Механизм повреждения энергетического обмена при гипоксии и возможные пути его коррекции фумаратсодержащими растворами
Л.В. Слепнева, Г.А. Хмылова
ФГБУ «Российский НИИ гематологи трансфузиологии ФМБА», г. Санкт-Петербург
Трансфузиология №2, 2013
Резюме
Статья посвящена вопросам механизма действия препаратов, влияющих на процессы энергообразования в организме. Показаны пути коррекции нарушений энергетического обмена и преимущества фумаратсодержащих инфузионных растворов.
Ключевые слова: гипоксия, цикл Кребса, сукцинат, фумарат, фумаратсодержащие инфузионные растворы, мафусол, полиоксифумарин, конфумин.
В настоящее время нарушения энергетического обмена рассматриваются как один из ведущих патологических процессов, приводящих к необратимым последствиям и гибели организма, что обусловливает исключительную важность рассматриваемой проблемы. Коррекция или устранение энергодефицита является обязательным компонентом в лечении большинства патологических состояний, и в связи с этим, понимание механизма действия препаратов, способных влиять на различные звенья энергообмена, для практикующих врачей приобретают особую ценность.
Жизнедеятельность организма с многообразием всех физиологических функций и биохимических процессов возможна лишь при условии его постоянного энергообеспечения. В настоящее время имеется значительный экспериментальный и клинический материал, свидетельствующий о том, что различные экстремальные воздействия на организм (тяжелая кровопотеря, ожог, травма, сердечная недостаточность, острое отравление и др.) вызывают однотипные повреждения в клеточной системе энергообразования. Это явление обусловлено тем, что результирующим эффектом различных по своей природе экстремальных факторов является развитие острого кислородного голодания тканей. Дефицит кислорода — акцептора электронов в митохондриальной дыхательной цепи, приводит к глубокому подавлению биоэнергетической функции митохондрий. Выходит из строя основная энергетическая система клетки, энергопродукция клетками резко снижается, и, как следствие, нарушается течение многочисленных энергозависимых процессов в организме [2, 6, 11, 13, 24, 34, 36, 37, 43].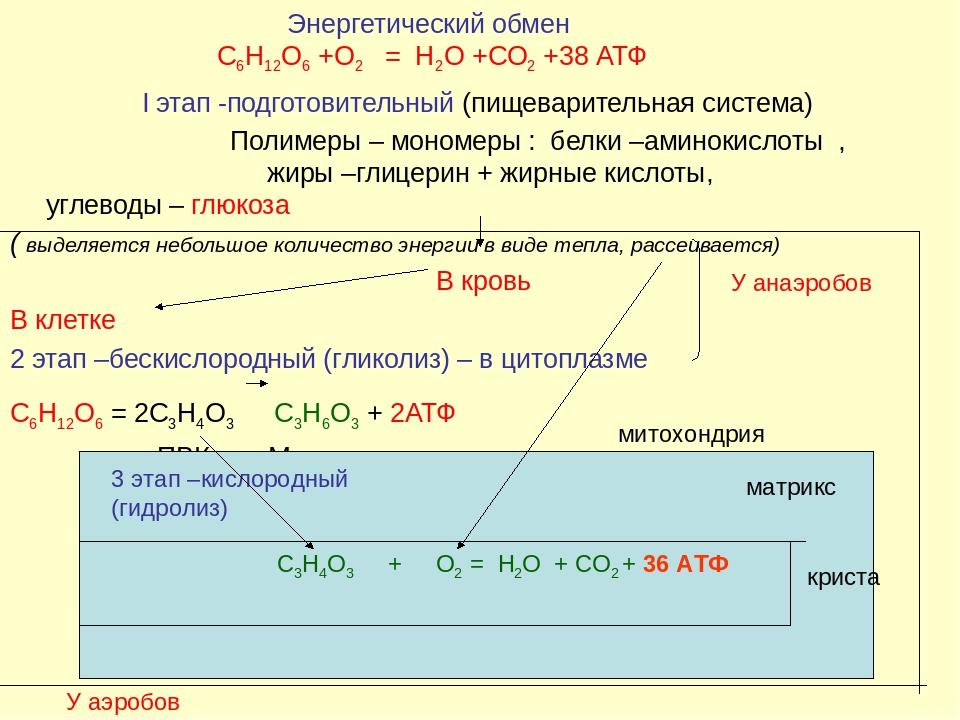
Недостаточность систем энергообразования в клетке составляет существенный элемент патогенеза многих заболеваний. По мнению ряда авторов, поддержание жизни в экстремальных условиях возможно до тех пор, пока дефицит энергии не достигнет критических величин. Истощение клеточных энергетических резервов ниже допустимого уровня сопровождается развитием в клетке необратимых процессов и гибелью организма.
Прежде чем перейти к рассмотрению вопросов, связанных с нарушением энергетического обмена в клетке при патологических состояниях и его коррекции применением различных лекарственных средств, кратко остановимся на описании процессов энергообразования в нормально функционирующей клетке [16, 41].
На рис. 1 схематически представлен сложный процесс распада питательных веществ, который обеспечивает ступенчатое постепенное освобождение энергии и аккумуляцию ее в виде макроэргической фосфатной связи аденозинтрифосфата (АТФ).
Распад сложных питательных веществ на более простые является необходимым условием для дальнейшего использования их в клетке в качестве источников энергии и пластического материала.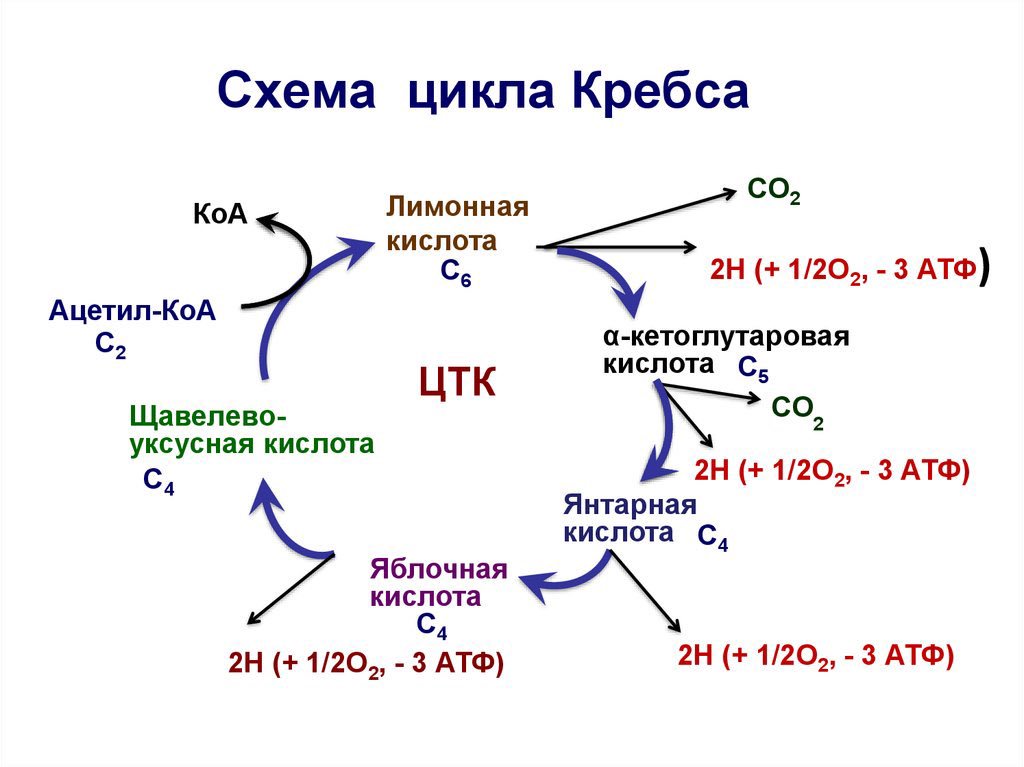 В катаболизме основных питательных веществ (углеводов, белков и жиров) можно выделить три основные стадии.
В катаболизме основных питательных веществ (углеводов, белков и жиров) можно выделить три основные стадии.
На первой стадии крупные молекулы под влиянием сложных ферментативных систем расщепляются на более простые. В результате действия этих ферментативных систем углеводы расщепляются до гексоз и пентоз, липиды – до глицерина и жирных кислот, из белков образуется около 20-ти аминокислот.
На второй стадии происходит дальнейшее расщепление образовавшихся соединений. Из 20-ти различных аминокислот образуется лишь несколько конечных продуктов, а именно, ацетил-коэнзим А, α-кетоглютаровая и щавелевоуксусная кислоты.
Жирные кислоты в процессе β-окисления превращаются в ацетил-КоА. Гексозы под действием ферментативных систем гликолиза расцепляются до пировиноградной кислоты, которая затем в процессе окислительного декарбоксилирования превращается также в ацетил-КоА.
Гликолиз является тем механизмом, посредством которого многие организмы получают химическую энергию из глюкозы и других субстратов в отсутствие молекулярного кислорода. У большинства аэробных организмов процесс гликолиза является предварительной ступенью для дальнейшего окисления продуктов брожения кислородом в процессе дыхания.
У большинства аэробных организмов процесс гликолиза является предварительной ступенью для дальнейшего окисления продуктов брожения кислородом в процессе дыхания.
Метаболиты, образовавшиеся на второй стадии распада питательных веществ (ацетил-КоА, α-кето-глютаровая, щавелевоуксусная кислоты) вступают в третью стадию, которая для них является общей и на которой они в конечном итоге окисляются до СО2 и Н2О.
Третья стадия – стадия терминального окисления питательных веществ, во время которой освобождается основная масса энергии, осуществляется в митохондриях через цикл трикарбоновых кислот (ЦТК) и митохондриальную дыхательную цепь. ЦТК – общий конечный путь окислительного катаболизма всех видов клеточного топлива в аэробных условиях. В этом цикле под действием специфических дегидрогеназ протекают процессы дегидрирования субстратов, восстановительные эквиваленты от которых (протоны и электроны) поступают на митохондриальную дыхательную цепь. Дегидрирование – отщепление молекул Н2 от интермедиатов цикла Кребса происходит, в основном, при помощи дегидрогеназ, простетической группой которых является никотинамидаденин-динуклеотид (НАД), и лишь дегидрирование янтарной кислоты осуществляется ФАД-зависимой дегидрогеназой (сукцинатдегидрогеназой).
Дыхательная цепь, состоящая из серии переносчиков электронов, передает восстановительные эквиваленты конечному акцептору электронов – молекулярному кислороду. Дыхательная цепь – это полиферментная система, локализованная во внутренней мембране митохондрий, основными компонентами которой являются НАД-зависимые дегидрогеназы, флавопротеиды и цитохромы (рис. 1).
Сопряженно с транспортом электронов протекает процесс окислительного фосфорилирования, в котором значительная часть свободной энергии электронов, передаваемых редокс-цепью на кислород, аккумулируется и трансформируется в специфическую макроэргическую связь АТФ. Таким образом, необходимая для нужд организма энергия образуется благодаря функционированию взаимосвязанных процессов гликолиза и дыхания. В процессе гликолиза высвобождается лишь незначительная часть той химической энергии, которая потенциально может быть извлечена из молекулы глюкозы. Полное окисление глюкозы до СО2 и Н 2О, осуществляемое в процессе дыхания, приводит к синтезу значительно большего количества макроэргов.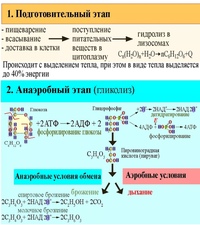 При окислении одной молекулы глюкозы в гликолитическом цикле образуется 2 молекулы АТФ, тогда как дальнейшее расщепление продуктов гликолиза в цикле Кребса сопровождается синтезом 38 молекул АТФ. Таким образом, митохондриальная дыхательная цепь является основным местом приложения и утилизации кислорода в клетке.
При окислении одной молекулы глюкозы в гликолитическом цикле образуется 2 молекулы АТФ, тогда как дальнейшее расщепление продуктов гликолиза в цикле Кребса сопровождается синтезом 38 молекул АТФ. Таким образом, митохондриальная дыхательная цепь является основным местом приложения и утилизации кислорода в клетке.
При дефиците кислорода – конечного акцептора электронов в редокс-цепи митохондрий – отмечается выраженная гиперредукция всех компонентов терминального звена окисления. Прекращаются транспорт электронов по дыхательной цепи и сопряженный с ним процесс образования макроэнергических фосфатов. Известно, что в условиях нормоксии НАД-звено дыхательной цепи митохондрий принимает восстановительные эквиваленты из различных источников:
1. от субстратов цикла Кребса при участии специфических дегидрогеназ;
2. оксиацил-КоА-дегидрогеназы поставляютионы водорода на НАД- звено при окислении жирных кислот;
3. сложная система пируват-дегидрогеназы, отщепляя ионы водорода в реакциях окислительного декарбоксилирования, передает их на НАД-звено редокс-цепи;
4. внемитохондриальный НАД-Н, образованный в центральной реакции гликолитической оксидоредукции, также отдает свои протоны на митохондриальную дыхательную цепь (рис.1).
внемитохондриальный НАД-Н, образованный в центральной реакции гликолитической оксидоредукции, также отдает свои протоны на митохондриальную дыхательную цепь (рис.1).
Мощный поток восстановительных эквивалентов в условиях кислородной недостаточности не может реализоваться из-за гипервосстановленности НАД-зависимого участка дыхательной цепи. Выключается из функционирования основная энергетическая система клетки, резко снижается продукция АТФ.
В анаэробных условиях клетка стремится восполнить энергетический дефицит за счёт активации гликолиза. Несмотря на то, что при анаэробном гликолизе продуцируется почти в 20 раз меньше АТФ, чем при полном сгорании глюкозы в цикле Кребса, потенциальная скорость процесса в основном может обеспечить энергозатраты организма. Однако для осуществления гликолитических реакций необходим постоянный приток окисленной формы НАД, который при нормоксии обеспечивается работой специфических челночных механизмов. Гликолитический НАД-Н проникает через митохондриальные мембраны посредством функционирования α-глицерофосфатного, β-оксибутиратного и других механизмов переноса восстановительных эквивалентов.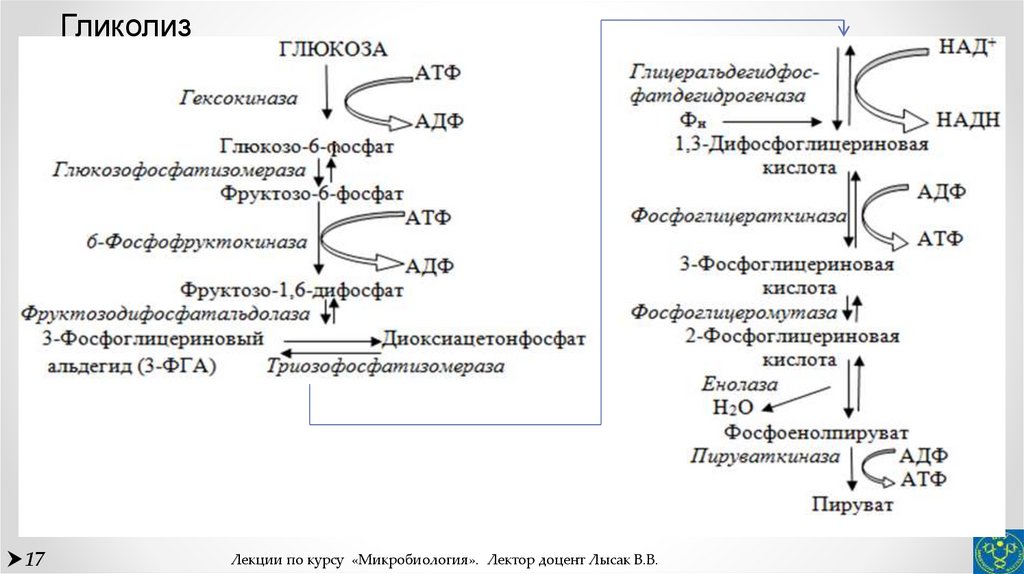
Оксибутиратный и глицерофосфатные шунты являются основными конкурентами лактатдегидрогеназного механизма окисления гликолитического НАД-Н, деятельность которого при нормальной концентрации кислорода в клетке подавлена более активными вышеназванными механизмами.
При нарушении электронтранспортной функции редокс-цепи и гиперредукции ее НАД-звена клетка вынуждена изыскивать другие пути реокисления цитоплазматического НАД-Н. В условиях острой гипоксии конечный продукт гликолиза – пируват – не подвергается декарбоксилированию и не вовлекается в цикл Кребса, а, принимая восстановительные эквиваленты от цитоплазматического НАД-Н, превращается в лактат с освобождением новых порций окисленной формы НАД (рис. 2).
Активация лактатдегидрогеназного механизма поставки НАД для гликолиза в конечном итоге приводит к истощению запасов гликогена и тканевому ацидозу вследствие накопления кислых продуктов метаболизма (лактата, пирувата, оксибутирата, глицерофосфата и др.). Избыточные концентрации конечного субстрата анаэробного гликолиза – лактата – тормозят последнюю реакцию гликолитического цикла.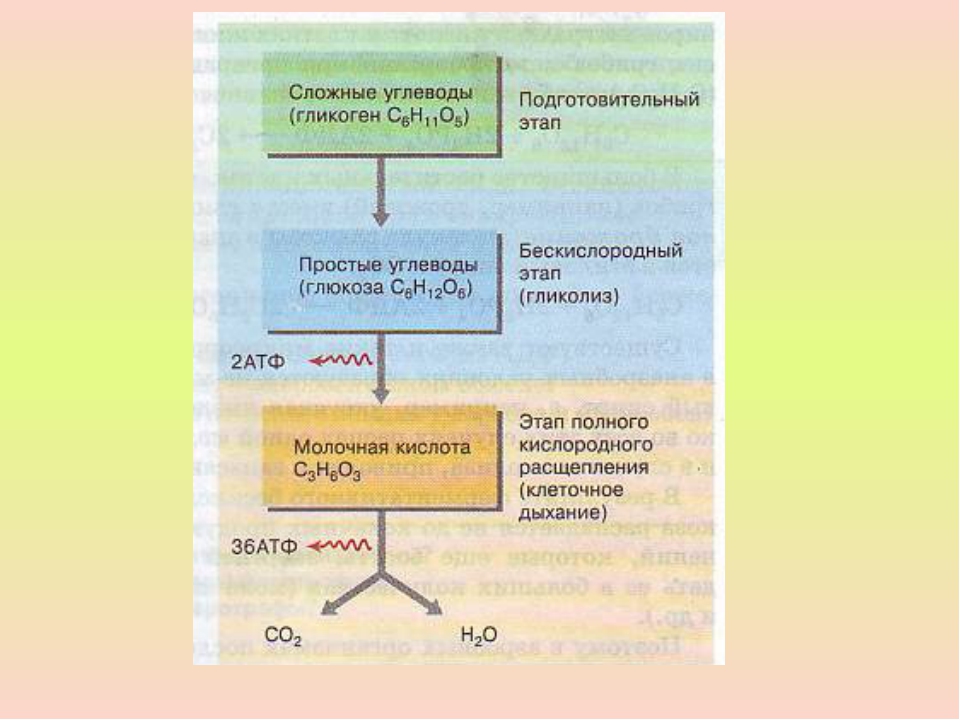
Регенерация НАД прекращается, и, именно, дефицит пиридиннуклеотида останавливает гликолиз и анаэробную продукцию АТФ. Клеточный ацидоз способствует нарушению проницаемости мембран, вплоть до разрушения лизосом. В цитоплазму поступают аутолитические ферменты. Развивается процесс аутолиза клеток, сопровождающийся повреждением тканей и органов. В организме формируются необратимые изменения.
Таким образом, степень повреждения митохондриального метаболизма в условиях тяжелой кислородной недостаточности определяет тяжесть многих патологических состояний. Накопленный опыт лечения шока и кровопотери показывает, что существующие инфузионнотрансфузионные среды, проявляя лечебное действие в стадиях легкой и средней тяжести, оказываются недостаточными на поздних стадиях процесса. Особенности течения поздних стадий геморрагического шока связывают главным образом с генерализованными нарушениями метаболизма и возникающими в результате этого расстройствами энергообмена.
В связи с этим применение совместно с кровезаменителями препаратов, способных повысить энергетический потенциал клетки в условиях гипоксии, рассматривается как один из путей повышения эффективности инфузионной терапии гиповолемических состояний.
В ликвидации энергетического дефицита большое значение придается антигипоксантам. К настоящему времени не выработано единого общепринятого определения антигипоксантов и их классификации, так как в ответ на гипоксическое воздействие вовлекаются самые разные системы организма. Препараты биоэнергетического действия можно разделить на несколько групп.
К первой группе следует отнести препараты, являющиеся источником энергетического сырья (глюкоза, сорбит, АТФ, фосфорилированные гексозы и др.). Использование их показано при патологических состояниях, сопровождающихся истощением энергетических ресурсов в клетке. Включение в состав противошоковых кровезаменителей 5-10% глюкозы или фосфорилированных гексоз [1, 42] для поддержания гликолиза в клетках не позволяет существенно повысить эффективность инфузионной терапии из-за неизбежно возникающего накопления кислых продуктов метаболизма и дефицита окисленной формы пиридиннуклеотида (НАД). Отсюда понятно, что введение таких субстратов окисления, как глюкоза или гексозы, при гипоксии целесообразно лишь с препаратами, ускоряющими утилизацию лактата.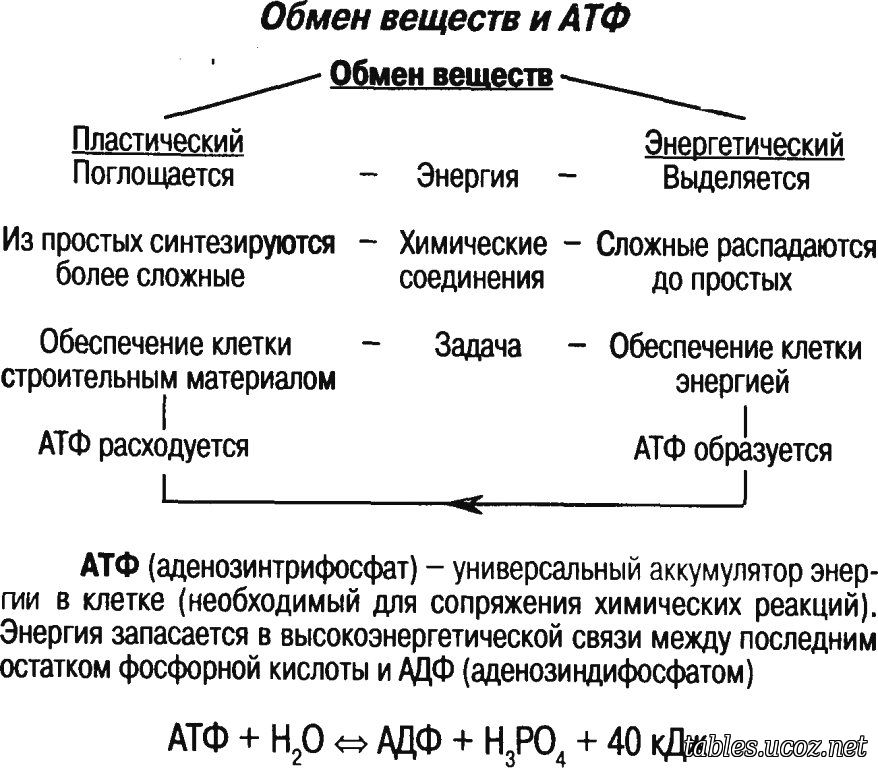 Таким свойством обладают соединения группы гутимина. В эксперименте показан антигипоксический эффект гутимина и амтизола при геморрагическом шоке [8].
Таким свойством обладают соединения группы гутимина. В эксперименте показан антигипоксический эффект гутимина и амтизола при геморрагическом шоке [8].
Ко второй группе препаратов можно отнести средства, которые, не являясь энергетически богатыми соединениями, способны активно воздействовать на энергетический обмен посредством коррекции отдельных звеньев многоступенчатого процесса аккумуляции энергии в клетке. Данные о нарушении транспорта электронов в дыхательной цепи митохондрий при шоке и кровопотери [28] являются теоретической предпосылкой для применения антигипоксантов с электрон-акцепторными свойствами. В литературе имеются довольно обширные сведения о применении естественных и искусственных антигипоксантов – переносчиков электронов. К числу первых относится цитохром С, который, как известно, является одним из компонентов дыхательной цепи митохондрий и служит мобильным переносчиком электронов. Играя важную роль в энергетическом метаболизме клетки, цитохром С показал высокую лечебную эффективность в клинической практике при терапии шока, кровопотери и постишемической гипоксии [12, 32].
Разработке и исследованию искусственных переносчиков электронов посвящено значительное количество работ. Эти соединения способны модифицировать дыхательную цепь митохондрий так, чтобы осуществлять «сброс» восстановительных эквивалентов непосредственно на кислород, минуя заблокированные участки дыхательной цепи. К числу таких веществ относится ряд соединений из класса хинонов (ортопарабензохиноны, нафтохиноны, гексогидрохиноны). Высокий редокс-потенциал этих препаратов определяет их способность к транспорту электронов [30]. При проведении экспериментов на животных многие из этих соединений оказались токсичными, что не позволило рекомендовать их в качестве лечебных средств. Из всех средств, формирующих искус- ственные редокс-системы, в медицинскую практику внедрен препарат «Гипоксен», представляющий собой синтетический полихинон [9].
Известно, что антигипоксанты группы хинонов осуществляют перенос электронов с НАД-Н звена на кислород, минуя все 3 пункта фосфорилирования в дыхательной цепи и, следовательно, устранение дефицита энергии при введении этих препаратов может происходить лишь за счет активации гликолитической выработки АТФ. Однако для полноценного проявления антигипоксических свойств этих соединений необходим акцептор электронов – кислород. Наибольший интерес для включения в состав новых комплексных кровезаменителей представляют, так называемые, истинные антигипоксанты или антигипоксанты прямого действия, непосредственно влияющие на митохондриальный метаболизм при гипоксии.
Однако для полноценного проявления антигипоксических свойств этих соединений необходим акцептор электронов – кислород. Наибольший интерес для включения в состав новых комплексных кровезаменителей представляют, так называемые, истинные антигипоксанты или антигипоксанты прямого действия, непосредственно влияющие на митохондриальный метаболизм при гипоксии.
Согласно теоретическим предпосылкам, одна из возможностей поддержания биоэнергетики клетки может быть реализована посредством стимуляции адаптационных механизмов к гипоксии, работающих на последних этапах цикла Кребса. Из всех субстратов цикла Кребса только влияние сукцината на энергетический обмен подробно изучено в эксперименте на животных.
Исследование механизма действия сукцината при гипоксических состояниях проведено в работах М.Н. Кондрашовой с соавторами [5, 6]. Исследователи считают, что в условиях гипоксии сукцинат, не являясь НАД-зависимым субстратом, «монополизирует» дыхательную цепь и активно в ней окисляется.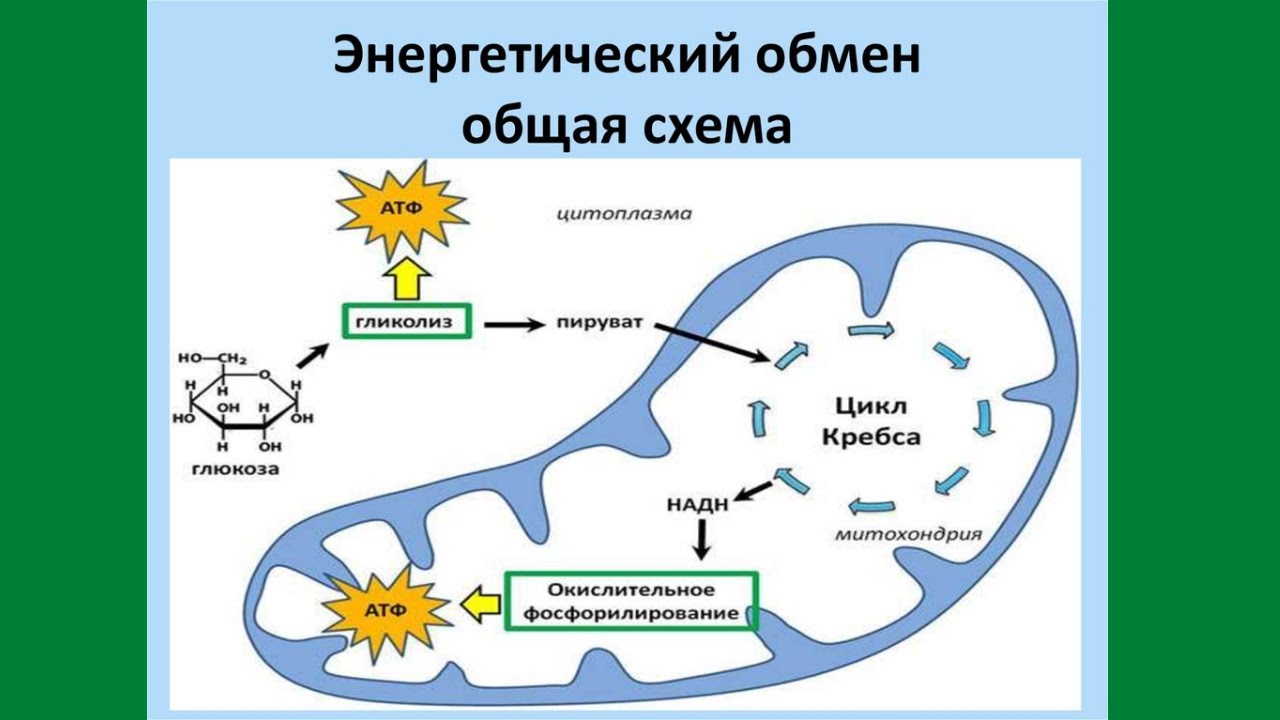 Высокая скорость реакции окисления сукцината, поставляющей 2 молекулы АТФ, позволяет компенсировать выработку 3-х молекул АТФ, образующихся при окислении НАД-зависимых субстратов. Однако в условиях прогрессирующей гипоксии дефицит кислорода, лимитирующий скорость окисления всех субстратов, снижает ценность сукцината и ставит его в один ряд с другими субстратами окисления. Поэтому применение сукцината в качестве антигипоксанта должно быть особенно эффективно в комплексе с препаратами, улучшающими кислородообеспечение организма. Преимущественное использование сукцината – естественная защита клетки против гипоксии. При этом пополнение фонда субстрата может происходить за счет реакций цикла Кребса, идущих как в прямом, так и в обратном направлениях (рис. 3).
Высокая скорость реакции окисления сукцината, поставляющей 2 молекулы АТФ, позволяет компенсировать выработку 3-х молекул АТФ, образующихся при окислении НАД-зависимых субстратов. Однако в условиях прогрессирующей гипоксии дефицит кислорода, лимитирующий скорость окисления всех субстратов, снижает ценность сукцината и ставит его в один ряд с другими субстратами окисления. Поэтому применение сукцината в качестве антигипоксанта должно быть особенно эффективно в комплексе с препаратами, улучшающими кислородообеспечение организма. Преимущественное использование сукцината – естественная защита клетки против гипоксии. При этом пополнение фонда субстрата может происходить за счет реакций цикла Кребса, идущих как в прямом, так и в обратном направлениях (рис. 3).
При обратном течении реакций имеющийся запас малата по мере необходимости превращается в фумарат, который восстанавливается в сукцинат. Восстановление фумарата сопровождается выработкой АТФ, и поэтому реакции обращения в системе «малат-фумарат-сукцинат» способны поддерживать окислительное фосфорилирование даже при аноксии.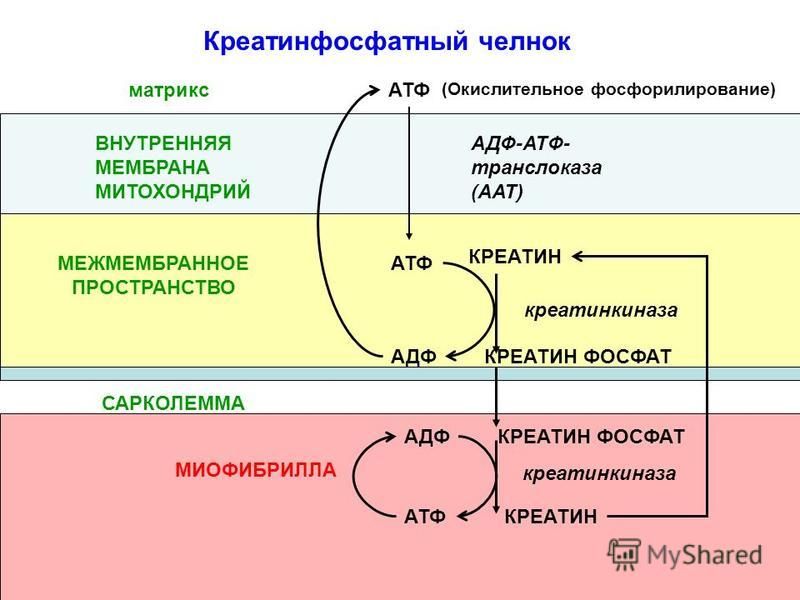
В условиях же гипоксии инверсивные превращения фумарата выполняют роль триггера, который, в зависимости от концентрации кислорода регулирует течение конечных реакций цикла Кребса в прямом либо в обратном направлениях, и эти реакции сопровождаются синтезом АТФ. Механизм инверсивных превращений фумарата в цикле Кребса объясняет эффективность применения фумаратсодержащих инфузионных сред, таких как кристаллоидный раствор – мафусол, коллоидный кровезаменитель – полиоксифумарин и концентрированный раствор фумарата натрия – конфумин. Эти препараты разработаны и основательно изучены в Российском НИИ гематологии и трансфузиологии. Лечебная эффективность была изучена на моделях геморрагического и ожогового шока, а также при экспериментальном перитоните [17–20, 23, 25,45]. Оценку эффективности инфузионных растворов определяли по совокупности показателей системной гемодинамики, кислородного режима, кислотно-основного состояния (КОС), перекисного окисления липидов и митохондриального метаболизма в печени и сердце животных. Полярографическое исследование митохондрий, выделенных из печени и сердца животных, леченных фумаратсодержащими растворами, свидетельствовало о полном восстановлении энергопродуцирующих функций этих органелл. Следует отметить, что летальность животных в контрольной группе (тяжелый шок) составляло 100%, при лечении мафусолом или полиоксифумарином – 17–20%.
Полярографическое исследование митохондрий, выделенных из печени и сердца животных, леченных фумаратсодержащими растворами, свидетельствовало о полном восстановлении энергопродуцирующих функций этих органелл. Следует отметить, что летальность животных в контрольной группе (тяжелый шок) составляло 100%, при лечении мафусолом или полиоксифумарином – 17–20%.
Результаты исследования митохондриального метаболизма позволяют предположить, что парентеральное введение фумарата индуцирует суперкомпенсацию адаптационного механизма к гипоксии, функционирующих на последних этапах цикла Кребса. Фумарат в системе «малат-фумарат-сукцинат» способен поддерживать синтез АТФ как в аэробных, так и в анаэробных условиях. При дефиците кислорода фумарат, восстанавливаясь ФАД∙Н2-группой сукцинатдегидрогеазой, превращается в сукцинат и освобождает новые порции окисленной формы ФАД. Принимая восстановительные эквиваленты от НАД-Н, ФАД способствует снятию гипервосстановленности НАД звена дыхательной цепи и синтезу АТФ в бескислородной среде. При поступлении кислорода в клетку сукцинат, синтезируемый из фумарата, монополизирует дыхательную цепь и, активно окисляясь в ней, продуцирует АТФ (рис. 3). К тому же, образование в этих реакциях окисленной формы НАД запускает также и механизм гликолитической продукции АТФ. Поддержание энергетического потенциала клетки при инфузия фумарата способствует удлинению периода обратимых изменений в организме и предотвращает развитие «необратимости» при патологических состояниях, отягощенных глубокой гипоксией.
При поступлении кислорода в клетку сукцинат, синтезируемый из фумарата, монополизирует дыхательную цепь и, активно окисляясь в ней, продуцирует АТФ (рис. 3). К тому же, образование в этих реакциях окисленной формы НАД запускает также и механизм гликолитической продукции АТФ. Поддержание энергетического потенциала клетки при инфузия фумарата способствует удлинению периода обратимых изменений в организме и предотвращает развитие «необратимости» при патологических состояниях, отягощенных глубокой гипоксией.
Парентеральное введение фумаратсодержащих растворов наряду с восстановлением биоэнергетики клетки, сопровождается «мягким» ощелачивающим действием препаратов на кислотно-основное состояние крови при ацидозе. Это действие обусловлено тем, что такие органические соли, как фумарат-, ацетат-, лактат-, сукцинат- и малат натрия являются соединениями, образованными сильным основанием (NaOH) и слабой кислотой. При гидролизе подобных солей в кровеносном русле освобождается соответствующая кислота и NaOH, который расходуется на нейтрализацию кислых продуктов метаболизма. Реакция гидролиза смещена вправо, так как постоянно происходит потребление продукта гидролитической реакции – NaOH (рис. 4).
Реакция гидролиза смещена вправо, так как постоянно происходит потребление продукта гидролитической реакции – NaOH (рис. 4).
Следует отметить, что вышеназванные соли оказывают мягкое ощелачивающие действие по сравнению с бикарбонатом натрия, широко используемым в клинической практике для ликвидации ацидоза. Реакция гидролиза NaHСО3 протекает значительно быстрее, так как в ходе реакции удаляются оба ее продукта: NaOH расходуется на нейтрализацию метаболитов, а второй продукт реакции – угольная кислота, нестоек и разлагается на Н2О и СО2. Образованная в избыточном количестве щелочь может способствовать развитию алкалоза, что имеет место в клинических условиях при передозировке бикарбоната натрия.
Все вышеперечисленные соли входят в состав различных инфузионных растворов (мафусол, полиоксифумарин, конфумин, лактасол, Рингер-лактат, ацесоль, реамберин, стерофундин и др.). Однако оказывая ощелачивающее действие при ацидозе, далеко не все эти препараты способны поддержать энергетический обмен при гипоксии. Восстановление показателей КОС «химическим путем» является недостаточным для успешной терапии шока.
Восстановление показателей КОС «химическим путем» является недостаточным для успешной терапии шока.
Следует к тому же учитывать, что при гидролизе лактата натрия выделяется молочная кислота, которая в сумме с эндогенной молочной кислотой, возникающей в больших концентрациях при гипоксии, могут способствовать подавлению реакций гликолиза, что, в свою очередь, вызывает снижение продукции гликолитической АТФ. Существуют также исследования, указывающие, что лактат может вызвать интерстициальный отек головного мозга и повышать агрегацию тромбоцитов и эритроцитов [14, 39, 44]. Лактатсодержащие инфузионные растворы нельзя использовать при печеночной недостаточности [35, 38, 40], а также в случаях шока, сопровождающегося гиперлактатемией или лактатным ацидозом [33].
Ацетат натрия, в отличие от лактата, не проявляет токсического действия при тяжелом шоке. Однако утилизация уксусной кислоты, образованной при гидролизе ацетата натрия, в условиях кислородной недостаточности затруднена вследствие постгипоксического дефекта в функционировании митохондриальной дыхательной цепи. Лечебное действие фумарата натрия в сравнении с лактатом и ацетатом представляется более физиологичным, так как при его введении наряду с ощелачивающим эффектом проявляется и его влияние на восстановление процессов генерации энергии в митохондриях, а, следовательно, устраняется причина возникновения метаболического ацидоза.
Лечебное действие фумарата натрия в сравнении с лактатом и ацетатом представляется более физиологичным, так как при его введении наряду с ощелачивающим эффектом проявляется и его влияние на восстановление процессов генерации энергии в митохондриях, а, следовательно, устраняется причина возникновения метаболического ацидоза.
Сукцинатсодержащие растворы, в частности «Реамберин», способствуют поддержанию энергетического обмена, однако, в условиях острого дефицита кислорода подавляется окисление сукцината и существенно снижается его энергопродуцирующая функция. Окисление малата в цикле Кребса осуществляется НАД-зависимой малатдегидрогеназой, и эта реакция тормозится из-за гипервосстановленности НАД-звена редокс-цепи митохондрий при гипоксии. Следовательно, в этих условиях субстрат не способен повысить энергетический потенциал клетки. К тому же, в инфузионном малатсодержащем растворе «Стерофундин» концентрация малата очень низкая, чтобы обеспечить достаточную продукцию АТФ. В условиях гипоксии повышение концентрации малата могло бы создать условия для обращения реакций в цикле Кребса с увеличением фонда фумарата, способного принимать восстановительные эквиваленты (Н2) и синтезировать АТФ. Однако концентрация малата в стерофундине (5 ммоль/л) незначительна для запуска реакций в цикле Кребса в обратном направлении.
Однако концентрация малата в стерофундине (5 ммоль/л) незначительна для запуска реакций в цикле Кребса в обратном направлении.
Фумаратсодержащие растворы (мафусол, полиоксифумарин) содержат высокие концентрации фумарата (86 ммоль/л), обеспечивающие как выработку АТФ, так и накопление сукцината, который активно окисляется при поступлении кислорода. Введение субстратов в организм при гипоксии показано еще и вследствие того, что кислородная недостаточность сопровождается значительным субстратным голодом клетки. Препараты «Мафусол» и «Полиоксифумарин» с высокой концентрацией фумарата и возможностью инфузий больших объемов этих растворов без побочных эффектов являются высокоэффективными средствами терапии шока различного генеза. Это подтверждено клинически. Так, кристаллоидный кровезаменитель «Мафусол» разрешен к медицинскому применению уже более 20 лет и широко используется в разных областях медицины (хирургия, неврология, кардиология, реаниматология, педиатрия, акушерство и гинекология, комбустиология, токсикология и др. ) [3, 15, 22, 27, 29, 31]. Отличительной особенностью этого препарата является то, что его можно переливать в больших количествах, не только внутривенно, но и внутриартериально, а также в смеси для заполнения контура АИК при открытых операциях на сердце. Ни один из существующих сейчас на фармацевтическом рынке инфузионных антигипоксических препаратов не обладает этими свойствами. Полифункциональный коллоидный плазмозаменитель «Полиоксифумарин» с 1999 года успешно применяется у взрослых и детей в клинической практике гиповолемических состояний различной степени тяжести [10, 21, 22]. Аналогов ему нет ни в России, ни зарубежом.
) [3, 15, 22, 27, 29, 31]. Отличительной особенностью этого препарата является то, что его можно переливать в больших количествах, не только внутривенно, но и внутриартериально, а также в смеси для заполнения контура АИК при открытых операциях на сердце. Ни один из существующих сейчас на фармацевтическом рынке инфузионных антигипоксических препаратов не обладает этими свойствами. Полифункциональный коллоидный плазмозаменитель «Полиоксифумарин» с 1999 года успешно применяется у взрослых и детей в клинической практике гиповолемических состояний различной степени тяжести [10, 21, 22]. Аналогов ему нет ни в России, ни зарубежом.
Применение концентрированного раствора фумарата натрия (препарата «Конфумин») в качестве антигипоксического компонента в схемах инфузионно-трансфузионной терапии существенно увеличивает уровень субстратов окисления в кровеносном русле и позволяет повысить лечебную эффективность общепринятых в клинической практике плазмозаменителей [3, 4, 22, 25-27]. Конфумин разрешен к широкому медицинскому применению у взрослых, промышленный выпуск препарата освоен в ОАО «Фирма Медполимер».
2. Энергетический обмен (катаболизм, диссимиляция)
В любой клетке главным источником энергии для всех протекающих в ней процессов служит универсальное энергетическое соединение — АТФ.
Рис. \(1\).Строение АТФ
АТФ образуется в результате присоединения к АДФ (аденозиндифосфату) одного остатка фосфорной кислоты. Этот процесс называется фосфорилированием:
АДФ + h4PO4+ 40 кДж = АТФ + h3O.
В молекуле АТФ есть две богатые энергией химические связи. Это связи между остатками фосфорной кислоты. Такие высокоэнергетические связи называют макроэргическими. При разрыве одной макроэргической связи АТФ превращается в АДФ и выделяется около \(40\) кДж/моль энергии.
Образование АТФ происходит в процессе энергетического обмена, или диссимиляции.
Энергетический обмен (диссимиляция, катаболизм) — это реакции расщепления и окисления органических веществ, протекающие с выделением энергии, частично расходующейся на образование АТФ.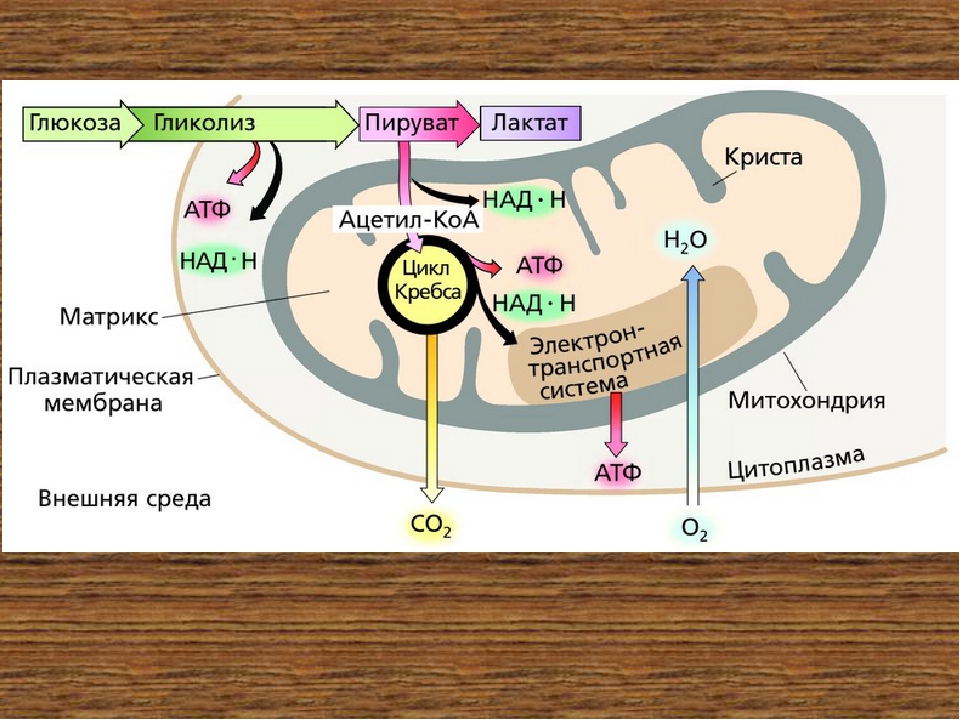
Энергетический обмен в клетках может быть двухэтапным или трёхэтапным.
В кислородсодержащей среде (у аэробных организмов) диссимиляция протекает в три этапа: подготовительный, бескислородный и кислородный. В результате образуются простые неорганические вещества.
Рис. \(2\). Этапы энергетического обмена
В бескислородной среде (у анаэробных организмов), а также при недостатке кислорода у аэробных организмов, энергетический обмен протекает в два этапа: подготовительный и бескислородный. В этом случае количество запасённой энергии намного меньше, чем в присутствии кислорода.
Первый этап — подготовительный
На подготовительном этапе большие молекулы органических веществ распадаются до более простых: из полисахаридов образуются моносахариды, из жиров — смесь глицерина и жирных кислот, а из белков — смесь аминокислот.
Этот процесс происходит в лизосомах и в органах пищеварения под действием пищеварительных ферментов.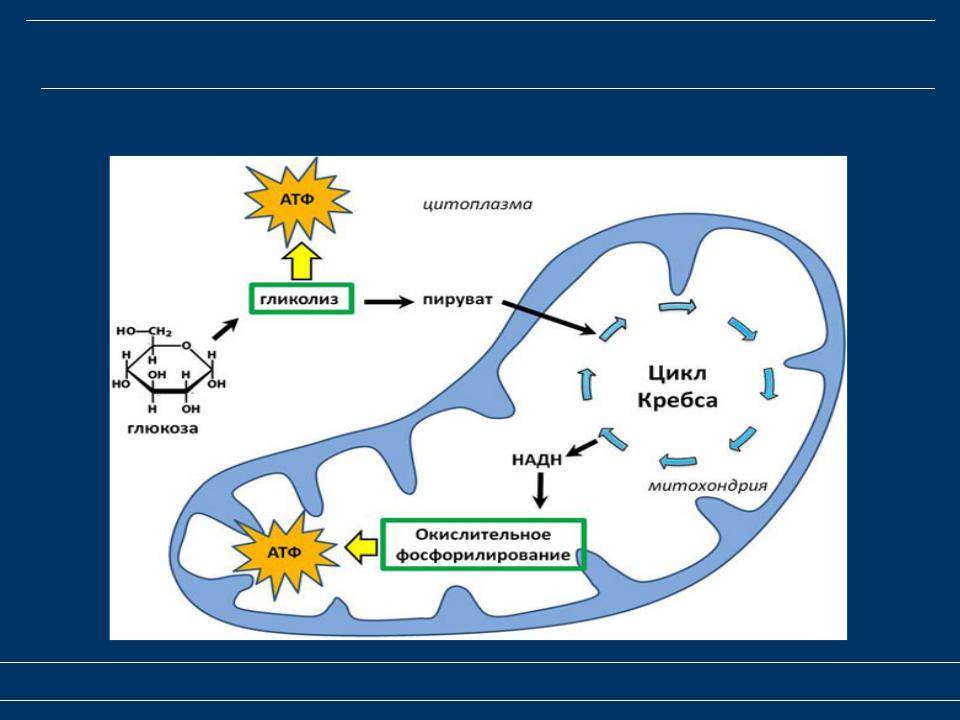
На подготовительном этапе АТФ не образуется, а небольшое количество выделяющейся энергии рассеивается.
Второй этап — бескислородный (гликолиз)
На втором этапе происходит расщепление продуктов подготовительного этапа под действием ферментов. Кислород при этом не используется.
Бескислородный этап расщепления глюкозы имеет название гликолиз. Этот процесс протекает в цитоплазме клеток.
При гликолизе происходит несколько реакций, в результате которых из молекулы глюкозы C6h22O6 образуется \(2\) молекулы пировиноградной кислоты (ПВК) C3h5O3, а также \(2\) молекулы АТФ. В них запасается около \(40\) % выделившейся энергии, остальные \(60\) % рассеиваются.
C6h22O6+2h4PO4+2АДФ=2C3h5O3+2АТФ +2h3O.
Образовавшаяся пировиноградная кислота в анаэробных условиях превращается в молочную кислоту C3H6O3:
HOOC−CO−Ch4→HOOC−CHOH−Ch4.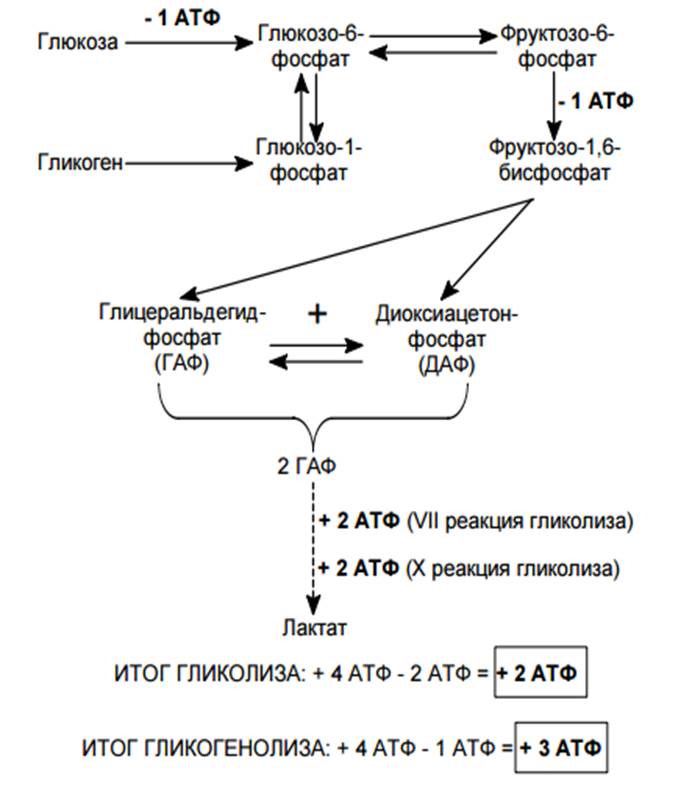
Такой процесс происходит в клетках животных, некоторых грибов и бактерий. Молочная кислота может образоваться и в мышцах человека при длительных нагрузках и недостатке кислорода. Тогда мышцы начинают болеть.
В растительных клетках и клетках дрожжей из пировиноградной кислоты образуется этиловый спирт C2H5OH и углекислый газ CO2, т. к. происходит спиртовое брожение:
C6h22O6+2h4PO4+2АДФ=2C2H5OH+2CO2+2АТФ+2h3O.
Третий этап — кислородный
В кислородной среде после гликолиза протекает третий этап энергетического обмена — кислородный, или клеточное дыхание. Полное кислородное расщепление осуществляется на мембранах митохондрий.
Этот этап тоже является многостадийным. В нём выделяют два процесса — цикл Кребса и окислительное фосфорилирование.
Сущность третьего (кислородного) этап заключается в окислении ПВК до углекислого газа и воды При этом выделившаяся энергия запасается в \(36\) молекулах АТФ (\(2\) — в цикле Кребса и \(34\) — при окислительном фосфорилировании).
Схема кислородного этапа:
2C3h5O3+6O2+36h4PO4+36АДФ=6CO2+42h3O+36АТФ.
Так как \(2\) молекулы АТФ образуются при гликолизе, то в сумме при полном окислении одной молекулы глюкозы образуется \(38\) молекул АТФ.
Суммарное уравнение энергетического обмена:
C6h22O6+6O2=6CO2+6h3O+38АТФ.
В реакциях энергетического обмена используется не только глюкоза, но и липиды, белки. Но главным источником энергии в большинстве клеток являются углеводы.
Источники:
Рис. 1. Строение АТФ. https://image.shutterstock.com/image-illustration/atp-energy-currency-cell-600w-1509423494
Рис. 2. Этапы энергетического обмена. © ЯКласс.
Учебная программа лечебного факультета
Содержание предмета биохимия. Биохимия белка. Строение, физико–химические и химические свойства протеиногенных аминокислот и белков. Классификации белков. Функции белков и пептидов. Биосинтез белка. Конформация белковой молекулы. Механизмы взаимодействия белок–лиганд. Денатурация белка. Методы исследования белков. Энзимология. Строение и механизмы функционирования ферментов. Классификация, индексация и номенклатура ферментов. Особенности ферментов в качестве биологических катализаторов. Кинетика ферментативного катализа. Уровни регуляции ферментов. Понятие о метаболических путях. Катаболизм и анаболизм. Энзимодиагностика и энзимотерапия. Типы и механизмы биологического окисления. Митохондриальное окисление: субстраты, цепи транспорта электронов, сопряжение с процессом окислительного фосфорилирования. Виды внемитохондриального окисления, их роль. Активные формы кислорода. Антиоксидантная система. Биологически важные углеводы человека. Переваривание и всасывание углеводов. Метаболические пути обмена углеводов, связанные с получением энергии в виде АТФ. Структурные углеводы, их синтез. Роль ГАГ, протеогликанов, углеводных компонентов гликопротеинов и гликолипидов.
Классификации белков. Функции белков и пептидов. Биосинтез белка. Конформация белковой молекулы. Механизмы взаимодействия белок–лиганд. Денатурация белка. Методы исследования белков. Энзимология. Строение и механизмы функционирования ферментов. Классификация, индексация и номенклатура ферментов. Особенности ферментов в качестве биологических катализаторов. Кинетика ферментативного катализа. Уровни регуляции ферментов. Понятие о метаболических путях. Катаболизм и анаболизм. Энзимодиагностика и энзимотерапия. Типы и механизмы биологического окисления. Митохондриальное окисление: субстраты, цепи транспорта электронов, сопряжение с процессом окислительного фосфорилирования. Виды внемитохондриального окисления, их роль. Активные формы кислорода. Антиоксидантная система. Биологически важные углеводы человека. Переваривание и всасывание углеводов. Метаболические пути обмена углеводов, связанные с получением энергии в виде АТФ. Структурные углеводы, их синтез. Роль ГАГ, протеогликанов, углеводных компонентов гликопротеинов и гликолипидов. Источники глюкозы в крови, регуляция ее концентрации. Механизмы гликирования и гликозилирования биомолекул. Липиды организма человека: определение, классификация. Переваривание и всасывание липидов. Строение и функции триглицеридов. Синтез и распад триглицеридов. Кетоновые тела: их синтез, утилизация и роль в организме человека. Мембранные липиды: строение, свойства, роль. Синтез и распад мембранных липидов. Системы транспорта липидов. Фракции липопротеинов, методы их исследования. Биологически активные продукты катаболизма липидов. Многообразие белков организма человека. Механизмы посттрансляционной модификации белка. Азотистый баланс. Пищевая ценность белков. Переваривание белков в желудочно–кишечном тракте, всасывание аминокислот. Этапы катаболизма белка. Протеолиз. Роль тотального и ограниченного протеолиза. Регуляция протеолитических систем. Трансаминирование и дезаминирование аминокислот. Синтез заменимых аминокислот. Пути образования и обезвреживания аммиака. Пути образования и обезвреживания биологически активных продуктов аминокислотного обмена.
Источники глюкозы в крови, регуляция ее концентрации. Механизмы гликирования и гликозилирования биомолекул. Липиды организма человека: определение, классификация. Переваривание и всасывание липидов. Строение и функции триглицеридов. Синтез и распад триглицеридов. Кетоновые тела: их синтез, утилизация и роль в организме человека. Мембранные липиды: строение, свойства, роль. Синтез и распад мембранных липидов. Системы транспорта липидов. Фракции липопротеинов, методы их исследования. Биологически активные продукты катаболизма липидов. Многообразие белков организма человека. Механизмы посттрансляционной модификации белка. Азотистый баланс. Пищевая ценность белков. Переваривание белков в желудочно–кишечном тракте, всасывание аминокислот. Этапы катаболизма белка. Протеолиз. Роль тотального и ограниченного протеолиза. Регуляция протеолитических систем. Трансаминирование и дезаминирование аминокислот. Синтез заменимых аминокислот. Пути образования и обезвреживания аммиака. Пути образования и обезвреживания биологически активных продуктов аминокислотного обмена. Синтез, распад и роль креатина. Роль аминокислот в синтезе гемма и нуклеиновых кислот. Строение и функции нуклеиновых кислот. Функции нуклеотидов. Биохимические основы синтеза нуклеиновых кислот. Посттранскрипционная модификации РНК. Синтез и распад нуклеотидов. Строение и классификация гемопротеинов. Молекулярные формы и производные гемоглобина. Синтез и распад гема. Прямой и непрямой билирубин. Обмен воды и минеральный веществ и их регуляция. Состав плазмы крови. Белковые фракции крови. Функции белковых и небелковых компонентов крови. Система свертывания крови и фибринолиза. Белки иммунной системы. Системы регуляции сосудистого тонуса. Особенности химического состава и метаболизма эритроцитов и лейкоцитов. Состав миоцитов. Механизм мышечного сокращения. Источники энергии для мышечной ткани. Особенности химического состава и метаболизма нервной ткани. Химическая классификация нейромедиаторов. Функции почек. Биохимические механизмы экскреторной и гомеостатичской функций почек. Гормоны почки.
Синтез, распад и роль креатина. Роль аминокислот в синтезе гемма и нуклеиновых кислот. Строение и функции нуклеиновых кислот. Функции нуклеотидов. Биохимические основы синтеза нуклеиновых кислот. Посттранскрипционная модификации РНК. Синтез и распад нуклеотидов. Строение и классификация гемопротеинов. Молекулярные формы и производные гемоглобина. Синтез и распад гема. Прямой и непрямой билирубин. Обмен воды и минеральный веществ и их регуляция. Состав плазмы крови. Белковые фракции крови. Функции белковых и небелковых компонентов крови. Система свертывания крови и фибринолиза. Белки иммунной системы. Системы регуляции сосудистого тонуса. Особенности химического состава и метаболизма эритроцитов и лейкоцитов. Состав миоцитов. Механизм мышечного сокращения. Источники энергии для мышечной ткани. Особенности химического состава и метаболизма нервной ткани. Химическая классификация нейромедиаторов. Функции почек. Биохимические механизмы экскреторной и гомеостатичской функций почек. Гормоны почки.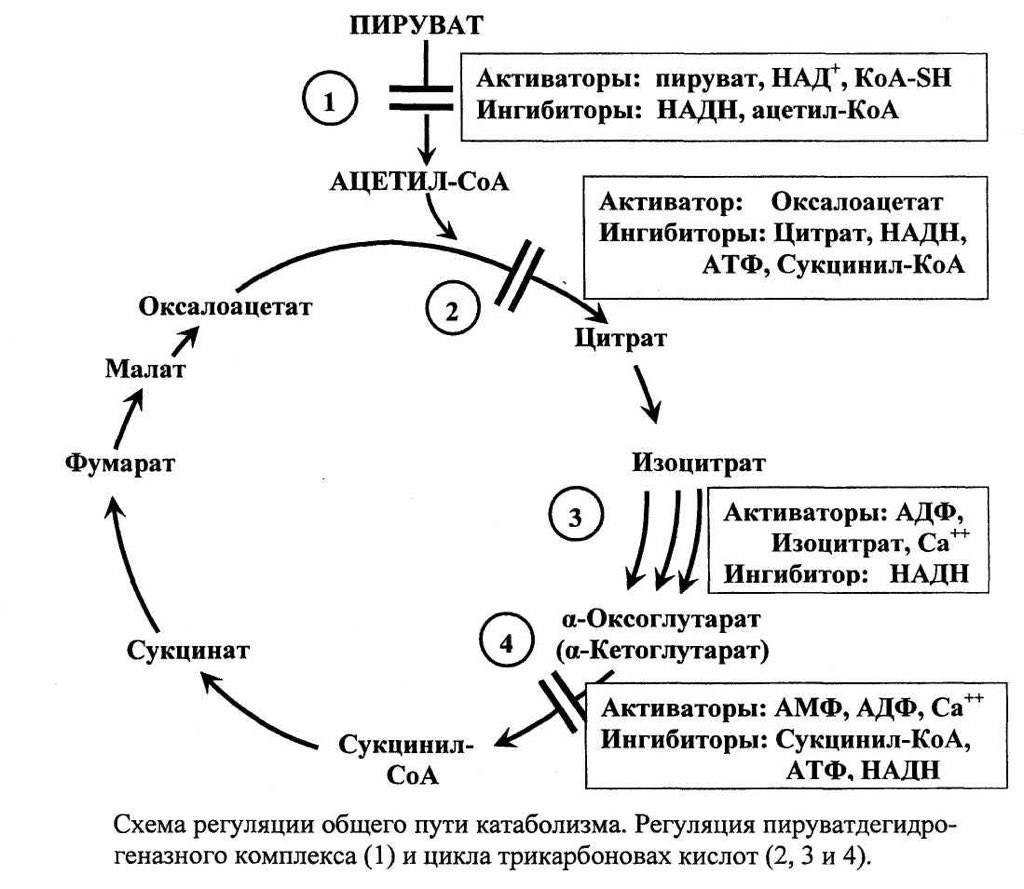 Гормональная регуляция мочеобразования. Химический состав мочи человека.
Гормональная регуляция мочеобразования. Химический состав мочи человека.
Онлайн урок: Энергетический обмен по предмету Биология 9 класс
Задача 1
В процессе гликолиза в клетках эукариот образовались 120 молекул пировиноградной кислоты (ПВК).
Какое количество молекул глюкозы подверглось расщеплению и сколько молекул АТФ образуется при полном ее окислении? Ответ поясните.
Решение:
1) В процессе гликолиза при расщеплении 1 молекулы глюкозы образуются 2 молекулы пировиноградной кислоты и выделяется энергия, которой хватает на синтез двух молекул АТФ
2) Так как образовалось 120 молекул пировиноградной кислоты, то расщеплению подверглось 120 : 2 = 60 молекул глюкозы
3) При полном окислении в расчете на одну молекулу глюкозы образуется 38 молекул АТФ
4) Следовательно, при полном окислении 60 молекул глюкозы образуются
38 х 60 = 2280 молекул АТФ
Задача 2
Гликолизу подверглись две молекулы глюкозы, окислению только одна.
Определите количество образовавшихся в результате этого процесса молекул АТФ и углекислого газа.
Решение:
Для решения используем уравнения 2 этапа (гликолиза) и 3 этапа (кислородного) энергетического обмена.
1) При гликолизе одной молекулы глюкозы образуются 2 молекулы АТФ
2) По условию задачи гликолизу подверглось 2 молекулы глюкозы:
2х2= 4 молекулы АТФ образовалось
3) При окислении (кислородный этап) из одной молекулы глюкозы образуются 36 молекул АТФ
4) По условию задачи окислению подверглась только одна молекула глюкозы:
1 х 36= 36 молекул АТФ образовалось
5) Таким образом образовалось 4 + 36 = 40 молекул АТФ
6) Углекислый газ образуется только на 3 этапе, при полном окислении одной молекулы глюкозы образуется 6 молекул СО2
Ответ: образовалось 40 молекул АТФ и 6 молекул СО2
Задача 3
В процессе окислительного фосфорилирования образовалось 1728 молекул АТФ.
Определите, какое количество молекул глюкозы подверглось расщеплению и сколько молекул АТФ образовалось в результате подготовительного и бескислородного этапов катаболизма.
Решение:
1) При окислительном фосфорилировании (третий этап энергетического обмена) из одной молекулы глюкозы образуется 36 молекул АТФ, следовательно, гидролизу подверглось:
1728 : 36 = 48 молекул глюкозы
2) При гликолизе она молекула глюкозы расщепляется до двух молекул пировиноградной кислоты (ПВК) с образованием двух молекул АТФ, следовательно, образовалось:
48 х 2 = 96 молекул АТФ
3) При полном окислении одной молекулы глюкозы образуется 38 молекул АТФ, следовательно, при полном окислении 48 молекул глюкозы образовалось:
48 х 38 = 1824 молекул АТФ (1728 АТФ третьего этапа + 96 АТФ второго этапа = 1824)
Задача 4
Сколько молекул АТФ будет синтезировано в клетках в процессе гликолиза, если происходит окисление участка молекулы крахмала, содержащего 50 остатков глюкозы?
Решение:
1) В процессе гликолиза при окислении одной молекулы глюкозы образуется 2молекулы АТФ
2) Из фрагмента молекулы крахмала, содержащего 50 остатков глюкозы в результате гликолиза образуется 100 молекул АТФ (50 х 2 = 100)
Задача 5
В процессе полного расщепления глюкозы образовалось 342 молекулы АТФ.
Сколько молекул глюкозы поверглось расщеплению?
Сколько молекул АТФ образовалось в результате гликолиза?
Объясните полученный результат.
Решение:
1) В результате полного расщепления одной молекулы глюкозы образуется 38 молекул АТФ
2) Расщеплению поверглись 342 : 38 = 9 молекул глюкозы
3) Гликолиз– бескислородный этап катаболизма, в результате которого из одной молекулы глюкозы образуется 2 молекулы АТФ
4) Следовательно, в результате гликолиза образовалось 9 х 2 = 18 молекул АТФ
Задача 6
При беге со средней скоростью мышцы ног расходуют за 1 минуту 24 кДж энергии.
Определите сколько граммов глюкозы израсходуют мышцы ног за 25 минут бега, если кислород доставляется кровью к мышцам в достаточном количестве?
Для сведения: один моль содержит 180 грамм глюкозы, и из этого количества образуется 38 молекул АТФ.
Решение:
1) Определим, сколько всего энергии было израсходовано за 25 минут бега:
25 минут х 24 кДж = 600 кДж энергии
2) Определим сколько молекул АТФ образовалось, учитывая, что 1 молекула АТФ образует 40 кДж энергии:
600 : 40 = 15 молекул АТФ
3) Определим сколько глюкозы было израсходовано. Из условия ясно, что 1 моль глюкозы образует 38 молекул АТФ:
Из условия ясно, что 1 моль глюкозы образует 38 молекул АТФ:
С6Н12О6 + 6 О2→ 6 СО2 + 6 Н2О + 38 АТФ
Составляем пропорцию:
1 моль (C6H12O6) — 38 (АТФ)
Х моль (C6H12O6) — 15 (АТФ)
Х=1 х 15/38
Х= 0,4 моль (C6H12O6)
4) Определяем, сколько грамм глюкозы содержится в 0,4 молях, для этого опять составляем пропорцию:
1 моль (C6H12O6) = 180 гр
0,4 моль (C6H12O6) = Х гр
Х=180 х 0,4/1
Х = 72 гр (C6H12O6)
Ответ: 71 гр глюкозы израсходуют мышцы при беге продолжительность 25 минут
5: Метаболизм I — Катаболические реакции
-
- Последнее обновление
- Сохранить как PDF
Без заголовков
Для жизни нужна энергия.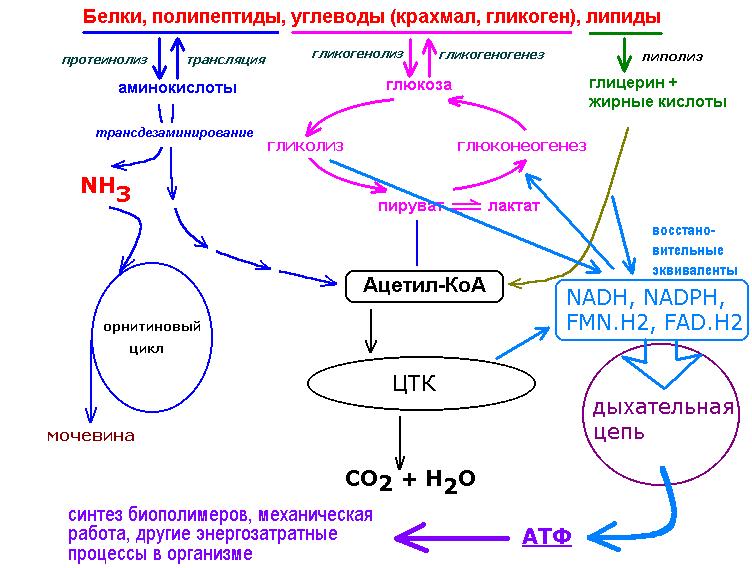 Как указывалось в нашем обсуждении биомолекул, основными функциональными компонентами клетки являются в основном полимеры — длинные цепи из более мелких индивидуальных молекулярных единиц.Каждое добавление небольшого звена в цепочку требует затрат энергии. Химические реакции, которые создают сложные молекулы из простых, известны как анаболические реакции. И наоборот, гетеротрофные организмы, такие как животные, потребляют пищу, состоящую из этих крупных полимеров, которые при расщеплении в процессе пищеварения высвобождают энергию для поддержания и построения этого организма. Такие химические реакции, в которых сложные молекулы распадаются на более простые компоненты, классифицируются как катаболические реакции.Взятые как группа реакций внутри клетки или даже организма, они могут быть названы анаболизмом или катаболизмом клетки или организма. Сумма обоих типов реакций составляет метаболизм клетки .
Как указывалось в нашем обсуждении биомолекул, основными функциональными компонентами клетки являются в основном полимеры — длинные цепи из более мелких индивидуальных молекулярных единиц.Каждое добавление небольшого звена в цепочку требует затрат энергии. Химические реакции, которые создают сложные молекулы из простых, известны как анаболические реакции. И наоборот, гетеротрофные организмы, такие как животные, потребляют пищу, состоящую из этих крупных полимеров, которые при расщеплении в процессе пищеварения высвобождают энергию для поддержания и построения этого организма. Такие химические реакции, в которых сложные молекулы распадаются на более простые компоненты, классифицируются как катаболические реакции.Взятые как группа реакций внутри клетки или даже организма, они могут быть названы анаболизмом или катаболизмом клетки или организма. Сумма обоих типов реакций составляет метаболизм клетки .
-
- 5.
 1: Гликолиз
1: Гликолиз
- 5.
-
- 5.2: Ферментация
- Гликолиз дал нам полезную энергию в виде АТФ, а также других продуктов, НАДН и пирувата. Если клетка эукариотическая и кислород доступен, то эти молекулы могут помочь вырабатывать больше АТФ.Если кислород недоступен или клетка представляет собой просто прокариот, она подвергается ферментации с образованием лактата или этилового спирта. Зачем клетке лактат или этанол? Это не так, хотя лактат может способствовать общему метаболизму.
-
- 5.3: 5.3 Цикл TCA
- Эукариоты производят огромное количество АТФ и, по-видимому, без особых усилий, используя только отбросы, оставшиеся после того, как гликолиз прошел через молекулу глюкозы: НАДН и пируват.Гликолиз у эукариот, как и его прокариотическое происхождение, происходит в цитоплазме. Цикл TCA (также называемый циклом лимонной кислоты) происходит внутри матрикса митохондрий, двухмембранной органеллы.

-
- 5.4: Окислительное фосфорилирование
- Окислительное фосфорилирование означает фосфорилирование АДФ в АТФ с использованием энергии от последовательных переносов электронов (отсюда «окислительное»). Основная идея заключается в том, что окисление НАДН, будучи сильно экзэргоничным, может генерировать энергию, необходимую для фосфорилирования АДФ.Поскольку окисление НАДН кислородом может потенциально высвободить 52 кКал / моль, а энергия, необходимая для фосфорилирования АТФ, составляет примерно 7,5 кКал / моль, мы можем ожидать образования нескольких АТФ на один окисленный НАДН.
-
- 5.5: Отключение переноса электронов от синтеза АТФ
-
- 5.6: Структура носителей электронов
-
- 5,7: 5,7 Крахмал и гликоген
- Чувствительная к гормонам липаза в жировой ткани гидролизует накопленный в этих клетках жир до глицерина и жирных кислот.
 Глицерин может войти в гликолитический цикл путем преобразования в дигидроксиацетонфосфат. Жирные кислоты секретируются из жировых клеток в кровоток, где они связываются с белком-носителем, альбумином. Этот комплекс затем может быть доставлен внутрь других клеток посредством эндоцитоза, где они могут быть расщеплены как источник энергии.
Глицерин может войти в гликолитический цикл путем преобразования в дигидроксиацетонфосфат. Жирные кислоты секретируются из жировых клеток в кровоток, где они связываются с белком-носителем, альбумином. Этот комплекс затем может быть доставлен внутрь других клеток посредством эндоцитоза, где они могут быть расщеплены как источник энергии.
- 900
-
Деполимеризация
-
8: 5.8 Распад жирных кислот
-
- 5.9: Аминокислотная деградация
- Белки расщепляются различными протеазами, которые гидролизуют пептидные связи с образованием более мелких пептидов и аминокислот. Те аминокислоты, которые не используются для создания новых белков, могут далее расщепляться, чтобы вступить в метаболические процессы, обсуждаемые в этой главе.
Миниатюра: Биохимические процессы, которые разрушают вещи от большего к меньшему, называются катаболическими процессами. Катаболические процессы часто носят окислительный характер и высвобождают энергию.Некоторая, но не вся эта энергия улавливается в виде АТФ. (CC BY-SA-NC; Кевин Ахерн и Индира Раджагопал).
(CC BY-SA-NC; Кевин Ахерн и Индира Раджагопал).
Катаболизм — обзор | Темы ScienceDirect
Некоторые косвенные индексы с использованием лимфоцитов и других факторов
Индекс катаболизма / анаболизма: Он выражает относительную часть активности катаболизма организма по отношению к его анаболической активности.
= генитально-тиреоидный индекс / GenitalratiocorrectedGenito-tyroidindex = нейтрофилы / лимфоциты = нейтрофилы / (генитальное соотношение × лимфоциты)
индекс анаболизма: Он отражает уровень анаболической активности организма.
= Индекс катаболизма / (Индекс катаболизма / анаболизма) = (Индекс катаболизма × Genitalratiocorrected × Лимфоциты) / Нейтрофилы
Индекс анаболизма оценивает абсолютную скорость анаболизма как результат кортикотропной, гонадотропной и относительной тиреотропной активности . (см. индекс катаболизма-анаболизма в разделе «Косвенные индексы с использованием нейтрофилов» и индекс катаболизма в разделе «Косвенные индексы с использованием ЛДГ или КФК» для дальнейшего обсуждения).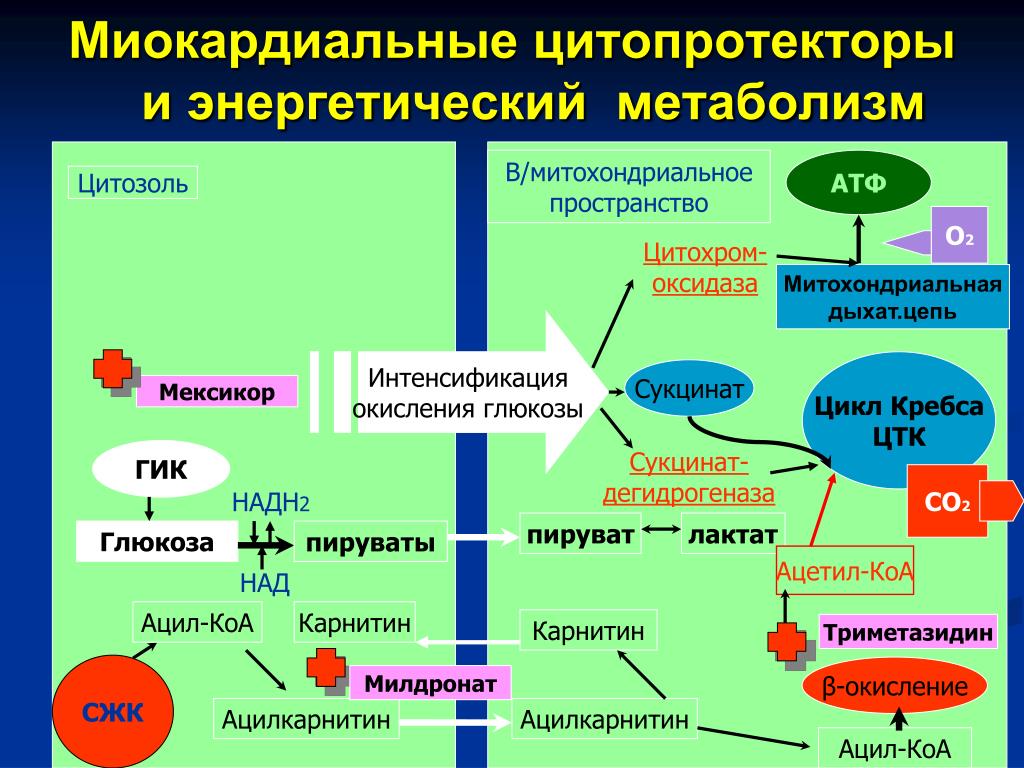 Низкая скорость катаболизма сама по себе не означает, что скорость анаболизма низкая.Каждый уровень активности может быть повышенным, низким или нормальным. Индекс анаболизма стремится оценить количественную скорость анаболизма. В числителе указан индекс катаболизма как количественная оценка катаболизма. Чем ниже абсолютный уровень катаболизма, тем выше может быть преобладание анаболизма. Однако относительная скорость катаболизма и анаболизма тем больше, чем выше преобладание анаболизма.
Низкая скорость катаболизма сама по себе не означает, что скорость анаболизма низкая.Каждый уровень активности может быть повышенным, низким или нормальным. Индекс анаболизма стремится оценить количественную скорость анаболизма. В числителе указан индекс катаболизма как количественная оценка катаболизма. Чем ниже абсолютный уровень катаболизма, тем выше может быть преобладание анаболизма. Однако относительная скорость катаболизма и анаболизма тем больше, чем выше преобладание анаболизма.
Как отмечалось выше, чем выше уровень лимфоцитов, тем хуже адаптирована катаболическая активность щитовидной железы, и, следовательно, тем ниже будет скорость катаболизма.Чем больше скорректированное генитальное соотношение, тем больше преобладание андрогенов по сравнению с эстрогенами в адаптации, что способствует завершению анаболизма.
Индекс апоптоза: Он выражает общий уровень апоптозной активности организма в целом.
= индекс структурного расширения / индекс расширения мембраны Индекс структурного расширения = индекс анаболизма × индекс нуклеомембранной активности Расширение мембраны = индекс катаболизма × скорректированный индекс роста = (анаболизм × индекс нуклеомембранной активности) / (индекс катаболизма × скорректированный индекс роста)
Апоптоз был впервые описан в 1847 г. В течение 140 лет (1847–1987) изучение апоптоза носило морфологический характер. С 1988 года, с открытием белка bcl-2, генетические механизмы апоптоза были в центре внимания. 367 С эндобиогенной точки зрения, поскольку эндокринная система управляет скоростью метаболизма клетки, она опосредует жизнь клетки и время апоптоза или некроза или их отсутствия, например, в случае раковых клеток.
В течение 140 лет (1847–1987) изучение апоптоза носило морфологический характер. С 1988 года, с открытием белка bcl-2, генетические механизмы апоптоза были в центре внимания. 367 С эндобиогенной точки зрения, поскольку эндокринная система управляет скоростью метаболизма клетки, она опосредует жизнь клетки и время апоптоза или некроза или их отсутствия, например, в случае раковых клеток.
Множество про- и антиапоптотических сигнальных факторов — это средства , регуляции апоптоза, и, хотя это интересно, они не являются детерминантом того, когда и с какой степенью интенсивности апоптоз происходит (или не происходит).Обоснованность такого индекса позволит применить глобальный подход к управлению апоптозом, который согласуется с общей схемой факторов, связанных с ростом рака, и далек от бесконечных поисков «серебряных пуль» в фармакотерапии — естественных или синтетических — которые в высшей степени эффективны. нацелены на конкретные механизмы апоптоза, но несут риск потенциально более серьезных побочных эффектов.
Числитель состоит из индекса анаболизма и индекса нуклеомембраны. Чем больше числитель, тем выше скорость апоптоза.Рост клеток происходит в результате анаболизма, который требует повышенной активности на уровне ядра в отношении транскрипции белка (представленной индексом нуклеомембраны) по сравнению с активностью мембраны. Чем выше анаболическая активность клетки, тем скорее она достигнет конца запрограммированного числа делений и, следовательно, умрет от апоптоза.
Знаменатель состоит из индекса расширения мембраны, который сам состоит из произведения катаболизма и индексов, скорректированных на индекс роста.При преобладании катаболизма 368, 369 и / или повышенной активности IGF 370, 371 мембрана расширяется. 372 Более высокая скорость расширения мембраны по сравнению со структурной активностью означает, что больше энергии тратится на клеточную гиперплазию, чем на клеточные деления, следовательно, тем больше времени требуется клетке, чтобы умереть из-за достижения запрограммированного времени смерти.
Таким образом, эндокринная система является регулятором апоптоза, а проапоптотические белки — механизмом апоптотической гибели клеток.С эндобиогенной точки зрения эндокринный подход к оценке общей физиологической скорости апоптоза позволяет оценить причину апоптоза (или его недостаточности) и определить причинные факторы, и, таким образом, позволяет разработать клинический план для устранения этих конкретных дисбалансов. . Напротив, простой подсчет количества активных про- или антиапоптозных факторов в настоящее время не предлагает пути клинического вмешательства.
Синтез АТФ
Синтез АТФ включает перенос электронов из межмембранного пространства через внутреннюю мембрану обратно в матрицу.Перенос электронов из матрицы в межмембранное пространство приводит к значительной разнице pH между двумя сторонами мембраны (около 1,4 единицы pH). Митчелл признал, что это представляет собой большую разницу в энергии, потому что хемиосмотический потенциал фактически состоит из двух компонентов. Одним из компонентов является разница в концентрации ионов водорода (pH из — pH из ), обозначенная термином ΔpH. Другой компонент следует из того факта, что протоны заряжены положительно, поэтому существует разница в электрическом потенциале , обозначенная термином ΔΨ.Протонный градиент приводит к состоянию, в котором межмембранное пространство является положительным и кислым по отношению к матрице. Условное обозначение этой ситуации: положительный выход, отрицательный вход; кислотный выход, основной в . Количественно градиент энергии через мембрану представляет собой сумму энергий этих двух компонентов градиента:
Одним из компонентов является разница в концентрации ионов водорода (pH из — pH из ), обозначенная термином ΔpH. Другой компонент следует из того факта, что протоны заряжены положительно, поэтому существует разница в электрическом потенциале , обозначенная термином ΔΨ.Протонный градиент приводит к состоянию, в котором межмембранное пространство является положительным и кислым по отношению к матрице. Условное обозначение этой ситуации: положительный выход, отрицательный вход; кислотный выход, основной в . Количественно градиент энергии через мембрану представляет собой сумму энергий этих двух компонентов градиента:
Комбинация двух компонентов обеспечивает достаточную энергию для выработки АТФ мультиферментным комплексом V митохондрии, более известным как АТФ-синтаза .(См. Рисунок 1.)
АТФ-синтаза содержит трансмембранный домен, иногда известный как субъединица F 0 , и узловатый выступ, который простирается в матрикс, субъединица F 1 . Механизм АТФ-синтазы — это не то, что можно было бы наивно предсказать. Субъединица АТФ-синтазы F 1 может выполнять свою функцию лигазы (производя АТФ из АДФ и фосфата) без протонов в матрицу; однако для высвобождения АТФ требуется поток протонов через мембрану .
Механизм АТФ-синтазы — это не то, что можно было бы наивно предсказать. Субъединица АТФ-синтазы F 1 может выполнять свою функцию лигазы (производя АТФ из АДФ и фосфата) без протонов в матрицу; однако для высвобождения АТФ требуется поток протонов через мембрану .
Существование АТФ-синтазы подразумевает, что транспорт электронов и синтез АТФ не связаны напрямую. Это подтверждается двумя экспериментальными наблюдениями: искусственный протонный градиент может привести к синтезу АТФ без транспорта электронов, а молекулы, называемые разобщителями , могут переносить протоны через мембрану, минуя АТФ-синтазу. В этом случае энергия метаболизма выделяется в виде тепла. Одним из таких разобщителей является динитрофенол, показанный на рисунке. Динитрофенол — это слабая кислота, которая достаточно гидрофобна, чтобы раствориться во внутренней мембране.Он протонируется в межмембранном пространстве и депротонируется на матричной стороне мембраны. Поскольку АТФ не производится, энергия из пищи недоступна для синтеза жира. Действительно, динитрофенол использовался в качестве диетического препарата до тех пор, пока побочные эффекты, включая токсичность для печени, не привели к его снятию с рынка.
Действительно, динитрофенол использовался в качестве диетического препарата до тех пор, пока побочные эффекты, включая токсичность для печени, не привели к его снятию с рынка.
Жирные кислоты также являются разобщителями — слабыми кислотами, которые могут проникать через внутреннюю мембрану. У младенцев тепло тела может вырабатываться так называемыми коричневыми жировыми тканями у основания шеи.Жир кажется коричневым, потому что он содержит большое количество митохондрий, а цитохромы придают ему коричневато-красный вид. Эти митохондрии находятся в естественном несвязанном состоянии. Жир окисляется, но вырабатывается очень мало АТФ; вместо этого метаболическая энергия преобразуется в тепло, чтобы мозг оставался в тепле и функционировал. К сожалению, с возрастом коричневая жировая ткань теряется, поэтому взрослые люди не могут сжечь лишние калории так легко и естественно.
Электронный транспорт и окислительное фосфорилирование
Если вы считаете, что контент, доступный через Веб-сайт (как определено в наших Условиях обслуживания), нарушает
или несколько ваших авторских прав, сообщите нам, отправив письменное уведомление («Уведомление о нарушении»), содержащее
в
информацию, описанную ниже, назначенному ниже агенту.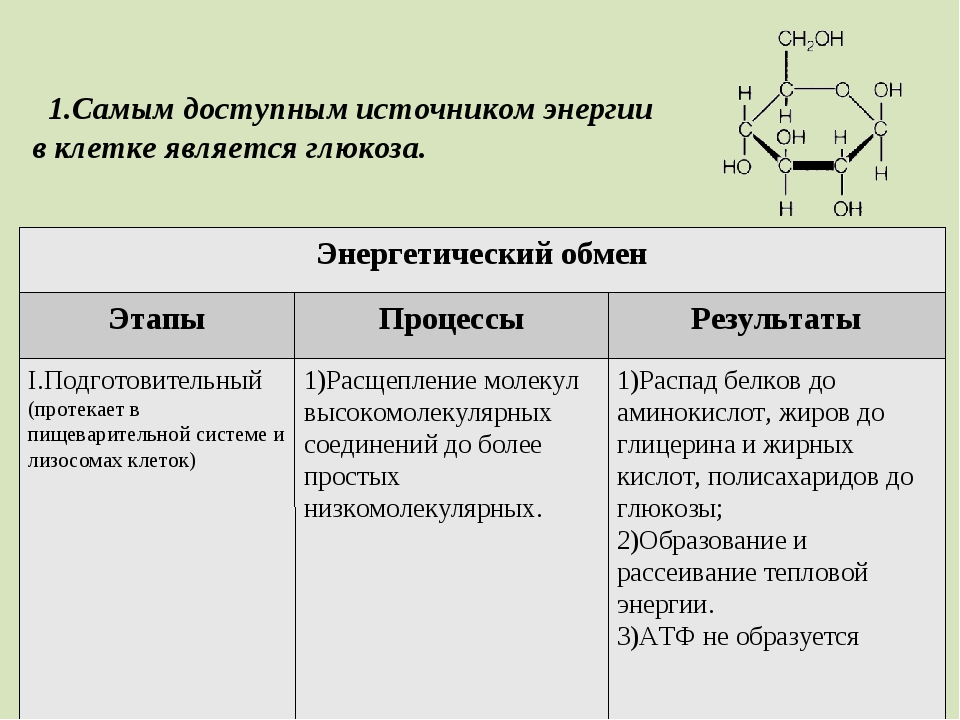 Если репетиторы университета предпримут действия в ответ на
Если репетиторы университета предпримут действия в ответ на
ан
Уведомление о нарушении, оно предпримет добросовестную попытку связаться со стороной, которая предоставила такой контент
средствами самого последнего адреса электронной почты, если таковой имеется, предоставленного такой стороной Varsity Tutors.
Ваше Уведомление о нарушении прав может быть отправлено стороне, предоставившей доступ к контенту, или третьим лицам, таким как
в качестве
ChillingEffects.org.
Обратите внимание, что вы будете нести ответственность за ущерб (включая расходы и гонорары адвокатам), если вы существенно
искажать информацию о том, что продукт или действие нарушает ваши авторские права.Таким образом, если вы не уверены, что контент находится
на Веб-сайте или по ссылке с него нарушает ваши авторские права, вам следует сначала обратиться к юристу.
Чтобы отправить уведомление, выполните следующие действия:
Вы должны включить следующее:
Физическая или электронная подпись правообладателя или лица, уполномоченного действовать от их имени;
Идентификация авторских прав, которые, как утверждается, были нарушены;
Описание характера и точного местонахождения контента, который, по вашему мнению, нарушает ваши авторские права, в \
достаточно подробностей, чтобы позволить репетиторам университетских школ найти и точно идентифицировать этот контент; например нам требуется
а
ссылка на конкретный вопрос (а не только на название вопроса), который содержит содержание и описание
к какой конкретной части вопроса — изображению, ссылке, тексту и т. д. — относится ваша жалоба;
д. — относится ваша жалоба;
Ваше имя, адрес, номер телефона и адрес электронной почты; а также
Ваше заявление: (а) вы добросовестно считаете, что использование контента, который, по вашему мнению, нарушает
ваши авторские права не разрешены законом, владельцем авторских прав или его агентом; (б) что все
информация, содержащаяся в вашем Уведомлении о нарушении, является точной, и (c) под страхом наказания за лжесвидетельство, что вы
либо владелец авторских прав, либо лицо, уполномоченное действовать от их имени.
Отправьте жалобу нашему уполномоченному агенту по адресу:
Чарльз Кон
Varsity Tutors LLC
101 S. Hanley Rd, Suite 300
St. Louis, MO 63105
Или заполните форму ниже:
III стадия катаболизма
20.4 Стадия III катаболизма
Цели обучения
- Опишите реакции цикла лимонной кислоты.

- Опишите функцию цикла лимонной кислоты и укажите производимые продукты.
- Опишите роль цепи переноса электронов в энергетическом обмене.
- Опишите роль окислительного фосфорилирования в энергетическом обмене.
Ацетильная группа вступает в циклическую последовательность реакций, известную под общим названием цикл лимонной кислоты (или цикл Кребса или цикл трикарбоновой кислоты [TCA]). Циклическая последовательность реакций, которая вызывает окисление двухкомпонентного звена до диоксида углерода и воды. .. Циклический дизайн этой сложной серии реакций, которые вызывают окисление ацетильной группы ацетил-КоА до диоксида углерода и воды, был впервые предложен Гансом Кребсом в 1937 году (он был удостоен Нобелевской премии по физиологии и медицине 1953 года. .) Вступление ацетил-КоА в цикл лимонной кислоты является началом III стадии катаболизма. Цикл лимонной кислоты производит аденозинтрифосфат (АТФ), восстановленный никотинамидадениндинуклеотид (НАДН), восстановленный флавинадениндинуклеотид (FADH 2 ) и промежуточные продукты метаболизма для синтеза необходимых соединений.
Этапы цикла лимонной кислоты
На первый взгляд цикл лимонной кислоты кажется довольно сложным (Рисунок 20.12 «Реакции цикла лимонной кислоты»). Однако все реакции являются знакомыми типами в органической химии: гидратация, окисление, декарбоксилирование и гидролиз. Каждая реакция цикла лимонной кислоты пронумерована, и на рисунке 20.12 «Реакции цикла лимонной кислоты» два ацетильных атома углерода выделены красным. Каждое промежуточное соединение в цикле представляет собой карбоновую кислоту, существующую в виде аниона при физиологическом pH.Все реакции происходят в митохондриях, которые представляют собой небольшие органеллы в клетках растений и животных. Мы более подробно рассмотрим структуру митохондрий в Разделе 20.5 «Стадия II углеводного катаболизма».
Рисунок 20.12 Реакции цикла лимонной кислоты
В первой реакции ацетил-КоА входит в цикл лимонной кислоты, и ацетильная группа переносится на оксалоацетат, давая цитрат. Обратите внимание, что на этом этапе высвобождается кофермент А.Реакция катализируется цитрат-синтазой .
На следующем этапе аконитаза катализирует изомеризацию цитрата в изоцитрат. В этой реакции третичный спирт, который не может быть окислен, превращается во вторичный спирт, который можно окислить на следующей стадии.
Изоцитрат затем подвергается реакции, известной как окислительное декарбоксилирование, потому что спирт окисляется, и молекула укорачивается на один атом углерода с высвобождением диоксида углерода (декарбоксилирование).Реакция катализируется изоцитратдегидрогеназой , а продуктом реакции является α-кетоглутарат. Важной реакцией, связанной с этим, является восстановление кофермента никотинамидадениндинуклеотида (NAD + ) до NADH. НАДН в конечном итоге повторно окисляется, и высвобождаемая энергия используется для синтеза АТФ, как мы увидим.
Четвертая стадия — это еще одно окислительное декарбоксилирование. На этот раз α-кетоглутарат превращается в сукцинил-КоА, а другая молекула NAD + восстанавливается до NADH.Комплекс α-кетоглутаратдегидрогеназы катализирует эту реакцию. Это единственная необратимая реакция в цикле лимонной кислоты. Таким образом, он предотвращает работу цикла в обратном направлении, в котором ацетил-КоА будет синтезироваться из диоксида углерода.
Комментарий : Пока что на первых четырех этапах два атома углерода вошли в цикл как ацетильная группа, а два атома углерода высвободились как молекулы диоксида углерода. Остальные реакции цикла лимонной кислоты используют четыре атома углерода сукцинильной группы для повторного синтеза молекулы оксалоацетата, который является соединением, необходимым для объединения с входящей ацетильной группой и начала следующего цикла цикла.
В пятой реакции энергия, выделяемая при гидролизе высокоэнергетической тиоэфирной связи сукцинил-КоА, используется для образования гуанозинтрифосфата (ГТФ) из дифосфата гуанозина (GDP) и неорганического фосфата в реакции, катализируемой сукцинил-КоА. синтетаза . Эта стадия — единственная реакция в цикле лимонной кислоты, которая непосредственно образует высокоэнергетическое фосфатное соединение. GTP может легко переносить свою концевую фосфатную группу на аденозиндифосфат (ADP) для генерации АТФ в присутствии нуклеозиддифосфокиназы .
Сукцинатдегидрогеназа затем катализирует удаление двух атомов водорода из сукцината с образованием фумарата. В этой окислительно-восстановительной реакции в качестве окислителя используется флавинадениндинуклеотид (FAD), а не NAD + . Сукцинатдегидрогеназа — единственный фермент цикла лимонной кислоты, расположенный внутри внутренней митохондриальной мембраны. Скоро мы увидим важность этого.
На следующем этапе молекула воды добавляется к двойной связи фумарата с образованием L-малата в реакции, катализируемой фумаразой .
Один оборот цикла завершается окислением L-малата до оксалоацетата, вызываемым малатдегидрогеназой . Это третья окислительно-восстановительная реакция, в которой в качестве окислителя используется NAD + . Оксалоацетат может принимать ацетильную группу от ацетил-КоА, позволяя циклу снова начинаться.
Клеточное дыхание
Дыхание можно определить как процесс, при котором клетки окисляют органические молекулы в присутствии газообразного кислорода с образованием углекислого газа, воды и энергии в форме АТФ.Мы видели, что два атома углерода входят в цикл лимонной кислоты из ацетил-КоА (этап 1), а два разных атома углерода выходят из цикла в виде диоксида углерода (этапы 3 и 4). Однако нигде в нашем обсуждении цикла лимонной кислоты мы не указывали, как используется кислород. Напомним, однако, что на четырех стадиях окисления-восстановления, происходящих в цикле лимонной кислоты, кофермент NAD + или FAD восстанавливается до NADH или FADH 2 соответственно. Кислород необходим для повторного окисления этих коферментов .Напомним также, что очень мало АТФ получается непосредственно из цикла лимонной кислоты. Вместо этого участие кислорода и значительная продукция АТФ происходят после цикла лимонной кислоты двумя путями, которые тесно связаны: перенос электронов и окислительное фосфорилирование.
Все ферменты и коферменты для цикла лимонной кислоты, повторного окисления NADH и FADH 2, и производства АТФ расположены в митохондриях Маленькие овальные органеллы с двойными мембранами; «электростанции» клетки., которые представляют собой маленькие овальные органеллы с двойными мембранами, часто называемые «энергетическими установками» клетки (рис. 20.13 «Дыхание»). Клетка может содержать от 100 до 5000 митохондрий, в зависимости от ее функции, и митохондрии могут воспроизводить себя, если потребности клетки в энергии увеличиваются.
Рисунок 20.13 Дыхание
В митохондриях происходит клеточное дыхание.
На рисунке 20.13 «Дыхание» показаны две мембраны митохондрии: внешняя и внутренняя .Внутренняя мембрана сильно сложена в серию внутренних гребней, называемых cristae . Таким образом, в митохондриях есть два компартмента: межмембранное пространство , , которое находится между мембранами, и матрица , , которая находится внутри внутренней мембраны. Наружная мембрана проницаема, тогда как внутренняя мембрана непроницаема для большинства молекул и ионов, хотя вода, кислород и углекислый газ могут свободно проникать через обе мембраны. Матрикс содержит все ферменты цикла лимонной кислоты, за исключением сукцинатдегидрогеназы, которая встроена во внутреннюю мембрану.Ферменты, которые необходимы для повторного окисления НАДН и ФАДН 2 и производства АТФ, также расположены во внутренней мембране. Они расположены на определенных позициях, поэтому действуют аналогично бригаде ковшей. Эта высокоорганизованная последовательность окислительно-восстановительных ферментов известна как цепь переноса электронов (или дыхательная цепь). Организованная последовательность окислительно-восстановительных реакций, которая в конечном итоге переносит электроны к кислороду, восстанавливая его до воды.
Электронный транспорт
Рисунок 20.14 «Митохондриальная цепь переноса электронов и АТФ-синтаза» иллюстрирует организацию цепи переноса электронов. Компоненты цепи организованы в четыре комплекса, обозначенные I, II, III и IV. Каждый комплекс содержит несколько ферментов, другие белки и ионы металлов. Ионы металлов можно восстанавливать, а затем многократно окислять по мере перехода электронов от одного компонента к другому. Вспомните из главы 5 «Введение в химические реакции», раздела 5.5 «Окислительно-восстановительные (окислительно-восстановительные) реакции», что соединение восстанавливается, когда оно приобретает электроны или атомы водорода, и окисляется, когда оно теряет электроны или атомы водорода.
Рисунок 20.14 Митохондриальная электронная транспортная цепь и АТФ-синтаза
Красная линия показывает путь электронов.
Электроны могут входить в цепь переноса электронов через комплекс I или II. Сначала мы рассмотрим электроны, входящие в комплекс I. Эти электроны происходят из НАДН, который образуется в трех реакциях цикла лимонной кислоты. Давайте возьмем стадию 8 в качестве примера, реакцию, в которой L-малат окисляется до оксалоацетата, а NAD + восстанавливается до NADH.Эту реакцию можно разделить на две половинные реакции:
Полуреакция окисления :
Полуреакция восстановления :
В ходе полуреакции окисления два иона водорода (H + ) и два электрона удаляются из подложки. В полуреакции восстановления молекула NAD + принимает оба этих электрона и один из ионов H + .Другой ион H + транспортируется из матрикса через внутреннюю митохондриальную мембрану в межмембранное пространство. НАДН диффундирует через матрицу и связывается комплексом I транспортной цепи электронов. В комплексе кофермент флавинмононуклеотид (FMN) принимает оба электрона от NADH. Пропуская электроны, НАДН окисляется обратно до НАД + , а FMN восстанавливается до FMNH 2 (восстановленная форма мононуклеотида флавина). Опять же, реакцию можно проиллюстрировать, разделив ее на соответствующие полуреакции.
Полуреакция окисления :
Полуреакция восстановления :
Комплекс I содержит несколько белков, которые имеют центры железо-сера (Fe · S). Электроны, восстановившие FMN до FMNH 2 , теперь переносятся на эти белки. Ионы железа в центрах Fe · S сначала находятся в форме Fe (III), но, принимая электрон, каждый ион восстанавливается до формы Fe (II).Поскольку каждый центр Fe · S может переносить только один электрон, необходимы два центра, чтобы принять два электрона, которые будут регенерировать FMN.
Полуреакция окисления :
FMNH 2 → FMN + 2H + + 2e —
Полуреакция восстановления :
2Fe (III) · S + 2e — → 2Fe (II) · S
Электроны из FADH 2 , образованные на стадии 6 цикла лимонной кислоты, входят в цепь переноса электронов через комплекс II.Сукцинатдегидрогеназа, фермент в цикле лимонной кислоты, который катализирует образование FADH 2 из FAD, является частью комплекса II. Электроны от FADH 2 затем переносятся на белок Fe · S.
Полуреакция окисления :
FADH 2 → FAD + 2H + + 2e —
Полуреакция восстановления :
2Fe (III) · S + 2e — → 2Fe (II) · S
Электроны из комплексов I и II затем переносятся от белка Fe · S к коферменту Q (CoQ), мобильному переносчику электронов, который действует как переносчик электронов между комплексами I или II и комплексом III.
Примечание
Коэнзим Q
также называют убихиноном , потому что он повсеместно встречается в живых системах.
Полуреакция окисления :
2Fe (II) · S → 2Fe (III) · S + 2e —
Полуреакция восстановления :
Комплексы III и IV включают несколько железосодержащих белков, известных как цитохромы. Белок, содержащий порфирин железа, в котором железо может чередоваться между Fe (II) и Fe (III).. Железо в этих ферментах находится в субструктурах, известных как порфирины железа (рис. 20.15 «Порфирин железа»). Подобно центрам Fe · S, характерной особенностью цитохромов является способность их атомов железа существовать либо в виде Fe (II), либо в виде Fe (III). Таким образом, каждый цитохром в его окисленной форме — Fe (III) — может принимать один электрон и восстанавливаться до формы Fe (II). Это изменение степени окисления обратимо, поэтому восстановленная форма может отдать свой электрон следующему цитохрому и так далее. Комплекс III содержит цитохромы b и c, а также белки Fe · S, причем цитохром c действует как переносчик электронов между комплексом III и IV.Комплекс IV содержит цитохромы а и 3 в ферменте, известном как цитохромоксидаза . Этот фермент обладает способностью передавать электроны молекулярному кислороду, последнему акцептору электронов в цепи реакций переноса электронов. На этом заключительном этапе образуется вода (H 2 O).
Полуреакция окисления :
4Cyt a 3 –Fe (II) → 4Cyt a 3 –Fe (III) + 4e —
Полуреакция восстановления :
O 2 + 4H + + 4e — → 2H 2 O
Рисунок 20.15 Железный порфирин
Порфирины железа присутствуют в цитохромах, а также в миоглобине и гемоглобине.
Окислительное фосфорилирование
Каждое промежуточное соединение в цепи переноса электронов восстанавливается путем добавления одного или двух электронов в одну реакцию, а затем восстанавливается до своей исходной формы путем доставки электронов к следующему соединению по цепи. Последовательные передачи электронов приводят к производству энергии.Но как эта энергия используется для синтеза АТФ? Процесс, который связывает синтез АТФ с работой цепи переноса электронов, называется окислительным фосфорилированием. Процесс, который связывает синтез АТФ с работой цепи переноса электронов.
Электронный транспорт тесно связан с окислительным фосфорилированием. Коферменты НАДН и ФАДН 2, окисляются дыхательной цепью только , если АДФ одновременно фосфорилируется до АТФ.В настоящее время принятая модель, объясняющая, как эти два процесса связаны между собой, известна как хемиосмотическая гипотеза , предложенная Питером Митчеллом, в результате чего Митчелл был удостоен Нобелевской премии по химии 1978 года.
Снова посмотрев на рисунок 20.14 «Митохондриальная цепь переноса электронов и АТФ-синтаза», мы видим, что по мере того, как электроны переносятся по цепи переноса электронов, ионы водорода (H + ) переносятся через внутреннюю митохондриальную мембрану из матрикса. в межмембранное пространство.Концентрация H + уже выше в межмембранном пространстве, чем в матрице, поэтому для транспортировки туда дополнительного H + требуется энергия. Эта энергия возникает из-за реакций переноса электрона в цепи переноса электронов. Но как тогда резкая разница в концентрации H + приводит к синтезу АТФ? Накопление ионов H + в межмембранном пространстве приводит к ионному градиенту H + , который является большим источником энергии, как вода за плотиной (потому что, если есть возможность, протоны будут вытекать из межмембранного пространства и в менее концентрированную матрицу).Текущие исследования показывают, что поток H + вниз по этому градиенту концентрации через пятый ферментный комплекс, известный как АТФ-синтаза, приводит к изменению структуры синтазы, вызывая синтез и высвобождение АТФ.
В клетках, которые используют энергию, оборот АТФ очень высок, поэтому эти клетки содержат высокие уровни АДФ. Поэтому они должны постоянно потреблять большое количество кислорода, чтобы иметь энергию, необходимую для фосфорилирования АДФ с образованием АТФ.Рассмотрим, например, что скелетные мышцы в состоянии покоя потребляют около 30% кислорода, потребляемого взрослым в состоянии покоя, но когда одни и те же мышцы интенсивно работают, на них приходится почти 90% общего потребления кислорода организмом.
Эксперимент показал, что 2,5–3 молекулы АТФ образуются на каждую молекулу НАДН, окисляемую в цепи переноса электронов, и 1,5–2 молекулы АТФ образуются на каждую молекулу окисленного FADH 2 . Таблица 20.2 «Максимальный выход АТФ от полного окисления 1 моля ацетил-КоА» суммирует теоретический максимальный выход АТФ, полученного при полном окислении 1 моль ацетил-КоА посредством последовательного действия цикла лимонной кислоты, электронного транспортная цепь и окислительное фосфорилирование.
Таблица 20.2 Максимальный выход АТФ от полного окисления 1 моль ацетил-КоА
| Реакция | Комментарии | Выход АТФ (моль) |
|---|---|---|
| Изоцитрат → α-кетоглутарат + CO 2 | производит 1 моль НАДН | |
| α-кетоглутарат → сукцинил-КоА + CO 2 | производит 1 моль НАДН | |
| Сукцинил-КоА → сукцинат | производит 1 моль GTP | +1 |
| сукцинат → фумарат | производит 1 моль FADH 2 | |
| Малат → оксалоацетат | производит 1 моль НАДН | |
| 1 FADH 2 из цикла лимонной кислоты | дает 2 моль АТФ | +2 |
| 3 НАДН из цикла лимонной кислоты | дает 3 моль АТФ / НАДН | +9 |
| Чистый выход ATP: | +12 | |
Упражнения по обзору концепции
-
Какова основная функция цикла лимонной кислоты?
-
Два атома углерода вводятся в цикл лимонной кислоты в виде ацетил-КоА.В какой форме два атома углерода удаляются из цикла?
-
Что такое митохондрии и какова их функция в клетке?
ответов
-
полное окисление атомов углерода до диоксида углерода и образование высокоэнергетического фосфатного соединения, богатых энергией восстановленных коферментов (НАДН и ФАДН 2 ) и промежуточных продуктов метаболизма для синтеза других соединений
-
Митохондрии — это небольшие органеллы с двойной мембраной, которые содержат ферменты и другие молекулы, необходимые для производства большей части АТФ, необходимого организму.
Основные выводы
- Ацетильная группа ацетил-КоА входит в цикл лимонной кислоты. Для каждого ацетил-КоА, который входит в цикл лимонной кислоты, продуцируются 2 молекулы диоксида углерода, 3 молекулы НАДН, 1 молекула АТФ и 1 молекула FADH 2 .
- Восстановленные коферменты (НАДН и ФАДН 2 ), продуцируемые циклом лимонной кислоты, повторно окисляются реакциями цепи переноса электронов.Эта серия реакций также вызывает градиент pH на внутренней митохондриальной мембране.
- Градиент pH, создаваемый цепью переноса электронов, стимулирует синтез АТФ из АДФ. На каждый повторно окисленный НАДН продуцируется 2,5–3 молекулы АТФ; на каждый повторно окисленный FADH 2 продуцируется 1,5–2 молекулы АТФ.
Упражнения
-
Замените каждый вопросительный знак правильным составом.
- ? → изоцитрат аконитазы
- ? +? → цитрат-синтаза цитрат + кофермент А
- фумарат → фумараза?
- изоцитрат + НАД + →? α-кетоглурат + НАДН + СО2
-
Замените каждый вопросительный знак правильным составом.
- малат + НАД + →? оксалоацетат + НАДН
- ? +? → нуклеозид дифосфокиназа GDP + ATP
- сукцинил-КоА → сукцинил-КоА синтетаза? +?
- сукцинат + FAD → сукцинатдегидрогеназа? + ФАДх3
-
Из реакций в упражнениях 1 и 2 выберите уравнение (я) по номеру и букве, в котором происходит каждый тип реакции.
- изомеризация
- гидратация
- синтез
-
Из реакций в упражнениях 1 и 2 выберите уравнение (я) по номеру и букве, в котором происходит каждый тип реакции.
- окисление
- декарбоксилирование
- фосфорилирование
-
Какую аналогичную роль кофермент Q и цитохром c выполняют в цепи переноса электронов?
-
Что такое акцептор электронов в конце цепи переноса электронов? В какой продукт восстанавливается это соединение?
-
Какова функция цитохромов в цепи переноса электронов?
-
- Что подразумевается под этим утверждением? «Электронный транспорт тесно связан с окислительным фосфорилированием.”
- Как связаны или связаны электронный транспорт и окислительное фосфорилирование?
ответов
-
- цитрат
- оксалоацетат + ацетил-КоА
- малат
- α-кетоглутарат гидрогеназный комплекс
-
- реакция в 1а
- реакция в 1c
- реакция в 1b
-
Обе молекулы служат переносчиками электронов между комплексами цепи переноса электронов.
-
Цитохромы представляют собой белки в цепи переноса электронов и служат одноэлектронными переносчиками.
Фотосинтез против дыхания | BioNinja
Во многих отношениях фотосинтез и клеточное дыхание являются взаимодополняющими реакциями в окружающей среде
- Продукты фотосинтеза действуют как входы клеточного дыхания (кислород и глюкоза)
- Продукты клеточного дыхания действуют как входы фотосинтеза (углерод диоксид и вода)
Большинство производителей (т.е.е. фотоавтотрофы) для выживания осуществляют как фотосинтез, так и клеточное дыхание
- Потребители (т.е. гетеротрофы) осуществляют только клеточное дыхание, но будут глотать или поглощать продукты фотосинтеза
Взаимосвязь между процессами
Сходства
Фотосинтез и клеточное дыхание включают производство химической энергии (АТФ)
- При фотосинтезе АТФ производится с помощью световой энергии (фотофосфорилирование) и используется для образования органических молекул
- В клеточном дыхании производится АТФ путем разрушения органических молекул (окислительное фосфорилирование)
В обоих случаях производство АТФ включает цепь переноса электронов и хемиосмос
- При фотосинтезе электроны отдают хлорофилл, а протоны накапливаются в просвете тилакоида
- При клеточном дыхании электроны передаются переносчиками водорода, а протоны накапливаются в межмембранном пространстве
Сравнение производства АТФ
Различия
Фотосинтез и клеточное дыхание во многих отношениях противоположны друг другу
- Фотосинтез — анаболический процесс, тогда как клеточное дыхание — катаболический процесс
Фотосинтез :
Вода расщепляется на кислород, чтобы высвободить электроны для цепи переноса электронов
Дыхание клеток :
- Использует цикл Кребса для расщепления глюкозы (высвобождает переносчики водорода и диоксид углерода
- Носители водорода высвобождают электроны для цепи переноса электронов (в частности, NADH и FADH 2 )
- Электроны из транспортная цепь поглощается кислородом (t o образуют воду)
Различия между фотосинтезом и дыханием клеток
Биосинтез АТФ
Аденозинтрифосфат (АТФ) — это высокоэнергетическая молекула, присутствующая в живых клетках.Это нуклеозидтрифосфат, который обеспечивает клетки энергией для метаболизма и используется в нескольких клеточных процессах, включая синтез ключевых биомолекул.
АТФ обычно присутствует в клетках в концентрации 1-10 мМ — он непрерывно рециркулируется в организмах, чтобы обеспечить постоянный запас энергии для клеточных процессов.
АТФ можно синтезировать с помощью окислительно-восстановительных реакций, в которых в качестве источника энергии используются простые и сложные липиды или углеводы. Сложные источники энергии необходимо переваривать в более простые молекулы, прежде чем использовать их в синтезе АТФ.Сложные углеводы обычно гидролизуются до глюкозы и фруктозы, а триглицериды метаболизируются с образованием глицерина и жирных кислот.
Биосинтез АТФ путем окислительного фосфорилирования и фотофосфорилирования является основным путем производства энергии животными, растениями и микробами. Производство АТФ эукариотами обычно происходит в митохондриях клетки.
Важными путями, с помощью которых эукариоты генерируют энергию, являются гликолиз, цикл лимонной кислоты (или цикл Креба) и цепь переноса электронов (или путь окислительного фосфорилирования).
Вместе эти 3 стадии называются клеточным дыханием. У людей клеточное дыхание превращает аденозиндифосфат (АДФ) в АТФ и, таким образом, высвобождает энергию из молекул, которые богаты энергией.
Гликолиз
Гликолиз включает метаболизм глюкозы и глицерина с образованием пирувата. Эти реакции происходят в цитоплазме большинства организмов и высвобождают 2 АТФ. Здесь глюкоза превращается в пируват путем фосфорилирования с помощью двух ключевых ферментов — фосфоглицераткиназы и пируваткиназы.
Общую реакцию можно представить следующим образом:
Глюкоза + 2 NAD + + 2 ADP + 2 P i à 2 пируват + 2 NADH + 2 H + + 2 ATP + 2 H 2 O
Таким образом, каждая молекула глюкозы подвергается гликолизу с образованием 2 пирувата. Две восстановленные молекулы никотинамидадениндинуклеотида (НАДН) и 2 молекулы воды (H 2 O) также образуются в результате гликолиза. Молекулы НАДН окисляются в цепи переноса электронов с образованием АТФ, а полученный пируват используется в качестве субстрата для цикла Креба.
Цикл Креба
Цикл Креба также известен как цикл трикарбоновой кислоты (ТСА) и происходит в митохондриях. Он включает серию реакций, в результате которых пируват разлагается на CO 2 , АТФ, воду и электроны.
Пируват, продуцируемый гликолизом в цитоплазме, превращается в ацетил-кофермент А (ацетил-КоА) в митохондриях. Ацетил-КоА превращается в цитрат, который затем подвергается серии реакций окислительно-восстановительного восстановления, гидратации, дегидратации и декарбоксилирования с образованием изоцитрата, альфа-кетоглютерата, сукцинл-КоА, фумерата и малата.
Эти реакции катализируются несколькими ключевыми ферментами пути, такими как цитратсинтаза, аконитаза, изоцитратдегидрогеназа и малатдегидрогеназа. В целом цикл Креба дает 2 молекулы АТФ, 6 молекул НАДН и 2 молекулы восстановленного флавинадениндинуклеотида (FADH 2 ).
Ацетил-КоА, полученный в результате метаболизма углеводов, жиров и белков в клетках, используется в цикле Креба для выработки энергии. Следовательно, это важный метаболический путь, объединяющий углеводный, жировой и белковый обмен в живых организмах.
НАДН и ФАДН 2 молекул, образующихся как побочные продукты цикла лимонной кислоты, подаются в цепь переноса электронов, где они окисляются с образованием АТФ с помощью фермента АТФ-синтазы. Этот фермент присутствует в митохондриях и катализирует производство АТФ, объединяя АДФ и неорганический фосфат.
АТФ-синтазу
часто называют сложной молекулярной машиной, у которой есть центральный ротор, который движется со скоростью 150 оборотов в секунду во время синтеза АТФ.
Всего каждая молекула глюкозы, подвергающаяся клеточному дыханию, производит 38 молекул АТФ — 2 АТФ из гликолиза, 2 АТФ из цикла Креба и 34 АТФ из цепи переноса электронов.