Когда и сколько протеина нужно принимать для достижения максимальных результатов?
В различных исследованиях изучалось необходимое суточное количество белков и их влияние на организм. В основном применяли 1,5 г белка на 1 кг массы. Только в нескольких исследованиях изучалось потребление более высоких порций белка примерно от 2,2 до 3,3 г / кг. При этом ни одно исследование не выявило негативного влияния протеина на здоровье людей. [17] В таблице ниже вы найдете основное распределение ежедневного потребления белка в соответствии с основными факторами. [10]
В таблице указано, что если вы активный спортсмен, не страдаете от избыточного веса и хотите достичь мышечного роста, вы должны принимать от 1,4 до 3,3 г протеина на килограмм веса в день. Это довольно широкий диапазон, поэтому нужно добавить еще один фактор – массу тела.
Сочетание интенсивной тренировки и правильного количества протеина – это золотой путь к успеху. Ниже мы привели таблицу правильного потребления протеина, основанное массе тела и подходящее как для женщин, так и для мужчин.
Опираясь на таблицу мы можем видеть, что спортсмены и активные взрослые люди могут свести к минимуму рост жировых отложений, увеличив потребление протеина до 3,3 грамма на килограмм веса. [10] Таким образом, здоровый взрослый спортсмен, который хочет достичь мышечного роста и в настоящее время весит 91 кг, может принимать до 299 граммов протеина в день.
Исследования показали, что если вы постепенно нагружаете свой организм во время тренировки и придерживаетесь гиперкалорической диеты (употребляете дополнительно 370-800 ккал), вы потеряете больше жира, если принимаете протеин в количестве 3,3 г / кг, чем при принятии 1,8 – 2,6 г / кг.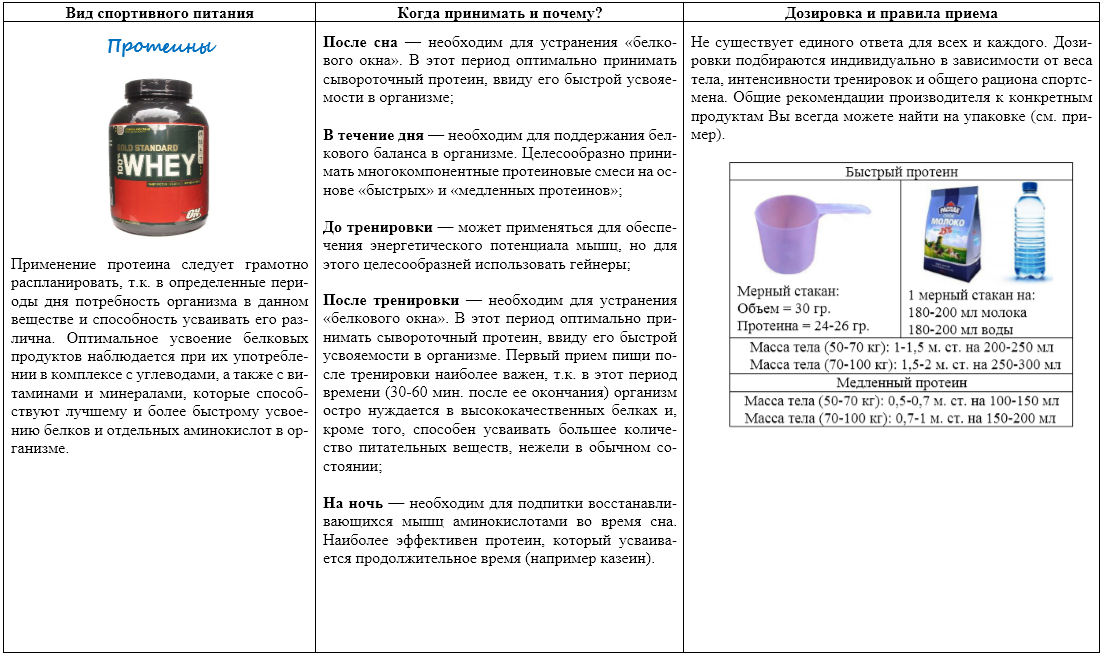 [18] [19] Следует отметить, что максимальная порция протеина не поможет нарастить больше мышц, чем минимальная порция. Тем не менее, это может свести к минимуму потребление жира из рациона питания, что поможет сбросить лишние килограммы.
[18] [19] Следует отметить, что максимальная порция протеина не поможет нарастить больше мышц, чем минимальная порция. Тем не менее, это может свести к минимуму потребление жира из рациона питания, что поможет сбросить лишние килограммы.
Также помните, что суточная доза белка должна основываться на весе тела, а не на потреблении калорий. Потребление калорий, наоборот, должно основыватся на весе и целях, которые вы для себя ставите.
Оптимальная дневная порция протеина при похудении
В одном из предыдущих исследований было установлено, что спортсмены должны употреблять около 1,8 – 2,7 грамм белка на килограмм массы. [20] Однако, недавние исследования показывают, что для того, чтобы минимизировать мышечную потерю во время диеты, спортсмены должны употреблять от 2,3 до 3,1 г протеина на килограмм массы. [21] Это значение также подтверждено Международным обществом спортивного питания (International Society of Sports Nutrition). [22]
[22]
Рекомендация, однако, применима только к относительно стройным людям, которые пытаются сформировать и укрепить свою фигуру. Некоторые исследования показали, что для людей с ожирением опритально подходит порция 1,2-1,5 г / кг, чтобы максимизировать потерю жира. [23] [24] [25] Европейская ассоциация по изучению ожирения (European Association for the Study of Obesity), также рекомендует принимать 1,5 г / кг протеина для взрослых людей, страдающих ожирением. [26]
Учитывая риски для здоровья, связанные с избыточным весом и ожирением, также интересно, что применение высокобелковых диет (около 27% ежедневного потребления калорий) может снизить кардиометаболические факторы риска, включая кровяное давление или уровень триглицеридов, а также помогает увеличить уровень холестерина ЛПВП. Но стоит признать, что эти эффекты наблюдались только в небольших количествах.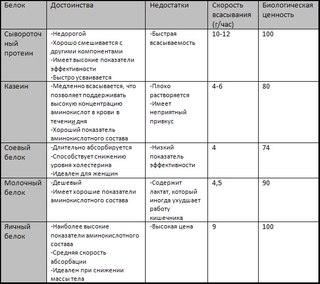 [27]
[27]
В таблице ниже вы найдете ежедневную порцию белка для здоровых взрослых и людей с избыточным весом. Значения разбиты по массе тела в килограммах. [10]
| Вес | При избыточном весе и ожирении | При здоровом весе | ||
|---|---|---|---|---|
| Минимальная порция | Максимальная порция | Минимальная порция | Максимальная порция | |
| 45 кг | 54 г | 68 г | 100 г | 150 г |
| 57 кг | 68 г | 85 г | 125 г | 187 г |
| 68 кг | 82 г | 102 г | 150 г | 225 г |
| 79 кг | 95 г | 119 г | 175 г | 262 г |
| 91 кг | 109 г | 136 г | 200 г | 299 г |
| 102 кг | 122 г | 153 г | 225 г | 337 г |
| 113 кг | 136 г | 170 г | 249 г | 374 г |
| 125 кг | 150 г | 187 г | 274 г | 412 г |
Помните, что вычисление суточной порции белка основано на вашей фактической массе тела, а не на идеальном весе, который вы хотите достичь. Поэтому, если вы находитесь на диете и хотите максимально уменьшить жировые отложения без потери мышечной массы, необходимо принимать 2,3 – 3,1 г / кг белка в день. Людям с избыточным весом или ожирением необходимо принимать 1,2-1,5 г / кг белка в течение дня. [10]
Поэтому, если вы находитесь на диете и хотите максимально уменьшить жировые отложения без потери мышечной массы, необходимо принимать 2,3 – 3,1 г / кг белка в день. Людям с избыточным весом или ожирением необходимо принимать 1,2-1,5 г / кг белка в течение дня. [10]
Могут ли белки быть вредными?
Возможно, вы слышали миф о том, что белки повреждают почки, вызывают печеночную недостаточность или остеопороз. Дело в том, что это утверждение не было подтверждено ни одним исследованием, и нет никаких доказательств негативного влияния белка в организме здоровых людей. [4] Конечно, если у вас проблемы с почками или печенью, вы должны проконсультироваться с врачом перед принятием белковых добавок.
Может ли чрезмерное потребление белка нанести вред вашему здоровью?
Использование чрезмерного количества протеина может сопровождать проблемы с пищеварением, такие как метеоризм или диарея. Поэтому необходимо употреблять оптимальную дозу протеина, разделенную на меньшие порции в течение дня.
Поэтому необходимо употреблять оптимальную дозу протеина, разделенную на меньшие порции в течение дня.
Сколько протеина стоит принимать после тренировки?
Были проведены многочисленные исследования в вопросе о том, сколько протеина стоит принимать в одной порции. Более старые исследования сошлись на том, что 20-25 грамм высококачественного протеина достаточно для максимизации белковой стимуляции в мышцах. [12] [13] [14]
Однако более новые исследования поставили под сомнение этот результат. Они ссылаются на то, что в предыдущих исследованиях не учитывалось количество чистой мышечной массы. Таким образом, человеку с более высокой долей чистой мышечной массы требуется более высокая пропорция аминокислот для максимизации мышечного роста и регенерации после тренировки. Эти исследования приходят к выводу, что синтез белка в мышцах повышается при употреблении 40 граммов сывороточного протеина после тренировки. [5]
[5]
В любом случае, чтобы ученые узнали о наиболее подходящем количестве белка в порции, необходимо большее количество исследований. В результате исследований, которые показали опримальную порцию в 20-25 г, проводилось исследование группы людей, которые тренировали только ноги. В то время как исследования, которые показали идеальную порцию в 40 г, основывались на группе людей, которые проводили комплексные тренировки всего тела. Поэтому показатели данных исследований не соответствуют друг другу. Также не стоит забывать, что повышенная мышечная активность во время тренировки требует большего количества аминокислот и белка для восстановления. [11]
В то же время мышечная ткань требует около 25-30% от общего потребления белка. Это означает, что потребление большего количества белка, не обязательно стимулирует рост мышц. Например, принятие порции в 70 г белка, окажет такое же влияние на рост мышц как и порция в 40 г белка. Однако, повышенное потребление белка не означает, что вы расходуете его впустую. [6] [15] Организм сохраняет белок в качестве источника энергии для дальнейших тренировок. [9]
Например, принятие порции в 70 г белка, окажет такое же влияние на рост мышц как и порция в 40 г белка. Однако, повышенное потребление белка не означает, что вы расходуете его впустую. [6] [15] Организм сохраняет белок в качестве источника энергии для дальнейших тренировок. [9]
Также во время проведения одного исследования ученые получали разные результаты, так как потребление белка для каждого человека зависит от индивидуальных потребностей. Например от возраста, веса, интенсивности тренировки, спортивных целей и уровня подготовки. Независимо от того, сколько составляет разовая порция белка (20 или 40 г), важно разделить потребление белка в течение дня – до 5 раз. Что поможет постоянно стимулировать синтез белка в организме. [8]
Когда лучше всего принимать протеин?
Определенное время приема протеина так же важно, как и правильная суточная порция. Как было сказано ранее, это зависит от ваших спортивных целей и текущего веса. Спортсменам необходимо регулярно получать белки в течении дня, в количестве 20-40 г. Эффективным решением является сочетание подходящего белка и времени. Оптимально разделить прием на 5 этапов – утром, днем, вечером, до тренировки и после тренировки.
Как было сказано ранее, это зависит от ваших спортивных целей и текущего веса. Спортсменам необходимо регулярно получать белки в течении дня, в количестве 20-40 г. Эффективным решением является сочетание подходящего белка и времени. Оптимально разделить прием на 5 этапов – утром, днем, вечером, до тренировки и после тренировки.
1. Утром
Исследования показывают, что организм наиболее чувствителен к воздействию аминокислот из белка через 24 часа после тренировки. [36] Высококачественный, быстро усваеваемый сывороточный белок является прекрасным началом нового дня и идеально подходит для мышц. Однако, если вы хотите полностью заменить свой завтрак протеином, то лучше использовать казеин. [37]
2. Днем
Чтобы получить желаемое количество белка в день, желательно использовать в качестве перекуса протеиновый коктейль, батончики или другие протеиновые снеки.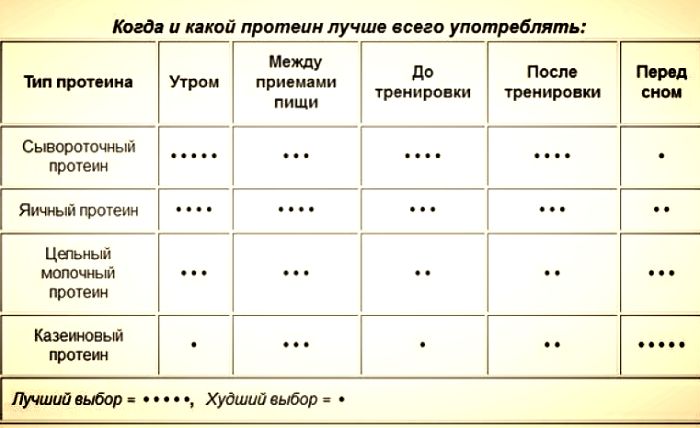 Добавление белка также можно осуществить с помощью протеинового напитка или природных источников белка.
Добавление белка также можно осуществить с помощью протеинового напитка или природных источников белка.
3. Вечером, перед сном
Протеиновый коктейль перед сном может быть очень эффективен для сжигания жиров, а также для увеличения силы и мышечной массы. В этом случае, наиболее подходящим выбором являются казеины, также называемые ночными протеинами, которые медленно усваиваются, обеспечивая мышцы белком пока вы спите. [33] [34] [35]
4. Перед тренировкой
При употреблении протеинового напитка перед тренировкой, вы получите тот же эффект, что и при употреблении после тренировки. [32] Быстро поглощаемый белок поможет вам восстановить мышечные волокна во время тренировки. Для употребления перед тренировкой лучше всего подойдет сывороточный протеин.
При этом, отличной альтернативой протеину перед тренировкой является BCAA (лейцин, изолейцин, валин). Лейцин – незаменимая аминокислота, которая обладает способностью стимулировать клетки, ответственные за рост мышц. [2]BCAA в растворимой форме следует принимать за 15 минут до тренировки, в форме таблеток за 35 – 40 минут до тренировки.
Лейцин – незаменимая аминокислота, которая обладает способностью стимулировать клетки, ответственные за рост мышц. [2]BCAA в растворимой форме следует принимать за 15 минут до тренировки, в форме таблеток за 35 – 40 минут до тренировки.
5. После тренировки
Тяжелые тренировки стимулируют мышечный синтез, но также увеличивают риск повреждения мышечного аппарата. Потребление протеина сразу после тренировки поможет нарастить мышцы и способствует их регенирации. Незаменимая аминокислота – лейцинн обладает способностью стимулировать клетки, ответственные за рост мышц. [2] Поэтому после тренировки рекомендованно употреблять сывороточный протеин.
Нет точного времени, в котрое необходимо употреблять протеин после тренировки, однако существует такое понятие, как анаболическое окно, которое занимает 30-45 минут после тренировки, соответственно лучше всего употреблять белок в это время. [16] Но даже если принять порцию протеина позже, это тоже обеспечит необходимый результат. В любом случае, необходимо принимать дозу белков после тренировки, чтобы дать мышцам аминокислоты, необходимые для роста и регенерации. [28] [29] [30] [31] При выборе подходящего протеина после тренировки, следует помнить, что скорость поглощения зависит от типа обработки конкретного протеина.
Потребление белка для вегетарианцев и веганов
Вегетарианцы и веганы должны обратить внимание на то, сколько белка и из какого источника они получают. Только соя и киноа содержат все 9 незаменимых аминокислот, которые необходимы для синтеза белка в организме. Остальные веганские и вегетарианские источники белка недостаточны для организма, поэтому часто такие протеины являются многокомпонентными, для обеспечения полного аминокислотного профиля. Ежедневная порция белка, варьирующаяся от 1,5 до 2 г на килограмм веса, наряду с богатой порцией бобовых и злаковых культур, может обеспечить вегетарианцев и веганов достаточным количеством белка для наращивания и регенерации мышечной ткани.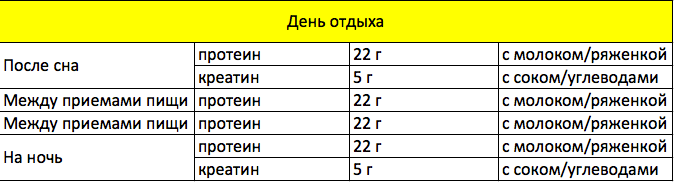 [2]
[2]
Узнали ли вы все необходимое о дозировке протеина и времени употребления? Напишите нам в комментариях, что из статьи вы знали ранее, а что прочитали впервые. Если вам понравилась статья, поделитесь ею с друзьями.
Источники:
[3] Bauer J, Biolo G, Cederholm T, Cesari M, ruz-Jentoft AJ, Morley JE, Philips S, Sieber C, Stehle P, Teta D, Visvanathan R, Volpi E, Boirie Y – Evidence-based recommendations for optimal dietary protein intake in older people: a position paper from the PROT-AGE Study Group. – https://www.ncbi.nlm.nih.gov/pubmed/23867520
[5] Macnaughton LS, Wardle SL, Witard OC, McGlory C, Hamilton DL, Jeromson S, Lawrence CE, Wallis GA, Tipton KD – The response of muscle protein synthesis following whole-body resistance exercise is greater following 40 g than 20 g of ingested protein. – https://www.ncbi.nlm.nih.gov/pubmed/27511985
[8] Mamerow MM, Mettler JA, English KL, Casperson SL, Arentson-Lantz E, Sheffield-Moore M, Layman DK, Paddon-Jones D – Dietary protein distribution positively influences 24-h muscle protein synthesis in healthy adults. – https://www.ncbi.nlm.nih.gov/pubmed/24477298
– https://www.ncbi.nlm.nih.gov/pubmed/24477298
[9] Symons TB, Sheffield-Moore M, Wolfe RR, Paddon-Jones D – A moderate serving of high-quality protein maximally stimulates skeletal muscle protein synthesis in young and elderly subjects. https://www.ncbi.nlm.nih.gov/pubmed/19699838
[13] Witard OC, Jackman SR, Breen L, Smith K, Selby A, Tipton KD – Myofibrillar muscle protein synthesis rates subsequent to a meal in response to increasing doses of whey protein at rest and after resistance exercise.- https://www.ncbi.nlm.nih.gov/pubmed/24257722
[19] Antonio J, Ellerbroek A? Silver T, Orris S, Scheiner M, Gonzalez A, Peacock CA – A high protein diet (3.4 g/kg/d) combined with a heavy resistance training program improves body composition in healthy trained men and women–a follow-up investigation. – https://www.ncbi.nlm.nih.gov/pubmed/26500462
[21] Helms ER, Zinn C, Rowlands DS, Brown SR – A systematic review of dietary protein during caloric restriction in resistance trained lean athletes: a case for higher intakes. – https://www.ncbi.nlm.nih.gov/pubmed/24092765
– https://www.ncbi.nlm.nih.gov/pubmed/24092765
[22] Aragon AA, Schoenfield BJ, Wildman R, Kleiner S, VanDusseldorp T, Taylor L, Earnest CP, Arciero PJ, Wilborn C, Kalman DS, Stouz JR, Willoughby DS, Campbell B, Arent SM, Bannock L, Smith-Ryan AE, Antonio J – International society of sports nutrition position stand: diets and body composition. – https://www.ncbi.nlm.nih.gov/pubmed/28630601
[23] Kim JE, O Connor LE? Sands LP, Slebodnik MB, Campbell WW – Effects of dietary protein intake on body composition changes after weight loss in older adults: a systematic review and meta-analysis. https://www.ncbi.nlm.nih.gov/pubmed/26883880
[24] Wycherley TP, Moran LJ, Clifton PM, Noakes M, Brinkworth GD – Effects of energy-restricted high-protein, low-fat compared with standard-protein, low-fat diets: a meta-analysis of randomized controlled trials. – https://www.ncbi.nlm.nih.gov/pubmed/23097268
[25] Krieger JW, Sitren HS, Daniels MJ, Langkamp-Henken B, – Effects of variation in protein and carbohydrate intake on body mass and composition during energy restriction: a meta-regression 1. – https://www.ncbi.nlm.nih.gov/pubmed/16469983
– https://www.ncbi.nlm.nih.gov/pubmed/16469983
[27] Santesso N, Akl EA, Bianchi M, Mente A, Mustafa R, Heels-Andell D? Schunemann HJ – Effects of higher- versus lower-protein diets on health outcomes: a systematic review and meta-analysis. – https://www.ncbi.nlm.nih.gov/pubmed/22510792
[28] Bart Pennings, Rene Koopman, Milou Beelen, Joan MG Senden, Wim HM Saris, Luc JC van Loon – Exercising before protein intake allows for greater use of dietary protein–derived amino acids for de novo muscle protein synthesis in both young and elderly men – https://academic.oup.com/ajcn/article/93/2/322/4597647
[29] Tieland M, Dirks ML, van der Zwaluw N, Verdijk LB, van de Rest O, de Groot LC, van Loon LJ – Protein supplementation increases muscle mass gain during prolonged resistance-type exercise training in frail elderly people: a randomized, double-blind, placebo-controlled trial. – https://www.ncbi.nlm.nih.gov/pubmed/22770932
[34] Tim Snijders, Peter T Res, Joey SJ Smeets, Stephan van Vliet, Janneau van Kranenburg, Kamiel Maase, Arie K Kies, Le B Verdijk, Luc JC van Loon – Protein Ingestion before Sleep Increases Muscle Mass and Strength Gains during Prolonged Resistance-Type Exercise Training in Healthy Young Men – https://academic. oup.com/jn/article/145/6/1178/4644372
oup.com/jn/article/145/6/1178/4644372
[35] Tim Snijders, Peter Res, Joey SJ Smeets, Stephan van Vliet, Janneau van Kranenburg, Kamiel Maase, Arie K Kies, Lex B Verijk, Luc JC van Loon – Protein Ingestion before Sleep Increases Muscle Mass and Strength Gains during Prolonged Resistance-Type Exercise Training in Healthy Young Men – https://academic.oup.com/jn/article/145/6/1178/4644372
[36] Burd NA, West DW, Moore DR, Atheton PJ, Staples AW, Prior T, Tang JE, REnnie MJ, Baker SK, Phillips SM – Enhanced amino acid sensitivity of myofibrillar protein synthesis persists for up to 24 h after resistance exercise in young men. – https://www.ncbi.nlm.nih.gov/pubmed/21289204
Как принимать протеин. Спортивные советы ЗонаСпорта
Протеин — одна из самых популярных добавок в спортивном питании для набора мышечной массы и удержания мышц при сбросе веса. Протеин на 70-90% состоит из белка, остальную массу составляют углеводы, немного полезных жиров, возможно наличие дополнительных компонентов, по типу витаминов и минералов.
Протеин — одна из самых популярных добавок в спортивном питании для набора мышечной массы и удержания мышц при сбросе веса. Протеин на 70-90% состоит из белка, остальную массу составляют углеводы, немного полезных жиров, возможно наличие дополнительных компонентов, по типу витаминов и минералов. Протеин, несмотря на свое натуральное происхождение, не может заменить белки, получаемые из обычной пищи, именно поэтому он принимается в качестве добавки к основному рациону, а не замены к нему.
Как принимать протеин?
У атлетов суточная потребность в белках значительно возрастает, так как им нужно не только “заполнить” суточную норму белка, но и получить излишек для наращивания мышечных тканей.
Протеин необходимо пить каждый день и в одинаковом количестве. Но сама дозировка протеина должна быть сугубо индивидуальной, так как производители берут достаточно усредненные значения потребности белка. Количество принимаемого протеина зависит от процентного содержания в нем белка, от количества белка, получаемого из пищи, и от веса спортсмена. Для роста мышц человек должен потреблять минимум 1,5 грамма белка ка килограмм веса в сутки, а в идеале — 2. При обычном питании человек в лучшем случае получает только 1 грамм белка. Исходя из этого нужно просчитать суточное количество белка, потребляемого из обычной пищи, и восполнить недостающее протеиновым коктейлем.
Для роста мышц человек должен потреблять минимум 1,5 грамма белка ка килограмм веса в сутки, а в идеале — 2. При обычном питании человек в лучшем случае получает только 1 грамм белка. Исходя из этого нужно просчитать суточное количество белка, потребляемого из обычной пищи, и восполнить недостающее протеиновым коктейлем.
Количество суточной нормы протеина (граммы)
Протеиновый коктейль готовится очень легко: смесь просто размешивают любом количестве воды, молока или сока до нужной консистенции. Единственное — нельзя размешивать смесь в кипятке, так как в нем белок теряет свои свойства.
Дневную дозу протеина лучше выпить в несколько приемов в течении дня. Можно, конечно, принять его и за раз, но тут сложность заключается в том, что часть белка просто не сможет усвоиться. Рассчитывайте количество приемов примерно так, чтобы в на один коктейль приходилось не более 30 грамм белка (получается 2-3 коктейля в сутки).
Схема приема может также отличаться в зависимости от целей употребления протеина:
Цель — набор мышечной массы
Тогда протеин принимается между основными приемами пищи, то есть как бы дополняет основной рацион.
Также в таком случае важен будет прием коктейля сразу после тренировки — это важный этап восстановления и наращивания мышц.
Цель — похудение
В таком случае протеиновым коктейлем нужно заменить некоторые основные приемы пищи, например, завтрак и перекус.
Это необходимо для того, чтобы наполнить организм белком, но не повышать общую калорийность рациона.
Прием протеина никак не зависит от его происхождения: растительные и животные белки принимаются по одинаковой, описанной выше схеме.
Сочетаемость протеина с другими спортивными добавками
Многих также интересует сочетаемость протеина с другими пищевыми добавками: гейнером, аминокислотами и креатином.
Гейнер — высококалорийная белково-углеводная смесь, которая принимается спортсменами худощавого телосложения для быстрого набора мышечной массы.
Гейнер содержит в среднем от 50 до 85% углеводов разной сложности, поэтому также используется в качестве источника энергии для занятий. Для процесса роста мышц одинаково необходимы как белки, так и углеводы: белки выступают материалом для создания новых тканей, а углеводы — дают энергию для этого процесса. Равноценно наполнить организм белками и углеводами можно с помощью гейнеров с высоким содержанием белка (30-35%), а можно приготовить собственную смесь из гейнера и протеина, где будет учтена индивидуальная потребность в белках.
Лучше готовить такую смесь из высокоуглеводного гейнера, тогда легче будет просчитать количество белка в коктейле.
Аминокислоты — это составляющие белков, поэтому у них тот же принцип действия: восстановление структуры мышц и наращивание мышечной массы.
Поэтому если вы уже принимаете протеин, то по сути смысла в приеме еще и аминокислот нет никакого. Но если все-таки хочется усилить действие протеина, то можно дополнить его приемом аминокислот BCAA. Прием обеих добавок можно объединить, лучше всего утром и после тренировки.
Креатин — кислота, которая принимает участие энергетическом обмене в мышечных клетках.
Сочетание креатина с протеином считается очень эффективным для набора мышечной массы: креатин наполнит мышцы энергией, а протеин выступит в качестве материала для их наращивания. Креатин и протеин можно принимать отдельно, а можно вместе. Для удобства креатин просто добавляют в протеиновый коктейль.
Получается, что в приеме протеина важно не время, а количество, потому что тут ключевой является возможность предоставить организму белок, а он “возьмет” столько, сколько ему нужно. Очень важно придерживаться нужной конкретно Вам дозировки, не уменьшать ее и не увеличивать, иначе может потеряться весь эффект. Цена качественного протеина часто превышает стоимость других спортивных добавок, например, гейнеров, тем не менее, прием протеиновых коктейлей — эффективный вариант для наращивания мышц.
Цена качественного протеина часто превышает стоимость других спортивных добавок, например, гейнеров, тем не менее, прием протеиновых коктейлей — эффективный вариант для наращивания мышц.
Как правильно принимать протеин | Блог
Протеин — это одна из важнейших добавок, необходимая для спортсменов желающих нарастить мускулатуру. Связано, это с тем, что он содержит до 95 % белка в своем составе, а белок в свою очередь используется нашим организмом для построения мышечной ткани. Принимают их, как правило, в виде коктейлей, которые довольно легко приготовить в домашних условиях. В состав таких коктейлей входит: протеин, молоко или вода и различные вкусовые добавки, например — мелкопорезанные фрукты.
Наиболее благоприятное время для усвоения белка приходится на утро, на время — сразу после сна. Прием протеина на ночь необходим для подпитки восстанавливающихся мышц аминокислотами во время сна, в такое время наиболее эффективен казеин, который усваивается продолжительное время.
На усвояемостью белковых добавок влияют два гормона: тиреоидный гормон и гормон роста. Всплеск выделения организмом этих гормонов, приходится на фазу быстрого сна, т.е время в течении полутора часов после засыпания. Таким образом, наиболее благоприятное время для приема приходится на утро, время в течении получаса после тренировки, когда открывается так называемое «протеиновое окно» и непосредственно перед сном.
Почему мы не рекомендуем принимать протеин непосредственно перед тренировкой? Связано это с тем, что нашему организму, для того что-бы усвоить белок и расщепить его на аминокислоты необходима энергия, которая после приема коктейля будет затрачена из внутренних резервов организма. И по этому, прием протеина может вызвать небольшую сонливость, что крайне не желательно перед тренировкой. Если вы хотите принять протеин перед тренировкой, то рекомендуем дополнить такой прием углеводами, например съесть 2-3шт банана.
Перед тренировкой можно принимать гейнеры, которые кроме белка содержат еще и углеводы. Углеводы из гейнера, принятого перед тренировкой, повысят запасы гликогена в ваших мышцах.
Углеводы из гейнера, принятого перед тренировкой, повысят запасы гликогена в ваших мышцах.
При принятии протеинового коктейля после тренировки, в работавшие и поврежденные микротравмами мышцы стягиваются аминокислоты, которые начинают процессы заживления микро травм мышечных волокон. Именно это время и принято называть «протеиновым окном», здесь прием легкоусвояемого белка будет как раз кстати.
Обобщенная таблица по применению протеина в течении дня в тренировочные и не тренировочные дни:
| Тренировочные дни: | Время | Дни отдыха: |
|
Натощак, порцию 25-40 грамм сывороточного белка |
Утро |
Натощак, порцию 25-40 грамм сывороточного белка |
|
В течении 30 минут после тренировки 30-50 грамм сывороточного белка |
День |
В течении дня «долгий» казеин или комбинированный протеин 30-50 грамм |
|
Перед сном, протеин с долгим временем усвоения — казеин 25-40 грамм |
Ночь |
Перед сном, протеин с долгим временем усвоения — казеин 25-40 грамм |
В каком количестве принимать протеин?
Ежедневная норма белка, необходимая для мышечного роста, это 2-2,5 грамма на 1 кг собственного веса. Следовательно, для спортсмена весом 90 кг суточная норма белка около 200 граммов! Если же спортсмен будет принимать всего 100-150 грамм белка, то он не сможет нарастить мышечной массы больше, чем у него сейчас есть, принимаемое количество протеина будет расходоваться на поддержание мышечной массы.
Следовательно, для спортсмена весом 90 кг суточная норма белка около 200 граммов! Если же спортсмен будет принимать всего 100-150 грамм белка, то он не сможет нарастить мышечной массы больше, чем у него сейчас есть, принимаемое количество протеина будет расходоваться на поддержание мышечной массы.
Под словосочетанием «ежедневная норма белка» — необходимо понимать суммарное количество принимаемой белковой пищи из вашего обычного пищевого рациона и от белка, получаемого из протеиновых коктейлей.
При этом, норма обычной пищи и протеиновых коктейлей в вашем рационе должна составлять 50% / 50 %. То-есть, тот-же спортсмен весом 90кг должен получать из пищи 100 грамм белка и из протеиновых коктейлей также 100 грамм белка. Исходя из этих простых расчетов, вы для себя определяете нужное количество приемов протеиновых коктейлей в течении дня.
Мы изложили обобщенную информацию о применении протеина, которая основывается на личном опыте наших сотрудников и многих клиентов нашего магазина. Если у вас есть что добавить к этой статье, то вы можете оставить свои комментарии внизу статьи. Искренне надеемся, что теперь вопрос — как принимать протеин, для вас полностью исчерпан и вы легко сможете спланировать ваш рацион таким образов, что бы получить максимум эффекта от приема белковых добавок.
Если у вас есть что добавить к этой статье, то вы можете оставить свои комментарии внизу статьи. Искренне надеемся, что теперь вопрос — как принимать протеин, для вас полностью исчерпан и вы легко сможете спланировать ваш рацион таким образов, что бы получить максимум эффекта от приема белковых добавок.
Как правильно пить протеин для набора мышечной массы
Рекомендуемые товары
-
Протеин, витамины и минералы
(Порошок 360г)
Больше энергии, сил и здоровья! Это смесь для приготовления диетического коктейля, который содержит сбалансированный состав соевого белка, витаминов, минералов, аминокислот. Подходит для мужчин и женщин, ведущих активных образ жизни, спортсменов, которые следят за здоровьем. Протеин коктейль способствует быстрому восстановлению сил, защищает от авитаминоза, дает энергию, полезен для кожи, волос, нормализует вес.
В корзину
2 900 р.
Если вы активно занимаетесь силовым спортом, качаете мышцы, то рано или поздно встает вопрос — каким специальным питанием поддержать этот процесс. Какой протеин пить и как это делать правильно, в каких дозах? Не вредит ли он здоровью? Об этом сейчас расскажем!
Какой протеин пить и как это делать правильно, в каких дозах? Не вредит ли он здоровью? Об этом сейчас расскажем!
Как и зачем принимать протеин
Начнем с того, что протеином называется спортивное питание, добавка, которая бывает разных видов. Каждому тренирующемуся рекомендуется подбирать свой, индивидуальный вид. В целом, протеин это важнейший белок, «строительный материал» для мышечной ткани, которые восполняет недостаток в организме.
По результатам спортивных исследований была выведена следующая дозировка: 1,5 гр протеина на 1 кг массы тела. Это суточная норма, которая необходима для роста мышц. Как употреблять протеин, если нагрузки каждый день очень большие, тренировки интенсивные? В этом случае дозировку можно увеличить на 0,5 гр. Главная задача — не допускать истощения организма, не нарушать процесс формирования мышц.
Можно ли «набрать» необходимое количество белка с пищей? К сожалению, это сделать сложно! Придется есть огромные порции, которые будут вызывать нагрузку на пищевод и желудок, и тяжесть в животе. Протеиновая добавка — это современное решение проблемы, регулярное пополнение белкового баланса при активных тренировках.
Протеиновая добавка — это современное решение проблемы, регулярное пополнение белкового баланса при активных тренировках.
Виды протеина
Существует несколько разновидностей, исходя из исходных компонентов добавки: быстрый (сывороточный), медленный (казеиновый). Первый вид быстро и практически полностью усваивается организмом, второй — дольше и хуже. Некоторые производители предлагают и комбинированные виды, которые сбалансированы по составу.
- «Быстрые» белки»- сывороточный, который быстро расщепляется, усваивается, имеет гипоаллергенные качества. В его составе незаменимые аминокислоты для организма. Злоупотреблять и превышать дозу «быстрых» белков нельзя! Суточная доза для мужчин — 340 гр, для женщин 300 гр.
- Медленно усваиваемые вещества — казеиновая добавка, обычно получают из молока. Усваивается долго, но хорошо сжигает жировые отложения, уменьшает чувство голода, наращивает мышечную массу. Еще одно важное качество казеина — он аккумулирует белковые запасы в человеческом организме.

Какая оптимальная доза белка
Прежде чем вы узнаете, как пить протеин для набора мышечной массы, мы расскажем вам об оптимальной дозировке белка. В ходе испытаний, которые проходили канадские спортсмены, было определено, что 20-30 гр белка в день составляют нужную дозу. В этом случае происходит максимальная стимуляция мышц. Важный нюанс! Не превышайте дозу, не пытайтесь употреблять более 35 г белка, чтобы быстрее набрать массу. Лишний белок просто не усвоится организмом, и он не пойдет впрок.
Как связать количество белка и протеина? Смотрите нашу табличку:
| Протеин(граммы) | Кол-во принятых белков (граммы) |
| 0,05 | |
| 10 | 0,08 |
| 20 | 0,11 |
| 30 | 0,11 |
| 40 | 0,12 |
Ориентируйтесь на эти данные и ваш организм всегда будет получать нужное (а не лишнее!) количество белка. Далее вы узнаете, как употреблять протеин в зависимости от времени суток и характера нагрузок.
Далее вы узнаете, как употреблять протеин в зависимости от времени суток и характера нагрузок.
Протеин утром до еды или после
В какое время дня оптимально принимать белок? Можно ли употреблять протеин с утра и стоит это делать натощак? Сначала вспомним, что обычно едят на завтрак спортсмены. Это блюда из яиц (яичница, омлет, отварное), которые хорошо усваиваются организмом. Но не всегда два, три яйца дадут нужное количество белка. Поэтому на завтрак можно предпочесть протеиновый коктейль, например с молочным белком. Что это даст? Во-первых, подкрепит организм в начале дня, во-вторых, пополнит баланс белка перед тренировками. Полезно также дополнить завтрак овсяной кашей, мюслями или другой зерновой или растительной пищей.
Протеин перед тренировкой
Если утром вы снабдили свой организм белком, то перед утренним занятием уже нет особой необходимости принимать протеин. Если тренировка проходит после обеда, то стоит позаботиться о том, чтобы еда была питательной и полноценной. В этом случае пить протеин тоже не нужно. Благодаря хорошему обеду, у вас будут силы, энергия активно качаться и заниматься.
В этом случае пить протеин тоже не нужно. Благодаря хорошему обеду, у вас будут силы, энергия активно качаться и заниматься.
Если же вместо обеда был «перекус» на бегу или после него прошло больше 3-х часов, то хорошо выпить белковый коктейль перед тренировкой. Тогда вы не будете качаться на голодный желудок и сжигать мышечную массу. Коктейль поможет скорейшему процессу восстановления белкового баланса.
Протеин после тренировки
Пить или не пить белковый коктейль сразу после занятия? Ответ однозначный — пить! Это самое подходящее время, чтобы ускорить синтез белка и рост мышечной массы. В течение первых 2-х часов после тренировки важно принять протеин, ведь мышцы в это время особенно чувствительны к аминокислотам и белку. В это время рекомендуется принимать сывороточный протеин, который быстро усваивается и ускорит мышечный рост. Не голодайте после тренировки! Это не полезно для ваших мышц!
Прием протеинов перед сном
Как «подкрепить» себя ночью? Хорошим вариантом будет употребление протеинового коктейля перед сном! Это полезно и будет питать ваши мышцы всю ночь, предотвращая распад белка. Полезнее в этом случае казеиновый протеин, который в желудке превратится в гелевую массу и будет медленно усваиваться организмом, аминокислоты будут поступать в кровь на протяжении всей ночи.
Полезнее в этом случае казеиновый протеин, который в желудке превратится в гелевую массу и будет медленно усваиваться организмом, аминокислоты будут поступать в кровь на протяжении всей ночи.
Подытожим, как пить протеин для набора мышечной массы:
- утром, с завтраком;
- в обед, если еда была неполноценной или порция маленькой;
- после тренировки;
- перед сном, на ночь;
Такая схема приема считается наиболее сбалансированной и полезной для активно тренирующихся людей.
Есть ли противопоказания?
Сам по себе любой протеин это белок, который очень нужен организму. Но учитывайте важные правила:
- не превышайте рекомендуемую дозу приема;
- если есть проблемы с почками, ЖКТ, то перед началом приема протеина лучше посоветоваться с врачом;
- если есть хронические болезни, то тоже важно спросить совета врача;
- если после приема наблюдается аллергия, стоит отказаться от этого протеина и попробовать другой вид.

Важно! Внимательно читайте инструкцию к добавке и анализируйте ее состав. Бывает так, что у человека возникает индивидуальная реакция на один из компонентов. Наблюдайте за собой и отслеживайте реакции организма на определенный протеиновый продукт
Заключение
Теперь вы знаете, как пить протеин для набора мышечной массы и можете делать это правильно! Только не стоит думать, что белковый коктейль может заменить активные тренировки. Здесь важен баланс спорта и питания, тогда результаты будут отличными!
правило употребления для роста мышц
Белок, который требуется организму, особенно необходим при желании набрать мышечную массу. Большая его часть содержится в протеине, который является основой спортивного питания. Но употреблять его нужно правильно. Давайте разберемся, как принимать протеин для набора мышечной массы.
Большая его часть содержится в протеине, который является основой спортивного питания. Но употреблять его нужно правильно. Давайте разберемся, как принимать протеин для набора мышечной массы.
Протеин следует смешивать с водой, соком или молоком. Важно следить за тем, чтобы жидкость была теплой или холодной, но не горячей, иначе белок свернется и потеряет свои свойства.
Приготовление протеиновой смеси с молоком. Вкусно, быстро и полезно, особенно для набора мышечной массы.
Периоды для употребления протеиновой смеси
Вопрос, когда лучше пить протеин, волнует многих спортсменов. Наиболее эффективен прием данной добавки в несколько раз. Давайте рассмотрим, когда следует употреблять протеин.
До начала тренировки
Для организма перед физическими нагрузками требуется белок, который надо употреблять с продуктами питания минимум за два часа до занятия, а также сложные углеводы. Если при приеме пищи не удалось получить нужное его количество, то имеется смысл употреблять за сорок минут до тренировки протеиновый коктейль, который сбалансирует рацион и запустит восстановительные и обменные процессы. В тот момент, когда мышцы начнут получать повреждения, протеин активизируется, заживляя травмы.
В тот момент, когда мышцы начнут получать повреждения, протеин активизируется, заживляя травмы.
Протеин и спорт — путь к идеальному телу. Он важен как для набора мышечной массы, так и для поддержания мышц в тонусе при похудении
После окончания тренировки
По окончании силовых нагрузок начинается голодание тела, и ему требуется подпитка энергией, и именно в этот временной промежуток мышцы поглощают все вещества, как губка, в ускоренном режиме. Наилучшим вариантом является принятие удвоенной дозы протеиновой смеси для набора мышечной массы совместно с углеводами простой формы. А через час нужно съедать полноценную еду. Это простое правило приема протеиновой смеси.
Прием протеинового порошка на ночь
Многие говорят, что есть на ночь вредно, но это не так. Белок — можно и даже нужно. Это обусловлено тем, что углеводы требуются для энергии, а принятые перед сном, они употребляются не для роста мышц, а больше для их восстановления. Протеиновая добавка будет поддержать уровень необходимого организму элемента — белка — на высоком уровне на протяжении всего периода.
Важно! Лучшим для приема перед тем, как лечь спать, является казеиновый протеин.
Употребление протеина по утрам
Давайте разберемся, как пить протеин в данное время суток. Завтрак является основным временем приема еды, так как после сна человеческий организм всегда готов к осуществлению «дозаправки». Но необязательно это будет протеин в виде коктейля. Лучше остановить свой выбор на обычной полноценной еде, богатой белком. Например, 2-3 яйца — это отличный протеиновый заряд.
Во время тренировки
Это плохая идея. Съедая в момент тренировки что-либо, человек активирует процесс, при котором кровь начинает стремиться к желудку с целью переваривания и усваивания продукта, вместо того чтобы поступить в мышцы с целью обеспечения их нутриентами, принятыми до начала занятия. Если хотите есть во время тренировки, и вам не нужно худеть, например, подкрепитесь бананом.
Ранее мы писали о том как принимать сывороточный протеин для набора и похудения и рекомендовали добавить эту статью в закладки.
Важно! Самым лучшим вариантом является прием белковой смеси для роста мышц в течение всего дня, но не во время тренировки.
Дозировка употребления разных типов протеина в разные промежутки времени. Для достижения желаемого результата следует придерживаться этих рекомендаций.
Как можно увидеть, имеется множество промежутков времени, когда следует и можно употреблять добавку. Интересен вопрос, как принимать для начинающих протеин, и есть ли разница в дозах с для них и для профессионалов.
Читатели считают данные материалы полезными:
- Основное назначение казеинового протеина: польза и вред
- Виды протеина: что выбрать для эффективного набора мышечной массы
Дозировка
Сколько принимать протеина — это вопрос, волнующий многих. Давайте разберемся в его тонкостях. Если целью тренировки является набор мышечной массы, то спортсмены должны осуществлять прием протеина в количестве 3 грамма на один килограмм веса. При этом правила приема диктуют необходимость деления всей дневной дозы на шесть равных частей.
При этом правила приема диктуют необходимость деления всей дневной дозы на шесть равных частей.
Важно! Если белка из полноценного рациона недостаточно, то протеиновые порошки станут отличным дополнением.
Например, творог является хорошим источником белка. Если в шейкер с молоком положить пачку творога и несколько бананов, получится неплохой углеводно-протеиновый коктейль. Единственным минусом является содержащийся в нем казеин и то, что его нельзя употреблять после тренировки. Тем не менее такой коктейль лучше всего пить на ночь или даже для осуществления замены одного из приемов еды.
Важно! Наилучшим вариантом будет прием двойной дозы сывороточного протеина после завершения тренинга, а перед тем как лечь спать — казеинового. В выходные от тренировок дни оптимальным вариантом станет употребление полноценной еды.
По данным врачей, для роста мышц человеку в сутки при нормальном образе жизни, без тяжелых нагрузок, требуется 2-3 грамма белка на 1 килограмм веса. Сколько раз в день им следует насыщать организм — вопрос актуальный. По аналогии со спортсменами лучше всего употребление белка разделить на 6 частей в долях 4-5 грамм на 1 кг веса суммарно в сутки. А так как по рекомендации диетологов прием пищи должен быть не менее шести раз в день мелкими порциями — это несложно.
Сколько раз в день им следует насыщать организм — вопрос актуальный. По аналогии со спортсменами лучше всего употребление белка разделить на 6 частей в долях 4-5 грамм на 1 кг веса суммарно в сутки. А так как по рекомендации диетологов прием пищи должен быть не менее шести раз в день мелкими порциями — это несложно.
Дозировка протеина в разное время суток в день тренировки для набора мышечной массы и отдыха.
Протеин поможет при желании набрать мышечную массу и удержать тонус мышц при похудении. При его употреблении важно основываться на правила приема как новичкам, так и профессионалам. Его можно получить как из полноценного рациона, так и из порошковой смеси. Несмотря на натуральность протеина, следует проконсультироваться с врачом и убедиться в отсутствии аллергической реакции на белок и его формы.
Статьи – АКАДЕМИЯ-Т
Содержание:
Известно, что при больших нагрузках организм требует больше питательных веществ, витаминов и минералов. То, что он получает с пищей, оказывается недостаточным для нормального функционирования и тренировочного прогресса. Увеличение же объёмов порций, чревато лишь несварением, ведь большая часть питательных веществ не может усвоиться и пропадает зря. Решение этой проблемы – приём легкоусвояемых диетических спортивных добавок. Основная их роль – дать организму то, чего он не может получить в достаточном количестве из обычной пищи.
То, что он получает с пищей, оказывается недостаточным для нормального функционирования и тренировочного прогресса. Увеличение же объёмов порций, чревато лишь несварением, ведь большая часть питательных веществ не может усвоиться и пропадает зря. Решение этой проблемы – приём легкоусвояемых диетических спортивных добавок. Основная их роль – дать организму то, чего он не может получить в достаточном количестве из обычной пищи.
Несмотря на то, что спортивное питание уже само по себе является отличным подспорьем для основного рациона спортсмена, благотворный эффект от его употребления можно усилить. Для этого стоит тщательно изучить все самые эффективные сочетания спортивных добавок, чтобы их приём отозвался максимальной пользой.
Одно из самых популярных сочетаний – связка протеин+креатин: протеин является материалом для построения мышц, а креатин – источником силы, энергетическим потенциалом для строительства.
Креатин, или иначе метил-гуанидо-уксусная кислота – это вещество, синтезированное на основе аминокислот: глицин, метионин, аргинин. Оно вырабатывается в печени, как энергетический запас. Креатин снабжает энергией мышечную ткань, участвуя в синтезе АТФ (когда АТФ в мышцах иссякает, креатин преобразуется в креатинфосфат, вступает в реакцию с АДФ и продуцирует новую АТФ). Креатин – достаточно нейтральное вещество, он сочетается практически с любым видом спортивного питания. Чаще всего он принимается в связке с BCAA, а также с протеином и гейнером.
Оно вырабатывается в печени, как энергетический запас. Креатин снабжает энергией мышечную ткань, участвуя в синтезе АТФ (когда АТФ в мышцах иссякает, креатин преобразуется в креатинфосфат, вступает в реакцию с АДФ и продуцирует новую АТФ). Креатин – достаточно нейтральное вещество, он сочетается практически с любым видом спортивного питания. Чаще всего он принимается в связке с BCAA, а также с протеином и гейнером.
Особенности приема креатина вместе с протеином
Главное правило приёма разнородных спортивных препаратов: употреблять вместе только те добавки, которые не противоречат друг другу. Если сочетание креатин+БСАА является оптимальным, так как обе добавки дают наилучший эффект при употреблении их до тренировки, то со связкой протеин+креатин дело обстоит иначе. Идеальный вариант: до тренировки выпить креатин, который снабдит организм энергией для тренировки, а после тренировки выпить протеин, дабы восполнить запасы строительного материала для восстановления мышечных волокон. Кроме того, креатин можно выпить дважды за день: порцию до тренировки и порцию, смешанную с протеиновым коктейлем после тренировки.
Кроме того, креатин можно выпить дважды за день: порцию до тренировки и порцию, смешанную с протеиновым коктейлем после тренировки.
Дозировка приема креатина с протеином
Часто спортсмены интересуются, можно ли смешивать креатин с протеином в одном коктейле, и есть ли оптимальная дозировка, при которой обе добавки будут действовать в синергии, обеспечивая положительный эффект.
Способ приёма зависит от конечной цели: набор веса за счет мышц, или сушка тела. В период набора мышечной массы хорошо подходят сочетания:
- за час до тренировки – 3-5 г креатина+30-40 г. протеина (можно заменить его гейнером)+сахар. Разбавляется этот коктейль на молоке или соке;
- сразу по окончании тренировки – коктейль в тех же пропорциях.
Сахар в коктейль добавляется с целью провоцирования инсулинового скачка, так как креатин хорошо усваивается при его наличии. Если коктейль замешивается на сладком соке с высоким гликемическим индексом (гранатовый, виноградный сок), сахар можно не добавлять.
Написал:
Хасанов Адам Алиевич подробнее
Протеины
Я не перестаю удивляться тому, как сывороточный протеин может быть столь популярным видом спортивного питания, но при этом вызывать такую неразбериху и противоречивость информации.
В чем же причина? Берусь утверждать, что это связано, в первую очередь, с недобросовестной рекламой некоторых производителей спортивного питания, а также появлением низкокачественных публикаций или “исследований” с “авторитетными” мнениями всякого рода специалистов в области диетологии. И, наконец, надо согласиться с тем фактом, что сывороточный протеин действительно “сложный” продукт.
В этой статье я рискну раз и навсегда все выяснить, снять, так сказать, завесу секретности и развеять мифы, окружающие этот популярный вид спортивного питания.
Прочитав мою статью, вы поймете разницу между разными формами сывороточного протеина: чем отличается концентрат от изолята, или белок, полученный методом микрофильтрации, от протеина, при производстве которого использовалась технология ионного обмена. Вы также получите ответы на многие другие непростые вопросы, вызывавшие у вас сомнения и неуверенность.
Что представляет собой сывороточный протеин?
Говоря о сывороточном протеине, мы подразумеваем комплексный продукт или смесь, состоящую из нескольких субфракций белка: бета-лактоглобулина, альфа-лактальбумина, иммуноглобулинов (IgGs), гликомакропептидов, альбумина бычьей сыворотки (BSA) и низших пептидов (ферментов): лактопероксидазы, лизоцима (мурамидазы) и лактоферрина. Каждой субфракции, выделенной из сыворотки, присущи уникальные биологические свойства.
До недавнего времени получить такие субфракции удавалось только в количестве, необходимом для проведения лабораторных исследований. Производство в промышленных масштабах было чрезвычайно дорогим и невыгодным предприятием. Современные технологии фильтрации значительно улучшились за последнее десятилетие, позволив выделять из сыворотки субфракции с очень высокой биологической активностью (например, лактоферрин и лактоперокисдазу).
Современные технологии фильтрации значительно улучшились за последнее десятилетие, позволив выделять из сыворотки субфракции с очень высокой биологической активностью (например, лактоферрин и лактоперокисдазу).
В коровьем молоке содержится очень незначительное количество этих субфракций (обычно менее 1%). К примеру, из всего сывороточного белка, содержащегося в коровьем молоке, лактоферрин составляет около 0,5% или того меньше. Это одна из многообещающих субфракций, которая может быть использована для профилактики многих заболеваний, и способствовать улучшению общего состояния здоровья. В грудном молоке содержится до 15% лактоферрина.
За последние десятилетия мы наблюдаем качественный и количественный рост сухих сывороточных протеинов (появление концентратов и изолятов).
Чем хорош сывороточный протеин?
Сывороточный протеин действительно замечательный продукт, иначе он бы не стал одним из основных элементов диеты для большинства бодибилдеров и других спортсменов. В последнее время этот продукт также становится популярным у людей, стремящихся вести здоровый образ жизни, которым известно о его иммуноукрепляющих свойствах.
В последнее время этот продукт также становится популярным у людей, стремящихся вести здоровый образ жизни, которым известно о его иммуноукрепляющих свойствах.
Результаты последних исследований подтверждают, что сывороточный протеин помогает в борьбе с раком, ВИЧ, повышает иммунитет, снижает стресс и уровень кортизола, повышает содержание серотонина в головном мозге, улучшает функцию печени у пациентов, страдающих от некоторых форм гепатита, снижает кровяное давление, улучшает общее самочувствие, не говоря уже о росте спортивных результатов у атлетов, представляющих разные виды спорта.
У сывороточного протеина исключительно высокая биологическая пищевая ценность (хотя производители спортивного питания СУЩЕСТВЕННО превышают значимость этого факта). Более того, в нем содержится много аминокислот с разветвленными цепями (BCAA).
Одним из основных свойств сывороточного протеина является его способность повышать уровень глутатиона (GSH) – важнейшего трипептида, регулирующего работу иммунной системы человека, который также является антиоксидантом.
Концентрация глутатиона внутри клетки напрямую связана со способностью лимфоцитов (важной составляющей иммунной системы) реагировать на угрозу для здоровья человека. Следовательно, изменение содержания внутриклеточного глутатиона можно считать одним из способов иммунной модуляции.
Глутатион – трипептид, состоящий из L-цистеина, L-глютамина и глицина. Цистеин содержит свободную сульфгидрильную группу GSH и является ограничивающим фактором в синтезе глутатиона (хотя влияние сыворотки на глутатион более сложное, чем просто его составляющей в виде цистеина).
Для сывороточного протеина найдется место в питании каждого человека, поскольку глутатион считается важным фактором для поддержания иммунитета (оксидативный стресс, общее состояние самочувствия, и пониженные уровни глутатиона, связанные с длинным списком болезней). Снижение уровня глутатиона также связано с синдромом перетренированности у спортсменов, поэтому сывороточный протеин как нельзя лучше походит для предупреждения, или по крайне мере смягчения состояния перетренированности.
Если говорить исключительно о спорте, то некоторые из последних исследований показывают, что сывороточный протеин может непосредственно влиять на общую результативность и рост мышечной массы у атлетов, но эти исследования принято считать в лучшем случае только предварительными. В ходе исследований также было выявлено, что поскольку оксидитативный стресс способствует появлению мышечной усталости, то высокий уровень глутатиона в крови даст возможность тренироваться дольше и жестче.
Разные типы сывороточного протеина
Больше всего путаницы вокруг сывороточного протеина возникает, когда речь идет о его формах: концентратах, изолятах, протеина, полученного способом ионной очистки, и т.д. Дальше я попытаюсь объяснить вам разницу.
Концентрат
В сухом сывороточном протеине первого поколения содержание чистого белка не превышало 30-40%. Кроме того смесь содержала лактозу, жиры и неденатурированные белки. Этот протеин считался “концентратом” и использовался в пищевой промышленности для выпечки и некоторых других продуктов.
Современные концентраты содержат до 70-80% белка, ограниченное количество лактозы и жиров. Многие считают, что сухой концентрат по качеству хуже изолята, но это неправда.
Хотя в концентрате действительно содержится меньше протеина на грамм смеси, если сравнивать его с изолятом, в их составе есть масса полезных веществ, которые отсутствуют в белковых изолятах.
Так, например, в хороших концентратах содержится несравнимо большее количество факторов роста ИФР-1, ТФР-2 и ТФР-2. В них также намного больше фосфолипидов и биоактивных липидов, таких как линолевая кислота (CLA), а также иммуноглобулинов и лактоферрина.
У нас нет достаточно достоверных сведений, чтобы судить о влиянии, которое оказывают эти соединения на рост мышечной массы и физическую подготовленность атлетов, но мы можем предположить, опять же на основании полученных результатов, что они могут способствовать повышению иммунитета, улучшать состояние желудочно-кишечного тракта и иметь ряд других положительных эффектов не только у спортсменов, но и у “обычных” людей.
К недостаткам сухих концентрированных сывороточных протеинов можно отнести меньшее содержание белка на грамм веса продукта в сравнении с изолятами, большее количество жиров (хотя это вопрос спорный, и здесь могут быть полезные жиры), а также повышенное содержание лактозы.
Вы не должны считать качественный сывороточный концентрат хуже любого изолята. Фактически концентрат может быть даже лучшим выбором в зависимости от ваших целей.
Например, некоторые люди не переносят лактозу и считают каждый грамм жира в своем рационе, тогда как другие не испытывают таких затруднений и могут захотеть использовать дополнительный свойства концентратов, благодаря их более сложной композиции.
Изоляты
Изоляты сывороточного протеина, как правило, содержат до 90-96% белка. Исследования показывают, что сывороточные протеины сохраняют биологическую активность, только находясь в своем природном неденатурированном состоянии (т.е. природном конформационном состоянии).
Производитель должен сильно постараться, чтобы при удалении из протеина лактозы, жиров и прочих компонентов, сохранилась его высокая биодоступность. Сохранение природной неденатурированной формы белков очень важно для их противораковой и иммуномодулирующей активности.
Чтобы сохранить природное состояние, протеин должен быть обработан при низкой температуре и/или в среде с пониженной кислотностью. Это очень ответственный этап во всей технологии получения изолята, которого нет в производстве концентратов.
В изолятах содержится не менее 90% белка с минимальным включением лактозы и практически полным отсутствием жиров. В этом отношении, и в плане содержания белка на грамм веса изолят превосходит концентрированный протеин.
Однако, читатель уже знает о том, что сывороточный протеин — это сложная смесь из многих компонентов, и здесь будет не совсем корректно судить о превосходстве того или иного продукта, основываясь лишь на количественном содержании белка.
При использовании технологии ионного обмена, например, получают изоляты с самым высоким содержанием белка. Значит ли это, что такой протеин лучший из всех изолятов? Вовсе нет, но многие компании до сих пор считают это своей “заветной целью”, пытаясь создать превосходный по качеству сывороточный протеин.
Значит ли это, что такой протеин лучший из всех изолятов? Вовсе нет, но многие компании до сих пор считают это своей “заветной целью”, пытаясь создать превосходный по качеству сывороточный протеин.
Ионный обмен
Сывороточный изолят, полученный способом ионного обмена, производится при прохождении концентрата протеина через специальную колонну. Звучит странно и непонятно, не так ли? При этом у ионного обмена есть один существенный недостаток.
Как ужу говорилось выше, сывороточный протеин — это комплексный белковый продукт, состоящий из множества пептидных субфракций, каждая из которых имеет свои уникальные свойства. Некоторые субфракции присутствуют в сыворотке в очень ограниченном количестве. Фактически, субфракции в итоге делают сывороточный протеин тем уникальным продуктом, которым он является от природы.
При ионном обмене многие из этих тонких и жизненно-важных субфракций разрушаются или истощаются, хотя содержания самого белка в конечном продукте увеличивается.
Вот, собственно, главный недостаток метода ионного обмена, который не может считаться оптимальным способом получения качественного сывороточного протеина третьего поколения. Хотя многие производители продолжают использовать эту технологию для достижения максимальной концентрации белка в своих продуктах.
В протеине, полученным способом ионного обмена, сохраняется от 70% и выше бета-лактоглобулина (по иронии самой неинтересной субфракции, которая при этом отличается высокими аллергенными свойствами), но при этом теряются многие другие биологически активные и интересные компоненты.
Итак, мы можем посоветовать этот тип сывороточного протеина тем людям, для которых важно именно высокое процентное содержание белка, но при этом они готовы пожертвовать отсутствием ряда важных биологически активных компонентов, разрушенных в процессе обработки.
По моему мнению, это нельзя будет назвать оптимальным решением, учитывая минимальную разницу между продуктами полученными с помощью ионного обмена и методом микрофильтрации, о котором говорится ниже.
Изоляты, полученные способом микрофильтрации
Итак, пришло время рассказать о сывороточных изолятах, полученных способом микрофильтрации. Прогресс не стоит на месте, и теперь производители научились получать уникальные изоляты сывороточных протеинов используя технологии перекрестной микрофильтрации (CFM®), сверх фильтрации (UF), микрофильтрации (MF), обратного осмоса (RO), динамической мембранной фильтрации (DMF), ионообменной хроматографии (IEC), электрической ультра фильтрации (EU), радиальной поточной хроматографии (RFC) и нано фильтрации (NF). Возможно, самым распространенным способом, о котором слышало большинство из нас, будет микрофильтрация (CFM®).
“Перекрестной микрофильтрацией” принято называть несколько вариантов обработки белка, при котором используется тонкая очистка концентрата в низкотемпературной среде. На выходе получается продукт с содержанием белка не менее 90%, сохраненными полезными субфракциями, очень низким уровнем жиров и лактозы и практически полным отсутствием неденатурированных белков.
Перекрестная микрофильтрация (CFM®) — естественный нехимический процесс, в котором применяют высокотехнологичные керамические фильтры, в отличие от ионного обмена, в котором используются катализаторы химических реакций типа хлористоводородной кислоты и едкий натр. Сывороточный изолят, полученный методом перекрестной микрофильтрации (CFM®) также содержит много кальция и мало натрия.
Будущее сывороточного протеина
Существует несколько перспективных направлений для создания следующего поколения сывороточных протеинов.
Увеличение процентного содержания полезных субфракций белка
Весьма перспективным видится развитие технологий, связанных с выделением отдельных биоактивных субфракций из сыворотки (например, лактоферрина или гликомакропептидов) в промышленных масштабах, используя уже имеющиеся способы обработки сырья.
Об этом можно было только мечтать еще несколько лет назад, но теперь некоторые производители сывороточного протеина уже используют самые современные технологии фильтрации.
Таким образом, можно было бы создавать продукты со специально подобранными свойствами: выделять субфракции и затем возвращать их в рафинированные продукты, чтобы восстановить их природные химико-биологические свойства (например, лактоферрин, который при существующих методах очистки просто удаляется из сыворотки).
В самых качественных продуктах сегодня содержится всего 0,5-1,0% этой редкой, но важной микрофракции. Ряд производителей уже сегодня могли бы искусственно повысить ее содержание, создавая по истине “дизайнерские” протеины с уникальными свойствами.
Стало известно, что одна компания активно изучает возможность создания изолятов с повышенным содержанием альфа-лактальбумина, еще одной полезной субфракции сыворотки, и практически полным отсутствием бета-лактальбумина, который часто вызывает аллергическую реакцию. Такие изоляты потенциально превосходят по качеству и полезным свойствам все остальные доступные на рынке сывороточные протеины.
Не стоит забывать и о концентратах. Производители могли бы повысить содержание в них факторов роста (ИФР-1, ТФР-1 и ТФР-2) и других биоактивных компонентов в виде фосфолипидов, сопряженной линолевой кислотой (CLA), иммуноглобулинов и лактоферрина. В таких продуктах также станет больше жиров (с 5-10% до приблизительно 15%), но только благодаря повышенному содержанию болезных субфракций.
Производители могли бы повысить содержание в них факторов роста (ИФР-1, ТФР-1 и ТФР-2) и других биоактивных компонентов в виде фосфолипидов, сопряженной линолевой кислотой (CLA), иммуноглобулинов и лактоферрина. В таких продуктах также станет больше жиров (с 5-10% до приблизительно 15%), но только благодаря повышенному содержанию болезных субфракций.
Возвращение гидролизованных белков
Многие из нас помнят бум, связанный с гидролизованными протеинами. Эти продукты были очень популярны несколько лет назад, они появились, сделали много шума и затем также быстро исчезли с полок магазинов. Под “гидролизом” мы понимаем частичное расщепление белков на пептидные цепочки разной длины.
Поскольку протеин поступает в организм уже в расщепленном виде, то и усваиваться он должен значительно быстрей, что может быть важным в некоторых обстоятельствах (например, для людей с тяжелыми ожогами, пациентов с болезнями системы пищеварения или при выкармливании недоношенных детей).
Насколько полезны гидролизованные формы протеина для атлетов — тема для отельного разговора. Ажиотаж вокруг гидролизованного протеина был связан с публикацией результатов лабораторных исследований, когда у мышей, питавшихся гидролизованной формой белка, наблюдалось повышенное задержание азота (положительный азотистый баланс) в отличие от другой группы грызунов, получавших обычный белок.
Ажиотаж вокруг гидролизованного протеина был связан с публикацией результатов лабораторных исследований, когда у мышей, питавшихся гидролизованной формой белка, наблюдалось повышенное задержание азота (положительный азотистый баланс) в отличие от другой группы грызунов, получавших обычный белок.
Прискорбно, но провести подобные эксперименты на спортсменах никто не решился. Как бы то ни было, но этот вид протеинов не задержался на рынке. Потребителям не нравился его отвратительный вкус, стоимость и отсутствие достоверно подтвержденных фактов его “чудотворного” влияния.
В то время при гидролизе протеин практически превращался в денатурат. Одна из компаний разработала собственную технологию расщепления. На выходе получается вполне естественный продукт с нормальными вкусовыми качествами.
Себестоимость производства гидролизованного протеина также снизилась. Не хватает только обширных исследований подобных продуктов на людях. Тем не менее, эта форма протеина может заинтересовать бодибилдеров и других спортсменов.
Минералы из молока?
Молоко может быть источником полезных минералов для бодибилдеров и других спортсменов.
В молоке содержится биологически активная форма кальция, которую можно извлекать без лактозы, содержащейся в кисломолочных продуктах. Здесь также есть магний, фосфор и цинк, необходимый для нормального остеогенеза и метаболизма. Последние проведенные исследования показывают, что дополнительный прием кальция, например, способствует снижению давления.
Бодибилдерам и другим атлетам будет интересен тот факт, что все больше исследований подтверждают взаимосвязь между повышенным содержанием кальция в организме и смещения метаболизма в сторону активного липолиза (расщепления жиров) и подавления липогенеза (жирообразования).
То есть, добавляя в спортивное питание кальций и прочие минералы, полученные из молочного сырья, мы получаем оптимальный продукт, способствующий ускорению метаболизма, набору сухой мышечной массы и укреплению костной ткани.
Заключение
Ну, вот и все. Я надеюсь, что моя статья помогла вам окончательно разобраться с теми вопросами, которые у вас возникали при упоминании о сывороточных протеинах. Теперь вы — опытный потребитель, который разбирается в сути проблемы, и, делая следующую покупку в магазине спортивного питания, вы теперь будете знать, что вам нужно. Не дайте себя одурачить!
Это очень хороший протеин во многих отношениях, но вам надо оставаться реалистами — вы не нарастите горы мышц за короткое время, просто включив этот продукт в ваш рацион питания. Я также советую вам держать руку на пульсе и постоянно интересоваться новыми разработками в сфере продуктов спортивного питания.
Сегодня многие люди стремятся к тому, чтобы вести здоровый образ жизни: в привычном распорядке дня появляются занятия спортом, изменяются в лучшую сторону пищевые привычки. Не всегда обычная еда по своим качествам удовлетворяет новым требованиям людей, ранее далеких от спорта.
С введением физической нагрузки организму требуется большее количество белка, часть которого можно получить с введением в свой рацион спортивного питания. А спортсмены профессионального уровня, да и любители тоже, почти всегда используют различные пищевые добавки. Одной из самых популярных можно смело назвать сывороточный протеин.
Сывороточный протеин – что это такое
Сывороточный протеин – это такой вид спортивного питания, который представляет собой белок, извлеченный из сыворотки посредством фильтрации и далее высушенный.
Когда в пищеварительную систему попадает белок, то аминокислоты, входящие в его состав используются организмом для восстановления различных тканей. Восемь из двадцати существующих аминокислот являются незаменимыми (могут поступать только с пищей). Когда белок содержит все восемь, тогда его называют полноценным. Сывороточный протеин как раз относится к их числу. Такие полноценные белки обязательно есть в мясе, рыбе, молочных продуктах и яйцах. Сывороточный протеин один из самых популярный видов спортивн
Сывороточный протеин один из самых популярный видов спортивн
Сывороточный протеин один из самых популярный видов спортивного питания, потому что он безопасен, полезен для здоровья, отлично подходит для набора массы. Его преимущества в том, что:
— в нем есть все необходимые аминокислоты для строительства мышечной массы;
— усиливает выработку инсулина, обладающего анаболическим действием;
— уменьшает выработку гормонов, разрушающих мышцы, адреналина и кортизола в их числе;
— дает необходимую энергию во время тренировки.
Руководство по оптимальному потреблению белка
| Examine.com
Обсуждаемые до сих пор потребности в белке были основаны на исследованиях, в которых использовались белковые добавки животного происхождения, такие как сывороточный или яичный белок, или проводились в основном на всеядных животных. Нет оснований полагать, что люди, которые получают протеин в основном или полностью из растений, по своей природе имеют разные потребности в протеине, но поскольку протеины растительного происхождения имеют тенденцию быть более низкого качества, чем протеины животного происхождения, если вы получаете большую часть протеина из растений. вам нужно будет обращать внимание не только на количество потребляемого белка, но и на его качество. [92]
вам нужно будет обращать внимание не только на количество потребляемого белка, но и на его качество. [92]
Качество белка определяется его перевариваемостью и аминокислотным профилем .
Усвояемость имеет значение, потому что, если вы не перевариваете и не усваиваете часть съеденного протеина, возможно, он не был съеден. Белки животного происхождения постоянно демонстрируют степень усвояемости выше 90%, тогда как белки из лучших растительных источников (бобовые и зерновые) демонстрируют степень усвояемости 60–80%. [93]
Усвояемость различных белков растительного и животного происхождения
Ссылка: FAO. Оценка качества протеина в питании человека . 2013. [94]
Растения содержат антипитательные вещества, которые препятствуют перевариванию и всасыванию белка, такие как ингибиторы трипсина, фитаты и дубильные вещества. [95] Хотя приготовление пищи действительно снижает концентрацию антипитательных веществ, она не устраняет их полностью. Однако порошки протеинов растительного происхождения в основном не содержат антинутриентов, поэтому их усвояемость аналогична показателям протеинов животного происхождения. [93]
[95] Хотя приготовление пищи действительно снижает концентрацию антипитательных веществ, она не устраняет их полностью. Однако порошки протеинов растительного происхождения в основном не содержат антинутриентов, поэтому их усвояемость аналогична показателям протеинов животного происхождения. [93]
аминокислотный профиль белка имеет значение, потому что все белки, включая белок, который вы едите, и белок в вашем теле, состоят из некоторой комбинации 20 аминокислот (AA).Ваше тело может производить 11 из этих аминокислот, что составляет заменимых аминокислоты и (NEAA). Ваше тело не может производить остальные 9, которые, следовательно, являются незаменимыми аминокислотами, (EAA), которые вы должны получать с пищей.
Для наращивания мышечной массы необходимо, чтобы в сумме синтез мышечного белка на (MPS) превышал распада мышечного белка (MPB), что приводит к чистому накоплению мышечного белка. Все 20 АК необходимы для наращивания мышечной ткани, [96] , но MPS стимулируется в первую очередь EAA в пище, которую вы принимаете. [97]
Все 20 АК необходимы для наращивания мышечной ткани, [96] , но MPS стимулируется в первую очередь EAA в пище, которую вы принимаете. [97]
Белки растительного происхождения, будь то цельные продукты питания или протеиновые порошки, содержат меньше ЕАА, чем белки животного происхождения.
Содержание EAA в растительных и животных белках
Ссылка: FAO. Оценка качества протеина в питании человека . 2013. [94]
В частности, растительные белки содержат меньше лейцина ЕАА, который, как полагают, действует как сигнал для «включения» анаболических сигнальных путей и МПС, [98] [ 99] , хотя для сохранения эффекта требуются все EAA. [100]
Содержание лейцина в растительных и животных белках
Ссылка: van Vliet et al. Дж Нутрь . 2015. [101]
Более низкое содержание лейцина и EAA в растительных белках помогает объяснить, почему в нескольких исследованиях сообщалось о более низких показателях MPS из порошков и напитков соевого белка, чем из сывороточного протеина, [102] [ 103] обезжиренное молоко, [104] цельное молоко с сыром, [105] и нежирная говядина. [106]
[106]
Однако, хотя различия в MPS, по-видимому, приводят к различиям в безжировой массе при использовании умеренных дополнительных доз белка (около 20 г / день), [107] [108] при более высоких дозах используются (33–50 г / день), пищевые добавки животного (сыворотка) и растительного происхождения (соя, рис) влияют на безжировую массу одинаково. [109] [110] [111] [112] Короче говоря, общее потребление большего количества белка, по-видимому, компенсирует более низкое качество белков растительного происхождения.
Белки растительного происхождения также содержат ограничивающих аминокислот , которые представляют собой EAA, присутствующие в таких небольших количествах, что они затрудняют синтез белка. Лизин является наиболее распространенной лимитирующей аминокислотой, особенно в зерновых культурах, таких как пшеница и рис. [113] Орехи и семена также имеют тенденцию содержать лизин в качестве ограничивающей аминокислоты. С другой стороны, фасоль и бобовые содержат достаточное количество лизина, но не содержат сернистых аминокислот, таких как метионин и цистеин. Комбинирование различных белков растительного происхождения может помочь восполнить их дефицит.
С другой стороны, фасоль и бобовые содержат достаточное количество лизина, но не содержат сернистых аминокислот, таких как метионин и цистеин. Комбинирование различных белков растительного происхождения может помочь восполнить их дефицит.
Белки растительного происхождения имеют более низкое качество (они менее биодоступны и содержат меньше EAA). Если вы получаете большую часть белка из растений, вам нужно будет потреблять больше белка, чтобы добиться такого же роста мышц, как и тем, кто придерживается более всеядной диеты.
Усиление растительных белков
Самый простой способ преодолеть дефицит ЕАА в растительном белке — это есть его больше. Как уже упоминалось, несколько исследований показали, что большие дозы (33–50 г / день) дополнительных белков животного (сыворотка) и растительного происхождения (соя, рис), по-видимому, одинаково увеличивают безжировую массу. [109] [110] [111] [112]
Еще один способ преодолеть дефицит ЕАА в растительных белках — комбинировать дополнительные профили ЕАА. [114] Исторические примеры таких комбинаций включают бобы с кукурузой в Северной и Южной Америке и рис с соей в Азии. Эти комбинации зерновых и бобовых работают, потому что бобовые поставляют лизин, отсутствующий в зернах, тогда как зерна поставляют метионин и цистеин, отсутствующие в бобовых.
[114] Исторические примеры таких комбинаций включают бобы с кукурузой в Северной и Южной Америке и рис с соей в Азии. Эти комбинации зерновых и бобовых работают, потому что бобовые поставляют лизин, отсутствующий в зернах, тогда как зерна поставляют метионин и цистеин, отсутствующие в бобовых.
Объединение неполных белков
По материалам: Woolf et al. PLoS Один . 2011. [114]
К сожалению, в большинстве растительных белков мало лейцина, а это означает, что комбинирование различных растительных белков не будет иметь большого преимущества, если только один из этих белков не является кукурузным белком (содержание лейцина в котором сопоставимо с содержанием лейцина сыворотки). .
Если в вашем белке меньше лейцина, вам нужно съесть его больше, чтобы максимизировать MPS, или вы можете принимать лейцин в качестве добавки. MPS был увеличен аналогичным образом на 25 граммов сывороточного протеина (обеспечивающий 3 грамма лейцина) и на комбинацию 6. 25 граммов сывороточного протеина и 4,25 грамма дополнительного лейцина (всего 5 граммов лейцина). [115] Исследование на грызунах с использованием растительных белков показало аналогичные результаты. [116]
25 граммов сывороточного протеина и 4,25 грамма дополнительного лейцина (всего 5 граммов лейцина). [115] Исследование на грызунах с использованием растительных белков показало аналогичные результаты. [116]
Дефицит EAA в растительных белках можно преодолеть, если есть больше, сочетать дополнительные белки и принимать лейцин.
Руководство по оптимальному потреблению белка | Examine.com
Обсуждаемые до сих пор потребности в белке были основаны на исследованиях, в которых использовались белковые добавки животного происхождения, такие как сывороточный или яичный белок, или проводились в основном на всеядных животных.Нет оснований полагать, что люди, которые получают протеин в основном или полностью из растений, по своей природе имеют разные потребности в протеине, но поскольку протеины растительного происхождения имеют тенденцию быть более низкого качества, чем протеины животного происхождения, если вы получаете большую часть протеина из растений. вам нужно будет обращать внимание не только на количество потребляемого белка, но и на его качество. [92]
вам нужно будет обращать внимание не только на количество потребляемого белка, но и на его качество. [92]
Качество белка определяется его перевариваемостью и аминокислотным профилем .
Усвояемость имеет значение, потому что, если вы не перевариваете и не усваиваете часть съеденного протеина, возможно, он не был съеден. Белки животного происхождения постоянно демонстрируют степень усвояемости выше 90%, тогда как белки из лучших растительных источников (бобовые и зерновые) демонстрируют степень усвояемости 60–80%. [93]
Усвояемость различных белков растительного и животного происхождения
Ссылка: FAO. Оценка качества протеина в питании человека .2013. [94]
Растения содержат антипитательные вещества, которые препятствуют перевариванию и всасыванию белка, такие как ингибиторы трипсина, фитаты и дубильные вещества. [95] Хотя приготовление пищи действительно снижает концентрацию антипитательных веществ, она не устраняет их полностью. Однако порошки протеинов растительного происхождения в основном не содержат антинутриентов, поэтому их усвояемость аналогична показателям протеинов животного происхождения. [93]
[95] Хотя приготовление пищи действительно снижает концентрацию антипитательных веществ, она не устраняет их полностью. Однако порошки протеинов растительного происхождения в основном не содержат антинутриентов, поэтому их усвояемость аналогична показателям протеинов животного происхождения. [93]
аминокислотный профиль белка имеет значение, потому что все белки, включая белок, который вы едите, и белок в вашем теле, состоят из некоторой комбинации 20 аминокислот (AA).Ваше тело может производить 11 из этих аминокислот, что составляет заменимых аминокислоты и (NEAA). Ваше тело не может производить остальные 9, которые, следовательно, являются незаменимыми аминокислотами, (EAA), которые вы должны получать с пищей.
Для наращивания мышечной массы необходимо, чтобы в сумме синтез мышечного белка на (MPS) превышал распада мышечного белка (MPB), что приводит к чистому накоплению мышечного белка. Все 20 АК необходимы для наращивания мышечной ткани, [96] , но MPS стимулируется в первую очередь EAA в пище, которую вы принимаете. [97]
Все 20 АК необходимы для наращивания мышечной ткани, [96] , но MPS стимулируется в первую очередь EAA в пище, которую вы принимаете. [97]
Белки растительного происхождения, будь то цельные продукты питания или протеиновые порошки, содержат меньше ЕАА, чем белки животного происхождения.
Содержание EAA в растительных и животных белках
Ссылка: FAO. Оценка качества протеина в питании человека . 2013. [94]
В частности, растительные белки содержат меньше лейцина ЕАА, который, как полагают, действует как сигнал для «включения» анаболических сигнальных путей и МПС, [98] [ 99] , хотя для сохранения эффекта требуются все EAA. [100]
Содержание лейцина в растительных и животных белках
Ссылка: van Vliet et al. Дж Нутрь . 2015. [101]
Более низкое содержание лейцина и EAA в растительных белках помогает объяснить, почему в нескольких исследованиях сообщалось о более низких показателях MPS из порошков и напитков соевого белка, чем из сывороточного протеина, [102] [ 103] обезжиренное молоко, [104] цельное молоко с сыром, [105] и нежирная говядина. [106]
[106]
Однако, хотя различия в MPS, по-видимому, приводят к различиям в безжировой массе при использовании умеренных дополнительных доз белка (около 20 г / день), [107] [108] при более высоких дозах используются (33–50 г / день), пищевые добавки животного (сыворотка) и растительного происхождения (соя, рис) влияют на безжировую массу одинаково. [109] [110] [111] [112] Короче говоря, общее потребление большего количества белка, по-видимому, компенсирует более низкое качество белков растительного происхождения.
Белки растительного происхождения также содержат ограничивающих аминокислот , которые представляют собой EAA, присутствующие в таких небольших количествах, что они затрудняют синтез белка. Лизин является наиболее распространенной лимитирующей аминокислотой, особенно в зерновых культурах, таких как пшеница и рис. [113] Орехи и семена также имеют тенденцию содержать лизин в качестве ограничивающей аминокислоты. С другой стороны, фасоль и бобовые содержат достаточное количество лизина, но не содержат сернистых аминокислот, таких как метионин и цистеин. Комбинирование различных белков растительного происхождения может помочь восполнить их дефицит.
С другой стороны, фасоль и бобовые содержат достаточное количество лизина, но не содержат сернистых аминокислот, таких как метионин и цистеин. Комбинирование различных белков растительного происхождения может помочь восполнить их дефицит.
Белки растительного происхождения имеют более низкое качество (они менее биодоступны и содержат меньше EAA). Если вы получаете большую часть белка из растений, вам нужно будет потреблять больше белка, чтобы добиться такого же роста мышц, как и тем, кто придерживается более всеядной диеты.
Усиление растительных белков
Самый простой способ преодолеть дефицит ЕАА в растительном белке — это есть его больше. Как уже упоминалось, несколько исследований показали, что большие дозы (33–50 г / день) дополнительных белков животного (сыворотка) и растительного происхождения (соя, рис), по-видимому, одинаково увеличивают безжировую массу. [109] [110] [111] [112]
Еще один способ преодолеть дефицит ЕАА в растительных белках — комбинировать дополнительные профили ЕАА. [114] Исторические примеры таких комбинаций включают бобы с кукурузой в Северной и Южной Америке и рис с соей в Азии. Эти комбинации зерновых и бобовых работают, потому что бобовые поставляют лизин, отсутствующий в зернах, тогда как зерна поставляют метионин и цистеин, отсутствующие в бобовых.
[114] Исторические примеры таких комбинаций включают бобы с кукурузой в Северной и Южной Америке и рис с соей в Азии. Эти комбинации зерновых и бобовых работают, потому что бобовые поставляют лизин, отсутствующий в зернах, тогда как зерна поставляют метионин и цистеин, отсутствующие в бобовых.
Объединение неполных белков
По материалам: Woolf et al. PLoS Один . 2011. [114]
К сожалению, в большинстве растительных белков мало лейцина, а это означает, что комбинирование различных растительных белков не будет иметь большого преимущества, если только один из этих белков не является кукурузным белком (содержание лейцина в котором сопоставимо с содержанием лейцина сыворотки). .
Если в вашем белке меньше лейцина, вам нужно съесть его больше, чтобы максимизировать MPS, или вы можете принимать лейцин в качестве добавки. MPS был увеличен аналогичным образом на 25 граммов сывороточного протеина (обеспечивающий 3 грамма лейцина) и на комбинацию 6. 25 граммов сывороточного протеина и 4,25 грамма дополнительного лейцина (всего 5 граммов лейцина). [115] Исследование на грызунах с использованием растительных белков показало аналогичные результаты. [116]
25 граммов сывороточного протеина и 4,25 грамма дополнительного лейцина (всего 5 граммов лейцина). [115] Исследование на грызунах с использованием растительных белков показало аналогичные результаты. [116]
Дефицит EAA в растительных белках можно преодолеть, если есть больше, сочетать дополнительные белки и принимать лейцин.
Руководство по оптимальному потреблению белка | Examine.com
Обсуждаемые до сих пор потребности в белке были основаны на исследованиях, в которых использовались белковые добавки животного происхождения, такие как сывороточный или яичный белок, или проводились в основном на всеядных животных.Нет оснований полагать, что люди, которые получают протеин в основном или полностью из растений, по своей природе имеют разные потребности в протеине, но поскольку протеины растительного происхождения имеют тенденцию быть более низкого качества, чем протеины животного происхождения, если вы получаете большую часть протеина из растений. вам нужно будет обращать внимание не только на количество потребляемого белка, но и на его качество. [92]
вам нужно будет обращать внимание не только на количество потребляемого белка, но и на его качество. [92]
Качество белка определяется его перевариваемостью и аминокислотным профилем .
Усвояемость имеет значение, потому что, если вы не перевариваете и не усваиваете часть съеденного протеина, возможно, он не был съеден. Белки животного происхождения постоянно демонстрируют степень усвояемости выше 90%, тогда как белки из лучших растительных источников (бобовые и зерновые) демонстрируют степень усвояемости 60–80%. [93]
Усвояемость различных белков растительного и животного происхождения
Ссылка: FAO. Оценка качества протеина в питании человека .2013. [94]
Растения содержат антипитательные вещества, которые препятствуют перевариванию и всасыванию белка, такие как ингибиторы трипсина, фитаты и дубильные вещества. [95] Хотя приготовление пищи действительно снижает концентрацию антипитательных веществ, она не устраняет их полностью. Однако порошки протеинов растительного происхождения в основном не содержат антинутриентов, поэтому их усвояемость аналогична показателям протеинов животного происхождения. [93]
[95] Хотя приготовление пищи действительно снижает концентрацию антипитательных веществ, она не устраняет их полностью. Однако порошки протеинов растительного происхождения в основном не содержат антинутриентов, поэтому их усвояемость аналогична показателям протеинов животного происхождения. [93]
аминокислотный профиль белка имеет значение, потому что все белки, включая белок, который вы едите, и белок в вашем теле, состоят из некоторой комбинации 20 аминокислот (AA).Ваше тело может производить 11 из этих аминокислот, что составляет заменимых аминокислоты и (NEAA). Ваше тело не может производить остальные 9, которые, следовательно, являются незаменимыми аминокислотами, (EAA), которые вы должны получать с пищей.
Для наращивания мышечной массы необходимо, чтобы в сумме синтез мышечного белка на (MPS) превышал распада мышечного белка (MPB), что приводит к чистому накоплению мышечного белка. Все 20 АК необходимы для наращивания мышечной ткани, [96] , но MPS стимулируется в первую очередь EAA в пище, которую вы принимаете. [97]
Все 20 АК необходимы для наращивания мышечной ткани, [96] , но MPS стимулируется в первую очередь EAA в пище, которую вы принимаете. [97]
Белки растительного происхождения, будь то цельные продукты питания или протеиновые порошки, содержат меньше ЕАА, чем белки животного происхождения.
Содержание EAA в растительных и животных белках
Ссылка: FAO. Оценка качества протеина в питании человека . 2013. [94]
В частности, растительные белки содержат меньше лейцина ЕАА, который, как полагают, действует как сигнал для «включения» анаболических сигнальных путей и МПС, [98] [ 99] , хотя для сохранения эффекта требуются все EAA. [100]
Содержание лейцина в растительных и животных белках
Ссылка: van Vliet et al. Дж Нутрь . 2015. [101]
Более низкое содержание лейцина и EAA в растительных белках помогает объяснить, почему в нескольких исследованиях сообщалось о более низких показателях MPS из порошков и напитков соевого белка, чем из сывороточного протеина, [102] [ 103] обезжиренное молоко, [104] цельное молоко с сыром, [105] и нежирная говядина. [106]
[106]
Однако, хотя различия в MPS, по-видимому, приводят к различиям в безжировой массе при использовании умеренных дополнительных доз белка (около 20 г / день), [107] [108] при более высоких дозах используются (33–50 г / день), пищевые добавки животного (сыворотка) и растительного происхождения (соя, рис) влияют на безжировую массу одинаково. [109] [110] [111] [112] Короче говоря, общее потребление большего количества белка, по-видимому, компенсирует более низкое качество белков растительного происхождения.
Белки растительного происхождения также содержат ограничивающих аминокислот , которые представляют собой EAA, присутствующие в таких небольших количествах, что они затрудняют синтез белка. Лизин является наиболее распространенной лимитирующей аминокислотой, особенно в зерновых культурах, таких как пшеница и рис. [113] Орехи и семена также имеют тенденцию содержать лизин в качестве ограничивающей аминокислоты. С другой стороны, фасоль и бобовые содержат достаточное количество лизина, но не содержат сернистых аминокислот, таких как метионин и цистеин. Комбинирование различных белков растительного происхождения может помочь восполнить их дефицит.
С другой стороны, фасоль и бобовые содержат достаточное количество лизина, но не содержат сернистых аминокислот, таких как метионин и цистеин. Комбинирование различных белков растительного происхождения может помочь восполнить их дефицит.
Белки растительного происхождения имеют более низкое качество (они менее биодоступны и содержат меньше EAA). Если вы получаете большую часть белка из растений, вам нужно будет потреблять больше белка, чтобы добиться такого же роста мышц, как и тем, кто придерживается более всеядной диеты.
Усиление растительных белков
Самый простой способ преодолеть дефицит ЕАА в растительном белке — это есть его больше. Как уже упоминалось, несколько исследований показали, что большие дозы (33–50 г / день) дополнительных белков животного (сыворотка) и растительного происхождения (соя, рис), по-видимому, одинаково увеличивают безжировую массу. [109] [110] [111] [112]
Еще один способ преодолеть дефицит ЕАА в растительных белках — комбинировать дополнительные профили ЕАА. [114] Исторические примеры таких комбинаций включают бобы с кукурузой в Северной и Южной Америке и рис с соей в Азии. Эти комбинации зерновых и бобовых работают, потому что бобовые поставляют лизин, отсутствующий в зернах, тогда как зерна поставляют метионин и цистеин, отсутствующие в бобовых.
[114] Исторические примеры таких комбинаций включают бобы с кукурузой в Северной и Южной Америке и рис с соей в Азии. Эти комбинации зерновых и бобовых работают, потому что бобовые поставляют лизин, отсутствующий в зернах, тогда как зерна поставляют метионин и цистеин, отсутствующие в бобовых.
Объединение неполных белков
По материалам: Woolf et al. PLoS Один . 2011. [114]
К сожалению, в большинстве растительных белков мало лейцина, а это означает, что комбинирование различных растительных белков не будет иметь большого преимущества, если только один из этих белков не является кукурузным белком (содержание лейцина в котором сопоставимо с содержанием лейцина сыворотки). .
Если в вашем белке меньше лейцина, вам нужно съесть его больше, чтобы максимизировать MPS, или вы можете принимать лейцин в качестве добавки. MPS был увеличен аналогичным образом на 25 граммов сывороточного протеина (обеспечивающий 3 грамма лейцина) и на комбинацию 6. 25 граммов сывороточного протеина и 4,25 грамма дополнительного лейцина (всего 5 граммов лейцина). [115] Исследование на грызунах с использованием растительных белков показало аналогичные результаты. [116]
25 граммов сывороточного протеина и 4,25 грамма дополнительного лейцина (всего 5 граммов лейцина). [115] Исследование на грызунах с использованием растительных белков показало аналогичные результаты. [116]
Дефицит EAA в растительных белках можно преодолеть, если есть больше, сочетать дополнительные белки и принимать лейцин.
Обзор, применение, побочные эффекты, меры предосторожности, взаимодействия, дозировка и отзывы
Abou-Samra, R., Keersmaekers, L., Brienza, D., Mukherjee, R., и Мейс, К. Влияние различных источников белка на сытость и кратковременное насыщение при употреблении в качестве закуски. Нутр Ж. 2011; 10: 139. Просмотреть аннотацию.
Ачесон, К.Дж., Блондель-Лубрано, А., Огей-Араймон, С., Бомонт, М., Эмади-Азар, С., Аммон-Цуффери, К., Моннар, И., Пино, С., Нильсен -Moennoz, C., и Bovetto, L. Выбор белков, направленных на термогенез и метаболизм. Am J Clin. Nutr 2011; 93 (3): 525-534. Просмотреть аннотацию.
Агин, Д., Галлахер, Д., Ван, Дж., Хеймсфилд, С.Б., Пирсон, Р. Н., младший, и Котлер, Д. П. Влияние сывороточного протеина и силовых упражнений на массу клеток тела, мышечную силу и качество жизни женщин с ВИЧ. СПИД 12-7-2001; 15 (18): 2431-2440. Просмотреть аннотацию.
Агин, Д., Котлер, Д. П., Папандреу, Д., Лисс, М., Ван, Дж., Торнтон, Дж., Галлахер, Д., и Пирсон, Р. Н., мл. Влияние сывороточного протеина и силовых упражнений на состав тела и силу мышц у женщин с ВИЧ-инфекцией. Ann.N.Y.Acad.Sci. 2000; 904: 607-609. Просмотреть аннотацию.
Ахаван Т., Луховый Б.Л. и Андерсон Г.Х. Влияние употребления алкоголя по сравнению с употреблением сахара или сывороточного протеина на краткосрочный аппетит и прием пищи. Int J Obes. (Лондон), 8-24-2010; Просмотреть аннотацию.
Ахаван, Т., Луховый, Б.Л., Браун, П.Х., Чо, К.Э. и Андерсон, Г.Х. Влияние потребления сывороточного протеина и его гидролизата перед приемом пищи на потребление пищи, гликемию после еды и реакцию на инсулин у молодых людей. Am J Clin Nutr 2010; 91 (4): 966-975. Просмотреть аннотацию.
Am J Clin Nutr 2010; 91 (4): 966-975. Просмотреть аннотацию.
Олдрич, Н.Д., Рейкс, М. М., Сибли, С. Д., Редмон, Дж. Б., Томас, В., и Раатц, С. К. Варьирование источника и количества белка не приводит к значительному улучшению потери веса, потери жира или насыщения при низкоэнергетической диете среди взрослых среднего возраста. Nutr Res 2011; 31 (2): 104-112. Просмотреть аннотацию.
Александер Д. Д. и Кабана М. Д. Частично гидролизованная детская смесь из 100% сывороточного белка и снижение риска атопического дерматита: метаанализ. Журнал Педиатр Гастроэнтерол. Нутр 2010; 50 (4): 422-430. Просмотреть аннотацию.
Александр, Д.Д., Шмитт, Д. Ф., Тран, Н. Л., Баррадж, Л. М., и Кушинг, С. А. Частично гидролизованная смесь для детского питания на основе 100% сывороточного белка и снижение риска атопического дерматита: систематический обзор литературы. Nutr Rev.2010; 68 (4): 232-245. Просмотреть аннотацию.
Alfenas, Rde C., Bressan, J. и Paiva, A.C. Влияние качества белка на аппетит и энергетический обмен у субъектов с нормальным весом. Arq Bras.Endocrinol Metabol. 2010; 54 (1): 45-51. Просмотреть аннотацию.
Алмаас, Х., Холм, Х., Лангсруд, Т., Flengsrud, R., and Vegarud, G.E. Исследования in vitro переваривания белков сыворотки крови козла желудочным и двенадцатиперстным соком человека и их воздействия на отдельные микроорганизмы. Br J Nutr 2006; 96 (3): 562-569. Просмотреть аннотацию.
Альварес, Л. И., Сомелл, К. А., Санчес, С. Ф., и Ланус, К. Е. Кинетика распределения метаболитов альбендазола в плазме у свиней, получавших разные диеты. Res Vet.Sci 1996; 60 (2): 152-156. Просмотреть аннотацию.
Андерсон, Г. Х., Текимер, С. Н., Шах, Д., и Зафар, Т.A. Источник белка, количество и время потребления определяют влияние белков на кратковременное потребление пищи у молодых мужчин. J Nutr 2004; 134 (11): 3011-3015. Просмотреть аннотацию.
Антонионе, Р., Калиандро, Э., Зорат, Ф., Гварниери, Г., Хеер, М., и Биоло, Г. Прием сывороточного протеина усиливает постпрандиальный анаболизм во время кратковременного постельного режима у молодых мужчин. J Nutr 2008; 138 (11): 2212-2216. Просмотреть аннотацию.
J Nutr 2008; 138 (11): 2212-2216. Просмотреть аннотацию.
Aoe, S., Koyama, T., Toba, Y., Itabashi, A., and Takada, Y. Контролируемое исследование влияния добавок основного белка молока (MBP) на метаболизм костей у здоровых женщин в менопаузе.Osteoporos.Int 2005; 16 (12): 2123-2128. Просмотреть аннотацию.
Aoe, S., Toba, Y., Yamamura, J., Kawakami, H., Yahiro, M., Kumegawa, M., Itabashi, A., and Takada, Y. Контролируемое испытание эффектов молочной основы добавка протеина (MBP) для метаболизма костей у здоровых взрослых женщин. Biosci.Biotechnol.Biochem. 2001; 65 (4): 913-918. Просмотреть аннотацию.
Arnberg, K., Molgaard, C., Michaelsen, KF, Jensen, SM, Trolle, E. и Larnkjaer, A. Обезжиренное молоко, сыворотка и казеин увеличивают массу тела, а сыворотка и казеин увеличивают C-пептид в плазме концентрация у подростков с избыточным весом.J Nutr 2012; 142 (12): 2083-2090. Просмотреть аннотацию.
Эстбери, Н. М., Стивенсон, Э. Дж., Моррис, П., Тейлор, М. А., и Макдональд, И. А. Дозозависимый эффект предварительной нагрузки сывороточного протеина на потребление энергии в течение дня у худых субъектов. Br J Nutr 2010; 104 (12): 1858-1867. Просмотреть аннотацию.
А. Дозозависимый эффект предварительной нагрузки сывороточного протеина на потребление энергии в течение дня у худых субъектов. Br J Nutr 2010; 104 (12): 1858-1867. Просмотреть аннотацию.
Atherton, PJ, Etheridge, T., Watt, PW, Wilkinson, D., Selby, A., Rankin, D., Smith, K., and Rennie, MJ Полный эффект мышц после перорального приема протеина: зависимость от времени и несоответствие между синтезом мышечного белка человека и передачей сигналов mTORC1.Am J Clin Nutr 2010; 92 (5): 1080-1088. Просмотреть аннотацию.
Баер, Д. Дж., Стоте, К. С., Пол, Д. Р., Харрис, Г. К., Румплер, В. В. и Клевиденс, Б. А. Добавка сывороточного протеина, но не соевого протеина, изменяет массу и состав тела у свободно живущих взрослых с избыточным весом и ожирением. J Nutr 2011; 141 (8): 1489-1494. Просмотреть аннотацию.
Баллард, К. Д., Бруно, Р. С., Зайп, Р. Л., Куанн, Э. Э., Фольк, Б. М., Фрейденрайх, Д. Дж., Кавецки, Д. М., Купчак, Б. Р., Чанг, М. Ю., Кремер, В. Дж., и Волек, Дж. С. Острый прием нового пептида, полученного из сыворотки, улучшает эндотелиальные реакции сосудов у здоровых людей: рандомизированное плацебо-контролируемое исследование. Нутр J 2009; 8: 34. Просмотреть аннотацию.
Острый прием нового пептида, полученного из сыворотки, улучшает эндотелиальные реакции сосудов у здоровых людей: рандомизированное плацебо-контролируемое исследование. Нутр J 2009; 8: 34. Просмотреть аннотацию.
Баллард, К.Д., Купчак, Б.Р., Волк, Б.М., Ма, Э., Шкрета, А., Липтак, К., Птолемей, А.С., Келлог, М.С., Бруно, Р.С., Зайп, Р.Л., Мареш, К.М., Кремер , WJ, и Volek, JS. Острые эффекты приема нового экстракта сыворотки на функцию эндотелия сосудов у мужчин и женщин среднего возраста с избыточным весом.Br.J Nutr 3-14-2013; 109 (5): 882-893. Просмотреть аннотацию.
Басу Дж., Сачдева А. и Нагпал Дж. Новые полезные свойства ферментированного молока и сывороточных белков: роль в искоренении Helicobacter pylori. J Clin Gastroenterol. 2009; 43 (10): 1011-1012. Просмотреть аннотацию.
Bauer, MP, Numan-Ruberg, SC, Bredewold, OW, Kuijper, EJ, Mooi-Kokenberg, EA, Debast, SB, and van Dissel, JT [Рецидив диареи, связанной с Clostridium difficile, предотвращенный введением сыворотки концентрат от специально иммунизированных коров; перспективное исследование]. Ned.Tijdschr.Geneeskd. 8-30-2008; 152 (35): 1919-1926. Просмотреть аннотацию.
Ned.Tijdschr.Geneeskd. 8-30-2008; 152 (35): 1919-1926. Просмотреть аннотацию.
Бауманн, Дж. М., Рунделл, К. В., Эванс, Т. М., и Левин, А. М. Влияние добавок доноров цистеина на бронхоспазм, вызванный физической нагрузкой. Медико-спортивные упражнения. 2005; 37 (9): 1468-1473. Просмотреть аннотацию.
Байрам Т., Пекмез М., Арда Н. и Ялцин А.С. Антиоксидантная активность фракций сывороточного белка, выделенных с помощью гель-эксклюзионной хроматографии и обработки протеазой. Таланта 5-15-2008; 75 (3): 705-709. Просмотреть аннотацию.
Beaulieu, J., Dupont, C., and Lemieux, P. Противовоспалительный потенциал податливого матрикса, состоящего из ферментированных белков сыворотки и молочнокислых бактерий, на модели атопического дерматита. J. Inflamm. (Лондон) 2007; 4: 6. Просмотреть аннотацию.
Beaulieu, J., Girard, D., Dupont, C., and Lemieux, P. Ингибирование нейтрофильной инфильтрации гибкой белковой матрицей белков сыворотки, ферментированных молочнокислыми бактериями, in vivo. Inflamm.Res 2009; 58 (3): 133-138. Просмотреть аннотацию.
Inflamm.Res 2009; 58 (3): 133-138. Просмотреть аннотацию.
Больё, Дж., Millette, E., Trottier, E., Precourt, L.P, Dupont, C., and Lemieux, P. Регулирующая функция пластичной белковой матрицы как нового ферментированного сывороточного продукта по признакам, определяющим метаболический синдром. J Med Food 2010; 13 (3): 509-519. Просмотреть аннотацию.
Беккер, А., Уотсон, В., Фергюсон, А., Димич-Уорд, Х., и Чан-Йунг, М. Канадское исследование первичной профилактики астмы: результаты в возрасте 2 лет. J Allergy Clin Immunol. 2004; 113 (4): 650-656. Просмотреть аннотацию.
Беллиссимо, Н., Десантадина, М. В., Пенчарц, П. Б., Бералл, Г. Б., Томас, С. Г., и Андерсон, Г. Х. Сравнение краткосрочного аппетита и потребления энергии у мальчиков с нормальным весом и тучных мальчиков после употребления глюкозы и сывороточного протеина. Int J Obes. (Лондон) 2008; 32 (2): 362-371. Просмотреть аннотацию.
Беллиссимо Н., Томас С. Г., Гуд Р. К. и Андерсон Г. Х. Влияние кратковременной физической активности и порога вентиляции на субъективный аппетит и кратковременное потребление энергии у мальчиков. Аппетит 2007; 49 (3): 644-651.Просмотреть аннотацию.
Аппетит 2007; 49 (3): 644-651.Просмотреть аннотацию.
Бембен М.Г., Виттен М.С. Картер Дж.М. Элиот К.А. Кнеханс А.В. Бембен Д.А. Влияние добавок креатина и протеина на мышечную силу после традиционной программы тренировок с отягощениями у мужчин среднего и старшего возраста. J Nutr Здоровье Старения. 2010; 14 (2): 155-159. Просмотреть аннотацию.
Бентон, М. Дж. И Свон, П. Д. Влияние приема белка на расход энергии и использование субстрата после упражнений у женщин среднего возраста. Int J Sport Nutr Exerc.Metab 2007; 17 (6): 544-555.Просмотреть аннотацию.
Берг, А., Крамер, У., Линк, Э., Боллрат, К., Генрих, Дж., Броков, И., Колецко, С., Грубль, А., Филипьяк-Питтрофф, Б., Вихманн , HE, Bauer, CP, Reinhardt, D., and Berdel, D. Влияние раннего кормления на детскую экзему: развитие после диетического вмешательства по сравнению с естественным течением — исследование GINIplus в возрасте до 6 лет. Clin Exp. Аллергия 2010; 40 (4): 627-636. Просмотреть аннотацию.
Бертольд, Х. К., Шульте, Д. М., Лапуант, Дж. Ф., Лемье, П., Krone, W., и Gouni-Berthold, I. Податливая белковая матрица продукта ферментации сыворотки снижает концентрацию триглицеридов у субъектов с гиперхолестеринемией: рандомизированное плацебо-контролируемое исследование. J Dairy Sci 2011; 94 (2): 589-601. Просмотреть аннотацию.
К., Шульте, Д. М., Лапуант, Дж. Ф., Лемье, П., Krone, W., и Gouni-Berthold, I. Податливая белковая матрица продукта ферментации сыворотки снижает концентрацию триглицеридов у субъектов с гиперхолестеринемией: рандомизированное плацебо-контролируемое исследование. J Dairy Sci 2011; 94 (2): 589-601. Просмотреть аннотацию.
Беттс, Дж. А., Тоун, Р. Дж., Стоукс, К. А. и Томпсон, Д. Системные индексы повреждения скелетных мышц и восстановления мышечной функции после упражнений: эффект комбинированного приема углеводов и белков. Appl Physiol Nutr Metab 2009; 34 (4): 773-784.Просмотреть аннотацию.
Беттс, Дж. А., Уильямс, К., Бубис, Л. и Цинцас, К. Повышенное окисление углеводов после приема углеводов с добавлением белка. Медико-спортивные упражнения. 2008; 40 (5): 903-912. Просмотреть аннотацию.
Беттс, Дж., Уильямс, К., Даффи, К. и Ганнер, Ф. Влияние приема углеводов и белков во время восстановления после длительных упражнений на последующие показатели выносливости. J Sports Sci 2007; 25 (13): 1449-1460. Просмотреть аннотацию.
Просмотреть аннотацию.
Бихуняк, Дж. Д., Simpson, C.A., Sullivan, R.R., Caseria, D.M., Kerstetter, J.E., Insogna, K.L. Повышение содержания кальция в моче, вызванное диетическим белком, сопровождается аналогичным увеличением азота и мочевины в моче: контролируемое клиническое испытание. J Acad.Nutr Diet. 2013; 113 (3): 447-451. Просмотреть аннотацию.
Бьоркман, М. П., Пилви, Т. К., Кекконен, Р. А., Корпела, Р. и Тилвис, Р. С. Схожие эффекты богатых лейцином и обычных молочных продуктов на мышечную массу и функции пожилых пациентов с ревматической полимиалгией: рандомизированное перекрестное исследование.J Nutr Health Aging 2011; 15 (6): 462-467. Просмотреть аннотацию.
Блэкер, С. Д., Уильямс, Н. С., Фаллоуфилд, Дж. Л., Билзон, Дж. Л., и Виллемс, М. Е. Углеводные и белковые добавки для восстановления нервно-мышечной функции после длительного ношения нагрузки. J Int Soc Sports Nutr 2010; 7: 2. Просмотреть аннотацию.
Bongers, ME, de, Lorijn F., Reitsma, JB, Groeneweg, M. , Taminiau, JA, and Benninga, MA Клинический эффект новой детской смеси для доношенных детей с запором: двойной слепой рандомизированный кросс — по делу.Нутр Ж 2007; 6: 8. Просмотреть аннотацию.
, Taminiau, JA, and Benninga, MA Клинический эффект новой детской смеси для доношенных детей с запором: двойной слепой рандомизированный кросс — по делу.Нутр Ж 2007; 6: 8. Просмотреть аннотацию.
Борденав, С., Саннье, Ф., Рикар, Г. и Пиот, Дж. М. Непрерывный гидролиз козьей сыворотки в реакторе ультрафильтрации: образование альфа-лакторфина. Prep.Biochem.Biotechnol. 1999; 29 (2): 189-202. Просмотреть аннотацию.
Borsheim, E., Aarsland, A. и Wolfe, R.R. Влияние смеси аминокислот, белков и углеводов на баланс чистого мышечного белка после упражнений с отягощениями. Int J Sport Nutr Exerc.Metab 2004; 14 (3): 255-271. Просмотреть аннотацию.
Бортолотти, М., Майоло, Э., Корацца, М., Ван, Дейк Э., Шнайтер, П., Босс, А., Каррел, Г., Джусти, В., Ле, К. А., Куо Чонг, Д.Г., Бюлер, Т., Крейс, Р., Бош, К., и Таппи, Л. Влияние добавок сывороточного протеина на внутрипеченочные липиды у женщин с ожирением. Клинический журнал 2011; 30 (4): 494-498. Просмотреть аннотацию.
Боуэн, Дж. , Ноукс, М., и Клифтон, П. М. Гормоны аппетита и потребление энергии у мужчин с ожирением после употребления фруктозы, глюкозы и напитков с сывороточным белком.Int J Obes. (Лондон) 2007; 31 (11): 1696-1703. Просмотреть аннотацию.
, Ноукс, М., и Клифтон, П. М. Гормоны аппетита и потребление энергии у мужчин с ожирением после употребления фруктозы, глюкозы и напитков с сывороточным белком.Int J Obes. (Лондон) 2007; 31 (11): 1696-1703. Просмотреть аннотацию.
Боуэн, Дж., Ноукс, М., и Клифтон, П. М. Реакции гормонов, регулирующих аппетит, на различные пищевые белки различаются в зависимости от статуса индекса массы тела, несмотря на аналогичное снижение потребления энергии ad libitum. J. Clin Endocrinol. Metab 2006; 91 (8): 2913-2919. Просмотреть аннотацию.
Боуэн, Дж., Ноукс, М., Тренерри, С. и Клифтон, П. М. Потребление энергии, грелин и холецистокинин после различных преднагрузок углеводов и белков у мужчин с избыточным весом.J. Clin Endocrinol Metab 2006; 91 (4): 1477-1483. Просмотреть аннотацию.
Браун-Фарландер К. и фон Мутиус Э. Может ли потребление молока на ферме предотвратить аллергические заболевания? Клиническая экспертиза аллергии 2011; 41 (1): 29-35. Просмотреть аннотацию.
Брин, Л. , Филп, А., Витард, О.К., Джекман, С.Р., Селби, А., Смит, К., Баар, К., и Типтон, К.Д. Влияние совместного приема углеводов и белков на выносливость упражнения на синтез миофибриллярных и митохондриальных белков. J. Physiol 8-15-2011; 589 (Pt 16): 4011-4025.Просмотреть аннотацию.
, Филп, А., Витард, О.К., Джекман, С.Р., Селби, А., Смит, К., Баар, К., и Типтон, К.Д. Влияние совместного приема углеводов и белков на выносливость упражнения на синтез миофибриллярных и митохондриальных белков. J. Physiol 8-15-2011; 589 (Pt 16): 4011-4025.Просмотреть аннотацию.
Браун, Э. К., ДиСилвестро, Р. А., Бабакния, А., Девор, С. Т. Соевые протеиновые батончики в сравнении с батончиками: влияние тренировок на мышечную массу и антиоксидантный статус. Нутр Ж 12-8-2004; 3: 22. Просмотреть аннотацию.
Брун, А.С., Стордал, К., Йоханнесдоттир, Г. Б., Бентсен, Б. С., и Медхус, А. В. Влияние белкового состава жидкой пищи на скорость опорожнения желудка у детей с церебральным параличом. Clin.Nutr 2012; 31 (1): 108-112. Просмотреть аннотацию.
Бакли, Дж.Д., Томсон, Р. Л., Коутс, А. М., Хоу, П. Р., ДеНичило, М. О. и Роуни, М. К. Добавка гидролизата сывороточного протеина ускоряет восстановление мышечной силы после эксцентрических упражнений. J Sci Med Sport 2010; 13 (1): 178-181. Просмотреть аннотацию.
Просмотреть аннотацию.
Burke, DG, Chilibeck, PD, Davidson, KS, Candow, DG, Farthing, J., and Smith-Palmer, T. Влияние добавок сывороточного протеина с моногидратом креатина и без него в сочетании с тренировками с отягощениями на мышечную массу и мышечная сила.Int J Sport Nutr. Exerc.Metab 2001; 11 (3): 349-364. Просмотреть аннотацию.
Кальбет, Дж. А. и Холст, Дж. Дж. Опорожнение желудка, желудочная секреция и энтерогастроновая реакция после введения белков молока или их пептидных гидролизатов людям. Eur.J Nutr 2004; 43 (3): 127-139. Просмотреть аннотацию.
Камфилд, Д. А., Оуэн, Л., Шоли, А. Б., Пипингас, А., и Стоу, С. Молочные компоненты и нейрокогнитивное здоровье в процессе старения. Br.J Nutr 2011; 106 (2): 159-174. Просмотреть аннотацию.
Канчиани, М.и Mastella, G. Поглощение новой полуэлементной диеты у младенцев с муковисцидозом. J Pediatr Gastroenterol. Nutr 1985; 4 (5): 735-740. Просмотреть аннотацию.
Карсилло, Дж. А., Дин, Дж. М., Голубков, Р. , Бергер, Дж., Меерт, К. Л., Ананд, К. Дж., Циммерман, Дж., Ньют, К. Дж., Харрисон, Р., Берр, Дж., Уилсон, Д. Ф. и Николсон, С. Рандомизированное сравнительное исследование профилактики иммуносупрессии, вызванной стрессом, критических заболеваний у детей (CRISIS). Pediatr.Crit Care Med. 2012; 13 (2): 165-173. Просмотреть аннотацию.
, Бергер, Дж., Меерт, К. Л., Ананд, К. Дж., Циммерман, Дж., Ньют, К. Дж., Харрисон, Р., Берр, Дж., Уилсон, Д. Ф. и Николсон, С. Рандомизированное сравнительное исследование профилактики иммуносупрессии, вызванной стрессом, критических заболеваний у детей (CRISIS). Pediatr.Crit Care Med. 2012; 13 (2): 165-173. Просмотреть аннотацию.
Карсилло, Дж., Голубков, Р., Дин, Дж. М., Бергер, Дж., Мерт, К.Л., Ананд, К.Дж., Циммерман, Дж., Ньют, С.Дж., Харрисон, Р., Уилсон, Д.Ф., и Николсон, C. Обоснование и дизайн исследования по профилактике иммунного подавления, вызванного стрессом, критических заболеваний у детей (CRISIS). JPEN J Parenter, Enteral Nutr 2009; 33 (4): 368-374. Просмотреть аннотацию.
Cepero, M. Влияние употребления напитков из казеинового белка и углеводов сыворотки на восстановление и выполнение теста на выносливость.Журнал Human Sport & Exercise 2010; 5 (2): 158.
Чан, Ю. Х., Шек, Л. П., Ав, М., Квак, С. Х., и Ли, Б. В. Использование гипоаллергенной смеси для профилактики атопических заболеваний среди азиатских детей.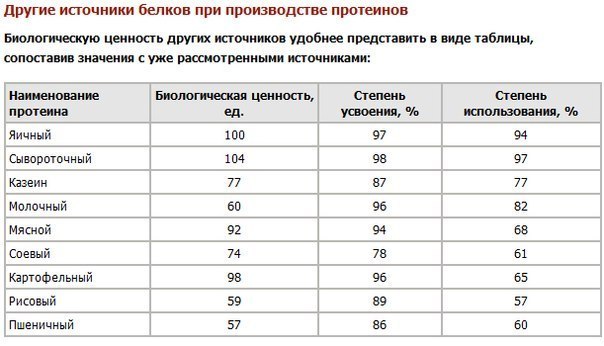 J Paediatr.Child Health 2002; 38 (1): 84-88. Просмотреть аннотацию.
J Paediatr.Child Health 2002; 38 (1): 84-88. Просмотреть аннотацию.
Chan-Yeung, M., Manfreda, J., Dimich-Ward, H., Ferguson, A., Watson, W., and Becker, A. Рандомизированное контролируемое исследование эффективности многогранной программы вмешательства в первичная профилактика астмы у младенцев из группы высокого риска.Arch Pediatr Adolesc.Med 2000; 154 (7): 657-663. Просмотреть аннотацию.
Chang, CY, Trehan, I., Wang, RJ, Thakwalakwa, C., Maleta, K., Deitchler, M., и Manary, MJ Дети, успешно вылеченные от умеренного острого недоедания, остаются в группе риска недоедания и смерти в следующий год после выздоровления. J Nutr 2013; 143 (2): 215-220. Просмотреть аннотацию.
Chen, JR, Singhal, R., Lazarenko, OP, Liu, X., Hogue, WR, Badger, TM, и Ronis, MJ Кратковременное влияние на качество костей, связанное с потреблением изолята соевого белка и других пищевых источников белка у быстрорастущих самок крыс.Exp.Biol Med (Maywood.) 2008; 233 (11): 1348-1358. Просмотреть аннотацию.
Кирико, Г., Гаспарони, А., Чиарделли, Л., Де, Амичи М., Коломбо, А., и Рондини, Г. Иммуногенность и антигенность частично гидролизованной смеси для грудных детей из коровьего молока. Аллергия 1997; 52 (1): 82-88. Просмотреть аннотацию.
Chitapanarux, T., Tienboon, P., Pojchamarnwiputh, S. и Leelarungrayub, D. Открытое пилотное исследование добавок изолятов сывороточного протеина, богатого цистеином, для пациентов с неалкогольным стеатогепатитом.J Гастроэнтерол.Гепатол. 2009; 24 (6): 1045-1050. Просмотреть аннотацию.
Чой, Ю.С., Гото, С., Икеда, И., и Сугано, М. Взаимодействие диетического белка, холестерина и возраста на липидный метаболизм крысы. Br J Nutr 1989; 61 (3): 531-543. Просмотреть аннотацию.
Чой, Ю.С., Икеда, И., и Сугано, М. Пищевые жиры модулируют возрастные эффекты пищевых белков на метаболизм холестерина у крыс. J Nutr Sci Vitaminol. (Токио) 1990; 36 Приложение 2: S181-S184. Просмотреть аннотацию.
Цой, Ю.С., Икеда И. и Сугано М. Комбинированное влияние пищевых белков и рыбьего жира на метаболизм холестерина у крыс разного возраста. Липиды 1989; 24 (6): 506-510. Просмотреть аннотацию.
Комбинированное влияние пищевых белков и рыбьего жира на метаболизм холестерина у крыс разного возраста. Липиды 1989; 24 (6): 506-510. Просмотреть аннотацию.
Chromiak, JA, Smedley, B., Carpenter, W., Brown, R., Koh, YS, Lamberth, JG, Joe, LA, Abadie, BR, и Altorfer, G. Эффект 10-недельной силовой тренировки программа и восстанавливающий напиток на композицию тела, мышечную силу и выносливость, а также анаэробную мощность и емкость. Питание 2004; 20 (5): 420-427.Просмотреть аннотацию.
Chung, C. S., Yamini, S., and Trumbo, P. R. Обзор заявлений FDA о здоровье: частично гидролизованные молочные смеси с сывороточным протеином и атопический дерматит. Педиатрия 2012; 130 (2): e408-e414. Просмотреть аннотацию.
Чунгчунлам, С. М., Моуган, П. Дж., Хенаре, С. Дж., И Ганеш, С. Влияние времени потребления предварительных нагрузок на показатели насыщения у здоровых женщин с нормальным весом. Аппетит 2012; 59 (2): 281-288. Просмотреть аннотацию.
Классенс, М., Каламе, В. , Сименсма, А.Д., ван Баак, М. А., и Сарис, В. Х. Влияние различных смесей гидролизата белка / углеводов на постпрандиальные реакции глюкагона и инсулина у здоровых субъектов. Eur.J Clin Nutr 2009; 63 (1): 48-56. Просмотреть аннотацию.
, Сименсма, А.Д., ван Баак, М. А., и Сарис, В. Х. Влияние различных смесей гидролизата белка / углеводов на постпрандиальные реакции глюкагона и инсулина у здоровых субъектов. Eur.J Clin Nutr 2009; 63 (1): 48-56. Просмотреть аннотацию.
Claessens, M., Saris, W.H. и van Baak, M.A. Ответы на глюкагон и инсулин после приема внутрь различных количеств интактных и гидролизованных белков. Br J Nutr 2008; 100 (1): 61-69. Просмотреть аннотацию.
Классенс, М., ван Баак, М.А., Моншеймер, С., и Сарис, В.H. Влияние диеты с низким содержанием жиров, высоким содержанием белка или углеводов ad libitum на поддержание потери веса и метаболические факторы риска. Int J Obes. (Лондон) 2009; 33 (3): 296-304. Просмотреть аннотацию.
Клаузен М. Р., Скибстед Л. Х. и Стагстед Дж. Характеристика основных видов поглотителей радикалов в коровьем молоке с помощью эксклюзионной хроматографии и функциональных анализов. J. Agric.Food Chem. 4-8-2009; 57 (7): 2912-2919. Просмотреть аннотацию.
Коберн, Дж. У., Хауш, Д. Дж., Хауш, Т. Дж., Малек, М.Х., Бек, Т. В., Крамер, Дж. Т., Джонсон, Г. О. и Донлин, П. Е. Влияние добавок лейцина и сывороточного протеина в течение восьми недель односторонних тренировок с отягощениями. J. Strength Cond. Res 2006; 20 (2): 284-291. Просмотреть аннотацию.
У., Хауш, Д. Дж., Хауш, Т. Дж., Малек, М.Х., Бек, Т. В., Крамер, Дж. Т., Джонсон, Г. О. и Донлин, П. Е. Влияние добавок лейцина и сывороточного протеина в течение восьми недель односторонних тренировок с отягощениями. J. Strength Cond. Res 2006; 20 (2): 284-291. Просмотреть аннотацию.
Коффи, В.Г., Мур, Д.Р., Бурд, Н.А., Ререч, Т., Стеллингверфф, Т., Гарнхэм, А.П., Филлипс, С.М., и Хоули, Дж. А. Обеспечение питательными веществами увеличивает передачу сигналов и синтез белка в скелетных мышцах человека после многократных спринтов . Eur.J Appl.Physiol 2011; 111 (7): 1473-1483.Просмотреть аннотацию.
Кук, М. Б., Рыбалка, Э., Статис, С. Г., Крибб, П. Дж. И Хейс, А. Изолят сывороточного протеина ослабляет снижение силы после эксцентрически индуцированного повреждения мышц у здоровых людей. J Int Soc Sports Nutr 2010; 7:30. Просмотреть аннотацию.
Cornish, SM, Candow, DG, Jantz, NT, Chilibeck, PD, Little, JP, Forbes, S., Abeysekara, S., и Zello, GA Конъюгированная линолевая кислота в сочетании с моногидратом креатина и добавлением сывороточного протеина во время силовых тренировок . Int J Sport Nutr Exerc.Metab 2009; 19 (1): 79-96. Просмотреть аннотацию.
Int J Sport Nutr Exerc.Metab 2009; 19 (1): 79-96. Просмотреть аннотацию.
Д’Агата, А., Бетта, П., Шакка, П., Морано, К., Пратико, Г., Каррери, Р., Куатрочки, О., и Шакка, Ф. [Роль диетической профилактики в новорожденные с риском атопии. Результаты контрольного исследования. Pediatr Med Chir 1996; 18 (5): 469-472. Просмотреть аннотацию.
Dangin, M., Guillet, C., Garcia-Rodenas, C., Gachon, P., Bouteloup-Demange, C., Reiffers-Magnani, K., Fauquant, J., Ballevre, O., and Beaufrere , B. Скорость переваривания белка по-разному влияет на прирост белка у людей в процессе старения.J. Physiol 6-1-2003; 549 (Pt 2): 635-644. Просмотреть аннотацию.
de, Seta L., Siani, P., Cirillo, G., Di, Gruttola M., Cimaduomo, L., и Coletta, S. [Профилактика аллергических заболеваний с помощью гипоаллергенной формулы: наблюдение в 24 мес. Предварительные результаты. Педиатр Мед Чир 1994; 16 (3): 251-254. Просмотреть аннотацию.
De, Simone C., Ferranti, P., Picariello, G., Scognamiglio, I. , Dicitore, A., Addeo, F., Chianese, L., и Stiuso, P. Пептиды из сыворотки сыра водяного буйвола, индуцированные гибель клеток старения через секрецию церамидов в клеточной линии аденокарциномы толстой кишки человека.Mol.Nutr Food Res 8-19-2010; Просмотреть аннотацию.
, Dicitore, A., Addeo, F., Chianese, L., и Stiuso, P. Пептиды из сыворотки сыра водяного буйвола, индуцированные гибель клеток старения через секрецию церамидов в клеточной линии аденокарциномы толстой кишки человека.Mol.Nutr Food Res 8-19-2010; Просмотреть аннотацию.
Демлинг, Р. Х. и ДеСанти, Л. Влияние гипокалорийной диеты, повышенного потребления белка и силовых тренировок на набор мышечной массы и потерю жировой массы у полицейских с избыточным весом. Энн Нутр Метаб 2000; 44 (1): 21-29. Просмотреть аннотацию.
den Hoed, CM, de Vries, AC, Mensink, PB, Dierikx, CM, Suzuki, H., Capelle, L., van, Dekken H., Ouwendijk, R., and Kuipers, EJ, орально на основе бычьих антител иммунотерапия для снижения внутрижелудочной колонизации Helicobacter pylori: рандомизированное клиническое исследование.Can.J Gastroenterol. 2011; 25 (4): 207-213. Просмотреть аннотацию.
Денисшен, К. А., Бертон, Х. У., Хорват, П. Дж., Ледди, Дж. Дж. И Браун, Р. У. Тренировка с отягощениями с добавками сои и сывороточного протеина у мужчин с гиперлипидемией. J Int Soc Sports Nutr 2009; 6: 8. Просмотреть аннотацию.
J Int Soc Sports Nutr 2009; 6: 8. Просмотреть аннотацию.
Deutz, NE, Safar, A., Schutzler, S., Memelink, R., Ferrando, A., Spencer, H., van, Helvoort A., and Wolfe, RR Можно стимулировать синтез мышечного белка у онкологических больных со специально разработанным лечебным питанием.Clin Nutr 2011; 30 (6): 759-768. Просмотреть аннотацию.
Дипвенс, К., Хаберер, Д., и Вестертерп-Плантенга, М. Различные белки и биопептиды по-разному влияют на чувство сытости и анорексигенные / орексигенные гормоны у здоровых людей. Int J Obes. (Лондон) 2008; 32 (3): 510-518. Просмотреть аннотацию.
Drouin, R., Lamiot, E., Cantin, K., Gauthier, SF, Pouliot, Y., Poubelle, PE, and Juneau, C. XP-828L (Dermylex), новый экстракт сывороточного протеина с потенциальной пользой для псориаза легкой и средней степени тяжести.Can J Physiol Pharmacol 2007; 85 (9): 943-951. Просмотреть аннотацию.
Ду, Б., Чай, В. З. и Лин, X. М. [Влияние основного белка сыворотки на минеральную плотность костей]. Чжунхуа Юй Фан Йи.Сюэ.За Чжи. 2007; 41 (2): 96-100. Просмотреть аннотацию.
Чжунхуа Юй Фан Йи.Сюэ.За Чжи. 2007; 41 (2): 96-100. Просмотреть аннотацию.
Дайер, А. Р., Бэрдок, Г. А., Карабин, И. Г., Хаас, М. К., Бойс, Дж., Алсакер, Р., и Рид, Л. С. Исследования безопасности патентованного экстракта сыворотки in vitro и in vivo. Food Chem.Toxicol. 2008; 46 (5): 1659-1665. Просмотреть аннотацию.
Элиа Д., Стадлер К., Хорват В. и Якус Дж. Влияние диеты с добавлением сои и изолята сывороточного белка на окислительно-восстановительные параметры обученных мышей. Eur.J Nutr 2006; 45 (5): 259-266. Просмотреть аннотацию.
Элиот, К. А., Кнеханс, А. В., Бембен, Д. А., Виттен, М. С., Картер, Дж. И Бембен, М. Г. Влияние добавок креатина и сывороточного протеина на состав тела у мужчин в возрасте от 48 до 72 лет во время силовых тренировок. J Nutr Health Aging 2008; 12 (3): 208-212. Просмотреть аннотацию.
Эльсадек, Х. М. и др.Действие зубной пасты и порошка с сывороточным протеином у нормальных и диабетиков. Журнал прикладных научных исследований 2009; 5 (9): 1259-1264.
Эриксен, Э.К., Холм, Х., Йенсен, Э., Обое, Р., Деволд, Т.Г., Якобсен, М., и Вегаруд, Г.Э. Различное переваривание белков сыворотки крови козьей сыворотки желудочно-кишечными ферментами человека и свиньи. Br J Nutr 2010; 104 (3): 374-381. Просмотреть аннотацию.
Errichiello, L., Pezzella, M., Santulli, L., Striano, S., Zara, F., Minetti, C., Mainardi, P., and Striano, P.Проверочное испытание сывороточного протеина альфа-лактальбумина при хроническом корковом миоклонусе. Mov Disord. 2011; 26 (14): 2573-2575. Просмотреть аннотацию.
Эстевес де Оливейра, Ф. К., Пиньейро Вольп, А. С. и Альфенас, Р. С. Влияние различных источников белка на гликемический и инсулинемический ответы. Nutr Hosp. 2011; 26 (4): 669-676. Просмотреть аннотацию.
Exl, BM, Deland, U., Secretin, MC, Preysch, U., Wall, M., and Shmerling, DH Улучшение общего состояния здоровья в неотобранной детской популяции после программы диетического вмешательства с пониженным содержанием аллергенов: ZUFF- УЧЕБНАЯ ПРОГРАММА. Часть II: рост ребенка и состояние здоровья до возраста 6 месяцев. ZUg-FrauenFeld. Eur J Nutr 2000; 39 (4): 145-156. Просмотреть аннотацию.
Часть II: рост ребенка и состояние здоровья до возраста 6 месяцев. ZUg-FrauenFeld. Eur J Nutr 2000; 39 (4): 145-156. Просмотреть аннотацию.
Exl, BM, Deland, U., Wall, M., Preysch, U., Secretin, MC, and Shmerling, DH Исследование питания в Цуг-Фрауэнфельде («Исследование Zuff»): питание с пониженным содержанием аллергенов в нормальном детском возрасте и его влияние на здоровье: результаты в возрасте шести месяцев. Nutr Res 1998; 18 (8): 1443-1462.
Фарнфилд, М. М., Тренерри, К., Кэри, К. А., и Кэмерон-Смит, Д.Аминокислотный ответ плазмы после приема различных фракций сывороточного протеина. Int J Food Sci Nutr 5-8-2008; 1-11. Просмотреть аннотацию.
Foltz, M., Ansems, P., Schwarz, J., Tasker, M.C., Lourbakos, A., and Gerhardt, C.C. Гидролизаты белков индуцируют высвобождение CCK из энтероэндокринных клеток и действуют как частичные агонисты рецептора CCK1. J. Agric.Food Chem. 2-13-2008; 56 (3): 837-843. Просмотреть аннотацию.
Фриман, С. Л., Фишер, Л., Герман, Дж. Б., Леунг, П. С., Принс, Х., Селми, К., Нагува, С. М., и Гершвин, М. Е. Молочные белки и реакция на пневмовакс у пожилых людей: рандомизированное, двойное слепое, плацебо-контролируемое пилотное исследование. Энн Н.Ю. Академик наук 2010; 1190: 97-103. Просмотреть аннотацию.
Б., Леунг, П. С., Принс, Х., Селми, К., Нагува, С. М., и Гершвин, М. Е. Молочные белки и реакция на пневмовакс у пожилых людей: рандомизированное, двойное слепое, плацебо-контролируемое пилотное исследование. Энн Н.Ю. Академик наук 2010; 1190: 97-103. Просмотреть аннотацию.
Фрестедт, Дж. Л., Зенк, Дж. Л., Кусковски, М. А., Уорд, Л. С. и Бастиан, Е. Д. Добавка сывороточного протеина увеличивает потерю жира и сохраняет мышечную массу у тучных субъектов: рандомизированное клиническое исследование на людях. Нутр Метаб (Лондон) 2008; 5: 8. Просмотреть аннотацию.
Фрид, А.Х., Нильссон, М., Холст, Дж. Дж. И Бьорк, И. М. Влияние сыворотки на глюкозу в крови и реакцию инсулина на комбинированные завтраки и обеды у пациентов с диабетом 2 типа. Am J Clin Nutr 2005; 82 (1): 69-75. Просмотреть аннотацию.
Гад, А.С., Хадрави, Ю.А., Эль-Некити, А.А., Мохамед, С.Р., Хассан, Н.С., и Абдель-Ваххаб, М.А. Антиоксидантная активность и гепатопротекторные эффекты сывороточного протеина и спирулины у крыс. Питание 8-12-2010; Просмотреть аннотацию.
Питание 8-12-2010; Просмотреть аннотацию.
Гаттас, В., Баррера, Г., Лейва, Л., де ла Маза, М.П., Банут, Д., Стинхаут, П., Классен, П., Восс, Т., и Хирш, С. [Гликемические и инсулиновые индексы смесей для зондового питания у здоровых взрослых]. Rev Med Chil. 2007; 135 (7): 879-884. Просмотреть аннотацию.
Гернигон, Г., Пиот, М., Бошер, Э., Жанте, Р., и Шук, П. Физико-химические характеристики сыворотки и стрейч воды из сыра моцарелла в сравнении с некоторыми другими сладкими сыворотками. J Dairy Sci 2009; 92 (11): 5371-5377. Просмотреть аннотацию.
Гилберт, Дж. А., Бендсен, Н. Т., Тремблей, А., и Аструп, А. Влияние белков из разных источников на состав тела. Нутр Метаб Кардиоваск. Дис. 2011; 21 Приложение 2: B16-B31. Просмотреть аннотацию.
Gouni-Berthold, I., Schulte, DM, Krone, W., Lapointe, JF, Lemieux, P., Predel, HG, and Berthold, HK. Податливая белковая матрица продукта ферментации сыворотки снижает концентрацию ТАГ у пациентов с метаболическими нарушениями. синдром: рандомизированное плацебо-контролируемое исследование. Br.J Nutr 2012; 107 (11): 1694-1706. Просмотреть аннотацию.
синдром: рандомизированное плацебо-контролируемое исследование. Br.J Nutr 2012; 107 (11): 1694-1706. Просмотреть аннотацию.
Граф, С., Эгерт, С., и Хеер, М. Влияние добавок сывороточного протеина на метаболизм: данные исследований вмешательства человека. Curr.Opin.Clin.Nutr Metab Care 2011; 14 (6): 569-580. Просмотреть аннотацию.
Грей В., Мохаммед С. Р., Смаунтас А. А., Бахул Р. и Лэндс Л. С. Улучшение глутатионового статуса у молодых взрослых пациентов с муковисцидозом, получавших сывороточный протеин. J.Cyst.Fibros. 2003; 2 (4): 195-198. Просмотреть аннотацию.
Гроблер, Л., Зигфрид, Н., Виссер, М. Э., Махлунгулу, С. С., и Волминк Дж. Вмешательства по питанию для снижения заболеваемости и смертности среди людей с ВИЧ. Cochrane.Database.Syst.Rev. 2013; 2: CD004536. Просмотреть аннотацию.
Guida, B., Piccoli, A., Trio, R., Laccetti, R., Nastasi, A., Paglione, A., Memoli, A., and Memoli, B. Ограничение количества фосфатов в диете у диализных пациентов: A новый подход к лечению гиперфосфатемии.
Gunnerud, U. J., Heinzle, C., Holst, J. J., Ostman, E.М., и Бьорк, И. М. Влияние напитков с белком и аминокислотами перед едой на гликемический и метаболический ответ при последующем комбинированном приеме пищи. PLoS.One. 2012; 7 (9): e44731. Просмотреть аннотацию.
Гуннеруд, У., Холст, Дж. Дж., Остман, Э. и Бьорк, И. Гликемическая, инсулинемическая и аминокислотная реакции в плазме крови на молочные продукты с равноуглеводным содержанием, пилотное исследование коровьего и человеческого молока. Нутр Ж, 2012; 11: 83. Просмотреть аннотацию.
Хакни, К. Дж., Брюнгер, А. Дж., И Леммер, Дж. Т. Определение времени приема белка увеличивает расход энергии через 24 часа после тренировки с отягощениями.Медико-спортивные упражнения. 2010; 42 (5): 998-1003. Просмотреть аннотацию.
Халкен, С., Хансен, К.С., Якобсен, HP, Эстманн, А., Фаеллинг, А.Э., Хансен, Л.Г., Кир, С.Р., Лассен, К., Линтруп, М., Мортенсен, С. , Ибсен, К.К. , Osterballe, O., и Host, A. Сравнение частично гидролизованной детской смеси с двумя экстенсивно гидролизованными смесями для профилактики аллергии: проспективное рандомизированное исследование. Pediatr Allergy Immunol. 2000; 11 (3): 149-161. Просмотреть аннотацию.
, Ибсен, К.К. , Osterballe, O., и Host, A. Сравнение частично гидролизованной детской смеси с двумя экстенсивно гидролизованными смесями для профилактики аллергии: проспективное рандомизированное исследование. Pediatr Allergy Immunol. 2000; 11 (3): 149-161. Просмотреть аннотацию.
Холл, У. Л., Миллуорд, Д.Дж., Лонг, С. Дж. И Морган, Л. М. Казеин и сыворотка по-разному влияют на аминокислотный профиль плазмы, секрецию желудочно-кишечных гормонов и аппетит. Br J Nutr 2003; 89 (2): 239-248. Просмотреть аннотацию.
Haraguchi, F. K., Pedrosa, M. L., de, Paula H., Dos Santos, R. C., и Silva, M. E. Оценка биологического и биохимического качества сывороточного протеина. J Med Food 2010; 13 (6): 1505-1509. Просмотреть аннотацию.
Хейс А. и Крибб П. Дж. Влияние изолята сывороточного протеина на силу, состав тела и гипертрофию мышц во время силовых тренировок.Curr Opin.Clin Nutr Metab Care 2008; 11 (1): 40-44. Просмотреть аннотацию.
Хейс, Н. П., Ким, Х., Уэллс, А. М. , Кайкенова, О., и Эванс, В. Дж. Влияние добавок сывороточного протеина и обогащенного гидролизата коллагена протеина на баланс азота и состав тела у пожилых женщин. J Am Diet.Assoc. 2009; 109 (6): 1082-1087. Просмотреть аннотацию.
, Кайкенова, О., и Эванс, В. Дж. Влияние добавок сывороточного протеина и обогащенного гидролизата коллагена протеина на баланс азота и состав тела у пожилых женщин. J Am Diet.Assoc. 2009; 109 (6): 1082-1087. Просмотреть аннотацию.
Эрнандес-Ледесма, Б., Мартин-Альварес, П. Дж., И Пуэйо, Е. Оценка спектрофотометрического метода определения активности ангиотензин-превращающего фермента: влияние типа ингибирования.J. Agric.Food Chem. 7-16-2003; 51 (15): 4175-4179. Просмотреть аннотацию.
Hoac, T., Lundh, T., Purup, S., Onning, G., Sejrsen, K., и Akesson, B. Разделение соединений селена, цинка и меди в бычьей сыворотке с использованием эксклюзионной хроматографии, связанной с масс-спектрометрия с индуктивно связанной плазмой. J. Agric.Food Chem. 5-16-2007; 55 (10): 4237-4243. Просмотреть аннотацию.
Ходжсон, Дж. М., Чжу, К., Льюис, Дж. Р., Керр, Д., Мэн, X., Солах, В., Девайн, А., Биннс, К. У., Вудман, Р. Дж., И Принс, Р.L. Долгосрочные эффекты диеты, обогащенной белком, на артериальное давление у пожилых женщин. Br.J Nutr 2012; 107 (11): 1664-1672. Просмотреть аннотацию.
Br.J Nutr 2012; 107 (11): 1664-1672. Просмотреть аннотацию.
Хоффман, Дж. Р., Ратамесс, Н. А., Транчина, К. П., Рашти, С. Л., Канг, Дж. И Файгенбаум, А. Д. Влияние патентованной белковой добавки на показатели восстановления после упражнений с отягощениями у силовых / силовых спортсменов. Amino.Acids 2010; 38 (3): 771-778. Просмотреть аннотацию.
Holmer-Jensen, J., Hartvigsen, M. L., Mortensen, L. S., Astrup, A., de, Vrese M., Холст, Дж. Дж., Томсен, К. и Хермансен, К. Острые дифференциальные эффекты пищевых белков, полученных из молока, на постпрандиальную липемию у субъектов с ожирением, не страдающих диабетом. Eur.J Clin.Nutr 2012; 66 (1): 32-38. Просмотреть аннотацию.
Hulmi, JJ, Kovanen, V., Lisko, I., Selanne, H., and Mero, AA Влияние сывороточного протеина на миостатин и ответы на экспрессию генов, связанных с клеточным циклом, на один бой с тяжелыми упражнениями у тренированных пожилых люди. Eur.J Appl Physiol 2008; 102 (2): 205-213. Просмотреть аннотацию.
Халми, Дж. Дж., Кованен, В., Селанне, Х., Кремер, В. Дж., Хаккинен, К., и Меро, А. А. Острые и долгосрочные эффекты упражнений с отягощениями с или без приема белка на гипертрофию мышц и экспрессию генов. Amino.Acids 2009; 37 (2): 297-308. Просмотреть аннотацию.
Hursel, R., van der Zee, L., and Westerterp-Plantenga, MS Влияние йогурта на завтрак с дополнительным общим сывороточным белком или обедненным казеиномакропептидом альфа-лактальбумином сывороточного белка на термогенез и аппетит, вызванный диетой подавление.Br J Nutr 2010; 103 (5): 775-780. Просмотреть аннотацию.
Hwangbo, S., Azuma, N., Kurisaki, J., and Kanno, C. Очистка и характеристика нового гликопротеина сыворотки WGP-88, который связывается с моноклональным антителом к гликопротеину PAS-4 в мембране глобулы коровьего молочного жира . Biosci.Biotechnol.Biochem. 1997; 61 (9): 1568-1574. Просмотреть аннотацию.
Искеджян, М., Шаевска, Х., Шпилденнер, Дж., Фарах, Б., и Бербари, Дж. Мета-анализ частично гидролизованной 100% -ной детской смеси на основе сыворотки по сравнению сдетское питание с интенсивным гидролизом для профилактики атопического дерматита. Curr.Med.Res Opin. 2010; 26 (11): 2599-2606. Просмотреть аннотацию.
Curr.Med.Res Opin. 2010; 26 (11): 2599-2606. Просмотреть аннотацию.
Ивамори, Т., Нукуми, Н., Ито, К., Кано, К., Наито, К., Курохмару, М., Яманучи, К., и Тоджо, Н. Бактериостатическая активность сывороточного кислого протеина (WAP ). Журнал ветеринарной медицины, 2010; 72 (5): 621-625. Просмотреть аннотацию.
Джеймс, Л. Дж., Гингелл, Р., Эванс, Г. Х. Добавление сывороточного протеина к раствору для регидратации углеводов и электролитов, принимаемому после тренировки в жару.J Athl. Поезд. 2012; 47 (1): 61-66. Просмотреть аннотацию.
Джеймс, Л. Молочный белок и восстановление баланса жидкости после тренировки. Med.Sport Sci 2012; 59: 120-126. Просмотреть аннотацию.
Джосс, А. Р. и Филипс, С. М. Влияние потребления молока и силовых тренировок на состав тела спортсменок. Med.Sport Sci 2012; 59: 94-103. Просмотреть аннотацию.
Ювонен, К. Р., Карунен, Л. Дж., Вуори, Э., Лилль, М. Э., Карху, Т., Хурадо-Акоста, А., Лааксонен, Д. Э., Микканен, Х.М., Нисканен, Л. К., Поутанен, К. С. и Херциг, К. Х. Модификация структуры модельной пищи на основе молочного белка влияет на высвобождение пептидов в кишечнике после приема пищи и их наполнение у здоровых молодых мужчин. Br.J Nutr 2011; 106 (12): 1890-1898. Просмотреть аннотацию.
К., Поутанен, К. С. и Херциг, К. Х. Модификация структуры модельной пищи на основе молочного белка влияет на высвобождение пептидов в кишечнике после приема пищи и их наполнение у здоровых молодых мужчин. Br.J Nutr 2011; 106 (12): 1890-1898. Просмотреть аннотацию.
Кальман, Д., Фельдман, С., Мартинес, М., Кригер, Д. Р., Таллон, М. Дж. Влияние источника белка и силовых тренировок на состав тела и половые гормоны. J.Int.Soc.Sports Nutr. 2007; 4: 4. Просмотреть аннотацию.
Каплан, Р. Дж., Гринвуд, К.E., Winocur, G., и Wolever, T. M. Диетический белок, углеводы и жир улучшают память у здоровых пожилых людей. Am J Clin Nutr 2001; 74 (5): 687-693. Просмотреть аннотацию.
Каппелер С., Фарах З. и Пухан З. Альтернативный сплайсинг мРНК лактофорина из лактирующей молочной железы верблюда (Camelus dromedarius). J Dairy Sci 1999; 82 (10): 2084-2093. Просмотреть аннотацию.
Касим-Каракас, С. Э., Альмарио, Р. У. и Каннингем, В. Влияние белка по сравнению с потреблением простого сахара на потерю веса при синдроме поликистозных яичников (согласно критериям Национального института здоровья). Fertil.Steril. 2009; 92 (1): 262-270. Просмотреть аннотацию.
Fertil.Steril. 2009; 92 (1): 262-270. Просмотреть аннотацию.
Кент, К. Д., Харпер, В. Дж. И Бомсер, Дж. А. Влияние изолята сывороточного белка на внутриклеточный глутатион и вызванную окислителем гибель клеток эпителия предстательной железы человека. Toxicol. In Vitro 2003; 17 (1): 27-33. Просмотреть аннотацию.
Керасиоти, Э., Кискини, А., Вескукис, А., Джамуртас, А., Цицимпику, К., Цацакис, А.М., Котедакис, Ю., Стагос, Д., Куретас, Д., и Каратанос, В. • Влияние особого углеводно-белкового жмыха на маркеры окислительного стресса у человека после изнурительной езды на велосипеде.Food Chem Toxicol. 2012; 50 (8): 2805-2810. Просмотреть аннотацию.
Kerksick, CM, Rasmussen, CJ, Lancaster, SL, Magu, B., Smith, P., Melton, C., Greenwood, M., Almada, AL, Earnest, CP, and Kreider, RB Влияние белка и добавление аминокислот для улучшения производительности и тренировочной адаптации в течение десяти недель тренировок с отягощениями. J Strength Cond. Res 2006; 20 (3): 643-653. Просмотреть аннотацию.
Просмотреть аннотацию.
Кернер, Дж. А., младший. Использование детских смесей для предотвращения или отсрочки атопических проявлений.J Pediatr Gastroenterol. Nutr 1997; 24 (4): 442-446. Просмотреть аннотацию.
Khoshoo, V., Zembo, M., King, A., Dhar, M., Reifen, R., and Pencharz, P. Частота возникновения гастроэзофагеального рефлюкса при применении смесей на основе сыворотки и казеина у младенцев и детей с тяжелые неврологические нарушения. J Pediatr Gastroenterol. Nutr 1996; 22 (1): 48-55. Просмотреть аннотацию.
Ким, Дж., Ко, Ю., Парк, Ю. К., Ким, Н. И., Ха, В. К., и Чо, Ю. Диетическое влияние ферментированного молока, обогащенного лактоферрином, на поверхностный липид кожи и клиническое улучшение вульгарных угрей.Питание 2010; 26 (9): 902-909. Просмотреть аннотацию.
Koopman, R., Verdijk, LB, Beelen, M., Gorselink, M., Kruseman, AN, Wagenmakers, AJ, Kuipers, H., and van Loon, LJ Совместное употребление лейцина с белком не увеличивает скорость синтеза мышечного белка после тренировки у пожилых мужчин. Br.J Nutr 2008; 99 (3): 571-580. Просмотреть аннотацию.
Br.J Nutr 2008; 99 (3): 571-580. Просмотреть аннотацию.
Крюгер, К.Л., Марано, К.М., Морита, Ю., Такада, Ю., Каваками, Х., Кобаяси, Т., Сунага, М., Фурукава, М., и Кавамура, К. Оценка безопасности молока основная белковая фракция.Food Chem.Toxicol. 2007; 45 (7): 1301-1307. Просмотреть аннотацию.
Кумар П., Ядав С., Сринивасан А., Бхатиа К. Л. и Сингх Т. П. Новый белок 40 кДа из секрета молочной железы коз: очистка, кристаллизация и предварительные исследования дифракции рентгеновских лучей. Acta Crystallogr.D.Biol Crystallogr. 2001; 57 (Pt 9): 1332-1333. Просмотреть аннотацию.
Куме, Х., Окадзаки, К. и Сасаки, Х. Гепатопротекторные эффекты сывороточного протеина на D-галактозамин-индуцированный гепатит и фиброз печени у крыс.Biosci.Biotechnol.Biochem. 2006; 70 (5): 1281-1285. Просмотреть аннотацию.
Lam, B.C. и Yeung, C.Y. Влияние грудного молока, детской смеси и гипоаллергенной смеси на частоту проявления атопии у младенцев с высоким риском. 1992;
Lan-Pidhainy, X. и Wolever, T. M. Гипогликемический эффект жиров и белков не ослабляется инсулинорезистентностью. Am J Clin Nutr 2010; 91 (1): 98-105. Просмотреть аннотацию.
и Wolever, T. M. Гипогликемический эффект жиров и белков не ослабляется инсулинорезистентностью. Am J Clin Nutr 2010; 91 (1): 98-105. Просмотреть аннотацию.
Лэндс, Л. К., Искандар, М., Бодуан, Н., Михан, Б., Даулетбаев, Н., и Berthiuame, Y. Пищевые добавки с сывороткой под давлением у пациентов с муковисцидозом. J Med Food 2010; 13 (1): 77-82. Просмотреть аннотацию.
Laviolette, L., Lands, LC, Dauletbaev, N., Saey, D., Milot, J., Provencher, S., LeBlanc, P., and Maltais, F. Комбинированный эффект диетических добавок с сывороткой под давлением и физические упражнения при хронической обструктивной болезни легких: рандомизированное контролируемое двойное слепое пилотное исследование. J Med Food 2010; 13 (3): 589-598. Просмотреть аннотацию.
Ли Ю.М., Скурк Т., Хенниг М. и Хаунер Х. Влияние молочного напитка с добавлением сывороточных пептидов на артериальное давление у пациентов с легкой гипертензией. Eur.J Nutr 2007; 46 (1): 21-27. Просмотреть аннотацию.
Lorenzen, J. , Frederiksen, R., Hoppe, C., Hvid, R., and Astrup, A. Влияние белков молока на регуляцию аппетита и термогенез, индуцированный диетой. Eur.J Clin.Nutr 2012; 66 (5): 622-627. Просмотреть аннотацию.
, Frederiksen, R., Hoppe, C., Hvid, R., and Astrup, A. Влияние белков молока на регуляцию аппетита и термогенез, индуцированный диетой. Eur.J Clin.Nutr 2012; 66 (5): 622-627. Просмотреть аннотацию.
Loss, G., Apprich, S., Waser, M., Kneifel, W., Genuneit, J., Buchele, G., Weber, J., Sozanska, B., Danielewicz, H., Horak, E., van Neerven, RJ, Heederik, D., Lorenzen, PC, von, Mutius E., and Braun-Fahrlander, C. Защитный эффект потребления молока на ферме по детской астме и атопии: исследование GABRIELA. J Allergy Clin.Immunol. 2011; 128 (4): 766-773. Просмотреть аннотацию.
Лотиан, Б., Грей, В., Кимофф, Р. Дж. И Лэндс, Л. С. Лечение обструктивного заболевания дыхательных путей добавкой белка-донора цистеина: описание случая. Chest 2000; 117 (3): 914-916. Просмотреть аннотацию.
Лотиан, Дж. Б., Грей, В., и Лэндс, Л. С. Влияние сывороточного протеина на модуляцию иммунного ответа у детей с атопической астмой. Int J Food Sci Nutr 2006; 57 (3-4): 204-211. Просмотреть аннотацию.
Low, P. P., Rutherfurd, K. J., Gill, H. S. и Cross, M. L. Влияние диетического концентрата сывороточного белка на первичные и вторичные реакции антител у иммунизированных мышей BALB / c. Int Immunopharmacol. 2003; 3 (3): 393-401. Просмотреть аннотацию.
Луховый Б. Л., Ахаван Т., Андерсон Г.H. Сывороточные белки в регуляции приема пищи и насыщения. J Am Coll. Nutr 2007; 26 (6): 704S-712S. Просмотреть аннотацию.
Lum, C. and et al. Исследование добавок сывороточного протеина к физической работоспособности и качеству жизни пожилых пациентов с хронической обструктивной болезнью легких. Австралазийский журнал по проблемам старения 2007; 26 (4): 168-172.
Ма, Дж., Стивенс, Дж. Э., Цукьер, К., Мэддокс, А. Ф., Уишарт, Дж. М., Джонс, К. Л., Клифтон, П. М., Хоровиц, М., и Рейнер, К. К. Влияние предварительной нагрузки белком на опорожнение желудка. гликемия и гормоны кишечника после углеводной еды при диабете 2 типа, контролируемом диетой.Уход за диабетом 2009; 32 (9): 1600-1602. Просмотреть аннотацию.
Просмотреть аннотацию.
Мабин, Д. К., Сайкс, А. Э. и Дэвид, Т. Дж. Контролируемое испытание диеты из нескольких продуктов при тяжелом атопическом дерматите. Arch Dis. Детский 1995; 73 (3): 202-207. Просмотреть аннотацию.
Мабин, Д. К., Сайкс, А. Э. и Дэвид, Т. Дж. Содержание питательных веществ в диете с небольшим количеством пищевых продуктов при атопическом дерматите. Arch Dis. Детский 1995; 73 (3): 208-210. Просмотреть аннотацию.
Macdermid, P. W. и Stannard, S. R. Диета с добавлением сыворотки и высоким содержанием белка по сравнению с диетой с высоким содержанием углеводов: влияние на выносливость при езде на велосипеде.Int J Sport Nutr Exerc.Metab 2006; 16 (1): 65-77. Просмотреть аннотацию.
Манкузо, М., Орсуччи, Д., Логерфо, А., Рокки, А., Петроцци, Л., Нести, К., Галетта, Ф., Санторо, Г., Мурри, Л., и Сицилиано, G. Биомаркеры оксидативного стресса при митохондриальных миопатиях, базально и после приема донорских добавок цистеина. J Neurol. 2010; 257 (5): 774-781. Просмотреть аннотацию.
Просмотреть аннотацию.
Манцони, П., Децембрино, Л., Стольфи, И., Пуни, Л., Ринальди, М., Каттани, С., Ромео, М. Г., Месснер, Х., Лафоргия, Н., Вагнарелли, Ф., Мемо, Л., Бординьон, Л., Сайя, О.С., Мауле, М., Галло, Э., Мостерт, М., Маньяни, К., Кверчия, М., Боллани, Л., Педичино, Р., Рензулло, Л., Бетта, П., Феррари, Ф., Магальди, Р., Моска, Ф., Стронати, М., и Фарина, Д. Лактоферрин и профилактика позднего сепсиса у недоношенных новорожденных. Early Hum Dev. 2010; 86 Приложение 1: 59-61. Просмотреть аннотацию.
Марини А., Агости М., Мотта Г. и Моска Ф. Влияние программы диетической и экологической профилактики на частоту аллергических симптомов у младенцев с высоким атопическим риском: трехлетнее наблюдение.Acta Paediatr.Suppl 1996; 414: 1-21. Просмотреть аннотацию.
Mark, A. B., Hoppe, C., Michaelsen, K. F. и Molgaard, C. Белки и минералы, полученные из молока, изменяют уровень остеокальцина в сыворотке крови у мальчиков препубертатного возраста через 7 дней. Nutr Res 2010; 30 (8): 558-564. Просмотреть аннотацию.
Просмотреть аннотацию.
Маркус К. Р., Оливье Б. и де Хаан Е. Х. Сывороточный протеин, богатый альфа-лактальбумином, увеличивает соотношение триптофана в плазме к сумме других крупных нейтральных аминокислот и улучшает когнитивные способности у подверженных стрессу субъектов.Am.J.Clin.Nutr. 2002; 75 (6): 1051-1056. Просмотреть аннотацию.
Маркус, ЧР, Оливье, Б., Панхуизен, Дж. Э., Ван дер Гугтен, Дж., Аллес, М. С., Туйтен, А., Вестенберг, Х. Г., Феккес, Д., Коппешар, Х. Ф., и де Хаан, Э Бычий белок альфа-лактальбумин увеличивает соотношение триптофана к другим крупным нейтральным аминокислотам в плазме, а у уязвимых субъектов повышает активность серотонина в мозге, снижает концентрацию кортизола и улучшает настроение в условиях стресса. Am J Clin Nutr 2000; 71 (6): 1536-1544.Просмотреть аннотацию.
Макинтош, Г. Х., Регестер, Г. О., Ле Леу, Р. К., Ройл, П. Дж. И Смитерс, Г. В. Молочные белки защищают от индуцированного диметилгидразином рака кишечника у крыс. J Nutr 1995; 125 (4): 809-816. Просмотреть аннотацию.
Просмотреть аннотацию.
Мертенс, Дж., Шток, С., Лунген, М., фон, Берг А., Крамер, У., Филипьяк-Питтрофф, Б., Генрих, Дж., Колецко, С., Грубль, А., Wichmann, HE, Bauer, CP, Reinhardt, D., Berdel, D., and Gerber, A. Является ли профилактика атопической экземы с помощью гидролизованных формул рентабельной? Оценка экономики здравоохранения из Германии.Pediatr.Allergy Immunol. 2012; 23 (6): 597-604. Просмотреть аннотацию.
Micke, P., Beeh, K. M., and Buhl, R. Влияние длительного приема сывороточных белков на уровни глутатиона в плазме у ВИЧ-инфицированных пациентов. Eur.J Nutr 2002; 41 (1): 12-18. Просмотреть аннотацию.
Micke, P., Beeh, K. M., Schlaak, J. F. и Buhl, R. Пероральный прием сывороточных белков повышает уровень глутатиона в плазме у ВИЧ-инфицированных пациентов. Eur.J Clin Invest 2001; 31 (2): 171-178. Просмотреть аннотацию.
Моджтахеди, М.К., Торп, М. П., Карампинос, Д. К., Джонсон, С. Л., Лейман, Д. К., Георгиадис, Дж. Г. и Эванс, Е. М. Влияние повышенного потребления белка во время ограничения энергии на изменения в составе тела и физических функциях у пожилых женщин. Ж. Геронтол. Биол. И мед. Науки 2011; 66 (11): 1218-1225. Просмотреть аннотацию.
Ж. Геронтол. Биол. И мед. Науки 2011; 66 (11): 1218-1225. Просмотреть аннотацию.
Молгаард, К., Ларнкьяер, А., Арнберг, К., и Михаэльсен, К. Ф. Молоко и рост у детей: эффекты сыворотки и казеина. Nestle.Nutr Workshop Ser.Pediatr.Программа. 2011; 67: 67-78. Просмотреть аннотацию.
Мур, Д. Р., Атертон, П. Дж., Ренни, М. Дж., Тарнопольски, М. А., и Филлипс, С. М. Упражнения с отягощениями усиливают передачу сигналов mTOR и MAPK в мышцах человека по сравнению с тем, что наблюдается в состоянии покоя после приема болюсного белка. Acta Physiol (Oxf), 27 сентября 2010 г .; Просмотреть аннотацию.
Мур, Д. Р., Танг, Дж. Э., Бурд, Н. А., Ререч, Т., Тарнопольски, М. А., и Филлипс, С. М. Дифференциальная стимуляция синтеза миофибриллярных и саркоплазматических белков с приемом белка в покое и после упражнений с отягощениями.J. Physiol 2-15-2009; 587 (Pt 4): 897-904. Просмотреть аннотацию.
Морено, Ю. Ф., Сгарбиери, В. К., да Силва, М. Н., Торо, А. А., и Вилела, М. М. Особенности приема добавок концентрата сывороточного протеина у детей с быстро прогрессирующей ВИЧ-инфекцией. Дж. Троп. Педиатр 2006; 52 (1): 34-38. Просмотреть аннотацию.
Особенности приема добавок концентрата сывороточного протеина у детей с быстро прогрессирующей ВИЧ-инфекцией. Дж. Троп. Педиатр 2006; 52 (1): 34-38. Просмотреть аннотацию.
Морифудзи, М., Ишизака, М., Баба, С., Фукуда, К., Мацумото, Х., Кога, Дж., Канеги, М., и Хигучи, М. Сравнение различных источников и степени гидролиза пищевого белка: влияние на аминокислоты, дипептиды и инсулиновые реакции в плазме у людей.J. Agric.Food Chem. 8-11-2010; 58 (15): 8788-8797. Просмотреть аннотацию.
Mortensen, LS, Hartvigsen, ML, Brader, LJ, Astrup, A., Schrezenmeir, J., Holst, JJ, Thomsen, C., and Hermansen, K. Дифференциальное влияние качества белка на постпрандиальную липемию в ответ на жирная пища при диабете 2 типа: сравнение сывороточного, казеинового, глютенового и трескового белка. Am J Clin Nutr 2009; 90 (1): 41-48. Просмотреть аннотацию.
Маллинз, Н. М. и Синнинг, У. Э. Влияние тренировок с отягощениями и протеиновых добавок на метаболизм костей у молодых взрослых женщин. Нутр Метаб (Лондон) 8-17-2005; 2: 19. Просмотреть аннотацию.
Нутр Метаб (Лондон) 8-17-2005; 2: 19. Просмотреть аннотацию.
Мерфи К. и Миллер Б. Ф. Потребление белка после аэробных упражнений увеличивает обмен белка во всем теле у пожилых людей. Appl Physiol Nutr Metab 2010; 35 (5): 583-590. Просмотреть аннотацию.
Мюррей, Б. А., Уолш, Д. Дж., И Фитцджеральд, Р. Дж. Модификация анализа фуранакрилоил-L-фенилаланилглицилглицина для определения активности ингибирования ангиотензин-I-превращающего фермента. Журнал биохимии. Биофиз. Методы 5-31-2004; 59 (2): 127-137.Просмотреть аннотацию.
Нагаока, К., Аоки, Ф., Хаяси, М., Мурои, Ю., Сакураи, Т., Ито, К., Икава, М., Окабе, М., Имакава, К., и Сакаи, Оксидаза S. L-аминокислоты играет решающую роль в защите хозяина в молочных железах. FASEB J 2009; 23 (8): 2514-2520. Просмотреть аннотацию.
Нарисава, Н., Фурукава, С., Кавараи, Т., Охиси, К., Канда, С., Кимидзима, К., Негиси, С., Огихара, Х., и Ямасаки, М. Эффект снятого обезжиренного материала молоко и его фракции на инактивацию Escherichia coli K12 обработкой высоким гидростатическим давлением. Int J Food Microbiol. 5-10-2008; 124 (1): 103-107. Просмотреть аннотацию.
Int J Food Microbiol. 5-10-2008; 124 (1): 103-107. Просмотреть аннотацию.
Nash, MS, Meltzer, NM, Martins, SC, Burns, PA, Lindley, SD, and Field-Fote, EC Пищевые добавки после передвижения у лиц с неполными повреждениями спинного мозга: рандомизированный, двойной слепой, плацебо-контролируемый случай серии. Arch Phys.Med Rehabil. 2007; 88 (2): 228-233. Просмотреть аннотацию.
Nentwich, I., Michkova, E., Nevoral, J., Urbanek, R., and Szepfalusi, Z. Специфические клеточные и гуморальные иммунные реакции коровьего молока и кожные симптомы атопии у младенцев из семей с атопией, которых кормили частично (pHF ) или экстенсивно (eHF) гидролизованная детская смесь.Аллергия 2001; 56 (12): 1144-1156. Просмотреть аннотацию.
Нильссон М., Холст Дж. Дж. И Бьорк И. М. Метаболические эффекты смесей аминокислот и сывороточного белка у здоровых субъектов: исследования с использованием напитков, эквивалентных глюкозе. Am J Clin Nutr 2007; 85 (4): 996-1004. Просмотреть аннотацию.
Numan, S. C., Veldkamp, P., Kuijper, E.J., van den Berg, R.J., van Dissel, J.T. Диарея, связанная с Clostridium difficile: бычий сывороточный белок против Clostridium difficile для помощи в предотвращении рецидивов.Кишечник 2007; 56 (6): 888-889. Просмотреть аннотацию.
C., Veldkamp, P., Kuijper, E.J., van den Berg, R.J., van Dissel, J.T. Диарея, связанная с Clostridium difficile: бычий сывороточный белок против Clostridium difficile для помощи в предотвращении рецидивов.Кишечник 2007; 56 (6): 888-889. Просмотреть аннотацию.
Онер, О. З., Огунк, А. В., Цинги, А., Уяр, С. Б., Ялчин, А. С. и Актан, А. О. Кормление сывороткой подавляет измерение окислительного стресса при экспериментальной ожоговой травме. Surg Today 2006; 36 (4): 376-381. Просмотреть аннотацию.
Осборн Д. А. и Синн Дж. Формулы, содержащие гидролизованный белок, для профилактики аллергии и пищевой непереносимости у младенцев. Cochrane.Database.Syst.Rev. 2006; (4): CD003664. Просмотреть аннотацию.
Паддон-Джонс, Д., Шеффилд-Мур, М., Катсанос, С. С., Чжан, Х. Дж., И Вулф, Р. Р. Дифференциальная стимуляция синтеза мышечного белка у пожилых людей после изокалорийного приема аминокислот или сывороточного белка. Exp.Gerontol. 2006; 41 (2): 215-219. Просмотреть аннотацию.
Пал С. и Эллис В. Острые эффекты изолята сывороточного протеина на артериальное давление, сосудистую функцию и маркеры воспаления у женщин в постменопаузе с избыточным весом. Br.J Nutr 2011; 105 (10): 1512-1519. Просмотреть аннотацию.
Пал С. и Эллис В.Острое влияние четырехразового белкового питания на инсулин, глюкозу, аппетит и потребление энергии у худых мужчин. Br J Nutr 2010; 104 (8): 1241-1248. Просмотреть аннотацию.
Пал, С. и Эллис, В. Хронические эффекты сывороточных белков на кровяное давление, сосудистую функцию и маркеры воспаления у людей с избыточным весом. Ожирение. (Серебро. Весна) 2010; 18 (7): 1354-1359. Просмотреть аннотацию.
Пал С., Эллис В. и Дхаливал С. Влияние изолята сывороточного протеина на состав тела, липиды, инсулин и глюкозу у людей с избыточным весом и ожирением.Br J Nutr 2010; 104 (5): 716-723. Просмотреть аннотацию.
Пал С., Эллис В. и Хо С. Острые эффекты изолята сывороточного протеина на факторы риска сердечно-сосудистых заболеваний у женщин с избыточным весом в постменопаузе. Атеросклероз 2010; 212 (1): 339-344. Просмотреть аннотацию.
Атеросклероз 2010; 212 (1): 339-344. Просмотреть аннотацию.
Пенттила, И.А., Чжан, М.Ф., Бейтс, Э., Регестер, Г., Рид, Л.С., и Зола, Х. Иммунная модуляция у детенышей сосущих крыс с помощью экстракта фактора роста, полученного из молочной сыворотки. J Dairy Res 2001; 68 (4): 587-599. Просмотреть аннотацию.
Петерсен, Б.Л., Уорд, Л. С., Бастиан, Э. Д., Дженкинс, А. Л., Кэмпбелл, Дж., И Вуксан, В. Добавка сывороточного протеина снижает гликемию после приема пищи. Nutr J 2009; 8: 47. Просмотреть аннотацию.
Пфеффер, Г., Маджамаа, К., Тернбулл, Д. М., Торберн, Д., и Чинери, П. Ф. Лечение митохондриальных нарушений. Кокрановская база данных.Syst.Rev. 2012; 4: CD004426. Просмотреть аннотацию.
Филлипс, С. М. Наука о мышечной гипертрофии: подсчет диетического белка. Proc.Nutr Soc. 2011; 70 (1): 100-103. Просмотреть аннотацию.
Пихланто-Леппала, А., Коскинен, П., Пийлола, К., Тупасела, Т., и Корхонен, Х. Свойства фермента, преобразующего ангиотензин I, в отношении переваривания сывороточного белка: концентрация и характеристика активных пептидов. J Dairy Res 2000; 67 (1): 53-64. Просмотреть аннотацию.
J Dairy Res 2000; 67 (1): 53-64. Просмотреть аннотацию.
Пинс, Дж. Дж. И Кинан, Дж. М. Эффекты сывороточных пептидов на факторы риска сердечно-сосудистых заболеваний. J. Clin Hypertens. (Гринвич) 2006; 8 (11): 775-782. Просмотреть аннотацию.
Планас, М., Альварес, Дж., Гарсия-Перис, П.А., де ла Куэрда, С., де, Лукас П., Кастелла, М., Кансеко, Ф., и Рейес, Л. Нутриционная поддержка и качество жизни у пациентов со стабильной хронической обструктивной болезнью легких (ХОБЛ). Clin Nutr 2005; 24 (3): 433-441. Просмотреть аннотацию.
Порч М.С., Шахан А. и Лейва Л. Влияние грудного молока, сои или двух гидролизованных смесей на развитие аллергических проявлений у младенцев из группы риска. Nutr Res 1998; 18 (1413): 1424.
Potier, M., Fromentin, G., Calvez, J., Benamouzig, R., Martin-Rouas, C., Pichon, L., Tome, D., and Marsset-Baglieri, A. Обычная сырная закуска с высоким содержанием белка, умеренной энергией, энергетически компенсируется у людей. Br J Nutr 2009; 102 (4): 625-631. Просмотреть аннотацию.
Просмотреть аннотацию.
Poulin, Y., Bissonnette, R., Juneau, C., Cantin, K., Drouin, R., and Poubelle, PE XP-828L в лечении псориаза легкой и средней степени тяжести: рандомизированное, двойное слепое, плацебо -контролируемое исследование. Альтернативная медицина Rev.2007; 12 (4): 352-359. Просмотреть аннотацию.
Пулен, Ю., Биссоннетт, Р., Juneau, C., Cantin, K., Drouin, R., and Poubelle, P. E. XP-828l в лечении псориаза легкой и средней степени тяжести: рандомизированное, двойное слепое, плацебо-контролируемое исследование. Дж. Кутан. Med Surg 2006; 10 (5): 241-248. Просмотреть аннотацию.
Принслоо, Дж. Г., Конради, Дж. М., Одендаал, В. А., и Ван дер Уолт, В. Х. [Начало лечения пациентов с квашиоркором с использованием молочного сывороточного продукта. Сравнение с коровьим молоком. S.Afr.Med J 10-22-1983; 64 (18): 710-712. Просмотреть аннотацию.
Рединг, Дж. Л., Мейерс, А.Ф. и Вякарнам А. Кислотные белки сыворотки (WAP): новые модуляторы врожденного иммунитета к ВИЧ-инфекции. Curr.Opin.HIV.AIDS 2012; 7 (2): 172-179. Просмотреть аннотацию.
Curr.Opin.HIV.AIDS 2012; 7 (2): 172-179. Просмотреть аннотацию.
Reitelseder, S., Agergaard, J., Doessing, S., Helmark, IC, Lund, P., Kristensen, NB, Frystyk, J., Flyvbjerg, A., Schjerling, P., van, Hall G. , Кьяер, М., и Холм, Л. Сыворотка и казеин, меченные L- [1-13C] лейцином, и синтез мышечного белка: эффект упражнений с отягощениями и приема белка. Am J Physiol Endocrinol. Metab 2011; 300 (1): E231-E242.Просмотреть аннотацию.
Райс Б. Х., Чифелли К. Дж., Пикоски М. А. и Миллер Г. Д. Молочные компоненты и факторы риска кардиометаболического синдрома: последние данные и возможности для будущих исследований. Adv. Nutr 2011; 2 (5): 396-407. Просмотреть аннотацию.
Розин, А. П., Шапира, Д., Браун-Москович, Ю., Марковиц, Д., Влодавский, Е., и Балбир-Гурман, А. Ультразвуковая визуализация фасциита, вызванного добавками для бодибилдинга. Am J Med Sci 2008; 335 (3): 242-245. Просмотреть аннотацию.
Ржехак, П., Саусенталер, С., Колецко, С., Рейнхард, Д., фон, Берг А. , Крамер, У., Бердел, Д., Боллрат, К., Грубл, А., Бауэр, С.П., Вихман, Х.Э., и Heinrich, J. Долгосрочное влияние смесей для грудных детей с гидролизованным белком на рост — расширенное наблюдение до возраста 10 лет: результаты исследования German Infant Nutritional Intervention (GINI). Am J Clin. Nutr 2011; 94 (6 приложений): 1803S-1807S. Просмотреть аннотацию.
, Крамер, У., Бердел, Д., Боллрат, К., Грубл, А., Бауэр, С.П., Вихман, Х.Э., и Heinrich, J. Долгосрочное влияние смесей для грудных детей с гидролизованным белком на рост — расширенное наблюдение до возраста 10 лет: результаты исследования German Infant Nutritional Intervention (GINI). Am J Clin. Nutr 2011; 94 (6 приложений): 1803S-1807S. Просмотреть аннотацию.
Салями, М., Мусави-Мовахеди, А. А., Эхсани, М. Р., Юсефи, Р., Хертле, Т., Чоберт, Дж. М., Разави, С.H., Henrich, R., Balalaie, S., Ebadi, S.A., Pourtakdoost, S., и Niasari-Naslaji, A. Улучшение антимикробной и антиоксидантной активности белков верблюжьей и коровьей сыворотки за счет ограниченного протеолиза. J. Agric.Food Chem. 3-24-2010; 58 (6): 3297-3302. Просмотреть аннотацию.
Саталофф, Р. Т., Биттерманн, Т., Маркс, Л., Лурье, Д. и Хокшоу, М. Влияние увеличения глутатиона на нейросенсорную тугоухость. Ухо-носовое горло J 2010; 89 (9): 422-433. Просмотреть аннотацию.
Саттлер, Ф.R., Rajicic, N., Mulligan, K. , Yarasheski, KE, Koletar, SL, Zolopa, A., Alston, Smith B., Zackin, R., and Bistrian, B. Оценка веса высокопротеиновых добавок -стабильные ВИЧ-положительные субъекты с историей потери веса: рандомизированное двойное слепое многоцентровое исследование. Am J Clin Nutr 2008; 88 (5): 1313-1321. Просмотреть аннотацию.
, Yarasheski, KE, Koletar, SL, Zolopa, A., Alston, Smith B., Zackin, R., and Bistrian, B. Оценка веса высокопротеиновых добавок -стабильные ВИЧ-положительные субъекты с историей потери веса: рандомизированное двойное слепое многоцентровое исследование. Am J Clin Nutr 2008; 88 (5): 1313-1321. Просмотреть аннотацию.
Сэвидж, К., Критас, С., Шварцер, А., Дэвидсон, Г., и Омари, Т. Энтеральная смесь на основе сывороточного казеина и функция желудочно-кишечного тракта у детей с церебральным параличом.JPEN J Parenter. Enteral Nutr 2012; 36 (1 приложение): 118S-123S. Просмотреть аннотацию.
Шмитт, Дж. А., Йориссен, Б. Л., Дай, Л., Маркус, К. Р., Дойц, Н. Е. и Ридель, В. Дж. Функция памяти у женщин с предменструальными жалобами и эффект серотонинергической стимуляции путем однократного введения альфа-лактальбумина. J. Psychopharmacol 2005; 19 (4): 375-384. Просмотреть аннотацию.
См. Д., Мейсон С. и Рошан Р. Повышение фактора некроза опухоли альфа (TNF-альфа) и функции естественных киллеров (NK) с использованием интегративного подхода при поздних стадиях рака. Иммунол. Инвест 2002; 31 (2): 137-153. Просмотреть аннотацию.
Иммунол. Инвест 2002; 31 (2): 137-153. Просмотреть аннотацию.
Шейхолеслами, Ватани Д. и Ахмади Кани, Гользар Ф. Изменения антиоксидантного статуса и факторов риска сердечно-сосудистых заболеваний у молодых мужчин с избыточным весом после шести недель приема изолята сывороточного протеина и силовых тренировок. Аппетит 2012; 59 (3): 673-678. Просмотреть аннотацию.
Сильва, Л. Б., Моурао, Л. Ф., Сильва, А. А., Лима, Н. М., Алмейда, С. Р., Франка-младший, М. С., Нуччи, А., и Амайя-Фарфан, Дж. Влияние пищевой добавки с белками молочной сыворотки на боковую амиотрофию. больные склерозом.Arq Neuropsiquiatr. 2010; 68 (2): 263-268. Просмотреть аннотацию.
Смит А. Э., Фукуда Д. Х., Кендалл К. Л. и Стаут Дж. Р. Влияние предтренировочной добавки, содержащей кофеин, креатин и аминокислоты, в течение трех недель высокоинтенсивных упражнений на аэробную и анаэробную производительность. J Int Soc Sports Nutr 2010; 7:10. Просмотреть аннотацию.
Смит, Т. Дж., Монтейн, С. Дж. , Андерсон, Д. и Янг, А. J. Аминокислотные реакции в плазме после употребления напитков с различным типом белка.Int J Sport Nutr Exerc.Metab 2009; 19 (1): 1-17. Просмотреть аннотацию.
, Андерсон, Д. и Янг, А. J. Аминокислотные реакции в плазме после употребления напитков с различным типом белка.Int J Sport Nutr Exerc.Metab 2009; 19 (1): 1-17. Просмотреть аннотацию.
Солах, В.А., Керр, Д.А., Адикара, С.Д., Мэн, X., Биннс, К.У., Чжу, К., Девайн, А., и Принс, Р.Л. Различия в эффекте насыщения продуктами на основе альгината и сывороточного протеина . Аппетит 2010; 54 (3): 485-491. Просмотреть аннотацию.
Соуза, Г. Т., Лира, Ф. С., Роза, Дж. К., де Оливейра, Э. П., Ояма, Л. М., Сантос, Р. В., и Пиментел, Г. Д. Диетический сывороточный протеин снижает несколько факторов риска метаболических заболеваний: обзор.Lipids Health Dis. 2012; 11: 67. Просмотреть аннотацию.
Spieldenner, J., Belli, D., Dupont, C., Haschke, F., Iskedjian, M., Nevot, Falco S., Szajewska, H., and von, Berg A. Частично гидролизованный 100% сыворотка детские смеси и профилактика атопического дерматита: сравнительный фармакоэкономический анализ. Ann.Nutr Metab 2011; 59 Дополнение 1: 44-52. Просмотреть аннотацию.
Просмотреть аннотацию.
Стелла, В. и Постер, Э. [Оценка антирадикального защитного эффекта мультиферментированной молочной сыворотки с повторной дозировкой у крыс].C.R.Seances Soc Biol Fil. 1995; 189 (6): 1191-1197. Просмотреть аннотацию.
Су, Дж., Прескотт, С., Синн, Дж., Танг, М., Смит, П., Хайне, Р. Г., Спилденнер, Дж., И Искеджян, М. Экономическая эффективность частично гидролизованной формулы для профилактика атопического дерматита в Австралии. J Med.Econ. 2012; 15 (6): 1064-1077. Просмотреть аннотацию.
Сугавара, К., Такахаши, Х., Кашивагура, Т., Ямада, К., Янагида, С., Хомма, М., Дайрики, К., Сасаки, Х., Кавагоши, А., Сатаке, М. ., и Шиоя, Т. Эффект противовоспалительной добавки с сывороточным пептидом и лечебной физкультуры у пациентов с ХОБЛ.Respir.Med. 2012; 106 (11): 1526-1534. Просмотреть аннотацию.
Санделл Дж., Халми Дж. И Росси Дж. [Сывороточный протеин и креатин в качестве пищевых добавок]. Duodecim 2011; 127 (7): 700-705. Просмотреть аннотацию.
Szajewska, H. и Horvath, A. Метаанализ доказательств использования частично гидролизованной смеси 100% сыворотки для профилактики аллергических заболеваний. Curr Med Res Opin. 2010; 26 (2): 423-437. Просмотреть аннотацию.
Тессари, П., Киванука, Э., Кристини, М., Зарамелла, М., Энслен, М., Зурло, С., и Гарсия-Роденас, С. Медленные белки по сравнению с быстрыми в стимуляции ответа бета-клеток и активации энтероинсулярной оси при диабете 2 типа. Diabetes Metab Res Rev.2007; 23 (5): 378-385. Просмотреть аннотацию.
Тестирование пищевой добавки при истощении. Res Initiat.Treat.Action. 1999; 5 (1): 18. Просмотреть аннотацию.
Томпсон, Дж. М., Бретт, А. и Роуз, С. Дж. Диетическое лечение кишечной лимфангиэктазии, осложненной синдромом короткой кишки. Hum Nutr Appl Nutr 1986; 40 (2): 136-140.Просмотреть аннотацию.
Типтон, К. Д., Эллиотт, Т. А., Кри, М. Г., Вольф, С. Е., Сэнфорд, А. П. и Вулф, Р. Р. Прием казеина и сывороточных белков приводит к анаболизму мышц после упражнений с отягощениями. Медико-спортивные упражнения. 2004; 36 (12): 2073-2081. Просмотреть аннотацию.
Медико-спортивные упражнения. 2004; 36 (12): 2073-2081. Просмотреть аннотацию.
Тоба, Ю., Такада, Ю., Мацуока, Ю., Морита, Ю., Мотури, М., Хираи, Т., Сугури, Т., Аое, С., Каваками, Х., Кумегава, М. ., Takeuchi, A., and Itabashi, A. Основной белок молока способствует формированию костей и подавляет резорбцию костей у здоровых взрослых мужчин.Biosci.Biotechnol.Biochem. 2001; 65 (6): 1353-1357. Просмотреть аннотацию.
Trocki, O., Mochizuki, H., Dominioni, L., and Alexander, J. W. Неповрежденный белок по сравнению со свободными аминокислотами в питательной поддержке термически травмированных животных. JPEN J. Parenter. Enteral Nutr 1986; 10 (2): 139-145. Просмотреть аннотацию.
Tsai, Y. T., Chou, C. C. и Hsieh, K. H. Влияние гипоаллергенной смеси на возникновение аллергических заболеваний у младенцев из группы высокого риска. Чжунхуа Минь Го Сяо, эр.Ке И Сюэ Хуэй За Чжи 1991; 32 (3): 137-144.Просмотреть аннотацию.
Tseng, YM, Chen, SY, Chen, CH, Jin, YR, Tsai, SM, Chen, IJ, Lee, JH, Chiu, CC и Tsai, LY Эффекты индуцированных алкоголем мононуклеарных клеток периферической крови человека (PBMC ) предварительно обработанный концентрат сывороточного протеина (WPC) на окислительное повреждение. J. Agric.Food Chem. 9-10-2008; 56 (17): 8141-8147. Просмотреть аннотацию.
J. Agric.Food Chem. 9-10-2008; 56 (17): 8141-8147. Просмотреть аннотацию.
Tseng, YM, Lin, SK, Hsiao, JK, Chen, IJ, Lee, JH, Wu, SH и Tsai, концентрат сывороточного белка LY способствует выработке глутатиона (GSH) GSH-редуктазой в клеточной линии PC12 после острое воздействие этанола.Food Chem.Toxicol. 2006; 44 (4): 574-578. Просмотреть аннотацию.
Уениши, К., Исида, Х., Тоба, Ю., Аое, С., Итабаши, А., и Такада, Ю. Основной белок молока увеличивает минеральную плотность костей и улучшает метаболизм костей у здоровых молодых женщин. Остеопорос. Int 2007; 18 (3): 385-390. Просмотреть аннотацию.
van Dissel, JT, de, Groot N., Hensgens, CM, Numan, S., Kuijper, EJ, Veldkamp, P., and van ‘t, Wout J. Сыворотка, обогащенная бычьими антителами, для предотвращения рецидив диареи, связанной с Clostridium difficile: доклинические и предварительные клинические данные.J Med Microbiol. 2005; 54 (часть 2): 197-205. Просмотреть аннотацию.
van Elswijk, DA, Diefenbach, O. , van der Berg, S., Irth, H., Tjaden, UR, and van der Greef, J. Быстрое обнаружение и идентификация ингибиторов ангиотензинпревращающего фермента с помощью жидкости в режиме онлайн хромато-биохимическое обнаружение в сочетании с масс-спектрометрией с электрораспылением. J Chromatogr.A 12-5-2003; 1020 (1): 45-58. Просмотреть аннотацию.
, van der Berg, S., Irth, H., Tjaden, UR, and van der Greef, J. Быстрое обнаружение и идентификация ингибиторов ангиотензинпревращающего фермента с помощью жидкости в режиме онлайн хромато-биохимическое обнаружение в сочетании с масс-спектрометрией с электрораспылением. J Chromatogr.A 12-5-2003; 1020 (1): 45-58. Просмотреть аннотацию.
ван Холл, Г., Сарис, В. Х., ван де Шур, П. А. и Вагенмакерс, А. Дж. Влияние приема свободного глутамина и пептидов на скорость ресинтеза мышечного гликогена у человека.Int.J.Sports Med. 2000; 21 (1): 25-30. Просмотреть аннотацию.
van, Hall G., Shirreffs, S. M., and Calbet, J. A. Ресинтез гликогена в мышцах во время восстановления после циклических упражнений: отсутствие эффекта от дополнительного приема белка. J. Appl Physiol 2000; 88 (5): 1631-1636. Просмотреть аннотацию.
Vandenplas, Y. Атопия в возрасте 3 лет у младенцев из группы высокого риска, получавших гидролизат сыворотки или обычную смесь. Ланцет 5-2-1992; 339 (8801): 1118..jpg) Просмотреть аннотацию.
Просмотреть аннотацию.
Vandenplas, Y., Deneyer, M., Sacre, L., and Loeb, H.Предварительные данные полевого исследования новой гипоаллергической формулы. Eur J Pediatr 1988; 148 (3): 274-277. Просмотреть аннотацию.
Vandenplas, Y., Hauser, B., Van den Borre, C., Clybouw, C., Mahler, T., Hachimi-Idrissi, S., Deraeve, L., Malfroot, A., and Dab, I. • Долгосрочное влияние формулы частичного гидролизата сыворотки на профилактику атопического заболевания. Eur J Pediatr 1995; 154 (6): 488-494. Просмотреть аннотацию.
Vandenplas, Y., Malfroot, A., and Dab, I. Краткосрочная профилактика аллергии на белок коровьего молока у младенцев.Иммунология и аллергологическая практика 1989; 11 (10): 430-437.
Veldhorst, MA, Nieuwenhuizen, AG, Hochstenbach-Waelen, A., van Vught, AJ, Westerterp, KR, Engelen, MP, Brummer, RJ, Deutz, NE, и Westerterp-Plantenga, MS Дозозависимый эффект насыщения сыворотка по сравнению с казеином или соей. Physiol Behav 3-23-2009; 96 (4-5): 675-682. Просмотреть аннотацию.
Просмотреть аннотацию.
Vermeirssen, V., Van Camp, J., Augustijns, P. и Verstraete, W. Пептиды, ингибирующие ангиотензин-I-превращающий фермент (ACE), полученные из горохового и сывороточного протеина.Meded.Rijksuniv.Gent Fak.Landbouwkd.Toegep.Biol.Wet. 2002; 67 (4): 27-30. Просмотреть аннотацию.
Vermeirssen, V., van der Bent, A., Van, Camp J., van, Amerongen A., and Verstraete, W. Количественный анализ in silico рассчитывает ингибирующую активность ангиотензин I превращающего фермента (АПФ) в горохе и переваривается сывороточный белок. Biochimie 2004; 86 (3): 231-239. Просмотреть аннотацию.
Vermeirssen, V., Van, Camp J., and Verstraete, W. Оптимизация и валидация анализа ингибирования ангиотензинпревращающего фермента для скрининга биоактивных пептидов.Журнал биохимии. Биофиз. Методы 3-4-2002; 51 (1): 75-87. Просмотреть аннотацию.
Вермейрссен, В., Ван, Кэмп Дж., Девос, Л. и Верстрете, В. Высвобождение ингибирующей активности ангиотензин-превращающего фермента (АПФ) во время желудочно-кишечного пищеварения in vitro: от серийного эксперимента к полунепрерывной модели. J. Agric.Food Chem. 9-10-2003; 51 (19): 5680-5687. Просмотреть аннотацию.
J. Agric.Food Chem. 9-10-2003; 51 (19): 5680-5687. Просмотреть аннотацию.
von Mutius, E. Воздействие на материнскую ферму / употребление непастеризованного коровьего молока и аллергические заболевания. Курр.Опин.Гастроэнтерол. 2012; 28 (6): 570-576.Просмотреть аннотацию.
фон, Берг А., Филипьяк-Питтрофф, Б., Крамер, У., Линк, Э., Боллрат, К., Броков, И., Колецко, С., Грубль, А., Генрих, Дж., Wichmann, HE, Bauer, CP, Reinhardt, D., and Berdel, D. Профилактический эффект гидролизованных смесей для младенцев сохраняется до 6 лет: долгосрочные результаты Немецкого исследования питания детей грудного возраста (GINI). J Allergy Clin Immunol. 2008; 121 (6): 1442-1447. Просмотреть аннотацию.
фон, Берг А., Колецко, С., Филипьяк-Питтрофф, Б., Лауберо, Б., Grubl, A., Wichmann, HE, Bauer, CP, Reinhardt, D., and Berdel, D. Некоторые гидролизованные смеси снижают частоту атопического дерматита, но не астмы: результаты трехлетнего исследования Немецкого исследования питания младенцев . J Allergy Clin Immunol. 2007; 119 (3): 718-725. Просмотреть аннотацию.
J Allergy Clin Immunol. 2007; 119 (3): 718-725. Просмотреть аннотацию.
von, Berg A., Koletzko, S., Grubl, A., Filipiak-Pittroff, B., Wichmann, HE, Bauer, CP, Reinhardt, D., and Berdel, D. Влияние гидролизованной смеси коровьего молока для профилактики аллергии на первом году жизни: Немецкое исследование питания младенцев, рандомизированное двойное слепое исследование.J Allergy Clin Immunol. 2003; 111 (3): 533-540. Просмотреть аннотацию.
Уокер, Т. Б., Смит, Дж., Эррера, М., Лебег, Б., Пинчак, А., и Фишер, Дж. Влияние 8-недельного приема сывороточного протеина и лейцина на физическую и когнитивную деятельность. Int J Sport Nutr Exerc.Metab 2010; 20 (5): 409-417. Просмотреть аннотацию.
Уолш, Д. Д., Бернард, Х., Мюррей, Б. А., Макдональд, Дж., Пентциен, А. К., Райт, Г. А., Уол, Дж. М., Струтерс, А. Д., Мейзел, Х. и Фитцджеральд, Р. Дж. Получение и стабильность in vitro фрагмента бета-лактоглобулина лактокинина (142-148).J Dairy Sci 2004; 87 (11): 3845-3857. Просмотреть аннотацию.
Ватанабэ, А., Окада, К., Симидзу, Ю., Вакабаяси, Х., Хигучи, К., Ниия, К., Кувабара, Ю., Ясуяма, Т., Ито, Х., Цукисиро, Т. ., Кондо, Ю., Эми, Н., и Кори, Х. Диетотерапия хронического гепатита с помощью сывороточного протеина (без подогрева). J.Med. 2000; 31 (5-6): 283-302. Просмотреть аннотацию.
Вайнерт, Д. Дж. Питание и синтез мышечного белка: описательный обзор. Журнал J Can Chiropr.Assoc 2009; 53 (3): 186-193. Просмотреть аннотацию.
Weinheimer, E.М., Конли, ТБ, Кобза, В.М., Сэндс, Л.П., Лим, Э., Джанл, Е.М., и Кэмпбелл, В.В. Добавка сывороточного протеина не влияет на вызванные тренировками изменения состава тела и индексы метаболического синдрома в среднем состоянии. пожилые люди с избыточным весом и ожирением. J Nutr 2012; 142 (8): 1532-1539. Просмотреть аннотацию.
Whitt, K. N., Ward, S. C., Deniz, K., Liu, L., Odin, J. A., and Qin, L. Холестатическое поражение печени, связанное с добавками сывороточного протеина и креатина. Семин. Дис. Печени. 2008; 28 (2): 226-231. Просмотреть аннотацию.
Просмотреть аннотацию.
Уайлд, К. Дж., Адди, К. В., Бодди, Л. М., и Пикер, М. Аутокринная регуляция секреции молока с помощью белка в молоке. Biochem.J 1-1-1995; 305 (Pt 1): 51-58. Просмотреть аннотацию.
Виллемс, Р., Дюшато, Дж., Магрез, П., Дени, Р. и Казимир, Г. Влияние гипоаллергенной молочной смеси на частоту ранних аллергических проявлений у младенцев, предрасположенных к атопическим заболеваниям. Энн Аллерджи 1993; 71 (2): 147-150. Просмотреть аннотацию.
Уиллоуби, Д. С., Стаут, Дж.Р. и Уилборн, С. Д. Влияние тренировок с отягощениями и добавок протеина и аминокислот на анаболизм, массу и силу мышц. Amino.Acids 2007; 32 (4): 467-477. Просмотреть аннотацию.
Уилмор, Д. В., Гудвин, К. В., Олик, Л. Х., Пованда, М. С., Мейсон, А. Д., младший, и Прюитт, Б. А., младший. Влияние травмы и инфекции на висцеральный метаболизм и кровообращение. Энн Сург 1980; 192 (4): 491-504. Просмотреть аннотацию.
Власюк, Г. и Верчелли, Д. Эффект фермы, или: когда, что и как сельскохозяйственная среда защищает от астмы и аллергических заболеваний. Curr.Opin.Allergy Clin.Immunol. 2012; 12 (5): 461-466. Просмотреть аннотацию.
Curr.Opin.Allergy Clin.Immunol. 2012; 12 (5): 461-466. Просмотреть аннотацию.
Ямамото Н., Маэно М. и Такано Т. Очистка и характеристика антигипертензивного пептида из йогуртоподобного продукта, ферментированного Lactobacillus helveticus CPN4. J Dairy Sci 1999; 82 (7): 1388-1393. Просмотреть аннотацию.
Янг, К. В., Манро, И. К., Тейлор, С. Л., Велдкамп, П., и ван Диссель, Дж. Т. Безопасность концентрата сывороточного белка, полученного из молока коров, иммунизированных против Clostridium difficile.Регул. Токсикол. Фармакол 2007; 47 (3): 317-326. Просмотреть аннотацию.
Земель, М. Б. Предполагаемая роль кальция и молочных пищевых компонентов в управлении весом и метаболическом здоровье. Физ. Спорт. 2009; 37 (2): 29-39. Просмотреть аннотацию.
Чжан, X. и Бейнен, А.С. Снижение эффекта диетического молочно-сывороточного протеина по сравнению с казеином на концентрацию холестерина в плазме и печени у крыс. Br J Nutr 1993; 70 (1): 139-146. Просмотреть аннотацию.
Чжао, X. Т., Миллер, Р. Х., МакКэмиш, М. А., Ван, Л., и Лин, Х.C. Всасывание белка зависит от подавления кишечного транзита в зависимости от нагрузки. Am J Clin Nutr 1996; 64 (3): 319-323. Просмотреть аннотацию.
Чжао, Ю., Мартин, Б. Р., Вастни, М. Е., Шоллум, Л., и Уивер, С. М. Острые и хронические эффекты сывороточных белков на абсорбцию кальция у растущих крыс. Exp.Biol Med (Maywood.) 2005; 230 (8): 536-542. Просмотреть аннотацию.
Zhu, K., Meng, X., Kerr, DA, Devine, A., Solah, V., Binns, CW, and Prince, RL Влияние двухлетнего рандомизированного контролируемого испытания добавок сывороточного протеина на структура костей, IGF-1 и экскреция кальция с мочой у пожилых женщин в постменопаузе.Дж. Костяной шахтер. Res 2011; 26 (9): 2298-2306. Просмотреть аннотацию.
Зоммара, М., Тубо, Х., Саконо, М., и Имаидзуми, К. Профилактика перекисного стресса у крыс, получавших диету с низким содержанием витамина Е, путем добавления препарата ферментированной сыворотки из коровьего молока: эффект молочной кислоты и бета-лактоглобулин на антипероксидантное действие. Biosci.Biotechnol.Biochem. 1998; 62 (4): 710-717. Просмотреть аннотацию.
Biosci.Biotechnol.Biochem. 1998; 62 (4): 710-717. Просмотреть аннотацию.
Zou, Z. Y., Lin, X. M., Xu, X. R., Xu, R., Ma, L., Li, Y. и Wang, M. F. Оценка добавок основного белка молока на плотность костной ткани и метаболизм костей у молодых китайских женщин.Eur.J Nutr 2009; 48 (5): 301-306. Просмотреть аннотацию.
Adebamowo CA, Spiegelman D, Berkey CS, et al. Потребление молока и прыщи у мальчиков-подростков. J Am Acad Dermatol. 2008 Май; 58 (5): 787-93. Просмотреть аннотацию.
Арчиеро П.Дж., Баур Д., Коннелли С. и др. Ежедневный прием сывороточного протеина и тренировки уменьшают массу висцеральной жировой ткани и повышают инсулинорезистентность: исследование PRIZE. J Appl Physiol (1985), 2014; 117 (1): 1-10. Просмотреть аннотацию.
Barak AJ, Beckenhauer HC, Tuma DJ.Бетаин, этанол и печень, обзор. Алкоголь 1996; 13: 395-8. Просмотреть аннотацию.
Baruchel S, Olivier R, Wainberg M. Анти-ВИЧ и антиапоптотическая активность концентрата сывороточного протеина: IMMUNOCAL. Int Conf AIDS 1994; 10: 32 (аннотация № 421A).
Int Conf AIDS 1994; 10: 32 (аннотация № 421A).
Белл К.Е., Снейдерс Т., Зулиняк М. и др. Многокомпонентная пищевая добавка на основе сывороточного протеина стимулирует рост мышечной массы и силы у здоровых пожилых мужчин: рандомизированное контролируемое исследование. PLoS One. 2017; 12 (7): e0181387.Просмотреть аннотацию.
Белл SJ. Концентраты сывороточного протеина с иммуноглобулинами и без них: обзор. J Med Food 2000; 3: 1-13.
Белобрайдик Д.П., МакИнтош Г.Х., Оуэнс Дж.А. Диета с высоким содержанием сывороточного белка снижает прибавку в весе и изменяет чувствительность к инсулину по сравнению с красным мясом у крыс линии Wister. J Nutr 2004; 134: 1454-8. Просмотреть аннотацию.
Bjørnshave A, Holst JJ, Hermansen K. Прием сывороточных белков перед приемом пищи вызывает различное влияние на метаболизм глюкозы и липидов у субъектов с метаболическим синдромом: рандомизированное перекрестное исследование.Eur J Nutr. 2019; 58 (2): 755-764. Просмотреть аннотацию.
Bounous G, Baruchel S, Falutz J, Gold P. Сывороточные белки в качестве пищевой добавки для ВИЧ-серопозитивных людей. Clin Invest Med 1993; 16: 204-9. Просмотреть аннотацию.
Bounous G, Batist G, Gold P. Сывороточные протеины в профилактике рака. Cancer Lett 1991; 7: 91-4. Просмотреть аннотацию.
Bumrungpert A, Pavadhgul P, Nunthanawanich P, Sirikanchanarod A, Adulbhan A. Добавка сывороточного протеина улучшает нутриционный статус, уровни глутатиона и иммунную функцию у больных раком: рандомизированное двойное слепое контролируемое исследование.J Med Food. 2018; 21 (6): 612-616. Просмотреть аннотацию.
Burton-Freeman, B.M. Гликомакропептид (GMP) не имеет решающего значения для насыщения, вызванного сывороткой, но может играть уникальную роль в регуляции потребления энергии посредством холецистокинина (CCK). Physiol Behav 2008; 93 (1-2): 379-387. Просмотреть аннотацию.
Candow DG, Burke NC, Smith-Palmer T, Burke DG. Эффект от приема добавок сывороточного и соевого протеина в сочетании с тренировками с отягощениями у молодых людей. Int J Sport Nutr Exerc Exerc Metab 2006; 16: 233-44. Просмотреть аннотацию.
Int J Sport Nutr Exerc Exerc Metab 2006; 16: 233-44. Просмотреть аннотацию.
Chanet A, Verlaan S, Salles J, et al. Дополнение завтрака лечебным питательным напитком из сывороточного протеина, обогащенного витамином D и лейцином, улучшает синтез мышечного протеина и мышечную массу после приема пищи у здоровых пожилых мужчин. J Nutr. 2017; 147 (12): 2262-2271. Просмотреть аннотацию.
Крибб П.Дж., Вильямс А.Д., Кэри М.Ф., Хейс А. Влияние изолята сыворотки и силовых тренировок на силу, состав тела и уровень глутамина в плазме. Int J Sport Nutr Exerc Exerc Metab 2006; 16: 494-509. Просмотреть аннотацию.
Крибб П.Дж., Уильямс А.Д., Статис К.Г. и др. Влияние изолята сыворотки, креатина и силовых тренировок на мышечную гипертрофию. Медико-научные спортивные упражнения 2007; 39: 298-307. Просмотреть аннотацию.
Даллас С., Стемпак Д., Корен Г. и др. Концентрат сывороточного протеина (WPC): модуляция уровня глутатиона в лимфоцитах in vitro. Clin Pharmacol Ther 2000; 67: 156 (аннотация PIII-56).
Дэвис Р.В., Карсон Б.П., Джейкман П.М. Влияние добавок сывороточного протеина на временное восстановление мышечной функции после тренировки с отягощениями: систематический обзор и метаанализ.Питательные вещества. 2018; 10 (2). Просмотреть аннотацию.
Энгельсон Э.С. и др. Влияние высокобелковой диеты на белковый обмен у ВИЧ-инфицированных мужчин и женщин. Int Conf AIDS 1998; 12: 553 (аннотация № 32166).
Энглунд Д.А., Кирн Д.Р., Кочек А. и др. Пищевые добавки с физической активностью улучшают состав мышц у пожилых людей с ограниченной подвижностью, исследование VIVE2: рандомизированное двойное слепое плацебо-контролируемое исследование. J Gerontol A Biol Sci Med Sci. 2017; 73 (1): 95-101. Просмотреть аннотацию.
Файер Р., Гидри А, Блэгберн Бл. Иммунотерапевтическая эффективность колостральных иммуноглобулинов крупного рогатого скота от гипериммунизированной коровы против криптоспоридиоза у новорожденных мышей. Инфекция иммунной 1990; 58: 2962-5. Просмотреть аннотацию.
FDA. Частично гидролизованная детская смесь из 100% сывороточного протеина снижает риск атопического дерматита. Май 2011 г. Доступно по адресу: https://wayback.archive-it.org/7993/20171114183703/https://www.fda.gov/Food/IngredientsPackagingLabeling/LabelingNutrition/ucm256731.htm
Fekete ÁA, Giromini C, Chatzidiakou Y, Гивенс Д.И., Лавгроув Д.А.Сывороточный протеин снижает артериальное давление и улучшает функцию эндотелия и липидные биомаркеры у взрослых с предгипертонией и легкой гипертензией: результаты хронического рандомизированного контролируемого исследования Whey2Go. Am J Clin Nutr. 2016; 104 (6): 1534-1544. Просмотреть аннотацию.
Фриланд-Грейвс JH, Lin PH. Поглощение марганца плазмой в зависимости от пероральных доз марганца, кальция, молока, фосфора, меди и цинка. J Am Coll Nutr 1991; 10: 38-43. Просмотреть аннотацию.
Fukushima Y, Kawata Y, Onda T, Kitagawa M.Длительное употребление смеси с гидролизатом сыворотки кормящими женщинами снижает перенос бета-лактоглобулина в грудное молоко. Nutr Sci Vitaminol (Токио) 1997; 43: 673-8. Просмотреть аннотацию.
Nutr Sci Vitaminol (Токио) 1997; 43: 673-8. Просмотреть аннотацию.
Гаффни К.А., Лусеро А., Стоунер Л. и др. Влияние нулевого сывороточного протеина на гликемический контроль после интенсивных тренировок в смешанном режиме при диабете 2 типа. Медико-спортивные упражнения. 2018; 50 (1): 11-17. Просмотреть аннотацию.
Giezenaar C, Hutchison AT, Luscombe-Marsh ND, Chapman I, Horowitz M, Soenen S. Влияние возраста на уровень глюкозы в крови и инсулин плазмы, глюкагон, грелин, CCK, GIP и GLP-1 в ответ на проглатывание сывороточного протеина.Питательные вещества. 2017; 10 (1). Просмотреть аннотацию.
Giezenaar C, Trahair LG, Luscombe-Marsh ND, et al. Влияние рандомизированных нагрузок сывороточного протеина на потребление энергии, аппетит, опорожнение желудка и концентрацию гормонов кишечника в плазме у мужчин и женщин пожилого возраста. Am J Clin Nutr. 2017; 106 (3): 865-877. Просмотреть аннотацию.
Hakkak R, Korourian S, Shelnutt SR, et al. Рационы, содержащие сывороточный протеин или изолят соевого протеина, защищают от индуцированных 7,12-диметилбенз (а) антраценом опухолей молочной железы у самок крыс. Эпидемиологические биомаркеры рака до 2000 г .; 9: 113-7. Просмотреть аннотацию.
Эпидемиологические биомаркеры рака до 2000 г .; 9: 113-7. Просмотреть аннотацию.
Хансен М., Бангсбо Дж., Дженсен Дж. И др. Влияние гидролизата сывороточного протеина на работоспособность и восстановление высококлассных бегунов по спортивному ориентированию. Int J Sport Nutr Exerc Exerc Metab 2015; 25 (2): 97-109. Просмотреть аннотацию.
Хван П.С., Андре Т.Л., МакКинли-Барнард С.К. и др. Повышение мышечной силы у тренированных мужчин, вызванное тренировкой с отягощениями, сохраняется после 2 недель отсутствия тренировок и не зависит от приема добавок сывороточного протеина.J Strength Cond Res. 2017; 31 (4): 869-881. Просмотреть аннотацию.
Якубович Д., Фрой О., Арен Б. и др. Инкретин, инсулинотропный и глюкозоснижающий эффекты предварительной нагрузки сывороточного протеина при диабете 2 типа: рандомизированное клиническое исследование. Диабетология 2014; 57 (9): 1807-11. Просмотреть аннотацию.
Кеннеди Р.С., Конок Г.П., Баунус Г. и др. Использование концентрата сывороточного протеина в лечении пациентов с метастатической карциномой: клиническое исследование фазы I-II. Anticancer Res 1995; 15: 2643-9. Просмотреть аннотацию.
Anticancer Res 1995; 15: 2643-9. Просмотреть аннотацию.
Кеог, Дж. Б. и Клифтон, П. Влияние заменителей пищи с высоким содержанием гликомакропептида на потерю веса и маркеры риска сердечно-сосудистых заболеваний. Am J Clin Nutr 2008; 87 (6): 1602-1605. Просмотреть аннотацию.
Кеог, Дж. Б., Вунтон, Б. В., Тейлор, К. М., Джанакиевски, Ф., Десилва, К., и Клифтон, П. М. Влияние фракций гликомакропептидов на холецистокинин и потребление пищи. Br J Nutr 2010; 104 (2): 286-290. Просмотреть аннотацию.
Kerstetter JE, Bihuniak JD, Brindisi J, et al.Влияние добавки сывороточного протеина на костную массу у пожилых людей европеоидной расы. J Clin Endocrinol Metab 2015; 100 (6): 2214-22. Просмотреть аннотацию.
Kim JM, Белый RH. Влияние витамина Е на антикоагулянтный ответ на варфарин. Am J Cardiol 1996; 77: 545-6. Просмотреть аннотацию.
Кита М., Обара К., Кондо С., Умеда С., Ано Й. Влияние добавки сывороточного пептида, богатого триптофан-тирозин-связанными пептидами, на когнитивные функции у здоровых взрослых: рандомизированное, двойное слепое, плацебо-контролируемое исследование . Питательные вещества. 2018 13 июля; 10 (7). Просмотреть аннотацию.
Питательные вещества. 2018 13 июля; 10 (7). Просмотреть аннотацию.
Лам С. М., Моуган П. Дж., Авати А. и Мортон Х. Р. Влияние сывороточного белка и гликомакропептида на чувство насыщения у взрослых людей. Physiol Behav 2009; 96 (1): 162-168. Просмотреть аннотацию.
Лаопрасерт Н., Валлен Н.Д., Джонс Р.Т. и др. Анафилаксия у ребенка с аллергией на молоко после приема лимонного шербета, содержащего следовые количества молока. J Food Prot 1998; 61: 1522-4. Просмотреть аннотацию.
Lopes Gomes D, Moehlecke M, Lopes da Silva FB, et al.Добавка сывороточного протеина способствует увеличению жировых отложений и снижению веса у женщин после бариатрической операции: рандомизированное контролируемое исследование. Obes Surg. 2017; 27 (2): 424-431. Просмотреть аннотацию.
Маккензи-Шалдерс К.Л., Бирн Н.М., Слейтер Г.Дж. и др. Влияние дозы сывороточного протеина на сытость и потребление пищи у спортсменов, тренирующихся с отягощениями. Аппетит 2015; 92: 178-84. Просмотреть аннотацию.
МакАдам Дж. С., Макгиннис К. Д., Бек Д. Т. и др. Влияние добавок сывороточного протеина на физическую работоспособность и состав тела солдат начальной подготовки армии.Питательные вещества. 2018; 10 (9). Просмотреть аннотацию.
МакИнтош GH. Рак толстой кишки: диетические изменения, необходимые для сбалансированной защитной диеты. Предыдущая Мед 1993; 22: 767-74. Просмотреть аннотацию.
Messina M, Lynch H, Dickinson JM, Reed KE. Нет разницы между влиянием добавок соевого протеина и животного протеина на прирост мышечной массы и силы в ответ на упражнения с отягощениями. Int J Sport Nutr Exerc Exerc Metab. 2018; 28 (6): 674-685. Просмотреть аннотацию.
Nabuco HCG, Tomeleri CM, Fernandes RR, et al.Влияние добавок сывороточного протеина до или после тренировки на окислительный стресс и антиоксидантные ферменты у пожилых женщин. Scand J Med Sci Sports. 2019; 29 (8): 1101-1108. Просмотреть аннотацию.
Набуко ХГЧ, Томелери С.М., Сугихара Джуниор П. и др. Влияние добавок сывороточного протеина до или после тренировки с отягощениями на мышечную массу, мышечную силу и функциональную способность у пожилых женщин в прекондиционировании: рандомизированное клиническое испытание. Питательные вещества. 2018 3 мая; 10 (5). Просмотреть аннотацию.
Влияние добавок сывороточного протеина до или после тренировки с отягощениями на мышечную массу, мышечную силу и функциональную способность у пожилых женщин в прекондиционировании: рандомизированное клиническое испытание. Питательные вещества. 2018 3 мая; 10 (5). Просмотреть аннотацию.
Нельсон Л., Рао А., Олсон П.Уникальные гидролизованные изоляты сывороточного протеина с антигипертензивной активностью. Институт Food Tech 2000 Ann Mtg & Food Expo: аннотация 38-6. Доступно по адресу: ift.confex.com/ift/2000/techprogram/paper_5129.htm
Niikawa M, Hayashi H, Sato T, et al. Выделение веществ из глянцевой бирючины (Ligustrum lucidum Ait.), Ингибирующих мутагенность бензо [а] пирена у бактерий. Mutat Res 1993; 319: 1-9. Просмотреть аннотацию.
Папенбург Р., Боунус Г., Флейзер Д., Голд П. Белки диетического молока подавляют развитие злокачественных новообразований, вызванных диметилгидразином.Tumor Biol 1990; 11: 129-36. Просмотреть аннотацию.
Pontes TC, Costa Fernandes Filho GM, Pereira Trindade AS, Sobral Filho JF. Заболеваемость вульгарными угрями у молодых взрослых, употребляющих белково-калорийные добавки в городе Хоатильде-Пессоа-ПБ. Бюстгальтеры Dermatol. 2013; 88 (6): 907-12. Просмотреть аннотацию.
Заболеваемость вульгарными угрями у молодых взрослых, употребляющих белково-калорийные добавки в городе Хоатильде-Пессоа-ПБ. Бюстгальтеры Dermatol. 2013; 88 (6): 907-12. Просмотреть аннотацию.
Power O, Hallihan A, Jakeman P. Инсулинотропный ответ человека на пероральное проникновение нативного и гидролизованного протреина сыворотки. Аминокислоты 2009; 37: 333-9. Просмотреть аннотацию.
Rencuzogullari I, Börekçi A, Karakoyun S, et al.Коронарный тромбоз трех коронарных артерий из-за сывороточного протеина. Am J Emerg Med. 2017; 35 (4): 664.e3-664.e4. Просмотреть аннотацию.
Ригамонти А.Е., Леончини Р., Касничи С. и др. Сывороточные протеины снижают аппетит, стимулируют анорексигенные желудочно-кишечные пептиды и улучшают глюкометаболический гомеостаз у молодых полных женщин. Питательные вещества. 2019; 11 (2). pii: E247. Просмотреть аннотацию.
Саломон С.Б., Юнг Дж., Восс Т. и др. Элементарный рацион, содержащий триглицериды со средней длиной цепи и ферментативно гидролизованный белок, может улучшить желудочно-кишечную толерантность у людей, инфицированных ВИЧ. J Am Diet Assoc 1998; 98: 460-2. Просмотреть аннотацию.
J Am Diet Assoc 1998; 98: 460-2. Просмотреть аннотацию.
Schroer AB, Saunders MJ, Baur DA и др. Прием сывороточного протеина и L-аланина во время длительных тренировок может ухудшить результаты велотренировок на время. Int J Sport Nutr Exerc Exerc Metab 2014; 24 (5): 507-15. Просмотреть аннотацию.
Семла Т.П., Бейзер Дж.Л., Хигби Мэриленд. Справочник по гериатрической дозировке. 4-е изд. Хадсон, Огайо: Lexicomp, 1998.
Шарп М.Х., Лоури Р.П., Шилдс К.А. и др. Влияние говядины, курицы или сывороточного протеина после тренировки на состав тела и работоспособность мышц.J Strength Cond Res. 2018; 32 (8): 2233-2242. Просмотреть аннотацию.
Сирато М., Цучия Ю., Сато Т. и др. Влияние комбинированного приема бета-гидрокси-бета-метилбутирата (HMB) и сывороточного протеина на симптомы повреждения мышц, вызванного эксцентрическими упражнениями. J Int Soc Sports Nutr. 2016; 13: 7. Просмотреть аннотацию.
Сильверберг NB. Сывороточный протеин вызывает умеренные и тяжелые обострения угрей у 5 спортсменов-подростков. Отчеты о случаях заболевания Cutis. 2012; 90 (2): 70-2. Просмотреть аннотацию.
Отчеты о случаях заболевания Cutis. 2012; 90 (2): 70-2. Просмотреть аннотацию.
Симонарт Т. Прием прыщей и сывороточного протеина среди бодибилдеров.Дерматология. 2012; 225 (3): 256-8. Просмотреть аннотацию.
Sinnott RA, Maddela RL, Nelson ED, et al. Модифицирующие эффекты добавки сывороточного протеина, богатой кальцием (порошок OsoLean), на потерю веса и окружность талии у субъектов с избыточным весом: предварительное исследование. OsoLean Open Nutraceuticals J 2009.
Smith GI, Commean PK, Reeds DN, Klein S, Mittendorfer B. Влияние протеиновых добавок во время похудания, вызванного диетой, на мышечную массу и силу: рандомизированное контролируемое исследование.Ожирение (Серебряная весна). 2018 Май; 26 (5): 854-861. Просмотреть аннотацию.
Srivastava Y, Venkatakrishna-Bhatt H, Verma Y, et al. Антидиабетические и адаптогенные свойства экстракта Momordica charantia: экспериментальная и клиническая оценка. Phytother Res 1993; 7: 285-9.
Сугихара Джуниор П., Рибейро А.С., Набуко ХГЧ и др. Влияние добавок сывороточного протеина, связанных с тренировками с отягощениями, на мышечную силу, гипертрофию и качество мышц у предварительно подготовленных пожилых женщин.Int J Sport Nutr Exerc Exerc Metab. 2018; 28 (5): 528-535. Просмотреть аннотацию.
Тахаворгар А., Вафа М., Шидфар Ф. и др. Предварительная загрузка сывороточного белка более полезна, чем предварительная загрузка соевого белка в регулировании аппетита, потребления калорий, антропометрии и состава тела мужчин с избыточным весом и ожирением. Nutr Res 2014; 34 (10): 856-61. Просмотреть аннотацию.
Тан Дж. Э., Манолакос Дж. Дж., Куйбида Г. В. и др. Минимальное содержание углеводов в сыворотке стимулирует синтез мышечного белка после упражнений с отягощениями у тренированных молодых мужчин.Appl Physiol Nutr Metab 2007; 32: 1132-8. Просмотреть аннотацию.
Тан Дж. Э., Мур Д. Р., Куйбида Г. В. и др. Прием гидролизата сыворотки, казеина или изолята соевого белка: влияние на синтез смешанного мышечного белка в состоянии покоя и после упражнений с отягощениями у молодых мужчин. J Appl Physiol 2009; 107: 987-92. Просмотреть аннотацию.
Taylor LW, Wilborn C, Roberts MD, White A, Dugan K. Восемь недель приема добавок сывороточного протеина до и после тренировки увеличивают мышечную массу и улучшают результаты у баскетболисток университетского дивизиона III.Appl Physiol Nutr Metab. 2016; 41 (3): 249-54. Просмотреть аннотацию.
Тосуховонг П., Бунла С., Диссаябутра Т. и др. Биохимические и клинические эффекты добавок сывороточного протеина при болезни Паркинсона: пилотное исследование. J Neurol Sci. 2016; 367: 162-70. Просмотреть аннотацию.
Vandenplas Y, Hauser B, Van den Borre C, et al. Влияние гидролизата сыворотки на профилактику атопического заболевания. Энн Аллерджи 1992; 68: 419-24. Просмотреть аннотацию.
Veldhorst, M. A., Nieuwenhuizen, A. G., Hochstenbach-Waelen, A., Westerterp, K. R., Engelen, M. P., Brummer, R.J., Deutz, N. E., and Westerterp-Plantenga, M. S. Влияние полноценного сывороточного завтрака по сравнению с сывороточным завтраком без GMP-завтраков на потребление энергии и насыщение. Аппетит 2009; 52 (2): 388-395. Просмотреть аннотацию.
Вергель Н.Р., Сальвато П., Муни М. Анаболические стероиды, упражнения с отягощениями и протеиновые добавки влияют на безжировую массу тела у ВИЧ-положительных пациентов. Int Conf AIDS 1998; 12: 557 (аннотация № 32185).
Verreijen AM, Verlaan S, Engberink MF и др.Добавка с высоким содержанием сывороточного протеина, лейцина и витамина D сохраняет мышечную массу во время преднамеренной потери веса у пожилых людей с ожирением: двойное слепое рандомизированное контролируемое исследование. Am J Clin Nutr 2015; 101 (2): 279-86. Просмотреть аннотацию.
Вилелла А.Л., Лимсуват С., Уильямс Д.Р., Зайферт С.Ф. Холестатическая желтуха в результате приема комбинированных пищевых добавок. Энн Фармакотер 2013; 47 (7-8): e33. Просмотреть аннотацию.
Volek JS, Volk BM, Gómez AL, et al. Добавление сывороточного протеина во время силовых тренировок увеличивает безжировую массу тела.J Am Coll Nutr 2013; 32 (2): 122-35. Просмотреть аннотацию.
Voss T, Rowe B, Graf L, et al. Управление потерей веса, связанной с ВИЧ, и диареей с помощью энтеральной смеси, содержащей пептиды сыворотки и триглицериды со средней длиной цепи. Int Conf AIDS 1991; 7: 223 (аннотация № WB2165).
Витард О.К., Джекман С.Р., Брин Л. и др. Скорость синтеза миофибриллярного мышечного протеина после еды в ответ на увеличение доз сывороточного протеина в покое и после упражнений с отягощениями. Am J Clin Nutr 2014; 99 (1): 86-95.Просмотреть аннотацию.
Вонг К.В., Лю А.Х., Регестер ГО и др. Влияние сывороточного и очищенного сывороточных белков на функции нейтрофилов у овец. J Dairy Res 1997; 64: 281-8. Просмотреть аннотацию.
Вонг CW, Ватсон DL. Иммуномодулирующие эффекты пищевых белков сыворотки у мышей. J Dairy Res 1995; 62: 359-68. Просмотреть аннотацию.
Wright CS, McMorrow AM, Weinheimer-Haus EM, Campbell WW. Добавки сывороточного протеина и более высокое общее потребление протеина не влияют на количество костей у взрослых с избыточным весом и ожирением после 36-недельных упражнений и диетического вмешательства.J Nutr. 2017; 147 (2): 179-186. Просмотреть аннотацию.
Чжу К., Керр Д.А., Мэн Х и др. Двухлетний прием сывороточного протеина не увеличивал мышечную массу и физические функции у здоровых пожилых женщин в постменопаузе с хорошим питанием. J Nutr 2015; 145 (11): 2520-6. Просмотреть аннотацию.
Сколько мне следует принимать сывороточного протеина? Простое руководство для достижения # целей
Если вы используете сывороточный протеин, вы, вероятно, задаетесь вопросом «сколько сывороточного протеина мне следует принимать?» Сумма зависит от ваших конкретных целей.В целом, если вашей целью является похудание, принимайте 10-20 граммов для умеренно активного образа жизни и 20-40 граммов, если вы очень активны. Если ваша цель — набрать сухую мышечную массу, употребляйте 20-48 граммов сывороточного протеина после тренировки.
Сывороточный протеин предназначен не только для бодибилдеров и профессиональных спортсменов. Это для всех, кто хочет похудеть, улучшить мышечную силу, уменьшить тягу к еде и получить больше белка в своем рационе.
Из этой статьи вы узнаете:
Дозировка сывороточного протеина для похудания
Хотите сжигать лишние 80-100 калорий в день и обуздать аппетит? Не ищите ничего, кроме сывороточного протеина! Он не только ускоряет метаболизм, но и снижает тягу к еде, что снижает тягу к перекусам и перееданию, что приводит к потере жира.Хорошая новость в том, что вам не нужно потреблять большое количество сывороточного протеина, чтобы воспользоваться преимуществами этой добавки для сжигания жира.
Рекомендуемая суточная доза белка составляет минимум 0,4-0,6 грамма белка на фунт вашего веса. Обратите внимание, что это минимальная сумма , необходимая для удовлетворения ваших основных потребностей в питании и предотвращения дефицита. Чтобы действительно получить пользу от похудения, лучше всего получать около 30% ежедневных калорий из белков.
Если вам интересно, одна мерная ложка нашего органического сывороточного протеина содержит 20 граммов чистого протеина.Имея это в виду, вот наша общая рекомендация о том, сколько сывороточного протеина нужно принимать в день для похудания:
- УМЕРЕННАЯ АКТИВНОСТЬ (3 тренировки или меньше в неделю): 10-20 граммов протеина (или 1 / 2-1 мерная ложка органического сывороточного протеина) помогут обуздать тягу к еде и повысить ваш метаболизм.
- ВЫСОКАЯ АКТИВНОСТЬ (4 и более тренировок в неделю): 20-40 граммов протеина (или 1-2 мерные ложки органического сывороточного протеина) подавят тягу к голоду и будут способствовать здоровому обмену веществ.
Поднимите свой метаболизм с помощью нашего восхитительно сытного шоколадного смузи с авокадо на завтрак!
В Магазин!
Дозировка сывороточного протеина для наращивания мышечной массы
Сывороточный протеин содержит все 9 незаменимых аминокислот, которые играют ключевую роль в способности вашего тела наращивать и восстанавливать мышцы. Кроме того, организм усваивает сыворотку быстрее, чем любой другой белок, что приводит к усилению синтеза мышечного белка (также известного как те мышцы, которые вам нужны).
Исследования предлагают сочетать HIIT или силовые тренировки с добавлением сывороточного протеина для достижения ваших целей. А для достижения максимальных результатов обязательно ознакомьтесь с нашими рекомендациями, когда лучше всего принимать сывороточный протеин.
Вот наша рекомендация относительно того, сколько сывороточного протеина нужно принимать для набора мышечной массы:
- СРЕДНЯЯ АКТИВНОСТЬ (3 тренировки или меньше в неделю): 20-25 граммов сыворотки (или 1 мерная ложка без горки на 1 большую мерную ложку органического сывороточного протеина) после тренировки должно дать вам протеин, необходимый для наращивания мышечной массы. .
- ВЫСОКАЯ АКТИВНОСТЬ (4 и более тренировок в неделю): Если вы спортсмен, вам может потребоваться до 0,9 грамма белка на фунт веса тела. Это много белка, поэтому, чтобы помочь вам достичь желаемых показателей белка и максимизировать набор и восстановление мышц, мы рекомендуем принимать 24-48 граммов сыворотки (или 1-2 ложки с горкой органического сывороточного протеина) после тренировки.
Чтобы быстро и легко получить протеиновый коктейль после тренировки, попробуйте наш Ванильный протеиновый коктейль с низким содержанием углеводов! На его приготовление уходит всего 30 секунд, поэтому вы сможете легко получить дозу белка в это решающее время после тренировки.
Заключение
Сывороточный протеин — это мощная добавка, которая может помочь вам достичь поставленных целей по сжиганию жира и увеличению мышечной массы (почти как личный тренер в тренажерном зале!). Хорошая новость в том, что его употреблять очень безопасно. Когда дело доходит до добавок для достижения ваших целей, не бойтесь экспериментировать с приведенными выше рекомендациями, пока не найдете свою золотую середину!
Сколько сывороточного протеина вы принимаете для достижения целей? Мы будем рады узнать в комментариях ниже!
Up Next: лучшее время для приема сывороточного протеина (если вы хотите максимальных результатов!)
(Хотите подобные статьи по электронной почте? Вот подписка!)
Когда, что и сколько?
Протеиновый коктейль после тренировки с отягощениями — это популярная стратегия питания, которую используют многие любители фитнеса и спортсмены для увеличения синтеза мышечного протеина (MPS).
Но подтверждают ли данные эту практику, и если да, то какой тип протеина лучше всего, сколько следует употреблять и когда нужно употреблять?
Сколько белка?
Согласно курсу NASM Nutrition, текущая рекомендуемая суточная норма потребления белка для здоровых взрослых составляет 0,8 г на килограмм (0,36 г на фунт) веса тела в день (1). Это составляет примерно 64 грамма белка в день для человека весом 176 фунтов (80 кг) (176 x 0.36). Однако этот RDA белка считается недостаточным для удовлетворения всех потребностей тренирующихся с отягощениями или даже людей, тренирующихся на выносливость (2,3). Впоследствии большинство авторитетных организаций, занимающихся вопросами здоровья, фитнеса и питания, пришли к единому мнению о том, что спортсменам действительно требуется большее количество белка в своем рационе, чем их коллегам, ведущим сидячий образ жизни, для поддержания MPS.
Распространенное заблуждение людей, занимающихся физическими упражнениями, заключается в том, что потребление большего количества белка поддерживает более высокий уровень MPS, но последние заявления о позиции Академии питания и диетологии (AND), диетологов Канады (DC), Американского колледжа спортивной медицины (ACSM) Национальная ассоциация силы и кондиционирования (NSCA) и Международное общество спортивного питания (ISSN) оспаривают это мнение.В каждом из них указаны рекомендуемые дозировки для людей, тренирующихся на выносливость и сопротивление, а также верхние пороги потребления белка (1,4,5). Идея установления верхнего порога основана на исследованиях, которые не смогли продемонстрировать повышенный уровень MPS при употреблении большего количества белка (то есть превышении дневного общего или рекомендуемого количества за один прием пищи, потому что он (а) не всасывается полностью или (б) может не использоваться для MPS телом). Хотя эти заявления о позиции несколько различаются между агентствами, ниже представлен консенсус в качестве руководства для практикующих специалистов:
- 2 к 1.4 грамма на килограмм веса тела (0,55 — 0,64 грамма / фунт) для тренированных на выносливость людей (определяется как более 10 часов в неделю тренировок на выносливость)
- 4–2,0 грамма на килограмм веса тела (0,64–0,91 грамма / фунт) для силовых тренировок *.
Например, атлет весом 154 фунта (70 кг) на выносливость потребляет от 84 до 98 г белка (154 фунта x 0,55–0,64), тогда как атлет весом 220 фунтов (100 кг) потребляет от 140 до 98 г белка. 200 граммов белка (220 фунтов.х 0,64 и 0,91).
* Традиционная дозировка, которую часто используют люди, составляет 1 грамм на фунт массы тела или 2,2 грамма на килограмм, что примерно на 10% превышает рекомендованный верхний порог.
Тем не менее, важно также признать, что существуют исследования, в которых изучали более высокое потребление белка у опытных бодибилдеров и людей с отягощениями, которые придерживались диеты с высоким содержанием белка (до 2,8 грамма на килограмм или 1,27 грамма на фунт). Эти люди продолжали демонстрировать МПС без каких-либо проблем со здоровьем, таких как нарушение функции почек (6).
Когда употреблять белок?
Другое обсуждение касается количества белка, которое следует потреблять за один прием пищи или один присест. Скорость всасывания протеина сильно различается между людьми и между источниками протеина. Хотя у мужчин, как правило, желудочно-кишечный тракт (ЖКТ) больше, поэтому они могут поглощать больше белка, чем женщины, в действительности очень трудно точно определить это количество, учитывая множество факторов, влияющих на переваривание и скорость всасывания белка, в том числе (7) :
- Эффективность усвояемости белков в пищевых продуктах (различается для растительных и животных источников).
- Размер тела и генетика.
- Размер и состав пищи (присутствие клетчатки и жиров может препятствовать всасыванию белка).
- Источники белка (например, сыворотка, казеин, яйца). Водорастворимая сыворотка всасывается быстрее, чем казеин, нерастворимый молочный белок, который образует творог в желудке, задерживая его опорожнение желудка и всасывание на несколько часов. С другой стороны, яйцо содержит от шести до семи граммов белка, но организм поглощает примерно 3 грамма вареного яйца каждый час.
- Диетический опыт (люди, потребляющие больше белка, могут адаптироваться к более эффективному перевариванию / поглощению белка).
В исследовании, проведенном Саймонсом и его коллегами (8), сравнивали 30 граммов протеина и 90 граммов протеина в группах молодого и пожилого возраста. Их исследование показало, что прием более 30 граммов за один прием пищи не усиливал стимуляцию МПС в обеих исследуемых популяциях. Это значение перекликается с мнением других исследователей, которые считают, что от 20 до 30 граммов представляют собой идеальное количество белка, которое может быть эффективно усвоено за один присест (9-11).Таким образом, дозы примерно 20 граммов для женщин и 25-30 граммов для мужчин за один присест кажутся идеальными, что впоследствии требует многократного кормления в течение дня для достижения желаемой суммы.
Несмотря на это, то, что, кажется, вызывает большой интерес в исследованиях MPS, так это количество потребляемого лейцина (12,13). Метаболизм аминокислот в скелетных мышцах обычно ограничивается шестью аминокислотами (глутамат, аспартат, аспарагин, BCAA — лейцин, изолейцин и валин), но наиболее значимые эффекты связаны с лейцином, который действует как критический регулятор MPS (например,g., путь m-Tor), дополняя сигнальные эффекты инсулина и производство других аминокислот в развитии мышц (подумайте о лейцине как о двигателе или переключателе для управления MPS). Текущий консенсус, по-видимому, согласен с тем, что на одно кормление требуется минимальная доза от 2 до 2,5 граммов лейцина, чтобы сигнализировать путям mTOR о наличии достаточного пищевого белка для поддержки MPS (14).
Время приема белка имеет решающее значение для улучшения MPS и восстановления мышц. Хотя было продемонстрировано, что потребление белка до и после тренировки приводит к увеличению частоты MPS по сравнению с утренним и вечерним кормлением (15), различия между до и после тренировки остаются менее очевидными.Типтон и его коллеги кормили активных людей аминокислотно-углеводным напитком, содержащим 6 граммов незаменимых аминокислот (EAA) * и 35 граммов высокогликемических углеводов (EAA-CHO) за 60 минут до высокоинтенсивной тренировки с отягощениями (80% 1 ПМ) и в течение 30-45 минут после этой же тренировки. Исследование показало, что чистый MPS, связанный с потреблением белка перед тренировкой, был больше, чем белок, потребляемый после тренировки (16). Однако этот результат не был воспроизведен в исследовании Schoenfeld и его коллег, которые не обнаружили различий между приемом 25 граммов белка до и после тренировки (17).Хотя это исследование также пришло к выводу, что оптимальное анаболическое окно после тренировки для максимизации MPS также не такое маленькое, как считалось ранее (т.е. 2-3 часа после тренировки), а, скорее, длится от 5 до 6 часов, другие исследования не подтверждают это же открытие, и рекомендуют потребление белка в течение первого часа после тренировки (18).
* 6 грамм EAA эквивалентны 17,5 граммам изолята гороха или 12 граммам изолята сыворотки (без учета всасываемости белка, когда изолят гороха составляет примерно 75% эффективности изолята сыворотки).
Применимый вывод для практикующих может заключаться в том, что доза до 20-25 граммов протеина потребляется в течение часа до тренировки, а затем снова потребляется в течение первого часа после тренировки. Согласно консенсусу исследований потребления белка в период после тренировки, доза составляет 20 граммов, но это количество можно более точно рассчитать, используя диапазон дозировки от 0,25 до 0,3 грамма на килограмм (от 0,11 до 0,14 грамма / фунт) (11 , 19). Например, человек весом 176 фунтов (80 кг) будет стремиться к потреблению от 20 до 24 граммов белка.
Какой белок?
В предыдущей статье были рассмотрены ключевые различия между качеством и усвояемостью растительных белков и белков животного происхождения, а также между белками в каждой пищевой категории. Самым важным результатом потребления белка до или сразу после тренировки является быстрая доставка к мышечным клеткам — «быстрые» белки доставляют аминокислоты в мышцы более эффективно. В то время как казеин может опуститься из желудка в течение нескольких часов, изоляты сыворотки могут попасть в кровь в течение 15-20 минут.Впоследствии людям будет лучше всего потреблять быстрый белок, такой как изолят сывороточного протеина, до и / или после тренировки. Но поскольку белок также следует употреблять через несколько часов (например, через 3-4 часа), альтернативная стратегия после тренировки состоит в том, чтобы для удобства потреблять смесь быстрых и медленных белков, учитывая, что некоторые люди могут не иметь склонность к способности снова есть через несколько часов после тренировки. Будет ли включение медленного белка в быстрый белок (т.е.е., смешанный белок) препятствует немедленному MPS, в значительной степени неизвестно. Тем не менее, потребление белка в течение дня в идеале должно соответствовать режиму частого приема меньших доз белка для поддержания более положительного баланса азота (то есть для сохранения мышечной массы, а не ее разрушения). Предпочтительно, это влечет за собой практику приема качественного белка каждые несколько часов (например, 3-4 часа) и дополняется приемом «медленного» белка, такого как казеин, перед сном, чтобы помочь снизить катаболическое состояние, которое организм испытывает во время ночного голодания.
В заключение, хотя потребление протеинового коктейля после тренировки остается жизнеспособной и эффективной практикой, практикующие врачи могут разработать и реализовать несколько научно обоснованных стратегий для оптимизации результатов, связанных с практикой потребления дополнительного белка.
Footnote: Считается, что потребление аминокислот с разветвленной цепью (BCAAS) во время упражнений снижает мышечную болезненность после упражнений и ускоряет восстановление, хотя это исследование не лишено противоречий.Учитывая эти противоречия и объем данной статьи, потребление BCAA во время упражнений обсуждаться не будет.
Список литературы
- Американская диетическая ассоциация (ADA), диетологи Канады (DC) и Американский колледж спортивной медицины (ACSM), Питание и спортивные результаты (2009). Журнал Американской диетической ассоциации , 109 (3): 509 — 527.
- Тарнопольский М.А., Аткинсон С.А., Макдугалл Д.Д., Челси А., Филлипс С. и Шварц Х.П. (1992).Оценка потребности в белке тренированных силовых атлетов. Журнал прикладной физиологии , 73 (5): 1986-1995.
- Тарнопольский М.А., Гибала М., Джеукендруп А.Е. и Филлипс С.М. (2005). Потребности в питании высококвалифицированных спортсменов на выносливость. Часть I: Углеводы и потребности в жидкости. Европейский журнал спортивной науки , 5: 314. 14.
- Kreider RB, Wilborn CD, Taylor L, Campbell B, Almada AL, Collins R, et al. (2010). Международное общество спортивного питания, упражнения и обзор спортивного питания: исследования и рекомендации. Журнал Международного общества спортивного питания , 7: 7.
- Кэмпбелл Б., Крайдер Р. Б., Зейгенфус Т., Ла Баунти П., Робертс М., Берк Д., и др. (2007). Позиция Международного общества спортивного питания: белок и упражнения. Журнал Международного общества спортивного питания , 4: 8.
- Poortmans JR, Dellalieux O (2000). Имеет ли регулярная диета с высоким содержанием белка потенциальный риск для здоровья почек у спортсменов? Международный журнал спортивного питания и метаболизма упражнений , 10 (1): 28-38.
- Антонио Дж. И Стаут Дж. Р. (ред.) (2001). Спортивное питание . Филадельфия, Пенсильвания. Липпинкотт, Уильямс и Уилкинс.
- Саймонс ТБ, Шеффилд-Мур М., Вулф Р.Р. и Паддон-Джонс Д. (2009). Умеренная порция высококачественного белка максимально стимулирует синтез белка в скелетных мышцах у молодых и пожилых людей. Журнал Американской диетической ассоциации , 109 (9): 1582-1586.
- Филлипс С.М. (2006). Диетический белок для спортсменов: от требований к метаболическому преимуществу. Прикладная физиология, питание и обмен веществ , 31: 647-54.
- Типтон К.Д. и Вулф Р.Р. (2004). Белки и аминокислоты для спортсменов. Journal of Sports Science , 22: 65-79.
- Мур Д. Р., Робинсон М. Дж., Фрай Дж. Л., Тан Дж. Э., Гловер Е. И., Уилкинсон С. Б., et al. (2009). Потребленный белок отвечает за синтез белка в мышцах и альбумина после упражнений с отягощениями у молодых мужчин. Американский журнал клинического питания, 89 (1): 161-168.
- Ренни MJ и Типтон KD (2000).Обмен белков и аминокислот во время и после тренировки и влияние питания. Ежегодный обзор питания, 20: 457-483.
- Мирянин ДК (2002). Роль лейцина в метаболизме белков во время упражнений и восстановления. Канадский журнал прикладной физиологии, 27: 646-662.
- Крозье С. Дж., Кимбалл С. Р., Эммерт С. В., Энтони Дж. С. и Джефферсон Л. С. (2005). Пероральное введение лейцина стимулирует синтез белка в скелетных мышцах крыс. Журнал питания , 135: 376-382.
- Крибб П.Дж. и Хейс А. (2006). Влияние времени приема добавок и упражнений с отягощениями на гипертрофию скелетных мышц. Медицина и наука в спорте и физических упражнениях , 38: 1918-1925.
- Типтон К.Д., Расмуссен Б.Б., Миллер С.Л., Вольф С.Е., Оуэнс-Стовалл С.К., Петрини Б.Е. и Вольф Р.Р. (2001). Выбор времени приема углеводов и аминокислот изменяет анаболический ответ мышц на упражнения с отягощениями. Американский журнал физиологии, эндокринологии и метаболизма , 281: E197-E206.
- Шенфельд Б.Дж., Арагон А.А., Уилборн С., Урбина С.Л., Хейворд С.Е. и Кригер Дж. (2017). Потребление белка до и после тренировки оказывает аналогичное влияние на мышечную адаптацию. PeerJ, 5: e2825 https://doi.org/10.7717/peerj.2825.
- Эсмарк Б., Андерсен Дж. Л., Олсен С., Рихтер Е. А., Мизуно М. и Кьяер М. (2001). Время потребления белка после тренировки важно для гипертрофии мышц при тренировках с отягощениями у пожилых людей. Журнал физиологии , 535: 301-311.
http: // blog.nasm.org/nutrition/latest-scoop-protein-supplementation/
Изучение сложности компенсации дозировки на уровне белка, которая точно настраивает стехиометрию мультибелковых комплексов
Abstract
Правильный контроль уровней экспрессии генов при различных возмущениях является фундаментальным аспектом устойчивости клетки. Компенсация дозировки на уровне белка — это один из механизмов буферизации нарушений стехиометрии мультибелковых комплексов за счет ускоренного протеолиза несобранных субъединиц.Хотя N-терминальное ацетилирование и убиквитин-опосредованная деградация протеасом с помощью пути правила Ac / N-конца делает возможным селективную компенсацию избыточных субъединиц, неясно, насколько широко распространенный этот путь вносит вклад в контроль стехиометрии. Здесь мы сообщаем, что дозовая компенсация лишь частично зависит от пути правила Ac / N-конца. Наш анализ генетических взаимодействий между 18 субъединицами и 12 факторами контроля качества у почкующихся дрожжей показал, что множественные E3 ubiquitin ligases и N-acetyltransferases участвуют в дозовой компенсации.Мы обнаружили, что компенсация, опосредованная N-ацетилтрансферазами, не просто предсказуема по N-концевой последовательности, несмотря на их специфичность последовательности для N-ацетилирования. Мы также обнаружили, что компенсация Pop3 и Bet4 в значительной степени обусловлена минорной N-ацетилтрансферазой NatD. Более того, канонические субстраты NatD гистон h3A / h5 были компенсированы даже в его отсутствие, что указывает на независимый от N-ацетилирования стехиометрический контроль. Наше исследование показывает сложность и надежность системы контроля стехиометрии.
Сведения об авторе
Контроль качества мультибелковых комплексов важен для поддержания гомеостаза в клеточных системах, основанных на функциональных комплексах. Правильная стехиометрия мультибелковых комплексов достигается за счет баланса между синтезом и распадом белка. Недавние исследования показали, что эффективность трансляции имеет тенденцию масштабироваться со стехиометрией их субъединиц. С другой стороны, хотя путь протеолиза, опосредованный N-концевым ацетилированием белка и убиквитином, вовлечен в селективную деградацию избыточных субъединиц, неясно, насколько широко этот путь способствует контролю стехиометрии из-за отсутствия систематических исследований с использованием эндогенных белков.Чтобы лучше понять ландшафт системы контроля стехиометрии, мы исследовали генетические взаимодействия между 18 субъединицами и 12 факторами контроля качества (убиквитинлигазы E3 и N-ацетилтрансферазы), всего 114 комбинаций. Наши данные предполагают, что N-ацетилтрансферазы частично ответственны за контроль стехиометрии и что независимый от N-ацетилирования путь также участвует в селективной деградации избыточных субъединиц. Таким образом, данное исследование показывает сложность и надежность системы контроля стехиометрии.Дальнейшее рассмотрение этой сложности поможет понять механизмы буферизации нарушений экспрессии генов и формирования стехиометрии протеома.
Образец цитирования: Исикава К., Исихара А., Мория Х. (2020) Изучение сложности компенсации дозировки на уровне белка, которая точно настраивает стехиометрию мультипротеиновых комплексов. PLoS Genet 16 (10):
e1009091.
https://doi.org/10.1371/journal.pgen.1009091
Редактор: Марко Йованович, Колумбийский университет, США
Поступило: 18 июля 2019 г .; Принята к печати: 31 августа 2020 г .; Опубликовано: 28 октября 2020 г.
Авторские права: © 2020 Ishikawa et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все соответствующие данные находятся в рукописи и ее файлах с вспомогательной информацией.
Финансирование: Эта работа финансировалась грантом для научного сотрудника JSPS (16J00852) (K.I.) и грантом JSPS для научных исследований (B) (262
) (H.М.). Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили, что конкурирующих интересов не существует.
Введение
Контроль внутриклеточной концентрации белка на должном уровне является критическим аспектом клеточных систем. Например, было высказано предположение, что клетки регулируют концентрацию протеома для постоянной скорости биохимических реакций [1–3].Более того, поскольку широкий спектр путей реакции на стресс динамически изменяет экспрессию белков, ответственных за выживание в различных сложных условиях, очевидно, что контроль концентрации белка важен для того, чтобы справляться с нарушениями и поддерживать клеточный гомеостаз.
Полногеномные исследования, измеряющие устойчивость клеток к генетическим нарушениям, показали, что клетки Saccharomyces cerevisiae являются хрупкими по отношению к сверхэкспрессии белка в подмножестве генома [4, 5].Это открытие указывает на то, что клеточная система обычно устойчива к генетическим нарушениям различных биологических процессов. Однако неясно, как клетки буферизируют такие изменения окружающей среды [6]. Недавно мы сообщили, что при увеличении числа копий гена примерно 10% генома дрожжей транскрибируется линейно в уровни мРНК, но не транслируется напрямую в уровни белка [7]. Комплексный анализ анеуплоидных клеток дрожжей и млекопитающих также показал, что примерно 20% генома обладают этим молекулярным фенотипом [8, 9].Следовательно, это явление, известное как дозовая компенсация на уровне белка, может частично объяснить буферизацию нарушений экспрессии генов.
Дозовая компенсация на уровне белка преимущественно нацелена на гены, кодирующие субъединицы мультибелковых комплексов [7–10]. Концентрация белков с компенсированной дозой изменяется двунаправленно в ответ на концентрацию его субъединиц-партнеров [7, 11]. Эти наблюдения согласуются с гипотезой, предсказывающей вредные эффекты из-за стехиометрического дисбаланса сложных субъединиц на рост клеток [5, 6, 12].Другое доказательство этого предсказания происходит из анализа профилей рибосом, раскрывающего стратегию пропорционального синтеза, которая позволяет масштабировать эффективность трансляции со стехиометрией субъединиц [13, 14]. Это открытие показывает, что стехиометрический баланс субъединиц мультибелковых комплексов в основном поддерживается на трансляционном уровне. Кроме того, эта стратегия сохраняется от бактерий к высшим эукариотам [14]. В этом контексте дозовая компенсация может быть признана безотказным механизмом, который точно регулирует стехиометрию протеома [15].
Механизм дозовой компенсации — ускоренная деградация несобранных субъединиц убиквитин-протеасомной системой [7, 11, 16, 17]. Множественные E3 ubiquitin ligases, участвующие в контроле стехиометрии, были идентифицированы ранее [11, 18, 19], но нам не хватает понимания их относительного вклада в компенсацию идентичных субъединиц [20]. Более того, основной механизм избирательной деградации избыточных субъединиц не совсем понятен. Хотя предыдущие исследования обнаружили доказательства избирательной компенсации Cog1 и Hcn1 у дрожжей и PLIN2 в клетках млекопитающих с помощью пути правила Ac / N-конца [11, 21], неясно, насколько широко распространенный этот путь вносит вклад в компенсацию.Путь правила Ac / N-конца является одним из механизмов деградации белка за счет ацетилирования N-концевой аминокислоты N-ацетилтрансферазами (NAT). На ограниченном количестве примеров было показано, что N-ацетилирование действует как сигнал деградации (N-degron), распознаваемый специфическими E3-убиквитинлигазами (N-распознаванием) [11, 18, 22]. Была выдвинута гипотеза, что N-степень несобранных субъединиц экспонируется и распознается N-распознаванием, тогда как собранная субъединица экранируется другими субъединицами в том же комплексе и недоступна для N-распознавания (рис. 1А) [11, 18 , 23].Эта модель согласуется с биологической ролью дозовой компенсации, но еще предстоит исследовать, зависит ли компенсация от этого пути. Более того, влияние пути правила Ac / N-конца на контроль стехиометрии различается в возмущенных и невозмущенных условиях [11, 18, 24], что побудило нас оценить, насколько широко распространенная NATs-обеспечиваемая компенсация происходит при генетических пертурбациях.
Рис. 1. Анализ генетических возмущений системы контроля стехиометрии.
( A ) Компенсация дозы на уровне белка преимущественно нацелена на субъединицы мультибелковых комплексов.Путь правила Ac / N-конца участвует в основном механизме избирательной компенсации несобранных субъединиц. ( B ) Экспериментальная установка для измерения вклада каждого фактора контроля качества в компенсацию дозировки. Немеченый белок с компенсированной дозой экспрессировали из многокопийной плазмиды pTOW40836, содержащей нативные регуляторные последовательности, включая промотор и 5´ и 3´ нетранслируемые области. Если целевой ген подвергается дозовой компенсации, уровень TAP-меченного целевого белка, экспрессируемого только из хромосомы, снижается при увеличении числа копий гена (Multi) по сравнению с векторным контролем (Single).Отношение Multi / Single выше, чем в клетках WT в делеционном мутанте гена, кодирующего убиквитинлигазу E3 или каталитическую субъединицу каждой NAT, если удаленный фактор способствует компенсации целевого гена.
https://doi.org/10.1371/journal.pgen.1009091.g001
В этом исследовании мы измерили вклад убиквитин-лигаз E3 и NAT в компенсацию дозировки субъединиц РНКазы P / MRP, которые представляют собой идентифицированные компенсированные белки. в нашем предыдущем исследовании [7], систематическим образом.Мы обнаружили, что в компенсации участвуют множественные E3-убиквитинлигазы и NAT. Мы также обнаружили, что зависимость компенсации от NAT варьируется среди тестируемых субъединиц. Наши результаты показывают, что дозовая компенсация не полностью зависит от пути правила Ac / N-конца, и подчеркивают сложность системы контроля стехиометрии.
Результаты
Множественные убиквитинлигазы E3 Tom1 и Not4 участвуют в дозовой компенсации идентичных субъединиц
В нашем предыдущем исследовании было проверено 54 гена на хромосоме I S . cerevisiae , чтобы объективно оценить, какая часть генома кодирует компенсированные белки, и мы идентифицировали пять белков (Rbg1, Mtw1, Pop5, Saw1 и Erp2) [7]. Чтобы исследовать, какие E3-убиквитин-лигазы участвуют в дозовой компенсации и их относительный вклад, мы исследовали, компенсируются ли эти белки в меньшей степени у пяти делеционных мутантов генов E3-убиквитин-лигазы ( DOA10 , NOT4 , TOM1 , SAN1 , и UBR1 ), участвующих в системе контроля качества белка, включая пути правила N-конца: (i) Doa10 и Not4 известны как N-распознаватели в пути правила Ac / N-конца [11, 18], (ii) Tom1 участвует в деградации широкого диапазона несобранных субъединиц рибосомы [19], (iii) San1 отвечает за протеасомную деградацию аберрантных ядерных белков [25], и (iv) Ubr1 является N-распознаванием в Arg / N-конце. путь правила, который, как предполагается, дополняет путь правила Ac / N-конца [26, 27].Используя ранее разработанный метод идентификации генов с дозовой компенсацией (S1A Fig.) [7], мы обнаружили, что количество Pop5 в tom1 Δ и not4 Δ и Saw1 в doa10 Δ было увеличено по сравнению с генами дикого типа. (WT) клетки (S1B и S1C на фиг.). Недавний систематический анализ стабильности белка в клетках doa10 Δ показал, что гидрофобность N-конца отрицательно коррелирует со стабильностью белка [24]. В соответствии с этим открытием, N-конец Saw1 более гидрофобен, чем другие белки, которые не стабилизируются в doa10 Δ клетках.Однако следует отметить, что Pop5 с гидрофобным N-концом не стабилизировался в отсутствие Doa10.
Pop5 — субъединица комплексов РНКаза P / MRP, состоящая из одиннадцати субъединиц [28]. Эти комплексы разделяют восемь субъединиц: Pop1, Pop3, Pop4, Pop5, Pop6, Pop7, Pop8 и Rpp1, в то время как Rpr2 включен только в РНКазу P, а Snm1 и Rmp1 включены только в MRP РНКазы. Ранее мы обнаружили, что эффективность трансляции мРНК POP5 не изменяется при увеличении числа копий POP5 с помощью профилирования рибосом [7].Кроме того, эффективность трансляции других субъединиц комплексов РНКазы P / MRP, включая Rpp1 и Pop8, которые образуют субкомплекс с Pop5 [29], также не затрагивается в тех же условиях. Следовательно, контроль стехиометрии этих комплексов зависит от деградации белка при генетических нарушениях, и можно более точно оценить его вклад в контроль стехиометрии, анализируя субъединицы РНКазы P / MRP.
Поскольку большинство субъединиц этих комплексов подвергаются дозовой компенсации [7], мы исследовали, участвуют ли Tom1 и Not4 в компенсации других субъединиц.Мы использовали экспериментальную установку, с помощью которой TAP-меченый целевой белок, экспрессируемый только из одного геномного локуса, выявляется при увеличении числа копий немаркированной версии того же гена (рис. 1B и S2A – S2D рис.). Сначала мы сравнили уровни белка между с избыточной экспрессией и без нее в клетках WT, tom1 Δ или not4 Δ (S2E фиг.). Этот анализ показал значительное снижение компенсации Pop4, Pop5, Pop6, Pop7, Pop8, Rpp1, Rpr2 и Rmp1 в tom1 Δ клетках (рис. 2A и 2B), а также Pop3, Pop4, Pop5, Pop7, Rpp1 и Rmp1 в , а не в 4 Δ ячейках (рис. 2D и 2E).Следовательно, Tom1 и Not4 участвуют в дозовой компенсации субъединиц РНКазы P / MRP: Tom1 и Not4 нацелены на по крайней мере 8 и 6 из 11 субъединиц, соответственно (Таблица 1).
Рис. 2. Tom1 и Not4 участвуют в компенсации дозировки.
( A ) Вестерн-блоттинг субъединиц РНКазы P / MRP в tom1 Δ клетках. Субъединица с меткой TAP, экспрессируемая в хромосоме, была обнаружена с помощью PAP. Pop1 и Snm1 в наших экспериментах не обнаружены. Показан репрезентативный блот из трех биологических повторов.( B ) Количественная оценка уровней белка в условиях Multi по сравнению с уровнями в условиях Single в клетках WT и tom1 Δ. Среднее кратное изменение ± стандартное отклонение. был рассчитан из трех биологических повторов. Статистическая значимость была определена с помощью двустороннего теста Велча t (* p <0,05, ** p <0,01). Пунктирная линия представляет один и тот же уровень выражения между условиями Single и Multi. ( C ) Сравнение уровней белка между клетками WT и tom1 Δ в одиночных или множественных условиях.Среднее кратное изменение ± стандартное отклонение. был рассчитан из трех биологических повторов. Пунктирная линия представляет одинаковый уровень экспрессии между клетками WT и tom1 Δ. Уровни Pop6 и Rpr2 были незначительно значимо увеличены в tom1 Δ в условиях Single ( p = 0,059 и 0,071, соответственно). ( D – F ) То же, что и в ( A – C ), за исключением того, что показаны вестерн-блоты субъединиц РНКазы P / MRP в not4 Δ клетках ( D ) и их количественное определение ( E, F ).Уровни Pop5, Pop6 и Rpr2 были значительно увеличены в not4 Δ по сравнению с клетками WT в условиях Single ( p <0,05).
https://doi.org/10.1371/journal.pgen.1009091.g002
Мы дополнительно исследовали, участвует ли Doa10 также в контроле стехиометрии субъединиц РНКазы P / MRP, и обнаружили, что все девять тестируемых субъединиц были компенсированы в doa10 Δ в той же степени в клетках WT (S1 и S3 фиг.). Следовательно, Doa10 не участвует в контроле стехиометрии субъединиц РНКазы P / MRP, что согласуется с предыдущим исследованием, показывающим, что Cog1 компенсируется Not4 вместо Doa10 [11].Эти наблюдения предполагают различные предпочтения субстратов для E3 ubiquitin ligases, ответственных за компенсацию. В самом деле, Tom1 имеет тенденцию компенсировать локализованные в ядре белки, как описано ниже (see Discussion).
Затем мы сравнили уровни белка между WT и tom1 Δ или not4 Δ клетками со сверхэкспрессией каждой субъединицы комплексов РНКаза P / MRP или без нее (S2F фиг.). Этот анализ показал более высокий уровень Pop6 и Rpr2 в tom1 Δ и Pop5, Pop6 и Rpr2 в not4 Δ, чем в клетках WT без увеличения числа копий гена (рис. 2C и 2F, одиночные).Поскольку это сравнение проводится между векторными контролями в WT и мутантных клетках, Tom1 и Not4 могут участвовать в деградации этих субъединиц в невозмущенных условиях. Кроме того, поскольку Pop6 не подвергался Not4-опосредованной компенсации, Not4 способствует его деградации, но это не ускоряется при сверхэкспрессии.
Наш анализ показал, что Not4 вносит вклад в компенсацию Rmp1, чья полилизиновая последовательность CKKKKKRKKKNK, близкая к C-концу (остатки 182–193), индуцирует арест трансляции, связанный с Not4-опосредованным протеолизом [30].Недавние исследования показали, что пептиды, останавливающие трансляцию, разрушаются комплексом контроля качества рибосом (RQC) [31]. RQC-опосредованная деградация арестованных пептидов и инициация этого пути опосредуются убиквитинлигазами E3 Ltn1 и Hel2 соответственно [32, 33]. Чтобы проверить, связан ли этот путь клиренса для арестованных пептидов с дозовой компенсацией, мы измерили степень компенсации Rmp1 у делеционных мутантов LTN1 или HEL2 , используя ту же экспериментальную установку, показанную на рис. 1В.Мы обнаружили, что степень компенсации Rmp1 в ltn1 Δ и hel2 Δ была совместима с таковой в клетках WT (S4 Рис.). Наши результаты предполагают, что нарушения пути контроля качества для клиренса арестованных пептидов не приводят к снижению уровня компенсации Rmp1.
Компенсация дозировки Pop6 и Pop7 происходит в ответ на их внутриклеточную концентрацию
Поскольку и Pop6, и Pop7, которые формируют гетеродимерный субкомплекс Pop6-Pop7 [34, 35], подвергаются дозовой компенсации (Рис. 2), количество Pop6 и Pop7 может изменяться в ответ на количество партнерских субъединиц.Мы предположили, что высокая дозировка POP7 приводит к изменениям доли стабильных / собранных и нестабильных / несобранных пулов Pop6 (рис. 3A, вверху). Более того, поскольку механизм дозовой компенсации — это ускоренная деградация несобранных субъединиц, баланс этих пулов субъединиц может быть нарушен в отсутствие Tom1 или Not4 (Рис. 3B, вверху). Как и ожидалось (рис. 3A и 3B, внизу), мы обнаружили, что Pop6 стабилизировался увеличением дозировки POP7 , и наоборот, в клетках WT, tom1 Δ или not4 Δ (рис. 3C и 3D) .Эти результаты показывают, что дозовая компенсация Pop6 и Pop7 происходит двунаправленно в ответ на изменения внутриклеточной концентрации каждой субъединицы: ускоренная деградация Pop6 / Pop7 при высокой дозировке POP6 / POP7 (нисходящая компенсация) и сниженная деградация Pop6 / Pop7 при высокой дозировке POP7 / POP6 (компенсация вверх).
Рис. 3. Двунаправленная дозовая компенсация гетеродимерного субкомплекса Pop6-Pop7.
( A ) Модель для контроля стехиометрии Pop6 и Pop7 в клетках WT (вверху) и экспериментальная установка для тестирования этой модели (внизу).Pop6 становится более стабильным, когда находится в комплексе с партнерской субъединицей Pop7, тогда как несобранный Pop6 менее стабилен (вверху слева). После многокопийной экспрессии Pop7 потенциально деградированный Pop6 становится стабильным за счет образования комплекса с избытком Pop7 (вверху справа). Уровень Pop6 в векторном управлении — это уровень устойчивого состояния (внизу слева). Если оборот Pop6 снижается за счет высокой дозировки POP7 , уровень Pop6 увеличивается по сравнению с таковым при векторном контроле (внизу справа). ( B ) То же, что в ( A ), за исключением того, что показано для ячеек tom1 Δ или not4 Δ.Потенциально деградированный Pop6 в клетках WT становится стабильным в отсутствие Tom1 или Not4, которые ответственны за деградацию Pop6 и Pop7 (вверху слева). При многокопийной экспрессии Pop7 эта не деградировавшая фракция и оставшийся нестабильный пул Pop6 становятся стабильными, образуя комплекс с избытком Pop7 (вверху справа). Устойчивый уровень Pop6 в клетках tom1, Δ или not4, Δ выше, чем в клетках WT (внизу слева). Если оборот Pop6 дополнительно снижается за счет высокой дозировки POP7 , это приводит к небольшому увеличению уровня Pop6 (внизу справа).( C, D ) Вестерн-блоты Pop6-TAP ( C ) и Pop7-TAP ( D ) в клетках WT, tom1 Δ и not4 Δ клеток. Белки, меченные TAP, детектировали с помощью PAP в условиях Single (+ Vector: pTOW40836) и Multi (+ POP6 : pTOW40836- POP6 , + POP7 : pTOW40836- POP7 ). Показана количественная оценка интенсивности полос относительно одного условия в каждом штамме. Окрашивание кумасси белка массой 50 кДа, соответствующего енолазе, показано как контроль нагрузки.
https://doi.org/10.1371/journal.pgen.1009091.g003
При увеличении числа копий гена субъединицы-партнера относительное соотношение Pop6 и Pop7 в tom1 Δ и not4 Δ было ниже, чем это в клетках WT. Это различие может отражать регуляцию степени компенсации, поскольку количество Pop6 и Pop7 было увеличено у обоих мутантов без изменения числа копий гена (фиг. 3C и 3D, + вектор). Следовательно, Tom1 и Not4 влияют на баланс пулов стабильных / нестабильных субъединиц гетеродимерного субкомплекса Pop6-Pop7.
NAT вносят комплексный вклад в дозовую компенсацию
S . cerevisiae имеет пять NAT: NatA, NatB, NatC, NatD и NatE, каталитическими субъединицами которых являются соответственно Naa10, Naa20, Naa30, Naa40 и Naa50 [36]. NAT имеют различную специфичность последовательностей для N-ацетилирования, и их субстраты могут быть классифицированы по первым двум N-концевым остаткам, за исключением NatD, который распознает мотивы последовательностей с приблизительно пятью остатками [18, 36, 37]. Как показано на фиг. 4A, на основании предсказания по N-концевым аминокислотам субъединицы РНКазы P / MRP могут быть N-ацетилированы с помощью NatA, NatB или NatC.Мы исследовали, являются ли субъединицы РНКазы P / MRP менее компенсированными в клетках naa10 Δ, naa20 Δ, naa30 Δ, naa40 Δ или naa50 Δ клетках, используя ту же экспериментальную установку, описанную на рис. Наши эксперименты показали, что Pop4 и Rmp1 в naa10 Δ (рис. 4B и 4C) и Pop6, Pop8, Rpr2 и Rmp1 в naa20 Δ (рис. 4D и 4E) были значительно менее компенсированы, чем в клетках WT, в то время как все тестируемые субъединицы были компенсированы в naa30, Δ или naa50, Δ, а также в клетках WT (рис. 4F, 4G, 4J и 4K).Только Pop3 не был компенсирован в naa40 Δ клетках (фиг. 4H и 4I), а период полужизни Pop3 был продлен при увеличении числа копий POP3 в naa40 Δ клетках (S5A и S5B фиг.). Следует отметить, что Pop3 был стабилизирован в naa10 Δ по сравнению с клетками WT с увеличением количества копий гена или без него (S6 Fig) ( p <0,05, двусторонний тест Велча t ), что предполагает участие NatA. в Pop3 деградации даже в невозмущенных условиях. Мы также обнаружили, что компенсация Pop5 была более выраженной в naa10 Δ, чем в клетках WT (рис. 4B и 4C), предполагая, что ускоренная деградация Pop5 во время дозовой компенсации блокируется NatA.
Рис. 4. NAT вносят сложный вклад в компенсацию дозировки.
( A ) Первые пять аминокислот субъединиц РНКазы P / MRP (слева). Основываясь на этих последовательностях, NatA, NatB или NatC ответственны за их N-ацетилирование (справа). NatA-опосредованное ацетилирование требует удаления N-концевого метионина с помощью Met-аминопептидаз Map1 и Map2 (MetAPs). ( B ) Вестерн-блоттинг субъединиц РНКазы P / MRP в naa10 Δ клетках. Субъединица с меткой TAP, экспрессируемая в хромосоме, была обнаружена с помощью PAP.Pop1 и Snm1 в наших экспериментах не обнаружены. Показан репрезентативный блот из трех биологических повторов. ( C ) Количественная оценка уровней белка в условиях Multi по сравнению с уровнями в условиях Single в клетках WT и naa10 Δ. Среднее кратное изменение ± стандартное отклонение. был рассчитан из трех биологических повторов. Статистическая значимость была определена с помощью двустороннего теста Велча t (* p <0,05, ** p <0,01). Пунктирная линия представляет один и тот же уровень выражения между условиями Single и Multi.( D – K ) То же, что и в ( B, C ), за исключением того, что показаны вестерн-блоты субъединиц РНКазы P / MRP и их количественное определение в naa20 Δ ( D, E ), naa30 Δ ( F, G ), naa40 Δ ( H, I ) или naa50 Δ ( J, K ) ячеек.
https://doi.org/10.1371/journal.pgen.1009091.g004
Чтобы оценить функциональную избыточность NAT во время компенсации доз, мы сгенерировали naa10 Δ naa20 Δ клеток, несущих TAP-tag POP4 или RMP1 в геноме.Однако у этого двойного мутанта обнаружен синтетический дефект роста (S7A фиг.), И мы не наблюдали образования колоний этого двойного мутанта при трансформации многокопийными плазмидами, несущими POP4 или RMP1 . Таким образом, мы измерили количество Pop4 и Rmp1 без увеличения числа копий гена и обнаружили, что оно не отличается от количества Pop4 и Rmp1, которое не отличается от такового у WT и одиночных мутантов (S7B фиг.).
Мы суммировали вклад каждого NAT в компенсацию субъединиц РНКазы P / MRP, что привело к нескольким находкам (Таблица 1).Во-первых, NatB-опосредованная компенсация Rmp1 — единственный случай, который можно ожидать от его N-концевой последовательности. Во-вторых, компенсация Pop4 и Pop6 была задействована с помощью NatA и NatB, соответственно, хотя компенсация Pop4 и Pop6 посредством NatB и NatC, соответственно, ожидалась из их специфичности последовательности для N-ацетилирования. В-третьих, Rmp1 компенсировался через несколько NAT (NatA и NatB). В-четвертых, компенсация Pop3 была в значительной степени за счет NatD, но не NatA. Наконец, NAT для компенсации Pop7 и Rpp1 не были идентифицированы.Взятые вместе, N-концевая последовательность не может просто объяснить вклад NAT в компенсацию. Более того, NATs-опосредованная компенсация субъединиц РНКазы P / MRP в основном опосредована NatA и NatB, хотя их влияние на уровни белка было ограниченным (Рис. 5). Таким образом, эти результаты демонстрируют сложность системы контроля стехиометрии, которая лишь частично зависит от опосредованной NAT компенсации дозировки.
Рис. 5. Относительный вклад Tom1, Not4 и NAT в дозовую компенсацию субъединиц РНКазы P / MRP.
Тепловая карта показывает количественную сводку вклада Tom1, Not4 и NAT в компенсацию дозировки субъединиц РНКазы P / MRP. Базальный уровень — это среднее кратное изменение в клетках WT. Вклад каждого фактора рассчитывали как среднее кратное изменение для каждого мутанта за вычетом базального уровня.
https://doi.org/10.1371/journal.pgen.1009091.g005
Кроме того, белки, запускающие MD, Pop4 и Rmp1 компенсировались с помощью NatA и Not4, тогда как NatB также участвовал в компенсации Rmp1 (рис. 2 и 4).Cog1 также является стартовым белком MD, который компенсируется NatB и Not4 при его сверхэкспрессии [11]. Эти наблюдения предполагают, что даже если первые два N-концевых остатка белков-субстратов одинаковы, компенсация посредством пути правила Ac / N-конца происходит с помощью различных комбинаций NAT и лигаз E3.
Канонические субстраты NatD, гистон h3A / h5, компенсируются в отсутствие NatD
Как показано на рис. 4H и 4I, NatD эффективно способствует компенсации Pop3, который не идентифицируется как субстрат NatD [38].Для дальнейшего изучения того, трудно ли предсказать дозовую компенсацию, опосредованную NAT, исходя из N-концевой последовательности, мы затем проанализировали канонические субстраты NatD. NatD более селективен, чем другие NAT [36], и только субъединицы гистона h3A / h5 идентифицированы как его субстраты [38-40]. Эти гистоновые субъединицы являются исходными белками MS, которые могут быть субстратами NatA, но они N-ацетилированы в отсутствие Naa10 или его вспомогательной субъединицы Naa15 [41]. Таким образом, мы сначала проверили, участвует ли NatD в компенсации субъединиц гистона h3A / h5: Hta2, Hhf1 и Hhf2.Мы обнаружили, что они подвергались дозовой компенсации даже в отсутствие Naa40 (рис. 6A – 6C), что позволяет предположить, что NatD участвует в их N-ацетилировании, но не в деградации. Кроме того, компенсация наблюдалась не только в naa40 Δ, но также в naa10 Δ, naa20 Δ или naa30 Δ, а также в клетках WT (S8 фиг.). Мы также обнаружили, что все они были значительно меньше компенсированы в клетках tom1 Δ, хотя степень компенсации у них была разной (Рис. 6A – 6C).Эти результаты предполагают, что Tom1-опосредованная компенсация Hta2, Hhf1 и Hhf2 не требует активности NAT и что N-ацетилирование и дозовая компенсация не обязательно связаны. Следовательно, дозовая компенсация гистона h3A / h5 может быть опосредована независимым от N-ацетилирования путем.
Рис. 6. Рассмотрение сложности компенсации дозы, опосредованной NAT, путем сосредоточения внимания на NatD.
( A – C ) Tom1-опосредованная компенсация дозировки субстратов NatD не требует Naa40.Вестерн-блоттинг гистона h3A / h5: Hta2-TAP ( A ), Hhf1-TAP ( B ) и Hhf2-TAP ( C ) в клетках WT, naa40 Δ или tom1 Δ. . Субъединицы с меткой TAP были обнаружены с помощью PAP. Среднее кратное изменение ± стандартное отклонение. был рассчитан из трех биологических повторов. ( D ) Вестерн-блоттинг Bet2-TAP в naa40 Δ клетках с использованием PAP. Среднее кратное изменение ± стандартное отклонение. был рассчитан из четырех биологических повторов. ( E ) Вестерн-блоттинг Bet4-TAP в naa10 Δ, naa20 Δ, naa30 Δ, naa40 Δ, naa50 Δ и naa20 Δ naa40 Δ клетках с использованием PAP .Среднее кратное изменение ± стандартное отклонение. рассчитывали из трех биологических повторов, за исключением анализа клеток naa40 и Δ в восьми биологических повторах. Статистическую значимость определяли с помощью двустороннего критерия Уэлча t (* p <0,05, ** p <0,01, *** p <0,001). Пунктирная линия представляет один и тот же уровень выражения между условиями Single и Multi. ( F ) Сравнение уровней Bet4-TAP между диким животным и каждым мутантом в одиночных или множественных условиях.Статистическую значимость определяли с помощью одностороннего критерия Уэлча t (* p <0,05, ** p <0,01). Пунктирная линия представляет одинаковый уровень экспрессии между WT и каждым мутантом.
https://doi.org/10.1371/journal.pgen.1009091.g006
NatD-опосредованная дозовая компенсация исходного белка MH Bet4
Селективность NatD-опосредованного N-ацетилирования происходит от мотива последовательности, распознаваемого NatD, и его сайта узнавания субстрата, адаптированного для этого мотива [37].N-концом Pop3 является SGSL после удаления N-конца Met с помощью Met-аминопептидаз, который отличается от известных мотивов, участвующих в гистоновых субъединицах: SGGK (h3A) и SGRG (h5) [38]. Таким образом, мы исследовали, подвергается ли Bet2, который является стартовым белком MSGSL и субъединицей гетеродимера Bet2-Bet4 [42], NatD-опосредованной компенсации. Мы обнаружили, что Bet2 компенсируется в одинаковой степени в naa40, Δ и клетках WT (фиг. 6D), указывая на то, что первого MSGSL недостаточно для определения субстратов опосредованной NatD компенсации.Неожиданно Bet4 был значительно меньше компенсирован в naa40 Δ по сравнению с клетками WT (рис. 6E), и действительно, Bet4 был стабильным при увеличении числа копий BET4 в naa40 Δ клетках из-за увеличенного периода полужизни (S5C и S5D Рис). Это явление было неожиданным, потому что Bet4 является исходным белком MH, который практически не N-ацетилирован, в то время как N-ацетилирование Bet4 было обнаружено, но его ответственные NAT не были идентифицированы [36, 43].
Далее мы исследовали вклад других NAT в компенсацию Bet4 и обнаружили, что Bet4 был значительно меньше компенсирован в ячейках naa20 Δ (рис. 6E).Таким образом, NatB и NatD участвовали в компенсации Bet4. Мы также обнаружили увеличение компенсации Bet4 в клетках naa10 и Δ (рис. 6E), что указывает на опосредованный NatA блок деградации Bet4 во время компенсации. Хотя эти наблюдения сходны с Rmp1 в naa10 Δ и naa20 Δ и Pop5 в naa10 Δ клетках (Fig 4B-4E), оба эффекта делеции NAT наблюдались только для Bet4. Следовательно, Bet4 был идентифицирован как компенсированный белок, стехиометрия которого двунаправленно контролируется множественными NAT при генетических нарушениях.
Чтобы проверить, контролируют ли NatB и NatD уровень Bet4 аддитивным образом, мы проанализировали naa20 Δ naa40 Δ клеток. Мы наблюдали, что Bet4 был компенсирован у этого двойного мутанта в той же степени у одиночных мутантов naa20 Δ и naa40 Δ (рис. 6E). Однако сравнение уровней белка между одиночными и двойными мутантами показало, что Bet4 был немного, но значительно более распространен в клетках naa20, Δ, naa40, Δ (Фиг.6F и S9 Фиг).Этот небольшой аддитивный эффект двойной делеции, по-видимому, согласуется с наблюдением, что уровень Bet4 был близок к некомпенсированному уровню даже у одиночных мутантов. Поскольку Bet4 был стабилизирован у двойного мутанта, несмотря на увеличение числа копий BET4 и , как NatB, так и NatD независимо вносят вклад в контроль уровня Bet4 даже в невозмущенных условиях. Эти результаты предполагают, что их функция в контексте деградации Bet4 не является избыточной.
Discussion
Чтобы исследовать, насколько широко распространенный путь правила Ac / N-конца объясняет дозовую компенсацию, мы систематически измеряли вклад E3 ubiquitin ligases и NATs в компенсацию субъединиц RNase P / MRP (Figs 2 and 4).Наши данные, состоящие из 18 субъединиц и 12 факторов контроля качества, всего 114 комбинаций, продемонстрировали, что в компенсации участвуют несколько убиквитин-лигаз E3 и NAT. Опосредованная NAT компенсация наблюдалась для 7 из 14 протестированных субъединиц: 6 из 9 субъединиц комплексов РНКаза P / MRP, ни одной из трех субъединиц гистона h3A / h5 и 1 из 2 субъединиц комплекса Bet2-Bet4 (рис. 4 и 6). ). Однако, учитывая, что отсутствие NAT не полностью снижает компенсацию тестируемых субъединиц, за исключением Pop3 в naa40 Δ и Bet4 в клетках naa20 Δ и naa40 Δ, контроль стехиометрии мультибелковых комплексов зависит только частично от Путь правила Ac / N-конца.
В этом исследовании мы манипулировали количеством копий гена, чтобы нарушить клеточные системы и выяснить механизмы буферизации нарушений экспрессии генов. Наши данные показали, что NAT вносят вклад в протеолиз при экзогенной сверхэкспрессии субъединиц P / MRP РНКазы, тогда как без сверхэкспрессии уровни эндогенного белка почти не затрагиваются у мутантов NAT по сравнению с мутантами E3 (рис. 2C и 2F и S6, рис.). Это согласуется с недавним всесторонним анализом, показывающим, что N-ацетилирование редко действует как сигнал деградации в физиологических условиях у дрожжей [24].Следовательно, протеолиз, опосредованный NAT, по-видимому, в основном запускается при генетических нарушениях. Следует отметить, что поскольку посттрансляционная дозовая компенсация широко используется для достижения надлежащей стехиометрии в невозмущенных дрожжевых клетках [44], физиологически индуцированный стехиометрический дисбаланс также регулируется компенсацией.
Мы идентифицировали Tom1 как фактор системы контроля стехиометрии (Рис. 2A и 2B), что согласуется с Tom1-опосредованной деградацией избыточных рибосомных субъединиц [19].Следует отметить, что, как обсуждалось в [19], основная изоэлектрическая точка является общей характеристикой известных субстратов Tom1 (Cdc6, Hht2, Yra1 и Rpl26a) и субъединиц РНКазы P / MRP, за исключением Pop8 (S10 Fig) [19, 45–47]. Слабое электрическое взаимодействие между Tom1 с кислой изоэлектрической точкой и Pop8 может объяснить, почему степень компенсации Pop8 ниже, чем у других субъединиц. Кроме того, эти субстраты, а также Tom1 связываются с нуклеиновыми кислотами и локализуются в ядре [48]. Эти характеристики предполагают возможность того, что Tom1 имеет тенденцию компенсировать субъединицы связывания нуклеиновой кислоты.
Мы описали сложность системы контроля стехиометрии (рис. 5, таблица 1), поскольку (i) несколько лигаз E3 Tom1 и Not4 участвуют в компенсации Pop4, Pop5, Pop7, Rpp1 и Rmp1, (ii) N- Конечная последовательность не обязательно является детерминантом того, какие NAT вносят вклад в компенсацию дозы, и (iii) несколько NAT участвуют в компенсации Rmp1 и Bet4. Таким образом, наши результаты показывают, что эта сложность отражает надежность системы контроля стехиометрии. Более того, как утверждается в [49], могут существовать другие механизмы для распознавания несобранных субъединиц и коррекции стехиометрического дисбаланса.Например, последнее исследование показало, что белковая агрегация избыточных субъединиц функционирует как путь компенсации в анеуплоидных дрожжевых клетках [50].
Наше наблюдение, что NatB и NatD независимо несут ответственность за компенсацию Bet4, согласуется с более высокой субстратной специфичностью NatD, чем другие четыре NAT у дрожжей (Рис. 6F и S9 Рис.), Хотя неясно, является ли это специфичным для Bet4 молекулярным фенотипом. . Если NatB может быть функционально избыточным с NatD, Pop3 должен компенсироваться посредством NatB в naa40 Δ, а Bet4 должен компенсироваться через NatB в naa40 Δ и через NatD в naa20 Δ клетках до аналогичных уровней в клетках WT.Поскольку это противоречит нашим результатам, мы предполагаем, что функции NatB и NatD не перекрываются, по крайней мере, в контексте контроля стехиометрии. Кроме того, поскольку мы наблюдали более сильное влияние удаления NAA20 , чем NAA10 на рост клеток и фенотип синтетического дефекта роста naa10 Δ naa20 Δ клеток (S7 рис.), NatA и NatB могут быть по крайней мере частично вовлечены. в другом процессе. Это наблюдение не исключает возможности функциональной избыточности, а скорее предполагает аддитивные функции NatA и NatB.Систематическое исследование мутантов, лишенных множественных NAT, необходимо для определения, существуют ли перекрывающиеся функции между NAT или между ними, а затем для раскрытия более общих принципов их функционального взаимодействия. Одна линия поддержки этой возможности исходит из аспекта эволюционной консервации NAT, поскольку экспрессия человеческого NatF (Naa60) увеличивает долю N-ацетилированных белков с 68% до 78% в дрожжах [51], и микроскопический анализ Arl3, субклеточная локализация которого зависит от NatC-опосредованного N-ацетилирования, демонстрирует функциональную избыточность между NatC дрожжей и NatF человека [52].
Компенсация Pop3 в ячейках naa40 Δ и Bet4 в клетках naa20 Δ и naa40 Δ была почти полностью снижена (рис. 4H, 4I и 6E), что подтверждает зависимость контроля стехиометрии от правила Ac / N-конца. путь [11]. Однако это редкие случаи в нашем исследовании и первые примеры NatD-опосредованной компенсации доз, и канонические субстраты NatD гистон h3A / h5 были компенсированы даже в отсутствие Naa40 и других NAT (Рис. 6A – 6C, S8 Рис.). Важно, что в соответствии со стабилизацией субъединицы гистона h4 Hht2 с помощью делеции TOM1 [46], h3A / h5 менее компенсируется в tom1 Δ клетках.Эти результаты предполагают, что несобранные субъединицы избирательно деградируют не только по правилу Ac / N-конца, но также и другими путями, независимыми от N-ацетилирования. Это дополнительно подтверждается наблюдением, что Rpp1 был почти полностью некомпенсирован в not4 Δ, но не во всех мутантах NATs (Figs 2D, 2E и 4), указывая тем самым, что Not4-опосредованная компенсация не обязательно требует NATs. Следовательно, выявление новых лигаз компенсации E3 важно для лучшего понимания механизма дозовой компенсации.Например, поскольку скрининг с использованием делеционных и чувствительных к температуре мутантов всех несущественных и нескольких основных генов лигазы E3 идентифицировал HEL1 , HEL2 , PEP5 и SNT2 , делеция которых приводит к дефектной деградации избытка гистон h4 / h5 [53], они, по-видимому, являются кандидатами на компенсацию E3, отвечающую за локализованные в ядре белки. В самом деле, хотя деградация избытка Rpl26a в основном опосредуется Tom1, уровень Rpl26a повышен в pep5 Δ по сравнению с клетками WT [19].
Мы также обнаружили, что Pop5 и Bet4 дестабилизируются, когда они сверхэкспрессируются в naa10 Δ клетках (рис. 4B, 4C и 6E). Аналогичным образом, предыдущие исследования показали, что белки с неацетилированным N-концом Met, за которым следует небольшой гидрофобный остаток, и исходные белки MN дестабилизируются в клетках naa30 Δ и naa20 Δ, соответственно, по пути правила Arg / N-конца [ 24, 54]. Однако поскольку N-концевые последовательности Pop5 (MV) и Bet4 (MH) не подходят для этих случаев, их деградация в клетках naa10 и Δ может быть ускорена неизвестными механизмами.Мы предполагаем, что сложная стехиометрия надежно поддерживается путем переключения путей деградации в ответ на возмущения в системе контроля стехиометрии. Действительно, взаимодействие между правилом Arg / N-конца и путями правила Ac / N-конца было предложено на основании наблюдения, что короткоживущий репортерный белок и Msn4 синергетически стабилизируются в двойных мутантах naa20 Δ ubr1 Δ и naa30 Δ ubr1 Δ соответственно [27, 54].
Контроль качества мультибелковых комплексов влияет на широкий спектр биологических процессов, поскольку клеточные системы основаны на функциональных комплексах.Комплексы РНКазы Р и MRP ответственны за созревание тРНК и рРНК соответственно [28]. Кроме того, Pop1, Pop6 и Pop7 являются общими с теломеразой [55]. Наше открытие надежного контроля баланса доз между Pop6 и Pop7, по-видимому, соответствует их функциональной важности (Рис. 3). Следовательно, возможно, что анализ дозовой компенсации по всему геному дает представление о взаимосвязи между функциональной важностью и существенностью мультибелковых комплексов и тем, насколько строго контролируется внутриклеточный уровень их субъединиц.Более того, тот же анализ у мутантов NATs приводит к более точной оценке того, насколько широко распространенный путь правила Ac / N-конца способствует контролю стехиометрии.
В более общем плане дозовая компенсация может играть роль отказоустойчивого механизма для формирования стехиометрии протеома, как обсуждалось выше. В соответствии с этой концепцией, субъединицы паралогичных комплексов имеют тенденцию компенсировать друг друга за модуляцию белкового интерактома [56, 57]. Более того, опосредованная протеолизом компенсация повсеместно буферизует физиологически вызванный стехиометрический дисбаланс во время мейоза [44], что может объяснить, как клетки справляются с субнабором субъединиц, сверхэкспрессируемых в невозмущенных условиях [13].Это открытие также предполагает, что дозовая компенсация оптимизирует стехиометрию протеома по ходу клеточного развития на каждой стадии мейоза. Взятые вместе, дальнейшее изучение сложности и рабочих принципов системы контроля стехиометрии поможет понять механизм буферизации для концентрации протеома против как внутренних, так и внешних генетических нарушений.
Материалы и методы
Штаммы, плазмиды и среды
Для конструирования штаммов с одиночной делецией, в которых отсутствует ген, кодирующий убиквитинлигазу E3 ( TOM1 , NOT4 , DOA10 , LTN1 или HEL2 ) или N-ацетилтрансферазы ( NAA10 , NAA20 , NAA30 , NAA40 или NAA50 ) и несущих интересующий ген, меченный TAP, использовали коллекцию делеций и коллекцию TAP гаплоидных дрожжей (Dharmacon).BY4741, несущий Pop4-TAP, Rmp1-TAP или Bet4-TAP, был сконструирован в этом исследовании. Геномная ДНК каждого штамма с одиночной делецией была извлечена, а затем каждый локус, замененный кассетой kanMX4 , был амплифицирован с помощью ПЦР с библиотекой праймеров, подготовленной в нашем предыдущем исследовании [5]. Для конструирования штаммов с двойной делецией ( naa10 Δ naa20 Δ и naa20 Δ naa40 Δ) кассету hphNT1 , амплифицированную из плазмиды pFA6-hphNT1, использовали для замены локуса NAA20 в геноме naa10 Δ, несущие Pop4-TAP или Rmp1-TAP, и naa40 Δ, несущие Bet4-TAP, соответственно.Штаммы, меченные ТАП, трансформировали продуктами ПЦР методом ацетата лития и отбирали на планшетах YPD, содержащих G418 (200 мкг / мл) или HygB (300 мкг / мл). Для конструирования плазмид фрагмент ДНК каждой целевой области амплифицировали из генома и клонировали в pTOW40836 путем гомологичной рекомбинации в клетках BY4741. Описанные выше штаммы с TAP-меткой трансформировали плазмидами и отбирали на среде SC без урацила.
Измерение количества копий плазмиды
Число копий плазмиды измеряли методом gTOW [58].Клетки, выращенные в логарифмической фазе для вестерн-блоттинга, собирали для анализа gTOW из той же культуры. Вкратце, 200 мкл культуры центрифугировали, осажденные клетки суспендировали в 100 мкл раствора зимолиазы [2,5 мг / мл зимолиазы-100Т, растворенной в 1,2 М сорбите, 10 мМ фосфата натрия (pH 7,5)] и инкубировали при 37 ° С. ° C в течение 15 мин для экстракции ДНК. Суспензию нагревали при 100 ° C в течение 10 мин, охлаждали при –80 ° C в течение 10 мин и снова нагревали при 100 ° C в течение 10 мин. После охлаждения до комнатной температуры (RT) с последующим центрифугированием супернатант подвергали количественной ПЦР в реальном времени с помощью Lightcycler 480 (Roche).Для ПЦР использовали праймеры для LEU3 в геноме или leu2d гена в pTOW40836 и SYBR Green I Master (Roche). Полученное число копий плазмиды было рассчитано на основе уровней экспрессии генов LEU3 и leu2d в соответствии с методом, описанным ранее [58].
Вестерн-блоттинг
Клетки выращивали при 30 ° C в 2 мл среды SC-Ura в течение ночи, а затем измеряли оптическую плотность при 600 нм (OD 600 ) и инокулировали в свежую среду при исходной OD 600 0.5 в 3 мл. Через 4 часа 1 единица OD 600 собирали в логарифмической фазе, когда OD 600 была около 1. Клетки обрабатывали 1 мл 0,2 н. NaOH в течение 5 минут при комнатной температуре и центрифугировали при 15000 об / мин в течение 1 минуты. Осажденные клетки суспендировали в 50 мкл буфера для образцов 1 × NuPAGE LDS (Invitrogen), содержащего 100 мМ DTT, и нагревали при 100 ° C в течение 5 минут. Для анализа белков, меченных ТАП, были проанализированы двукратные серийно разведенные лизаты, как описано ниже, и мы подтвердили, что 0,025 OD 600 единиц обеспечивают соответствующий сигнал в линейном диапазоне.Экстракт, разбавленный в 8 раз буфером для образцов NuPAGE LDS, соответствующим 0,025 OD 600 единиц, разделяли электрофорезом в полиакриламидном геле с додецилсульфатом лития (SDS-PAGE) на NuPAGE 4% –12% Bis-Tris Gel ( Invitrogen). Для анализа белков, меченных GFP, 0,2 OD 600 единиц подвергали SDS-PAGE. Разделенные белки переносили на PVDF-мембрану с использованием PVDF-мембраны iBlot Transfer Stack (Invitrogen). Блоттированную мембрану обрабатывали PBST [1 × PBS, 0.1% Tween 20] в течение 10 минут, а затем блокировали 4% обезжиренным молоком в PBST в течение 1 часа при комнатной температуре. Белки, меченные TAP, исследовали с помощью PAP (Sigma-Aldrich) (1: 2000) в течение 1 ч при комнатной температуре. Белки, меченные GFP, исследовали с помощью антитела против GFP (Roche) (1: 1000) и вторичного антитела, конъюгированного с пероксидазой (Nichirei Biosciences) (1: 1000), в течение 1 ч при комнатной температуре. После трехкратной промывки мембраны PBST в течение 5 мин хемилюминесценцию индуцировали добавлением 500 мкл субстрата максимальной чувствительности SuperSignal West Femto Maximum Sensitivity Substrate (Thermo Scientific) на мембрану и детектировали с помощью анализатора изображений LAS-4000 (Fujifilm) и ImageQuant LAS 4000 ( GE Healthcare).Количественную оценку интенсивности полосы проводили с использованием ImageQuant TL (GE Healthcare), а кратное изменение рассчитывали после вычитания фона. После трехкратной промывки мембраны стерильной водой в течение 5 мин, общий белок визуализировали путем окрашивания Кумасси с помощью SimplyBlue SafeStain (Invitrogen), чтобы подтвердить равную загрузку белков. Окрашенную мембрану оцифровывали с использованием анализатора изображений LAS-4000 и ImageQuant LAS 4000. Типичные блоты и соответствующие пятна Кумасси показаны на S11 рис.
Для анализа гистона h3A / h5 и гетеродимера Bet2-Bet4 собранные клетки обрабатывали 1 мл 0,2 н. NaOH в течение 5 мин при комнатной температуре. Клетки суспендировали в буфере для образцов 1 × NuPAGE LDS (Invitrogen), а затем нагревали при 70 ° C в течение 10 минут. Супернатант, соответствующий 0,5 OD 600 единиц, метили EzLabel FluoroNeo (ATTO) и подвергали SDS-PAGE с последующим вестерн-блоттингом с PAP (Sigma-Aldrich) (1: 2000), как описано выше.
Циклогексимидный анализ
Скорость разложения Pop3-TAP и Bet4-TAP измеряли, как описано ранее [7].Вкратце, культуру в логарифмической фазе, соответствующую 1 единице клеток OD 600 , собирали для временной точки 0, а затем добавляли CHX до конечной концентрации 200 мкг / мл. Клетки собирали через 1, 2, 4 и 6,5 ч с последующей экстракцией общего белка в буфере для образцов 1 × NuPAGE LDS. Супернатант, соответствующий 0,5 OD 600 единиц, подвергали Вестерн-блоттингу с использованием PAP, как описано выше. Оставшийся уровень белка в каждый момент времени рассчитывали по сравнению с уровнем в момент времени 0.
Дополнительная информация
S1 Рис. Идентификация лигаз убиквитина E3, участвующих в дозовой компенсации.
( A ) Экспериментальная установка для скрининга убиквитин-лигаз E3, участвующих в дозовой компенсации. Белки с компенсированной дозировкой, меченные зеленым флуоресцентным белком (GFP), были экспрессированы из многокопийной плазмиды pTOW40836, содержащей нативные регуляторные последовательности, включая промотор и 5´ и 3´ нетранслируемые области. Если тестируемая лигаза E3 не отвечает за деградацию целевого белка, уровень белка одинаков для мутантных клеток WT и E3 (левая и средняя панели).С другой стороны, если целевой белок разрушается с помощью тестируемой лигазы E3, уровень белка увеличивается в мутанте E3 по сравнению с клетками WT (правая и левая панели). ( B ) Вестерн-блоттинг GFP-меченных доза-компенсированных белков в мутантах E3 с использованием антитела против GFP. Окрашивание кумасси белка массой 50 кДа, соответствующего енолазе, показано как контроль нагрузки. ( C ) Номер копии гена во время компенсации дозировки. Вестерн-блоттинг обнаружил повышенное количество Pop5 в tom1 Δ и not4 Δ и Saw1 в doa10 Δ по сравнению с таковыми в клетках WT, хотя количество копий плазмиды было почти таким же среди тестируемых штаммов.Таким образом, Tom1 и Not4 и Doa10 были идентифицированы как E3 ubiquitin ligases, участвующие в деградации Pop5 и Saw1, соответственно. Столбчатая диаграмма представляет количество копий pTOW40836, несущего каждый из указанных генов, у мутантов WT или E3. Среднее количество копий ± стандартное отклонение. были рассчитаны из четырех технических повторений.
https://doi.org/10.1371/journal.pgen.1009091.s001
(TIF)
S2 Рис. Количественная оценка данных вестерн-блоттинга.
( A, B ) PVDF-мембрана, окрашенная Кумасси ( A ) после вестерн-блоттинга с PAP ( B ) двукратных серийно разведенных лизатов клеток, полученных из tom1 Δ Pop3-TAP или Pop8-TAP клеток культивируется в Единичном состоянии.( C ) Количественное определение площади белка массой 50 кДа в ( A ). Интенсивность сигнала каждой полосы измеряли после вычитания фона, а суммарную интенсивность откладывали на оси ординат. Количество лизата из tom1 Δ Pop3-TAP и Pop8-TAP имело коэффициент корреляции ( R 2 ) 0,99 и 0,97 с чистой интенсивностью, соответственно. ( D ) Количественное определение площади Pop3-TAP и Pop8-TAP в ( B ).Интенсивность сигнала каждой полосы измеряли после вычитания фона, а суммарную интенсивность откладывали на оси ординат. Количество лизата из tom1, Δ Pop3-TAP и Pop8-TAP имело R 2 0,99 и 0,99 с чистой интенсивностью, соответственно. ( E ) Сравнение уровней белка между одиночными и множественными условиями у WT или каждого мутанта. В качестве примера показан Pop4-TAP в ячейках WT и tom1 Δ. Интенсивность полосы Pop4-TAP в условиях Multi была разделена на таковую в условиях Single в каждом штамме.Для нормализации использовали полную полосу Кумасси на мембране из ПВДФ. ( F ) Сравнение уровней белка между диким животным и каждым мутантом в одиночных или множественных условиях. Интенсивность полосы Pop4-TAP в tom1 Δ была разделена на интенсивность полосы в клетках WT в каждом условии количества копий. Данные взяты из рис. 2A – 2C.
https://doi.org/10.1371/journal.pgen.1009091.s002
(TIF)
S3 Рис. Вестерн-блоттинг субъединиц РНКазы P / MRP в
doa10 Δ клетках.
( A – C ) Doa10 не участвует в компенсации дозы субъединиц РНКазы P / MRP. Вестерн-блоттинг Pop3-TAP в двух биологических повторностях ( A ) и других субъединиц РНКазы P / MRP, за исключением Pop5 ( B, C ), с использованием PAP в doa10 Δ клетках. Пунктирная линия представляет один и тот же уровень выражения между условиями Single и Multi.
https://doi.org/10.1371/journal.pgen.1009091.s003
(TIF)
S5 Рис. CHX эксперименты по преследованию Pop3-TAP и Bet4-TAP в
naa40 Δ клетках.
( A, B ) CHX эксперименты по преследованию Pop3-TAP в клетках WT и naa40 Δ. Вестерн-блоттинг с PAP и SDS-PAGE белка 50 кДа в качестве контроля загрузки ( A ). Количественная оценка уровней Pop3-TAP в условиях одиночного (слева) или множественного (справа) ( B ). Средний уровень протеина ± стандартное отклонение. был рассчитан из трех биологических повторов. ( C, D ) То же, что в ( A, B ), за исключением того, что показаны вестерн-блоттинг и количественное определение Bet4-TAP.
https://doi.org/10.1371/journal.pgen.1009091.s005
(TIF)
S6 Рис. Влияние NAT на уровень эндогенного белка субъединиц РНКазы P / MRP.
( A – E ) Сравнение уровней белка между WT и naa10 Δ ( A ), naa20 Δ ( B ), naa30 Δ ( C ), naa40 Δ ( D ) или naa50 Δ ( E ) ячеек в одиночных или множественных условиях.Среднее кратное изменение ± стандартное отклонение. был рассчитан из трех биологических повторов. Пунктирная линия представляет одинаковый уровень экспрессии между WT и мутантными клетками. Данные взяты с рис. 4B – 4K.
https://doi.org/10.1371/journal.pgen.1009091.s006
(TIF)
S7 Рис. Синтетический дефект роста клеток
naa10 Δ naa20 Δ.
( A ) Точечный тест клеток naa10 Δ naa20 Δ. Ночные культуры указанных штаммов в среде YPD, разбавленной до 0.OD 600 и его 10-кратные серийные разведения наносили на чашки YPD и инкубировали при 30 ° C в течение 6 дней. ( B ) Вестерн-блоты Pop4-TAP (слева) и Rmp1-TAP (справа) в двойном мутанте naa10 Δ naa20 Δ. Все штаммы культивировали в среде YPD из-за отсутствия образования колоний клеток naa10 Δ naa20 Δ, трансформированных многокопийными плазмидами pTOW40836, несущими POP4 или RMP1 на планшете SC-Ura. Количественный анализ из трех биологических повторов показан ниже.
https://doi.org/10.1371/journal.pgen.1009091.s007
(TIF)
S8 Рис. Компенсация дозировки Hhf2 у мутантов NAT.
Вестерн-блоттинг Hhf2-TAP в клетках WT, naa10 Δ, naa20 Δ, naa30 Δ и naa40 Δ клетках. Hhf2-TAP был обнаружен с помощью PAP (вверху) и определен количественно (внизу). Пунктирная линия представляет один и тот же уровень выражения между условиями Single и Multi.
https://doi.org/10.1371/journal.pgen.1009091.s008
(TIF)
S9 Рис. Bet4 было немного, но значительно больше в клетках
naa20, Δ naa40 Δ.
( A, B ) Более высокий уровень Bet4-TAP у двойных мутантов naa20 Δ naa40 Δ по сравнению с одиночными мутантами в условиях Single ( A ) и Multi ( B ). Показана количественная оценка трех биологических повторов. Данные взяты из фиг. 6F.
https://doi.org/10.1371/journal.pgen.1009091.s009
(TIF)
S11 Рис. Репрезентативные блоты и соответствующие окрашивания Кумасси вестерн-блотов субъединиц РНКазы P / MRP в лигазах E3 или мутантах NAT.
(A – G) Показаны полноразмерные блоты и пятна Кумасси, использованные для анализа tom1 Δ ( A ), not4 Δ ( B ), naa10 Δ ( C ) ), naa20 Δ ( D ), naa30 Δ ( E ), naa40 Δ ( F ) или naa50 Δ ( G ) мутанты, соответствующие рис. 2A, 2D , 4B, 4D, 4F, 4H и 4J.
https://doi.org/10.1371/journal.pgen.1009091.s011
(TIF)
Благодарности
Благодарим сотрудников лаборатории «Мория» за поддержку.
Список литературы
- 1.
Чжэн XY, О’Ши Е.К. Цианобактерии поддерживают постоянную концентрацию белка, несмотря на вариацию числа копий генома. Cell Rep. 2017; 19 (3): 497–504. pmid: 28423314 - 2.
Кафри М., Метцл-Раз Э, Йона Г., Баркай Н. Стоимость производства белка.Cell Rep. 2016; 14 (1): 22–31. pmid: 26725116 - 3.
Дилл К.А., Гош К., Шмит Дж.Д. Физические пределы клеток и протеомов. Proc Natl Acad Sci U S. A. 2011; 108 (44): 17876–82. pmid: 22006304 - 4.
Сопко Р., Хуанг Д., Престон Н., Чуа Г., Папп Б., Кафадар К. и др. Картирование путей и фенотипов с помощью систематической сверхэкспрессии генов. Mol Cell. 2006. 21 (3): 319–30. pmid: 16455487 - 5.
Маканае К., Кинтака Р., Макино Т., Китано Х., Мория Х. Идентификация дозозависимых генов в Saccharomyces cerevisiae с использованием генетического метода перетягивания каната.Исследование генома. 2013. 23 (2): 300–11. pmid: 23275495 - 6.
Мория Х. Количественная природа экспериментов со сверхэкспрессией. Mol Biol Cell. 2015; 26 (22): 3932–9. pmid: 26543202 - 7.
Исикава К., Маканаэ К., Ивасаки С., Инголия Н. Т., Мория Х. Посттрансляционные буферы для компенсации дозировки генетических нарушений стехиометрии белковых комплексов. PLoS Genet. 2017; 13 (1): e1006554. pmid: 28121980 - 8.
Stingele S, Stoehr G, Peplowska K, Cox J, Mann M, Storchova Z.Глобальный анализ генома, транскриптома и протеома показывает ответ на анеуплоидию в клетках человека. Молекулярная системная биология. 2012; 8: 608. pmid: 22968442 - 9.
Дефур Н., Хван С., О’Салливан С., Доджсон С.Е., Гайги С.П., Амон А. и др. Количественный протеомный анализ выявляет посттрансляционные реакции на анеуплоидию у дрожжей. eLife. 2014; 3: e03023. pmid: 25073701 - 10.
Торрес Э.М., Сокольски Т., Такер С.М., Чан Л.Й., Бозелли М., Данхэм М.Дж. и др. Влияние анеуплоидии на физиологию клеток и деление клеток у гаплоидных дрожжей.Наука (Нью-Йорк, Нью-Йорк). 2007. 317 (5840): 916–24. pmid: 17702937 - 11.
Шеморри А., Хванг С.С., Варшавский А. Контроль качества и стехиометрии белка с помощью N-концевого ацетилирования и пути правила N-конца. Mol Cell. 2013; 50 (4): 540–51. pmid: 23603116 - 12.
Папп Б, Пал С, Херст Л.Д. Чувствительность к дозировке и эволюция семейств генов у дрожжей. Природа. 2003. 424 (6945): 194–7. pmid: 12853957 - 13.
Li GW, Burkhardt D, Gross C, Weissman JS.Количественная оценка абсолютных скоростей синтеза белка раскрывает принципы, лежащие в основе распределения клеточных ресурсов. Клетка. 2014. 157 (3): 624–35. pmid: 24766808 - 14.
Таггарт Дж. К., Ли Г. В.. Производство компонентов белкового комплекса является стехиометрическим и не имеет общей регуляции обратной связи у эукариот. Клеточные системы. 2018. pmid: 30553725 - 15.
Ingolia NT, Hussmann JA, Weissman JS. Профилирование рибосом: глобальные взгляды на перевод. Cold Spring Harb Perspect Biol. 2018 г.pmid: 30037969 - 16.
Сун МК, Рейтсма Дж. М., Суэредоски М. Дж., Хесс С., Деше Р. Дж. Рибосомные белки, продуцируемые в избытке, разрушаются убиквитин-протеасомной системой. Mol Biol Cell. 2016; 27 (17): 2642–52. pmid: 27385339 - 17.
Янагитани К., Юшкевич С., Хегде Р.С. UBE2O — это фактор контроля качества для сирот мультибелковых комплексов. Наука (Нью-Йорк, Нью-Йорк). 2017; 357 (6350): 472–5. pmid: 28774922 - 18.
Hwang CS, Shemorry A, Варшавский А.N-концевое ацетилирование клеточных белков создает специфические сигналы деградации. Наука (Нью-Йорк, Нью-Йорк). 2010. 327 (5968): 973–7. pmid: 20110468 - 19.
Сун М.К., Поррас-Якуши Т.Р., Рейтсма Дж. М., Хубер Ф. М., Суэредоски М. Дж., Хельц А. и др. Консервативный путь контроля качества, который опосредует деградацию несобранных рибосомных белков. eLife. 2016; 5. pmid: 27552055 - 20.
Акснес Х., Ри Р., Арнесен Т. Ко-трансляционная, пост-трансляционная и некаталитическая роль N-концевых ацетилтрансфераз.Mol Cell. 2019; 73 (6): 1097–114. pmid: 30878283 - 21.
Нгуен К.Т., Ли К.С., Мун С.Х., Чыонг Н.Т., Парк С.К., Хван С.С. N-концевое ацетилирование и путь правила N-конца контролируют деградацию липидного капельного белка PLIN2. J Biol Chem. 2019; 294 (1): 379–88. pmid: 30425097 - 22.
Варшавский А. Путь правила N-конца и регуляция протеолизом. Protein Sci. 2011. 20 (8): 1298–345. pmid: 21633985 - 23.
Харпер Дж. У., Беннетт Э. Дж. Сложность протеома и силы, которые вызывают дисбаланс протеома.Природа. 2016; 537 (7620): 328–38. pmid: 27629639 - 24.
Кац И., Хмелинский А., Кшонсак М., Хубер Ф., Книсс Р.А., Бартосик А. и др. Картирование сигналов деградации и путей в N-терминальном эукариотическом. Mol Cell. 2018; 70 (3): 488–501 e5. pmid: 29727619 - 25.
Gardner RG, Nelson ZW, Gottschling DE. Контроль качества белка, опосредованного деградацией, в ядре. Клетка. 2005. 120 (6): 803–15. pmid: 15797381 - 26.
Бартель Б, Ваннинг I, Варшавский А.Компонент распознавания пути правила N-конца. Embo j. 1990. 9 (10): 3179–89. pmid: 2209542 - 27.
Нгуен К.Т., Ким Дж.М., Парк С.Е., Хван К.С. Удаление белков метионином на N-конце создает третичные дестабилизирующие N-дегроны пути правила Arg / N-конца. J Biol Chem. 2019; 294 (12): 4464–76. pmid: 30674553 - 28.
Есакова О, Красильников АС. Из белков и РНК: семейство РНКазы P / MRP. РНК. 2010. 16 (9): 1725–47. pmid: 20627997 - 29.
Передерина А, Березин И, Красильников АС.Восстановление in vitro и анализ РНП эукариотической РНКазы P. Nucleic Acids Res. 2018. 46 (13): 6857–68. pmid: 29722866 - 30.
Димитрова Л.Н., Куроха К., Татемацу Т., Инада Т. Возрождающаяся пептидозависимая остановка трансляции приводит к Not4p-опосредованной деградации протеина протеасомой. J Biol Chem. 2009. 284 (16): 10343–52. pmid: 101
- 31.
Brandman O, Hegde RS. Контроль качества белка, связанного с рибосомами. Nat Struct Mol Biol. 2016; 23 (1): 7–15. pmid: 26733220 - 32.Брэндман О., Стюарт-Орнштейн Дж., Вонг Д., Ларсон А., Уильямс С.С., Ли Г.В. и др. Связанный с рибосомами комплекс контроля качества запускает деградацию растущих пептидов и сигнализирует о стрессе трансляции. Клетка. 2012. 151 (5): 1042–54. pmid: 23178123
- 33.
Мацуо Ю., Икеучи К., Саеки Ю., Ивасаки С., Шмидт С., Удагава Т. и др. Убиквитинирование застопорившейся рибосомы запускает связанный с рибосомой контроль качества. Nat Commun. 2017; 8 (1): 159. pmid: 28757607 - 34.
Передерина А, Есакова О, Коч Х, Шмитт М.Е., Красильников АС.Специфическое связывание гетеродимера Pop6 / Pop7 со стеблем P3 дрожжевой РНКазы MRP и РНК РНКазы P. РНК. 2007. 13 (10): 1648–55. pmid: 17717080 - 35.
Передерина А, Есакова О, Куан С, Ханова Э, Красильников АС. Рибонуклеазы эукариот P / MRP: кристаллическая структура домена P3. Embo j. 2010. 29 (4): 761–9. pmid: 20075859 - 36.
Акснес Х., Дразич А., Мари М., Арнесен Т. В первую очередь: важные белковые метки N-концевыми ацетилтрансферазами. Trends Biochem Sci.2016; 41 (9): 746–60. pmid: 27498224 - 37.
Magin RS, Liszczak GP, Marmorstein R. Молекулярная основа для гистонового h5- и h3A-специфичного аминоконцевого ацетилирования с помощью NatD. Структура. 2015; 23 (2): 332–41. pmid: 25619998 - 38.
Song OK, Wang X, Waterborg JH, Sternglanz R. Нальфа-ацетилтрансфераза, ответственная за ацетилирование N-концевых остатков гистонов h5 и h3A. J Biol Chem. 2003. 278 (40): 38109–12. pmid: 120 - 39.
Hole K, Van Damme P, Dalva M, Aksnes H, Glomnes N, Varhaug JE и др.Человеческая N-альфа-ацетилтрансфераза 40 (hNaa40p / hNatD) является консервативной из дрожжей и ацетилирует на N-конце гистоны h3A и h5. PLoS One. 2011; 6 (9): e24713. pmid: 21935442 - 40.
Полевода Б., Хоскинс Дж., Шерман Ф. Свойства Nat4, N (альфа) -ацетилтрансферазы Saccharomyces cerevisiae, которая модифицирует N-концы гистонов h3A и h5. Mol Cell Biol. 2009. 29 (11): 2913–24. pmid: 19332560 - 41.
Mullen JR, Kayne PS, Moerschell RP, Tsunasawa S, Gribskov M, Colavito-Shepanski M и др.Идентификация и характеристика генов и мутантов N-концевой ацетилтрансферазы дрожжей. Embo j. 1989. 8 (7): 2067–75. pmid: 2551674 - 42.
Jiang Y, Rossi G, Ferro-Novick S. Bet2p и Mad2p являются компонентами пренилтрансферазы, которая добавляет геранилгеранил к Ypt1p и Sec4p. Природа. 1993. 366 (6450): 84–6. pmid: 8232542 - 43.
Ван Дамм П., Ласа М., Полевода Б., Газкес С., Элосеги-Артола А., Ким Д.С. и др. Анализ N-концевого ацетилома и функциональное понимание N-концевой ацетилтрансферазы NatB.Proc Natl Acad Sci U S. A. 2012; 109 (31): 12449–54. pmid: 22814378 - 44.
Айзенберг А.Р., Хигдон А., Кескин А., Ходапп С., Йованович М., Брар Г.А. Точная посттрансляционная настройка происходит для большинства компонентов белкового комплекса во время мейоза. Cell Rep. 2018; 25 (13): 3603–17.e2. pmid: 305 - 45.
Ким DH, Чжан В., Кепп DM. Домен Hect E3, лигаза Tom1 и белок F-бокса Dia2 контролируют деградацию Cdc6 в фазе G1. J Biol Chem. 2012. 287 (53): 44212–20. pmid: 23129771 - 46.Сингх Р.К., Каббадж М.Х., Пайк Дж., Гунджан А. Уровни гистонов регулируются фосфорилированием и зависимым от убиквитилированием протеолизом. Nat Cell Biol. 2009. 11 (8): 925–33. pmid: 19578373
- 47.
Иглесиас Н., Тутуччи Э., Гвиздек С., Винчигерра П., Фон Дах Э., Корбетт А. Х. и др. Убиквитин-опосредованная динамика мРНП и наблюдение до экспорта мРНК почкующихся дрожжей. Genes Dev. 2010. 24 (17): 1927–38. pmid: 20810649 - 48.
Ха В.К., Фалво СП, Герке Л.К., Кэрролл А.С., Хоусон Р.В., Вайсман Дж.С. и др.Глобальный анализ локализации белка у почкующихся дрожжей. Природа. 2003. 425 (6959): 686–91. pmid: 14562095 - 49.
Юшкевич S, Hegde RS. Контроль качества орфанных белков. Mol Cell. 2018. 71 (3): 443–57. pmid: 30075143 - 50.
Бреннан С.М., Вайтес Л.П., Уэллс Дж. Н., Сантагуида С., Пауло Дж. А., Сторчова З. и др. Агрегация белков опосредует стехиометрию белковых комплексов в анеуплоидных клетках. Genes Dev. 2019. pmid: 31196865 - 51.
Ван Дамм П., Отверстие К, Пимента-Маркес А., Хелсенс К., Вандекеркхове Дж., Мартиньо Р.Г. и др.NatF вносит вклад в эволюционный сдвиг в ацетилировании N-конца белка и важен для нормальной сегрегации хромосом. PLoS Genet. 2011; 7 (7): e1002169. pmid: 21750686 - 52.
Осберг С., Акснес Х., Нинзима С., Мари М., Арнесен Т. Модель комплементации Saccharomyces cerevisiae, основанная на микроскопии, выявляет функциональную консервацию и избыточность N-концевых ацетилтрансфераз. Научный доклад 2016; 6: 31627. pmid: 27555049 - 53.
Сингх Р.К., Гонсалес М., Каббадж М.Х., Гунджан А.Новые лигазы убиквитина E3, которые регулируют уровни гистонового белка в почкующихся дрожжах Saccharomyces cerevisiae. PLoS One. 2012; 7 (5): e36295. pmid: 22570702 - 54.
Kim HK, Kim RR, Oh JH, Cho H, Varshavsky A, Hwang CS. N-концевой метионин клеточных белков как сигнал деградации. Клетка. 2014. 156 (1–2): 158–69. pmid: 24361105 - 55.
Lemieux B, Laterreur N, Perederina A, Noel JF, Dubois ML, Krasilnikov AS, et al. Активная теломераза дрожжей делит субъединицы с рибонуклеопротеинами, РНКазой Р и РНКазой MRP.Клетка. 2016; 165 (5): 1171–81. pmid: 27156450 - 56.
Diss G, Dube AK, Boutin J, Gagnon-Arsenault I, Landry CR. Системный подход к генетическому разделению белковых комплексов в живых клетках. Cell Rep. 2013; 3 (6): 2155–67. pmid: 23746448 - 57.
Diss G, Gagnon-Arsenault I, Dion-Cote AM, Vignaud H, Ascencio DI, Berger CM и др. Дублирование генов может придавать хрупкость, а не устойчивость сети взаимодействия дрожжевых белков.