до, во время и после забега
Очередной выпуск рубрики «Азбука бегуна» в социальных сетях мы недавно посвятили слову «гликоген». Это энергетический запас спортсмена. Выносливость стайера напрямую зависит от объёмов гликогена в его организме. В комментариях к этому посту нам задали вопрос: какие продукты максимально помогают обогатить организм гликогеном — до, во время и после забега? Сегодня на него отвечает спортивный диетолог и нутрициолог Тина Белякова.
Гликогеновое «депо» — это энергетический пул человека, который расходуется в первую очередь. Он базируется в печени и внутри мышц. Соответственно, чем больше мышечная масса, тем выше запасы гликогена. Когда вы начинаете интенсивно тренироваться, происходит постепенное расщепление молекул глюкозы для обеспечения организма энергией. Вы наверняка слышали выражение «загрузиться углеводами перед тренировкой». Чем же это лучше всего делать? Любой углевод, сложный или простой — глюкоза.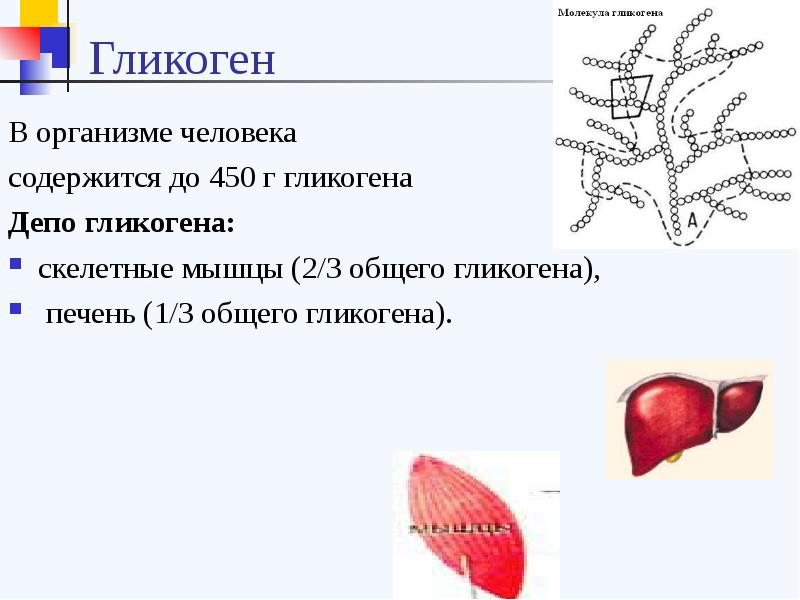 Разница в длине цепочки. Простые углеводы содержат односложные молекулы. Усваиваются быстро, в течение получаса. Сложные действуют до 6-ти часов. Они расщепляются постепенно, отсоединяя единичные молекулы на энергию. Но суть, как вы догадались, одна. Да и белки в результате глюконеогенеза могут распадаться на глюкозу. Приведу пример: гречка усваивается и даёт энергию в течение пяти часов. Рафинированный сахар — за 15-20 минут.
Разница в длине цепочки. Простые углеводы содержат односложные молекулы. Усваиваются быстро, в течение получаса. Сложные действуют до 6-ти часов. Они расщепляются постепенно, отсоединяя единичные молекулы на энергию. Но суть, как вы догадались, одна. Да и белки в результате глюконеогенеза могут распадаться на глюкозу. Приведу пример: гречка усваивается и даёт энергию в течение пяти часов. Рафинированный сахар — за 15-20 минут.
Всем известно, что мозг питается исключительно глюкозой. Ему требуется 2 грамма сахара в час. Сладкоежки могут пользоваться этой уловкой и при этом оставаться стройными. Будьте внимательны с фруктозой! Её избыток ведёт к ожирению быстрее, чем рафинад. Всё дело — в специфике влияния на инсулин. Если глюкоза способна использоваться всеми клетками организма в качестве источника энергии, то фруктоза может перерабатываться только печенью. Поэтому я не рекомендую заедать основные приёмы пищи фруктами, тем более запивать соками.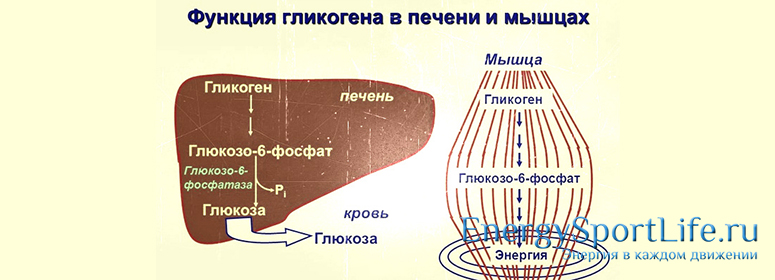 Если запасы гликогена истощаются, скорость и выносливость спортсмена начинают падать. Подкожный жир также может использоваться для работы. Для окисления ему необходим кислород. Этот вариант подходит бегунам, а для силовых нагрузок (анаэробная работа) такой вид топлива недоступен.
Если запасы гликогена истощаются, скорость и выносливость спортсмена начинают падать. Подкожный жир также может использоваться для работы. Для окисления ему необходим кислород. Этот вариант подходит бегунам, а для силовых нагрузок (анаэробная работа) такой вид топлива недоступен.
Поговорим о питании. Людям, ориентированным на спортивные показатели, нужно придерживаться высокоуглеводного рациона. До 60% общей калорийности будет приходиться на углеводы, 20% на белковую пищу и примерно столько же на жиры. За час-полтора до тренировки рекомендую комплексный приём пищи, содержащей сложные углеводы. К ним относятся гречка, киноа, булгур, полба, перловка, макароны из твёрдых сортов пшеницы, необдирные сорта риса и так далее. Как источник углеводов и белка можно использовать бобовые культуры, если нет пищевой непереносимости. Такие как нут, маш, чечевица и фасоль.
Если есть задача убрать несколько лишних килограммов, стоит взять на контроль потребление сухофруктов, бананов, винограда, хурмы, хлеба (в том числе бездрожжевого). Не говоря уже о сладостях, в них организм и вовсе не нуждается. Он и без того найдёт, где взять глюкозу (смотрите выше). Во время забега на помощь придёт изотоник. Об этих напитках я уже рассказывала.
Не говоря уже о сладостях, в них организм и вовсе не нуждается. Он и без того найдёт, где взять глюкозу (смотрите выше). Во время забега на помощь придёт изотоник. Об этих напитках я уже рассказывала.
Сразу после забега многие используют простые углеводы. Хотя, на мой взгляд, нужно подпитать мышцы, которые обеспечивали движение. Как вариант подойдёт протеиновый коктейль. Через пару часов после интенсивной нагрузки включите в рацион белки и углеводы. Организм нуждается в восстановлении. Это может быть овощной салат, а также рис с птицей (или нежирной говядиной, кроликом, рыбой, морепродуктами).
Не забывайте про питьевой режим, чтобы не допустить дегидратации тканей. Спортсмен вдвойне нуждается в богатом рационе. А это и витамины, и минералы. Поэтому своевременно проверяйте анализы и включайте всё необходимое, чтоб не было сбоев в работе организма.
фото: yogobe. сom, личный архив Тины Беляковой, myslide. ru, lasvegasnews.
 media, vrachmedik. ru, img.fabryka. fit, multivkus. ru
media, vrachmedik. ru, img.fabryka. fit, multivkus. ru
Энергетические депо организма
Ограниченность запасов АТФ и, как следствие, непрерывный процесс её ресинтеза, осуществляемый разными путями, обуславливает существование в организме человека форм и механизмов запасания и хранения энергии.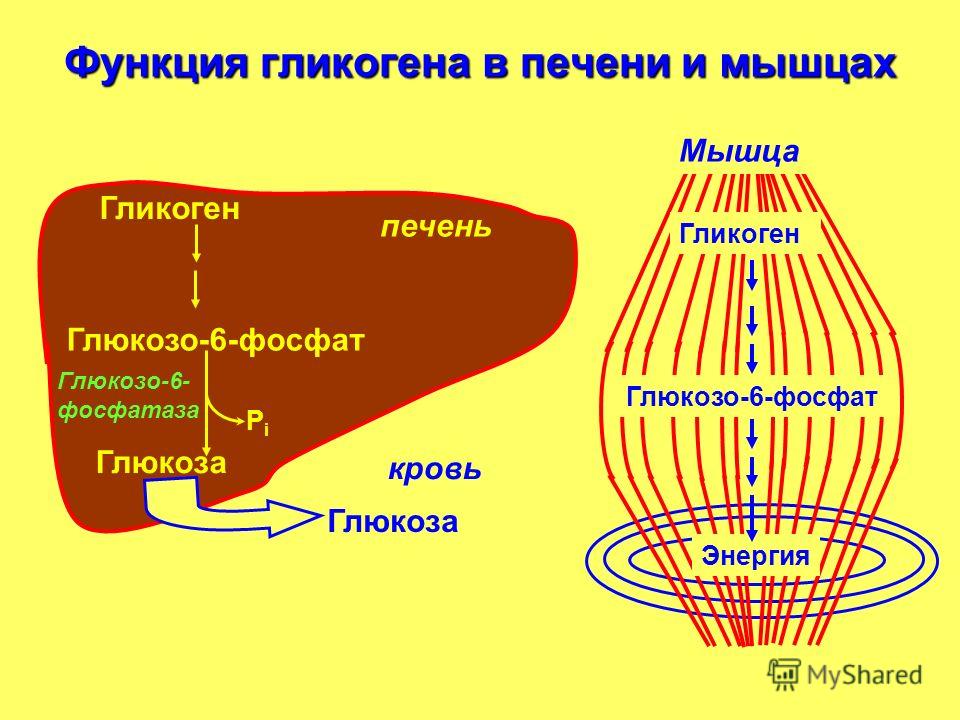 В независимости от субстратов, участвующих в конечных биоэнергетических реакциях, 99% образующейся энергии появляется за счёт расщепления углеводов, триглицеридов (жиров) и белков. Таким образом, именно депонирование стабильных форм данных типов химических соединений является необходимым условием для оптимального функционирования органов и систем в различных условиях.
В независимости от субстратов, участвующих в конечных биоэнергетических реакциях, 99% образующейся энергии появляется за счёт расщепления углеводов, триглицеридов (жиров) и белков. Таким образом, именно депонирование стабильных форм данных типов химических соединений является необходимым условием для оптимального функционирования органов и систем в различных условиях.
Резерв углеводов в организме ограничен многими факторами и существенно варьируется у конкретного индивида в зависимости от режима и качества питания и нагрузок. Метаболизм сахаров в отсутствии выраженной физической активности и при достаточном поступлении их с пищей позволяет не испытывать необходимости в запасании. Ситуация меняется при появлении дефицита глюкозы как самой распространенной формы углеводного источника энергообеспечения. В таком случае появляется необходимость наличия стабильных источников запасённых сахаров.
Адаптированной формой долгосрочного хранения углеводов является гликоген — полисахарид, который образован остатками глюкозы, соединенными альфа ‒1,4 связями. Полимерная структура гликогена даёт возможность формировать резерв углеводов без повышения осмотического давления, которое бы имело место в случае использования для этих целей молекул глюкозы. Резерв гликогена присутствует в печени и мышечной ткани.
Полимерная структура гликогена даёт возможность формировать резерв углеводов без повышения осмотического давления, которое бы имело место в случае использования для этих целей молекул глюкозы. Резерв гликогена присутствует в печени и мышечной ткани.
Рисунок 1 | Строение гликогена
Гликоген хранится в мышечной ткани в виде гранул, расположенных в саркоплазме мышечной клетки, в количестве 14‒18 г/кг веса человека. В печени гликоген содержится в количестве 80‒120 г и может в любой момент быть высвобожден в кровоток для поддержания оптимальной концентрации глюкозы в крови.
Во время выполнения физической работы высокой интенсивности и длительной продолжительности запасы гликогена печени истощаются, обеспечивая нормальный уровень глюкозы крови; таким образом, глюкоза крови не является сама по себе депо углеводов, а лишь служит транспортным звеном в цепи энергообеспечения.
В мышечной ткани гликоген метаболизируется до глюкозы с последующим включением её в реситнез АТФ, либо используется самостоятельно как энергетический субстрат.
Основным запасаемым источником энергии человеческого организма являются липиды (жиры), хранящиеся в жировой ткани в виде триглицеридов. Ёмкость депо триглицеридов велика и у нетренированных людей может составлять значительную долю массы всего тела (у женщин до 30%). Количество жировой ткани у тренированных атлетов обоих полов значительно снижено (вплоть до 2‒3 %) и находится в корреляции от целого набора факторов, связанных с конкретными спортивными и задачами и стадиями тренировочного процесса. Триглицериды, мобилизованные их подкожной и висцеральной жировой ткани, расщепляются на свободные жирные кислоты (ЖК) с помощью ферментов — липаз — и с кровотоком попадают к работающей мышце.
Триглицериды также хранятся в самих мышечных клетках в количестве примерно 300 г. В ММВ (медленных или окислительных мышечных волокнах) потенциал для запасания жиров значительно больше, чем в БМВ (быстрых мышечных волокнах). У тренированных индивидов вследствие низкого содержания подкожного жира увеличивается количество внутримышечных липидов. Такие внутримышечные триглицериды могут обеспечить до 20% всех энергетических запасов и до 50% всех жиров, используемых в реакциях аэробного окислительного ресинтеза АТФ при продолжительной работе в средних значениях интенсивности. Использование липидов приобретает также существенное значение для биоэнергетических процессов в период после выполнения работы.
Такие внутримышечные триглицериды могут обеспечить до 20% всех энергетических запасов и до 50% всех жиров, используемых в реакциях аэробного окислительного ресинтеза АТФ при продолжительной работе в средних значениях интенсивности. Использование липидов приобретает также существенное значение для биоэнергетических процессов в период после выполнения работы.
Рисунок 2 | Жировая ткань
Белки практически не запасаются в человеческом организме в энергетически усвояемых формах, а концентрация свободных аминокислот имеет слишком низкие значения, чтобы считаться влиятельным фактором депонирования. Существуют данные об использовании в качестве энергетического субстрата продуктов распада белков слизистой оболочки кишечника. Указанные белки синтезируются со скоростью, превышающей скорость синтеза белков скелетных мышц в 30 раз, и могут использоваться при сильном дефиците иных соединений. Однако окончательной структурно выстроенной теории депонирования белков для использования их как источника энергии пока не существует.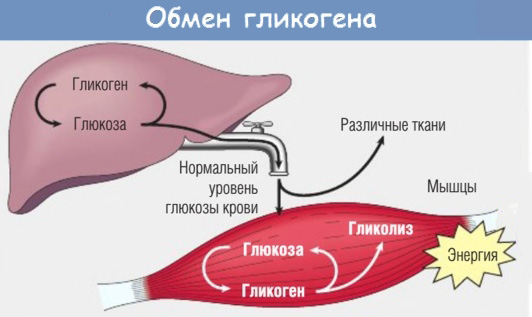
Ниже приведены данные о запасах энергетических субстратов у тренированного человека весом 70 кг, обеспеченного адекватным питанием, в фазе отдыха.
Глюкоза крови — 3‒5 г;
Гликоген печени — 80‒100 г;
Гликоген мышц — 300‒400 г;
Жировая ткань — 3‒20 кг;
Триглицериды мышц — 500 г.
Объективно известно, что депо энергетически важных веществ у людей разного пола, возраста и уровня тренированности могут быть выражены различными структурами, однако общая пластичность и адаптивность метаболизма человека позволяет пользоваться несколькими стабильными источниками энергообеспечения в зависимости от возникшей задачи.
Источники
- BIOCHEMISTRY — VOET.D, VOET .J.G., JOHN WILEY & SONS INC, 2011
- Михайлов С.С. Спортивная биохимия: Учебник для вузов и колледжей. – М.: Советский спорт, 2004
- Агаджанян Н.А., Власова И.Г., Ермакова Н.В. Торшин В.И. Основы физиологии человека: Учебник. – Изд.
 2-е, испр. – М.: Изд-во РУНД, 2004
2-е, испр. – М.: Изд-во РУНД, 2004
Гликоген: для чего он нужен?. Что такое гликоген? | by Efim Klinovsky
Что такое гликоген?
Гликоген — это одна из основных форм запасной энергии в организме человека. Структура представляет собой сотни связанных между собой молекул глюкозы.
В случае снижения уровня глюкозы в крови, тело начинает вырабатывать специальные ферменты, в результате чего, накопленный гликоген в мышцах начинает расщепляться на молекулы глюкозы, становясь источником энергии.
Важность углеводов для организма.
В процессе пищеварения, углеводы расщепляются в глюкозу, после чего она поступает в кровь.
Жиры и белки в глюкозу конвертироваться не могут!
Далее глюкоза используется телом для текущих энергозатрат, либо откладывается в резервные запасы — жир. При этом, организм сначала связывает глюкозу в молекулы гликогена и заполняет ими гликогеновые депо, как только они переполняются, тело преобразует избыток в жир.
Где накапливается гликоген?
Гликоген накапливается преимущественно в печени (~100–120г) и в мышечных тканях (~1% от общего веса мышц). Суммарно запасов гликогена в теле ~200–300г, однако, чем больше мышечной массы, тем больше накоплений (вплоть до 500г).
Гликоген в печени служит источником энергии для всего тела, а запасы гликогена в мышцах доступны исключительно для локального использования (у каждой группы мышц свой запас).
Функции гликогена в мышцах.
Биологически, гликоген накапливается не в мышечных волокнах, а в окружающей их жидкости — саркоплазме. Мышечная структура схожа с губкой, которая впитывает в себя саркоплазму и увеличивается в размерах.
Регулярные силовые тренировки увеличивают размер гликогеновых депо и количество саркоплазмы, визуально мышцы становятся больше и объемнее.
Число мышечных волокон задается прежде всего генетическим типом телосложения и практически не меняется в течение жизни.

Влияние гликогена на мышцы: Биохимия.
Для набора мышечной массы требуется два условия: 1.Достаточное количество запасов гликогена в мышцах ДО тренировки. 2.Успешное восстановление гликогеновых депо ПОСЛЕ тренировки. Выполняя силовые упражнения без запасов гликогена в надежде «просушиться», прежде всего тело сжигает мышечные ткани.
Важно! Для роста мышц не так важно употребление белка и аминокислот BCAA, как наличие достаточного количества правильных углеводов в рационе, в особенности, достаточное потребление быстрых углеводов по окончании тренировки.
Организм не сможет нарастить мышцы, находясь на безуглеводной диете.
Как повысить запасы гликогена?
Пополнить можно углеводами из продуктов питания, либо с помощью спортивного гейнера (смесь белков и углеводов). Чем ниже гликемический индекс (ГИ — скорость усваивания организмом углеводов и повышения сахара в крови), тем медленнее углеводы отдают свою энергию в кровь и тем выше вероятность конвертации в гликогеновые депо.
Влияние гликогена на сжигание жира.
Для эффективного избавления от подкожного жира нужно помнить, что первым делом тело расходует запасы гликогена, а лишь затем переходит к запасам жира. Эффективнее всего для сжигания жира будет тренировка на пустой желудок, тренировка должна быть не менее 40 минут с поддержанием умеренного пульса (кратковременный отдых, только для восстановления дыхания).
Лучше всего взять в привычку бег на дистанцию не менее двух миль на пустой желудок.
Гликоген — что это и где запасается? Функции для работы мышц
Гликоген — это накапливаемый в мышцах (и в печени) резерв углеводов, используемый в качестве первичного источника энергии при физических тренировках. Источником гликогена являются употребляемые с пищей (или со спортивными напитками) углеводы.
По сути, чем больше человек занимается спортом, тем эффективнее его организм запасает углеводы в мышцах в виде гликогена — тогда как при малоподвижном образе жизни они отправляются в жир.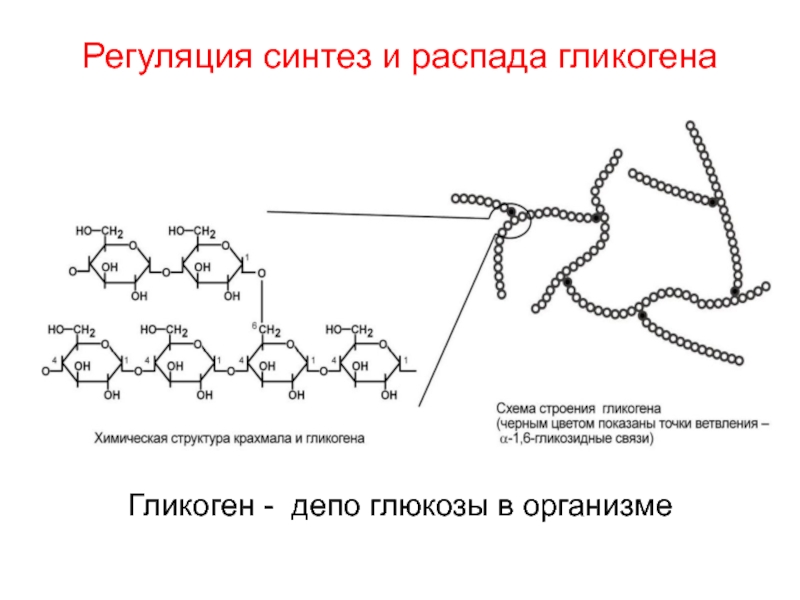 Кроме этого, сжигание жира также достигается после опустошения гликогеновых депо.
Кроме этого, сжигание жира также достигается после опустошения гликогеновых депо.
// Гликоген — что это?
Гликоген — это тип углеводов, накапливаемый в организме человека. Вещество иногда называют «животным крахмалом», поскольку по своей структуре гликоген похож на обычный крахмал и состоит из сотен и тысяч связанных между собой молекул глюкозы.
Источником для гликогена являются углеводы из продуктов питания. Напомним, что в чистом виде организм не может хранить глюкозу — ее высокое содержание в клетках создает гипертоническую среду, приводя к притоку воды и развитию сахарного диабета. В свою очередь, гликоген не растворим в воде.
После того, как уровень глюкозы в крови снижается (например, через несколько часов после приема пищи или при физических тренировках), организм начинает расщеплять накопленный в мышах гликоген до глюкозы, становясь источником для быстрой энергии.
// Функции гликогена:
- продукт пищеварения углеводов
- главное топливо для работы мышц
- источник быстрой энергии для организма
// Читать дальше:
Гликоген и гликемический индекс еды
В процессе пищеварения углеводы из продуктов питания расщепляются до глюкозы (жиры и белки в нее конвертироваться не могут) — после чего она попадает в кровь.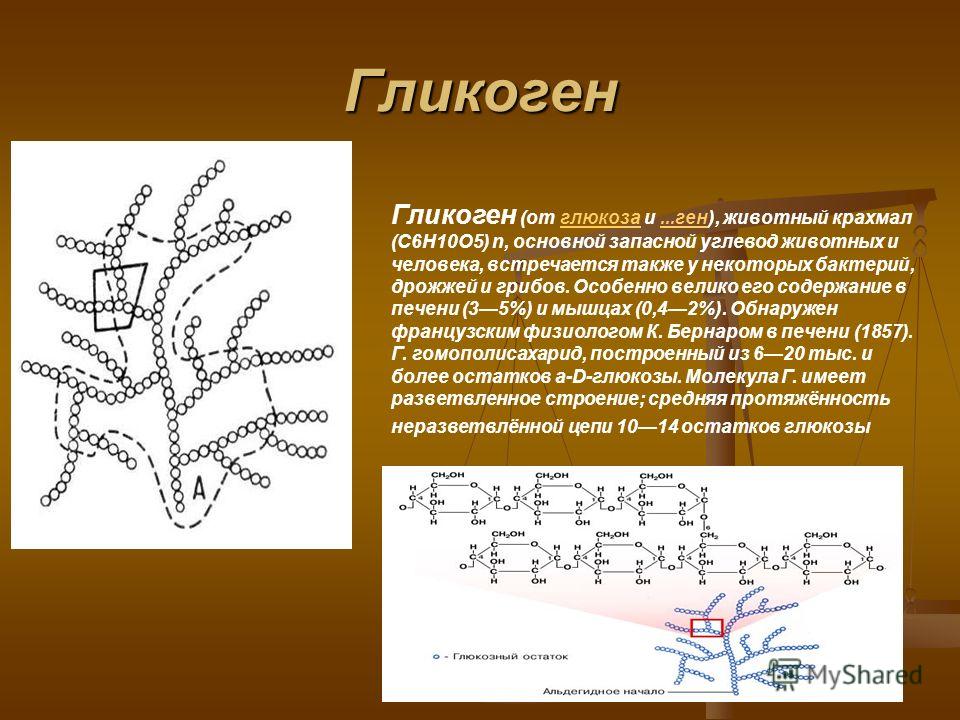 Глюкоза может быть использована телом либо для текущих нужд метаболизма, либо быть преобразованной в гликоген — или в жир.
Глюкоза может быть использована телом либо для текущих нужд метаболизма, либо быть преобразованной в гликоген — или в жир.
Чем ниже гликемический индекс пищи, тем лучше содержащиеся в ней углеводы конвертируются в гликоген. Несмотря на то, что простые углеводы максимально быстро повышают уровень глюкозы в крови, значительная их часть конвертируется в жировые запасы.
В свою очередь, энергия сложных углеводов, получаемся организмом постепенно, более полно конвертируется в гликоген, содержащийся в мышцах. Именно поэтому диета для набора сухой массы подразумевает употребление углеводов с низким и средним ГИ.
// Читать дальше:
Где накапливается гликоген?
В организме гликоген накапливается преимущественно в печени (порядка 100-120 г) и в мышечной ткани (от 200 до 600 г)¹. Считается, что примерно 1% от общего веса мышц приходится именно на это вещество. Неспортивный человек может иметь запасы гликогена в 200-300 г, мускулистый спортсмен — до 600 г.
Также важно, что если запасы гликогена в печени используются для покрытия энергетических потребностей в глюкозе по всему телу, тогда как запасы гликогена в мышцах доступны исключительно для локального потребления. Говоря простыми словами, во время приседаний тело использует депо мышц ног, а не бицепса.
Функции гликогена в мышцах
Говоря более точно, гликоген накапливается не в самих мышечных волокнах, а в саркоплазме — окружающей их питательной жидкости. Рост мускулатуры связан с увеличением объема именно этой питательной жидкости — по своей структуре мышцы похожи на губку, впитывающей саркоплазму для увеличения размера.
Регулярные силовые тренировки положительно влияют на размер гликогеновых депо и количество саркоплазмы, делая мышцы визуально более большими и объемными. При этом число мышечных волокон задается прежде всего типом телосложения и практически не меняется в течение жизни человека вне зависимости от тренировок — меняется лишь способность организма накапливать больше гликогена.
// Читать дальше:
Гликоген в печени
Печень — это главный фильтрующий орган организма. В том числе, он перерабатывает поступающие с пищей углеводы — однако за раз печень способна переработать не более 100 г глюкозы. В случае хронического избытка быстрых углеводов в питании, эта цифра повышается.
В результате клетки печени могут превращать сахар в жирные кислоты. В этом случае исключается стадия гликогена, и начинается жировое перерождение печени.
Влияние на мышцы — биохимия
Успешная тренировка для набора мускулатуры требует двух условий — во-первых, наличия достаточного содержания запасов гликогена в мышцах до тренировки, а, во-вторых, успешное восстановление гликогеновых депо по ее окончанию.
Выполняя силовые упражнения без запасов гликогена (или без подпитки аминокислотами BCAA) в надежде «просушиться», вы вынуждаете тело сжигать мышцы. Для роста мышц важно не столько употребление белка, сколько наличие в рационе существенного количества углеводов.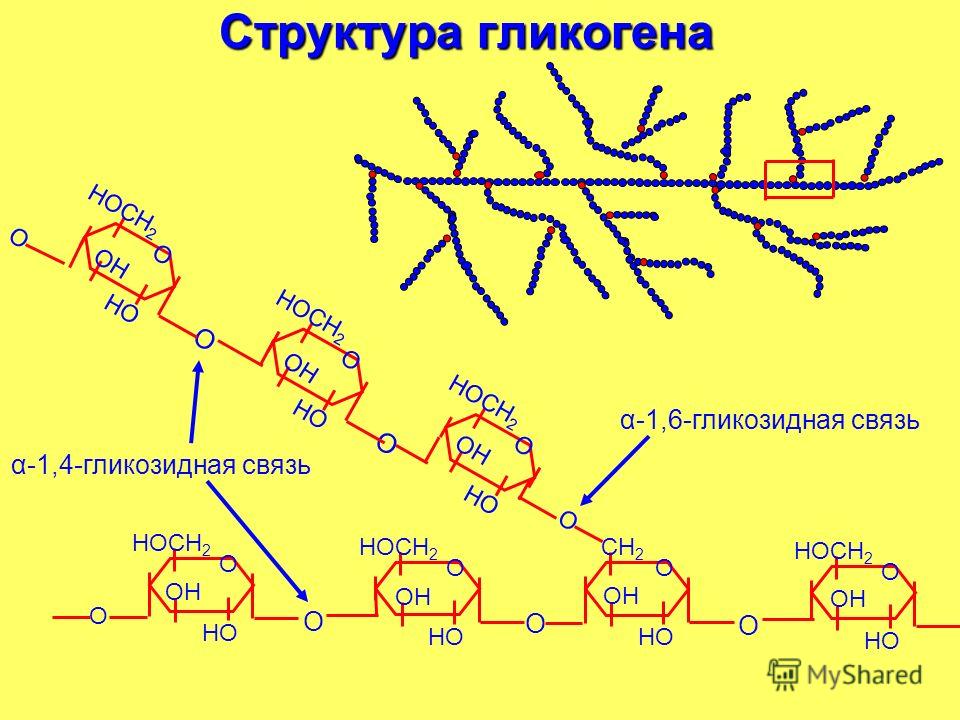
В особенности, достаточное потребление углеводов сразу по окончанию тренировки в период “углеводного окна” — это нужно для восполнения запасов гликогена и остановки катаболических процессов. В противоположность этому, на безуглеводной диете нарастить мышцы нельзя.
// Читать дальше:
Как повысить запасы гликогена?
Запасы гликогена в мышцах пополняются либо углеводами из продуктов питания, либо употреблением спортивного гейнера (смеси протеина и углеводов в виде мальтодекстрина). Как мы уже упоминали выше, в процессе пищеварения сложные углеводы расщепляются до простых; сперва они попадают в кровь в виде глюкозы, а затем переработаются организмом до гликогена.
Чем ниже гликемический индекс конкретного углевода, тем медленнее он отдает свою энергию в кровь и тем выше его процент конвертации именно в гликогеновые депо, а не в подкожную жировую клетчатку. Особенную важность это правило имеет в вечернее время — к сожалению, простые углеводы, съеденные за ужином, пойдут прежде всего в жир на животе.
// Что повышает содержание гликогена в мышцах:
- Регулярные силовые тренировки
- Употребление углеводов с низким гликемическим индексом
- Прием гейнера после тренировки
- Восстанавливающий массаж мышц
Влияние на сжигание жира
Если вы хотите сжечь жир с помощью тренировок, помните о том, что тело сперва расходует запасы гликогена, а лишь затем переходит к запасам жира. Именно на этом факте и строится рекомендация о том, что эффективная жиросжигающая тренировка должна проводиться не менее 40-45 минут при умеренном пульсе — сперва организм тратит гликоген, затем переходит на жир.
Практика показывает, что жир быстрее всего сгорает при кардиотренировках утром на пустой желудок или использовании интервального голодания. Поскольку в этих случаях уровень глюкозы в крови уже находится на минимальном уровне, с первых минут тренинга тратятся запасы гликогена из мышц (а затем и жира), а вовсе не энергия глюкозы из крови.
***
Гликоген является основной формой хранения энергии глюкозы в животных клетках (в растениях гликогена нет). В теле взрослого человека накапливается примерно 200-300 г гликогена, запасаемого преимущественно в печени и в мышцах. Гликоген тратится при силовых и кардиотренировках, а для роста мышц чрезвычайно важно правильно восполнять его запасы.
Научные источники:
- Fundamentals of glycogen metabolism for coaches and athletes, source
В продолжение темы
Дата последнего обновления материала — 8 июня 2020
Гликоген и жир — рассуждения дилетанта
Много-много-много раз я слышал от людей, приверженных фитнесу, что после сна гликогеновые депо истощены. Потому что сон это 8 часов голодания. На мой взгляд это херня и вот почему: гликогеновые депо, хранилище высокопотенциального и легкодоступного топлива — углеводов. Запасается его не так много, ориентировочно у взрослых людей от 400 до 800 грамм во всём теле или 1600-3200 ккал. Гликоген позволяет высвобождать химическую энергию с очень большой скоростью, т.е. это топливо, предназначенное для развития больших мощностей.
Гликоген позволяет высвобождать химическую энергию с очень большой скоростью, т.е. это топливо, предназначенное для развития больших мощностей.
Причём расходуется гликоген как аэробно, так и анаэробно. Аэробно во время довольно интенсивных аэробных нагрузок, когда жиры уже не позволяют развивать заданные мощности. Анаэробно, когда развиваются пиковые усилия: это спринт и поднятие тяжестей. При этом при использовании гликогена анаэробно выделяется всего лишь 7% запасённой в нём химической энергии, остальная энергия остаётся в молекулах продуктов распада и становится на некоторое время недоступной для использования. Аэробно с соревновательными усилиями, если не кушать углеводы по ходу, гликогена обычно хватает примерно на два-три часа (бега, велосипеда, лыж и т.п.). С помощью спринтов можно остаться полностью закисленным и без гликогена буквально за 10 минут.
Ещё один интересный факт о кликогене, он хранится в сугубо связанном с водой состоянии. «Сироп» с калорийностью 1-1.3 ккал/г (при калорийности углеводов 4 ккал/г). Так что масса топлива при полностью заряженных гликогеновых депо может достигать 1.6-2 кг у небольших и нетренированных людей и до 3-4 кг у крупных людей или людей с хорошо тренированных.
Так что масса топлива при полностью заряженных гликогеновых депо может достигать 1.6-2 кг у небольших и нетренированных людей и до 3-4 кг у крупных людей или людей с хорошо тренированных.
В общем гликоген это краткосрочное хранилище энергии, которое позволяет развивать максимальные усилия в той или иной области деятельности, стратегический запас. А вот в случае невысокой скорости расходования энергии (мощности): покой, сон, сидячая работа, ходьба, очень медленный бег, энергия добывается преимущественно из жиров. Из жировых депо, да. И калорийность жирового топлива существенно выше: 7.7 ккал/г и хранится у обычных людей топлива столько, что хватит на многие дни, недели (И ДАЖЕ МЕСЯЦЫ) обеспечения жизнедеятельности. Жиры — основной источник топлива.
Итак, по какой причине во сне, когда требуется порядка 1 ккал на 1 килограмм массы тела в час, будет расходоваться гликоген, который может выделять энергию с темпом в 15-20 раз больше? Наоборот, если вы хорошо поужинали, скушали адекватное количество [преимущественно медленных] углеводов, во сне гликогеновые депо будут восполняться, если они по какой-то причине опустошены (например, после интенсивной беговой или силовой тренировок).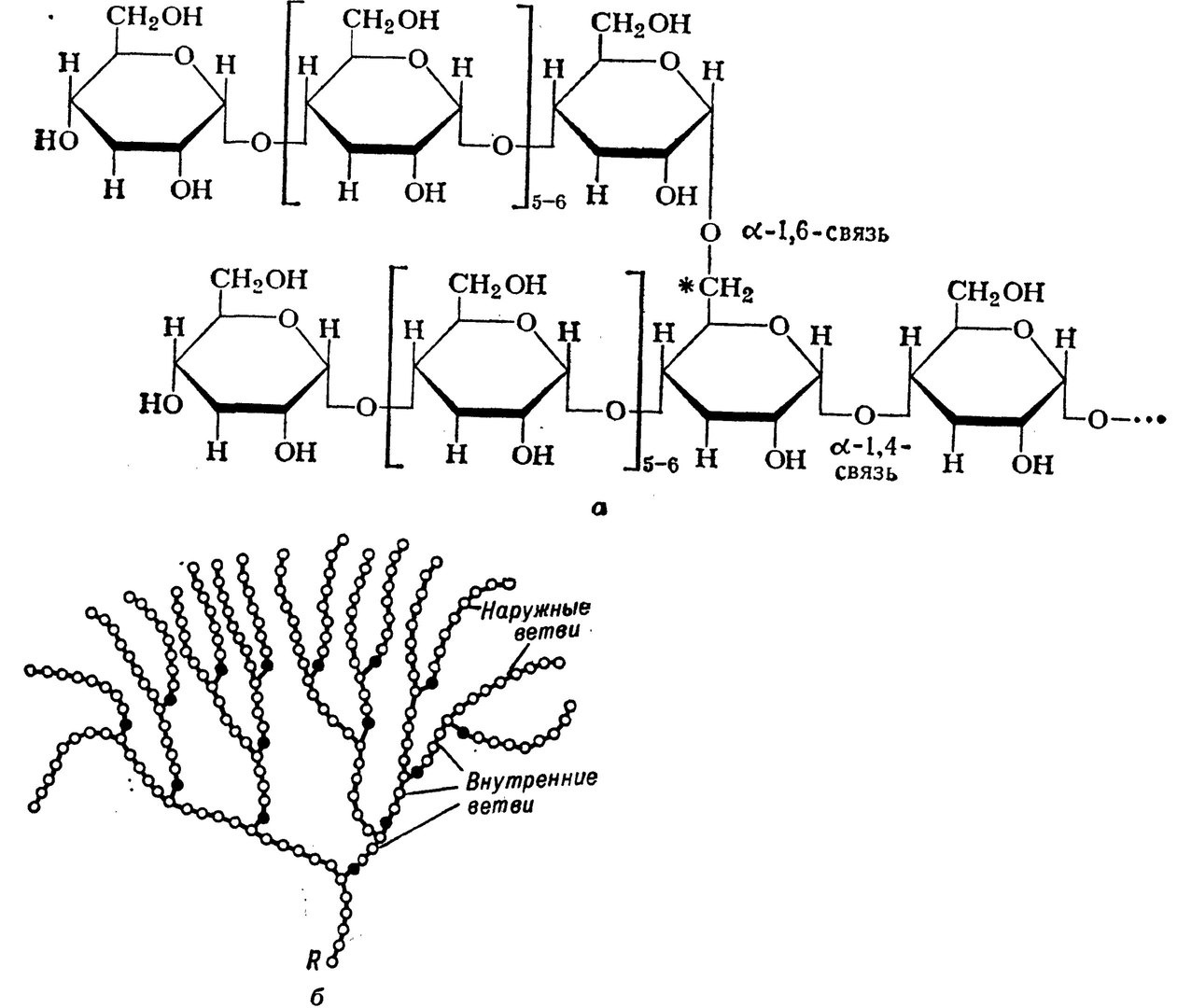 И это действительно так, я тренируюсь по 2-3 раза в день и выбираю гликогеновые депо довольно глубоко, так что хорошо понимаю о чём говорю 🙂
И это действительно так, я тренируюсь по 2-3 раза в день и выбираю гликогеновые депо довольно глубоко, так что хорошо понимаю о чём говорю 🙂
Единственная причина, по которой может расходоваться гликоген во сне — необходимость поставлять глюкозу мозгу (ну если вы не ели после шести и легли спать сугубо-голодными или по какой-то дурости избегаете потребления углеводов в принципе). Ну так мозг потребляет ~20% основного обмена веществ. Это 0.2 ккал на 1 кг массы тела в час, или 14 ккал в час для 70 кг человека. Или 28 грамм углеводов за 8 часов соннного голодания. В сравнении даже с 400 граммами максимальной ёмкости — капля в море.
Ещё одно упражнение: сколько углеводов может уместиться крови у человека? Молярная масса глюкозы 180.2 г/моль. В норме после еды это 7.7 ммоль/литр максимум (натощак в норме 3.5-5.5). Примем средний объём крови в 5 литров. Тогда получается 5 л * 180.2 г/моль * 0.0077 моль/литр = 6.9 грамм сахара. Нижняя граница нормы натощак — половина этого.
Отсюда, кстати, следует причина, по которой «быстрые» углеводы легко пополняют хранилища энергии, преимущественно жировые. Если вы сожрали 50 грамм рафинированного сахара за несколько минут (что легко), усваивается он стремительно, тоже по порядку величины это 10 минут. А в покое вам нужен эквивалент 13-18 грамм в час. В кровь больше семи грамм не запихаешь. Куда девать остальное богатство? Куда-то, если повезёт, то пополянтся гликоеновые депо, но они не слишком быстро перезаряжаются. Да и далеко не всегда в них есть место… А вот жировые всегда рады принять излишек, места там полно, поджелудочная инсулин выбрасывает добросовестно (до поры до времени). И оттуда эти калории доставать уже совсем не легко, т.к. высокопотенциальные угли превратились в низкопотенциальный жир — топливо покоя.
Если вы сожрали 50 грамм рафинированного сахара за несколько минут (что легко), усваивается он стремительно, тоже по порядку величины это 10 минут. А в покое вам нужен эквивалент 13-18 грамм в час. В кровь больше семи грамм не запихаешь. Куда девать остальное богатство? Куда-то, если повезёт, то пополянтся гликоеновые депо, но они не слишком быстро перезаряжаются. Да и далеко не всегда в них есть место… А вот жировые всегда рады принять излишек, места там полно, поджелудочная инсулин выбрасывает добросовестно (до поры до времени). И оттуда эти калории доставать уже совсем не легко, т.к. высокопотенциальные угли превратились в низкопотенциальный жир — топливо покоя.
И ещё одно соображение, в связи с тем, почему у людей от высокоинтенсивной тренировки и/или низкоуглеводных диет возникает иллюзия быстрой потери веса, особенно в начале. Многие говорят, что интенсивные упражнения «сушат». Дело опять же в том, что они действительно сушат, т.к. расходуется гликоген а, в месте с ним, выходит значительный объём воды.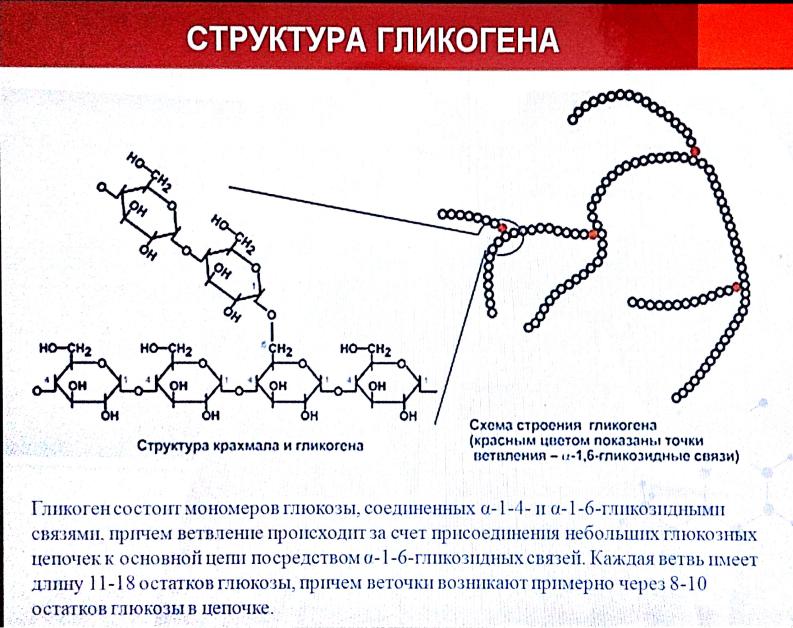 Но это не долгосрочная потеря веса, у неистощённого человека гликогеновые должны быть где-то на максимуме и в любом случае больше 2-3-4 кг ими не потерять. Так же имеет место иллюзия быстрого набора веса при отмене низкоуглеводной/низкокалорийной диеты, когда на фоне долгой нехватки углеводов возникает явление гиперкоменсации и тело может запасти до двух раз больше гликогена, чем в норме. Во сколько это может вылиться килограмм, см. выше.
Но это не долгосрочная потеря веса, у неистощённого человека гликогеновые должны быть где-то на максимуме и в любом случае больше 2-3-4 кг ими не потерять. Так же имеет место иллюзия быстрого набора веса при отмене низкоуглеводной/низкокалорийной диеты, когда на фоне долгой нехватки углеводов возникает явление гиперкоменсации и тело может запасти до двух раз больше гликогена, чем в норме. Во сколько это может вылиться килограмм, см. выше.
А при похудении, конечно же, необходимо истощать жировые депо, в которых и хранятся основные десятки и даже сотни тысяч килокалорий. А, в силу высокой плотности содержания энергии (7700 ккал на килограмм) процесс этот безусловно небыстрый. И да, чудес не бывает. Чтобы облегчить жировые депо на 2 кг за месяц, нужно за этот месяц недоесть 15400 ккал или создать дефицит в 513 ккал в день. И ничего другого. Суммарная масса тела может изображать вообще странные финты, но, что касается жира, тут всё работает с математической точностью, в особнности конечно на достаточно долгом интервале времени. Никаких чудес нет. Масса и энергия сохраняются. И точка 🙂
Никаких чудес нет. Масса и энергия сохраняются. И точка 🙂
И вот ещё один фитнесс-психоз, по поводу разоблачения «зоны жирожигания». Она действительно существует. При невысоких интенсивностях львиная доля энергетических потребностей прямо в процессе выполнения упражнения (ходьба, лёгкая трусца, прогулочный темп на велосипеде) обеспечивается за счёт жиров. Естественно килокалорий потратится немного, но это будут преимущественно калории из жиров. Высокоинтенсивная тренировка задействует углеводы и тем бОльшую их долю, чем интенсивнее работа. Естественно, калорий может тратиться в два, а то и в три раза больше за одно и то же время. НО. У нетренированных или плохо тренированных людей имеет место тенденция к полному подавлению окисления жиров уже на очень умеренных интенсивностях выполнения упражнения. Т.е. при выходе по пульсу за коридор пресловутой зоны жиросжигания.
Так-то в общем-то ничего плохого, но штука в том, что пустые гликогеновые депо и низкий уровень сахара в крови делают (нередко) человека маниаком, ищущим углеводов. Вспомним, что огромная масса современных людей плотно сидит на сладком, в особенности худеющая часть, можете быть уверены, что при углеводном голоде удежать дефицит калорий и не сорваться в зажор очень и очень тяжело. А вот потеря энергии из жировых депо, проходит довольно «незаметно» для мозга, он в жире не слишком заинтересован, так что потратить за час 300 ккал ходьбой или 700-750 ккал быстрым бегом, ещё неизвестно что лучше в смысле похудения. Пользоваться ли зоной жиросжигания или не пользоваться, это уже дело десятое, но как понятие она вполне оправдана. Ну и если уж поднимать интенсивность, то надо подкладывать в топку быстрые углеводы, так чтобы держать гликогеновые депо на максимуме, а уж что из жировых сгорит, то сгорит.
Вспомним, что огромная масса современных людей плотно сидит на сладком, в особенности худеющая часть, можете быть уверены, что при углеводном голоде удежать дефицит калорий и не сорваться в зажор очень и очень тяжело. А вот потеря энергии из жировых депо, проходит довольно «незаметно» для мозга, он в жире не слишком заинтересован, так что потратить за час 300 ккал ходьбой или 700-750 ккал быстрым бегом, ещё неизвестно что лучше в смысле похудения. Пользоваться ли зоной жиросжигания или не пользоваться, это уже дело десятое, но как понятие она вполне оправдана. Ну и если уж поднимать интенсивность, то надо подкладывать в топку быстрые углеводы, так чтобы держать гликогеновые депо на максимуме, а уж что из жировых сгорит, то сгорит.
Но вообще основное жировое похудение конечно происходит в покое. На спорт мы получаем адекватное количество углеводов, на восстановление адекватное количество белков и незаменимых жиров, а на всё остальное медленные «пустые калории» поступают из жировых депо, если имеет место превышение общего расхода над общим приходом.
И вот ещё одна памятка: 1 кал = 4.187 Дж, 1 ккал/ч = 1.163 Вт.
И ещё, если велосипедист развивает целый час на педалях 250 Вт, то при типичном для такого уровня велосипедном КПД=23% он потратит за час 935 ккал. Это приблизительный уровень первого разряда по велоспорту.
Гликоген
Дано определение основному углеводу организма человека – гликогену. Описан синтез и распад гликогена в печени и скелетных мышцах. Приводится эмпирическая формула гликогена.
Гликоген
Гликоген является одним из основных углеводов, типичным для человека и животных.
Определение
Гликоген – полисахарид, состоящий из большого количества (до n=30000) остатков глюкозы (рис. 1).
Эмпирическая формула гликогена – (С6Н10О5)n, где: С6Н10О5 – остаток глюкозы, n — количество остатков глюкозы.
Где содержится в организме человека
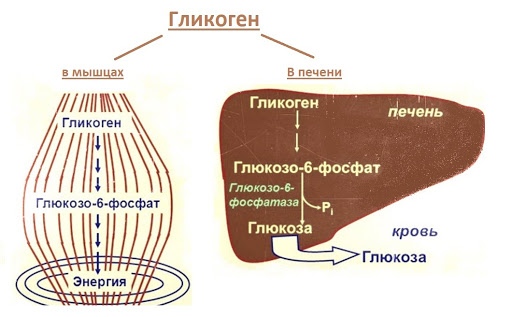
В организме человека содержится около 450 г гликогена. Треть этого количества (то есть около150 г) накапливается в печени, остальные две трети (около 300 г) накапливается в мышцах (Я. Кольман, К.-Г. Рём, 2004), рис. 2. Другими словами в печени содержится 5-6% от массы печени, в мышцах — 2-3% от массы мышц. Содержание гликогена в других органах незначительно. Гликоген печени служит прежде всего для поддержания уровня глюкозы в крови. Гликоген мышц служит резервом энергии и не участвует регуляции уровня глюкозы в крови.
Треть этого количества (то есть около150 г) накапливается в печени, остальные две трети (около 300 г) накапливается в мышцах (Я. Кольман, К.-Г. Рём, 2004), рис. 2. Другими словами в печени содержится 5-6% от массы печени, в мышцах — 2-3% от массы мышц. Содержание гликогена в других органах незначительно. Гликоген печени служит прежде всего для поддержания уровня глюкозы в крови. Гликоген мышц служит резервом энергии и не участвует регуляции уровня глюкозы в крови.
Рис. 2. Баланс гликогена в организме человека (Я. Кольман, К.-Г. Рём, 2004)
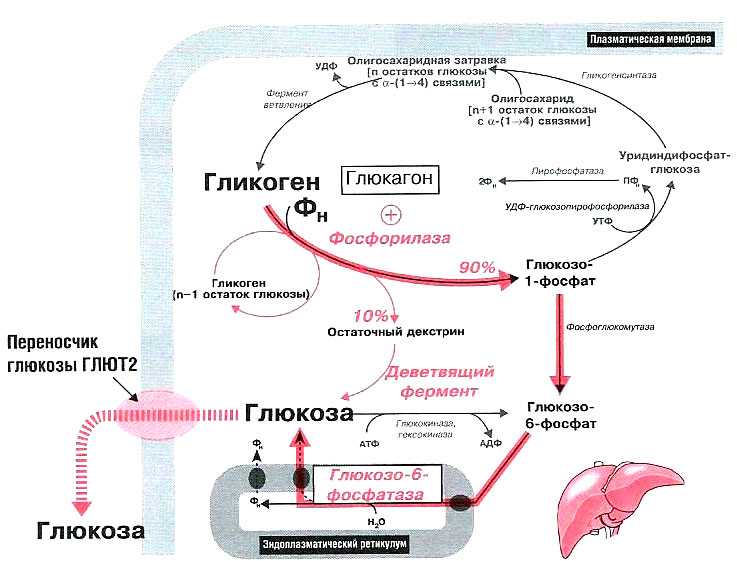
Синтез гликогена
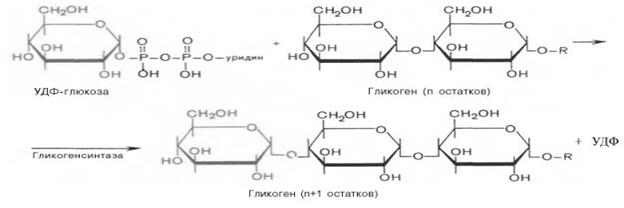
Гликоген синтезируется в печени и мышцах из глюкозы, поступающей по кровеносным сосудам. Собственно, в печени гликоген представляет собой запасную, резервную форму глюкозы или депо глюкозы.
Свободная глюкоза не может накапливаться в печени и мышцах. Это связано с тем, что молекулы глюкозы имеют малые размеры и легко проходят через внешнюю оболочку клеток печени (гепатоцитов) и через сарколемму мышечных волокон (С. С. Михайлов, 2009). Синтез гликогена требует затрат энергии. Для присоединения к гликогену одного остатка глюкозы необходимо 41 кДж энергии. Синтез гликогена усиливает гормон инсулин.
С. Михайлов, 2009). Синтез гликогена требует затрат энергии. Для присоединения к гликогену одного остатка глюкозы необходимо 41 кДж энергии. Синтез гликогена усиливает гормон инсулин.
О взаимосвязи гормонов и мышечной массы можно прочесть в моей книге «Гормоны и гипертрофия скелетных мышц человека»
Распад гликогена
В печени распад (лизис) гликогена называется гликогенолизом. Так как в гликоген печени распадается на глюкозу, этот процесс называется глюкогенезом. Он ускоряется гормонами глюкагоном, адреналином и норадреналином. При мышечной деятельности скорость мобилизации гликогена в печени зависит от интенсивности выполненной нагрузки. Так, например, при умеренной физической нагрузке скорость мобилизации гликогена возрастает в 2-3 раза, а при интенсивной – в 7-10 раз по сравнению с состоянием покоя.
Распад гликогена в печени происходит и во время отдыха. В результате этого образующаяся глюкоза способствует восстановлению запасов гликогена в сердечной мышце и скелетных мышцах (Н. И. Волков с соавт., 2000).
И. Волков с соавт., 2000).
В мышцах гликоген обычно распадается при выполнении физической нагрузки. Распад гликогена стимулирует гормон адреналин. Если распад гликогена происходит в анаэробных условиях, этот процесс называется гликолизом.
Литература
- Кольман Я., Рём К.-Г. Наглядная биохимия.- М.: Мир, 2004.- 469 с.
- Мак-Комас, А. Дж. Скелетные мышцы. – Киев: Олимпийская литература, 2001.- 407 с.
- Михайлов, С. С. Спортивная биохимия. – М.: Советский спорт, 2009.– 348 с.
С уважением, А.В. Самсонова
|
По мере […] волокнах, они начинают рассчитывать на жиры. herbalifenutritioninstitute.com |
As the glycogen levels drop in the red muscle […]
fibers, they will rely more on fat. herbalifenutritioninstitute. |
|
Снижение уровней мышечного гликогена приводит к переутомлению […]
и к накоплению лактата в мышцах. herbalifenutritioninstitute.com |
Depletion of muscle glycogen causes fatigue and is […]
associated with a build-up of muscle lactate. herbalifenutritioninstitute.com |
|
Во время […] herbalifenutritioninstitute.com |
During exercise lasting for more than 20 to 30 minutes, blood glucose becomes important as a fuel to spare muscle glycogen breakdown.
herbalifenutritioninstitute.com |
Глютамин играет важную роль в ряде обменных процессов, включая межорганный транспорт азота, синтез почечного аммиака, гепатический глюконеогенез и синтез гликогена в печени.
herbalifenutritioninstitute.com |
Glutamine plays an essential role in a number of metabolic processes, including interorgan transfer of nitrogen, renal ammonia synthesis, hepatic gluconeogenesis and hepatic glycogen synthesis.
herbalifenutritioninstitute.com |
|
Ксилитол указывает […] укрепление ATP, рост освобождания и потребляния […] и антикатаболические, антиоксдацийные […] эффекты разрешая на уничтожание усталости мышцев. ksylitol.com |
Xylitol shows strengthening ATP, the increase […] of freeing and using up fat, the […] catabolic, and at last antioxidizing […] effects, allowing one to fight the feeling of the cardiac muscle’s tiredness. ksylitol.com |
|
И аэробные и силовые тренировки ведут […] ферментов, а также к увеличению […] количества и объема митохондрий. herbalifenutritioninstitute.com |
Both aerobic and endurance training lead to increases in glycogen stores, […] and size of mitochondria. herbalifenutritioninstitute.com |
|
Так как во время тяжелых […] herbalifenutritioninstitute.com |
Since during heavy exercise fat is a primary fuel, this is taken to enhance fat utilization and sparing of glycogen stores.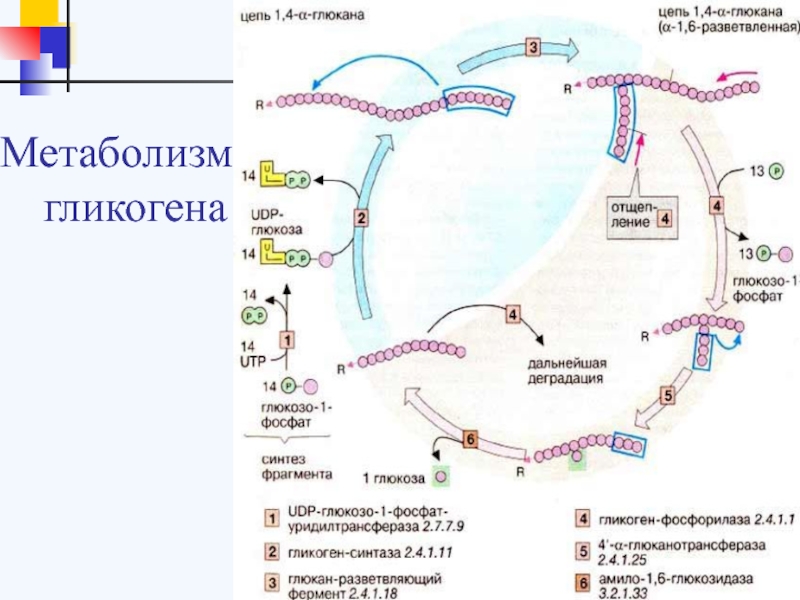
herbalifenutritioninstitute.com |
|
Пророщенные зерна пшеницы, богатые жизненной энергией молодых ростков, мед, масла кедрового ореха и зародышей пшеницы обеспечивают антиоксидантную защиту и […] насыщают организм энергией, создавая […] для дальнейшего использования в качестве […] источника энергии. bioline247.ru |
Germinated wheat, rich vitality of young sprouts, honey, cedar oil and wheat germ provide antioxidant protection and saturate […] the body with energy, creating carbohydrate depot as glycogen in […] bioline247.com |
|
Фосфатные соли у спортсменов […] также предназначены для улучшения процесса утилизации [. фосфорилирование. herbalifenutritioninstitute.com |
Phosphate salts in the athlete are also meant to enhance glucose utilization for […] herbalifenutritioninstitute.com |
|
По этой причине тренируемый спортсмен при длительных […] тренировках умеренной интенсивности будет сжигать жиры, но все же захочет […] herbalifenutritioninstitute.com |
Therefore, the trained athlete will […] burn fat with long-term moderate-intensity exercise, but will also […] herbalifenutritioninstitute. |
|
Прежде всего энергией снабжаются мышцы и органы в зависимости от того, что им необходимо в данный момент, затем […] пополняется уровень сахара в крови, […] энергии преобразуется в жир. waiworld.com |
First the muscles and organs are supplied with the energy they need at that moment, then the blood sugar […] level is replenished, next the glycogen depots are filled, and […] waiworld.com |
|
Короткие серии тренировок высокой интенсивности сжигают в первую очередь углеводы и требуют больших запасов гликогена в мышцах.
herbalifenutritioninstitute.com |
Short bursts of high-intensity exercise burn primarily carbohydrates and require large stores of glycogen in the muscle.
herbalifenutritioninstitute.com |
|
Тренировка […] herbalifenutritioninstitute.com |
Training causes an increase in the mitochondrial capacity for fat oxidation, which spares glycogen utilization.
herbalifenutritioninstitute.com |
|
Витамин B3, очень важен в процессе углеводного метаболизма и белкового, для клеточного дыхания, способоствует нормализации функции нервной системы, некоторых желез внутренней секреции, стимулирует эритропоез, снижает […] уровень шлюкозы в крови, […] холестерола в крови, […] улучшает поступление железа. hyllan.ro |
Vitamin B3, plays a role in the metabolism of carbohydrates, proteins, in cell respiration, contributes to the normalization of the nervous system function, of some glands of internal secretion, stimulates erythropoiesis, lowers the level of glucose in blood, contributes to the increase of [. the reserve of glycogen in the liver, […] encourages iron supply. hyllan.ro |
|
Для спортсмена, […] herbalifenutritioninstitute.com |
For a performance athlete, it is important to maintain the efficiency edge provided by carbohydrate as long as glycogen is available in the muscles.
herbalifenutritioninstitute.com |
|
Мышечный гликоген является наиболее предпочтительным […]
углеводным топливом во время тренировок, длящихся меньше двух часов, […] как при аэробном, так и при анаэробном метаболизме. herbalifenutritioninstitute. |
Muscle glycogen is the preferred carbohydrate […]
fuel for events lasting less than two hours for both aerobic and anaerobic metabolism. herbalifenutritioninstitute.com |
|
Гликоген печени может быть использована для различных целей, но мышечный гликоген может быть использован только для мышечной деятельности.
waiworld.com |
Glycogen from the liver can be used for different purposes, but that from muscles can only be used for muscle activity.
waiworld.com |
|
Главным преимуществом менее […] результативной анаэробной схемы является то, что она более быстро снабжает […] мышцах. herbalifenutritioninstitute. |
The major advantage of the less efficient anaerobic pathway is that it more rapidly provides ATP […] herbalifenutritioninstitute.com |
|
При наличии у человека сахарного диабета его печень и мышцы уже не в состоянии превращать поступающий с пищей сахар в гликоген.
bioline247.ru |
In the presence of human diabetes and had liver and muscles are no longer able to convert sugar from food into glycogen.
bioline247.com |
|
Для того, чтобы держаться на этом пике и избегать ненужной секреции инсулина (дополнительный […] инсулин нужен, когда глюкоза должна […] только пополнять уровень сахара в […] крови в случае необходимости. waiworld.com |
In order to keep near this peak and avoid unnecessary insulin secretion (extra insulin is needed [. when glucose needs to be converted into […] the blood sugar when needed. waiworld.com |
|
Гликоген может быть преобразован […]
снова в глюкозу, когда это необходимо, например, когда мышцы максимально активны. waiworld.com |
Glycogen can be reconverted into […]
glucose whenever needed, for instance when muscles are maximally active. waiworld.com |
 com
com
 ..]
..] com
com ..]
..]  com
com com
com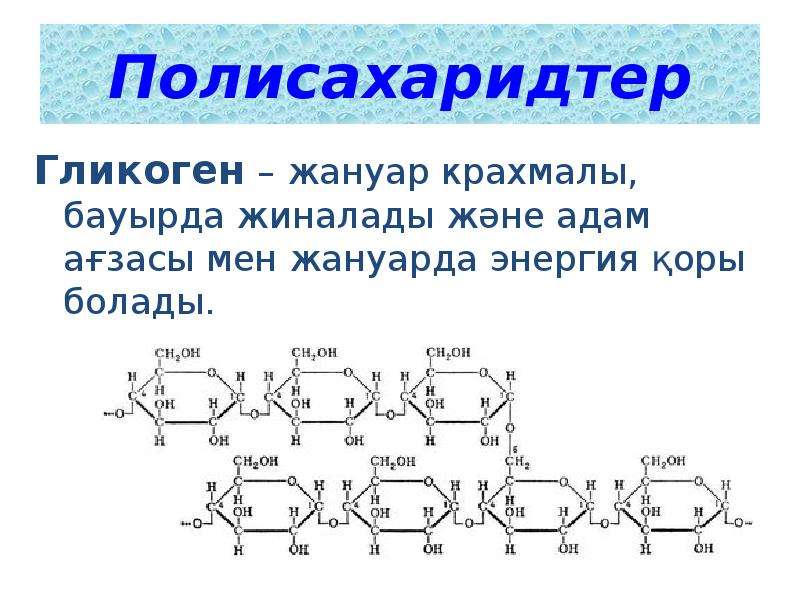 ..]
..] Метаболизм гликогена в организме человека — ScienceDirect
Основные моменты
- •
-
Гликоген — это разветвленный полимер глюкозы, который позволяет хранить глюкозу в организме человека.
- •
-
Сборка и распад гликогена — это сложные процессы, для которых требуется несколько ферментов.
- •
-
Дефицит ферментов, участвующих в метаболизме гликогена, вызывает различные гликогенозы.
- •
-
Эффективный метаболизм гликогена важен во время голодания и сокращения мышц.
- •
-
Помимо использования в качестве топлива, глюкоза, полученная из гликогена, выполняет и другие функции.
Реферат
В организме человека гликоген представляет собой разветвленный полимер глюкозы, хранящийся в основном в печени и скелетных мышцах, который поставляет глюкозу в кровоток во время голодания и в мышечные клетки во время сокращения мышц. Гликоген был обнаружен в других тканях, таких как мозг, сердце, почки, жировая ткань и эритроциты, но функция гликогена в этих тканях в основном неизвестна.Синтез гликогена требует ряда реакций, которые включают поступление глюкозы в клетку через переносчики, фосфорилирование глюкозы до глюкозо-6-фосфата, изомеризацию до глюкозо-1-фосфата и образование уридин-5ʹ-дифосфат-глюкозы, которая является прямым донором глюкозы для синтез гликогена.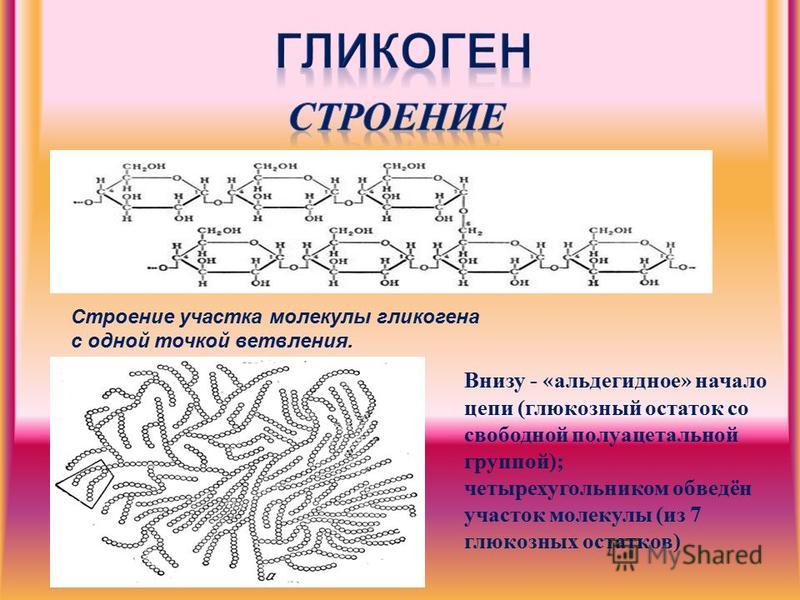 Гликогенин катализирует образование короткого полимера глюкозы, который удлиняется действием гликогенсинтазы. Фермент разветвления гликогена создает точки разветвления в частице гликогена через равные промежутки времени.Лафорин и малин являются белками, участвующими в сборке гликогена, но их специфическая функция у людей остается неуловимой. Гликоген накапливается в печени в основном в постпрандиальный период и в скелетных мышцах — в основном после упражнений. В цитозоле расщепление гликогена или гликогенолиз осуществляется двумя ферментами: гликогенфосфорилазой, которая высвобождает глюкозо-1-фосфат из линейных цепей гликогена, и ферментом, разветвляющим гликоген, который распутывает точки ветвления. В лизосомах деградация гликогена катализируется α-глюкозидазой.Система глюкозо-6-фосфатазы катализирует дефосфорилирование глюкозо-6-фосфата до глюкозы, что является необходимым этапом для выхода свободной глюкозы из клетки. Мутации в генах, кодирующих ферменты, участвующие в метаболизме гликогена, вызывают болезни накопления гликогена.
Гликогенин катализирует образование короткого полимера глюкозы, который удлиняется действием гликогенсинтазы. Фермент разветвления гликогена создает точки разветвления в частице гликогена через равные промежутки времени.Лафорин и малин являются белками, участвующими в сборке гликогена, но их специфическая функция у людей остается неуловимой. Гликоген накапливается в печени в основном в постпрандиальный период и в скелетных мышцах — в основном после упражнений. В цитозоле расщепление гликогена или гликогенолиз осуществляется двумя ферментами: гликогенфосфорилазой, которая высвобождает глюкозо-1-фосфат из линейных цепей гликогена, и ферментом, разветвляющим гликоген, который распутывает точки ветвления. В лизосомах деградация гликогена катализируется α-глюкозидазой.Система глюкозо-6-фосфатазы катализирует дефосфорилирование глюкозо-6-фосфата до глюкозы, что является необходимым этапом для выхода свободной глюкозы из клетки. Мутации в генах, кодирующих ферменты, участвующие в метаболизме гликогена, вызывают болезни накопления гликогена.
Ключевые слова
Глюкоза
Глюкокиназа
Фосфоглюкомутазы
Гликогенсинтаза
Гликогенфосфорилаза
α-Глюкозидаза
Резюме
0008 статей о накоплении гликогенаОпубликовано Elsevier B.V.
Рекомендуемые статьи
Цитирующие статьи
Накопление гликогена в связи с циклом линьки у двух ракообразных Emerita asiatica и Ligia exotica
Баумбергер, Дж. П. и Д. B. Укроп: исследование содержания гликогена и сахара и осмотического давления крабов во время цикла линьки. Physiol. Zoöl. 1 , 545–549 (1928).
Google ученый
Бернар, К.: Phenomènes de la vie, Париж: 1879.
Брэдфилд, Дж. Р. Г .: Гликоген эпидермиса позвоночных. Природа, Лондон. 167 , 40–41 (1951).
Р. Г .: Гликоген эпидермиса позвоночных. Природа, Лондон. 167 , 40–41 (1951).
Google ученый
Кэрролл, Н. В., Р. В. Лонгли и Дж. Х. Роу: Определение гликогена в печени и мышцах с использованием антронового реагента. J. biol. хим. 220 , 583–593 (1956).
PubMed
Google ученый
Хоэт, Дж.П. и П. М. Т. Керридж: Наблюдения за мышцами нормальных и линяющих ракообразных. Proc. R. Soc. (Сер. B) 100 , 116–119 (1926).
Google ученый
Мартин, А. Л .: Гистохимия цикла линьки Gammarus pulex (Crustacea, Amphipoda). J.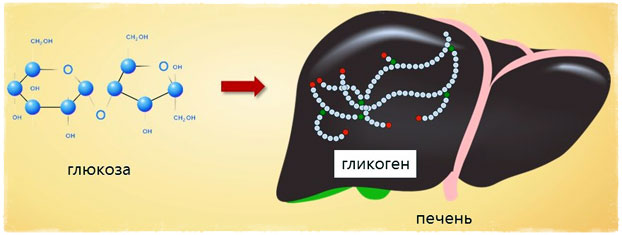 Zool. 147 , 185–200 (1965).
Zool. 147 , 185–200 (1965).
Google ученый
Парвати, К.: Исследования крови в связи с линькой у некоторых членистоногих, докторская диссертация, Мадрасский университет, 1968.
-: Уровень сахара в крови по отношению к синтезу хитина во время образования кутикулы в Emerita asiatica . Mar. Biol. 5 , 108–112 (1970).
Google ученый
-: Углеводный обмен двух ракообразных во время голодания. Mar. Biol. 8 , 1–5 (1971).
Google ученый
Пассано, Л. М .: Линька и борьба с ней. В : Физиология ракообразных, Vol. 1. С. 473–536. Эд. byT. Х. Уотерман. Нью-Йорк: Academic Press, 1960.
М .: Линька и борьба с ней. В : Физиология ракообразных, Vol. 1. С. 473–536. Эд. byT. Х. Уотерман. Нью-Йорк: Academic Press, 1960.
Google ученый
Patane, L .: Primi deti su gli aspetti Metabolici del fonomeno della muta negli isopodi. Атти. Accad. gioenia Sci. физ. 10 , 39–43 (1954).
Google ученый
Шеер, Б. Т.: Гормональный контроль обмена веществ у ракообразных. IX. Углеводный обмен при переходе от промежуточной к предварительной у Carcinides maenas . Биол. Бык. мар. биол. Lab., Woods Hole, , 116, , 175–183 (1959).
Google ученый
Schwabe, C. W., B. T. Scheer и M. А. Р. Шеер: Цикл линьки у Panulirus japonicus . Часть II. О гормональной регуляции обмена веществ у ракообразных. Physiologia comp.Oecol. 2 , 310–320 (1952).
W., B. T. Scheer и M. А. Р. Шеер: Цикл линьки у Panulirus japonicus . Часть II. О гормональной регуляции обмена веществ у ракообразных. Physiologia comp.Oecol. 2 , 310–320 (1952).
Google ученый
Рено, Л .: Mue et cycle d’intermue chez les crustacés décapodés. Annls Int. océanogr., Монако, 19 , 103–391 (1949).
Google ученый
Трэвис, Д. Ф .: Цикл линьки колючего лобстера, Panulirus argus
Латрей. II Преддизиальные, гистологические и гистохимические изменения в гепатопанкреасе и покровных тканях.Биол. Бык. мар. биол. Lab., Woods Hole, 108 , 88–112 (1955a).
Google ученый
-: Цикл линьки колючего лобстера Panulirus argus
Латрей.IV Постдизиальные гистологические и гистохимические изменения гепатопанкреаса и покровных тканей. Биол. Бык. мар. биол. Lab., Woods Hole, , 113, , 451–479 (1957).
Google ученый
-: Матрица и минеральные отложения в скелетных структурах десятиногих ракообразных. В : Кальцификация в биологических системах, Vol. 64. С. 57–116. Вашингтон, округ Колумбия: Американская ассоциация развития науки, 1960.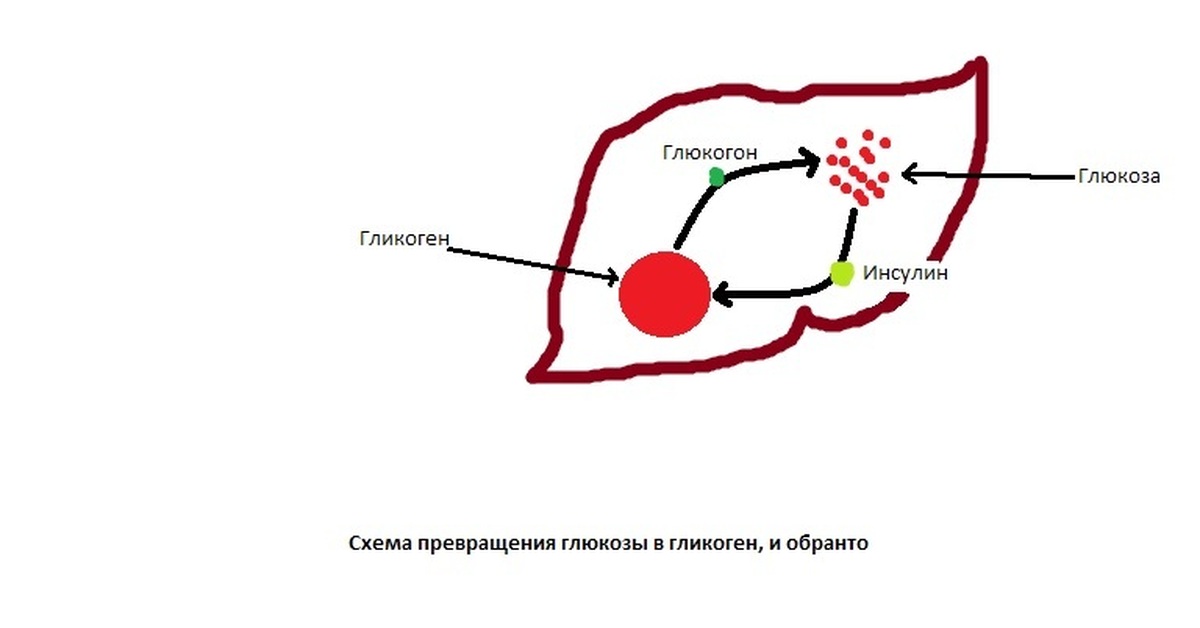
Google ученый
-: Отложение скелетных структур у ракообразных II.Гистохимические изменения, связанные с развитием неминерализованных компонентов скелета раков Orconectes virilis . Acta histochem. 15 , 251–268 (1963a).
PubMed
Google ученый
-: Отложение скелетной структуры у ракообразных. III Гистохимические изменения, связанные с развитием минерализованного гастролита у раков Orconectes virilis .Acta histochem. 15 , 269–284 (1963b).
PubMed
Google ученый
Верн, Дж .: Обратите внимание на гистохимический анализ метаболизма гликогена la mue chéz les crustacés. C. r. Séanc. Soc. Биол. 90 , 186–188 (1924).
C. r. Séanc. Soc. Биол. 90 , 186–188 (1924).
Google ученый
(PDF) Метаболизм гликогена у жвачных
Fernandez JM, Croom WJ Jr, Johnson AD, Jaquette RD, Edens FW (1988)
Субклиническая токсичность аммиака у бычков: влияние на метаболиты в крови
и концентрации регулирующих гормонов.Journal of Animal Science 66,
3259–3266.
Franch J, Aslesen R, Jensen J (1999) Регулирование синтеза гликогена в скелетных мышцах крысы
после гликоген-истощающей сократительной активности: эффекты
адреналина на синтез гликогена и активацию гликогенсинтазы
и гликогенфосфорилазы. Журнал биохимии 344, 231–235.
doi: 10.1042 / 0264-6021: 3440231
Gardner GE, Kennedy L, Milton JTB, Pethick DW (1999) Glycogen
метаболизм и конечный pH мышц у мериносов при первом скрещивании и
при втором скрещивании ягнята, пострадавшие от стресса перед убоем.
Австралийский журнал сельскохозяйственных исследований 50, 175–181. DOI: 10.1071 /
DOI: 10.1071 /
A98093
Gardner GE, Jacobs RH, Pethick DW (2001a) Влияние добавок оксида магния
на метаболизм гликогена в мышцах до и после упражнений
и при убое овец. Австралийский сельскохозяйственный журнал
Research 52, 723–729. DOI: 10.1071 / AR00128
Gardner GE, McIntyre BL, Tudor G, Pethick DW (2001b) Влияние питания
на метаболизм гликогена в мышцах крупного рогатого скота после упражнений.
Австралийский журнал сельскохозяйственных исследований 52, 461–470. DOI: 10.1071 /
AR00108
Gardner GE, McIntyre BL, Tudor GD, Pethick DW (2001c) Nutritional
влияет на восстановление мышечного гликогена после упражнений у овец
и крупного рогатого скота. В «Последние достижения в области питания животных в Австралии».
(Ed. JL Corbett), стр. 145–52. (Университет Новой Англии:
Армидейл, Новый Южный Уэльс)
Gardner GE, McGilchrist P, Thompson JM, Martin KM (2009) Selection
для мускулатуры снижает мышечную реакцию на адреналин. В «Международном симпозиуме
В «Международном симпозиуме
по физиологии жвачных животных». (Редакторы Y Chilliard, F Glasser, Y
Faulconnier, F Bocquier, I. Veissier, M. Doreau) стр. 430–431.
(Wageningen Academic Publishers: Clermont Ferrand, Франция)
Gardner GE, McIntyre BL, Tudor GD, Pethick DW (2013) Диеты с высоким содержанием
разлагаемого азота в рубце снижают концентрацию гликогена в мышцах крупного рогатого скота
при убое. В «59-м международном мясном конгрессе
науки и техники».(Редакторы М. Сердароглу, Б. Озтюрк, Т. Аккан)
(Материалы электронной книги: Измир, Турция)
Goodyear LJ, Hirshman MF, Smith RJ, Horton ES (1991) Транспортер глюкозы
Номер
, активность и содержание изоформы в плазматических мембранах красной
и белой скелетной мышцы. Американский журнал физиологии 261,
E556 – E561.
Greenwood PL, Gardner GE, Hegarty RS (2006) На характеристики миофибры ягненка
влияет оценочная племенная ценность производителей
и пастбищная система питания. Австралийский сельскохозяйственный журнал
Австралийский сельскохозяйственный журнал
Research 57, 627–639. doi: 10.1071 / AR04318
Gruber SL, Tatum JD, Engle TE, Chapman PL, Belk KE, Smith GC (2010)
Взаимосвязь поведенческих и физиологических симптомов предубойного стресса
с болезненностью длинных мышц говядины. Журнал зоотехники
88, 1148–1159. doi: 10.2527 / jas.2009-2183
Gunja-Smith Z, Marshall JJ, Mercier C, Smith EE, Whelan WJ (1970)
Пересмотр модели гликогена и амилопектина Мейера – Бернфельда.
FEBS Letters 12, 101–104. DOI: 10.1016 / 0014-5793 (70) 80573-7
Haman F, Peronnet F, Kenny GP, Massicotte D, Lavoie C, Weber JM (2005)
Разделение окислительного топлива во время воздействия холода на человека: мышцы
гликоген становится доминирующим по мере усиления дрожи. Журнал
Физиология 566, 247–256. doi: 10.1113 / jphysiol.2005.086272
Holness MJ, MacLennan PA, Palmer TN, Sugden MC (1988) Распределение
углеводов между гликогенезом, липогенезом и окислением в печени
во время перехода от голодания к кормлению.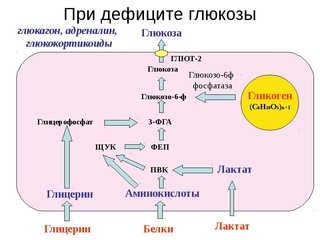 Биохимический журнал 252, 325–330.
Биохимический журнал 252, 325–330.
Jacob RH, Pethick DW, Chapman HM (2005) Концентрация мышечного гликогена
в коммерческих партиях австралийской баранины
, измеренная на ферме и после убоя после трех разных периодов хранения
. Австралийский журнал экспериментального сельского хозяйства 45, 543–552.
doi: 10.1071 / EA03216
Jacob RH, Gardner GE, Pethick DW (2009) Восполнение запасов гликогена в мышцах
предшествует восполнению запасов гликогена в печени свиней мериноса
.Животноводство 49, 131–138. DOI: 10.1071 / EA07387
Johnson RR (1976) Влияние растворимости углеводов на небелковую утилизацию азота
у жвачных животных. Journal of Animal Science 43,
184–191.
Кей Р.Н. (1983) Функция и физиология рубца. Ветеринарный учет
113,6–9. doi: 10.1136 / vr.113.1.6
Knee BW, Cummins LJ, Walker P, Warner R (2004) Сезонные колебания гликогена в мышцах
у бычков. Австралийский экспериментальный журнал
Agriculture 44, 729–734.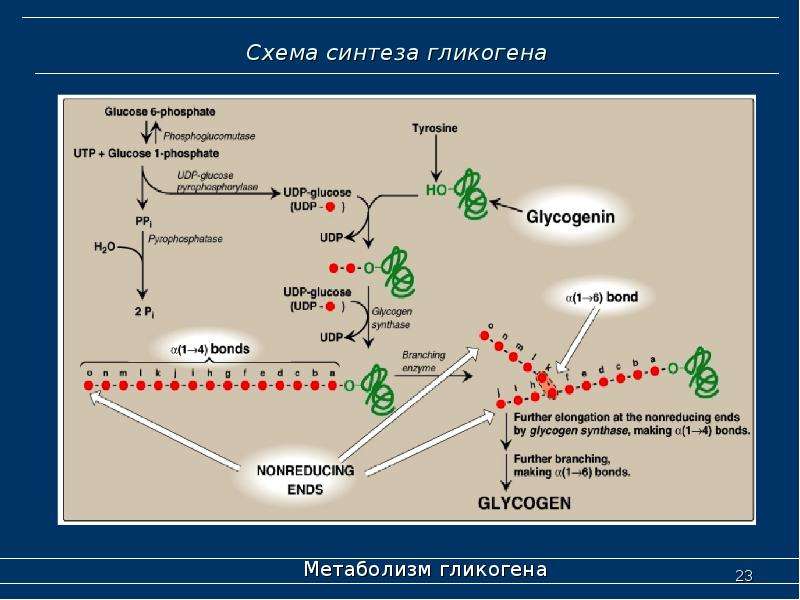 doi: 10.1071 / EA03044
doi: 10.1071 / EA03044
Knee BW, Cummins LJ, Walker PJ, Kearney GA, Warner RD (2007)
Уменьшение темноты стрижки у бычков, выращиваемых на пастбищах, с помощью высокоэнергетических добавок
. Австралийский журнал экспериментального сельского хозяйства 47,
1277–1283. DOI: 10.1071 / EA05362
Kumar R (1998) «Экспрессия гликогенина в мышцах овцы при различных метаболических условиях
» (Школа ветеринарных и биомедицинских наук,
Университет Мердока: Мердок, Вашингтон)
Maehlum S, Feliglum S. P, Wahren J (1978) Спланхническая глюкоза и мышцы
метаболизм гликогена после кормления глюкозой во время послетренировочного
восстановления.Американский журнал физиологии 235, E255 – E260.
Martin WH 3rd, Murphree SS, Safftz JE (1989) Распределение бета-адренергических рецепторов
среди типов мышечных волокон и резистентных артериол белого,
красного цвета и промежуточных скелетных мышц. Circulation Research 64,
1096–1105.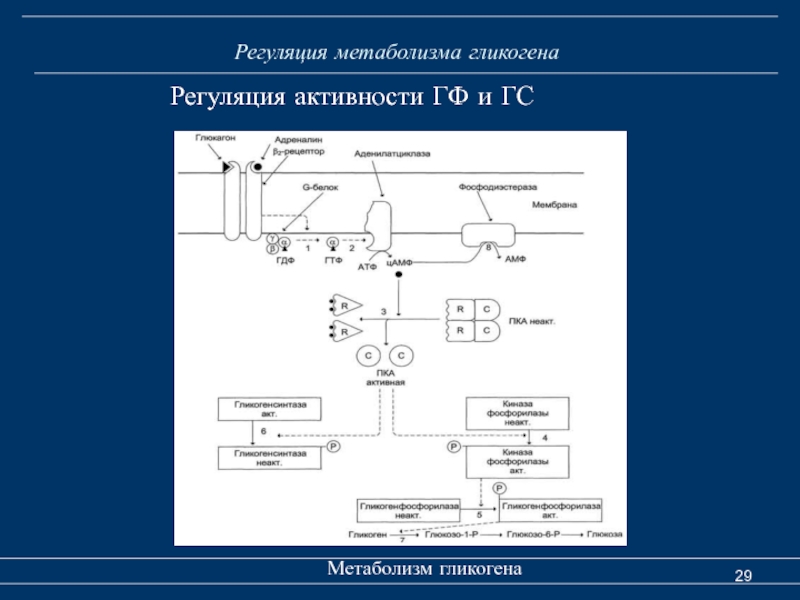 doi: 10.1161 / 01.RES.64.6.1096
doi: 10.1161 / 01.RES.64.6.1096
Martin KM, Gardner GE, Thompson JM, Hopkins DL (2004) Nutritional
влияние на метаболизм гликогена в мышцах ягнят, отобранных для мускулатуры.
Journal of Animal and Feed Sciences 13, 639–642.
Мартин К.М., МакГилкрист П., Томпсон Дж.М., Гарднер Г.Е. (2011) Потомство
-быков с высокой мускулатурой
имеет пониженный мышечный ответ на адреналин у овец.
Животное 5, 1060–1070. DOI: 10.1017 / S1751731110002764
McGilchrist P (2011) «Выбор мышечной ткани для углеводного и жирного
метаболизма кислоты у мясного крупного рогатого скота» (Школа ветеринарии и биомедицины
Sciences, Университет Мердока: Мердок, Вашингтон)
, Pethick DW, Bonny SPF, Greenwood PL, Gardner GE (2011a)
Мясной рогатый скот, отобранный из-за повышенной мускулатуры, имеет пониженную мышечную реакцию
и повышенную реакцию жировой ткани на адреналин.Животное 5,
875–884. doi: 10.1017 / S1751731110002508
McGilchrist P, Pethick DW, Bonny SPF, Greenwood PL, Gardner GE (2011b)
Чувствительность к инсулину всего тела выше у бычков, отобранных для увеличения мускулатуры
. животное 5, 1579–1586. doi: 10.1017 / S1751731111
животное 5, 1579–1586. doi: 10.1017 / S1751731111
000450
McGilchrist P, Alston CL, Gardner GE, Thomson KL, Pethick DW (2012)
Говяжьи туши с большей площадью глазных мышц, нижней частью костной ткани
и улучшенным питанием имеют более низкую частоту встречаемости темная резка.Мясо
Наука 92, 474–480. DOI: 10.1016 / j.meatsci.2012.05.014
Meinke MH, Edstrom RD (1991) Гликогенолиз мышц. Регламент
циклического взаимопревращения фосфорилазы а и фосфорилазы b.
Журнал биологической химии 266, 2259–2266.
Меллгрен Р.Л., Коулсон М. (1983) Скоординированная регуляция мышечной системы с обратной связью
Метаболизм гликогена: ингибирование очищенной фосфорилазы фосфатазы
гликогеном. Сообщения о биохимических и биофизических исследованиях
114, 148–154.DOI: 10.1016 / 0006-291X (83) 91606-6
MLA (2013) Ежегодный отчет о результатах деятельности Австралийских стандартов мяса за 2012–2013 годы.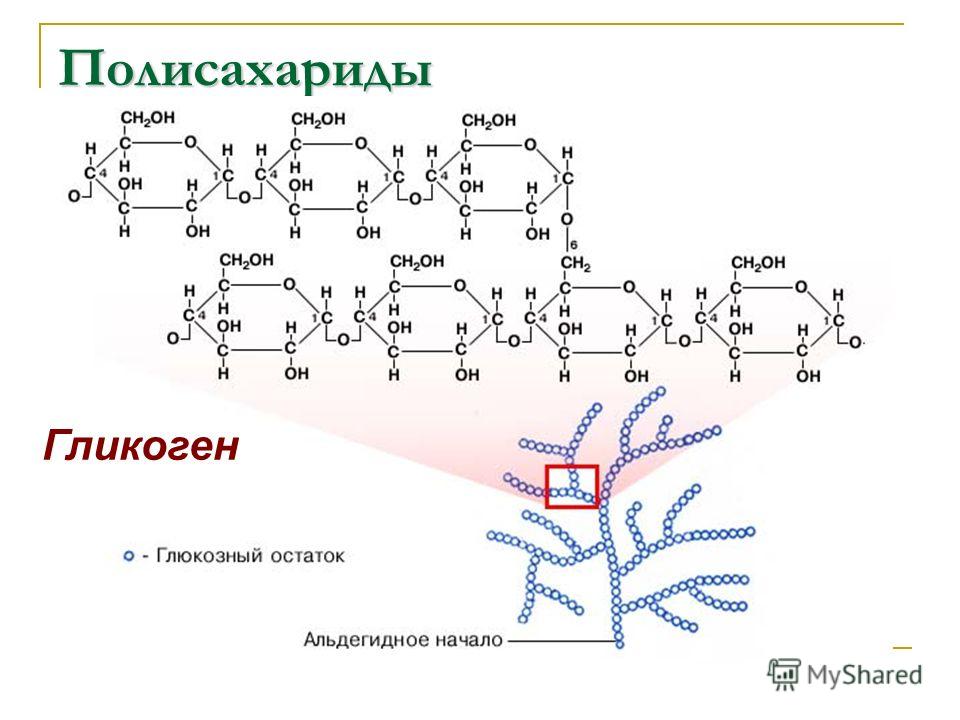
В «Годовых отчетах о результатах». (Ред. MLA) Доступно по адресу http: //www.mla.
com.au/files / 7bd7071d-bf9c-482e-b2e4-a24a0094d090 / MSA_AOR12-
13_web.pdf [проверено 4 июля 2014 г.]
Орсков Е.Р., Райл М (1990) «Энергетическое питание в Эльсеянц Applied
Science: London)
Pethick DW (1984) Энергетический метаболизм в скелетных мышцах.В «Физиология жвачных
: концепции и последствия». (Eds SK Baker, JM Gawthorne,
JB Mackintosh, DB Purser) стр. 277–287. (Западный университет
Австралия: Перт)
1582 Наука о животноводстве G. E. Gardner et al.
Лаборатория Депо | Гликоген, порошок
Химические вещества
Ваш выбор Laboratory Chemicals всегда должен быть сделан с учетом ценности. Предлагаемые здесь лабораторные химикаты, а также более 750 000 других лабораторных предметов были выбраны The Lab Depot в качестве ценных лабораторных принадлежностей.
Химические характеристики
Ultratrace, ACS: Подходит для использования в элементном анализе ultratrace. Соответствует или превосходит спецификации, перечисленные в документе Reagent Chemicals , опубликованном Американским химическим обществом.
Соответствует или превосходит спецификации, перечисленные в документе Reagent Chemicals , опубликованном Американским химическим обществом.
ACS: Отвечает или превосходит спецификации, перечисленные в Reagent Chemicals , опубликованном Американским химическим обществом.
USP: Отвечает требованиям Фармакопеи США или превосходит их.
NF: Отвечает требованиям Национального формуляра или превосходит их.
BP: Отвечает требованиям Британской фармакопеи или превосходит их.
EP: Отвечает требованиям Европейской фармакопеи или превосходит их.
JP: Отвечает требованиям Японской фармакопеи или превосходит их.
FCC: Отвечает требованиям Кодекса пищевых химикатов или превосходит их.
KSA: Сертифицированный кошерный.
UniSolv *: Универсальный растворитель чрезвычайно высокой чистоты, превосходящий требования для высокоэффективной жидкостной хроматографии (ВЭЖХ), газовой хроматографии (ГХ), спектрофотометрии и анализа остатков. Превосходит спецификации ACS. Контролируется на высокий уровень анализа и низкое УФ-поглощение, флуоресценцию, остатки и воду. Фактический анализ партии на этикетке.
Превосходит спецификации ACS. Контролируется на высокий уровень анализа и низкое УФ-поглощение, флуоресценцию, остатки и воду. Фактический анализ партии на этикетке.
HPLC: Растворитель высокой чистоты, подходящий для HPLC, спектрофотометрии или общего применения.
PestiSolv *: Растворитель высокой чистоты для анализа остатков пестицидов. Специально контролируется для методов ГЖХ с захватом электронов.
SpectroSolv *: Растворитель высокой чистоты для ультрафиолетовой спектрофотометрии.
BioSolv *: Растворитель высокой чистоты для биотехнологических приложений, включая биосинтез, секвенирование и разделение.
GCSolv *: Растворитель высокой чистоты для газовой хроматографии.
TraceGrade: Подходит для анализа следов металлов.
BiotechGrade: Продукты высокой чистоты, подходящие для использования в биотехнических целях.
UltraPure: Продукты для применений, в которых чистота имеет решающее значение.
Электроника / Чистое помещение: Продукты высокой чистоты, упакованные в чистом помещении для контроля твердых частиц.
Гистологический: Подходит для использования в гистологических лабораториях.
Реагент: Химикаты высокой чистоты, подходящие для использования во многих аналитических и общих лабораторных применениях.
C.P .: Chemically Pure обозначает химические вещества высокой чистоты, подходящие для использования во многих областях.
Очищено: Обозначает химические вещества хорошего качества, для которых нет официальных стандартов.
Практично: Относительно высокого качества, но может содержать небольшое количество примесей.
Технический: Обозначает химические вещества избранного коммерческого или промышленного качества.
Метаболизм гликогена у жвачных животных — Исследовательский репозиторий Университета Мердока
Гарднер, G.E.ORCID: 0000-0001-7499-9986, МакГилкрист, П. и Петик, Д.В. ORCID: 0000-0002-3255-7677
и Петик, Д.В. ORCID: 0000-0002-3255-7677
(2014)
Метаболизм гликогена у жвачных животных.
Животноводство, 54
(10).
С. 1575-1583.
Аннотация
Биохимия метаболизма гликогена хорошо изучена, она широко изучена на лабораторных грызунах и людях, и из этого проистекает основная часть наших знаний о метаболизме гликогена у жвачных животных.Что касается промежуточного метаболизма, ключевыми тканями являются печень и мышцы. Депо гликогена в печени играет центральную роль в промежуточном метаболизме, накапливая и мобилизуя гликоген во время метаболических состояний после еды и натощак, причем эти реакции модулируются во время беременности, кормления грудью и физических упражнений. В качестве альтернативы депо гликогена в мышцах особенно важно для местного энергетического гомеостаза и, вероятно, будет менее важным в качестве ключевого постпрандиального стока глюкозы в крови, учитывая пониженное всасывание глюкозы из кишечника у жвачных животных.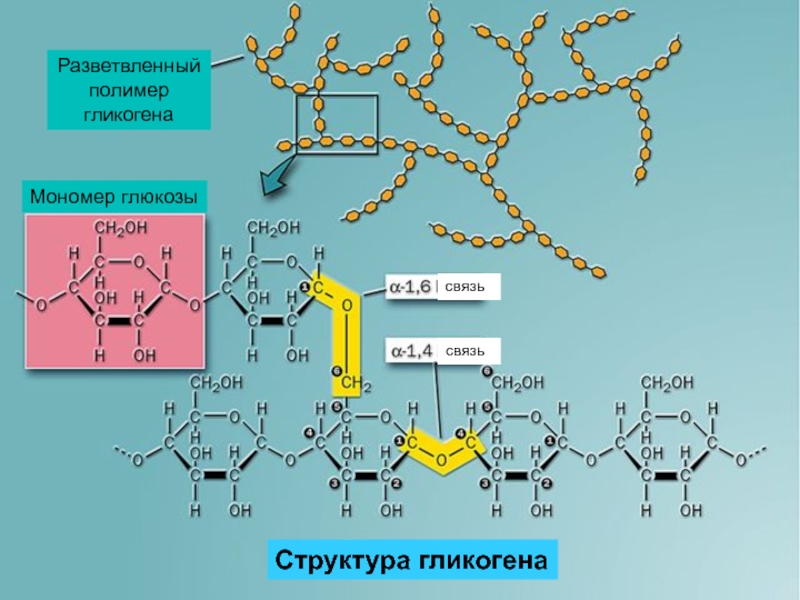 Однако, как и печень, это депо также находится в постоянном состоянии оборота, при этом концентрация гликогена в мышцах в любой момент времени отражает скорость синтеза и распада гликогена. Метаболизм мышечного гликогена привлекает особое внимание, учитывая его важность для посмертного закисления мышечной ткани, при нехватке на убое, что приводит к темной разделке мяса. Проще говоря, концентрация мышечного гликогена при убое является функцией двух ключевых факторов: начальные уровни гликогена на ферме за вычетом количества, истощенного во время предубойной фазы.Концентрации мышечного гликогена на ферме в значительной степени являются отражением потребления метаболизируемой энергии, что способствует увеличению скорости синтеза мышечного гликогена. По сравнению с видами с простыми желудками скорость синтеза гликогена у жвачных животных относительно низкая. Тем не менее, похоже, есть различия между овцами и рогатым скотом, когда их кормят диетами с одинаковой метаболизируемой энергией, при этом у крупного рогатого скота более медленное восполнение мышечного гликогена после истощения его в результате упражнений.
Однако, как и печень, это депо также находится в постоянном состоянии оборота, при этом концентрация гликогена в мышцах в любой момент времени отражает скорость синтеза и распада гликогена. Метаболизм мышечного гликогена привлекает особое внимание, учитывая его важность для посмертного закисления мышечной ткани, при нехватке на убое, что приводит к темной разделке мяса. Проще говоря, концентрация мышечного гликогена при убое является функцией двух ключевых факторов: начальные уровни гликогена на ферме за вычетом количества, истощенного во время предубойной фазы.Концентрации мышечного гликогена на ферме в значительной степени являются отражением потребления метаболизируемой энергии, что способствует увеличению скорости синтеза мышечного гликогена. По сравнению с видами с простыми желудками скорость синтеза гликогена у жвачных животных относительно низкая. Тем не менее, похоже, есть различия между овцами и рогатым скотом, когда их кормят диетами с одинаковой метаболизируемой энергией, при этом у крупного рогатого скота более медленное восполнение мышечного гликогена после истощения его в результате упражнений. Хотя потребление метаболической энергии является ключевым фактором, было показано, что генетические и возрастные факторы также влияют на восполнение запасов гликогена.Количество мышечного гликогена, истощенное во время предубойной фазы, в значительной степени связано со стрессом и высвобождением адреналина, и несколько недавних исследований охарактеризовали важность таких факторов, как физические упражнения, возраст и генетика, которые модулируют эту реакцию на стресс. В этой статье представлен обзор недавних экспериментов с крупным рогатым скотом и овцами, которые подчеркивают текущие достижения в понимании этого признака.
Хотя потребление метаболической энергии является ключевым фактором, было показано, что генетические и возрастные факторы также влияют на восполнение запасов гликогена.Количество мышечного гликогена, истощенное во время предубойной фазы, в значительной степени связано со стрессом и высвобождением адреналина, и несколько недавних исследований охарактеризовали важность таких факторов, как физические упражнения, возраст и генетика, которые модулируют эту реакцию на стресс. В этой статье представлен обзор недавних экспериментов с крупным рогатым скотом и овцами, которые подчеркивают текущие достижения в понимании этого признака.
| Страница управления предметами |
Болезнь накопления гликогена, тип V
УЧЕБНИКИ
Weinstein DA, Koeberl DD, Wolfsdorf JI.Болезнь накопления гликогена типа V. В: Справочник НОРД по редким заболеваниям. Липпинкотт Уильямс и Уилкинс. Филадельфия, Пенсильвания. 2003: 454-55.
ЖУРНАЛЬНЫЕ СТАТЬИ
de Luna N, Brull A, Guiu JM, Lucia A, Martin MA, Arenas J, Martí R, Andreu AL, Pinós T. Вальпроат натрия увеличивает изоформу гликогенфосфорилазы в мозге: поиск механизма компенсации в McArdle болезнь с использованием первичной культуры скелетных мышц мышей in vitro. Dis Model Mech. 2015 1 мая; 8 (5): 467-72
Nogales-Gadea G, Santalla A, Brull A, de Luna N, Lucia A, Pinós T.Патогеномика болезни Макардла — гены, ферменты, модели и терапевтическое значение. J Inherit Metab Dis. 2015 Март; 38 (2): 221-30.
Санталла А., Ногалес-Гадеа Г., Эртенблад Н., Брулл А., де Луна Н., Пинос Т., Люсия А. Болезнь Макардла: уникальная модель исследования в спортивной медицине. Sports Med. 2014 ноя; 44 (11): 1531-44
Люсия А, Руис Дж. Р., Санталла А, Ногалес-Гадеа Дж., Рубио Дж. К., Гарсия-Консуэгра I, Кабельо А, Перес М, Тейейра С., Виитес I, Наварро С., Аренас Дж., Мартин М.А., Андреу А.Л. Генотипические и фенотипические особенности болезни Макардла: информация из национального реестра Испании. J Neurol Neurosurg Psychiatry. 2012 Март; 83 (3): 322-8.
J Neurol Neurosurg Psychiatry. 2012 Март; 83 (3): 322-8.
Quinlivan R, Buckley J, James M, Twist A, Ball S, Duno M, Vissing J, Bruno C, Cassandrini D, Roberts M, Winer J, Rose M, болезнь Сьюри К. Макардла: клинический обзор. J Neurol Neurosurg Psychiatry. 2010 Nov; 81 (11): 1182-
Howell JM, Walker KR, Davies L, Dunton E, Everaardt A, Laing N, Karpati G.
Аденовирус и аденоассоциированная вирусная доставка кДНК миофосфорилазы человека и LacZ кДНК в мышцу в модели болезни Макардла у овец: экспрессия и повторная экспрессия гликогенфосфорилазы.Нейромышечное расстройство. Март 2008; 18 (3): 248-58.
Hadjigeorgiou GM, Sadeh M, Musumeci O, et al. Две новые мутации в гене миофосфорилазы у пациента с болезнью Макардла. Нейромышечное расстройство. 2002; 12: 824-27.
Халлер Р.Г., Виссинг Дж. Спонтанное «второе дыхание» и индуцированное глюкозой «второе дыхание» при болезни Макардла: окислительные механизмы. Arch Neurol. 2002; 59: 1395-402.
Каземи-Эсфарджани П. , Скоморовска Э., Йенсен Т.Д. и др. Тест с физической нагрузкой для предплечий без ишемии при болезни Макардла.Энн Нейрол. 2002; 52: 153-59.
, Скоморовска Э., Йенсен Т.Д. и др. Тест с физической нагрузкой для предплечий без ишемии при болезни Макардла.Энн Нейрол. 2002; 52: 153-59.
Jensen TD, Kazemi-Esfarjani P, Skomorowska E, et al. Скрининговый тест с нагрузкой для предплечий на митохондриальную миопатию. Неврология. 2002; 58: 1533-38.
Воргерд М., Занге Дж., Клей Р. и др. Влияние терапии высокими дозами креатина на симптомы непереносимости физических упражнений при болезни Макардла. Двойное слепое плацебо-контролируемое перекрестное исследование. Arch Neurol. 2002; 59: 97-101.
Martinuzzi A, Schievano G, Nascimbeni A, et al. Болезнь Макардла. Неразгаданная загадка вновь появляющегося фермента.Am J Pathol. 1999; 154: 1893-97.
DiMauro S, Bruno C. Болезни накопления гликогена в мышцах. Curr Opin Neurol. 1998; 11: 477-84.
ИНТЕРНЕТ
Мартин М.А., Люсия А., Аренас Дж. И др. Болезнь накопления гликогена, тип V. 19 апреля 2006 г. [Обновлено 26 июня 2014 г.]. В: Pagon RA, Adam MP, Ardinger HH, et al. , Редакторы. GeneReviews [Интернет]. Сиэтл (Вашингтон): Вашингтонский университет, Сиэтл; 1993-2015. Доступно по адресу: http://www.ncbi.nlm.nih.gov/books/NBK1344/ По состоянию на 10 июня 2015 г.
, Редакторы. GeneReviews [Интернет]. Сиэтл (Вашингтон): Вашингтонский университет, Сиэтл; 1993-2015. Доступно по адресу: http://www.ncbi.nlm.nih.gov/books/NBK1344/ По состоянию на 10 июня 2015 г.
McKusick VA, Ed.Интернет-Менделирующее наследование в человеке (OMIM). Болезнь накопления гликогена V. Университет Джона Хопкинса. Регистрационный номер; 232600. Доступно по адресу http://omim.org/entry/232600. : Дата последнего редактирования: 29.01.2015. Доступ 10 июня 2015 г.
Андерсон В. Болезнь накопления гликогена типа V. Обновлено: 25 августа 2014 г. Доступно по адресу: www.emedicine.com/med/topic911.htm По состоянию на 10 июня 2015 г.
CSIRO PUBLISHING | Животноводство
Список литературы
Баллард Ф.Дж., Хансон Р.В., Кронфельд Д.С.
(1969 ) Глюконеогенез и липогенез в тканях жвачных и нежвачных животных. Federation Proceedings 28 , 218–231.
Бассет Дж. М.
(1972 ) Концентрации глюкагона в плазме овец: их регуляция и связь с концентрациями инсулина и гормона роста. Австралийский журнал биологических наук 25 , 1277–1287.
Австралийский журнал биологических наук 25 , 1277–1287.
Белл А. В., Гарднер Дж. В., Мэнсон В., Томпсон Дж. Э.
(1975 ) Острое воздействие холода и метаболизм глюкозы в крови, лактата и пирувата, а также аминокислот в плазме в задней ноге кормленного и голодного молодого быка. Британский журнал питания 33 , 207–217.
| Острое воздействие холода и метаболизм глюкозы в крови, лактата и пирувата, а также аминокислот в плазме в задних лапах кормленного и голодного молодого быка.Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DyaE2MXhtV2lu7w% 3D & md5 = c64b0d66bbe2d051569865b5479174c1CAS | 1115760PubMed |
Брандстеттер А.М., Пикард Б., Гей Y
(1998 ) Характеристики мышечных волокон четырех мышц растущих быков. I. Постнатальная дифференциация. Наука животноводства 53 , 15–23.
| Характеристики мышечных волокон четырех мышц растущих бычков. I. Постнатальная дифференциация. Crossref | GoogleScholarGoogle Scholar |
Briand M, Talmant A, Braind Y, Monin G, Durand R
(1981 ) Метаболические типы мышц у овец: I. Миозин-АТФаза, гликолитическая и митохондриальная ферментативная активность. Европейский журнал прикладной физиологии 46 , 347–358.
Миозин-АТФаза, гликолитическая и митохондриальная ферментативная активность. Европейский журнал прикладной физиологии 46 , 347–358.
| Метаболические типы мышц у овец: I.Миозин-АТФаза, активность гликолитических и митохондриальных ферментов.Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DyaL3MXlvVCntL8% 3D & md5 = 4d3344e60cb0f900b2956a35874bd5c4CAS |
Брокман Р.П., Лаарвельд Б.
(1986 ) Гормональная регуляция метаболизма у жвачных животных — обзор. Наука животноводства 14 , 313–334.
| Гормонально-регуляция обмена веществ у жвачных животных — обзор.Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DyaL28XkvFWnsLc% 3D & md5 = 6c49e8dcfb716fbb9efc9fce3e79e043CAS |
Бердик, NC, Кэрролл Дж. А., Халберт Л. Е., Дейли Дж. В., Уиллард, С. Т., Ванн, Р. К., Уэльс, Т. Д., Рандел, Р. Д.
(2010 ) Связь между темпераментом и транспортировкой с ректальной температурой и концентрацией кортизола и адреналина в сыворотке у быков.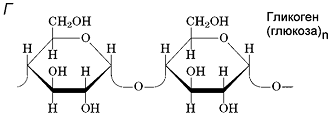 Животноводство 129 , 166–172.
Животноводство 129 , 166–172.
| Взаимосвязь темперамента и транспорта с ректальной температурой и концентрацией кортизола и адреналина в сыворотке у быков.Crossref | GoogleScholarGoogle Scholar |
Бердик, NC, Кэрролл Дж. А., Халберт Л. Е., Дейли Дж. В., Баллоу, Массачусетс, Рэндел Р. Д., Уиллард С. Т., Ванн Р. К., Валлийский TH
(2011 ) Темперамент влияет на вызванные эндотоксинами изменения ректальной температуры, болезненного поведения и концентрации адреналина в плазме у быков. Врожденный иммунитет 17 , 355–364.
| Темперамент влияет на вызванные эндотоксинами изменения ректальной температуры, болезненное поведение и концентрацию адреналина в плазме у быков.Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DC% 2BC3MXhtFGisrvK & md5 = 48bdc6597619b42dd5ae7bf3fa6cf2dcCAS | 20682590PubMed |
Кафе LM, Robinson DL, Ferguson DM, McIntyre BL, Geesink GH, Greenwood PL
(2011 ) Темперамент крупного рогатого скота: постоянство оценок и ассоциаций с производительностью, продуктивностью, показателями качества туши и мяса. Журнал зоотехники 89 , 1452–1465.
Журнал зоотехники 89 , 1452–1465.
| Темперамент крупного рогатого скота: стойкость оценок и ассоциаций с производительностью, продуктивностью, показателями качества туши и мяса. GoogleScholarGoogle Scholar | 1: CAS: 528: DC% 2BC3MXls1Oiu74% 3D & md5 = 428fb5ad67bc4579d593e39d0492fae8CAS | 21169516PubMed |
Кумбс С.В., Гарднер Дж. Э., Петик Д. В., МакГилкрист П.
(2014 ) Влияние темперамента животного на гликоген в мышцах, лактат в мышцах и концентрацию лактата в плазме при убое. Meat Science в печати.
Керли К.О., Кэрролл Д.А., Ванн Р.К., Рэндел Р.Д., Валлийский TH
(2010 ) Сравнение чувствительности надпочечников к кортикотропин-рилизинг-гормону (CRH) у бычков ангуса и брахмана с разным темпераментом. Журнал молочной промышленности 93 , 19
Дэли Б.Л., Гарднер Г.Е., Фергюсон Д.М., Томпсон Дж. М.
(2006 ) Влияние перерыва в кормлении перед убоем на метаболизм гликогена в мышцах и скорость снижения pH. Австралийский журнал сельскохозяйственных исследований 57 , 1229–1235.
| Влияние перерыва в кормлении перед убоем на метаболизм гликогена в мышцах и скорость снижения pH.Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DC% 2BD28XhtFeqsr7M & md5 = 78d7e290ba25ed0f23be96aea41ed49bCAS |
Давидович А., Бартли Э., Чепмен Т. Е., Бехтл Р. М., Дейтон А. Д., Фрей Р. А.
(1977 ) Токсичность аммиака для крупного рогатого скота. II. Изменения компонентов сонной и яремной крови, связанные с токсичностью. Журнал зоотехники 44 , 702–709.
Эбнер К., Вотяк К. Т., Ландграф Р., Энгельманн М.
(2005 ) Нейроэндокринная и поведенческая реакция на социальную конфронтацию: жители против злоумышленников, активные и пассивные стили совладания. Гормоны и поведение 47 , 14–21.
| Нейроэндокринная и поведенческая реакция на социальную конфронтацию: жители против злоумышленников, активные и пассивные стили совладания. Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DC% 2BD2cXhtVaqt73O & md5 = 3dc2ca1f66b7146848281647599034f5CAS | 15579261PubMed |
Фелиг П.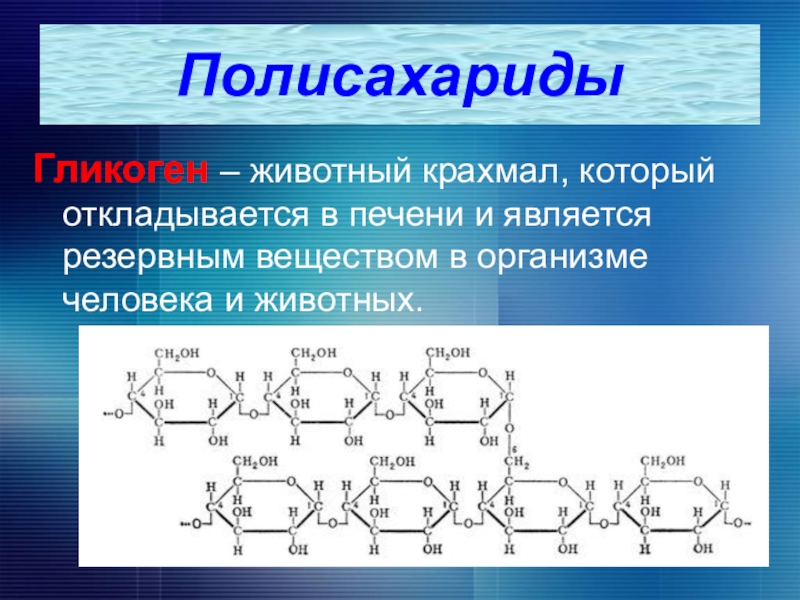 , Варен Дж., Хендлер Р.
, Варен Дж., Хендлер Р.
(1975 ) Влияние перорального приема глюкозы на внутреннюю глюкозу и метаболизм глюконеогенных субстратов у человека. Диабет 24 , 468–475.
| Влияние перорального приема глюкозы на внутренний метаболизм глюкозы и глюконеогенных субстратов у человека.Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DyaE2MXkvFCnu74% 3D & md5 = a5e8dfe30bb5111
bb3923689156CAS | 1126590PubMed |
Фергюсон Д.М., Брюс Х.Л., Томпсон Дж. М., Иган А.Ф., Перри Д., Короткие рукава WR
(2001 ) Факторы, влияющие на вкусовые качества говядины — от фермы до охлажденной тушки. Австралийский журнал экспериментального сельского хозяйства 41 , 879–891.
| Факторы, влияющие на вкусовые качества говядины — от фермы до охлажденной тушки.Crossref | GoogleScholarGoogle Scholar |
Фернандес Дж. М., Крум В. Дж., Джонсон А. Д., Жакетт Р. Д., Эденс Ф. В.
(1988 ) Субклиническая токсичность аммиака у бычков: влияние на метаболиты в крови и концентрации регулирующих гормонов. Журнал зоотехники 66 , 3259–3266.
Журнал зоотехники 66 , 3259–3266.
Франч Дж., Аслесен Р., Дженсен Дж.
(1999 ) Регулирование синтеза гликогена в скелетных мышцах крысы после истощающей гликоген сократительной активности: влияние адреналина на синтез гликогена и активацию гликогенсинтазы и гликогенфосфорилазы. Журнал биохимии 344 , 231–235.
| Регуляция синтеза гликогена в скелетных мышцах крысы после истощающей гликоген сократительной активности: влияние адреналина на синтез гликогена и активацию гликогенсинтазы и гликогенфосфорилазы.Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DyaK1MXns1yjtL0% 3D & md5 = 2450917806adb5fc32cc793eafd2c53dCAS |
Гарднер Дж. Э., Кеннеди Л., Милтон Дж. Т. Б., Петик Д. В.
(1999 ) Метаболизм гликогена и конечный pH мышц мериносовых ягнят, ягнят первого и второго скрещивания, подверженных стрессу перед убой. Австралийский журнал сельскохозяйственных исследований 50 , 175–181.
| Метаболизм гликогена и предельное значение pH в мышцах мериносовых ягнят, ягнят первого и второго скрещивания под воздействием стресса перед убоем. Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DyaK1MXhslCrs7s% 3D & md5 = acb7351768acf5b8f8c94566bdb0161fCAS |
Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DyaK1MXhslCrs7s% 3D & md5 = acb7351768acf5b8f8c94566bdb0161fCAS |
Гарднер Г.Е., Джейкобс Р.Х., Петик Д.В.
(2001 a ) Влияние добавок оксида магния на метаболизм гликогена в мышцах до и после физических упражнений и при убое овец. Австралийский журнал сельскохозяйственных исследований 52 , 723–729.
| Влияние добавок оксида магния на метаболизм гликогена в мышцах овец до и после упражнений, а также при убое. GoogleScholarGoogle Scholar | 1: CAS: 528: DC% 2BD3MXlvVCrsLo% 3D & md5 = 0a21a1ae3735fa5f530f7108a76fb380CAS |
Гарднер Г.Е., Макинтайр Б.Л., Тюдор Дж., Петик Д.В.
(2001 b ) Влияние питания на метаболизм гликогена в мышцах крупного рогатого скота после физических упражнений. Австралийский журнал сельскохозяйственных исследований 52 , 461–470.
| Влияние питания на метаболизм гликогена в мышцах крупного рогатого скота после физических упражнений.Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DC% 2BD3MXjsFShsrY% 3D & md5 = fca8f6b85d561f0462ae8f5f4209a685CAS |
Гарднер Г.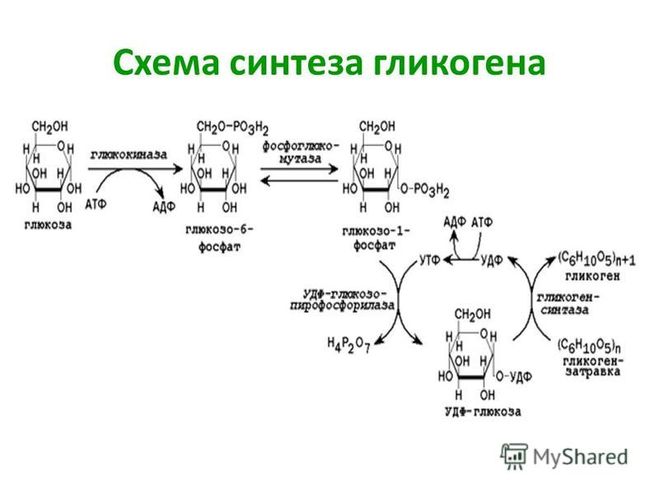 Е., Макинтайр Б.Л., Тюдор Г.Д., Петик Д.В. (2001 c ) Влияние питания на восстановление мышечного гликогена после упражнений у овец и крупного рогатого скота. В «Последние достижения в области питания животных в Австралии».(Ред. Дж. Л. Корбетт) стр. 145–52. (Университет Новой Англии: Армидейл, Новый Южный Уэльс)
Е., Макинтайр Б.Л., Тюдор Г.Д., Петик Д.В. (2001 c ) Влияние питания на восстановление мышечного гликогена после упражнений у овец и крупного рогатого скота. В «Последние достижения в области питания животных в Австралии».(Ред. Дж. Л. Корбетт) стр. 145–52. (Университет Новой Англии: Армидейл, Новый Южный Уэльс)
Gardner GE, McGilchrist P, Thompson JM, Martin KM (2009) Выбор мышечной массы снижает мышечную реакцию на адреналин. В «Международном симпозиуме по физиологии жвачных животных». (Редакторы Y. Chilliard, F Glasser, Y Faulconnier, F Bocquier, I. Veissier, M. Doreau) стр. 430–431. (Вагенингенское академическое издательство: Клермон-Ферран, Франция)
Gardner GE, McIntyre BL, Tudor GD, Pethick DW (2013) Диеты с высоким содержанием разлагаемого в рубце азота снижают концентрацию гликогена в мышцах крупного рогатого скота при убое.В «59-м международном конгрессе мясной науки и технологий». (Редакторы М. Сердароглу, Б. Озтюрк, Т Аккан) (Труды электронной книги: Измир, Турция)
Goodyear LJ, Hirshman MF, Smith RJ, Horton ES
(1991 ) Число переносчиков глюкозы, активность и содержание изоформ в плазматических мембранах красных и белых скелетных мышц. Американский журнал физиологии 261 , E556 – E561.
Американский журнал физиологии 261 , E556 – E561.
Greenwood PL, Gardner GE, Hegarty RS
(2006 ) Характеристики миофибры ягненка зависят от оценочной племенной ценности производителей и пастбищной системы питания. Австралийский журнал сельскохозяйственных исследований 57 , 627–639.
| На характеристики миофибры ягненка влияют оценочные племенные ценности производителей и пастбищная система питания. Crossref | GoogleScholarGoogle Scholar |
Грубер С.Л., Татум Дж.Д., Энгл Т.Э., Чапман П.Л., Белк К.Е., Смит Г.С.
(2010 ) Связь поведенческих и физиологических симптомов предубойного стресса с болезненностью длинных мышц говядины. Журнал зоотехники 88 , 1148–1159.
| Взаимосвязь поведенческих и физиологических симптомов предубойного стресса с болезненностью длиннейшей мышцы говядины.Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DC% 2BC3cXktVyjs7w% 3D & md5 = cdcb1baf13b10efcc65e7b768b0b9257CAS | 19897634PubMed |
Гунджа-Смит З. , Маршалл Дж. Дж., Мерсье К., Смит Э., Уилан В. Дж.
, Маршалл Дж. Дж., Мерсье К., Смит Э., Уилан В. Дж.
(1970 ) Пересмотр модели Мейера – Бернфельда гликогена и амилопектина. Письма FEBS 12 , 101–104.
| Пересмотр модели гликогена и амилопектина Мейера – Бернфельда.Crossref | GoogleScholarGoogle Scholar | 11945551PubMed |
Хаман Ф., Перонне Ф., Кенни Г. П., Массикотт Д., Лавуа С., Вебер Дж. М.
(2005 ) Разделение окислительного топлива на человека при воздействии холода: гликоген в мышцах становится доминирующим по мере усиления дрожи. Журнал физиологии 566 , 247–256.
| Разделение окислительного топлива на человека при воздействии холода: гликоген в мышцах становится доминирующим по мере усиления дрожи.Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DC% 2BD2MXmvVyqsbw% 3D & md5 = 0a007fbdbe0e49b10a550718dca98fceCAS | 15831534PubMed |
Холнесс MJ, MacLennan PA, Palmer TN, Sugden MC
(1988 ) Распределение углеводов между гликогенезом, липогенезом и окислением в печени во время перехода от голодания к кормлению. Биохимический журнал 252 , 325–330.
Биохимический журнал 252 , 325–330.
Джейкоб Р. Х., Петик Д. В., Чепмен Х. М.
(2005 ) Концентрации мышечного гликогена в коммерческих партиях австралийского ягненка, измеренные на ферме и после убоя после трех различных периодов содержания. Австралийский журнал экспериментального сельского хозяйства 45 , 543–552.
| Концентрация гликогена в мышцах в коммерческих партиях австралийского ягненка, измеренная на ферме и после убоя после трех различных периодов хранения.Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DC% 2BD2MXlsFOmtbc% 3D & md5 = 86c15fc40d04e9464b58aaf90cca762aCAS |
Джейкоб Р.Х., Гарднер Г.Е., Петик Д.В.
(2009 ) Восполнению запасов гликогена в мышцах предшествует восполнение запасов гликогена в печени свиней-мериносов. Наука о животноводстве 49 , 131–138.
| Восполнению запасов гликогена в мышцах предшествует восполнение запасов гликогена в печени мериносовых свиней. Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DC% 2BD1MXhtVehsb4% 3D & md5 = 211faabdf3b2e332662053633d32dce9CAS |
Джонсон Р. Р.
Р.
(1976 ) Влияние растворимости углеводов на использование небелкового азота у жвачных животных. Журнал зоотехники 43 , 184–191.
Кей Р.Н.
(1983 ) Функция и физиология рубца. Ветеринарная карта 113 , 6–9.
| Функция и физиология рубца. Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DyaL3sXkvVGmtr8% 3D & md5 = 958b652018c92870ecbc45fe6556adaeCAS | 6880003PubMed |
Колено BW, Cummins LJ, Walker P, Warner R
(2004 ) Сезонные колебания мышечного гликогена у бычков. Австралийский журнал экспериментального сельского хозяйства 44 , 729–734.
| Сезонные колебания мышечного гликогена у говяжьих бычков.Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DC% 2BD2cXnslShurg% 3D & md5 = 6d31d9cd66e7f13a5b92e37e7e05b083CAS |
Колено BW, Cummins LJ, Walker PJ, Kearney GA, Warner RD
(2007 ) Сокращение темного покоса у бычков, выращиваемых на пастбищах, с помощью высокоэнергетических добавок. Австралийский журнал экспериментального сельского хозяйства 47 , 1277–1283.
Австралийский журнал экспериментального сельского хозяйства 47 , 1277–1283.
| Сокращение темного покоса у бычков, выращиваемых на пастбищах, с помощью высокоэнергетических добавок.Crossref | GoogleScholarGoogle Scholar |
Кумар Р. (1998) «Экспрессия гликогенина в мышцах овцы при различных метаболических условиях». (Школа ветеринарных и биомедицинских наук, Университет Мердока: Мердок, Вашингтон)
Маэлум С., Фелиг П., Варен Дж.
(1978 ) Спланхнический метаболизм глюкозы и мышечного гликогена после приема глюкозы во время восстановления после нагрузки. Американский журнал физиологии 235 , E255 – E260.
Мартин WH, Мерфри СС, Саффитц Дж. Э.
(1989 ) Распределение бета-адренорецепторов среди типов мышечных волокон и артериол сопротивления белой, красной и промежуточной скелетных мышц. Исследования циркуляции 64 , 1096–1105.
| Распределение бета-адренорецепторов среди типов мышечных волокон и артериол сопротивления белой, красной и промежуточных скелетных мышц. Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DyaL1MXksFSqsb4% 3D & md5 = c0a6f243b41c336f798189d498a8e9c9CAS | 2541942PubMed |
Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DyaL1MXksFSqsb4% 3D & md5 = c0a6f243b41c336f798189d498a8e9c9CAS | 2541942PubMed |
Мартин К.М., Гарднер Г.Е., Томпсон Дж.М., Хопкинс Д.Л.
(2004 ) Влияние питания на метаболизм гликогена в мышцах ягнят, отобранных для мускулатуры. Журнал наук о животных и кормах 13 , 639–642.
Мартин К.М., МакГилкрист П., Томпсон Дж. М., Гарднер Дж. Э.
(2011 ) Потомство быков с высокой мускулатурой снизило мышечную реакцию на адреналин у овец. Животное 5 , 1060–1070.
| Потомство быков с высокой мускулатурой снизило мышечную реакцию на адреналин у овец.Crossref | GoogleScholarGoogle Scholar | 1: STN: 280: DC% 2BC38vovV2isQ% 3D% 3D & md5 = 9654afb82fccc89b714ac9ffe8314742CAS | 22440101PubMed |
МакГилкрист П. (2011 г.) «Отбор по влиянию мышечной массы на метаболизм углеводов и жирных кислот у мясного скота.’(Школа ветеринарных и биомедицинских наук, Университет Мердока: Мердок, Вашингтон)
McGilchrist P, Pethick DW, Bonny SPF, Greenwood PL, Gardner GE
(2011 a ) У мясного крупного рогатого скота, отобранного из-за повышенной мускулатуры, снижен мышечный ответ и повышен ответ жировой ткани на адреналин.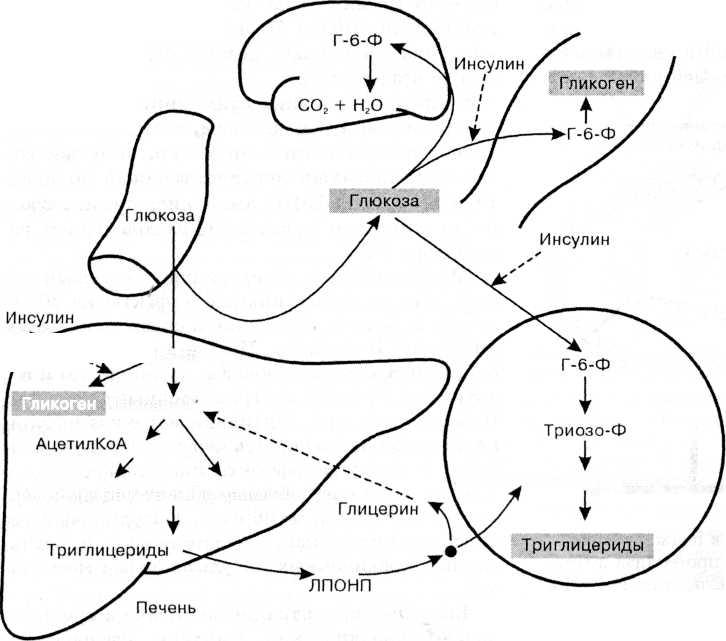 Животное 5 , 875–884.
Животное 5 , 875–884.
| Мясной скот, отобранный по повышенной мускулатуре, имеет пониженный мышечный ответ и повышенный ответ жировой ткани на адреналин.Crossref | GoogleScholarGoogle Scholar | 1: STN: 280: DC% 2BC38vovVyktg% 3D% 3D & md5 = 86616898e73a3fd2e532db40797de427CAS | 22440027PubMed |
McGilchrist P, Pethick DW, Bonny SPF, Greenwood PL, Gardner GE
(2011 b ) Чувствительность к инсулину всего тела выше у бычков, отобранных для увеличения мускулатуры. животное 5 , 1579–1586.
| Чувствительность к инсулину всего тела выше у бычков, отобранных для увеличения мышечной массы.Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DC% 2BC3MXht1yms7rI & md5 = d967b79e694e911793b7d5919dc63dedCAS | 22440349PubMed |
МакГилкрист П., Олстон К.Л., Гарднер Г.Е., Томсон К.Л., Петик Д.В.
(2012 ) Говяжьи туши с большей площадью глазных мышц, более низкими показателями окостенения и улучшенным питанием имеют меньшую вероятность темного разделывания.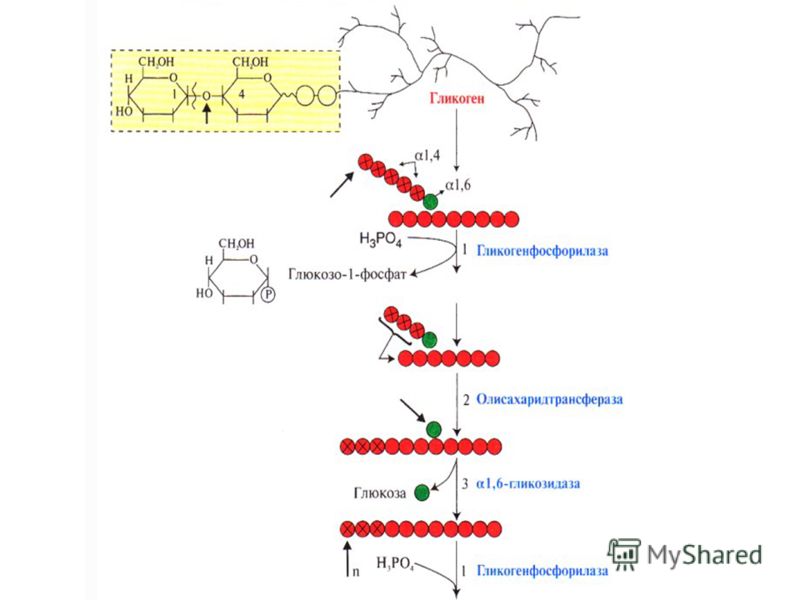 Meat Science 92 , 474–480.
Meat Science 92 , 474–480.
| Туши говядины с увеличенной площадью глазных мышц, более низкими показателями окостенения и улучшенным питанием имеют меньшую вероятность темного разделки.Crossref | GoogleScholarGoogle Scholar | 1: STN: 280: DC% 2BC38jjsVKhuw% 3D% 3D & md5 = fe9793a8548011feaa3267536b957463CAS | 22717222PubMed |
Meinke MH, Edstrom RD
(1991 ) Гликогенолиз мышц. Регулирование циклического взаимопревращения фосфорилазы а и фосфорилазы b. Журнал биологической химии 266 , 2259–2266.Меллгрен Р.Л., Колсон М.
(1983 ) Скоординированная обратная связь регуляции метаболизма мышечного гликогена: ингибирование очищенной фосфорилазы фосфатазы гликогеном. Сообщения о биохимических и биофизических исследованиях 114 , 148–154.
| Скоординированная обратная связь регуляции метаболизма гликогена в мышцах: ингибирование очищенной фосфорилазы фосфатазы гликогеном. Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DyaL3sXks1KmtLk% 3D & md5 = 946ccae40e340850eb74c26757beccc3CAS | 6309163PubMed |
MLA (2013) Годовой отчет Австралии по стандартам мяса, 2012–2013 гг. В «Годовых отчетах о результатах». (Ред. MLA) Доступно на http://www.mla.com.au/files/7bd7071d-bf9c-482e-b2e4-a24a0094d090/MSA_AOR12-13_web.pdf [Проверено 4 июля 2014 г.]
В «Годовых отчетах о результатах». (Ред. MLA) Доступно на http://www.mla.com.au/files/7bd7071d-bf9c-482e-b2e4-a24a0094d090/MSA_AOR12-13_web.pdf [Проверено 4 июля 2014 г.]
Орсков Э. Р., Райл М. (1990) «Энергетическое питание жвачных животных». (Elsevier Applied Science: Лондон)
Петик Д.В. (1984) Энергетический метаболизм в скелетных мышцах. В книге «Физиология жвачных животных: концепции и последствия». (Под ред. С. К. Бейкера, Дж. М. Гоуторна, Дж. Б. Макинтоша, Д. Б. Персера) стр. 277–287. (Университет Западной Австралии: Перт)
Петик DW
(1993 ) Окисление углеводов и липидов во время упражнений. Австралийский журнал сельскохозяйственных исследований 44 , 431–441.
| Окисление углеводов и липидов во время упражнений.Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DyaK2cXltVGit78% 3D & md5 = 0bd322a1d60710a22d674c21b5055fb6CAS |
Петик Д. В., Роу Дж. Б.
(1996 ) Влияние питания и физических упражнений на параметры туши и уровень гликогена в скелетных мышцах овец породы меринос.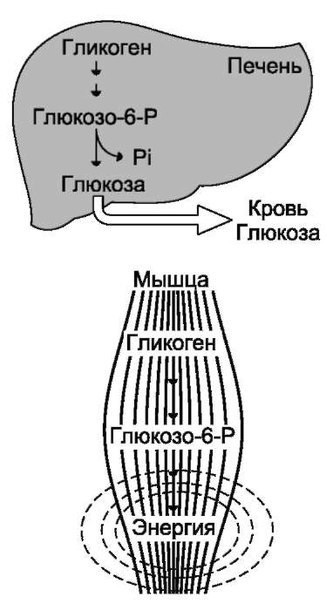 Австралийский журнал сельскохозяйственных исследований 47 , 525–537.
Австралийский журнал сельскохозяйственных исследований 47 , 525–537.
| Влияние питания и физических упражнений на параметры туши и уровень гликогена в скелетных мышцах мериносовой овцы. Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DyaK28XislKgs7k% 3D & md5 = 7fa6e2c694e102640fca98930aaCAS |
Петик Д.В., Линдси Д.Б., Баркер П.Дж., Нортроп А.Дж.
(1983 ) Метаболизм циркулирующих неэтерифицированных жирных кислот всем животным, задними конечностями и маткой беременных овец. Британский журнал питания 49 , 129–143.
| Метаболизм циркулирующих неэтерифицированных жирных кислот всем животным, задними конечностями и маткой беременных овец.Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DyaL3sXks1yju7o% 3D & md5 = eba1649c892438e3a317aecf04474230CAS | 6821682PubMed |
Петик Д.В., Миллер С.Б., Харман Н.Г.
(1991 ) Физические упражнения у овец породы меринос — взаимосвязь между интенсивностью работы, выносливостью, анаэробным порогом и метаболизмом глюкозы.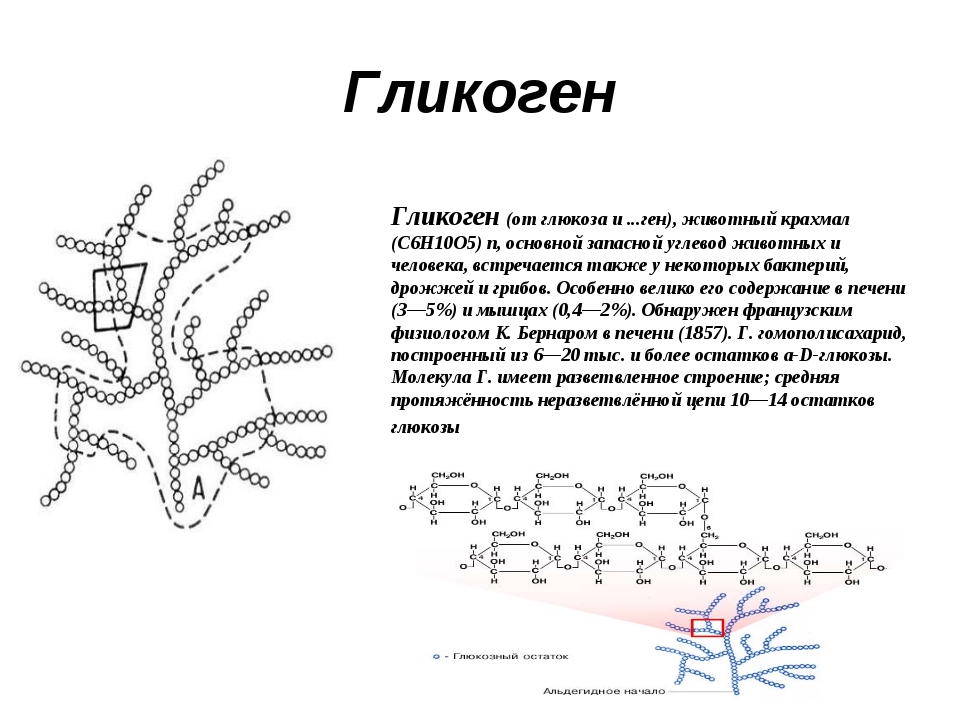 Австралийский журнал сельскохозяйственных исследований 42 , 599–620.
Австралийский журнал сельскохозяйственных исследований 42 , 599–620.
| Физические упражнения у овец мериноса — взаимосвязь между интенсивностью работы, выносливостью, анаэробным порогом и метаболизмом глюкозы. Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DyaK3MXktVGrurg% 3D & md5 = acbb7309bea181af71a99c379cdc7a9dCAS |
Петик Д.В., Камминз Л., Гарднер Г.Е., Джейкобс Р.Х., Колено Б.В., Макдауэлл М., Макинтайр Б.Л., Тюдор Дж., Уокер П.Дж., Уорнер Р.Д.
(2000 ) Регулирование уровня гликогена в мышцах жвачных животных с помощью питания. Труды Новозеландского общества животноводства 60 , 94–97.Петик Д.В., Харпер Дж., Даншеа Ф.Д. (2005) Метаболизм и обмен жиров. В «Количественные аспекты пищеварения и метаболизма жвачных животных». (Редакторы Дж. Дейкстра, Дж. М. Форбс, Дж. Франция) стр. 345–371. (Издательство CABI: Уоллингфорд, Великобритания)
Петтерсон Дж. А., Даншеа Ф. Р., Эрхардт Р. А., Белл А. В.
(1993 ) Беременность и недостаточное питание изменяют метаболические реакции глюкозы на инсулин у овец. Журнал питания 123 , 1286–1295.
Журнал питания 123 , 1286–1295.
Рагги Ф, Кронфельд Д.С., Кляйбер М
(1960 ) Активность глюкозо-6-фосфатазы в различных тканях овец. Труды Общества экспериментальной биологии и медицины 105 , 485–486.
| Активность глюкозо-6-фосфатазы в различных тканях овец. Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DyaF3MXksVSmsg% 3D% 3D & md5 = 7cfa7f56f935dc7aae5ec614c0aad7f9CAS | 13739125PubMed |
Рэндл П.Дж., Гарланд ПБ, Хейлз С.Н., Ньюсхолм Е.А.
(1963 ) Цикл глюкозы и жирных кислот. Его роль в чувствительности к инсулину и метаболических нарушениях при сахарном диабете. Ланцет 281 , 785–789.
| Жирно-кислотный цикл глюкозы. Его роль в чувствительности к инсулину и метаболических нарушениях при сахарном диабете.Crossref | GoogleScholarGoogle Scholar |
Рихтер Э., Гальбо Х.
(1986 ) Высокий уровень гликогена усиливает распад гликогена в изолированных сокращающихся скелетных мышцах. Журнал прикладной физиологии 61 , 827–831.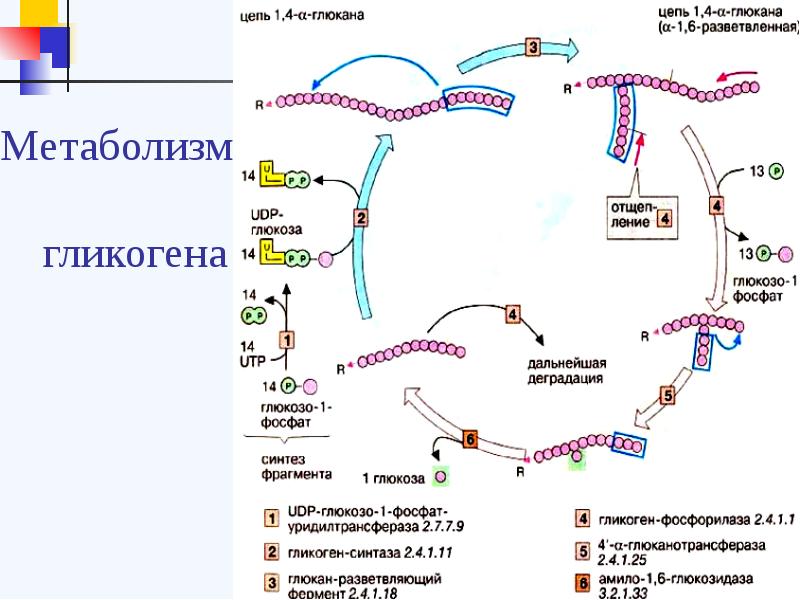
Roach PJ
(1990 ) Контроль гликогенсинтазы посредством иерархического фосфорилирования белков. Журнал FASEB 4 , 2961–2968.
Saltin B, Gollnick PD (1983) Адаптивность скелетных мышц: значение для метаболизма и производительности. В «Справочнике по физиологии, раздел 10, скелетные мышцы». (Редакторы Л. Д. Пичи, Р. Х. Адриан, С. Р. Гейгер) стр. 555–631. (Американское физиологическое общество: Bethesda, MD)
Smetana R, Huber K, Hartter E, Yang P, Meisinger V, Konnaris C, Gabriel H, Zehetgruber M, Klappacher G, Spona J, Vierhapper H (1995) Ишемическая болезнь сердца — взаимодействия магния и катехоламинов во время стрессовой нагрузки.В «Достижения в исследованиях магния: магний в кардиологии»
. (Ред. Р. Сметана) стр. 61–65. (John Libbey and Company Ltd: Лондон)
Stangassinger M, Geisecke D (1986) Спланхнический метаболизм глюкозы и связанных с ней энергетических субстратов. В «Контроль пищеварения и обмена веществ у жвачных животных». (Редакторы Л. П. Миллиган, В. Л. Гровум, А. Добсон) стр. 347–66. (Prentice-Hall: Englewood Cliffs, Нью-Джерси)
Л. Гровум, А. Добсон) стр. 347–66. (Prentice-Hall: Englewood Cliffs, Нью-Джерси)
Симондс Х.В., Мазер Д.Л., Коллис К.А.
(1981 ) Максимальная способность печени взрослой дойной коровы усваивать аммиак. Британский журнал питания 46 , 481–486.
| Максимальная способность печени взрослой дойной коровы усваивать аммиак.Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DyaL38XotlagtQ% 3D% 3D & md5 = c1f0a209d0b814f216e0173b243f5217CAS | 7317343PubMed |
Tarrant PV (1981) Возникновение, причины и экономические последствия темновой резки говядины — обзор текущей информации. В «Проблема темной нарезки говядины». (Редакторы DE Hood, PV Tarrant), стр.25–62. (Издательство Martinus Nijhoff: Лейден, Нидерланды)
Таррант П.В. (1988) Поведение животных и окружающая среда в условиях темной стрижки. В «Темной рубке крупного рогатого скота и овец»; труды австралийского семинара ». (Редакторы СУ Фабианссон, WR Shorthose, RD Warner) стр. 8–18. (Австралийская корпорация исследований и развития мяса и животноводства: Сидней)
Тюдор G, Coupar FJ, Pethick DW
(1996 ) Влияние силосных рационов на концентрацию гликогена в мышцах молодняка крупного рогатого скота. Труды Австралийского общества животноводства 21 , 451
Труды Австралийского общества животноводства 21 , 451
Ван ден Топ AM, Гилен MJH, Венсинг Т., Вентинк Г.Х., ВантКлоостер А.Т., Бейнен А.С.
(1996 ) Более высокие концентрации триацилглицерина в печени в послеродовом периоде у дойных коров со свободным, а не с ограниченным доступом корма в сухой период связаны с более низкой активностью глицеролфосфатацилтрансферазы в печени. Журнал питания 126 , 76–85.
Ванн Р.К., Бердик, Северная Каролина, Лион Дж. Дж., Уэльсский TH, Рэндел Р. Д.
(2010 ) Влияние темперамента крупного рогатого скота на гормоны стресса и концентрацию IgG у телят ангусского кросса. Journal of Dairy Science 93 , 464–465.
Висек WJ
(1984 ) Аммиак: его влияние на биологические системы, метаболические гормоны и репродуктивную функцию. Journal of Dairy Science 67 , 481–498.
| Аммиак: его влияние на биологические системы, метаболические гормоны и репродуктивную функцию. Crossref | GoogleScholarGoogle Scholar | 1: CAS: 528: DyaL2cXhs12guro% 3D & md5 = d176700be2034bfa8a1a37fff3cbdfe2CAS | 6371080PubMed |
Вуазине Б. Д., Грандин Т., О’Коннор С.Ф., Татум Д.Д., Дисинг М.Дж.
Д., Грандин Т., О’Коннор С.Ф., Татум Д.Д., Дисинг М.Дж.
(1997 )
Bos indicus — кросс-откормочный скот с возбудимым темпераментом, имеет более жесткое мясо и более высокий процент пограничных темных разделчиков. Meat Science 46 , 367–377.
|
Bos indicus — кросс-откормочный скот с возбудимым темпераментом, имеет более жесткое мясо и чаще встречается пограничная темная резка.Crossref | GoogleScholarGoogle Scholar | 1: STN: 280: DC% 2BC3MbnsVynsg% 3D% 3D & md5 = 89a7600a833a21f94e0800a6b3e04286CAS | 22062320PubMed |
Weekes TEC (1979) Углеводный обмен. В «Физиологии пищеварения и питании жвачных животных». (Ред. DC Church) стр. 187–209. (Компания O&B Books Inc.: Корваллис, Орегон)
Вегнер Дж., Альбрехт Э., Фидлер I, Тойшер Ф., Папштейн Х.-Дж., Эндер К.
(2000 ) Рост и связанные с породой изменения характеристик мышечных волокон крупного рогатого скота. Журнал зоотехники 78 , 1485–1496.
Рэй-Кахен Д., Даншеа ФР, Бойд Р.