Гликоген — Glycogen — qaz.wiki
полимер глюкозы, используемый в качестве накопителя энергии у животных
Схематическое двумерное поперечное сечение гликогена: основной белок гликогенина окружен ответвлениями единиц глюкозы . Вся глобулярная гранула может содержать около 30 000 единиц глюкозы.
Гликоген (черные гранулы) в сперматозоидах плоского червя; просвечивающая электронная микроскопия, масштаб: 0,3 мкм
Гликоген является multibranched полисахарида из глюкозы , которая служит в качестве формы хранения энергии в животных , грибов и бактерий. Структура полисахарида представляет собой основную форму хранения глюкозы в организме.
Гликоген функционирует как одна из двух форм энергетических резервов: гликоген — кратковременный, а другая — запасы триглицеридов в жировой ткани (т. Е. Жировой ткани ) для длительного хранения. У человека гликоген вырабатывается и хранится в основном в клетках печени и скелетных мышц . В печени гликоген может составлять 5–6% от сырой массы органа, а печень взрослого человека весом 1,5 кг может хранить примерно 100–120 граммов гликогена. В скелетных мышцах гликоген содержится в низкой концентрации (1-2% от мышечной массы), а скелетные мышцы взрослого человека весом 70 кг хранят примерно 400 граммов гликогена. Количество гликогена, хранящегося в организме, особенно в мышцах и печени, в основном зависит от физической подготовки, основного обмена веществ и привычек питания. Небольшие количества гликогена также обнаруживаются в других тканях и клетках, включая почки , красные кровяные тельца , лейкоциты и глиальные клетки головного мозга . Во время беременности матка также накапливает гликоген для питания эмбриона.
В печени гликоген может составлять 5–6% от сырой массы органа, а печень взрослого человека весом 1,5 кг может хранить примерно 100–120 граммов гликогена. В скелетных мышцах гликоген содержится в низкой концентрации (1-2% от мышечной массы), а скелетные мышцы взрослого человека весом 70 кг хранят примерно 400 граммов гликогена. Количество гликогена, хранящегося в организме, особенно в мышцах и печени, в основном зависит от физической подготовки, основного обмена веществ и привычек питания. Небольшие количества гликогена также обнаруживаются в других тканях и клетках, включая почки , красные кровяные тельца , лейкоциты и глиальные клетки головного мозга . Во время беременности матка также накапливает гликоген для питания эмбриона.
В крови человека постоянно присутствует примерно 4 грамма глюкозы ; У голодных людей уровень глюкозы в крови поддерживается постоянным на этом уровне за счет запасов гликогена в печени и скелетных мышцах. Запасы гликогена в скелетных мышцах служат формой хранения энергии для самих мышц; однако распад мышечного гликогена препятствует поглощению мышечной глюкозы из крови, тем самым увеличивая количество глюкозы в крови, доступной для использования в других тканях.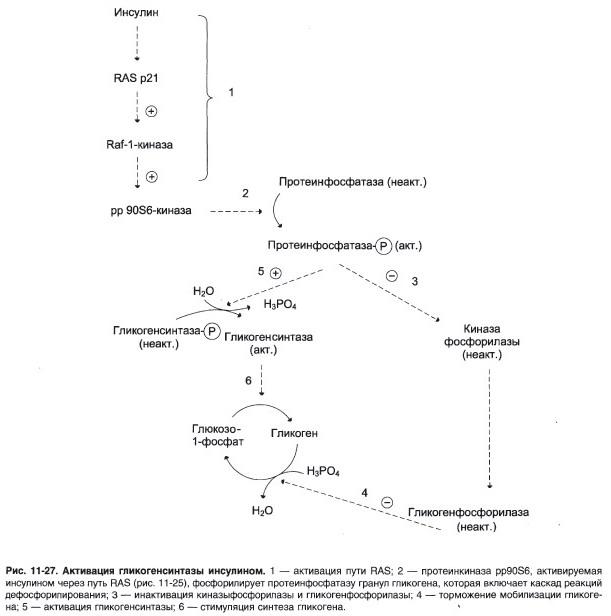 Запасы гликогена в печени служат хранилищем глюкозы для использования во всем организме, особенно в центральной нервной системе . Человеческий мозг потребляет около 60% глюкозы в крови натощак, оседлых особей.
Запасы гликогена в печени служат хранилищем глюкозы для использования во всем организме, особенно в центральной нервной системе . Человеческий мозг потребляет около 60% глюкозы в крови натощак, оседлых особей.
Гликоген — это аналог крахмала , полимера глюкозы, который функционирует как накопитель энергии в растениях . Он имеет структуру, аналогичную амилопектину (компонент крахмала), но более разветвленный и компактный, чем крахмал. Оба представляют собой белые порошки в сухом состоянии. Гликоген находится в форме гранул в цитозоле / цитоплазме многих типов клеток и играет важную роль в цикле глюкозы . Гликоген образует запас энергии, который можно быстро мобилизовать для удовлетворения внезапной потребности в глюкозе, но он менее компактный, чем запасы энергии триглицеридов (липидов). Как таковой, он также является резервом хранения у многих паразитических простейших.
Структура
1,4-α-гликозидные связи в олигомере гликогена
1,4-α-гликозидные и 1,6-гликозидные связи в олигомере гликогена
Гликоген — это разветвленный биополимер, состоящий из линейных цепей остатков глюкозы со средней длиной цепи примерно 8–12 глюкозных единиц и 2000-60 000 остатков на одну молекулу гликогена.
Единицы глюкозы связаны друг с другом линейно α (1 → 4) гликозидными связями от одной глюкозы к другой. Разветвления связаны с цепями, от которых они ответвляются, посредством α (1 → 6) гликозидных связей между первой глюкозой новой ветви и глюкозой в цепи ствола.
Из-за того, как гликоген синтезируется, каждая гранула гликогена имеет в своей основе белок гликогенин .
Гликоген находится в мышцах, печени и жировых клетках, хранящихся в гидратированной форме, состоящей из трех или четырех частей воды на часть гликогена, связанного с 0,45 миллимолями (18 мг) калия на грамм гликогена.
Глюкоза — это осмотическая молекула, которая может оказывать сильное влияние на осмотическое давление в высоких концентрациях, что может приводить к повреждению или гибели клетки, если она хранится в клетке без модификации. Гликоген — это неосмотическая молекула, поэтому его можно использовать в качестве раствора для хранения глюкозы в клетке без нарушения осмотического давления.
Функции
Печень
Когда пища, содержащая углеводы или белок, съедается и переваривается , уровень глюкозы в крови повышается, а поджелудочная железа выделяет инсулин . Глюкоза крови из воротной вены попадает в клетки печени ( гепатоциты ). Инсулин действует на гепатоциты, стимулируя действие нескольких ферментов , в том числе гликогенсинтазы . Молекулы глюкозы добавляются к цепочкам гликогена до тех пор, пока инсулин и глюкоза остаются в изобилии. В этом постпрандиальном или «сытом» состоянии печень забирает из крови больше глюкозы, чем выделяет.
После того, как еда переваривается и уровень глюкозы начинает падать, секреция инсулина снижается, и синтез гликогена прекращается. Когда он необходим для получения энергии , гликоген расщепляется и снова превращается в глюкозу. Гликогенфосфорилаза является основным ферментом распада гликогена. В течение следующих 8–12 часов глюкоза, полученная из гликогена печени, является основным источником глюкозы в крови, используемой остальным телом в качестве топлива.
Глюкагон , еще один гормон, вырабатываемый поджелудочной железой, во многих отношениях служит контрсигналом для инсулина. В ответ на то, что уровень инсулина ниже нормы (когда уровень глюкозы в крови начинает падать ниже нормального диапазона), глюкагон секретируется в увеличивающихся количествах и стимулирует как гликогенолиз (расщепление гликогена), так и глюконеогенез (производство глюкозы из других источников). .
Мышцы
Мышечные клетки гликоген , как представляется , функции в качестве непосредственного резервного источника доступных глюкоз для мышечных клеток. Другие ячейки, содержащие небольшие количества, также используют его локально. Поскольку в мышечных клетках отсутствует глюкозо-6-фосфатаза , которая необходима для передачи глюкозы в кровь, гликоген, который они хранят, доступен исключительно для внутреннего использования и не передается другим клеткам. Это контрастирует с клетками печени, которые при необходимости легко расщепляют накопленный гликоген на глюкозу и отправляют ее через кровоток в качестве топлива для других органов.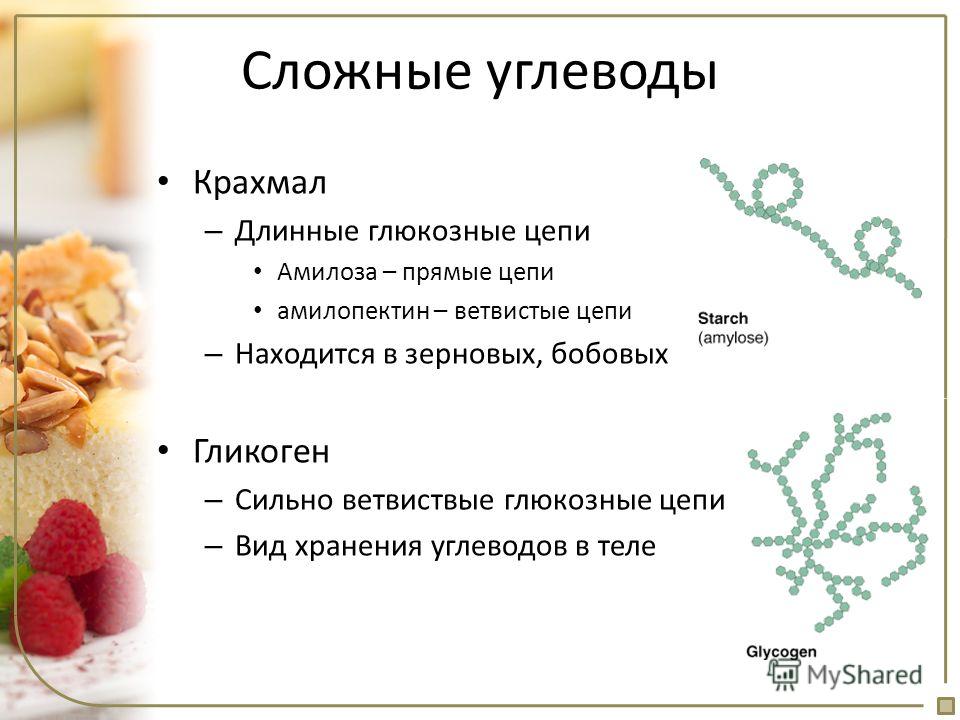
История
Гликоген был открыт Клодом Бернаром . Его эксперименты показали, что в печени содержится вещество, которое может приводить к снижению уровня сахара за счет действия «фермента» в печени. К 1857 году он описал выделение вещества, которое он назвал « la matière glycogène », или «сахарообразующее вещество». Вскоре после открытия гликогена в печени А. Сансон обнаружил, что мышечная ткань также содержит гликоген. Эмпирическая формула гликогена ( C
6 ЧАС
10 О
5 ) n была основана Кекуле в 1858 году.
Метаболизм
Синтез
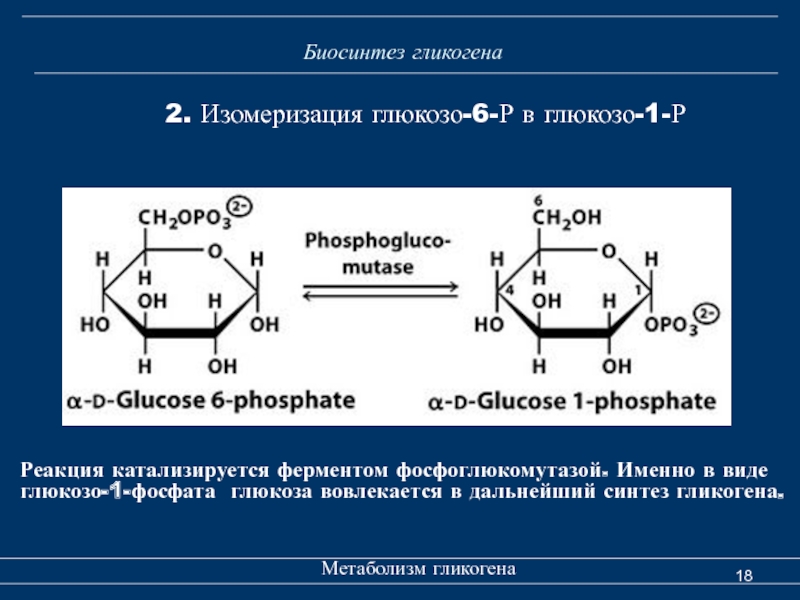
Синтез гликогена, в отличие от его расщепления, является эндергоническим — он требует затрат энергии. Энергия для синтеза гликогена поступает от уридинтрифосфата (UTP), который реагирует с глюкозо-1-фосфатом , образуя UDP-глюкозу , в реакции, катализируемой UTP — глюкозо-1-фосфатуридилтрансферазой . Гликоген синтезируется из мономеров UDP-глюкозы первоначально белком гликогенином , который имеет два тирозиновых якоря для восстанавливающего конца гликогена, поскольку гликогенин является гомодимером. После добавления примерно восьми молекул глюкозы к остатку тирозина фермент гликогенсинтаза постепенно удлиняет цепь гликогена с помощью UDP-глюкозы, добавляя α (1 → 4) -связанную глюкозу к восстанавливающему концу цепи гликогена.
Гликоген синтезируется из мономеров UDP-глюкозы первоначально белком гликогенином , который имеет два тирозиновых якоря для восстанавливающего конца гликогена, поскольку гликогенин является гомодимером. После добавления примерно восьми молекул глюкозы к остатку тирозина фермент гликогенсинтаза постепенно удлиняет цепь гликогена с помощью UDP-глюкозы, добавляя α (1 → 4) -связанную глюкозу к восстанавливающему концу цепи гликогена.
Гликоген ветвления фермента катализирует перенос концевого фрагмента шесть или семь остатков глюкозы из невосстанавливающего конца к С-6 гидроксильной группы остатка глюкозы глубже внутрь молекулы гликогена. Фермент разветвления может действовать только на ответвление, имеющее по крайней мере 11 остатков, и фермент может переноситься на ту же самую цепь глюкозы или соседние цепи глюкозы.
Авария
Гликоген отщепляется от невосстанавливающих концов цепи ферментом гликогенфосфорилазой с образованием мономеров глюкозо-1 фосфата:
В естественных условиях, фосфорилаза протекает в направлении распада гликогена , поскольку отношение фосфата и глюкозо-1-фосфата, как правило , больше , чем 100. Глюкозы-1 фосфат затем преобразуется в глюкозо-6-фосфат (G6P) по фосфоглюкомутазам . Для удаления α (1-6) разветвлений разветвленного гликогена и преобразования цепи в линейный полимер необходим специальный разветвляющий фермент . Произведенные мономеры G6P имеют три возможных судьбы:
Глюкозы-1 фосфат затем преобразуется в глюкозо-6-фосфат (G6P) по фосфоглюкомутазам . Для удаления α (1-6) разветвлений разветвленного гликогена и преобразования цепи в линейный полимер необходим специальный разветвляющий фермент . Произведенные мономеры G6P имеют три возможных судьбы:
Клиническая значимость
Нарушения обмена гликогена
Наиболее частым заболеванием, при котором метаболизм гликогена становится ненормальным, является диабет , при котором из-за ненормального количества инсулина гликоген в печени может ненормально накапливаться или истощаться. Восстановление нормального метаболизма глюкозы обычно также нормализует метаболизм гликогена.
При гипогликемии, вызванной чрезмерным количеством инсулина, уровни гликогена в печени высоки, но высокие уровни инсулина препятствуют гликогенолизу, необходимому для поддержания нормального уровня сахара в крови. Глюкагон — распространенное средство для лечения этого типа гипогликемии.
Различные врожденные нарушения метаболизма вызваны недостатком ферментов, необходимых для синтеза или распада гликогена. Все это называется болезнями накопления гликогена .
Истощение гликогена и упражнения на выносливость
Спортсмены на длинные дистанции, такие как марафонцы , лыжники и велосипедисты , часто испытывают истощение гликогена, когда почти все запасы гликогена у спортсмена истощаются после длительных периодов нагрузки без достаточного потребления углеводов. Это явление называется « удар о стену ».
Истощение запасов гликогена можно предотвратить тремя способами:
- Во-первых, во время тренировки постоянно потребляются углеводы с максимально возможной скоростью преобразования в глюкозу крови (высокий гликемический индекс ). Наилучший возможный результат этой стратегии — замена примерно 35% глюкозы, потребляемой при ЧСС выше примерно 80% от максимальной.
- Во-вторых, за счет адаптации тренировок на выносливость и специальных режимов (например, голодание, тренировки на выносливость с низкой интенсивностью) организм может кондиционировать мышечные волокна типа I, чтобы улучшить как эффективность использования топлива, так и нагрузочную способность, чтобы увеличить процент жирных кислот, используемых в качестве топлива, экономя углеводы.
 использовать из всех источников.
использовать из всех источников. - В-третьих, потребляя большое количество углеводов после истощения запасов гликогена в результате физических упражнений или диеты, организм может увеличить емкость внутримышечных запасов гликогена. Этот процесс известен как углеводная загрузка . В общем, гликемический индекс источника углеводов не имеет значения, так как мышечная чувствительность к инсулину увеличивается в результате временного истощения гликогена.
Испытывая дефицит гликогена, спортсмены часто испытывают сильную усталость до такой степени, что им становится трудно двигаться. Для справки, самые лучшие профессиональные велосипедисты в мире обычно заканчивают 4–5- часовой этап гонки на пределе истощения гликогена, используя первые три стратегии.
Когда спортсмены потребляют углеводы и кофеин после изнурительных упражнений, их запасы гликогена, как правило, пополняются быстрее; однако минимальная доза кофеина, при которой наблюдается клинически значимый эффект на восполнение запасов гликогена, не установлена.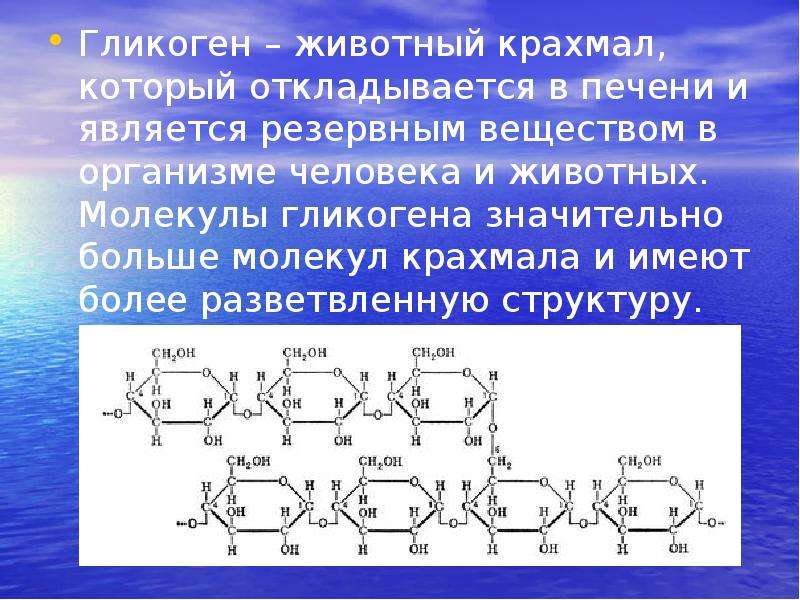
Смотрите также
Рекомендации
внешняя ссылка
Detecting Glycogen in Peripheral Blood Mononuclear Cells with Periodic Acid Schiff Staining
Критические шаги этого видео статьи были во время стирки и амилазы обработки клеток. При промывке слайды, ключевым шагом было с помощью пластиковой податливый стиральная бутылку и вода мягко запустить через образец на слайде и не стремимся непосредственно на образцах. Даже малейшее прямое давление воды может вызвать клетки оторваться от слайда. Другим ключевым шагом было использовать тот же слайд для ± условиях амилазы. После того, как РВМС прикреплен к слайд, слайд был осторожно помещают в химический стакан так что только половина мазок подвергается воздействию амилазы раствора. Этот шаг обеспечивает надежный контроль, потому что клетки крови от одной и той же слайде, таким образом минимизируя искажающих факторов, которые могут возникнуть в связи с небольшими вариациями синхронизации. Клетки застрял и без обработки, так что не будет оправданным дополнительные затраты и время связаны с поли-L-лизин или полиэтиленгликоль, например, покрытия.
Через TRoubleshooting оптимального деятельность амилазы определена. Для секций мышцы, было отмечено, что оптимальная активность амилазы наблюдалось в течение 1 ч инкубации. При более длительном разделы мышечные бы Медленно снимите слайд, в то время как более короткие интервалы времени амилазы не в полной мере удалить Па-сигнал. Время для амилазы инкубации в течение РВМС слайдов должно было быть уменьшена по сравнению с мышечной-образец синхронизации (который был 1 час) до всего лишь 15 мин. Более длительное время вызвало МНПК оторваться слайд, в то время как более короткие времена не эффективно воздействовать па-сигнал. Одна из модификаций со стандартным протоколом окрашивания PAS меняется метод промывки слайдов. Инструкции производителя указано мыть слайды с проточной водой, что вызывало клетки оторваться слайды. Модификация была мыть слайды с податливый стиральной бутылки, чтобы сохранить клетки. Как описано в шагах раздела критической, это было очень важно не непосредственно применять водуДавление на клетках.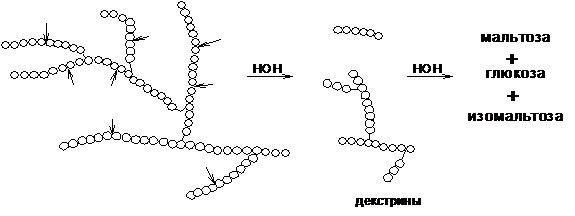
Есть некоторые ограничения в этой технике. Мышечных клеток мембрану окрашивают наряду с точечными гранул в цитоплазме мышечных клеток. Гранулы были устранены путем обработки амилазы и, следовательно, скорее всего, будет гликогена, в то время как окрашивание мембраны был нечувствителен к амилазы. Идентичность частиц PAS-положительных на клеточной мембране, не известно. Это может быть мышечная подвал (perimysium) мембрана. Эта мембрана окружает мышечные пучки и из-за высокого содержания гликопротеина известно, что ПАС положительным. Другим ограничением окрашивания PAS в том, что гранулы гликогена должно быть не менее 50 нм в диаметре, чтобы быть видимыми с помощью обычного светового микроскопа. Таким образом, мелкие гранулы гликогена может присутствовать в клетке, но все еще зарегистрировать отрицательный тест на PAS. Второй метод преодолевает это ограничение, обеспечивая обнаружение гликогена в лизате. В сочетании со стандартной кривой это может быть использовано, чтобы Determine точное количество гликогена. Хотя этот метод является очень количественно и не ограничивается размером гранул, она сама по себе имеет некоторые ограничения. Гликоген гранулы представляют собой сложные разветвленные структуры со многими вспомогательными белками и химических поперечных связей, что делает маловероятным, что гидролиз ферменты полного растворения всей молекулы гликогена 13. Таким образом, ферментативная обнаружения (например, техники PAS) может при-представляют собой реальную сумму гликогена в образце. Единственный способ справиться с это ограничение с электронной микроскопии, который решает даже самые маленькие гранулы гликогена 14. Можно было бы использовать комбинацию этих методов, PAS-окрашивание, ферментативного расщепления и электронной микроскопии для наиболее полной характеристики гликогена.
Хотя этот метод является очень количественно и не ограничивается размером гранул, она сама по себе имеет некоторые ограничения. Гликоген гранулы представляют собой сложные разветвленные структуры со многими вспомогательными белками и химических поперечных связей, что делает маловероятным, что гидролиз ферменты полного растворения всей молекулы гликогена 13. Таким образом, ферментативная обнаружения (например, техники PAS) может при-представляют собой реальную сумму гликогена в образце. Единственный способ справиться с это ограничение с электронной микроскопии, который решает даже самые маленькие гранулы гликогена 14. Можно было бы использовать комбинацию этих методов, PAS-окрашивание, ферментативного расщепления и электронной микроскопии для наиболее полной характеристики гликогена.
Эта статья и видео демонстрирует кислоты Шифф (PAS) метод периодического окрашивания, предназначенную для использования на МНПК. Значение этого исследования видно в выборе использованияМНПК более мазке крови, что сделало его более возможным перечислить лимфоцитов. Первоначально классический метод крови мазок был протестирован, однако большинство клеток на слайде были красные кровяные клетки и, возможно, нейтрофилы (рис 4в). Концентрировать лимфоциты, РВМС выделяли из крови с использованием стандартной техники в градиенте плотности. Используя методику, аналогичную кроваво-мазке, РВМС легко придерживаться на протяжении процедуре PAS, которая имеет множество активные стадии промывки. Техника PAS был использован на протяжении десятилетий для определения уровня гликогена в тканях биопсии мышц, которые разрезают на тонкие секции и присоединились к слайдам. Окрашивание PAS был выбран по сравнению с другими химическими веществами окрашивания углеводов из-за его высокой надежностью и наличием ожидаемых результатов в литературе. Мышь (Mus spretus) участки мышц был использован в качестве положительного контроля и обнаружили, как и ожидалось, что 37% клеток были PAS-положительных 9. С точки зрения стоимости и времени, готовясь PBMС более обременительным, чем мазке крови, но есть несколько преимуществ. Во-первых, препарат более обогащены лимфоцитов, которые являются клетки, представляющие интерес для проектов, которые сосредотачиваются на аутоиммунных заболеваний. Если были использованы мазки крови, лимфоциты будет меньшинство, что делает его сложно найти ячейку интерес. Можно было бы готовить и анализировать гораздо больше слайдов, чтобы получить те же самые номера вы получите с несколькими горками РВМС. Исследователи были бы очень заинтересованы в использовании наш новый оптимизированный метод в АУТОИММУНИТЕТ проектов, связанных с сахарным диабетом 1 типа, рассеянный склероз, волчанка, ревматоидный артрит, и т.д., где Т- и В-лимфоциты и NK-клетки играют роль. Конечно, для других применений, где эритроциты или нейтрофилов интерес мазка крови будет рекомендовано. Другим преимуществом является то, что РВМС могут быть использованы для изучения биологии лимфоцитов человека в лабораторных условиях.
Текущие исследования расследует источник гликогена в МНПК. Яп будущее, планируется для измерения содержания гликогена в контексте рассеянный склероз, Т-лимфоцитами аутоиммунные заболевания, поражающего насчитывалось 2,3 миллиона человек во всем мире в 2013 15,16. Каковы малые и большие клетки в образцах РВМС? Клетки в 5 мкм согласуются с лимфоидного в их состоянии покоя. Т-лимфоцитов, В-лимфоциты, природные клетки-киллеры и другие незначительные подмножества в пределах этого диапазона размера. Более крупные РВМС, вероятно, состоит из активированных лимфоцитов, которые растут больше, так как они получают воспалительные сигналы от иммунной системы, а также моноцитов с миелоидной линии. Передовые технологии сортировки клеток, например, люминесцентных активирована сортировки клеток или магнитного активирована сортировки клеток требуется для улучшения результативности подмножество, которое выражает гликогена.
Гематоксилин счетчика окрашивание используется, когда окрашивание было сделано на цельной крови. Это позволило нам выделить моноцитов эритроцитов (Фиг.4С). Когда окрашивание было сделано на РВМС, так как все клетки мононуклеарных, нет никакой необходимости, чтобы противостоять пятна гематоксилином выявления клеток. Также гематоксилин вмешивается с PAS сигнала в клетках. Таким образом, мы показали, процедуру PAS-окрашивание, приспособленный для РВМС. Этот метод полезен для ученых и клиницистов, исследующих АУТОИММУНИТЕТ, инфекции и аллергии.
Subscription Required. Please recommend JoVE to your librarian.
сеть ветеринарных лабораторий «Шанс Био»
Гликогеноз (Glycogen Storage Disease, GSD)
– группа аутосомно-рецессивных заболеваний, возникающих вследствие нарушения метаболизма гликогена.
Основным и наиболее универсальным источником энергии для человека, животных и растений является глюкоза. В организме животных глюкоза хранится в форме гликогена, который откладывается в виде гранул в цитоплазме клеток, преимущественно в клетках печени и мышц. При недостатке глюкозы гликоген расщепляется, и глюкоза попадает в кровь. Превращение гликогена в глюкозу – многоступенчатый процесс, протекающий под воздействием ряда ферментов, нарушение работы которых приводит к избыточному накоплению гликогена в виде аномальных гранул и нарушению гомеостаза глюкозы.
Клинические проявления гликогеноза бывают разными в зависимости от исходных ферментативных нарушений, в связи с чем выделяют разные типы заболевания. Номера типам присваивались в хронологическом порядке по мере их открытия.
Для собак известны следующие породоспецифичные типы: Ia тип описан для мальтийских болонок (дефицит глюкозо-6-фосфотазы, болезнь фон Гирке), II тип – для лапландской собаки (дефицит лизосомальной кислоты альфа-глюкозидазы, болезнь Помпе), III тип – для немецкой овчарки (дефицит гликоген-дебранчинг фермента, болезнь Кори) и VII тип – для английских спрингер-спаниелей (дефицит фосфофруктокиназы, PFK).
Недавно гликогеноз был описан для собак породы курчавошерстный ретривер (CCR) и классифицирован как тип IIIa. На первом году жизни GSD IIIa сложно детектируем и внешне почти никак не проявляется. С возрастом гликоген все больше и больше накапливается в клетках печени и мышц, начинают появляться непереносимость физических нагрузок, эпизодическая гипогликемия, вялость, миопатии. Люди, болеющие гликогенозом III типа, страдают от гипогликемии, возможно развитие скелетной миопатии, кардиомиопатии, цирроза печени, несмотря на это многие пациенты доживают до старости.Причина GSD IIIa у курчавошерстных ретриверов – мутация в гене AGL (amylo-alpha-1, 6-glucosidase, 4-alpha-glucanotransferase). Несмотря на сходство GSD IIIa CCR с болезнью Кори, показано, что для немецких овчарок заболевание имеет другую генетическую природу.
Диагностика
Для диагностики GSD IIIa курчавошерстных ретриверов разработан генетический тест. Исследование можно проводить в любом возрасте. При тестировании анализируется мутация в ДНК, приводящая к развитию заболевания. ДНК-тест позволяет определить дефектную (мутантную) копию гена и нормальную копию гена. Результат теста – это определение генотипа, которое позволяет разделить животных на три группы: здоровые (гомозиготы по нормальной копии гена), носители (гетерозиготы) и больные (гомозиготы по мутации).
Код 844
Genomia: Тестирование кошек: GSD IV
Описание:
Гликогеноз (Glycogen Storage Disease, GSD) – группа аутосомно-рецессивных заболеваний, возникающих вследствие нарушения метаболизма гликогена.
Основным и наиболее универсальным источником энергии для человека, животных и растений является глюкоза. В организме животных глюкоза хранится в форме гликогена, который откладывается в виде гранул в цитоплазме клеток, преимущественно в клетках печени и мышц. При недостатке глюкозы гликоген расщепляется, и глюкоза попадает в кровь. Превращение гликогена в глюкозу – многоступенчатый процесс, протекающий под воздействием ряда ферментов, нарушение работы которых приводит к избыточному накоплению гликогена в виде аномальных гранул и нарушению гомеостаза глюкозы.
Больные котята погибают от гипогликемии вскоре после рождения. В некоторых случаях животное живет до полугода, страдая при этом от прогрессирующей мышечной дегенерации.
Наследственность:
Мутация передается
аутосомно-рецессивным наследованием.
Заболевание проявляется у особей,
которые получили мутированный ген от
обоих родителей. Данные особи обозначаются
как P/P (мутированный гомозигот). Носители
мутированного гена, обозначаемые как
N/P (гетерозигот), получили мутированный
ген лишь от одного из родителей,
клинические признаки заболевания у них
отсутствуют; однако носители передают
заболевание своим потомкам. Теоретически
в результате спаривания двух гетерозигот
(N/P) 25% потомства будут здоровыми, 50%
будут носителями, а 25 % потомства
унаследуют от своих родителей мутированные
гены и будут страдать данным генетическим
заболеванием.
Тестируемая мутация: p.Y34X в гене GBE1
Образец:
кровь в EDTA (1,0 мл) или мазок из ротовой
полости. Подробная информация об отборе
образцов приведена здесь
Общая информация о генетическом
тесте:
Генетический тест позволяет
обнаружить больную особ или носителя
мутации. Тест может быть выполнен в
любом возрасте и действителен на
протяжении всей жизни. Генетический
тест методом полимеразной цепной реакции
(PCR) является очень точным, результаты
анализа позволяют определить больных
животных, здоровых носителей мутации
и здоровых животных.
С учетом наличия мутаций
собаки делятся на три группы:
-
P/P = позитивный / позитивный
= больной, особь унаследовала мутацию
от обоих родителей „affected“ -
N/P = негативный / позитивный
= особь унаследовала мутацию от одного
родителя, является носителем мутации
„Carrier“, болезнь у него не проявится -
N/N = негативный / негативный
= особь без мутаций, болезнь у особи не
проявится = нормальный генотип „wildtype“
Результаты спаривания
особей с различными генотипами:
|
|
Affected (P/P)
|
||
|
Wild type (N/N)
|
аллель
|
P
|
P
|
|
N
|
N/P (carrier)
|
N/P (carrier)
|
|
|
N
|
N/P (carrier)
|
N/P (carrier)
|
|
Все
потомки являются носителями мутации
.
|
|
Carrier (N/P)
|
||
|
Wild type (N/N)
|
аллель
|
N
|
P
|
|
N
|
N/N (здоровый)
|
N/P (carrier)
|
|
|
N
|
N/N (здоровый)
|
N/P (carrier)
|
|
Статистически
50 % потомков будут носителями, а 50 % будут здоровыми.
.
|
|
Carrier (N/P)
|
||
|
Carrier (N/P)
|
аллель
|
N
|
P
|
|
N
|
N/N (здоровый)
|
N/P (carrier)
|
|
|
P
|
N/P (carrier)
|
P/P (больной)
|
|
Статистически
25 % потомства будут здоровы, 25 % больны, а 50 % будут носителями.
.
Литература:
John C. Fyfe, Rebeccah L. Kurzhals, Michelle G. Hawkins, Ping Wang,
Naoya Yuhki, Urs Giger, Thomas J. Van Winkle, Mark E. Haskins, Donald F.
Patterson, Paula S. Henthorn: A complex rearrangement in GBE1 causes
both perinatal hypoglycemic collapse and late-juvenile-onset
neuromuscular degeneration in glycogen storage disease type IV of
Norwegian forest cats, Molecular Genetics and Metabolism 90 (2007)
383-392
Гликоген – определение, структура, функции и примеры
Определение гликогена
Гликоген большой, разветвленный полисахарид это основная форма хранения глюкозы у животных и человека. Гликоген является важным энергетическим резервуаром; когда организму требуется энергия, гликоген расщепляется до глюкозы, которая затем попадает в гликолитический или пентозофосфатный путь или выделяется в кровоток. Гликоген также является важной формой хранения глюкозы в грибы а также бактерии.
Структура гликогена
Гликоген является разветвленным полимером глюкозы. Глюкозные остатки линейно связаны α-1,4-гликозидными связями, и приблизительно каждые десять остатков цепь остатков глюкозы разветвляется через α-1,6-гликозидные связи. Α-гликозидные связи приводят к спиральной структуре полимера. Гликоген гидратируется тремя-четырьмя частями воды и образует гранулы в цитоплазма это 10-40 нм в диаметре. Белок гликогенин, который участвует в синтезе гликогена, находится в ядре каждой гранулы гликогена. Гликоген является аналогом крахмала, который является основной формой хранения глюкозы в большинстве растений, но крахмал имеет меньше ветвей и менее компактен, чем гликоген.
Функция гликогена
У животных и людей гликоген находится в основном в мускул а также печень клетки. Гликоген синтезируется из глюкозы, когда кровь уровень глюкозы высокий и служит готовым источником глюкозы для тканей всего тела, когда уровень глюкозы в крови снижается.
Клетки печени
Гликоген составляет 6-10% печени по массе. При приеме пищи уровень глюкозы в крови повышается, а выделение инсулина из поджелудочной железы способствует поглощению глюкозы клетками печени. Инсулин также активирует ферменты, участвующие в синтезе гликогена, такие как гликогенсинтаза. Хотя уровни глюкозы и инсулина достаточно высоки, гликогеновые цепи удлиняются путем добавления молекул глюкозы, процесс, называемый гликонеогенезом. По мере снижения уровня глюкозы и инсулина синтез гликогена прекращается. Когда уровень глюкозы в крови падает ниже определенного уровня, глюкагон, высвобождаемый из поджелудочной железы, сигнализирует клеткам печени о расщеплении гликогена. Гликоген расщепляется через гликогенолиз в глюкозо-1-фосфат, который превращается в глюкозу и выделяется в кровоток. Таким образом, гликоген служит в качестве основного буфера уровней глюкозы в крови, сохраняя глюкозу, когда его уровни высокие, и выделяя глюкозу, когда уровни низкие. Расщепление гликогена в печени имеет решающее значение для обеспечения глюкозы для удовлетворения энергетических потребностей организма. В дополнение к глюкагону, кортизол, адреналин и норадреналин также стимулируют расщепление гликогена.
Мышечные клетки
В отличие от клеток печени, гликоген составляет только 1-2% мышечной массы. Однако, учитывая большую массу мышц в организме, общее количество гликогена, хранящегося в мышцах, больше, чем запасенного в печени. Мышцы также отличаются от печени тем, что гликоген в мышцах обеспечивает глюкозу только мышечная клетка сам. Мышечные клетки не экспрессируют фермент глюкозо-6-фосфатазу, который необходим для высвобождения глюкозы в кровоток. Глюкозо-1-фосфат, образующийся в результате расщепления гли��огена в мышечных волокнах, превращается в глюкозо-6-фосфат и обеспечивает энергию для мышц во время тренировки или в ответ на стресс, как в реакции «сражайся или беги».
Другие ткани
Помимо печени и мышц, гликоген обнаруживается в меньших количествах в других тканях, включая эритроциты, лейкоциты, почечные клетки и некоторые глиальные клетки. Кроме того, гликоген используется для хранения глюкозы в матка обеспечить энергетические потребности эмбрион.
Грибы и бактерии
Микроорганизмы обладают механизмами накопления энергии, чтобы справиться с ними в случае ограниченных природных ресурсов, а гликоген представляет собой главную форму накопления энергии. Ограничение питательных веществ (низкий уровень углерода, фосфора, азота или серы) может стимулировать образование гликогена в дрожжах, в то время как бактерии синтезируют гликоген в ответ на легкодоступные источники энергии углерода с ограничением других питательных веществ. Рост бактерий и споруляция дрожжей также связаны с накоплением гликогена.
Гликоген гомеостаз это строго регулируемый процесс, который позволяет организму накапливать или выделять глюкозу в зависимости от его энергетических потребностей. Основными этапами метаболизма глюкозы являются гликогенез или синтез гликогена и гликогенолиз, или расщепление гликогена.
гликогенеза
Для синтеза гликогена требуется энергия, которую обеспечивает уридин трифосфат (UTP). Гексокиназы или глюкокиназы сначала фосфорилируют свободную глюкозу с образованием глюкозо-6-фосфата, который превращается в глюкозо-1-фосфат фосфоглюкомутазой. UTP-глюкозо-1-фосфат-уридилилтрансфераза затем катализирует активацию глюкозы, при которой UTP и глюкозо-1-фосфат реагируют с образованием UDP-глюкозы. При синтезе гликогена de novo белок гликогенин катализирует прикрепление UDP-глюкозы к себе. Гликогенин представляет собой гомодимер, содержащий остаток тирозина в каждой субъединице, который служит якорем или точкой присоединения для глюкозы. Дополнительные молекулы глюкозы впоследствии добавляются к восстанавливающему концу предыдущей глюкозы молекула сформировать цепь из примерно восьми молекул глюкозы. Затем гликогенсинтаза удлиняет цепь, добавляя глюкозу через α-1,4-гликозидные связи.
Разветвление катализируется амило- (от 1,4 до 1,6) -трансглюкозидазой, также называемой гликоген-разветвляющим ферментом. Гликоген-разветвляющий фермент переносит фрагмент из шести-семи молекул глюкозы от конца цепи к С6 молекулы глюкозы, расположенной далее внутри молекулы гликогена, образуя гликозидные связи α-1,6.
гликогенолиз
Глюкоза удаляется из гликогена с помощью гликогенфосфорилазы, которая фосфоролитически удаляет одну молекулу глюкозы с невосстанавливающего конца с образованием глюкозо-1-фосфата. Глюкозо-1-фосфат, образующийся при расщеплении гликогена, превращается в глюкозо-6-фосфат, процесс, который требует фермента фосфоглюкомутазы. Фосфоглюкомутаза переносит фосфатная группа из фосфорилированного остатка серина в пределах активный сайт до С6 глюкозо-1-фосфата с образованием глюкозо-1,6-бисфосфата. Затем фосфат глюкозы С1 присоединяют к активный сайт серин в фосфоглюкомутазе и глюкозо-6-фосфат высвобождается.
Гликогенфосфорилаза не способна отщеплять глюкозу от точек ветвления; для разветвления требуется амило-1,6-глюкозидаза, 4-α-глюканотрансфераза или гликогендеразрушающий фермент (GDE), который обладает глюкотрансферазной и глюкозидазной активностями. Примерно в четырех остатках от точки ветвления гликогенфосфорилаза не способна удалять остатки глюкозы. GDE расщепляет последние три остатка ветви и присоединяет их к С4 молекулы глюкозы в конце другой ветви, а затем удаляет последний α-1,6-связанный глюкозный остаток из точки ветвления. GDE не удаляет α-1,6-связанную глюкозу из точки ветвления фосфорилически, что означает, что высвобождается свободная глюкоза. Эта свободная глюкоза теоретически может высвобождаться из мышц в кровоток без действия глюкозо-6-фосфатазы; однако эта свободная глюкоза быстро фосфорилируется гексокиназой, предотвращая ее попадание в кровоток.
Глюкозо-6-фосфат, полученный в результате расщепления гликогена, может превращаться в глюкозу под действием глюкозо-6-фосфатазы и высвобождаться в кровоток. Это происходит в печени, кишечнике и почках, но не в мышцах, где этот фермент отсутствует. В мышцах глюкозо-6-фосфат входит в гликолитический путь и обеспечивает энергию для клетка, Глюкоза-6-фосфат также может проникать в пентозофосфатный путь, что приводит к выработке NADPH и пять углеродных сахаров.
Упражнения и истощение гликогена
В упражнениях на выносливость спортсмены могут испытывать истощение гликогена, при котором большая часть гликогена истощается из мышц. Это может привести к сильной усталости и затруднению движения. Истощение гликогена может быть уменьшено путем непрерывного потребления углеводов с высоким гликемическим индексом (высокая скорость превращения в глюкозу крови) во время физических упражнений, которые заменит часть глюкозы, используемой во время физических упражнений. Могут также использоваться специализированные режимы упражнений, которые приводят в действие мышцу жирные кислоты как источник энергии с большей скоростью, тем самым разрушая меньше гликогена. Спортсмены могут также использовать углеводную загрузку, потребление большого количества углеводов, чтобы увеличить емкость для хранения гликогена.
Примеры болезней накопления гликогена
Существуют две основные категории заболеваний, связанных с накоплением гликогена: те, которые возникают в результате нарушения гомеостаза гликогена в печени, и те, которые возникают в результате нарушения гомеостаза гликогена в мышцах. Заболевания, возникающие в результате неправильного хранения гликогена в печени, обычно вызывают гепатомегалию (увеличение печени), гипогликемию и цирроз печени (рубцевание печени). Заболевания, возникающие из-за дефектного накопления гликогена в мышцах, обычно вызывают миопатии и нарушение обмена веществ. Примеры заболеваний накопления гликогена включают болезнь Помпе, болезнь Макардла и болезнь Андерсена.
Болезнь Помпе
Болезнь Помпе вызвана мутациями в GAA ген, который кодирует лизосомальную кислотную α-глюкозидазу, также называемую кислой мальтазой, и влияет на скелет и сердечная мышца, Кислотная мальтаза участвует в расщеплении гликогена, а вызывающие заболевания мутации приводят к пагубному накоплению гликогена в клетке. Существует три типа болезни Помпе: взрослая форма, ювенильная форма и инфантильная форма, которые становятся все более тяжелыми. Инфантильная форма приводит к смерти в возрасте от одного до двух лет, если ее не лечить.
Болезнь Макардла
Болезнь Макардла вызвана мутациями в гене PYGM, который кодирует миофосфорилазу, изоформу гликогенфосфорилазы, присутствующую в мышцах. Симптомы часто наблюдаются у детей, но болезнь не может быть диагностирована до зрелого возраста. Симптомы включают мышечную боль и усталость, и если болезнь не лечится, болезнь может быть опасной для жизни.
Болезнь Андерсена
Болезнь Андерсена вызвана мутация в гене GBE1, который кодирует гликоген, разветвляющий фермент, и влияет на мышцы и печень. Симптомы обычно наблюдаются в возрасте нескольких месяцев и включают в себя задержку роста, увеличение печени и цирроз печени. Осложнения заболевания могут быть опасными для жизни.
викторина
1. Что лучше всего описывает функцию гликогена?A. Обеспечивает структурную поддержку мышечных клетокB. фактор транскрипции который регулирует дифференцировка клеток C. Хранит глюкозу в растенияхD. Буферы уровня глюкозы в крови и служит легко мобилизованным источником энергии
Ответ на вопрос № 1
D верно. Гликоген является основной формой хранения глюкозы у животных и человека. Гликоген синтезируется при высоком уровне глюкозы в крови и расщепляется при низком уровне глюкозы в крови, что делает его важным буфером уровня глюкозы в крови. Когда энергия требуется клеткой или организм Гликоген служит критическим источником энергии, обеспечивая глюкозу тканями по всему организму.
2. Что является основным гормон что стимулирует распад гликогена?A. глюкагонB. Щитовидная железаC. инсулинD. эстроген
Ответ на вопрос № 2
верно. Глюкагон, который вырабатывается в ответ на низкий уровень сахара в крови, стимулирует расщепление гликогена. Инсулин, вырабатываемый в ответ на высокий уровень сахара в крови, стимулирует поглощение глюкозы и синтез гликогена.
3. Каковы возможные судьбы глюкозо-1-фосфата, образующегося при гликогенолизе?A. Превращение в глюкозо-6-фосфат с последующим вступлением в гликолитический путьB. Превращение в глюкозо-6-фосфат с последующим вступлением в пентозофосфатный путьC. Преобразование в глюкозу с последующим выделением в кровотокD. Все вышеперечисленное
Ответ на вопрос № 3
D верно. В мышечных клетках глюкозо-1-фосфат превращается в глюкозо-6-фосфат с помощью фосфоглюкомутазы, после чего он может вступать в гликолитический или пентозофосфатный путь. В клетках печени глюкозо-6-фосфат превращается в глюкозу глюкозо-6-фосфатазой и выделяется в кровоток.
Ссылки
- Eicke, S., Seung, D., Egli, B., Devers, E.A., и Streb, S. (2017) «Повышение способности растений хранить углеводы путем создания пула, подобного гликогеноподобному полимеру, в цитозоль «. Метаболическая инженерия. 40: 23-32.
- Харгривз М. и Рихтер Е.А. (1988) «Регулирование скелетная мышца гликогенолиз во время тренировки ». Канадский журнал спортивных наук. 13 (4): 197-203.
- Ivy, J.L. (1991). «Синтез мышечного гликогена до и после тренировки». Спортивная медицина. 11 (1): 6-19.
Почему животные используют гликоген для хранения полисахаридов, а растения используют крахмал?
Резюме
Ключевое различие между гликогеном и амилопектином (основным компонентом крахмала) заключается не в количестве 1,6-гликозидных ветвей, а в их расположении .
В гликогене ветви последовательно подразделяются, образуя относительно небольшую глобулярную структуру, которая не способна расти дальше. Он растворим в водной среде и с его многочисленными открытыми концами может быстро метаболизироваться — подходит для клеток животных, в которых необходимо мобилизовать запасы энергии в ответ на насущные потребности, например, для сокращения мышц.
В амилопектине имеется длинная центральная полисахаридная цепь, от которой ветки ограниченного размера простираются с интервалами. В результате образуются гораздо большие полукристаллические частицы (крахмальные зерна), форма, особенно подходящая для длительного хранения в больших количествах в семенах и клубнях.
Химия
Это общая черта гликогена и амилопектиновой части крахмала. (Часть амилозы неразветвленная.) В гликогене содержится ок. одна точка ветвления на 10 единиц глюкозы, тогда как у амилопектина эта цифра равна 1 на 24–30 (источник: Wikipedia ).
Топография
Контрастная ветвящаяся топография двух полисахаридов, упомянутых выше, схематически показана ниже:
Это двумерное представление. В трех измерениях гликоген распространяется во всех направлениях от центральной точки — на самом деле праймерный фермент, гликогенин . В трех измерениях нити амилопектина в основном лежат рядом.
Макроструктура
Иллюстрация ниже, измененная от Bell et al. , показывает различные формы и размеры макромолекулярных структур. Следует отметить, что полукристаллической природе амилопектина способствует спиральная конформация цепей.
Вместо того, чтобы предоставить информацию о обзоре Bell et al. (Журнал «Экспериментальная ботаника», том 62, стр. 1775–1801, 2011). Я приведу их непосредственно (не упоминая их цитаты).
Что касается гликогена, они пишут:
Каждая цепь, за исключением внешних неразветвленных цепей, поддерживает две ветви. Эта схема ветвления обеспечивает сферический рост уровней, генерирующих частицы (уровень соответствует сферическому пространству, отделяющему две последовательные ветви от всех цепей, расположенных на одинаковом расстоянии от центра частицы). Этот тип роста приводит к увеличению плотности цепей на каждом уровне, что приводит к прогрессивно более тесной структуре к периферии.
Математическое моделирование предсказывает максимальное значение для размера частиц, выше которого дальнейший рост невозможен, поскольку не было бы достаточно места для взаимодействия цепей с каталитическими центрами ферментов метаболизма гликогена. В результате образуется частица, состоящая из 12 ярусов, соответствующих максимальному диаметру 42 нм, включая 55 000 остатков глюкозы. 36% от этого общего количества находится в наружной (неразветвленной) оболочке и, таким образом, легко доступен для катаболизма гликогена без разветвления. Таким образом, in vivo частицы гликогена присутствуют в форме гранул предельного размера (макрогликоген), а также гранул меньшего размера, представляющих промежуточные состояния биосинтеза и деградации гликогена (прогликоген). Частицы гликогена полностью растворимы в воде и, следовательно, определяют состояние, при котором глюкоза становится менее активной осмотически, но в то же время легко доступной для быстрой мобилизации через ферменты катаболизма гликогена, как если бы она находилась в растворимой фазе.
Относительно амилопектина они пишут:
Амилопектин определяет один из, если не самый большой, известный биологический полимер и содержит от 105 до 106 остатков глюкозы. Не существует теоретического верхнего предела размера, достигаемого отдельными молекулами амилопектина. Это не связано с несколько меньшей степенью общего разветвления молекулы по сравнению с гликогеном. Скорее это связано с тем, как ветви распределяются внутри структуры. Ветви сконцентрированы в участках молекулы амилопектина, приводящих к скоплениям цепей, которые обеспечивают неограниченный рост полисахарида. Другая важная особенность кластерной структуры амилопектина состоит в плотной упаковке цепей, образующихся в корне кластеров, где плотность ветвей локально достигает или превышает плотность гликогена. Эта плотная упаковка ветвей генерирует плотно упакованные глюкановые цепи, которые достаточно близки для выравнивания и образования параллельных двойных спиральных структур. Спирали внутри одного кластера и соседних кластеров выравнивают и образуют участки кристаллических структур, разделенных участками аморфного материала (содержащего ветви), тем самым генерируя полукристаллическую природу амилопектина и последующей гранулы крахмала. Действительно, кристаллизованные цепи становятся нерастворимыми и обычно разрушаются в макрогранулированное твердое вещество. Эта осмотически инертная гранула крахмала позволяет хранить неограниченное количество глюкозы, которая становится метаболически недоступной. В самом деле, ферменты синтеза и мобилизации крахмала не могут напрямую взаимодействовать с твердой структурой, за исключением заметного гранулярно-связанной крахмалсинтазы, единственного фермента, необходимого для синтеза амилозы.
кода
Недостаток информации о метаболизме растительного крахмала, по-видимому, отражает комбинацию того, что они менее известны о биохимии растений и менее интересны из-за общего внимания к медицинской биохимии и биохимии животных. Хотя сам биохимик по животным (и, таким образом, ранее неосведомленный об информации в этом ответе) я чувствую, что пришло время исправить этот дисбаланс.
цитоплазматические гранулы гликогена — это… Что такое цитоплазматические гранулы гликогена?
- цитоплазматические гранулы гликогена
-
Medicine: alpha units
Универсальный русско-английский словарь.
Академик.ру.
2011.
- цитоплазматические глыбки гликогена
- цитоплазматические тельца-включения
Смотреть что такое «цитоплазматические гранулы гликогена» в других словарях:
-
лейкоциты — ов; мн. (ед. лейкоцит, а; м.) [от греч. leukos белый и kytos клетка] Физиол. Бесцветные клетки крови человека и животных. * * * лейкоциты (от лейко… и …цит), бесцветные клетки крови человека и животных. Все типы лейкоцитов (лимфоциты,… … Энциклопедический словарь
-
Патологическая анатомия слюнных желёз — Сиалоаденопатии различные патологические изменения, развивающиеся в слюнных железах. Содержание 1 Пороки развития 1.1 Аномалии размера и аномалады жаберных дуг … Википедия
-
ГОРМОНЫ — органические соединения, вырабатываемые определенными клетками и предназначенные для управления функциями организма, их регуляции и координации. У высших животных есть две регуляторных системы, с помощью которых организм приспосабливается к… … Энциклопедия Кольера
-
ГИСТОЛОГИЯ — наука, занимающаяся изучением тканей животных. Тканью называют группу клеток, сходных по форме, размерам и функциям и по продуктам своей жизнедеятельности. У всех растений и животных, за исключением самых примитивных, тело состоит из тканей,… … Энциклопедия Кольера
-
Строение и химический состав бактериальной клетки — Общая схема строения бактериальной клетки показана на рисунке 2. Внутренняя организация бактериальной клетки сложна. Каждая систематическая группа микроорганизмов имеет свои специфические особенности строения. Клеточная стенка.… … Биологическая энциклопедия
-
Кожа — I Кожа (cutis) сложный орган, являющийся наружным покровом тела животных и человека, выполняющий разнообразные физиологические функции. АНАТОМИЯ И ГИСТОЛОГИЯ У человека площадь поверхности К. равна 1,5 2 м2 (в зависимости от роста, пола,… … Медицинская энциклопедия
(PDF) Динамическая жизнь гранулы гликогена
E. I. (1999) Фрактальная структура гликогена: умное решение для оптимизации клеточного метаболизма.
Biophys. J. 77, 1327–1332
31. Мелендес, Р., Мелендес-Хевиа, Э., Мас, Ф., Мах, Дж., Касканте, М., Р., М., Мелендес-Хевиа, Э. ,
F., M., Mach, J. и Cascnte, M. (1998) Физические ограничения в синтезе гликогена, которые влияют на его структурную однородность
: двумерный подход.Биофиз. J. 75, 106–114
32. Baqué, S., Guinovart, J. J., and Ferrer, J. C. (1997) Гликогенин, праймер для синтеза гликогена,
связывается с актином. FEBS Lett. 417, 355–359
33. Cid, E., Cifuentes, D., Baqué, S., Ferrer, J.C. и Guinovart, J.J. (2005) Детерминанты ядерно-цитоплазматического перемещения мышечной гликогенсинтазы
. FEBS J. 272, 3197–213
34. Wilson, WA, Boyer, MP, Davis, KD, Burke, M., and Roach, PJ (2010) Субклеточная локализация дрожжевой гликогенсинтазы
зависит от содержания гликогена. .Может. J. Microbiol.
56, 408–20
35. Ou, H., Yan, L., Osmanovic, S., Greenberg, CC, and Brady, MJ (2005) Пространственная реорганизация
гликоген-синтазы при активации в 3T3-L1 Адипоциты. Эндокринология. 146, 494–502
36. Фернандес-Новелл, Дж. М., Беллидо, Д., Виларо, С. и Гуиноварт, Дж. Дж. (1997). Глюкоза индуцирует транслокацию
гликогенсинтазы в кору клеток гепатоцитов крыс. Biochem. J. 321, 227–31
37.Prats, C., Cadefau, JA, Cussó, R., Qvortrup, K., Nielsen, JN, Wojtaszewski, JFP,
Wojtaszewki, JFP, Hardie, DG, Stewart, G., Hansen, BF, and Ploug, T. . (2005)
Зависимая от фосфорилирования транслокация гликогенсинтазы в новую структуру во время
ресинтеза гликогена. J. Biol. Chem. 280, 23165–72
38. Prats, C., Helge, JW, Nordby, P., Qvortrup, K., Ploug, T., Dela, F., and Wojtaszewski, JFP
(2009) Двойное регулирование мышечная гликогенсинтаза во время упражнений путем активации и
компартментализации.J. Biol. Chem. 284, 15692–700
39. Маршан, И., Чернейко, К., Тарнопольски, М., Гамильтон, С., Ширер, Дж., Потвин, Дж., И Грэм,
TE (2002). субклеточный гликоген в мышцах человека в состоянии покоя: размер гранул, количество,
и расположение. J. Appl. Physiol. 93, 1598–1607
40. Элснер П., Кисторфф Б., Хансен Г. Х. и Граннет Н. (2002) Частично упорядоченный синтез и
деградация гликогена в культивируемых мышечных трубках крыс. Дж.Биол. Chem. 277, 4831–4838
41. Девос П. и Херс Х. Г. (1979) Молекулярный порядок в синтезе и разложении гликогена
в печени. Евро. J. Biochem. 99, 161–7
42. Oe, Y., Baba, O., Ashida, H., Nakamura, KC, and Hirase, H. (2016) Распределение гликогена в головном мозге мышей
, фиксированных с помощью микроволн, выявляет гетерогенные астроциты. узоры. Глия. 64, 1532–45
43. Накамура-Цурута, С., Ясуда, М., Накамура, Т., Шинода, Э., Фуруясики, Т., Kakutani, R.,
Takata, H., Kato, Y., and Ashida, H. (2012) Сравнительный анализ связывания углеводов
гостем 11 мая 2020 г. http://www.jbc.org/ Скачал с
Glycogen | SpringerLink
- Margit Pavelka
- Jürgen Roth
Abstract
Глюкоза является важным источником энергии, а гликоген — его клеточная форма хранения, которой больше всего в печени и мышцах. Гликоген находится в цитоплазме в виде гранул диаметром от 10 до 40 нм, так называемых β-частиц, типичных для мышечных клеток.В гепатоцитах β-частицы собираются в характерные розетки гликогена — α-частицы (стрелки). Α-частицы состоят не только из гликогена, но, кроме того, содержат различные ферментные белки, участвующие в синтезе гликогена, отсюда и название гликосомы. Во время синтеза гликогена гликогенин, который инициирует синтез, и гликогенсинтаза, удлиняющая цепь глюкозы, образуют комплекс с глюкозой.
Ключевые слова
Заболевание накопления гликогена Метаболизм гликогена Заболевание накопления гликогена Тип цепи глюкозы Нормальная концентрация глюкозы в крови
Эти ключевые слова были добавлены машиной, а не авторами.Это экспериментальный процесс, и ключевые слова могут обновляться по мере улучшения алгоритма обучения.
Ссылки
-
Burchell A и Waddell ID (1991) Молекулярные основы микросомальной глюкозо-6-фосфатазной системы печени. Biochim Biophys Acta 1092: 129
CrossRefPubMedGoogle Scholar
-
Рыбицка К.К. (1996) Гликосомы — органеллы метаболизма гликогена. Tissue Cell 28: 253
CrossRefPubMedGoogle Scholar
Информация об авторских правах
© Springer-Verlag / Wien 2010
Авторы и аффилированные лица
- Маргит Павелка
- Юрген Рот
- 1.Центр анатомии и клеточной биологии, Департамент клеточной биологии и исследований ультраструктуры, Венский медицинский университет, Вена, Австрия,
- 2. Аспирантура Университета Йонсей, Университетская программа мирового класса, Департамент биомедицинских наук поток аутофагии
- 1.
Браун, AM Мозговый гликоген пробудился. J. Neurochem. 89 , 537–552 (2004).
CAS
Статья
PubMedGoogle ученый
- 2.
Vilchez, D. et al. Механизм подавления синтеза гликогена в нейронах и его гибель при прогрессирующей миоклонической эпилепсии. Nat. Neurosci. 10 , 1407–1413 (2007).
CAS
Статья
PubMedGoogle ученый
- 3.
Триведи, Дж. Р. и др. Полиглюкозановая болезнь тела взрослых, связанная с непристойными телами и тремором. Arch. Neurol. 60 , 764–766 (2003).
Артикул
PubMedGoogle ученый
- 4.
Иноуэ, М., Ягишита, С., Ито, Ю., Амано, Н. и Мацусита, М. Сосуществование парных спиральных нитей и полиглюкозановых тел в одном нейроне при вскрытии болезни Альцгеймера. Acta Neuropathol. 92 , 511–514 (1996).
CAS
Статья
PubMedGoogle ученый
- 5.
Робитайл, Ю., Карпентер, С., Карпати, Г. и Ди Мауро, С. Д. Отдельная форма заболевания тела полиглюкозана у взрослых с массовым поражением центральных и периферических нейрональных процессов и астроцитов: отчет о четырех случаях и обзор возникновения полиглюкозановых тел при других состояниях, таких как болезнь Лафора и нормальное старение. Мозг 103 , 315–336 (1980).
CAS
Статья
PubMedGoogle ученый
- 6.
Saez, I. et al. Нейроны обладают активным метаболизмом гликогена, что способствует устойчивости к гипоксии. J. Cereb. Кровоток. Метаб. 34 , 945–955 (2014).
CAS
Статья
PubMed
PubMed CentralGoogle ученый
- 7.
Wang, Y. et al. Лафорин предотвращает вызванное стрессом образование полиглюкозановых тел и прогрессирование болезни Лафора в нейронах. Mol. Neurobiol. 48 , 49–61 (2013).
CAS
Статья
PubMed
PubMed CentralGoogle ученый
- 8.
Duran, J. et al. Вредные эффекты накопления гликогена в нейронах у мух и мышей. EMBO Mol. Мед 4 , 719–729 (2012).
CAS
Статья
PubMed
PubMed CentralGoogle ученый
- 9.
Миттал С. и Ганеш С. Механизмы контроля качества белка и нейродегенеративные расстройства: сдержки, противовесы и тупики. Neurosci. Res 68 , 159–166 (2010).
CAS
Статья
PubMedGoogle ученый
- 10.
Yerbury, J. J. et al. По канату: протеостаз и нейродегенеративные заболевания. J. Neurochem. 137 , 489–505 (2016).
CAS
Статья
PubMedGoogle ученый
- 11.
Дэвис, Дж. Б. и Махер, П.Активация протеинкиназы C подавляет цитотоксичность, вызванную глутаматом, в линии нейрональных клеток. Brain Res. 652 , 169–173 (1994).
CAS
Статья
PubMedGoogle ученый
- 12.
Кумар А. и Ратан Р. Р. Окислительный стресс и болезнь Хантингтона: хорошее, плохое и уродливое. Дж. Хантингт. Дис. 5 , 217–237 (2016).
Артикул
Google ученый
- 13.
Kumar, A. et al. Снижение О-связанного GlcNA-цилирования защищает от цитотоксичности, опосредованной фрагментом белка экзона 1 хантинтина. J. Biol. Chem. 289 , 13543–13553 (2014).
CAS
Статья
PubMed
PubMed CentralGoogle ученый
- 14.
Garyali, P. et al. Комплекс малин-лафорин подавляет клеточную токсичность неправильно свернутых белков, способствуя их деградации через систему убиквитин-протеасома. Hum. Мол. Genet. 18 , 688–700 (2009).
CAS
Статья
PubMedGoogle ученый
- 15.
Баба О. [Производство моноклональных антител, распознающих гликоген, и его применение в иммуногистохимии]. Kokubyo Gakkai Zasshi 60 , 264–287 (1993).
CAS
Статья
PubMedGoogle ученый
- 16.
Goswami, A. et al. Окислительный стресс способствует агрегации мутантного хантинтина и зависимой от мутантного хантингтина гибели клеток, имитируя протеасомные нарушения. Biochem Biophys. Res. Commun. 342 , 184–190 (2006).
CAS
Статья
PubMedGoogle ученый
- 17.
Mangiarini, L. et al. Экзона 1 гена HD с увеличенным повторением CAG достаточно, чтобы вызвать прогрессирующий неврологический фенотип у трансгенных мышей. Cell 87 , 493–506 (1996).
CAS
Статья
PubMedGoogle ученый
- 18.
Carter, R.J. et al. Характеристика прогрессирующих моторных дефицитов у мышей, трансгенных по мутации болезни Гентингтона человека. J. Neurosci. 19 , 3248–3257 (1999).
CAS
PubMedGoogle ученый
- 19.
Сингх П.K., Singh, S. & Ganesh, S. Комплекс лафорин-малин отрицательно регулирует синтез гликогена, модулируя поглощение глюкозы клетками через переносчики глюкозы. Mol. Cell Biol. 32 , 652–663 (2012).
CAS
Статья
PubMed
PubMed CentralGoogle ученый
- 20.
Юнг, К. Ю. и Рампал, А. Л. Сайты связывания цитохалазина B и переносчик глюкозы в призраках эритроцитов человека. Дж.Биол. Chem. 252 , 5456–5463 (1977).
CAS
PubMedGoogle ученый
- 21.
Мора, А., Сакамото, К., МакМанус, Э. Дж. И Алесси, Д. Р. Роль пути PDK1-PKB-GSK3 в регулировании гликогенсинтазы и захвата глюкозы сердцем. FEBS Lett. 579 , 3632–3638 (2005).
CAS
Статья
PubMedGoogle ученый
- 22.
Гринберг, К. С., Мередит, К. Н., Ян, Л. и Брэди, М. Дж. Нацеливание белка на сверхэкспрессию гликогена приводит к специфическому усилению накопления гликогена в адипоцитах 3T3-L1. J. Biol. Chem. 278 , 30835–30842 (2003).
CAS
Статья
PubMedGoogle ученый
- 23.
Klionsky, D. J. et al. Рекомендации по использованию и интерпретации анализов для мониторинга аутофагии. Аутофагия 8 , 445–544 (2012).
CAS
Статья
PubMed
PubMed CentralGoogle ученый
- 24.
Танида И. и Вагури С. Измерение аутофагии в клетках и тканях. Methods Mol. Биол. 648 , 193–214 (2010).
CAS
Статья
PubMedGoogle ученый
- 25.
Ginet, V. et al. Умирающие нейроны таламуса доношенных новорожденных и крыс являются аутофагами. Ann. Neurol. 76 , 695–711 (2014).
CAS
Статья
PubMedGoogle ученый
- 26.
Ginet, V. et al. Участие аутофагии в гипоксически-эксайтотоксической гибели нейронов. Аутофагия 10 , 846–860 (2014).
CAS
Статья
PubMed
PubMed CentralGoogle ученый
- 27.
Ginet, V., Puyal, J., Кларк, П. Г. и Труттманн, А. С. Усиление аутофагического потока после неонатальной церебральной гипоксии-ишемии и его регионально-зависимая связь с механизмами апоптоза. Am. J. Pathol. 175 , 1962–1974 (2009).
CAS
Статья
PubMed
PubMed CentralGoogle ученый
- 28.
Чжан З., Сингх Р. и Ашнер М. Методы обнаружения аутофагии в клетках млекопитающих. Curr. Protoc.Toxicol. 69 , 20 12 21–20 12 26 (2016).
Google ученый
- 29.
Чен С. и Ветцель Р. Солюбилизация и дезагрегация полиглутаминовых пептидов. Protein Sci. 10 , 887–891 (2001).
CAS
Статья
PubMed
PubMed CentralGoogle ученый
- 30.
Чен, С., Бертелье, В., Гамильтон, Дж. Б., О’Нуаллен, Б.& Ветцель, Р. Амилоидоподобные особенности агрегатов полиглутамина и их кинетика сборки. Биохимия 41 , 7391–7399 (2002).
CAS
Статья
PubMedGoogle ученый
- 31.
Chen, S., Berthelier, V., Yang, W. & Wetzel, R. Поведение агрегации полиглутамина in vitro поддерживает механизм набора цитотоксичности. J. Mol. Биол. 311 , 173–182 (2001).
CAS
Статья
PubMedGoogle ученый
- 32.
Джаяраман, М., Такур, А. К., Кар, К., Кодали, Р. и Ветцель, Р. Анализы для изучения ядерной агрегации полиглутаминовых белков. Методы 53 , 246–254 (2011).
CAS
Статья
PubMed
PubMed CentralGoogle ученый
- 33.
Scherzinger, E. et al. Самосборка полиглутаминсодержащих фрагментов хантингтина в амилоидоподобные фибриллы: последствия для патологии болезни Хантингтона. Proc. Natl. Акад. Sci. США 96 , 4604–4609 (1999).
CAS
Статья
PubMed
PubMed CentralGoogle ученый
- 34.
Harish, G. et al. Метаболизм глутатиона регулируется посмертным интервалом, гендерными различиями и агональным состоянием в посмертном человеческом мозге. Neurochem. Int. 59 , 1029–1042 (2011).
CAS
Статья
PubMedGoogle ученый
- 35.
Li, I.H. et al. Активация аутофагии участвует в нейротоксичности, вызванной 3,4-метилендиоксиметамфетамином («экстази») в культивируемых корковых нейронах. PLoS ONE 9 , e116565 (2014).
Артикул
PubMed
PubMed CentralGoogle ученый
- 36.
Хиггинс, Г. К., Девениш, Р. Дж., Беарт, П. М. и Нагли, П. Аутофагическая активность корковых нейронов в условиях острого окислительного стресса напрямую способствует гибели клеток. Cell Mol. Life Sci. 68 , 3725–3740 (2011).
CAS
Статья
PubMedGoogle ученый
- 37.
Shi, R. et al. Чрезмерная аутофагия способствует гибели нейронов при церебральной ишемии. CNS Neurosci. Ther. 18 , 250–260 (2012).
CAS
Статья
PubMedGoogle ученый
- 38.
Рами, А. и Когель, Д.Апоптоз встречается с гибелью клеток, подобной аутофагии, в ишемической полутени: две стороны одной медали? Аутофагия 4 , 422–426 (2008).
CAS
Статья
PubMedGoogle ученый
- 39.
Котулас, О. Б., Каламидас, С. А., Кондомеркос, Д. Дж. Аутофагия гликогена. Microsc. Res Tech. 64 , 10–20 (2004).
CAS
Статья
PubMedGoogle ученый
- 40.
Сингх П. К., Сингх С. и Ганеш С. Активация сывороточной / глюкокортикоид-индуцированной киназы 1 (SGK1) лежит в основе повышенных уровней гликогена, активации mTOR и дефектов аутофагии при болезни Лафора. Mol. Биол. Ячейка 24 , 3776–3786 (2013).
Артикул
PubMed
PubMed CentralGoogle ученый
- 41.
DeBosch, B.J. et al. Трегалоза подавляет белки-носители растворенного вещества 2A (SLC2A), вызывая аутофагию и предотвращая стеатоз печени. Sci. Сигнал. 9 , ra21 (2016).
Артикул
PubMed
PubMed CentralGoogle ученый
- 42.
Zirin, J., Nieuwenhuis, J. & Perrimon, N. Роль аутофагии в распаде гликогена и ее отношение к хлорохиновой миопатии. PLoS Biol. 11 , e1001708 (2013).
Артикул
PubMed
PubMed CentralGoogle ученый
- 43.
Halse, R., Fryer, L.G., McCormack, J.G., Carling, D. & Yeaman, S.J. Регулирование гликогенсинтазы глюкозой и гликогеном: возможная роль AMP-активированной протеинкиназы. Диабет 52 , 9–15 (2003).
CAS
Статья
PubMedGoogle ученый
- 44.
Харди Д. Г. AMPK: ключевой регулятор энергетического баланса в отдельной клетке и во всем организме. Int J. Obes. (Лондон) 32 , S7 – S12 (2008).
CAS
СтатьяGoogle ученый
- 45.
Ха, Дж., Гуан, К. Л. и Ким, Дж. AMPK и аутофагия в метаболизме глюкозы / гликогена. Mol. Asp. Мед 46 , 46–62 (2015).
CAS
СтатьяGoogle ученый
- 46.
Duran, J., Gruart, A., García-Rocha, M., Delgado-García, J. M. & Guinovart, J. J. Накопление гликогена лежит в основе нейродегенерации и нарушения аутофагии при болезни Лафора. Hum. Мол. Genet 23 , 3147–3156 (2014).
CAS
Статья
PubMedGoogle ученый
- 47.
Puri, R., Suzuki, T., Yamakawa, K. & Ganesh, S. Дисфункции эндосомно-лизосомных путей и аутофагии лежат в основе невропатологии на мышиной модели болезни Лафоры. Hum. Мол. Genet 21 , 175–184 (2012).
Артикул
PubMedGoogle ученый
- 48.
Aguado, C. et al. Лафорин, самый распространенный белок, мутировавший при болезни Лафора, регулирует аутофагию. Hum. Мол. Genet. 19 , 2867–2876 (2010).
CAS
Статья
PubMed
PubMed CentralGoogle ученый
- 49.
Puri, R., Jain, N. & Ganesh, S. Повышенная концентрация глюкозы приводит к снижению протеасомной активности и образованию гликоген-положительных агресомных структур. FEBS J. 278 , 3688–3698 (2011).
CAS
Статья
PubMedGoogle ученый
- 50.
Миттал, С. и Сингх, Л. Р. Макромолекулярное скопление замедляет агрегацию бета-богатого белка, бычьей карбоангидразы: тематическое исследование. J. Biochem. 156 , 273–282 (2014).
CAS
Статья
PubMedGoogle ученый
- 51.
Перуц, М. Ф., Джонсон, Т., Сузуки, М. и Финч, Дж. Т. Глютамин повторы как полярные застежки-молнии: их возможная роль в наследственных нейродегенеративных заболеваниях. Proc. Natl. Акад. Sci. США 91 , 5355–5358 (1994).
CAS
Статья
PubMed
PubMed CentralGoogle ученый
- 52.
Groen, A.C., Coughlin, M. & Mitchison, T.J. Сборка микротрубочек в мейотическом экстракте требует гликогена. Mol.Биол. Ячейка 22 , 3139–3151 (2011).
CAS
Статья
PubMed
PubMed CentralGoogle ученый
- 53.
Waelter, S. et al. Накопление мутантных фрагментов хантингтина в агресомоподобных телец включения в результате недостаточной деградации белка. Mol. Биол. Ячейка 12 , 1393–1407 (2001).
CAS
Статья
PubMed
PubMed CentralGoogle ученый
- 54.
Джонстон, Дж. А., Уорд, К. Л. и Копито, Р. Р. Агресомы: клеточный ответ на неправильно свернутые белки. J. Cell Biol. 143 , 1883–1898 (1998).
CAS
Статья
PubMed
PubMed CentralGoogle ученый
- 55.
Хартл, П., Олсон, Э., Данг, Т. и Форбс, Д. Дж. Сборка ядер с лямбда-ДНК во фракционированных экстрактах яиц Xenopus: неожиданная роль гликогена в образовании промежуточного хроматина более высокого порядка. J. Cell Biol. 124 , 235–248 (1994).
CAS
Статья
PubMedGoogle ученый
- 56.
Патель, М. Ориентация на окислительный стресс при расстройствах центральной нервной системы. Trends Pharmacol. Sci. 37 , 768–778 (2016).
CAS
Статья
PubMed
PubMed CentralGoogle ученый
- 57.
Siwach, P., Сенгупта, С., Парихар, Р. и Ганеш, С. Пространственные положения гомополимерных повторов в протеоме человека и их влияние на клеточную токсичность. Biochem Biophys. Res Commun. 380 , 382–386 (2009).
CAS
Статья
PubMedGoogle ученый
- 58.
Goenka, A. et al. Некодирующие РНК сателлита-III человека модулируют репрессию транскрипции, вызванную тепловым шоком. J. Cell Sci. 129 , 3541–3552 (2016).
CAS
Статья
PubMedGoogle ученый
- 59.
Upadhyay, M., Bhadauriya, P. & Ganesh, S. Тепловой шок модулирует субклеточную локализацию, стабильность и активность HIPK2. Biochem. Биофиз. Res Commun. 472 , 580–584 (2016).
CAS
Статья
PubMedGoogle ученый
- 60.
Siwach, P., Sengupta, S., Parihar, R.& Ganesh, S. Пролиновые повторы в цис- и транс-положениях обеспечивают защиту от токсичности неправильно свернутых белков в клеточной модели млекопитающих. Neurosci. Res 70 , 435–441 (2011).
CAS
Статья
PubMedGoogle ученый
- 61.
Ganesh, S. et al. Целенаправленное нарушение гена Epm2a вызывает образование телец включения Lafora, нейродегенерацию, атаксию, миоклоническую эпилепсию и нарушение поведенческой реакции у мышей. Hum. Мол. Genet 11 , 1251–1262 (2002).
CAS
Статья
PubMedGoogle ученый
- 62.
Maheshwari, M. et al. Дексаметазон вызывает реакцию теплового шока и замедляет прогрессирование болезни у мышей и мух, моделирующих болезнь Хантингтона. Hum. Мол. Genet 23 , 2737–2751 (2014).
CAS
Статья
PubMedGoogle ученый
- 63.
Шанкар, С. К., Махадеван, А., Хариш, Г. и Шринивас Бхарат, М. М. Репозиторий ткани человеческого мозга: Национальное учреждение, содействующее исследованиям в области нейробиологии. Proc. Natl. Акад. Sci., India Sect. В: Биол. Sci. 84 , 239–250 (2014).
CAS
СтатьяGoogle ученый
- 64.
Джоши, А. С. и Такур, А. К. Биоразлагаемая система доставки, содержащая пептидный ингибитор агрегации полиглутамина: шаг на пути к терапевтическим разработкам при болезни Хантингтона. J. Pept. Sci. 20 , 630–639 (2014).
CAS
Статья
PubMedGoogle ученый
- 2N; главный исследователь, Химадри Пакраси) и, частично, стипендией Билсленда Дэвида Г. Велки, предоставленной Департаментом биологических наук Университета Пердью.
Мы благодарим Дебру М. Шерман из DS Imaging, LLC за электронные микрофотографии.
СНОСКИ
- Получено 21 октября 2015 г.
- Принято 30 ноября 2015 г.
- Принятая рукопись размещена в Интернете 14 декабря 2015 г.
- Адресная переписка Луи А. Шерману, lsherman {at} purdue.edu.
-
↵ * Текущий адрес: Дэвид Г. Велки, Центр циркадной биологии, Отделение биологических наук, Калифорнийский университет в Сан-Диего, Ла-Хойя, Калифорния, США.
-
Citation Велки Д.Г., Ли Б.Х., Шерман Л.А. 2016. Изменение структуры гранул хранения углеводов у цианобактерий Synechocystis sp.штамм PCC 6803 посредством усечений фермента ветвления. J Bacteriol 198: 701–710. DOI: 10.1128 / JB.00830-15.
-
Дополнительные материалы к этой статье можно найти по адресу http://dx.doi.org/10.1128/JB.00830-15.
- Авторские права © Американское общество микробиологии, 2016 г. Все права защищены.
Производство нерастворимых крахмалоподобных гранул в Escherichia coli путем модификации пути синтеза гликогена
Abstract
При исследовании превращения целлюлозной биомассы в крахмалоподобные материалы для промышленного использования было обнаружено, что сверхэкспрессия нативного АДФ -глюкозопирофосфорилаза GlgC в Escherichia coli привела к образованию нерастворимых гранул полисахарида в цитоплазме, занимающих большую часть объема клетки, а также к общему увеличению содержания полисахаридов в клетках.Электронная микроскопия показала, что гранулы не имели ламеллярной структуры крахмала, а имели неправильную кластерную структуру. При голодании клетки, сверхэкспрессирующие GlgC, оказались неспособными полностью разложить свой полисахаридный материал, и гранулы все еще были четко видны в культурах после 8 дней голодания. Интересно, что дополнительная сверхэкспрессия фермента ветвления GlgB устраняет образование гранул и приводит к дальнейшему увеличению клеточных полисахаридов. Обычно считается, что GlgC отвечает за лимитирующую стадию синтеза гликогена.Наша интерпретация этих результатов заключается в том, что избыточная активность GlgC может вызывать удлинение цепей гликогена, опережающее добавление боковых ответвлений, позволяя цепям соседних молекул гликогена достигать длины, на которой они спонтанно переплетаются, образуя плотные кластеры, которые в значительной степени недоступны для хозяин. Однако при дополнительной активации фермента ветвления GlgB разветвление полисахарида способно поддерживать скорость синтеза линейных цепей, устраняя фенотип гранул.Это исследование предлагает потенциальные возможности для увеличения производства и восстановления бактериального полисахарида.
Значение В этой работе запасы полисахаридов Escherichia coli были изменены путем добавления дополнительных копий собственных генов синтеза полисахаридов бактерий. Таким образом были созданы бактерии, которые продуцировали более чем вдвое больший уровень запасающего полисахарида, чем контрольный штамм, в форме гранул, которые потенциально могут облегчить сбор урожая.Была создана другая форма мутанта Escherichia coli , которая продуцировала более чем в семь раз превышающий нормальный уровень запасающего полисахарида, а также росла до более высоких плотностей клеток в жидкой культуре. В дополнение к расширению нашего понимания синтеза гликогена предполагается, что аналогично модифицированные бактерии, выращенные на недорогих отходах, могут быть полезным источником крахмалоподобных полисахаридов для промышленного или сельскохозяйственного использования. В частности, в последнее время вызывает интерес использование цианобактериального гликогена в качестве источника углерода для биотоплива, и представленная здесь работа может быть применима в этой области.
Введение
Гранулы крахмала, накапливаемые растительными клетками, составляют основу значительной части продуктов питания человека и являются ценным ресурсом во многих отраслях промышленности, включая производство бумаги и биотоплива. В последние годы огромные объемы зерна направляются на производство биоэтанола; По имеющимся данным, в 2013-2014 гг. 40% урожая кукурузы в США использовалось для производства биотоплива (1). По некоторым данным, это было одним из факторов роста цен на продукты питания во многих частях мира в 2008 году (2, 3).Кроме того, было высказано предположение, что расчистка земель, необходимая для выращивания сельскохозяйственных культур, используемых в качестве биотоплива, может произвести больше выбросов CO 2 , чем сэкономлено за счет сокращения использования ископаемого топлива (4, 5).
Так или иначе, но очевидно, что по мере роста населения человечества будет становиться все более неприемлемым переключение пищевых материалов, пригодных для людей, на непродовольственные цели. Таким образом, требуется источник крахмалоподобных полимеров на непищевой основе. Одним из текущих планов по сокращению утечки пищевых материалов может быть постепенный переход на биотопливо «второго поколения», такое как этанол, производимый непосредственно из обильных целлюлозных отходов генетически модифицированными штаммами Saccharomyces cerevisiae , Escherichia coli или другими. такие организмы, как Geobacillus thermoglucosidasius (6–11).Несмотря на затраты огромных ресурсов, значительные государственные субсидии и другие стимулы, производство целлюлозного этанола в настоящее время остается в довольно небольших масштабах. Одной, по-видимому, неизученной альтернативой может быть преобразование сахаров из целлюлозного материала в легко разлагаемую крахмалоподобную форму, которая может быть включена в текущую, очень крупномасштабную систему производства этанола на основе зерна. Такой двухстадийный процесс может иметь значительные преимущества с точки зрения кинетики процесса, поскольку преобразование целлюлозной биомассы в сахара неизбежно является «медленным» этапом по сравнению с ферментацией сахаров в этанол.Начальное медленное превращение целлюлозных полимеров в нерастворимый, но легко разлагаемый крахмалоподобный материал, который можно легко извлечь из ферментационной среды, промыть, транспортировать и хранить для последующего использования с последующим быстрым осахариванием и ферментацией с использованием стандартных высокоэффективных процессов. может изменить экономику производства целлюлозного этанола. Кроме того, микробные крахмалоподобные полимеры также могут быть полезны в других отраслях промышленности и даже могут частично заменить использование зерна в кормлении домашнего скота, тем самым высвобождая больше зерна для употребления в пищу людьми.
По этим причинам производство нерастворимых крахмалоподобных полимеров было исследовано на бактериях. В целом производство настоящего крахмала ограничивается растениями, зелеными и красными водорослями, хотя в настоящее время он также идентифицирован по крайней мере у одного штамма диазотрофных цианобактерий подгруппы V (12, 13). Обычные бактерии, такие как Escherichia coli , производят гликоген, который имеет структуру, аналогичную амилопектиновой фракции крахмала, то есть α-1,4-связанные цепи глюкозы с α-1,6-связанными боковыми ответвлениями, но с более высокий уровень ветвления около 9%.Уровень разветвления амилопектина ниже 6% и происходит прерывисто, ярусами, тем самым создавая чередующиеся аморфные и кристаллические ламеллы (14, 15). Кристаллические ламели, вероятно, состоят из неразветвленных глюкановых цепей длиной от 12 до 20 глюкозильных остатков, которые спонтанно соединяются вместе, образуя двойные спирали со своими соседями, в то время как аморфные ламели считаются повторяющимися областями молекулы, которые содержат точки ветвления. и как таковые не могут образовывать спирали из-за интерференции α-1,6-связей.Предполагается также, что двойные спирали кристаллической ламеллы закручиваются в суперпрепараты, хотя детали этого еще предстоит выяснить (16–18).
Фракция амилозы обычно составляет около 15% кристаллов крахмала в Chloroplastida и считается вкрапленной в аморфную пластинку амилопектина, хотя ее точное местоположение все еще обсуждается (19, 20). Он намного проще, чем амилопектин, поскольку в основном представляет собой неразветвленные цепи α-1,4 связанных глюкозильных остатков.Он также намного меньше, с оценками молекулярной массы, варьирующимися от 10 5 до 10 6 Дальтон (18). Из-за редкости ответвлений амилоза скручивается в отдельные спирали, причем каждый виток спирали, как считается, состоит из 6 остатков глюкозила (21).
Йод часто используется в колориметрическом анализе крахмала, и это отдельные спирали фракции амилозы, которые реагируют с образованием интенсивного сине-черного цвета, типичного для этого анализа. Результирующий цвет зависит от длины комплекса спираль-йод, от коричневого до красного и до синего с увеличением длины (20, 22, 23).Амилопектин также окрашивается раствором йода, хотя с гораздо более низким сродством (<1% по сравнению с примерно 20% с амилозой), поскольку йод не образует комплексов с двойными спиралями кристаллической ламеллы. Поэтому комплекс амилопектин-йод имеет тенденцию быть очень бледным, с красновато-пурпурным цветом. Между тем, гликоген обычно образует комплекс темно-красного цвета с йодом, что свидетельствует об изобилии более коротких одиночных спиралей, хотя это значительно варьируется между разными источниками из-за более низкой структурной организации гликогена (24).
Из-за высокого уровня разветвления гликогена предполагается, что его гранулы имеют ограниченный размер примерно до 42 нм в диаметре по стерическим соображениям и хорошо растворимы, тогда как гранулы крахмала при их многоуровневом росте могут иметь диаметр в сотни микрометров. и нерастворимы, обеспечивая более плотный накопитель энергии (25, 26). Основные цепи гликогена синтезируются гликоген-синтазой (GlgA), которая добавляет остатки сахара в форме АДФ-глюкозы, продуцируемой из глюкозо-1-фосфата и АТФ с помощью АДФ-глюкозопирофосфорилазы (GlgC).Поэтому активность GlgC обычно рассматривается как первая обязательная стадия, а также стадия, ограничивающая скорость синтеза гликогена (14). Между тем, α-1,6-связанные боковые цепи добавляются разветвляющим ферментом GlgB, который отщепляет около четырех остатков цепи на концевом конце и присоединяет их к 6-гидроксилу остатка, расположенного дальше по цепи. Сильно разветвленная структура гликогена обеспечивает оптимальный доступ к остаткам глюкозы для расщепляющих ферментов гликогенфосфорилазы (GlgP) и фермента, разветвляющего гликоген (GlgX) (27) (рисунок 1).
Рисунок 1:
Метаболизм гликогена в E. coli . GlgC и GlgA работают в тандеме, чтобы вырастить невосстанавливающие концы внешних цепей глюкана по одной единице глюкозы за раз, в то время как GlgP удаляет остатки глюкозы с невосстанавливающих концов цепей, пока его активность не будет прервана близостью к ветки, созданные GlgB. GlgX может разорвать связи α-1,6 на ответвлении этих ветвей, но только на ответвлениях короче тех, которые изначально были присоединены GlgB, тем самым предотвращая попадание ферментов в бесполезный цикл последовательных ветвлений и разветвлений (Cenci et al., 2014).
Чтобы изучить возможность превращения бактериального гликогена в нерастворимую крахмалоподобную форму для облегчения извлечения и обработки, это исследование первоначально было направлено на усиление выработки гликогена. В соответствии с идеей о том, что GlgC катализирует лимитирующую стадию синтеза гликогена, известно, что усиление выработки гликогена может быть достигнуто за счет сверхэкспрессии GlgC или, более конкретно, мутантной формы GlgC16 (G335D), которая является устойчивы к нормальному ингибированию АМФ по обратной связи (28).Однако при сверхэкспрессии GlgC в E. coli MG1655 JM109 мы наблюдали большие нерастворимые тельца внутри клеток, по-видимому, состоящие из полимеров глюкозы. Насколько нам известно, об этом явлении ранее не сообщалось, хотя синтез гликогена широко изучался. Здесь дается предварительная характеристика этих включений.
Результаты
Для создания клеток с более высоким содержанием полисахаридов, GlgC и GlgC16 были сверхэкспрессированы в Escherichia coli JM109 из плазмиды с высоким числом копий, pSB1C3, с использованием промотора lac (pJW-glgC и pJW- , соответственно).Такую же плазмиду с промотором lac и только lacZ ’(pJW-lacZ) использовали в качестве контроля (таблица 3). Было обнаружено, что клетки, несущие pJW-glgC или pJW-glgC16, выращенные в присутствии лактозы, содержат более высокие уровни полисахарида, чем контрольные, как ожидалось, и, соответственно, окрашивают йод в более темный цвет (рис. 2). Клетки обрабатывали CuSO 4 и H 2 O 2 для стабилизации изменения цвета йодом (29). Влияние этой обработки на интенсивность окрашивания реакций крахмал-йод исследовали, чтобы убедиться, что она не прерывает линейную тенденцию интенсивности окраски к содержанию полисахарида, и было обнаружено, что она не нарушает эту линейность.
Рисунок 2:
(A) общее содержание сахара гексозы в E. coli JM109 / pJW-lacZ по сравнению с E. coli JM109 / pJW-glgC и E. coli JM109 / pJW-glgC16, f rom антроновые анализы, проводимые на культурах, выращенных таким образом, чтобы максимально увеличить содержание в них полисахаридов. Планки погрешностей представляют собой стандартную ошибку среднего при n = 3. (B) Фазово-контрастная световая микроскопия, показывающая фенотип «телец включения» в окрашенных йодом клетках E. coli JM109 / pJW-glgC16 до (вверху) и после (внизу) лизиса КОН, с результатами анализа йода тех же культур, до и после лизиса с КОН. Культуры выращивали в течение ночи в LB с добавлением IPTG и 1% лактозы. Лизис достигается при 30% КОН и кипячении в течение 5 минут. Масштабная шкала соответствует 5 мкм. Столбец «i»: E. coli, JM109 / pJW-lacZ. Колонка «ii»: E. coli, JM109 / pJW-glgC.
Неожиданно было обнаружено, что клетки, трансформированные pJW-glgC или pJW-glgC16, также содержат большие включения, ясно видимые при фазово-контрастной микроскопии, которые отсутствовали в клетках, содержащих контрольную плазмиду. Эти включения были окрашены йодом в темный цвет на фоне неокрашенного тела клетки, выглядя ярко-синим при фазовом контрасте (рис. 2В) или коричневым при микроскопии светового поля (не показано).Также было замечено, что окрашивающий йод материал присутствует в клеточных остатках, а не в растворимой фракции (рис. 2В). Анализ SDS-PAGE не показал больших новых полос в цитоплазме или клеточных остатков по сравнению с контролем, что указывает на то, что включения не состояли из неправильно свернутого белка.
Было высказано предположение, что повышенное поступление АДФ-глюкозы, обеспечиваемое повышающей регуляцией glgC , позволило активности гликогенсинтазы достичь ранее нереализованного потенциала и опередить активность фермента ветвления.Для проверки этой гипотезы был сконструирован pJW-glgCB, содержащий glgC , за которым следует glgB , оба под контролем промотора lac (таблица 3). В соответствии с предсказанием, E. coli JM109 / pJW-glgCB продуцировали повышенные уровни полисахарида по сравнению с контролем, хотя неожиданно было обнаружено, что это повышение было еще более значительным, чем для клеток, трансформированных одним дополнительным glgC ( рисунок 3А). Также в соответствии с предсказаниями, клетки, трансформированные pJW-glgCB, окрашивали типичный красно-коричневый цвет гликогена в реакции с йодом (рис. 3, B и C) и не содержали видимых включений при световой микроскопии.Все культуры были проанализированы с помощью просвечивающей электронной микроскопии, где оба новых трансформанта показали заметный фенотип, отличный от контрольных (рис. 4). Было обнаружено, что включения, обнаруженные в клетках E. coli JM109 / pJW-glgC, имеют зернистую внутреннюю структуру. Было замечено, что клетки, трансформированные pJW-glgCB, содержат большое количество инклюзивного вещества, рассредоточенного по клеткам. В случае обоих трансформантов инклюзивное вещество наблюдалось примерно в трети клеток.
Рисунок 3:
(A) Общее содержание гексозного сахара в трансформированных клетках JM109. Анализы на антрон, выполненные на культурах, равных OD λ600 нм 3,5, после примерно 30-часового роста в среде Корнберга с последующим M9 таким образом, чтобы максимально увеличить содержание в них полисахаридов. Планки погрешностей представляют собой стандартную ошибку среднего при n = 3. (B и C) Анализ йода трансформированного JM109. B) пластыри из культур, выращенных до стационарной фазы в LB с добавлением лактозы и IPTG, нанесенные на чашку M9, выращенные в течение ночи при 37 ° C и залитые йодом Люголя (0.2%) .; C) Осадки клеток из 1,4 мл тех же культур, что и на изображении B, осажденные и ресуспендированные в 200 мкл PBS, плюс 50 мкл CuSO 4 (100 мМ), 50 мкл H 2 O 2 (6%) и 25 мкл йода Люголя (0,2%), перенесенные в лунки 48-луночного планшета и визуализированные с помощью планшетного сканера. i) E. coli JM109 / pJW-lacZ; ii) E. coli JM109 / pJW-glgC; iii) E. coli JM109 / pJW-glgB; iv) E. coli JM109 / pJW-glgCB.
Рисунок 4:
Электронные микрофотографии (ПЭМ) E.coli штамм JM109, трансформированный «контрольной» плазмидой pJW-lacZ (A, B), плазмидой pJW-glgC16 (C, D) или плазмидой pJW-glgCB (E, F). Многие из «контрольных» клеток показывают отрицательно окрашенное инклюзивное вещество. Однако считается, что определенные зернистые тела, видимые во многих трансформированных клетках pJW-glgC16 и pJW-glgCB, отличаются как от контроля, так и друг от друга, а в случае pJW-glgC16 соответствуют найденным телам. окрашивать йодом при просмотре под световым микроскопом.
Изображения выбраны из 14 захваченных просвечивающих электронных микрофотографий pJW-lacZ, 20 pJW-glgC16 и 25 pJW-glgCB и считаются репрезентативными.
Также было обнаружено, что культур E.coli JM109 / pJW-glgCB достигли более высокой плотности, чем таковые в контроле. Для дальнейшего исследования были получены кривые роста для различных трансформантов как в среде LB, так и в модифицированной среде Корнберга, замещенной лактозой и IPTG (рис. 5). E. coli JM109 / pJW-glgCB показала более медленный начальный рост, чем контроль, в течение первых пяти часов рост LB, но затем продолжал увеличивать плотность клеток и выходил на плато с более высокой плотностью, чем любой из других трансформантов.Клетки, трансформированные pJW-glgC, показали еще более медленную начальную скорость роста, достигая той же плотности, что и контроль, примерно через восемь часов роста в LB, и никогда не наблюдали превышения плотности клеток контроля в этой среде. Аналогичная картина наблюдалась в модифицированной среде Корнберга, хотя все культуры достигли более высокой конечной плотности, и различия между конечной плотностью культур были менее четкими. Клетки, трансформированные pJW-glgB, демонстрировали почти идентичную кривой роста контролю во всех обстоятельствах.
Рисунок 5:
Кривые роста для четырех трансформантов, выращенных в среде LB (A) или модифицированной среде Корнберга (B), дополненной лактозой и IPTG, в течение 24 часов. Серые линии показывают показания встряхиваемой колбы на 50 мл. Планки погрешностей представляют собой стандартную ошибку среднего при n = 3. Черные линии показывают показания 96-луночного планшета с 3 планшетов. Планки погрешностей представляют собой стандартную ошибку среднего при n = 6.
Впоследствии было проверено, доступны ли полисахариды клеткам в условиях голодания.Культуры всех трансформантов выращивали до одинаковой оптической плотности таким образом, чтобы максимально увеличить содержание в них полисахаридов, прежде чем клетки промывали и ресуспендировали в минимальной среде без добавления источника углерода. Показания плотности культур снимались с интервалами более 216 часов, в то время как антроновые анализы проводились на аликвотах культур в те же моменты времени, чтобы определить, сколько полисахарида, присутствующего в исходных осадках клеток, метаболизировалось с течением времени (рис. ).Все культуры показали снижение плотности культуры и содержания полисахаридов. В соответствии с ожиданиями, E. coli JM109 / pJW-glgC показали гораздо большее снижение плотности культуры, чем контроль, за это время, и никогда не наблюдалось полного истощения своих запасов полисахаридов. Однако наибольшее снижение плотности культуры наблюдалось у E. coli JM109 / pJW-glgCB. Кроме того, эти культуры также никогда не могли полностью истощить свои запасы полисахаридов, даже когда эксперимент повторяли в течение 264 часов, чтобы прояснить неоднозначные результаты (рисунок 6C).Во всех случаях снова наблюдали поведение E. coli JM109 / pJW-glgB, аналогичное контролю. Белковые анализы были выполнены на всех культурах для последних двух временных точек эксперимента по голоданию, и было замечено, что они подтверждают показания оптической плотности. Культуры также наблюдали под световой микроскопией после окрашивания йодом в каждый момент времени эксперимента с голоданием. Тельца включения наблюдались в аналогичном соотношении в клетках E. coli JM109 / pJW-glgC на протяжении обоих экспериментов (фигура 7).
Рисунок 6:
(A) Кривые роста для трансформированного JM109 в течение 216 часов в условиях голодания . Начиная с плотности клеток 1,5 OD λ600 нм . (B) Общее содержание сахара гексозы в культурах в ходе эксперимента по голоданию. (C) Повторение эксперимента по общему содержанию сахара в гексозе. Планки погрешностей представляют собой стандартную ошибку среднего значения при n = 3.
Рисунок 7:
Световая микроскопия окрашенного йодом, E. coli JM109 / pJW-glgC до и после голодания. Из культур E. coli JM109 / pJW-glgC, использованных в эксперименте по голоданию, изображения на день 1 (A) и день 8 (B), показывающие прозрачные тельца включения в обоих случаях. Масштабная полоса представляет 5 мкм (прибл.)
Обсуждение
Эта работа показала, что трансформация E. coli MG1655 JM109 дополнительными копиями его нативной glgC , экспрессированной на плазмиде с большим числом копий под lac промотор, приводил к образованию телец включения внутри клеток, когда они выращивались в культуре с добавлением лактозы и IPTG.Тельца включения также наблюдались у тех же трансформантов при выращивании в среде с добавлением глюкозы вместо лактозы, хотя и в меньшей степени, что, как полагают, связано с ингибирующим действием глюкозы на промотор lac (данные не показаны). . Тот же фенотип «тельца включения» не наблюдался в клетках, трансформированных той же плазмидой, но лишенных glgC , ни в одном из тестируемых условий роста. Тельца включения, если они присутствуют, окрашиваются в темно-коричневый цвет в результате реакции с йодом, что указывает на обилие одиночных спиралей, более коротких, чем те, которые обнаруживаются в гликогене, который имеет тенденцию окрашиваться в красный цвет.Анализ SDS-PAGE клеток E. coli JM109 / pJW-glgC16 показал, что наблюдаемые включения не состояли из одного сверхэкспрессированного белка.
Анализы антрона, проведенные на трансформантах в этой работе, последовательно показали более высокое содержание гексозного сахара для клеток, трансформированных glgC16 и glgC , чем для контроля. Было высказано предположение, что, поскольку активность GlgC обычно выполняет лимитирующую стадию синтеза гликогена, повышая его экспрессию в JM109 E.coli , выращенная в среде, богатой сахаром, приводит к появлению телец включения во многих клетках, а также к более высокой средней внутриклеточной концентрации сахара, чем в контрольной группе, выращенной в тех же условиях, поскольку GlgC увеличивает доступность субстрата для GlgA , гликогенсинтаза. Считается, что этот увеличенный субстрат позволяет гликогенсинтазе соответствовать ранее нереализованному потенциалу и синтезировать линейные цепи глюкана в грануле полисахарида с большей скоростью, чем GlgB может добавлять к ним ответвления, так что они вырастают длинные неразветвленные цепи, способные спонтанно образовывать двойные -спирали, и это переплетение соседних цепей вызывает агрегацию гранул в наблюдаемые большие тела включения.Эти двойные спирали не связывают йод. Однако, поскольку общее содержание полисахаридов также увеличивается примерно вдвое в результате этой манипуляции, мы предполагаем, что это также приводит к появлению большого количества коротких одиночных спиралей, дающих наблюдаемое темно-коричневое пятно. Гранулярная внутренняя структура, наблюдаемая во включениях при ПЭМ, которая отличается от ламеллярной структуры, наблюдаемой в гранулах крахмала, также поддерживает гипотезу о том, что они образованы кластерами гликогеноподобных полисахаридов.
Если эта гипотеза верна, считалось, что добавление glgB к той же плазмиде, что и glgC , позволило бы разветвлению полисахарида поддерживать скорость синтеза линейных цепей. Ожидалось, что это остановит образование телец включения, поскольку неразветвленные области глюкановых цепей больше не смогут расти достаточно долго, чтобы свернуться в двойные спирали, и, кроме того, стерическое вмешательство, вызванное плотной упаковкой ответвлений глюкана, что теоретически ограничивает рост гранул природного гликогена, будет повторно введен.Однако ожидалось, что клетки будут демонстрировать высокое содержание сахара, наблюдаемое у клеток, трансформированных glgC , поскольку гликогенсинтаза все еще будет способна синтезировать цепи с повышенной скоростью. Все эти фенотипы наблюдались с дополнительным наблюдением, что вместо того, чтобы синтезировать такие же высокие уровни полисахарида, как у клеток, трансформированных glgC , клетки, трансформированные как glgC , так и glgB , показали более чем удвоенное общее содержание сахара.
Кроме того, эта работа показала, что E. coli JM109 / pJW-glgCB вырастают до значительно более высокой плотности культуры, чем E. coli JM109 / pJW-lacZ, выращенные в тех же условиях. Однако, вопреки прогнозам, они также оказались гораздо более уязвимыми в условиях голодания, когда плотность культур упала до менее чем одной трети от контроля через девять дней (значения оптической плотности, которые привели к такому выводу, хорошо коррелируют с белком. анализы на одних и тех же культурах, предполагая, что они дают точное измерение биомассы, а не, например, являются артефактом повышенного светорассеяния, вызванного тельцами включения).Следовательно, избыточный запас сахара, возможно, является дополнительным стрессом для этих клеток в таких условиях. E. coli JM109 / pJW-glgC также были более уязвимы к условиям голодания, чем контроль E. coli JM109 / pJW-lacZ, а также показали самую низкую скорость роста среди всех трансформантов. Если действительно тельца включения, накопленные во время предшествующего роста в богатой углеродом среде, образованы из полисахаридов, похоже, что эти трансформанты были неспособны их переваривать. Наличие тел включения в пределах E.Считается, что трансформанты coli JM109 / pJW-glgC в начале и в конце периода голодания подтверждают эту гипотезу. Хотя необходимы дальнейшие испытания, эти результаты могут иметь значение для синтеза полисахаридов в промышленности, поскольку большие тельца включения, которые бактерии не могут переварить, должно быть легче собрать, чем гранулы нативного гликогена.
Эксперименты с голоданием дополнительно, по-видимому, предполагают, что E. coli JM109 / pJW-glgCB содержали сахара гексозы, которые он не мог переваривать, поскольку анализы антрона для этого трансформанта показали, что его общий уровень содержания сахара гексозы примерно такой же, как и у трансформанта. модель E.coli JM109 / pJW-glgC. Под микроскопом в этих клетках не наблюдаются тельца включения, хотя клетки показывают плотную концентрацию инклюзивного вещества под ТЕМ. Предполагается, что это гликоген. Непонятно, почему эти трансформанты не могут полностью переваривать содержащийся в них гексозный сахар.
В E. coli отсутствуют дикиназы (глюкан-водная дикиназа и фосфоглюкан-водная дикиназа), которые, как считается, способствуют разложению крахмала организмами, метаболизирующими крахмал, возможно, за счет извлечения α-1,4-связанных цепей глюкана из их двойной спирали. чтобы сделать их доступными для водорастворимых ферментов, таких как фосфорилаза и фермент разветвления (GlgP и GlgX, соответственно, в E.coli ) (30, 31). Следовательно, добавление gwd и pwd к батарее трансгенов, экспрессируемых в этих клетках, может обеспечить переваривание телец включения. Если это так, это подтверждает теорию о том, что дополнительный glgC ведет к росту более длинных неразветвленных областей глюкановых цепей, которые затем спонтанно скручиваются в двойные спирали с соседними цепями, вызывая агрегацию полисахарида.
Таким образом, эта работа демонстрирует, что простые модификации нативного аппарата синтеза гликогена у E.coli может приводить к образованию гранул нерастворимого полисахарида, которые в условиях голодания не перевариваются. Такие полисахариды могут быть легко извлечены из культуральной среды и как таковые могут представлять собой полезный промышленный метод улавливания сахаров, высвобождаемых из целлюлозных материалов, в форме, которая может быть легко преобразована в глюкозу для последующей обработки с целью получения биотоплива или химического сырья. Такой «псевдокрахмал», если он образован из неперевариваемой биомассы или сахарных отходов, также может быть испытан в качестве кормовой добавки для домашнего скота, потенциально высвобождая большие количества зерна для использования в пищу человеком.
Материалы и методы
Химическая трансформация
Компетентные клетки E. coli JM109 получали и трансформировали, как описано Chung et al. (32).
Рост для максимального увеличения содержания полисахаридов
Ночные культуры (выращенные в LB (таблица 1), 37 ° C, 200 об / мин) разбавляли 1: 100 и выращивали до фазы экспоненциального роста (0,5–0,6 OD λ600 нм ) в модифицированном Kornberg Средняя (таблица 1) при соответствующем выборе при 37 ° C и 200 об / мин. Затем в культуры добавляли изопропил-β-D-1-тиогалактопиранозид (IPTG) до 1 мМ, чтобы индуцировать P lac , и инкубировали еще 20 часов при 22 ° C и 200 об / мин.Затем их центрифугировали при 3000 × g в течение 15 минут при комнатной температуре, супернатант удаляли и осадок промывали раствором M9 × 1 (таблица 1). Затем осадки клеток ресуспендировали в среде M9 с IPTG (90 мг / л), хлорамфениколом (40 мг / л) и лактозой (20 г / л) и инкубировали еще 4 часа при 37 ° C и 200 об / мин (модифицировано из Sundberg. и др., 33). Затем культуры можно центрифугировать при 3000 × g в течение 15 минут при комнатной температуре, дважды промыть в PBS (таблица 1) и уравнять оптическую плотность при 600 нм перед анализом.
Таблица 1:
Среда для выращивания и аналитические растворы
Кривые роста
Три ночные культуры LB с хлорамфениколом (40 мг / л) были созданы для каждого исследуемого трансформанта, каждая инокулированная отдельной колонией из планшета для трансформации. Их инкубировали в течение ночи при 37 ° C и 200 об / мин, затем OD λ600 нм выровняли до 3,0 (± 0,09). Из каждой ночной культуры использовали 100 мкл для инокуляции свежих 10 мл культур: LB с IPTG (90 мг / л), хлорамфениколом (40 мг / л) и лактозой (20 г / л) и средой Корнберга с IPTG (90 мг / л). / л), хлорамфеникол (40 мг / л) и лактоза (20 г / л).Культуры встряхивали для перемешивания перед тем, как аликвоты по 100 мкл были перенесены из каждой свежей культуры в лунки 96-луночного планшета с плоским дном Costar 3628 в порядке, разработанном для рандомизации распределения культур и устранения краевого эффекта. Остальные лунки, окружающие образцы культур, были заполнены 100 мкл стерильной среды. В первом случае 100 мкл переносили из каждой культуры в 96-луночный планшет дважды, используя два отдельных планшета, которые запускались одновременно. Во втором случае из каждой культуры переносили только одну аликвоту, так что использовали одну чашку.96-луночные планшеты инкубировали в микропланшете Tecan Sunrise Absorbance Reader 30041769 при 37 ° C и «нормальном» встряхивании (4,4 мм, 9,2 Гц). Показания оптической плотности снимали каждые 15 минут в течение 24 часов с использованием режима измерения «точность». В то же время оставшиеся 9,98 мл культур инкубировали при 37 ° C и 200 об / мин. Показания оптической плотности снимали с этих культур каждые два часа в течение первых 14 часов, затем через 24 часа. Каждое показание оптической плотности получали путем смешивания 100 мкл культуры с 900 мкл стерильной среды в стандартной кювете, затем измерения OD λ600 нм против 1 мл стерильной среды на многомодовом считывающем устройстве Modulus Single Tube Multimode Reader (Turner Biosystems BS040271), оборудованном датчиком абсорбции. Модуль E6076.
Световая микроскопия
Для каждой культуры 10 мкл фиксировали нагреванием на предметном стекле микроскопа. Термофиксированный мазок замачивали в 20 мкл 5% йода Люголя (таблица 1) на 2 минуты, затем промывали избытком 100% этанола. Слайды просматривали с помощью настольного микроскопа Nikon eclipse E200 в фазовом контрасте при увеличении 1000 ×. Изображения световой микроскопии получали с помощью цифровой камеры Canon 1XUS9501S непосредственно через окуляр микроскопа. Все изображения считаются типичными для культур.
Просвечивающая электронная микроскопия
Для каждого штамма две культуры по 50 мл выращивали в течение ночи в LB с IPTG (90 мг / л), хлорамфениколом (40 мг / л) и лактозой (10 г / л) при 37 ° C и 200 ° C. об / мин. Затем целые культуры центрифугировали при 6000 × g в течение 10 минут. Супернатант удаляли, а осадок дважды промывали 25 мл PBS. Затем суспензию клеток снова центрифугировали при 6000 × g в течение 10 минут, супернатант удаляли и осадок, наконец, ресуспендировали в 5 мл PBS. Затем из каждой культуры 1 мл переносили в микроцентрифужную пробирку.Их центрифугировали при 6000 × g в течение 10 минут при комнатной температуре и фиксировали в 3% -ном глутаральдегиде в буфере C (0,1 М буфер какодилат натрия, pH 7,3) в течение 2 часов, затем промывали тремя 10-минутными сменами буфера C. затем постфиксировали в 1% тетроксиде осмия в буфере C в течение 45 минут, затем промыли тремя 10-минутными сменами буфера C. Затем эти образцы обезвоживали в 50%, 70%, 90% и 100% ацетонах нормальной чистоты в течение 10 минут. каждый, затем еще две 10-минутные смены ацетона аналара.Затем образцы заливали аралдитовой смолой. Срезы толщиной 1 мкм вырезали на ультрамикротоме Reichert OMU4, окрашивали толуидиновым синим и просматривали в световом микроскопе для выбора подходящих областей для исследования. Ультратонкие срезы толщиной 60 нм вырезали из выбранных участков, окрашивали уранилацетатом и цитратом свинца, а затем просматривали в просвечивающем электронном микроскопе Philips CM120. Снимки сделаны на камеру Gatan Orius CCD.
Анализ йода
Для каждой культуры аликвоты объемом 1 мл центрифугировали в настольной центрифуге при приблизительно 6000 × g в течение 3 минут при комнатной температуре.Осадки клеток ресуспендировали в 200 мкл PBS, к которому добавляли 50 мкл 100 мМ CuSO 4 , 50 мкл 6% об. / Об. H 2 O 2 и 25 мкл 0,2% йода Люголя. Суспензии могут быть перенесены в 48-луночные планшеты для визуализации с помощью планшетного сканера. Растущие колонии на чашках с агаром заливали 0,2% раствором йода Люголя для проверки наличия крахмала.
Влияние CuSO 4 и H 2 O 2 на интенсивность окрашивания исследовали путем смешивания 1 мл раствора крахмала при различных концентрациях с 25 мкл 0.2% йода Люголя с предварительным добавлением или без добавления 50 мкл 100 мМ CuSO 4 и 50 мкл 6% об. H 2 O 2 . Оптическую плотность каждой культуры измеряли при 620 нм. Растворимый крахмал был от Scientific Laboratory Supplies (CHE3620).
Антроновые анализы (индивидуальные)
Осадки промытых клеток ресуспендировали в PBS до объема, на котором они были выращены. Оптические плотности всех культур были уравновешены. Для каждой клеточной суспензии 3 аликвоты по 0.33 мл переносили в микроцентрифужные пробирки и в каждую добавляли 0,66 мл реагента антрон (таблица 1). Стандартные растворы глюкозы (0 мкг мл -1 , 2 мкг мл -1 , 10 мкг мл -1 , 20 мкг мл -1 , 50 мкг мл -1 и 100 мкг мл — 1 ), и для каждого стандарта 3 аликвоты по 0,33 мл смешивали с 0,66 мл реагента Антрон, чтобы получить стандартную кривую концентрации сахара. Порядок, в котором реагент добавлялся к образцам, был установлен таким образом, чтобы один набор стандартных растворов глюкозы вступал в реакцию в начале, в середине и в конце процесса анализа.Порядок, в котором реагент добавлялся к каждому набору образцов культуры, также менялся. Образцы оставляли на льду на 45 минут. Затем все образцы переносили в стандартные кюветы и измеряли при OD λ620 нм против 0 мкг мл -1 стандартного образца на спектрофотометре Hitachi Digilab U-1800. Концентрация глюкозного эквивалента сахара в гексозе для каждого образца оценивалась по стандартной кривой концентраций глюкозы. Таким образом, анализы измеряли общее содержание сахара в гексозе в клетках как эквивалент глюкозы.
Эксперимент по голоданию
В начале периода голодания клетки, которые культивировали для максимального увеличения содержания полисахаридов, извлекали центрифугированием, промывали и ресуспендировали в М9 с хлорамфениколом (40 мг / л) и доводили до OD λ600 нм 1,5. Затем каждую культуру инкубировали при 37 ° C и 200 об / мин без источника углерода и отбирали пробы в заданные моменты времени. В каждый момент времени 3 аликвоты по 0,33 мл каждой культуры переносили в микроцентрифужные пробирки и анализировали, как описано выше.Поэтому анализы Anthrone для эксперимента с голоданием измеряли общее содержание сахара-гексозы в культурах, чтобы измерить, сколько сахара-гексозы, обнаруженного в исходных осадках промытых клеток в начале эксперимента, метаболизировалось с течением времени.
Электрофорез в полиакриламидном геле с SDS
Обработанные ультразвуком аликвоты культуры, содержащие 30 мкг белка в каждом случае (рассчитанные с помощью анализов Брэдфорда), разделяли по размеру на предварительно отлитых полиакриламидных гелях Mini-Protean TGX TM (4-15%) при температуре 120 В, согласно инструкциям BioRad Laboratories.
Конструкция BioBrick ™
Конструкция плазмиды соответствует методу BioBrick RFC10, как впервые было описано Knight (34). Штамм E. coli JM109, стандартная плазмида синтетической биологии pSB1C3 и BioBrick BBa_K523005 (P LAC -RBS-LacZ-RBS) использовали для всех процедур клонирования и размножения плазмидной ДНК. Для glgC и glgC16 использовали части BioBrick BBa_K118015 и BBa_K118016 (соответственно), давая гены с удаленными сайтами EcoRI и заменой G336D в случае glgC16 .В таблице 2 показаны праймеры, использованные для амплификации glgB и glgC из хромосомы E. coli , а также праймеры, использованные для удаления сайтов EcoRI (glgCm1 и glgCm2). Все рестрикционные ферменты были приобретены в New England Biolabs (NEB) и использовались в соответствии с инструкциями производителя.
Благодарности
Эта работа была поддержана Исследовательским советом по биотехнологии и биологическим наукам, номер гранта BB / F017073 / 1.
гранулы гликогена | SGD
Аннотации
Отобран вручную
Увеличьте общее количество строк, отображаемых на этой странице, используя раскрывающийся список, расположенный под таблицей, или используйте страницу
прокрутите в правом верхнем углу таблицы, чтобы просмотреть ее страницы; используйте стрелки справа от заголовка столбца
отсортировать по этому столбцу; отфильтруйте таблицу, используя поле «Фильтр» в верхней части таблицы.Идентификатор доказательства Анализировать ID Джин Систематическое название гена Квалификация Идентификатор термина генной онтологии Термин онтологии гена Аспект Расширение аннотации Доказательства Метод Источник Назначено Номер ссылки Высокая пропускная способность
Увеличьте общее количество строк, отображаемых на этой странице, используя раскрывающийся список, расположенный под таблицей, или используйте страницу
прокрутите в правом верхнем углу таблицы, чтобы просмотреть ее страницы; используйте стрелки справа от заголовка столбца
отсортировать по этому столбцу; отфильтруйте таблицу, используя поле «Фильтр» в верхней части таблицы.Идентификатор доказательства Анализировать ID Джин Систематическое название гена Квалификация Идентификатор термина генной онтологии Термин онтологии гена Аспект Расширение аннотации Доказательства Метод Источник Назначено Номер ссылки Вычислительный
Увеличьте общее количество строк, отображаемых на этой странице, используя раскрывающийся список, расположенный под таблицей, или используйте страницу
прокрутите в правом верхнем углу таблицы, чтобы просмотреть ее страницы; используйте стрелки справа от заголовка столбца
отсортировать по этому столбцу; отфильтруйте таблицу, используя поле «Фильтр» в верхней части таблицы.Идентификатор доказательства Анализировать ID Джин Систематическое название гена Квалификация Идентификатор термина генной онтологии Термин онтологии гена Аспект Расширение аннотации Доказательства Метод Источник Назначено Номер ссылки болезнь накопления гликогена (GSD); Симптомы, причины, лечение
Обзор
Что такое гликоген и болезнь накопления гликогена (GSD)?
Клеткам тела требуется постоянный запас топлива для правильного функционирования.Это топливо представляет собой простой сахар, называемый глюкозой. Глюкоза образуется в результате расщепления пищи, которую мы едим. Организм использует столько глюкозы, сколько ему необходимо для функционирования, а остальное сохраняет, чтобы использовать позже.
Прежде чем его можно будет хранить, организм должен объединить простые единицы глюкозы в новый сложный сахар, называемый гликогеном. Затем гликоген накапливается в печени и мышечных клетках. Когда организму требуется дополнительное топливо, он расщепляет гликоген, хранящийся в печени, обратно на единицы глюкозы, которые клетки могут использовать. Специальные белки, называемые ферментами, помогают производить и расщеплять гликоген в процессе, называемом метаболизмом гликогена.
Иногда человек рождается без фермента, необходимого для этого процесса, или он может работать неправильно. Тогда организм не сможет хранить или расщеплять гликоген должным образом. Это может привести к очень низкому уровню глюкозы в крови во время голодания. Мышцам и органам для правильной работы необходим определенный уровень глюкозы в крови.
Когда в организме отсутствует фермент или имеется дефектный фермент и он не может правильно использовать гликоген, это приводит к состоянию, называемому болезнью накопления гликогена (GSD).Для обработки гликогена организм использует множество различных ферментов. И, как следствие, существует несколько типов GSD.
Какие бывают типы GSD?
Каждый тип GSD основан на определенном ферменте или наборе ферментов, участвующих в хранении или распаде гликогена. Существует как минимум 13 типов болезни накопления гликогена. Врачи знают о некоторых типах больше, чем о других. GSD в основном поражает печень и мышцы. Некоторые типы вызывают проблемы и в других частях тела. Типы GSD и части тела, на которые они влияют больше всего, включают:
- Тип 0 (болезнь Льюиса) — Печень.
- Тип I (болезнь фон Гирке) Тип Ia — печень, почки, кишечник; Тип Ib — печень, почки, кишечник, клетки крови.
- Тип II (болезнь Помпе) — мышцы, сердце, печень, нервная система, кровеносные сосуды.
- Тип III (болезнь Форбса-Кори) — печень, сердце, скелетные мышцы, клетки крови.
- Тип IV (болезнь Андерсена) — печень, мозг, сердце, мышцы, кожа, нервная система.
- Тип V (болезнь Макардла) — Скелетные мышцы.
- Тип VI (болезнь Герса) — Печень, клетки крови.
- Тип VII (болезнь Таруи) — Скелетные мышцы, клетки крови.
- Тип IX — Печень.
- Тип XI (синдром Фанкони-Бикеля) — печень, почки, кишечник.
Насколько они распространены?
Расстройство накопления гликогена встречается примерно у одного из 20 000–25 000 младенцев. Наиболее распространенными типами GSD являются типы I, II, III и IV, причем тип I является наиболее распространенным. Считается, что почти 90% всех пациентов с GSD имеют типы от I до IV. Считается, что около 25% пациентов с GSD имеют тип I.Однако GSD типов VI и IX могут иметь очень легкие симптомы и не могут быть диагностированы.
Большинство тяжелых форм GSD диагностируется у младенцев и детей. Некоторые из более мягких типов могут не обнаружиться, пока человек не станет взрослым.
Симптомы и причины
Каковы симптомы болезни накопления гликогена (GSD)?
Симптомы зависят от типа GSD.Некоторые GSD поражают в основном печень. К ним относятся типы 0, I, III, IV, VI и IX. Однако иногда они могут иметь частично совпадающие симптомы, влияющие на мышцы и сердце. Эти типы (за исключением GSD типа 0) могут вызывать увеличение печени. Увеличение печени связано с низким уровнем глюкозы в крови, потому что избыток гликогена хранится в печени, а не выделяется в виде глюкозы в кровоток. Симптомы низкого уровня глюкозы в крови или гипогликемии включают потливость, тремор, сонливость, спутанность сознания, а иногда и судороги.Некоторые GSD, такие как типы V и VII, в основном поражают скелетные мышцы. Мышечная слабость и мышечные судороги являются наиболее частыми симптомами этих типов.
Другие симптомы, которые могут возникнуть, включают:
- Усталость.
- Очень медленный рост.
- Ожирение (очень лишний вес).
- Проблемы с кровотечением и свертыванием крови.
- Проблемы с почками.
- Низкая устойчивость к инфекциям.
- Проблемы с дыханием.
- Проблемы с сердцем.
- Язвы во рту.
- Подагра.
Что вызывает GSD?
GSD возникают, когда есть проблема с геном, который имеет инструкции по производству фермента, который отсутствует или работает неправильно. Ген передается от родителей к детям. В большинстве случаев, чтобы иметь GSD, ребенок должен получить плохой ген от обоих родителей. Тот факт, что у обоих родителей есть этот ген, не всегда означает, что они оба передадут его своим детям.
Диагностика и тесты
Как выявляются типы болезней накопления гликогена (GSD)?
Есть четыре симптома, по которым врач может заподозрить тип GSD, поражающий печень.
Сюда входят:
- Низкий уровень глюкозы в крови.
- Увеличенная печень.
- Отставание в росте.
- Аномальные анализы крови.
Поскольку GSD могут работать в семьях, подробный медицинский анамнез также может дать врачу первую подсказку. Он или она может предложить несколько тестов, которые могут включать:
- Анализы крови — чтобы узнать уровень глюкозы в крови и увидеть, как работают ваша печень, почки и мышцы.
- УЗИ брюшной полости — чтобы увидеть, увеличена ли ваша печень.
- Биопсия ткани — исследование образца ткани мышцы или печени для измерения уровня гликогена или ферментов.
- Тестирование генов — поиск проблем с генами различных ферментов. Генное тестирование может подтвердить GSD.
Ведение и лечение
Как лечится болезнь накопления гликогена (GSD)?
Лечение зависит от типа GSD.При типах GSD, поражающих печень, лечение направлено на поддержание нужного уровня глюкозы в крови. Этого часто бывает достаточно, чтобы поддерживать потребности элементов в топливе и предотвращать долгосрочные осложнения, связанные с плохо контролируемым GSD. Лечение заключается в регулярном приеме сырого кукурузного крахмала и / или пищевых добавок. Кукурузный крахмал — сложный углевод, который трудно переваривать организмом; поэтому он поддерживает нормальный уровень сахара в крови в течение более длительного периода времени, чем большинство углеводов в пище.Употребление большого количества небольших блюд с низким содержанием сахара может помочь поддерживать нормальный уровень сахара в крови, предотвращая при этом избыточное хранение гликогена в печени.
Растворы углеводов можно давать постоянно в течение ночи, чтобы предотвратить падение уровня глюкозы в крови во время сна, но это несет больший риск тяжелой гипогликемии по сравнению с круглосуточным употреблением сырого кукурузного крахмала.
GSD типа IV с прогрессирующим заболеванием печени, возможно, потребуется рассмотреть для трансплантации печени после тщательной оценки.
Профилактика
Можно ли предотвратить болезнь накопления гликогена (GSD)?
GSD передаются от родителей к детям через их гены. Следовательно, их нельзя предотвратить. Родители могут узнать с помощью генетического тестирования, несут ли они ген GSD. У обоих родителей должен быть ген одного и того же типа GSD, чтобы ребенок унаследовал заболевание.
Перспективы / Прогноз
Каковы перспективы для человека с болезнью накопления гликогена (GSD)?
Прогноз зависит от типа GSD и пораженных органов. Благодаря недавним достижениям в терапии, лечение очень эффективно при лечении болезней накопления гликогена, поражающих печень.GSD типа I считался смертельным заболеванием до 1971 года, когда было обнаружено, что постоянный источник глюкозы улучшает метаболические нарушения, связанные с этим заболеванием. В 1982 году сырой кукурузный крахмал начал использоваться в качестве постоянного источника глюкозы, и прогноз для пациентов, страдающих этим заболеванием, резко улучшился. Люди с этими типами теперь могут жить полноценной нормальной жизнью.
GSD, которые не лечить должным образом, могут привести к таким проблемам, как печеночная недостаточность, сердечная недостаточность и легочная недостаточность.В этих случаях снижается качество жизни и ожидаемая продолжительность жизни. Поскольку они влияют на очень многие системы органов, GSD типа II (болезнь Помпе) и GSD типа IV (болезнь Андерсена) очень трудно поддаются лечению и могут привести к летальному исходу.
Исследования в области заместительной ферментной терапии и генной терапии являются многообещающими, что может улучшить перспективы на будущее.
Жить с
Когда мне следует позвонить своему врачу по поводу GSD?
Позвоните своему врачу, если у вас необъяснимая мышечная слабость или какие-либо симптомы низкого уровня глюкозы в крови, такие как потливость, тремор, сонливость и спутанность сознания.
.
Изменение структуры гранул хранения углеводов у Cyanobacterium Synechocystis sp. Штамм PCC 6803 посредством усечений ветвящихся ферментов
РЕФЕРАТ
Хранение углеводов является важным элементом метаболизма цианобактерий и хлоропластов растений. Понимание того, как управлять метаболизмом и хранением углеводов, также является важным фактором в использовании цианобактерий для производства энергии.В то время как большинство цианобактерий производят гликоген, было обнаружено, что некоторые из них накапливают полисахариды в форме нерастворимого в воде α-глюкана, аналогичного амилопектину. Примечательно, что эта альтернативная форма, называемая «полуамилопектин», образуется у видов цианобактерий, несущих три гомолога фермента ветвления (BE), обозначенные BE1, BE2 и BE3. В этом исследовании мутагенез генов ветвления, обнаруженных у Synechocystis sp. штамм PCC 6803 был выполнен, чтобы охарактеризовать их возможное влияние на морфологию полисахаридных гранул хранения.N-концевые усечения были сделаны для природного гена BE Synechocystis sp. PCC 6803. Кроме того, один из двух генов нативного фермента разветвления был заменен геном гетерологичного фермента разветвления из полуамилопектин-образующего штамма. Рост и содержание гликогена у мутантных штаммов существенно не отличались от таковых у дикого типа, а ультраструктурный анализ выявил лишь незначительные изменения морфологии гранул. Однако анализ распределения длины цепи с помощью анионообменной хроматографии выявил умеренные изменения профиля длины разветвленной цепи.Полученный гликоген имеет общие структурные характеристики, аналогичные характеристикам гранул, выделенных из штаммов, продуцирующих полуамилопектин.
ВАЖНОСТЬ Это первое исследование, в котором изучается влияние усечения ферментов ветвления на структуру запасных углеводов у цианобактерий. Результаты этого исследования являются важным вкладом в понимание взаимосвязи между ферментативным репертуаром цианобактерий и морфологией запасаемых ими углеводов.
ВВЕДЕНИЕ
Ассимиляция фотосинтетического углерода и последующее хранение образующихся углеводов являются важнейшим процессом в клетках цианобактерий во время суточного роста. Этот процесс происходит в непосредственной близости от тилакоидных мембран, которые обычно располагаются вокруг центрального цитоплазматического пространства или радиально выступают из полюсов клеток (1). Большинство бактерий, включая цианобактерии, производят растворимый гликоген (2–4). Фактически, использование гликогена в качестве запасного полимера полисахарида широко распространено в природе и может быть обнаружено у бактерий, архей и эукариот (5).Биосинтез гликогена происходит за счет последовательного действия гликогенсинтазы (GlgA) АДФ-глюкозопирофосфорилазы (GlgC) и фермента ветвления (BE; GlgB) (рис. 1). Вкратце, глюкоза активируется путем добавления АДФ, чтобы стать АДФ-глюкозой, которая затем полимеризуется на невосстанавливающем конце α-1,4-связанной цепи глюкана. Затем растущие полимерные цепи глюкозы связываются вместе в положении α-1,6 с помощью фермента разветвления (GlgB) посредством реакции гидролитического расщепления, в результате чего связь α-1,4 разрывается, и меж- или внутримолекулярный перенос повторно соединяет цепь.Эти действия продолжаются последовательно, в конечном итоге образуя гранулы гликогена. Разрушение гранулы гликогена происходит под действием двух ферментов: фермента разветвления (DBE; GlgX) и гликогенфосфорилазы (GPase и GlgP). (См. Обзоры структур гликогена [6] и крахмала [7].)
FIG 1
Пути биосинтеза и деградации гликогена. (A) Процесс биосинтеза гликогена происходит за счет активности трех важнейших ферментов: АДФ-глюкозопирофосфорилазы (GlgC), гликоген / крахмалосинтазы (GlgA) и фермента ветвления (GlgB).Разложение происходит через фермент разветвления (GlgX) и гликогенфосфорилазу (GlgP). (B) Короткоцепочечное разветвление бактериального гликогена приводит к экспоненциальному увеличению плотности цепей дальше от ядра, что приводит к максимуму 42 нм. Этот тип накопительных гранул наблюдается у Synechocystis sp. PCC 6803 (C) и Cyanothece sp. РСС 7822 (D). Для крахмала (E) кластерная структура амилопектина создается за счет асимметричного распределения ветвей, что позволяет формировать гораздо более крупные гранулы.Полуамилопектины из Cyanothece sp. ATCC 51142 и ATCC 51472 показаны на панелях F и G соответственно. Панели B и E адаптированы из справочного материала 8 с разрешения.
Морфология углеводов гликогена и крахмала весьма различна. Паттерн ветвления, обнаруженный в гликогене, обеспечивает более сферический рост. Эта характеристика приводит к появлению плотно упакованных гликозильных цепей, которые постепенно становятся более тесными к периферии. Из-за этой модели рост гранул в конечном итоге становится невозможным, поскольку пространство для взаимодействия с каталитическими центрами становится ограниченным.Математическое моделирование предсказало, что из-за этих характеристик максимальное значение для гликогена составляет 42 нм в диаметре (6). С другой стороны, крахмал состоит из амилопектина и амилозы и может образовывать гораздо более крупные гранулярные структуры из осмотически инертной и метаболически недоступной (до переваривания) глюкозы (рис. 1B и E) (7–9).
В то время как большинство цианобактерий образуют бактериальный гликоген, некоторые виды цианобактерий производят другую форму запасного полисахарида, которая является более нерастворимой и аналогичной амилопектину, содержащемуся в крахмале.Эта разновидность гранул для хранения полисахаридов называется «цианобактериальным крахмалом» и считается полуамилопектином из-за более низкого содержания в нем более длинных цепей глюкана по сравнению с цепями глюкана, обнаруженными в истинном амилопектине. Накопление полуамилопектина в качестве запасных полисахаридов было обнаружено у ограниченного, но растущего числа видов цианобактерий (10). Одно из первых описаний полуамилопектина было сделано для одноклеточного диазотрофного штамма Cyanothece sp. штамм ATCC 51142 (11).Считалось, что все представители рода Cyanothece хранят углеводный материал в этой форме, пока мы не сообщили о наличии небольших гранул гликогена в штамме Cyanothece sp. PCC 7822 аналогичен тем, которые хранятся у других видов бактерий и цианобактерий, таких как Synechocystis sp. штамм PCC 6803 (12). Примеры цианобактериального гликогена из генетических модельных видов Synechocystis sp. PCC 6803 и Cyanothece sp. PCC 7822, а также полуамилопектин из Cyanothece sp. ATCC 51142 и Cyanothece sp.ATCC 51472, можно увидеть на электронных микрофотографиях на рис. 1.
Распространенным методом характеристики молекулярной архитектуры полисахарида является анализ распределения длин цепи глюкана с использованием анионообменной хроматографии (13). Этот метод позволяет измерить степень полимеризации (DP) или количество моносахаридных единиц (например, глюкозы), связанных в полисахаридной цепи. Полуамилопектин, по сравнению с гликогеном, имеет высокое относительное количество более длинных α-1,4-связанных цепей глюкана с DP ≥37, составляющим от 2 до 6%.Напротив, обычный гликоген содержит <1% α-1,4-связанных цепей глюкана с DP ≥37, тогда как истинный амилопектин из риса имеет от ~ 6 до 7% этих длинных цепей. Когда анализируется профиль DP цианобактериального гликогена, средний относительный процент находится между DP 6 и 8. Среднее значение DP амилозы колеблется от 11 до 12. Среднее значение DP от 11 до 12 также сообщалось для видов цианобактерий. которые образуют полуамилопектин. Анализ запасных полисахаридов Cyanothece sp. ATCC 51142, штамм Cyanobacterium Clg1 и штамм Cyanobacterium NBRC 102756 показали, что их запасающие гранулы имеют молекулярную массу, практически неотличимую от молекулярной массы амилопектина (9, 14).Более того, термические свойства, кристалличность и разветвленная структура аналогичны амилопектину, а полуамилопектиновый материал, синтезируемый этими штаммами, организован в тандемные кластерные структуры.
Неизвестно, почему некоторые цианобактерии образуют различные гранулы для хранения углеводов. Биоинформатический анализ видов Cyanothece, Cyanobacterium и Synechocystis выявил различное количество копий всех генов, участвующих в синтезе и деградации запасных полисахаридов (таблица 1).Примечательно, что все штаммы, образующие полуамилопектин, содержат три копии гена фермента ветвления, тогда как бактериальные штаммы, образующие гликоген, содержат либо одну, либо две. Кроме того, среди подгрупп штаммов можно увидеть высокую вариабельность генов ферментов разветвления. В этом исследовании были изучены различия между аминокислотными последовательностями ферментов ветвления, обнаруженных у полуамилопектин- и неполуамилопектин-образующих цианобактерий. Целью этого исследования было использование генетических модификаций для преобразования штамма цианобактерий, продуцирующего бактериальный (β-гранулы) гликоген, в штамм, продуцирующий крахмалоподобный полуамилопектин.
ТАБЛИЦА 1
Число копий генов метаболизма гликогена у различных представителей Cyanothece, Synechocystis и цианобактерий, которые, как известно, продуцируют крахмалоподобные гранулы
МАТЕРИАЛЫ И МЕТОДЫ
Рост и конструирование мутантов. Все цианобактериальные штаммы, использованные в этом исследовании, перечислены в таблице 2. Synechocystis sp. Штаммы PCC 6803 выращивали в колбах объемом 100 мл, содержащих среду BG-11 (15), тогда как Cyanothece sp. ATCC 51142 (16) выращивали в среде ASP2 (17, 18), оба со встряхиванием при ∼125 об / мин при освещении от ∼30 до 40 мкмоль фотонов m −2 с −1 света от холодных белых флуоресцентных ламп. .Для экспериментальных процедур культуры инокулировали в идентичные среды и, если указано, модифицировали путем пересева в безнитратную среду (BG-11 NF) или в среде с 0,01 × нитратом. Рост анализировали с использованием оптической плотности при 730 нм (спектрофотометр Perkin-Elmer UV-Vis Lambda 40).
ТАБЛИЦА 2
Бактериальные штаммы и плазмиды, использованные в этом исследовании
Для генерации мутаций геномную ДНК экстрагировали из 100 мл культуры (~ 1 × 10 8 клеток / мл). Гомогенизатор Braun и 0,5 мл стеклянных шариков (0.1–0,2 мм) использовали для лизиса клеток четырьмя 30-секундными импульсами на высокой скорости с последующим центрифугированием. Затем супернатант переносили в новые пробирки и последовательно промывали встряхиванием 500 мкл фенола и хлороформа. ДНК осаждали добавлением 2 об. Ледяного абсолютного этанола. Для конструкций фермента ветвления pUC19 использовали в качестве основы клонирования для последовательной вставки фрагментов размером ~ 500 нуклеотидов (нт), расположенных непосредственно перед нативным стартовым кодоном и ниже кодирующей последовательности гена фермента ветвления sll0158 .Сначала восходящий фрагмент амплифицировали с использованием праймеров BEup / F и BEup / R и вставляли в сайты PstI и XmaI. Нижележащая область, амплифицированная с использованием праймеров BEdown / F и BEdown / R, была вставлена в сайты XmaI и EcoRI. Кодирующая последовательность sll0158 была амплифицирована с помощью ПЦР с прямыми праймерами BE97 / f и BE112 / f и обратным праймером BE / R1. Эти праймеры амплифицировали область гена, соответствующую 97-й или 112-й аминокислоте, вводя стартовый кодон в положение 97 (давая мутантный ген sll0158 Δ97) или 112 (давая мутантный ген sll0158 Δ112), соответственно.Мутант sll0158 фрагментов вставляли между вышестоящим и нижележащим фрагментами с использованием сайтов XmaI и BglII. Кассету устойчивости к канамицину из pRL448, амплифицированную с использованием праймеров BEKanF и BRKanR, использовали для положительной селекции и вставляли между усеченным геном фермента ветвления и нижележащей областью.
Процедуру, аналогичную описанной выше, использовали для замены нативного фермента разветвления (DBE), кодируемого геном slr0237 , на DBE от Cyanothece sp.PCC 8801. Вышестоящие и нижележащие фрагменты нативного DBE были амплифицированы с использованием праймеров U0237F / R и D0237F / R и вставлены через сайты BamHI и SalI и сайты SalI и PstI в pUC19 соответственно. Кодирующая последовательность DBE из Cyanothece sp. PCC 8801 амплифицировали с использованием пары праймеров 8801-1045F / 8801-1045R и вставляли между передним и нижележащим фрагментами через сайт SalI. Затем кассета спектиномицина (~ 2 т.п.н.), выделенная из плазмиды pRL453, была вставлена между геном DBE, PCC8801_1045 , и нижележащим фрагментом slr0237 в сайте EcoRI.Synechocystis sp. PCC 6803 трансформировали с использованием стандартных протоколов трансформации (19). Трансформированные колонии отбирали на чашках с антибиотиками и переносили на 2-4 месяца для разделения. Полную сегрегацию мутантов подтверждали с помощью ПЦР, а затем через регулярные промежутки времени. Последовательности для используемых праймеров можно найти в Таблице 3.
ТАБЛИЦА 3
Последовательности праймеров, использованные в исследовании
Определение содержания гликогена. Внутриклеточное содержание гликогена измеряли с помощью колориметрического биохимического анализа с использованием антрона, описанного ранее (11, 12, 16, 20, 21).Образцы измеряли в одинаковые ежедневные временные точки с использованием 2 мл культуры. Полученные значения поглощения при 625 нм сравнивали со стандартной кривой, как описано Welkie et al. (12).
Приготовление полисахаридов. Культуры (500 мл) выращивали в течение 3-4 дней в условиях постоянного света и низкого содержания азота (0,01 × нитратная среда BG-11) для увеличения общего запаса гликогена. Клетки собирали центрифугированием при 10000 × g и суспендировали в 20 мл буфера для разрушения, содержащего 50 мМ имидазол-HCl (pH 7.4), 8 мМ MgCl 2 и 12,5% (об. / Об.) Глицерина (9) и помещали на лед. Суспензию пропускали через ячейку под давлением French три раза при 16000 фунтов / дюйм 2 (16), при этом пробирки для сбора были погружены в лед для максимального подавления ферментативной активности. После лизиса раствор кипятили в течение 10 мин, а затем центрифугировали при 8000 × g в течение 15 мин. Гликоген осаждали из супернатанта добавлением 5 объемов ледяного абсолютного этанола (22).Гликоген выделяли центрифугированием при 8000 × g в течение 20 минут, сушили в центробежном вакуумном испарителе (9) и хранили при -20 ° C до обработки.
Анализ распределения длин цепей с помощью HPAEC. Чтобы изучить паттерн ветвления гликогена, продуцируемого диким типом и мутантными штаммами Synechocystis sp. PCC 6803, выделенный полисахаридный материал из клеточного лизата (100 мг) растворяли в 10 мл H 2 O (10% раствор [вес / объем]) и перемешивали в течение ночи для диспергирования.Затем образцы кипятили в течение 1 ч с последующим центрифугированием при 12000 × g в течение 10 мин. Супернатант разбавляли 10 раз 10 мМ натрий-ацетатным буфером (pH 4,5) и добавляли по 1 мкл изоамилазы (0,1 ед.) И пулланазы (0,72 ед.) (Megazyme, Wicklow, Ирландия) для гидролиза α-1,6 связей. . Реакционную смесь инкубировали при 40 ° C при встряхивании при 120 об / мин в течение 48 ч. Распределение длин разветвленных цепей гликогена определяли с помощью системы высокоэффективной анионообменной хроматографии (HPAEC) (GP40; Thermo Scientific, Саннивейл, Калифорния), оснащенной электрохимическим детектором (ED40; Thermo Scientific).Анионообменная аналитическая колонка CarboPac PA100 (4 на 250 мм; Thermo Scientific) с защитной колонкой (4 на 50 мм; Thermo Scientific) использовалась для разделения 25 мкл проб от разветвленных проб. Элюенты A и B состояли из 150 мМ NaOH и 150 мМ NaOH, содержащих 600 мМ ацетата натрия, соответственно. Градиент элюента B на 70 мин (увеличиваясь на 1% / мин) составлял от 0 до 70%.
Анализ ультраструктуры. Ячейки визуализировали двумя способами с помощью просвечивающей электронной микроскопии (ПЭМ). Для регулярной морфологии клетки фиксировали замораживанием под высоким давлением (HPF), как описано ранее (12).Чтобы определить, содержат ли гранулы гликоген, использовали гистохимический метод PATO (периодическая кислота-тиокарбогидразид-осмий) (23, 24). Клетки отбирали в одинаковые моменты времени после 72 часов роста в среде BG-11 NF. Клетки предварительно окрашивали 0,15% рутениевым красным в 0,1 М натрий-какодилатном буфере, pH 7,0 (NaCaB) в течение 1 ч при комнатной температуре, а затем промывали. Первичное окрашивание состояло из 3,6% глутарового альдегида и 0,15% рутениевого красного в NaCaB, инкубированных в течение 1 ч при комнатной температуре.После этого следовала промывка NaCaB и затем процедура вторичного окрашивания 1% OsO 4 с 0,15% рутениевым красным в NaCaB в течение 1 часа при 4 ° C. Фиксированные образцы осаждали и покрывали 2% агарозой, и полученные гранулы разрезали на блоки для обезвоживания и заделки. Образцы дегидратировали с помощью серии градуированных этанолов и заливали свежей смолой Spurr. Тонкие срезы были вырезаны и извлечены с помощью никелевой сетки 100 меш, покрытой углеродной пленкой Formvar. Постокрашивание сеток осуществляли плаванием по каплям 1% -ной периодической кислоты в течение 1 ч при комнатной температуре, промыванием водой, плаванием по каплям 1% -ного тиосемикарбазида (Sigma-Aldrich, St.Луи, Миссури) в течение 1 ч при комнатной температуре, снова промыли водой и дали высохнуть на воздухе. Затем срезы подвергали воздействию паров OsO 4 в небольших закрытых чашках Петри в течение 7 часов при комнатной температуре, окрашивали 2% водным уранилацетатом в течение 5 минут, промывали водой и сушили на воздухе. Контроль для каждой временной точки был приготовлен путем повторного приготовления без периодной кислоты и тиосемикарбазида перед окончательной экспозицией паров осмия. Образцы получали с помощью ПЭМ Philips CM-100, работающего при 100 кВ, с пятном 3, апертурой конденсатора 200 мкм и апертурой объектива 50 мкм.Изображения были сняты на пленку Kodak SO-163.
Филогенетический анализ ферментов биосинтеза гликогена у Cyanothece, Synechocystis и Cyanobacterium spp. Аминокислотные последовательности для ферментов разветвления и разветвления были обнаружены в эталонных геномах с помощью анализа BLASTP (http://www.ncbi.nlm.nih.gov/BLAST) для получения всех связанных последовательностей, показывающих высокое сходство. Белки, которые оказались неполными, были исключены из анализа. Затем последовательности были выровнены с помощью программы Clustal Omega (ClustalO) с использованием настроек по умолчанию во время анализа с числом итераций, установленным на пять (http: // www.ebi.ac.uk/Tools/msa/clustalo/). Полученный файл выравнивания был использован для создания дерева соединения соседей с помощью программы MEGA-6 для Mac (http://www.megasoftware.net/index.php). Чтобы прийти к сгенерированному дереву, был проведен анализ начальной загрузки с 1000 повторов. Дополнительное выравнивание N-концевых и каталитических доменов ферментов ветвления проводили аналогично ClustalO. Затем файл выравнивания был импортирован в ESPript 3 (http://espript.ibcp.fr/ESPript/ESPript/), и функции вторичной структуры были визуализированы с использованием PDB Chain ID 1M7X в качестве ссылки (http: // www.rcsb.org/pdb), аналогично анализу, выполненному Suzuki et al. (14).
РЕЗУЛЬТАТЫ
Аминокислотные последовательности и филогенетический анализ BE1, BE2 и BE3. Аминокислотные последовательности для всех ветвящихся ферментов, обнаруженных в геномах штаммов Cyanothece, включая известные полуамилопектин-образующие штаммы Cyanobacterium sp. Clg1 и Cyanobacterium sp. NBRC 102756, были выровнены для сравнения структур. Были идентифицированы три отдельные группы, названные BE1, BE2 и BE3 (см. Рис. S1 в дополнительном материале).Эти группы были основаны на том факте, что BE2 и BE3 не содержали 97 или 111 аминокислот на N-конце, что согласуется с наблюдениями, сделанными другими (10, 12), и что были различные вставки и делеции в каталитической области A-домена. (см. рис. S2) (14). Некоторые обычные гликоген-образующие виды, такие как Synechocystis sp. PCC 6803 несут только один ген BE для BE1, тогда как другие, такие как Cyanothece sp. PCC 7822, содержат два (таблица 1). Все подтвержденные штаммы цианобактерий, о которых известно, что они образуют полуамилопектин, за исключением Cyanothece sp.PCC 8801 содержал третью изоформу фермента ветвления (BE3).
На рис. 2 показано выравнивание белковой последовательности на амино-концах ферментов ветвления. BE2 от Cyanothece sp. ATCC 51142 и Cyanothece sp. Оба PCC 7822 содержат усечение из 97 аминокислот, тогда как BE3 из Cyanothece sp. ATCC 51142 содержит усечение из 111 аминокислот. Изоформы BE1 из Cyanothece sp. PCC 7822 и Cyanothece sp. ATCC 51142 были идентичны друг другу на 77%, тогда как изоформы BE2 этих штаммов были идентичны на 65%.Кроме того, изоформы BE1, BE2 и BE3 демонстрируют высокое сходство последовательностей с их соответствующими аналогами у разных видов (см. Таблицу S1 в дополнительном материале).
FIG 2
Множественное выравнивание аминокислотной последовательности N-концевого домена из BE из штаммов Cyanothece. Ec, Escherichia coli; S.6803 , Synechocystis sp. PCC 6803; C.7822 , Cyanothece sp. PCC 7822; C.51142 , Cyanothece sp. ATCC 51142; C.Clg1 , Cyanobacterium sp.Clg1; C.NBRC , Cyanobacterium sp. NBRC 102756. Выравнивание последовательностей было произведено с использованием ClustalO (43) и ESPript (http://espript.ibcp.fr) (44). Показаны области каталитического домена на основе трехмерной структуры BE из E. coli (PDB: 1M7X: D). Обозначены бета-нити (β).
Аминокислотные последовательности и филогенетический анализ DBE и DBE-X2. Некоторые исследования также показали корреляцию между морфологией гранул полисахарида и активностью фермента разветвления.Например, Suzuki et al. (25) сообщили, что делеция гена glgX в Synechococcus elongatus PCC 7942 привела к обогащению очень коротких цепей значениями DP в диапазоне от 2 до 4 и увеличению скорости поглощения кислорода в темноте. Кроме того, было продемонстрировано, что вариантная копия гена расщепляющего фермента, обнаруженная в крахмалообразующем штамме Cyanobacterium Clg1, имеет решающее значение для накопления крупных крахмалоподобных полисахаридов. Этот штамм содержит ген, получивший название glgX2 .Клетки с мутированным геном glgX2 накапливают небольшие β-гранулы гликогена с морфологией, напоминающей Synechocystis sp. PCC 6803 и Cyanothece sp. РСС 7822 (26). Интересно, что glgX2 отсутствует в Cyanothece sp. ATCC 51142. Это свидетельствует о том, что различные комбинации генов BE и DBE могут действовать согласованно, образуя различные структуры гранул полисахарида. Выравнивание белковых последовательностей для всех ферментов удаления ветвей из геномов, включенных в это исследование, показано на рис.S3 в дополнительном материале.
Число генов, кодирующих фермент разветвления, было более вариабельным, чем для генов фермента ветвления. Например, Synechocystis sp. PCC 6803 и Cyanothece sp. ATCC 51142 имеет два гена DBE, тогда как Cyanothece sp. В PCC 7822 их три. Все пять генов фермента разветвления показывают низкое сходство последовательностей с glgX2 из штамма Cyanobacterium Clg1 (<50%). Тем не менее, существует относительно близкая идентичность последовательностей между glgX2 и генами фермента разветвления из Cyanothece sp.PCC 8801, PCC 8802, PCC 7822 и PCC 7424, самые высокие из которых принадлежат генам Cyanothece sp. PCC 8801 с> 65%. Интересно, что электронные микрофотографии Cyanothece sp. PCC 8801 показывает большие запасающие гранулы, присутствующие в непосредственной близости от тилакоидов, подобные тем, которые обнаруживаются у Cyanothece sp. АТСС 51142 (27).
Мутагенез ферментов ветвления и разветвления у Synechocystis sp. PCC 6803. Чтобы исследовать роль N-концевой области фермента ветвления, усечения, подобные тем, которые обнаружены в BE2 и BE3 Cyanothece sp.ATCC 51142 были использованы для замены единственной копии BE (BE1) Synechocystis sp. PCC 6803. Это было достигнуто с использованием праймеров, которые амплифицировали кодирующую область фермента ветвления, начиная с 97 или 112 аминокислот ниже обычного стартового кодона. Нативная аминокислота в 97-м или 112-м положении была заменена (с помощью праймера для ПЦР) новым стартовым кодоном (AUG) (рис. 3). Сегрегацию мутантов подтверждали гель-электрофорезом ДНК амплифицированных ПЦР геномных областей, содержащих встроенную ДНК.
Рис. 3.
Мутагенез с ферментами разветвления и ферментами разветвления. (A) Карта, показывающая сайт мутагенеза с усечением аминоконцевых концов. (B) Гель-электрофорез, показывающий фрагменты ДНК, амплифицированные с помощью ПЦР, указывает на полную сегрегацию модифицированных генов в мутантных штаммах Δ97 и Δ112. (C) Карта, показывающая замену двойной гомологичной рекомбинации фермента разветвления slr0237 на PCC8801_1045. сша , верхний по течению регион; DS , низовья; нуклеотидов , нуклеотид, Sp R , ген устойчивости к спектиномицину; ×, расположение двойного гомологичного кроссовера.Стрелки указывают места, где были отожжены праймеры ПЦР, используемые для проверки мутагенеза генов.
Поскольку ген DBE PCC8801_1045 от Cyanothece sp. PCC 8801 имел наибольшее сходство последовательности с геном glgX2 у Cyanobacterium sp. Clg1, он также был интродуцирован в Synechocystis sp. PCC 6803. Один из двух генов DBE Synechocystis ( slr0237 ) был заменен на PCC8801_1045 посредством гомологичной рекомбинации, поддерживая регуляцию транскрипции от природного промотора.Мутантные клетки были отделены и проверены, как описано для мутаций усечения BE. Генетическая карта для конструкции, использованной для введения гена glgX2 из Cyanothece sp. PCC 8801 ( PCC8801_1045 ) показан на рис. 3.
Рост и содержание гликогена в мутантных штаммах. Сначала мы исследовали эффекты укороченного БЭ (Δ97 и Δ112) и добавления DBE из Cyanothece sp. PCC 8801 дикого типа и мутанты Δ97 и Δ112 по росту и физиологии клеток.Никакого существенного влияния на рост или содержание гликогена в любом из мутантных штаммов не наблюдалось при стандартных условиях непрерывного света. Через 72 часа рост культур снизился с 0,15 до 0,38 ± 0,02. Это также имело место, когда клетки, выращенные в стандартных условиях, промывали и переносили в безнитратную среду BG-11 при непрерывном освещении в течение 72 часов до конечной оптической плотности 0,33 ± 0,02. При стрессе с истощением азота содержание гликогена увеличивалось во всех штаммах без существенной разницы в накоплении гликогена между штаммами дикого типа и мутантными штаммами; содержание гликогена было 7.4 ± 2,0 и 43,6 ± 5,5 мкг / оптическая плотность при 730 нм (OD 730 ) через 72 часа в обычном и безнитратном BG-11, соответственно (данные не показаны).
Ультраструктурные характеристики мутантов Δ97, Δ97x и Δ112. Для определения наличия мутаций в BE и DBE Synechocystis sp. PCC 6803 влиял на морфологию гранул гликогена, мы исследовали клетки, выращенные в безазотных условиях в течение 72 часов, с помощью просвечивающей электронной микроскопии. Известно, что азотный стресс вызывает накопление гликогена в Synechocystis и позволяет наблюдать клетки с высоким уровнем гликогена.Клетки фиксировали замораживанием под высоким давлением (HPF), процедурой, которая, как было доказано, обеспечивает наилучшее сохранение клеточной ультраструктуры (12). Для окрашивания углеводов методом PATO (12, 23, 24) мы также использовали химическую фиксацию с помощью микроволновой процедуры. На рис. 4 показано изображение в тонком срезе Synechocystis sp. Дикого типа. Мутантные клетки PCC 6803 (фиг. 4A) и Δ97 (фиг. 4B), Δ97x (фиг. 4C) и Δ112 (фиг. 4D) не показывают заметных различий по морфологии гранул. Окрашенные PATO клетки дикого типа (рис.4E) подтвердил, что небольшие гранулы, видимые в тилакоидном и цитоплазматическом пространствах, заполняющих клетку, представляют собой углеводный материал. Контроль окрашивания PATO (фиг. 4F) показывает клетки дикого типа, полученные идентично, но без периодической обработки кислотой и тиосемикарбазидом перед окончательной экспозицией паров осмия.
Фиг. 4.
ТЕМ клеток Synechocystis дикого типа и мутантных клеток. Электронные микрофотографии Synechocystis sp. Показаны мутантные штаммы PCC 6803, выращенные в безазотной среде BG-11 и зафиксированные замораживанием под высоким давлением (HPF).Изображения показывают Synechocystis sp. Дикого типа. PCC 6803 (A) и мутанты Δ97 (B), Δ97x (C), Δ112 (D), а также мутанты дикого типа, окрашенные с использованием метода PATO с помощью химической фиксации в микроволновой печи (E) и контроля PATO дикого типа (F) . Увеличение, × 25000.
Измерения размера гранул, образующихся внутри каждого штамма, были выполнены с использованием программного обеспечения ImageJ (28). Диаметр отдельных гранул гликогена измеряли вручную ( n > 50, на основании 3+ различных изображений ПЭМ каждое) и сравнивали с таковыми для дикого типа.Средняя гранула гликогена Synechocystis sp. Дикого типа. PCC 6803 имел диаметр ~ 30 нм. Средняя гранула гликогена из мутантных клеток Δ97 была сравнима с гранулами гликогена дикого типа, но мутантные клетки Δ97x и Δ112 имели гранулы гликогена с небольшим увеличением размера, причем гранулы мутанта Δ112 были самыми крупными, со средним диаметром ∼35. нм (таблица 4). Размеры углеводных гранул в мутантных штаммах, описанных здесь, находятся в пределах диапазона диаметров, характерных для обычных гранул гликогена, и меньше, чем у полуамилопектина, обнаруженного у Cyanothece sp.ATCC 51142 и CCY 0110 и штаммы Cyanobacterium Clg1 и NBRC 102756 (9).
ТАБЛИЦА 4
Сравнение размеров гранул гликогена
Распределение длины цепи хранимых полисахаридов. Был проведен ряд исследований, описывающих судьбу накопления углеводов и физиологические эффекты на клетки-хозяева при изменении генов метаболизма гликогена. Ю и др. (29) показали, что делеция фермента ветвления у Synechocystis sp. PCC 6803 приводил к увеличению количества цепей α-глюкана с DP от 2 до 16 и уменьшению в более крупных цепях с DP от 17 до 44.Судзуки и др. (25) показали, что нарушение гена glgX в Synechococcus elongatus PCC 7942 привело к обогащению коротких цепей DP в диапазоне от 2 до 4. Это указывает на то, что фермент разветвления у цианобактерий может вносить вклад в структуру ветвления полисахарида. Кроме того, работа Cenci et al. на Cyanobacterium cp. Clg1 предоставил дополнительные доказательства того, что ферменты разветвления могут влиять на структуру хранения полисахаридов (26). Более того, экспрессия синтазы картофельного крахмала III в цианобактериальном хозяине привела к неожиданно более коротким и менее длинным α-1,4 цепям, чем у дикого типа (30).
В этом исследовании распределение длин разветвленных цепей экстрагированных α-глюканов дикого типа Synechocystis sp. PCC 6803, Cyanothece sp. ATCC 51142 и мутантные клетки Synechocystis анализировали с помощью HPAEC (фиг. 5). Полисахариды, выделенные из Synechocystis sp. Дикого типа. PCC 6803 (фиг. 5A) показал ожидаемое распределение длины цепи, при этом наиболее распространенная глюкановая цепь имела DP, равную 6, тогда как наиболее распространенная длина цепи глюкана в полисахаридах, выделенных из Cyanothece sp.АТСС 51142 имеет DP 11. Это сопоставимо с экспериментальными результатами, описанными ранее (25, 31, 32). Сравнение этих двух штаммов показывает, как полуамилопектин, синтезируемый Cyanothece sp. У ATCC 51142 было меньше коротких цепей (DP, от 2 до 10) и было больше длинных цепей (DP, от 11 до 35), чем у нормального гликогена β-гранул Synechocystis sp. PCC 6803. Материалы из мутантных штаммов Δ97 (рис. 5B), Δ112 (рис. 5C) и Δ112x (рис. 5D) были подобны материалам дикого типа в том, что наибольшая длина цепи соответствовала DP 6, но Δ97 имеет пониженные полиглюканы с короткой цепью и увеличение длинных цепей с DP> 16.Оба мутанта Δ112 и Δ112x показали результаты, аналогичные таковым для штамма Δ97, но с увеличением полиглюкановых цепей с DP> 22 и меньше, что связано с уменьшением короткоцепочечных глюканов. Вычитающий анализ относительного процента каждой DP Cyanothece sp. Дикого типа. PCC 51142 (фиг. 5E) и Δ97 (фиг. 5F), Δ112 (фиг. 5G) и Δ112x (фиг. 5H) выявили уменьшение коротких цепей (DP, от ~ 3 до 8) и увеличение длинных -цепи (DP, от ~ 13 до 30) в мутантных штаммах, заметно сходное с содержанием полуамилопектина из Cyanothece sp.ATCC 51142. Числовые (DP n ) и средневзвешенные (DP w ) значения DP для Synechocystis sp. Дикого типа. PCC 6803 и мутанты Δ97, Δ112 и Δ112x, а также Cyanothece sp. ATCC 51142 отражает результаты распределения (Таблица 5). DP n для дикого типа составлял 8,9 ± 0,1, в то время как для Cyanothece sp. ATCC 51142, DP n был 11,3. Значения DP n для мутантов Δ97, Δ112 и Δ112x были промежуточными и составляли 9,6 ± 0,1, 9,3 и 9,4 соответственно.Мы пришли к выводу, что описанные здесь ферментативные модификации привели к накопительной структуре α-глюкана у Synechocystis sp. PCC 6803 имеет сходство с полуамилопектином, что подразумевает вариацию ферментов ветвления как фактор, влияющий на морфологию хранения α-глюкана.
Рис. 5
Распределение длин цепей разветвленных α-глюканов дикого типа и мутантных Synechocystis sp. Штаммы PCC 6803. После обработки изоамилазой для гидролиза α-1,6 связей, α-глюкановые цепи были разделены с помощью HPAEC-PAD для PCC 6803 дикого типа (столбцы) и Cyanothece sp.ATCC 51142 (заштрихованная область) (A) и мутанты Δ97 (B), Δ112 (C) и Δ112x (D). Площадь пика рассчитывали и нормализовали по хроматограмме профиля цепи. Вычитающий анализ, показывающий разницу в нормированной площади пика между Cyanothece sp. PCC 51142 (E) и мутанты Δ97 (F), Δ112 (G) и Δ112 (H) выявили снижение содержания коротких цепей (DP, от ∼3 до 8) и увеличение содержания длинных цепей (DP, От ~ 13 до 30) в мутантных штаммах, что заметно сходно с таковым у цианобактериального α-глюкансодержащего Cyanothece sp.ATCC 51142. Изменения в коротких и длинных разветвленных цепях (заштрихованные области) показаны горизонтальными скобками.
ТАБЛИЦА 5
Влияние модификаций ферментов разветвления и разветвления гликогена на запасы и структуру гликогена a
ОБСУЖДЕНИЕ
Природа накопления углеводов является ключевым элементом метаболизма цианобактерий и хлоропластов растений. Этот процесс накопления углерода находится на пересечении фотосинтеза, дыхания и центрального метаболизма и оказывает глубокое влияние на рост цианобактерий и поддержание дневного гомеостаза.Метаболизм гликогена также имеет решающее значение для установления надлежащей внутриклеточной среды для фиксации N 2 у диазотрофных видов, особенно у одноклеточных цианобактерий. Его накопление во всех фотосинтетических ветвях древа жизни из зеленых растений, водорослей и цианобактерий представляет собой поглотитель углерода, который может накапливаться до высоких уровней по сравнению с биомассой, а затем расщепляться и подаваться в биосинтетические пути для химического производства. Поскольку большинство видов цианобактерий, образующих крахмал и полуамилопектин (например,g., Cyanothece sp.) были невосприимчивы к сайт-специфическим генетическим модификациям (33), выяснение функций определенных ферментов, которые вносят вклад в крахмалоподобную структуру хранения, является сложной задачей. Хотя методы генетической модификации Cyanothece sp. Были разработаны PCC 7822 (33), Synechocystis sp. PCC 6803 представляет собой штамм с превосходной генетической системой и более простой системой биосинтеза гликогена, только с одним ферментом ветвления и двумя DBE. Мутации, внесенные в BE и DBE в Synechocystis PCC 6803, описанные здесь, явно повлияли на метаболизм гликогена в умеренной степени.Усечения на N-конце нативного фермента ветвления у Synechocystis sp. PCC 6803 привел к паттерну разветвления гликогена с увеличением количества более длинных цепей глюкана и уменьшением количества более коротких по сравнению с диким типом. Наблюдаемое увеличение и уменьшение каждого соответствующего полимера, содержащего гранулы гликогена, экстрагированных из мутантных штаммов, согласуется с профилями длины цепи полуамилопектина, хотя процент от общего распределения не так велик.Предыдущая работа Yoo et al. Введение синтазы III картофельного крахмала привело к противоположным изменениям в схеме разветвления с уменьшением длинных цепей и увеличением количества коротких в масштабе, аналогичном наблюдаемому в этом исследовании (30). Таким образом, мы предполагаем, что модифицированные в данной работе ферменты BE играют важную роль в процессе образования крахмала, и считаем наши результаты шагом в правильном направлении для определения энзимологии производства крахмала. Добавление гетерологичного DBE могло также оказать дополнительное влияние на профиль длины цепи.Тем не менее, объем изменений указывает на то, что другие белки или потенциально белок-белковые взаимодействия (30, 34) должны быть задействованы, если весь гликоген у Synechocystis sp. PCC 6803 должен быть преобразован в полуамилопектин, как в Cyanothece sp. ATCC 51142.
Одной из проблем при сравнении биохимии крахмал-, полуамилопектин- и гликоген-образующих видов водорослей и бактерий является сложность метаболической сети запасающих полисахаридов у разных видов. По сравнению со многими эубактериями, цианобактерии имеют необычно большое количество изоформ для каждой стадии метаболического пути гликогена.Ball et al. (35) недавно исследовали геномы многих секвенированных цианобактерий и эукариотических водорослей, которые продуцируют крахмал или полуамилопектин, и разработали более полную и сложную картину эволюции производства крахмала. Действительно, они отвергли свою более раннюю гипотезу о том, что одно событие привело к приобретению крахмала хозяином после эндосимбиотического события, которое привело к развитию пластид у эукариотических водорослей, а затем и у растений (26, 31, 36, 37). Они продемонстрировали путем тщательного генетического скрининга, что изоамилазоподобный DBE (названный GlgX2) в Cyanobacterium Clg1 отвечает за образование крахмалоподобного материала.Их геномный анализ привел их к выводу, что приобретение крахмала часто происходило у цианобактерий и эукариот и что переход гликогена в крахмал, вероятно, происходил много раз (35). Этот продукт гена glgX2 или любой Gh23-подобный DBE, по-видимому, полностью отсутствует у Cyanothece sp. ATCC 51142, что, вероятно, означает, что ферменты, ответственные за превращение гликогена в крахмал (или крахмалоподобный материал), у этих двух организмов различны. Этот вывод во многом согласуется с нашим анализом количества копий и типа изоформ ферментов для хранения полисахаридов, присутствующих в геномах Cyanothece.Это говорит о том, что конвергентная эволюция происходила много раз, чтобы произвести ферменты со сходными свойствами.
Одним из возможных факторов эволюционного давления, которое привело к образованию полуамилопектина и крахмала, могло быть большое событие оксигенации (38). Этот глобальный экологический переход вызвал широкое разнообразие цианобактерий. По мере повышения уровня кислорода в окружающей среде чувствительные к кислороду ферменты нитрогеназы, присутствующие в некоторых азотфиксирующих бактериях, нуждались в защите. Одноклеточным штаммам требовался способ эффективного локального снижения уровня кислорода посредством сильных всплесков дыхания.Один из способов сделать это — разработать более эффективную структуру полисахаридов для хранения, которая максимизирует запасы углеводов. Эта идея поддерживается несколькими способами. Во-первых, исследование шести штаммов Cyanothece выявило тесную связь между частотой дыхания, поддержанием внутриклеточного уровня кислорода и активностью фиксации N 2 (39). Например, штаммы Cyanothece (такие как Cyanothece sp. ATCC 51142), содержащие полуамилопектин, обладали респираторной активностью, которая начиналась в начале темного периода и достигала пика активности нитрогеназы через несколько часов.С другой стороны, такие штаммы, как Cyanothece sp. PCC 7822, которые образуют гликоген, не были так хорошо синхронизированы в отношении фотосинтеза, дыхания или продукции и деградации гликогена. Таким образом, измененный запас углеводов обеспечил множество эволюционных преимуществ с точки зрения более устойчивого роста в неблагоприятных условиях. Во-вторых, мутанты зеленых водорослей с дефектной активностью фермента разветвления изоамилазы вернулись к образованию гликогена вместо крахмала, и, как следствие, их аноксическая гидрогеназная продукция водорода была отменена.И снова продукт хранения углеводов обеспечивает значительные метаболические преимущества.
В этой области остается много открытых вопросов. Например, если Cyanothece sp. АТСС 51142 не содержит DBE, подобного изоамилазе, тогда что делает возможным образование полуамилопектина? Насколько активен белок GlgX2 из Cyanothece sp. PCC 8801 по сравнению с хостом GlgX1? Кроме того, анализ экспрессии на уровне белка и более полная библиотека мутантов могут усилить эти результаты и, возможно, выявить дополнительные возможности для пластичности структуры углеводов в этих организмах.Более того, подробный анализ гипотетических генов и генов, расположенных на внехромосомных плазмидах в штаммах Cyanothece, является оправданным и может выявить потенциальные гидролазы и ферменты, влияющие на структуру хранения полисахаридов. Например, у Cyanothece sp. Имеется 4 потенциальных фермента амилазы. АТСС 51142 (35). Какую роль они играют в образовании полуамилопектина у этого штамма? Наконец, исследования случайного мутагенеза Cyanothece sp. ATCC 51142, аналогичный описанному для штамма Cyanobacterium Clg1 (26), который привел к идентификации генов, ответственных за продукцию амилозы, может быть использован для идентификации неизвестных генов, участвующих в образовании полуамилопектина.
БЛАГОДАРНОСТИ
Эта работа была поддержана грантом программы DOE Genomics: GTL (DE 09-19 PO 2