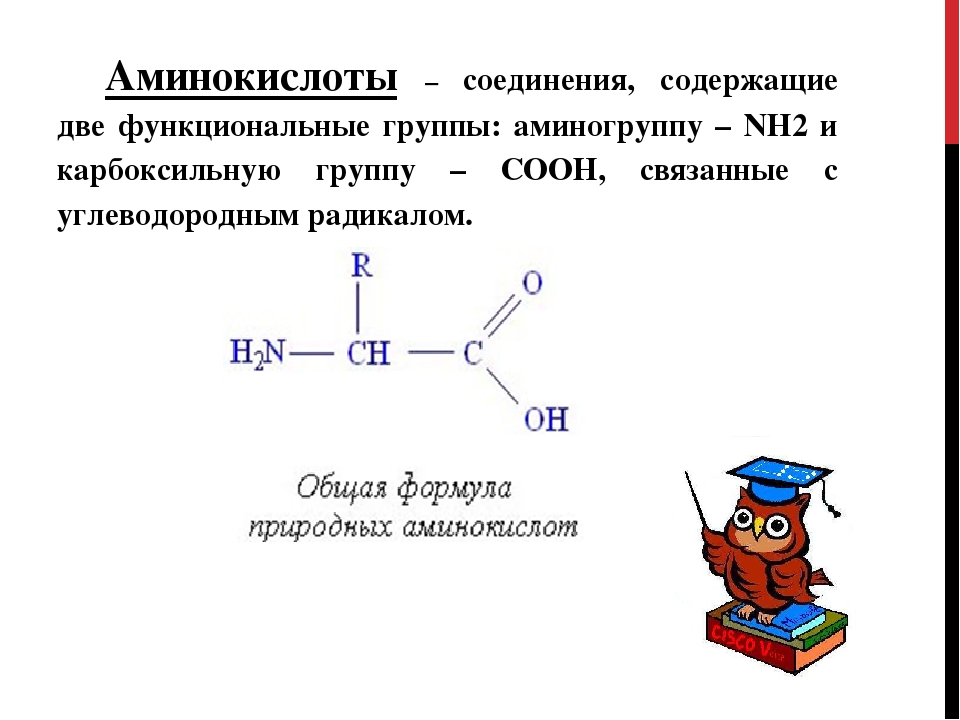 Tree Physiol. 32, 232–244. doi: 10.1093 / treephys / tpr142
Tree Physiol. 32, 232–244. doi: 10.1093 / treephys / tpr142
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Canales, J., Bautista, R., Label, P., Gómez-Maldonado, J., Lesur, I., Fernández-Pozo, N., et al. (2014). De novo сборка транскриптома морской сосны: значение для лесоводства и биотехнологии. Plant Biotechnol. J. 12, 286–299. doi: 10.1111 / pbi.12136
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цао, П., Ким, С. Дж., Син, А., Шенк, К. А., Лю, Л., Цзян, Н., и др. (2019). Гомеостаз аминокислот с разветвленной цепью имеет решающее значение для активности передачи сигналов TOR у Arabidopsis. eLife 8, e50747. doi: 10.7554 / eLife.50747
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чен, Х., Сакса, К., Чжао, Ф., Цю, Дж., Сюн, Л. (2010). Генетический анализ регуляции пути повышения биосинтеза аминокислот с разветвленной цепью у растений. Завод J. 63, 573–583. doi: 10.1111 / j.1365-313X.2010.04261.x
doi: 10.1111 / j.1365-313X.2010.04261.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Chen, K., Wang, Y., Zhang, R., Zhang, H., Gao, C. (2019). Редактирование генома CRISPR / Cas и точная селекция растений в сельском хозяйстве. Annu. Rev. Plant Biol. 70, 667–697. doi: 10.1146 / annurev-arplant-050718-100049
PubMed Реферат | CrossRef Полный текст | Google Scholar
Коэн, Х., Исраэль, Х., Матитьяху, И., Амир, Р. (2014). Специфическая для семян экспрессия нечувствительной к обратной связи формы ЦИСТАТИОНИН-γ-СИНТАЗЫ в Arabidopsis стимулирует метаболические и транскриптомные реакции, связанные со стрессом высыхания. Plant Physiol. 166, 1575–1592. doi: 10.1104 / pp.114.246058
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Datta, A., Bhattacharyya, D., Singh, S., Ghosh, A., Schmidtchen, A., Malmsten, M., et al. (2016). Роль ароматических аминокислот в липополисахаридах и мембранных взаимодействиях антимикробных пептидов для использования в борьбе с болезнями растений. J. Biol. Chem. 291, 13301–13317. doi: 10.1074 / jbc.M116.719575
J. Biol. Chem. 291, 13301–13317. doi: 10.1074 / jbc.M116.719575
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Falco, S.C., Guida, T., Locke, M., Mauvais, J., Sanders, C., Ward, R.T., et al. (1995). Трансгенные семена канолы и сои с повышенным содержанием лизина. Нац. Biotechnol. 13, 577–582. doi: 10.1038 / nbt0695-577
CrossRef Полный текст | Google Scholar
Галили, Г., Амир, Р., Ферни, А. Р. (2016). Регуляция синтеза и накопления незаменимых аминокислот в растениях. Annu. Rev. Plant Biol. 67, 153–178. doi: 10.1146 / annurev-arplant-043015-112213
PubMed Реферат | CrossRef Полный текст | Google Scholar
Гонг, Б., Ли, X., VandenLangenberg, K.M, Wen, D., Sun, S., Wei, M., et al. (2014). Сверхэкспрессия S-аденозил-L-метионинсинтетазы увеличивала устойчивость томатов к щелочному стрессу за счет метаболизма полиаминов. Plant Biotechnol. J. 12, 694–708. doi: 10.1111 / pbi.12173
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Готор, К. , Лауреано-Марин, А. М., Морено, И., Арока, А., Гарсия, И., Ромеро, Л. К. (2015). Сигнал в цитозоле растения: цистеин или сульфид? Аминокислоты 47, 2155–2164.doi: 10.1007 / s00726-014-1786-z
, Лауреано-Марин, А. М., Морено, И., Арока, А., Гарсия, И., Ромеро, Л. К. (2015). Сигнал в цитозоле растения: цистеин или сульфид? Аминокислоты 47, 2155–2164.doi: 10.1007 / s00726-014-1786-z
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гу Л., Джонс А. Д., Ласт Р. Л. (2010). Широкие связи в метаболической сети семян арабидопсиса, выявленные с помощью профилирования метаболитов мутанта по катаболизму аминокислот. Plant J. 61, 579–590. doi: 10.1111 / j.1365-313X.2009.04083.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Huang, S., Kruger, D. E., Frizzi, A., D’Ordine, R. L., Florida, C.А., Адамс, В. Р. и др. (2005). Кукуруза с высоким содержанием лизина, продуцируемая комбинацией усиленного биосинтеза лизина и сниженного накопления зеина. Plant Biotechnol. J. 3, 555–569. doi: 10.1111 / j.1467-7652.2005.00146.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ингл, Р. А., Магфорд, С. Т., Риз, Дж. Д., Кэмпбелл, М. М., Смит, Дж. А. С. (2005). Постоянно высокая экспрессия пути биосинтеза гистидина способствует устойчивости к никелю у растений-гипераккумуляторов. Растительная клетка 17, 2089–2106. doi: 10.1105 / tpc.104.030577
Т., Риз, Дж. Д., Кэмпбелл, М. М., Смит, Дж. А. С. (2005). Постоянно высокая экспрессия пути биосинтеза гистидина способствует устойчивости к никелю у растений-гипераккумуляторов. Растительная клетка 17, 2089–2106. doi: 10.1105 / tpc.104.030577
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цзя, М., Ву, Х., Клэй, К. Л., Юнг, Р., Ларкинс, Б. А., Гиббон, Б. С. (2013). Идентификация и характеристика белков, богатых лизином, и генов биосинтеза крахмала в мутанте opaque2 с помощью транскрипционного и протеомного анализа. BMC Plant Biol. 13, 60. doi: 10.1186 / 1471-2229-13-60
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джоб, Т.О., Зензен, И., Карвансара, П. Р., Коприва, С. (2019). Интеграция ассимиляции сульфатов с метаболизмом углерода и азота при переходе от фотосинтеза C 3 к C 4 . J. Exp. Бот. 70, 4211–4221. doi: 10.1093 / jxb / erz250
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Коприва, С. , Малаголи, М., Такахаши, Х. (2019). Серное питание: влияет на развитие растений, метаболизм и реакцию на стресс. J. Exp. Бот. 70, 4069–4073.doi: 10.1093 / jxb / erz319
, Малаголи, М., Такахаши, Х. (2019). Серное питание: влияет на развитие растений, метаболизм и реакцию на стресс. J. Exp. Бот. 70, 4069–4073.doi: 10.1093 / jxb / erz319
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Koslowsky, S., Riegler, H., Bergmüller, E., Zrenner, R. (2008). Повышенное накопление биомассы за счет увеличения активности фосфорибозилпирофосфатсинтетазы у Arabidopsis thaliana и Nicotiana tabacum. Plant Biotechnol. J. 6, 281–294. doi: 10.1111 / j.1467-7652.2007.00314.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лю, X., Zhang, C., Wang, X., Лю, К., Юань, Д., Пан, Г., и др. (2016). Получение риса с высоким содержанием лизина посредством специфической для эндосперма экспрессии чужеродного гена LYSINE RICH PROTEIN . BMC Plant Biol. 16, 147. doi: 10.1186 / s12870-016-0837-x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Llebrés, M. T., Pascual, M. B., Debille, S. , Trontin, J. F., Harvengt, L., Avila, C., et al. (2018). Роль метаболического пути аргинина в эмбриогенезе и прорастании сосны приморской ( Pinus pinaster Ait.). Tree Physiol. 38, 471–484. doi: 10.1093 / treephys / tpx133
, Trontin, J. F., Harvengt, L., Avila, C., et al. (2018). Роль метаболического пути аргинина в эмбриогенезе и прорастании сосны приморской ( Pinus pinaster Ait.). Tree Physiol. 38, 471–484. doi: 10.1093 / treephys / tpx133
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Long, X., Liu, Q., Chan, M., Wang, Q., Sun, S. S. M. (2013). Метаболическая инженерия и профилирование риса с повышенным содержанием лизина. Plant Biotechnol. J. 11, 490–501. doi: 10.1111 / pbi.12037
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ma, C., Wang, Y., Gu, D., Nan, J., Chen, S., Li, H. (2017). Сверхэкспрессия S-аденозил-L-метионинсинтетазы 2 из сахарной свеклы M14 увеличивала устойчивость Arabidopsis к соли и окислительному стрессу. Внутр. J. Mol. Sci. 18, 847. doi: 10.3390 / ijms18040847
CrossRef Полный текст | Google Scholar
Медичи, Л. О., Газиола, С. А., Вариси, В. А., Паула, Дж. А. С., Феррейра, Р. Р., Азеведо, Р. А. (2009). Диаллельный анализ содержания лизина и масла в зернах кукурузы. Sci. Agric. 66, 204–209. doi: 10.1590 / S0103-
Р., Азеведо, Р. А. (2009). Диаллельный анализ содержания лизина и масла в зернах кукурузы. Sci. Agric. 66, 204–209. doi: 10.1590 / S0103-
00
00009
CrossRef Полный текст | Google Scholar
Moseler, A., Kruse, I., Maclean, A. E., Pedroletti, L., Wagner, S., Wehler, R., et al. (2020). Катаболизм аминокислот с разветвленной цепью зависит от GRXS15 через гомеостаз митохондриального липоильного кофактора. BioRxiv. 2020.02.13.947697. doi: 10.1101 / 2020.02.13.947697
CrossRef Полный текст | Google Scholar
Peng, C., Uygun, S., Shiu, S.H., Last, R.L. (2015). Влияние комплекса дегидрогеназы кетокислот с разветвленной цепью на аминокислотный гомеостаз у Arabidopsis. Plant Physiol. 169, 1807–1820. doi: 10.1104 / pp.15.00461
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пирес, М.В., Джуниор, А. А. П., Медейрос, Д. Б., Далосо, Д. М., Фам, П. А., Баррос, К. А. и др. (2016). Влияние альтернативных путей дыхания, в которых используются аминокислоты с разветвленной цепью, после нехватки воды у Arabidopsis. Среда растительных клеток. 39, 1304–1319. doi: 10.1111 / pce.12682
Среда растительных клеток. 39, 1304–1319. doi: 10.1111 / pce.12682
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Reinhart, P. E., Rouhier, K. A. (2019). Метаболическое отслеживание и количественное определение β-аланина в пшенице. FASEB J. 33, 486.9. doi: 10.1096 / fasebj.2019.33.1_supplement.486.9
CrossRef Полный текст | Google Scholar
Роблин, Г., Октав, С., Фаучер, М., Флёрат-Лессар, П., Бержео, Дж. (2018). Цистеин: многогранная аминокислота, участвующая в передаче сигналов, устойчивости растений и развитии противогрибковых средств. Plant Physiol. Bioch. 129, 77–89. doi: 10.1016 / j.plaphy.2018.05.024
CrossRef Полный текст | Google Scholar
Rocha, M., Licausi, F., Araújom, W. L., Nunes-Nesim, A., Sodek, L., Ферни, А. Р. и др. (2010). Гликолиз и цикл трикарбоновых кислот связаны с помощью аланинаминотрансферазы во время гипоксии, вызванной переувлажнением Lotus japonicus . Plant Physiol. 152, 1501–1513. doi: 10.1104 / pp.109.150045
152, 1501–1513. doi: 10.1104 / pp.109.150045
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Rouhier, K. A., Perrett, M. E., Goldfarb, M. H., Reinhart, P. E., Wilczewski-Shirai, K. C., Raymond-Schmidt, L., et al. (2019). Изолейцин служит предшественником синтеза β-аланина в растениях. FASEB J. 33, 486.10. doi: 10.1096 / fasebj.2019.33.1_supplement.486.10
CrossRef Полный текст | Google Scholar
Руберти, К., Брандици, Ф. (2018). «Методы в молекулярной биологии» в Unfolded protein response in Arabidopsis, в The Plant Endoplasmic Reticulum (Нью-Йорк: Humana Press).
Google Scholar
Заутер, М., Моффатт, Б., Саехао, М.С., Ад, Р., Виртц, М. (2013). Спасение метионина и S-аденозилметионин: важные связи между биосинтезом серы, этилена и полиамина. Biochem. J. 451, 145–154. doi: 10.1042 / BJ20121744
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Song, S., Hou, W., Godo, I., Wu, C. , Yu, Y., Matityahu, I., et al. (2013). Семена сои, экспрессирующие нечувствительную к обратной связи цистатионин-γ-синтазу, демонстрируют более высокое содержание метионина. J. Exp. Бот. 64, 1917–1926. doi: 10.1093 / jxb / ert053
, Yu, Y., Matityahu, I., et al. (2013). Семена сои, экспрессирующие нечувствительную к обратной связи цистатионин-γ-синтазу, демонстрируют более высокое содержание метионина. J. Exp. Бот. 64, 1917–1926. doi: 10.1093 / jxb / ert053
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Sun, S. S. M., Liu, Q.(2004). Трансгенные подходы к улучшению питательных свойств растительных белков. Vitro Cell. Dev. Pl. 40, 155–162. doi: 10.1079 / IVP2003517
CrossRef Полный текст | Google Scholar
Тровато, М., Форлани, Г., Синьорелли, С., Функ, Д. (2019). «Метаболизм пролина и его функции в развитии и устойчивости к стрессу» в Устойчивость растений к абиотическому стрессу, опосредованная осмопротекторами. Ред. Хоссейн, М., Кумар, В., Буррит, Д., Фуджита, М., Мякеля, П. Нью-Йорк: Springer.doi: 10.1007 / 978-3-030-27423-8_2
CrossRef Полный текст | Google Scholar
Vanholme, R., Meester, B.D., Ralph, J., Boerjan, W. (2019). Биосинтез лигнина и его интеграция в метаболизм. Curr. Opin. Биотех. 56, 230–239. doi: 10.1016 / j.copbio.2019.02.018
Биосинтез лигнина и его интеграция в метаболизм. Curr. Opin. Биотех. 56, 230–239. doi: 10.1016 / j.copbio.2019.02.018
CrossRef Полный текст | Google Scholar
Wang, W., Xu, M., Wang, G., Galili, G. (2018). Новые сведения о метаболизме аминокислот семейства аспартатов в семенах растений. Завод Репродукт. 31, 203–211. doi: 10.1007 / s00497-018-0322-9
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wang, W., Dai, Y., Wang, M., Yang, W., Zhao, D. (2019). Динамика транскриптома двойного рецессивного мутанта o2o2o16o16 выявляет транскрипционные механизмы увеличения содержания его лизина и триптофана в кукурузе. Гены 10, 316. doi: 10.3390 / genes10040316
CrossRef Полный текст | Google Scholar
Yang, Q., Zhang, C., Chan, M. L., Zhao, D., Chen, J., Wang, Q., et al. (2016). Биообогащение риса незаменимой аминокислотой лизином: молекулярная характеристика, оценка питательности и полевые показатели. J. Exp. Бот. 67, 4285–4296. doi: 10.1093 / jxb / erw209
Exp. Бот. 67, 4285–4296. doi: 10.1093 / jxb / erw209
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Yang, Q., Zhao, D., Zhang, C., Wu, H., Li, Q., Gu, M., et al. (2018). Связь между метаболизмом лизина и серотонина в эндосперме риса. Plant Physiol. 176, 1965–1980. doi: 10.1104 / pp.17.01283
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ying, H., He, X., Li, Y., Chen, K., Ouyang, P. (2014). Оптимизация условий культивирования для увеличения производства лизина с использованием сконструированной Escherichia coli . заявл. Biochem. Biotechnol. 172, 3835–3843. doi: 10.1007 / s12010-014-0820-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Zhang, Z., Zheng, X., Yang, J., Messing, J., Ву, Ю. (2016). Факторы транскрипции, специфичные для эндосперма кукурузы O2 и PBF , регулируют синтез белка и крахмала. Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 113, 10842–10847. doi: 10.1073 / pnas.1613721113
CrossRef Полный текст | Google Scholar
Zhong, M., Liu, X., Liu, F., Ren, Y., Wang, Y., Zhu, J., et al. (2019). FLOURY ENDOSPERM12 , кодирующая аланинаминотрансферазу 1, регулирует метаболизм углерода и азота в рисе. J. Plant Biol. 62, 61–73. doi: 10.1007 / s12374-018-0288-z
CrossRef Полный текст | Google Scholar
Чжу, X., Галили, Г. (2004). Метаболизм лизина одновременно регулируется синтезом и катаболизмом как в репродуктивных, так и в вегетативных тканях. Plant Physiol. 135, 129–136. doi: 10.1104 / pp.103.037168
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Различные характеристики экскреции аминокислот с мочой у людей и использование аминокислотных добавок для снижения усталости и ухудшения здоровья у взрослых | Журнал питания
Boirie Y. Физиопатологический механизм саркопении. J Nutr Здоровье Старения. 2009; 13: 717–23.
CAS
Статья
PubMed
Google ученый
Jagoe RT, Engelen MPKJ. Мышечное истощение и изменения метаболизма мышечных белков при хронической обструктивной болезни легких. Eur Respir J. 2003; 22: 52s – 63s.
CAS
Статья
Google ученый
Oehler R, Roth E.Метаболизм глутамина. В: Кинобер Л.А., редактор. Метаболические и терапевтические аспекты аминокислот в лечебном питании. Издание второе. Нью-Йорк: CRC Press; 2004. с. 169–82.
Google ученый
Udenfriend S, Wyngaarden JB. Предшественники адреналина и норадреналина in vivo. Biochim Biophys Acta. 1956; 20: 48–52.
CAS
Статья
PubMed
Google ученый
де Конинг Т.Дж., Снелл К., Дюран М., Бергер Р., Опрос-Б-Т, Сёртиз Р. L-серин в болезни и развитии. Биохим Дж. 2003; 371: 653–61.
Артикул
PubMed
PubMed Central
Google ученый
Poortmans JR, Carpentier LO, Pereira-Lancha LO, Lancha Jr A. Оборот белков, потребности в аминокислотах и рекомендации для спортсменов и активных групп населения. Braz J Med Biol Res. 2012; 45: 875–90.
CAS
Статья
PubMed
PubMed Central
Google ученый
Филипс СМ. Потребность в белке и добавки в силовых видах спорта. Питание. 2004. 20: 689–95.
CAS
Статья
PubMed
Google ученый
Waterlow JC, Jackson AA. Питание и белковый обмен в человеке. Br Med Bull. 1981; 37: 5–10.
CAS
Статья
PubMed
Google ученый
Corsetti R, Barassi A, Perego S, Sansoni V, Rossi A, Damele CAL, d’Eril GM, Banfi G, Lombardi G.Изменения экскреции аминокислот с мочой в зависимости от маркеров мышечной активности во время профессиональных соревнований по велоспорту: в поисках маркеров усталости. Аминокислоты. 2016; 48: 183–92.
CAS
Статья
PubMed
Google ученый
Dunstan RH, Sparkes DL, Dascombe BJ, Evans CA, Macdonald MM, Crompton MJ, Franks J, Murphy G, Gottfries J, Carlton B. Sweat способствовал потере аминокислот у лошадей стандартной породы и применению добавок стратегии для поддержания кондиции во время тренировок.Comp Exercise Physiol. 2015; 11: 201–12.
Артикул
Google ученый
Данстан Р.Х., Спаркс Д.Л., Даскомб Б.Дж., Макдональд М.М., Эванс К.А., Стивенс С.Дж., Кромптон М.Дж., Готфрис Дж., Фрэнкс Дж., Мерфи Дж. И др. Пот способствует потере аминокислот у спортсменов-мужчин во время упражнений при 32-34 ° C. PLoS One. 2016; 11: 1–16.
Артикул
Google ученый
Dunstan RH.Измененный аминокислотный гомеостаз и развитие утомляемости пациентов лучевой терапии рака груди: пилотное исследование. Clin Biochem. 2011; 44: 208–15.
Niblett SN. Гематологические аномалии и аномалии мочевыделения у пациентов с синдромом хронической усталости. Exp Biol Med. 2007; 232: 1041–9.
CAS
Статья
Google ученый
Liappis N, Hungerland H. Количественное исследование свободных аминокислот в эккринном поте человека при нормальных условиях.Am J Clin Nutr. 1972; 25: 661–3.
CAS
PubMed
Google ученый
Армстронг, доктор медицины, Став У. Исследование уровней свободных аминокислот в плазме. II. Нормальные значения для детей и взрослых. Обмен веществ. 1973; 22: 561–9.
CAS
Статья
PubMed
Google ученый
Weschler LB. Концентрация электролитов пота, полученная из окклюзионных покрытий, ложно высока, потому что пот сам вымывает электролиты кожи.J Appl Physiol. 2008; 105: 1376–7.
Артикул
PubMed
Google ученый
Scott IR, Harding CR, Barrett JG. Богатый гистидином белок гранул кератогиалина. Источник свободных аминокислот, урокановой кислоты и пирролидонкарбоновой кислоты в роговом слое. Biochim Biophys Acta. 1982; 719: 110–7.
CAS
Статья
PubMed
Google ученый
Kingsbury KJ, Kay L, Hjelm M. Контрастные образцы свободных аминокислот в плазме крови у элитных спортсменов: связь с усталостью и инфекцией. Br J Sports Med. 1998. 32: 25–32.
CAS
Статья
PubMed
PubMed Central
Google ученый
Паддон-Джонс Д. Атрофия и нарушение синтеза мышечного белка при длительном бездействии и стрессе. J Clin Endocrinol Metabol. 2006; 91: 4836–41.
CAS
Статья
Google ученый
Данстан Р.Х., Спаркс Д.Л., Робертс Т.К., Кромптон М.Дж., Готфрис Дж., Даскомб Б.Дж. Разработка комплексной аминокислотной добавки fatigue reviva ™ для перорального приема: первоначальная оценка концепции продукта и влияние на симптомы слабого здоровья в группе мужчин. Нутр Дж. 2013; 12: 115.
CAS
Статья
PubMed
PubMed Central
Google ученый
Данстан Р.Х., Спаркс Д.Л., Робертс Т.К., Даскомб Б.Дж. Предварительные оценки комплексной аминокислотной добавки Reviva TM для снижения утомляемости в группе профессиональных спортсменов-мужчин и в группе мужчин, набранных из числа населения.Food Nutr Sci. 2014; 5: 231–5.
Артикул
Google ученый
Данстан Р.Х., МакГрегор Н.Р., Батт Х.Л., Робертс Т.К., Клайнберг И.Дж., Ниблетт С.Н., Роткирх Т.Б., Баттфилд I. Характеристика дифференциального гомеостаза аминокислот среди подгрупп населения: основа для определения конкретных потребностей в аминокислотах. J Nutr Environ Med. 2000; 10: 211–23.
CAS
Статья
Google ученый
Watanabe N, Stewart R, Jenkins R, Bhugra DK, Furukawa TA. Эпидемиология хронической усталости, соматических заболеваний и симптомов распространенных психических расстройств: перекрестное исследование из второго британского национального исследования психиатрической заболеваемости. J Psychosom Res. 2008. 64: 357–62.
Артикул
PubMed
Google ученый
Li G, Xie F, Yan S, Hu X, Jin B, Wang J, Wu J, Yin D, Xie Q: Subhealth: определение, критерии диагностики и потенциальная распространенность в центральном регионе Китая.BMC Public Health. 2013; 13.
Эванс К., Данстан Р.Х., Роткирх Т., Робертс Т.К., Райхелт К.Л., Косфорд Р., Дид Дж., Эллис Э.Б., Спаркс Д.Л. Нарушение экскреции аминокислот у детей с аутизмом. Nutr Neurosci. 2008; 11: 9–17.
CAS
Статья
PubMed
Google ученый
Чалдер Т., Береловиц Г., Павликовска Т., Уоттс Л., Уэссели С., Райт Д., Уоллес Е.П. Развитие шкалы утомляемости. J Psychosom Res.1993; 37: 147–53.
CAS
Статья
PubMed
Google ученый
МакГрегор Н.Р., Данстан Р.Х., Зербес М., Батт Х.Л., Робертс Т.К., Клайнберг И.Дж. Предварительное определение связи между выражением симптомов и метаболитами в моче у субъектов с синдромом хронической усталости. Biochem Mol Med. 1996. 58: 85–92.
CAS
Статья
PubMed
Google ученый
Данстан Р.Х., МакГрегор Н.Р., Батт Г.Л., Робертс Т.К. Биохимические и микробиологические аномалии при синдроме хронической усталости: разработка лабораторных тестов и возможная роль токсичных химикатов. J Nutr Environ Med. 1999; 9: 97–108.
CAS
Статья
Google ученый
Ричардс Р.С., Робертс Т.К., МакГрегор Н.Р., Данстан Р.Х., Батт Х.Л. Параметры крови, указывающие на окислительный стресс, связаны с выражением симптомов синдрома хронической усталости.Redox Rep. 2000; 5: 35–41.
CAS
Статья
PubMed
Google ученый
Коды пищевых стандартов Австралии и Новой Зеландии [http://www.comlaw.gov.au/Search/Australia%20New%20Zealand%20Food%20Standards]. По состоянию на 13 июня 2013 г.
Закон о пищевых продуктах 2003 г. № 43 [http://www.legislation.nsw.gov.au/viewtop/inforce/act+43+2003+FIRST+0+N/]. По состоянию на 13 июня 2013 г.
Food Regulation 2010 [http: // www.законодательство.nsw.gov.au/viewtop/inforce/subordleg+250+2010+cd+0+N]. По состоянию на 13 июня 2013 г.
Clemens RA, Kopple JD, Swendseid ME. Метаболические эффекты диет с дефицитом гистидина, скармливаемых растущим крысам через желудочный зонд. J Nutr. 1984; 114: 2138–46.
CAS
PubMed
Google ученый
Куперман Дж. М., Лопес Р. Роль гистидина при анемии дефицита фолиевой кислоты. Exp Biol Med. 2002; 227: 998–1000.
CAS
Google ученый
Виру А., Виру А. Биохимический мониторинг спортивной тренировки. 1-е изд. Champaign: Human Kinetics Publishers; 2001.
Google ученый
Хараламби Г., Берг А. Изменения мочевины в сыворотке и аминного азота в зависимости от продолжительности упражнений. Eur J Phys Occup Physiol. 1976; 36: 39–48.
CAS
Статья
Google ученый
Hedges REM, Clement JG, Thomas DL, O’Connell TC.Обмен коллагена в средней части диафиза бедренной кости у взрослых: моделируется на основе измерений антропогенных радиоуглеродных индикаторов. Am J Phys Anthropol. 2007; 133: 808–16.
Артикул
PubMed
Google ученый
Wang W, Wu Z, Dai Z, Yang Y, Wang J, Wu G. Метаболизм глицина у животных и людей: последствия для питания и здоровья. Аминокислоты. 2013; 45: 463–77.
Артикул
PubMed
Google ученый
Кори JG. Метаболизм пуриновых и пиримидиновых нуклеотидов. В: Devlin TM, редактор. Учебник биохимии с клиническими корреляциями. 3-е изд. Нью-Йорк: Вили-Лисс; 1992. стр. 529–73.
Google ученый
Fessas PH, Koniavitis A, Zeis A. Экскреция бета-аминоизомасляной кислоты с мочой при талассемии. J Clin Pathol. 1969; 22: 154–7.
CAS
Статья
PubMed
PubMed Central
Google ученый
Энхьяргал Т., Цереннадмид С. Выведение с мочой бета-аминоизомасляной кислоты при гематологических заболеваниях (аннотация). Риншо Бёри Jpn J Clin Pathol. 2004; 52: 17–21.
CAS
Google ученый
Мацумото AM. Андропауза: клинические последствия снижения уровня тестостерона в сыворотке с возрастом у мужчин. J Gerontol. 2002; 57A: M76–99.
CAS
Статья
Google ученый
Новак А., Брод М., Эльберс Дж. Андропауза и качество жизни: результаты фокус-групп пациентов и клинических экспертов. Maturitas. 2002; 43: 231–7.
CAS
Статья
PubMed
Google ученый
Bain J. Заместительная терапия тестостероном для пожилых мужчин. Может Фам Врач. 2001; 47: 91–7.
CAS
PubMed
PubMed Central
Google ученый
Ногейра Аде С., Вале Р.Г., Гомеш А.Л., Дантас Э.Влияние мышечных воздействий на уровень повреждения соединительной ткани. Res Sports Med. 2011; 19: 259–70.
PubMed
Google ученый
Prockop DJ, Sioerdsma D. Значение гидроксипролина в моче у человека. Clin Invest. 1961; 40: 843–9.
CAS
Статья
Google ученый
Кьяер М., Хансен М. Тайна женской соединительной ткани. J Appl Physiol.2008; 105: 1026–7.
Артикул
PubMed
Google ученый
Джейсон Л.А., Ричман Дж. А., Радемейкер А. В., Джордан К. М., Плиоплис А. В., Тейлор Р. Р., Маккриди В., Хуанг С. Ф., Плиоплис С. Исследование синдрома хронической усталости на уровне сообщества. Arch Intern Med. 1999; 159: 2129–37.
CAS
Статья
PubMed
Google ученый
Уэссели С. Эпидемиология синдрома хронической усталости.Epidemiologic Rev.1995; 17: 139–51.
CAS
Статья
Google ученый
Будес П. Комплаенс в терапевтических испытаниях: обзор. Контрольные клинические испытания. 1998. 19: 257–68.
CAS
Статья
PubMed
Google ученый
Селла М., Чалдер Т. Измерение утомляемости в клинических условиях и в общественных местах. J Psychosom Res. 2010; 69: 17–22.
Артикул
PubMed
Google ученый
Current Opinion in Clinical Nutrition & Metabolic Care
Основная функция 20 распространенных аминокислот — встраиваться в белки.Однако, помимо этого, аминокислоты играют много разных ролей; действительно, они являются наиболее универсальными питательными веществами. Это является следствием их очень разных боковых цепей, которые допускают широкий спектр химических модификаций и реакций, гораздо больше, чем в случае с другими питательными веществами. Конечно, эти разные боковые цепи играют в белках особую и критическую роль. Аминокислоты с разветвленной цепью открывают возможности для гидрофобных взаимодействий в ядрах глобулярных белков. Остатки глицина вносят гибкость в конформацию пептидных цепей, тогда как пролин вносит излом.Остатки со спиртовыми группами, такими как серин, треонин и тирозин, обеспечивают локус для введения фосфорильных групп. Есть много других примеров. Эта универсальность функции различных аминокислотных остатков в белках отражается в небелковых функциях аминокислот. Многие из этих «вспомогательных» функций касаются передачи сигналов либо напрямую, либо посредством использования в качестве субстратов для синтеза ключевых сигнальных молекул. Вряд ли случайно, что три из четырех известных газовых сигнальных молекул образуются из аминокислот или их производных: этилен из S-аденозилметионина, оксид азота из аргинина и сероводород из S-содержащих аминокислот.Другие важные функции свободных аминокислот включают роль аминокислот и их производных в качестве нейротрансмиттеров (например, глутамата и ГАМК), их роль в качестве сигнальных агентов (например, активация mTOR) и, через S-аденозилметионин, их роль в широком спектре реакций метилирования. Возможно, самым заметным достижением в физиологии аминокислот за последнее десятилетие или около того стало появление роли ряда D-аминокислот. За исключением глицина, все аминокислоты являются хиральными молекулами, и один из ключевых вопросов, с которыми сталкиваются исследователи, заключается в том, играет ли только горстка D-аминокислот функциональную роль или существует гораздо более широкая, хотя и нераспознанная, физиология D-аминокислот. аминокислоты.
Эти темы, касающиеся аминокислот как субстратов для синтеза белка и предшественников ключевых регуляторных молекул, хорошо представлены в исследованиях в этом разделе журнала. В двух обзорах исследуется синтез белка при патологических ситуациях, сердечной недостаточности и диабете 2 типа. Два обзора касаются метаболизма метионина, один — об изменении метаболизма этой аминокислоты при алкогольном поражении печени, а другой — о пищевой и метаболической нагрузке, связанной с реакциями метилирования.Два обзора специально исследуют синтез и функцию ключевых сигнальных молекул (оксида азота и D-серина). Наконец, два обзора касаются различных аспектов передачи сигналов: один вводит новые концепции о роли, которую играет транспорт аминокислот в передаче сигналов; В этом обзоре также представлены новые концепции относительно роли лизосом в аминокислотной активации mTOR. Другой исследует интеграцию пищевых, молекулярных и нейрофизиологических подходов, которые так сильно продвинули наши знания об адаптации животных к недостатку аминокислот.Аминокислоты всегда открывали новые горизонты и концепции; они продолжают делать.
Благодарности
Нет .
Конфликт интересов
Нет конфликта интересов.
Аминокислоты как регуляторы и компоненты небелковых путей | Журнал питания
9″> Примеры аминокислот как регуляторов метаболизма
Хрестоматийный пример контроля метаболизма с помощью аминокислот — ингибирование пируваткиназы L-типа аланином. Считается, что это имеет значение при голодании, потому что этот регуляторный механизм, в дополнение к другим механизмам, помогает предотвратить одновременное возникновение глюконеогенеза и гликолиза в гепатоцитах.
Другой известный пример — каталитическая роль глутамата и аспартата в опосредовании переноса восстанавливающих эквивалентов через митохондриальную мембрану через челнок малат / аспартат (1), например, во время аэробного гликолиза, в сердце, скелетных мышцах и головном мозге, а также во время окисление этанола в печени. Редокс-челнок работает в одном направлении, против направления разности окислительно-восстановительных потенциалов через митохондриальную мембрану из-за свойств транслокатора глутамата / аспартата.Эта транспортная система опосредует отток отрицательно заряженного аспартата из митохондрий в обмен на одну молекулу глутамата и протон, который отбирается из митохондриального протонного градиента, генерируемого дыхательной цепью (2,3).
Третий пример регуляции метаболизма аминокислот лежит в основе катаболизма аминокислот, то есть синтеза мочевины (4). Хотя две N-донорные молекулы для синтеза одной молекулы мочевины, аммиака и аспартата являются производными аминокислотной деградации с глутаматом в качестве промежуточного соединения, большинство учебников игнорируют тот факт, что глутамат также является субстратом для синтеза N-ацетилглутамата в митохондрии, важный активатор карбамоилфосфатсинтазы.Примечательно, что в дополнение к его роли в качестве компонента орнитинового цикла, аргинин ускоряет синтез N-ацетилглутамата, потому что после его попадания в митохондрии аргинин стимулирует активность N-ацетилглутаматсинтазы. Таким образом, увеличение поступления аминокислот в воротную вену не только увеличивает скорость производства аммиака и аспартата для синтеза мочевины, но одновременно увеличивает активность существующих молекул карбамоилфосфатсинтазы в митохондриях. Это приводит к сигмоидальной зависимости между концентрацией аммиака в клетке печени и скоростью синтеза мочевины, что позволяет значительно варьировать синтез мочевины при относительно постоянных концентрациях аммиака (4).Это важно, потому что аммиак токсичен не только для мозга, но и является важным промежуточным метаболитом.
Четвертым и прекрасным примером регуляции метаболизма, в котором принимают участие аминокислоты, является адаптация N-метаболизма к метаболическому ацидозу (4,5; для обзора). В этих условиях синтез мочевины снижается, чтобы сэкономить бикарбонат для нейтрализации избытка протонов. В этом случае одновременно работают несколько механизмов управления. К ним относятся снижение транспорта аминокислот через плазматическую мембрану гепатоцитов, снижение синтеза N-ацетилглутамата, снижение активности карбоангидразы V (которая продуцирует CO 2 в митохондриях для карбамоилфосфатсинтазы), снижение активности печеночной глутаминазы, повышение активности синтеза глутамина в перицентральных гепатоцитах и в мышечных клетках, повышение почечной глутаминазы (для выработки аммиака для выведения с мочой) и, наконец, усиление деградации белка в мышцах (для производства большего количества аминокислот, которые после окисления их отрицательно заряженных карбоксилатных групп дают больше бикарбонат).
5″> Аминокислоты и изменение объема клеток.
Новый путь регуляции метаболизма аминокислот в клетках млекопитающих был открыт более десяти лет назад с открытием того факта, что увеличение объема клетки, связанное либо с транспортом аминокислот, либо с метаболизмом аминокислот, оказывает инсулиноподобное воздействие на потоки через основные метаболические пути, и что эффект может имитироваться гипоосмотически индуцированным набуханием клеток (6-8).Представление о том, что изменения осмолярности могут влиять на метаболизм, само по себе не ново, поскольку давно известно, что микроорганизмы могут адаптироваться к большим изменениям внеклеточной осмолярности, изменяя активность ферментов и / или транспортных систем, цель которых заключается в предотвращении большой разницы в осмотическом давлении на клеточной мембране (9; для обзора). Новое наблюдение, однако, заключалось в том, что тот же принцип используется клетками млекопитающих.
В нашем собственном исследовании мы руководствовались старым наблюдением Каца и его коллег (10), которые показали, что синтез гликогена из глюкозы в гепатоцитах зависит от присутствия аминокислот в инкубационной среде.Прошло почти 15 лет, прежде чем механизм этого эффекта был раскрыт. В сотрудничестве с Баке и Хью из Брюсселя (8; также указано в ссылке 11) мы показали, что способность аминокислот стимулировать синтез гликогена в гепатоцитах связана с их способностью увеличивать объем клеток. Позднее этот результат был подтвержден на мышечных клетках (12). Одновременно и независимо Хойссингер и его коллеги изучили влияние набухания клеток на деградацию гликогена в перфузируемой печени и продемонстрировали, что набухание клеток ингибируется, а сокращение клеток стимулирует гликогенолиз (6,11).Эти анаболические и антикатаболические эффекты набухания клеток позже были распространены на другие метаболические пути, такие как деградация белка (13) и синтез (14), синтез жирных кислот (15-17) и окисление (18), а также на процессы, зависящие от мембранный поток, такой как аутофагия (ингибирование), экзоцитоз (стимулирование), рецептор-опосредованный эндоцитоз (стимулирование) и отток желчи (стимулирование) (19,20; для обзора). Эти эффекты могут быть как краткосрочными, так и долгосрочными через изменения транскрипции генов (19,20).Следует подчеркнуть, что анаболические эффекты вызванного аминокислотами набухания клеток не ограничиваются гепатоцитами, но обнаруживаются во многих различных типах клеток (19,20).
Вызванное аминокислотами набухание клеток происходит из-за концентрированного, зависимого от Na + транспорта аминокислот через плазматическую мембрану (управляемого трансмембранным градиентом Na + , например, транспорта глутамина, аланина, таурина) и внутриклеточное производство непроницаемых катаболитов, таких как глутамат и аспартат (8, 19–22).В ответ на начальное набухание клеток, клетки подвергаются регулятивному уменьшению объема и высвобождают KCl в попытке восстановить свой первоначальный объем (19, 20). Механизм, ответственный за стимуляцию аминокислотами синтеза гликогена из глюкозы, может, по крайней мере частично, включать уменьшение хлорида, который де-ингибирует гликоген-синтазную фосфатазу, и повышение внутриклеточного глутамата, который также активирует гликоген-синтазную фосфатазу (21). ). Тот же механизм может быть ответственным за стимуляцию ацетилСоА-карбоксилазы аминокислотами с аналогичным действием хлорида и глутамата на протеинфосфатазу, которая опосредует дефосфорилирование ацетилСоА-карбоксилазы (16,17).Существуют также различия в регуляции двух систем с помощью аминокислот: например, лейцин, который не вызывает набухания клеток, поскольку его транспорт не связан с Na + (8), активирует активность ацетил-СоА-карбоксилазы в гепатоцитах. но не влияет на активность гликогенсинтазы (23) или на синтез гликогена (8). Однако в мышечных клетках лейцин действительно стимулирует синтез гликогена (24).
Внутриклеточный глутамат также может быть ответственным за ингибирование окисления жирных кислот за счет вызванного аминокислотами набухания клеток, поскольку он непосредственно ингибирует карнитин-пальмитоилтрансферазу (18).Было высказано предположение, что продуцируемый митохондриями глутамат может действовать как внутриклеточный мессенджер, который связывает метаболизм глюкозы с секрецией инсулина β -клетками, потому что после его оттока из митохондрий глутамат накапливается в инсулинсодержащих гранулах, заставляя их набухать. и стимулирует экзоцитоз (25,26). Хотя это очень привлекательный механизм, следует подчеркнуть, что высокие концентрации глутамата в цитозоле не всегда коррелируют с высокой скоростью продукции инсулина (27).
Помимо этих специфических механизмов, включающих изменения концентраций хлорида и глутамата, вызванное аминокислотами набухание клеток также может влиять на метаболизм за счет изменения макромолекулярного скопления. Из-за их высокой внутриклеточной концентрации белки часто могут находиться в непосредственной близости друг от друга, и изменения, вызванные объемом клетки, могут влиять на их взаимодействие и, таким образом, на кинетические свойства ферментов (20). В этом контексте также возможно, что цитоскелет участвует в восприятии изменений объема клетки и что это влияет на связанные с ним ферменты (19,20).Примерами ферментов, связанных с цитоскелетом, являются гликогенсинтаза (28) и ацетилCoA-карбоксилаза (29). Было бы очень интересно выяснить, действительно ли вызванные аминокислотами изменения объема клетки влияют на компартментацию ферментов в цитозоле (30).
Иллюстрацию влияния набухания клеток на продукцию гликогена можно найти при печеночной энцефалопатии, когда астроциты синтезируют и накапливают большое количество глутамина из-за повышения уровня аммиака в крови (31). В этих условиях астроциты наполнены гликогеном (32).Поскольку гликоген содержит большое количество кристаллической воды, это может еще больше усилить набухание астроцитов и, следовательно, повышение внутричерепного давления, которое наблюдается в этих условиях.
5″> Подавление передачи сигналов инсулина с помощью зависимой от аминокислот передачи сигналов.
Несмотря на то, что аминокислоты и инсулин взаимодействуют друг с другом при активации последующих мишеней mTOR, появляется все больше доказательств того, что в мышечных клетках, адипоцитах и клетках гепатомы аминокислоты вызывают чувствительное к рапамицину подавление инсулино-индуцированной активации PI 3-киназы. , протеинкиназа B и транспорт глюкозы (67–71).Это вызвано повышенным фосфорилированием ser / th IRS-1 (субстрат-1 инсулинового рецептора), снижением связывания регуляторной субъединицы p85 PI 3-киназы с IRS-1 с последующей (предположительно протеасомной) деградацией IRS-1 ( 68,69). Поскольку индуцированное аминокислотами подавление IRS-1 вызывает ингибирование транспорта глюкозы (см. Выше), этот механизм может лежать в основе снижения потребления глюкозы во время кормления с высоким содержанием белка (67,68).
Таким образом, кажется, что аминокислоты и инсулин действуют синергически в отношении активации mTOR и его последующих мишеней и тем самым влияют на метаболизм белка; тем не менее, аминокислоты, по-видимому, ингибируют начальную часть сигнального пути инсулина.Эта часть, как известно, контролирует транспорт и метаболизм глюкозы (67,72). Как это ни парадоксально, как обсуждалось ранее, аминокислоты действительно стимулируют превращение глюкозы в гликоген. Хотя это сбивает с толку, парадокс можно разрешить за счет различий в аминокислотной специфичности. Лейцин особенно эффективен для активации mTOR-зависимой передачи сигналов и подавления метаболизма глюкозы, тогда как аминокислоты, увеличивающие объем клеток (например, глутамин, аланин), стимулируют синтез гликогена. Интересно, что кормление рыбным белком защищает от непереносимости глюкозы и инсулинорезистентности, вызванных диетой с высоким содержанием жиров, а концентрации лейцина, тирозина и некоторых других аминокислот в плазме были значительно ниже у крыс, получавших белок трески, по сравнению с теми, кто получал казеин (68 ; и ссылки в нем).
0″> Заключительные замечания
Из предыдущего обсуждения ясно, что аминокислоты, помимо их роли в качестве промежуточных метаболитов, являются важными анаболическими сигналами.Механизмы основаны на способности некоторых аминокислот вызывать набухание клеток (например, глутамин и аланин) и на способности других аминокислот (в частности, лейцина) имитировать некоторые, но не все эффекты инсулина на передачу сигналов ( см. сводку на рис. 1). Способность стимулировать передачу сигналов и выработку инсулина в β -клетках поджелудочной железы дополнительно усиливает анаболические свойства аминокислот.
3″> Сокращения
-
4E-BP
фактор инициации трансляции эукариотических белков 4E-связывающий белок-1
-
mTOR
мишень рапамицина у млекопитающих
-
PI 3-киназа
фосфатидная (3) P
фосфатидилинозитол 3-фосфат
-
PI (4,5) P 2
фосфатидилинозитол 4,5-бисфосфат
-
000 PI (3,4,5) P 3 3 0 фосфатидилинозитол 3,4,5-трифосфат
-
p70S6k
© 2003 Американское общество диетологии
D-аминокислот в нервной и эндокринной системах
Аминокислоты являются важными компонентами пептидов и белков и действуют как передатчики сигналов.Для млекопитающих, включая человека, необходимы только L-аминокислоты. Однако у млекопитающих обнаружены различные D-аминокислоты, такие как D-серин, D-аспартат, D-аланин и D-цистеин. Постепенно выясняется физиологическая роль этих D-аминокислот не только в нервной, но и в эндокринной системе. Рецепторы N-метил-D-аспартата (NMDA) связаны с обучением и памятью. D-серин, D-аспартат и D-аланин могут связываться с рецепторами NMDA. H 2 S, образующийся из D-цистеина, уменьшает дисульфидные связи в рецепторах и усиливает их активность.Аберрантная активность рецепторов связана с заболеваниями центральной нервной системы (ЦНС), такими как болезнь Альцгеймера, боковой амиотрофический склероз и шизофрения. Кроме того, D-аминокислоты обнаруживаются в частях эндокринной системы, таких как шишковидная железа, гипоталамус, гипофиз, поджелудочная железа, надпочечники и яички. D-аспартат исследуется на предмет регуляции высвобождения гормонов из различных эндокринных органов. Здесь мы сосредоточились на недавних открытиях, касающихся синтеза и физиологических функций D-аминокислот в нервной и эндокринной системах.
1. Введение
Аминокислоты важны не только как важные компоненты для строительных блоков пептидов и белков, но также как биохимические регуляторы, такие как нейротрансмиттеры [1–4] и регуляторы аутофагии [5–8]. D-аминокислоты являются энантиомерами L-аминокислот и долгое время считались отсутствующими и неприродными аминокислотами у млекопитающих. Однако недавнее развитие чувствительных аналитических методов выявило присутствие D-аминокислот, таких как D-серин, D-аспартат и D-аланин, у млекопитающих [9–11].Более того, исследования ферментов, которые синтезируют или метаболизируют D-аминокислоты, также прояснили локализацию и функции D-аминокислот в нервной и эндокринной системах и обнаружили, что синтез и метаболизм D-аминокислот регулируются физиологически [12-15] . Здесь мы сосредоточились на последних достижениях в понимании синтеза, метаболизма и физиологической роли D-аминокислот в нервной и эндокринной системах.
2. D-серин
Отмеченные уровни D-серина были обнаружены в центральной нервной системе (ЦНС) грызунов и людей.Областями ЦНС, богатыми D-серином, были кора головного мозга, гиппокамп и полосатое тело. Кроме того, D-серин также обнаруживается в других регионах, таких как средний мозг, мозжечок и спинной мозг, у грызунов и людей [16-19]. Внеклеточные уровни D-серина в медиальной префронтальной коре и полосатом теле крыс составляют примерно 20% от общего уровня серина [20]. D-серин биосинтезируется серинрацемазой (SR) в ЦНС грызунов и человека [21–23]. Кроме того, уровень D-серина в ЦНС значительно снижен у мышей с нокаутом Sr [24, 25].
Было показано, что SR локализуется как в астроцитах, так и в нейронах, и D-серин высвобождается из обоих [22, 23, 26–29]. Кроме того, D-серин в нейронах генерируется из L-серина, полученного из астроцитов [27]. Модель D-серинового челнока описывает оптимальную D-серин-опосредованную активность рецептора N-метил-D-аспартата (NMDA), предполагая, что D-серин транспортируется между нейронами и астроцитами [30, 31]. Эта модель челнока D-серина выглядит следующим образом. Астроциты захватывают глюкозу из кровеносных сосудов через переносчик глюкозы 1, а затем через 3-фосфоглицератдегидрогеназу (Phgdh), которая в основном локализуется в астроцитах и превращает глюкозу в L-серин.L-серин экспортируется из астроцитов и импортируется в нейроны через транспортеры аланина / серина / цистеина / треонина (ASCT). В нейроне L-серин превращается в D-серин с помощью SR. D-серин высвобождается из нейронов через транспортер-1 аланин-серин-цистеин (Asc-1) или другими путями в синапс, где он может регулировать активность рецептора NMDA. Высвобожденный D-серин также может быть импортирован в астроциты через ASCT. Способность SR рацемизировать L-серин в D-серин положительно регулируется пиридоксаль-5′-фосфатом (PLP), двухвалентными катионами и АТФ [22, 32, 33].С другой стороны, активность SR подавляется его перемещением из цитозоля на мембраны, содержащие фосфатидилинозитол-4,5-бисфосфат (PIP 2 ), такие как ядерный, эндоплазматический ретикулум (ER) и плазматические мембраны [34 –37]. SR также регулируется посредством взаимодействия с другими белками (рис. 1). Было обнаружено, что белок 1, взаимодействующий с глутаматным рецептором (GRIP1) [38], и белок, взаимодействующий с С-киназой (PICK1) [39, 40], активируют SR, взаимодействуя с ним в астроцитах грызунов.GRIP1 связывается с рецептором α -амино-3-гидрокси-5-метилизоксазол-4-пропионовой кислоты (AMPA) и высвобождается после стимуляции L-глутаматом. Освободившийся GRIP1 связывает и активирует SR [38]. Было обнаружено, что белок постсинаптической плотности 95 (PSD95) / Discs large / ZO-1 (PDZ) в С-концевой области GRIP1 отвечает за взаимодействие с SR [38, 41]. PICK1 имеет домен PDZ, который связывается с SR [39], и домен Bin / amphiphysin / Rvs (BAR), который взаимодействует с мембранами [42]. PICK1 связывается с эритропоэтин-продуцирующим гепатоцеллюлярным рецептором (Eph) B3 или EphA4 в астроцитах.После того, как ephrinB3, который экспрессируется на постсинаптических нейронах, связывается с рецептором EphB3 или EphA4, PICK1 высвобождается в цитозоль. Усиление ассоциации между высвобожденным PICK1 и SR увеличивает синтез D-серина в астроцитах мышей [40]. Старгазин и белок 22, содержащий только F-бокс (FBXO22), также регулируют активность SR, влияя на субклеточную локализацию SR. Старгазин образует комплекс с покоящимся рецептором AMPA, PSD-95 и SR, ингибируя активность SR, способствуя локализации мембран в нейронах мышей.После активации рецептора AMPA SR высвобождается из старгазина и плазматической мембраны, что приводит к активации SR [43]. FBXO22 связывает и активирует SR, предотвращая его связывание с мембраной ER в нейронах и астроцитах крыс. FBXO22 может блокировать липид-связывающую область SR [35].
D-серин катализируется SR и оксидазой D-аминокислот (DAO). SR имеет α , β -элиминирующую и рацемизирующую активности. SR превращает как D-серин, так и L-серин в пируват и аммиак, удаляя воду из этих аминокислот путем α , β -элиминирования [44].Следовательно, SR может регулировать физиологический уровень D-серина за счет активности рацемизации для синтеза D-серина и активности α , β -элиминации для разрушения D-серина. DAO — это флавопротеин, который превращает D-серин с образованием соответствующей альфа-кетокислоты, перекиси водорода и аммиака [11].
Было обнаружено, что распределение D-серина в ЦНС сходно с распределением рецепторов NMDA, которые являются ионотропными рецепторами глутамата (iGluR) [18, 45], а для активации рецепторов NMDA требуется связывание как глутамата, так и D -серин или глицин.Активированные рецепторы NMDA индуцируют поток Ca 2+ , что приводит к регуляции долговременной потенциации (LTP) и долговременной депрессии (LTD) в различных областях ЦНС. Таким образом, регуляция рецепторов NMDA тесно связана с синаптической активностью, обучением и памятью [46–49].
Рецептор NMDA в основном состоит из двух субъединиц GluN1 и двух субъединиц GluN2 одной и той же или разных субъединиц GluN2. Глутамат связывается с GluN2, тогда как D-серин или глицин связывается с GluN1 [50–52].Хотя D-серин или глицин могут действовать как коагонист рецепторов NMDA [46], снижение уровня D-серина снижает активность рецептора NMDA, а добавление D-серина отменяет эту инактивацию, приводя к LTP [46, 53]. Более того, D-серин индуцирует LTP, а деградация D-серина под действием DAO приводит к подавлению этой индукции [49]. Следовательно, D-серин играет важную роль в регуляции синаптической активности, обучения и памяти, регулируя активацию рецепторов NMDA.Однако сообщалось о нейротоксических эффектах D-серина. По сравнению с мышами дикого типа, сниженные уровни D-серина у взрослых мышей с нокаутом Sr снижали нейротоксичность, опосредованную рецептором NMDA, и индуцированную β -амилоидом 1–42 [24]. Более того, повышенные уровни D-серина у взрослых мышей с нокаутом Dao приводили к дегенерации двигательных нейронов [54]. Аберрантные уровни D-серина связаны с заболеваниями, вызванными аномальной активностью рецептора NMDA. Сообщалось, что уровни D-серина в спинномозговой жидкости (CSF) пациентов с болезнью Альцгеймера выше, чем у нормальных контролей [55, 56].Кроме того, нейротоксичность, вызванная бета-амилоидом, подавляется у мышей с нокаутом Sr , которые показали снижение уровня D-серина в головном мозге на 90% [24]. Шизофрения связана с гипофункцией рецепторов NMDA [57]. Снижение уровня D-серина приводит к гипофункции рецепторов NMDA, что приводит к симптомам шизофрении [25, 58]. Более того, введение D-серина улучшает положительные, отрицательные и когнитивные симптомы у пациентов с шизофренией [59]. Боковой амиотрофический склероз (БАС) связан с повышенным уровнем D-серина [60, 61].Следовательно, контроль уровня D-серина может быть одной из терапевтических целей при этих заболеваниях.
С другой стороны, D-серин связывается с глутаматным рецептором δ 2-типа (GluD2), который представляет собой iGluR, и вызывает уменьшение количества рецепторов AMPA в клетках Пуркинье в мозжечке за счет эндоцитоза. Это приводит к продвижению компании cerebellar LTD. Более того, связывание D-серина с GluD2 приводит к приобретению двигательного обучения у мышей [62]. Недавно сообщалось, что возрастное увеличение активности пируваткарбоксилазы в глиальных клетках вызывает снижение уровня D-серина, что приводит к возрастному ухудшению памяти у мух [63].Пируваткарбоксилаза образует щавелевоуксусную кислоту, которая может быть преобразована в аспарагиновую кислоту. И щавелевоуксусная кислота, и аспарагиновая кислота могут ингибировать SR, что приводит к снижению уровня D-серина [64].
D-серин также был обнаружен в эндокринных органах, таких как надпочечники и гипофиз, поджелудочная железа и яички крыс [65, 66]. Однако уровни D-серина в эндокринной системе намного ниже, чем в ЦНС, и физиологическая роль D-серина в эндокринной системе остается неясной.
3. D-аспартат
D-аспартат обнаружен в ЦНС грызунов и людей [16, 19, 67–72], а также в эндокринных органах, включая шишковидную железу [69, 72, 73], гипофиз. железы [68, 69, 71, 72, 74], поджелудочной железы [66], надпочечников [19, 67, 72, 74, 75] и семенников крыс [74, 76]. D-аспартат активирует рецепторы NMDA посредством связывания с сайтом агониста субъединиц GluN2 (2A – D) [77–79], а также может активировать метаботропный рецептор глутамата 5 (mGlu5) [80]. Хотя фермент, который генерирует D-аспартат, еще не идентифицирован [81–83], образование D-аспартата может зависеть от PLP [67].
С другой стороны, было продемонстрировано, что D-аспартат разлагается до оксалоацетата, пероксида водорода и аммония пероксисомальной флавопротеин D-аспартатоксидазой (DDO) [84–86]. Хотя D-аспартат в изобилии присутствует в мозге грызунов и людей во время развития, уровни D-аспартата сильно снижаются на постнатальных стадиях, и это снижение сохраняется на протяжении всей взрослой жизни [16, 67, 68, 71, 87, 88]. Напротив, активность DDO и его мРНК чрезвычайно низки на постнатальных стадиях и повышаются после рождения у грызунов [88, 89].Более того, было показано, что уровни D-аспартата были значительно повышены в ЦНС мышей с нокаутом Ddo [88, 90–93]. Кроме того, D-аспартат обнаружен исключительно в нейронах крыс [67]. Улучшение пространственной памяти было продемонстрировано у мышей с нокаутом Ddo [77, 78]. Более того, мыши, получавшие D-аспартат, и мыши с нокаутом Ddo продемонстрировали усиление LTP [77–79] и увеличение длины дендритов и плотности шипов в нейронах префронтальной коры и гиппокампа [94].Следовательно, значительное увеличение уровней D-аспартата, усиление LTP и увеличение длины дендритов и плотности шипов нейронов у взрослых мышей с нокаутом Ddo указывают на то, что D-аспартат также может генерироваться и функционировать в ЦНС во взрослом возрасте. Однако стойкое повышение уровней D-аспартата у взрослых мышей с нокаутом Ddo имеет нейротоксические эффекты, такие как активация каспазы-3 и апоптоз в нейронах [88]. Экспрессия мРНК Ddo в посмертном мозге пациентов с шизофренией была значительно выше, чем у контрольных лиц [95].Более того, сообщалось, что уровни D-аспартата были значительно снижены в посмертном мозге пациентов с шизофренией по сравнению с таковыми в контрольной группе [96]. Мыши с нокаутом Ddo демонстрируют сниженное шизофреническое поведение, вызванное фенциклидином, такое как двигательная гиперактивность и предымпульсное торможение [95]. Эти данные показывают, что аномальный уровень D-аспартата может быть связан с шизофренией.
В эндокринной системе D-аспартат контролирует синтез и высвобождение гормонов.D-аспартат подавляет синтез мелатонина в шишковидной железе крысы [97], а также снижает высвобождение мелатонина из культивируемых пинеалоцитов крысы [98]. В гипоталамусе крыс D-аспартат индуцирует экспрессию генов окситоцина и вазопрессина [99] и может увеличивать высвобождение гонадотропин-рилизинг-гормона [100]. D-аспартат также вызывает высвобождение пролактина [101], гормона роста и лютеинизирующего гормона [100, 102] из передней доли гипофиза крысы. Напротив, увеличение уровней D-аспартата связано со значительным снижением проопиомеланокортина и α -меланоцит-стимулирующего гормона в промежуточной доле мыши [93].В семенниках крыс D-аспартат индуцирует синтез и высвобождение тестостерона [102–104]. Повышенная экспрессия андрогенов и пониженная экспрессия рецепторов эстрогена при введении D-аспартата были показаны в семенниках крыс [104]. Более того, пероральный прием D-аспартата вызывает высвобождение тестостерона в сыворотке человека [102] и улучшает количество и подвижность сперматозоидов у человека [105]. Следовательно, ожидается, что D-аспартат может быть кандидатом для лечения бесплодия.
4.Другие D-аминокислоты
Различные D-аминокислоты, кроме D-серина и D-аспартата, также были обнаружены в нервной и эндокринной системах.
D-Аланин обнаружен в головном мозге, гипофизе, поджелудочной железе, надпочечниках и семенниках грызунов [106]. Кроме того, он обнаруживается также в головном мозге человека [107]. Большая часть D-аланина у грызунов происходит из кишечных бактерий [108, 109]. D-аланин метаболизируется DAO [11], а уровень D-аланина у грызунов зависит от циркадного ритма [9, 110].Кроме того, сообщалось, что D-аланин связывается с рецептором NMDA и улучшает симптомы у пациентов с шизофренией [111].
D-Цистеин оказался одним из источников H 2 S в головном мозге (рис. 2). D-цистеин, вероятно, хотя бы частично абсорбируется с пищей, хотя источник D-цистеина в организме до сих пор не выяснен [112]. В ЦНС D-цистеин расщепляется DAO с образованием 3-меркаптопирувата (3MP). Затем 3-МП катализируется 3-меркаптопируват-серотрансферазой (3МСТ) с образованием H 2 S [113].3MST обнаружен в синаптосомах и нейронах мозга мышей [114]. Более того, полисульфиды водорода (H 2 ; –5) обнаруживаются в мозге мышей и также генерируются 3MST из 3MP [115]. H 2 S усиливает активность рецепторов NMDA за счет уменьшения дисульфидных связей в рецепторах NMDA и индуцирует LTP у крыс [116, 117]. Напротив, H 2 активирует каналы A1 переходного рецепторного потенциала (TRP) в астроцитах грызунов и индуцирует поток Ca 2+ [118].Активация каналов TRPA1 приводит к высвобождению D-серина из астроцитов грызунов и усиливает зависимый от рецептора NMDA LTP [119]. Кроме того, Паркин является одним из ключевых факторов болезни Паркинсона [7], а H 2 S усиливает активность Паркина и приводит к защитным эффектам против болезни Паркинсона [120]. В эндокринной системе H 2 S ингибирует индуцированное глюкозой высвобождение инсулина из клеток поджелудочной железы β [121].
D-лейцин и D-пролин обнаружены в головном мозге, шишковидной железе и гипофизе грызунов [122, 123].D-глутамат обнаружен в головном мозге грызунов [70, 124]. Однако детальные физиологические роли этих аминокислот остаются неясными.
5. Выводы
Считается, что только L-аминокислоты используются у млекопитающих, включая человека. Однако из-за недавней разработки чувствительных и селективных аналитических методов обнаружения хиральных аминокислот [125] в тканях млекопитающих были обнаружены различные D-аминокислоты. Физиологические функции этих D-аминокислот постепенно выясняются.Было продемонстрировано, что D-аминокислоты, такие как D-серин, D-аспартат, D-аланин и D-цистеин, играют важную роль в нервной и эндокринной системах. Поэтому очень важно, чтобы механизмы синтеза и метаболизма, а также физиологические функции D-аминокислот были дополнительно исследованы. Эти исследования предоставят новые терапевтические и диагностические стратегии для заболеваний, связанных с нервной и эндокринной системами.
Конкурирующие интересы
Авторы заявляют, что у них нет конкурирующих интересов.
Белки и аминокислоты | Рекомендуемые диетические нормы: 10-е издание,
стр. 74
Fomon, S.J. 1974. 74. Детское питание, , 2-е изд., . W.B. Сондерс, Филадельфия. 575 с.
Fomon, S.J. 1986. Потребность доношенных детей в белке. Стр. 55-68 в S.J. Фомон и У. Heiod, eds. Потребности в энергии и белке в младенчестве. Академическая пресса. Нью-Йорк.
Фомон, С.Дж., и Л.Дж. Filer, Jr. 1967. Потребность в аминокислотах для нормального роста.Стр. 391-101 в W.I .. Nyhan ,. изд. Аминокислотный метаболизм и генетическая изменчивость. Макгроу-Хилл, Нью-Йорк.
Гарза, К., Н.С. Скримшоу и В. Молодой. 1977 г. Потребности в человеческом белке: долгосрочное исследование метаболического баланса азота у молодых мужчин для оценки безопасного уровня потребления яичного белка, установленного ФАО / ВОЗ в 1973 году. J. Nutr. 107: 335-352.
Герсовиц М., Х.Н. Манро, Дж. Удалл и В. Молодой. 1980. Синтез альбумина у молодых и пожилых людей с использованием новой методологии стабильных изотопов: реакция на уровень потребления белка.Метаболизм 29: 1075-1086.
Герсовиц, М., К. Мотил, Х.Н. Манро, Н.С. Скримшоу и В. Молодой. 1982. Потребности в человеческом белке: оценка адекватности действующей Рекомендуемой диетической нормы пищевого белка для пожилых мужчин и женщин. Являюсь. J. Clin. Nutr. 35: 6-14.
Hegsted, D.M. 1976. Балансовые исследования. J. Nutl. 106: 307-311.
Hegsted, D.M., S.A. Schuette, M.B. Земель, Х. Linkswiler. 1981. Кальций и баланс кальция в моче у молодых мужчин в зависимости от уровня потребления белка и фосфора.J. Nutr. 111: 553-562.
Хейрд, W.C., Дж. Ф. Николсон, Дж. М. Дрисколл, мл., Н. Дж. Шуллингер и Р. В. Винтерс. 1972. Гипераммонемия в результате внутривенного введения смеси синтетических l-аминокислот: предварительный отчет. J. Pediatr. 81: 162-165.
Хиггинс А.К., Ф. В. Крэмптон и Дж. Ф. Моксли. 1973. Питание и исход беременности. Стр. 1071-1077 в Р.О. Scow, изд. Эндокринология. Труды Четвертого Международного конгресса эндокринологов, Вашингтон, Д.C., 18-24 июня 1972 г. Excerpta Medica, Амстердам.
Хопкинс, Д. Т. 1981. Влияние изменения усвояемости белка. Стр. 169-193 в C.F. Бодвелл, Дж. Адкинс, Д.Т. Хопкинс, ред. Качество белка у людей: оценка и оценка in vitro. Издательство AVI, Вестпорт, штат Коннектикут,
Horowitz, J.H., F.B. Рыпинс, Дж.М. Хендерсон, С. Хеймсфилд, С. Моффитт, Р.П. Бейн, Р.К. Чавла, Дж. К. Блейер и Д. Рудман. 1981. Доказательства нарушения пути транссульфурации при циррозе печени.Гастроэнтерология 81: 668-675.
Hytten, F.E., and I. Leitch. 1971. Физиология беременности человека, 2-е изд. Научные публикации Блэквелла, Оксфорд. 599 с.
Inone, G.T., Komatsu, K. Kishi, and Y. Fujita. 1988. Аминокислотные потребности японских юношей. Стр. 55-62 в издании G.L. Blackburn, J.P. Grant и V.R. Янг, ред. Аминокислоты: метаболизм и медицинское применение. Джон Райт, Бостон.
Ирвин, М.И. и Д.М. Хегстед. 1971. Обзор исследований потребности человека в белке.J. Nutr. 101: 387-429.
Джексон, А.А. 1987. Измерение белкового обмена во время беременности. Human Nutr. Clin. Nutr. 4IC: 497-498.
Джонстон, F.D., D.M. Кэмпбелл и И. Макгилливрей. 1981. Исследования баланса азота при беременности человека. J. Nutr. 111: 1884–1893.
Kindt, F., and S. Halvorsen. 1980. Потребность в незаменимых аминокислотах у детей. Оценка, основанная на приеме фенилаланина, тирозина, лейцина, изолейцина и валина у детей с фенилкетонурией, дефектом тирозин-аминотрансферазы и болезнью мочи кленового сиропа.Являюсь. J. Clin. Nutr. 33: 279-286.
Кинг, Дж. К. 1975. Метаболизм белков во время беременности. Clin. Перинатол. 2: 243-254.
Amino Acids Reference Tool
Аминокислоты — это органические соединения, содержащие в основном атомы углерода, водорода, кислорода и азота. У них также есть аминные (–NH 2 ) и карбоксильные (–COOH) функциональные группы с боковыми цепями (R-группы), которые отличают одну аминокислоту от другой. Сера и другие дополнительные элементы можно найти в некоторых боковых цепях.
Аминокислоты являются строительными блоками белков. Они могут соединяться пептидными связями с образованием полипептидных цепей. В зависимости от взаимодействия соседних аминокислот и их боковых цепей полипептиды сворачиваются и образуют трехмерные молекулы, известные как белки. Форма белка и его аминокислотные компоненты помогут определить его функцию.
Помимо своей важной роли в образовании белка, аминокислоты важны для биосинтеза, транспорта нейротрансмиттеров и других биохимических процессов.
Категории аминокислот
Существует около 500 аминокислот, но только 20 присутствуют в нашем генетическом коде. Эти 20 протеиногенных аминокислот известны как стандартные аминокислоты.
Полярность аминокислоты зависит от структуры ее боковых цепей. Молекула будет более полярной, если разница в электроотрицательности между атомами больше. Например, алкановые ответвления и бензольные кольца считаются неполярными, а спирты и амины полярными.
Аминокислоты классифицируются по:
- Заряд (положительный vs.отрицательный)
- pH (кислотный против основного)
- Растворимость (полярный против неполярного)
- Расположение основных функциональных групп: альфа (α), бета (β), гамма (γ) или дельта (δ)
- Структура боковой цепи (алифатическая, ароматическая и т. Д.)
Гидрофобность
| Очень гидрофобная | Гидрофобная | Нейтральная | Гидрофильная | |
|---|---|---|---|---|
| 9364 93 644 9364 Тирозол 4364 Тирозол 4364 Тиреол 4364 Фенилал43644 9364 Тирозин | Цистеин | Гистидин | Лизин | |
| Триптофан | аланин | Глицин | аспарагин | |
| Лейцин | серин | Глутаминовая кислота | ||
| валин | глютамин | пролин | ||
| Метионин | Аспарагиновая кислота |
На основе на значениях при pH 7.
Справочная таблица аминокислот
AlanineAla & nbsp A9.872.35
| Полярность боковой цепи | Неполярная | ||
| Молекулярная 3 | Нейтральная масса | Нейтральная масса 3 | |
| Классификация | Алкил |
Описание: Аланин представляет собой небольшую заменимую аминокислоту с относительно нереактивной одной боковой цепью метильной группы.Это одна из простейших аминокислот и вторая по содержанию белков. Он кодируется кодонами, которые начинаются с GC.
Магазинный аланин и производные
ArginineArg & nbsp R9.092.18
| Полярность боковой цепи | Полярность | ||
| Заряд | |||
| Заряд | моль / моль | ||
| Классификация | Основной |
Описание: Аргинин представляет собой химически сложную аминокислоту с 3-углеродной алифатической боковой цепью, оканчивающейся гуанидиновой группой.Обычно он опосредует взаимодействия между белками и отрицательно заряженными остовами ДНК и РНК.
Магазин аргинина и производных
AsparagineAsn & nbsp N8.802.02
| Полярность боковой цепи | Полярность | ||||
| Заряд | Молекулярная масса | 936 936 | Классификация | Нейтральная |
Описание: Аспарагин — это аминокислота с карбоксамидной боковой цепью.Амидная группа сохраняет свою полярность и является богатым источником водородных связей, что делает аспарагин обычным сайтом связывания белка и субстратом для гликозилтрансфераз.
Магазин аспарагин и производные
Аспарагиновая кислота Asp & nbsp D9.601.88
| Полярность боковой цепи | Полярность | ||||
| Заряд | |||||
| Заряд | Молекулярная масса | Отрицательная молекулярная масса | |||
| Классификация | Кислая |
Описание: Аспарагиновая кислота имеет боковую цепь карбоновой кислоты, которая взаимодействует с другими аминокислотами, ферментами и белками.Его отрицательный заряд и большой потенциал водородных связей позволяют ему участвовать во многих молекулярных взаимодействиях.
CysteineCys & nbsp C10.781.71
| Полярность боковой цепи | Полярность |
| Заряд | Нейтральная |
| Нейтральная масса4 | |
| 93,16 93,16 93,16 моль |
Описание: Цистеин — это полузаменимая протеиногенная аминокислота с тиоловой боковой цепью, которая участвует в ферментативных реакциях.Его сульфгидрильная группа образует дисульфидные связи с другими цистеинами или молекулами со свободными сульфгидрилами.
Glutamic AcidGlu & nbsp E9.672.19
| Полярность боковой цепи | Полярность | ||||
| Заряд | Отрицательная | ||||
| Молекулярная масса3 14364 | Молекулярная масса3 14364 | Молекулярная масса3 4 г / моль |
Описание: Глутаминовая кислота имеет одну амино и две карбоксильные группы, которые придают ей отрицательный заряд практически во всех биологических системах.Это позволяет глутаминовой кислоте взаимодействовать с положительно заряженными объектами, включая ионы металлов.
Shop Глутаминовая кислота и производные
GlutamineGln & nbsp Q9.132.17
| Полярность боковой цепи | Полярность |
| Заряд | |
| Заряд | Мол. |
| Классификация | Нейтрально |
Описание: Глютамин имеет боковую цепь, подобную цепи глутаминовой кислоты; одна карбоксильная группа просто заменяется амидом.Он важен для метаболизма азота и помогает регулировать токсический уровень аммиака в организме.
Заводской глутамин и производные
GlycineGly & nbsp G9.602.34
| Полярность боковой цепи | Неполярная | |||
| Заряд | Молекулярная масса | Нейтральная масса | Нейтральная масса | |
| Классификация | Алкил |
Описание: Глицин — простейшая аминокислота с одним атомом водорода в боковой цепи.Это делает его идеальным для размещения крутых поворотов некоторых полипептидных цепей. Глицин также действует как тормозящий нейротрансмиттер.
HistidineHis & nbsp H8.971.78
| Полярность боковой цепи | Полярность |
| Заряд | Положительный |
| Молекулярный вес (г / моль) 9364 9364 9364 | |
Описание: Гистидин встречается редко и имеет частично протонированную ароматическую имидазольную боковую цепь.Он может быть нейтральным или положительно заряженным и может служить естественным катализатором или преобладающим остатком в активных центрах ферментов благодаря своему потенциалу связывания.
Заводской гистидин и производные
ИзолейцинIle & nbsp I9.762.32
| Полярность боковой цепи | Неполярная |
| Заряд | |
| Заряд | Мол. |
| Классификация | Алкил |
Описание: Изолейцин имеет разветвленную углеводородную боковую цепь, расположенную в L-образной форме, и считается одной из крупнейших алифатических аминокислот.Он сконцентрирован в мышечной ткани и кодируется кодонами AUU, AUC и AUA.
Shop Изолейцин и производные
LeucineLeu & nbsp L9.602.36
| Полярность боковой цепи | Неполярная |
| Заряд | Мол. |
| Классификация | Алкил |
Описание: Лейцин имеет боковую цепь изобутильной группы с четырьмя полностью протонированными атомами углерода, расположенными в объемной Y-образной форме.Это также самая распространенная аминокислота в белках и одна из крупнейших алифатических аминокислот.
LysineLys & nbsp K10.288.90
| Полярность боковой цепи | Полярность |
| Заряд | Положительный |
| Молекулярный вес (г / моль3) 14632 9364 9364 9364 9364 Базовый класс 146329364 |
Описание: Лизин является незаменимой аминокислотой с лизильной боковой цепью.Он важен для протеиногенеза, сшивания полипептидов коллагена, поглощения минеральных питательных веществ и выработки карнитина, ключевого для метаболизма жирных кислот.
MethionineMet & nbsp M9.212.28
| Полярность боковой цепи | Неполярная |
| Заряд | Нейтральная |
| Молекулярная масса1 143,21 93,21 Алкил |
Описание: Метионин имеет линейную боковую цепь и содержит сульфидную группу.Это позволяет метионину быть плодовитым донором метильных групп. Он также играет важную роль в обмене веществ, росте новых кровеносных сосудов и восстановлении тканей.
Заводской метионин и производные
PhenylalaninePhe & nbsp F9.242.58
| Полярность боковой цепи | Неполярная |
| Заряд | Нейтральная | Нейтральная масса |
| Классификация | Ароматическая |
Описание: Фенилаланин содержит боковую цепь бензильной группы и считается самой простой и наиболее распространенной ароматической аминокислотой в белках.Это предшественник синтеза тирозина, дофамина, норадреналина, адреналина и меланина.
Shop Фенилаланин и производные
ProlinePro P10.601.99
| Полярность боковой цепи | Неполярная |
| Заряд | |
| Заряд | Мол. |
| Классификация | Алкил |
Описание: Пролин представляет собой протеиногенную аминокислоту с циклической пирролидиновой боковой цепью.Он не является незаменимым для человека и может быть синтезирован из глутаминовой кислоты. Незаменим для иммунной системы, пролин также играет ключевую роль в функционировании суставов и сухожилий.
Заводской пролин и производные
SerineSer & nbsp S9.152.21
| Полярность боковой цепи | Полярность | |
| Заряд | Нейтральный | |
| Нейтральный | ||
| Нейтральный | ||
| Классификация | Нейтральная |
Описание: Серин — это аминокислота с боковой цепью гидроксиметильной группы.Это простейшая гидроксиламинокислота и третья по распространенности в белках. Серин можно синтезировать из глицина и некоторых других метаболитов.
Shop Серин и производные
ThreonineThr & nbsp T9.122.15
| Полярность боковой цепи | Полярность | |||
| Заряд | Нейтральная | г | Классификация | Нейтральная |
Описание: Треонин — незаменимая аминокислота, которая содержит гидроксильную группу, что делает ее полярной и незаряженной.Он имеет много общих свойств с серином и является субстратом киназ и гликозилтрансфераз. Он кодируется кодонами, начинающимися с AC.
ТриптофанTrp & nbsp W9.392.38
| Полярность боковой цепи | Неполярная |
| Заряд | Нейтральная |
| Молекулярная432364 9364 | Молекулярный класс Ароматический |
Описание: Триптофан — самая большая и самая редкая аминокислота, содержащаяся в белках.Обычно находящийся в гидрофобном ядре белков, триптофан часто участвует в ароматической укладке. В его боковой цепи преобладает индольная группа.
TyrosineTyr & nbsp Y9.112.20
| Полярность боковой цепи | Полярность | |
| Заряд | Нейтральная | |
| Нейтральная масса4
4 | Нейтральная масса4 | |
Описание: Тирозин представляет собой ароматическую аминокислоту, подобную фенилаланину с гидроксильной группой вместо фенильной группы в пара-положении.Он обладает уникальными химическими свойствами и служит катализатором и субстратом для множества киназ.
ValineVal & nbsp V9.722.29
| Полярность боковой цепи | Неполярная | ||
| Заряд | Нейтральная | ||
| Молекулярная масса4
2 | 643117643 | 643113643 Алкил |
Описание: Валин — это алифатическая аминокислота среднего размера с боковой цепью, состоящей из пропильной группы, присоединенной к CA на C2.Как и другие алифатические остатки, валин часто находится в гидрофобных ядрах белков.
Незаменимые аминокислоты
Человеческий организм может производить 10 из 20 необходимых аминокислот. Другая половина, называемая незаменимыми аминокислотами, поступает из вашего рациона и должна потребляться регулярно.
| Вырабатывается организмом (несущественное) | Поставляется продуктами питания (незаменимое) | ||
|---|---|---|---|
| Аланин | Глутамин | Аргинин ** | Метионин |
| Asparagine | Фенилаланин | ||
| аспарагиновая кислота | пролин | изолейцин | Треонин |
| Цистеин | серин | Лейцин | Триптофан |
| Глутаминовая кислота | Тирозин * | Лизин | валин |
* Для производства требуется фенилаланин.
** Нужен детям, но не взрослым
Дополнительные ресурсы
Новый способ покупки химикатов
Проведите поиск, нарисовав свою молекулу или загрузив ее химическую структуру.