Аргинин | Химия онлайн
Аргинин – условно незаменимая аминокислота. У взрослого и здорового человека аргинин вырабатывается организмом в достаточном количестве. В то же время, у детей и подростков, у пожилых и больных людей уровень синтеза аргинина часто недостаточен.
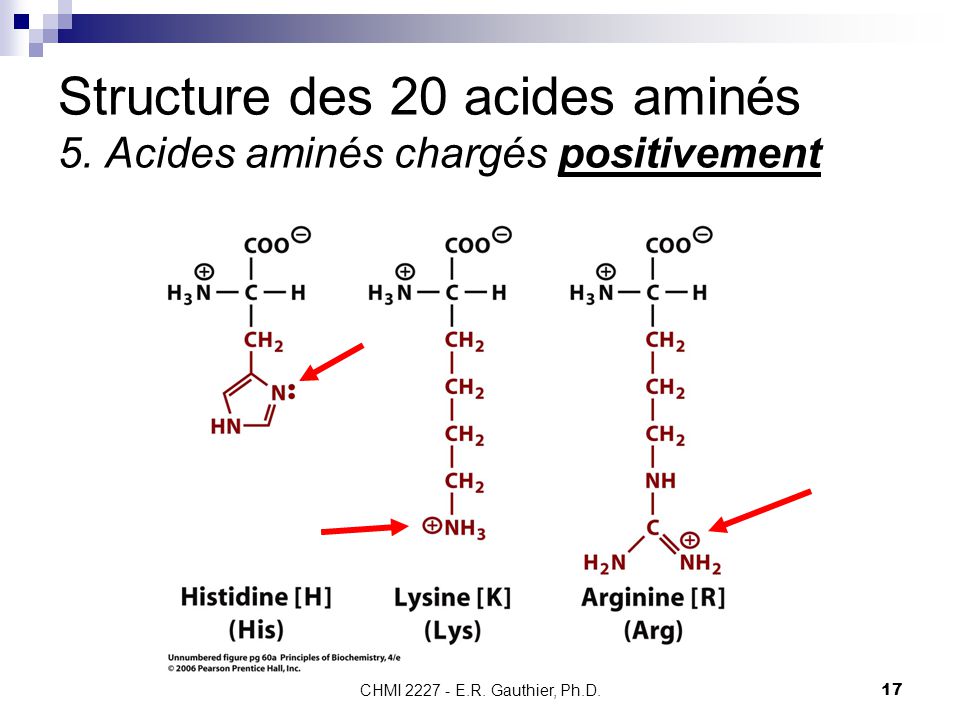
Аргинин – диаминкарбоновая аминокислота, в молекуле которой, помимо аминогруппы, есть амидиновая группа (NH2-C=NH).
Аргинин входит в состав пептидов и белков, особенно высоко содержание аргинина в основных белках – гистонах (обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в пигенетической регуляции таких ядерных процессов, как транскрипция, репликация и репарация) и протаминах (низкомолекулярные белки в ядрах сперматозоидов большинства групп животных) (до 85 %). Встречается аргинин также и в других белках.
Аргинином богаты белки ядер клеток, а также белки растущих тканей (эмбриональная ткань, опухоли).
Аргинин — 2-амино-5-гуанидинпентановая или α-амино-δ-гуанидил-н-валериановая кислота.
Аргинин (Арг, Arg, R) — алифатическая основная α-аминокислота, химическая формула NH-C (NH2) NH (CH2)3CH (NH2) -COOH.
Аргинин впервые выделен в 1886 г. из проростков люпина E. Schulze и E.
Steiger, а структура его установлена E. Schulze и E. Winterstein в 1897 г. Название этой аминокислоты произошло от латинского argentums-серебро, так как аргинин впервые был получен в виде серебряной соли.
Суточная потребность аргинина составляет 6 грамм.
Физиологическая потребность тканей и органов большинства млекопитающих в аргинине удовлетворяется его эндогенным синтезом и/или поступлением с пищей.
Физические свойства
Аргинин представляет собой бесцветные кристаллы, растворимые в воде. Температура плавления аргинина 2440С (с разл.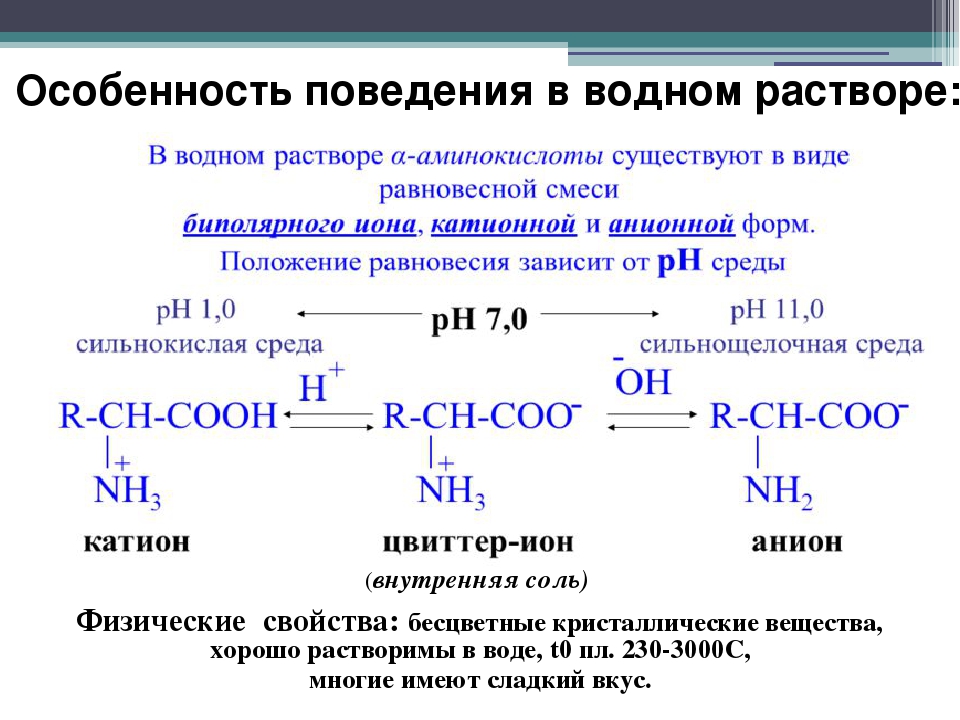 ). Аргинин растворим в воде, не растворяется в эфире. Аргинин имеет основные свойства
). Аргинин растворим в воде, не растворяется в эфире. Аргинин имеет основные свойства
Биологическая роль
Аргинин обладает широким спектром биологических свойств, многофункционален.
Аргинин служит необходимым предшественником для синтеза белков и многих биологически важных молекул, в том числе и пролина. Однако главная роль аргинина в организме человека – быть субстратом для синтеза оксида азота. Поступивший с пищей аргинин всасывается в тонком кишечнике и транспортируется в печень.
Аргинин играет важную роль в процессе образования мочевины.
Аргинин входит в состав многих белков и является одним из предшественников в синтезе креатина, промежуточный продукт синтеза мочевины в печени.
Он способствует детоксикации и выведению аммиака, принимает активное участие в регуляции обмена веществ в организме.
Активизирует процессы регенерации в посттравматическом периоде при заживлении переломов, при ожогах (восстанавливает белковый баланс при тяжѐлых ожогах), при заживлении трофических язв.
Аргинин участвует в процессах образования коллагена, входит в состав пептидного гормона гипофиза вазопрессина. Способствует функционированию вилочковой железы (тимуса), увеличивает еѐ размер и активность.
Аргинин выполняет важные иммунные функции, участвуя в образовании антител, стимулирует выработку Т-лимфоцитов, повышает антибактериальную активность нейтрофилов, повышает содержание гормона роста в крови.
Аргинин снижает уровень жира в организме, способствует синтезу гликогена в печени и мышцах. Увеличивает сперматогенез – участвует в образовании и формировании семенной жидкости (составляет почти 80% от еѐ объѐма), улучшает качество и продолжительность эрекции, усиливает половое влечение у обоих полов, увеличивает частоту и интенсивность оргазмов у женщин.
Аргинин регулирует кровяное давление, обладает противоопухлевой активностью (при переходе аргинина в окись азота (NO) – свободнорадикальное соединение, разрушающее опухлевые клетки). NO тормозит развитие остеопороза и способствует повышению плотности костей.
Аргинин в сочетании с карнитином, способен стимулировать образование гормона роста в человеческом организме: наблюдается уменьшение подкожного жира и увеличение мышечной массы.
Большинство эффектов аргинина связывают с тем, что он является предшественником оксида азота (NО). Молекула NО найдена во всех тканях тела и играет важную роль в функционировании сердечно-сосудистой, иммунной и нервной систем. Пероральное введение аргинина улучшает состояние больных стенокардией.
Аргинин увеличивает секрецию инсулина, стимулирует высвобождение гистамина, который вызывает сосудорасширяющий ответ.
Аргинин усиливает обезвреживание аммиака в печени, способствуя превращению аммиака в мочевину, связывает токсичные ионы аммония, образующиеся при катаболизме белков в печени.
Аргинин нормализует показатели перекисного окисления и антиоксидантной системы. Он проявляет антиоксидантную активность, в результате которой снижается усиление перекисного окисления липидов и повреждающее влияние свободных радикалов на органы.
Природные источники аргинина
Тыквенные семечки, орехи (арахис, миндаль, кедровые, грецкие, фундук), горох, рис нешлифованный, проростки пшеницы, пшеничная и кукурузная мука, мясо (свинина, индейка, курица), рыба (лосось, горбуша), яйца, чечевица, маш, кальмар филе, фасоль белая, молоко, желатин, шоколад.
Чрезмерное потребление аргинина иммунными клетками, которые обычно защищают мозг, является причиной возникновения болезни Альцгеймера.
Области применения
Аргинин — профилактическое средство от физического и умственного переутомления, предотвращает преждевременное старение.
Введение аргинина приводит к расширению сосудов и существенному
снижению артериального давления при гипертонической болезни.
Регулярное назначение аргинина способствует улучшению клинического состояния больных ишемической болезнью сердца.
Применение аргинина эффективно при травмах, ожогах, голоде, стрессах, нарушениях функции печени.
Одним из важных свойств аргинина является способность обеспечивать восстановление организма при астении, которая значительно ухудшает качество жизни больных, приводит к снижению трудоспособности.
Аргинин снижает вязкость крови и уменьшает риск тромбоза сосудов.
При физической нагрузке аргинин способствует улучшению кровотока в коронарных артериях.
Аргинин ускоряет регенерацию печени при ее токсическом поражении и может применяться при лечении цирроза печени и жирового гепатоза.
Аргинин помогает при астматическом бронхите.
Назначение аргинина больным сахарным диабетом 2-го типа существенно повышает чувствительность к инсулину рецепторов клеток организма.
У лиц с повышенной физической нагрузкой аргинин усиливает действие инсулина и увеличивает рост кровеносных капилляров в скелетных мышцах.
Аргинин стимулирует выработку гормона роста, способствует уменьшению жировых отложений, росту мышечной массы, увеличивает активность и физическую выносливость, улучшает настроение.
Аргинин не рекомендуют беременным и кормящим женщинам, больным шизофренией.
Он может стимулировать свободнорадикальные процессы, поэтому его следует принимать в сопровождении антиоксидантов.
Для профилактики и в комплексной терапии сердечнососудистых заболеваний показана биологически активная добавка в форме капсул на основе аргинина. Средство рекомендуется также при холециститах, желчнокаменной болезни, гепатитах, циррозах; после лечения алкоголизма, длительного приема лекарств, больших физических нагрузок. Активизирует иммунитет, оказывает стимулирующее влияние на половую систему мужчин и женщин всех возрастных групп.
Способность аргинина корректировать эндотелиальную дисфункцию сосудов обуславливает его включение в препарат для профилактики атеросклероза, облегчения недомоганий во время менопаузы. Препарат содержит аргинин, лизин, пролин, витамин С, магний, экстракт зеленого чая, N-ацетил-цистеин, Se, Cu, Mn. Средство используют для перорального, внутривенного и парентерального введения.
Аминокислоты
Классификация аминокислот
Аргинин — это… Что такое Аргинин?
Аргинин (2-амино-5-гуанидиновалериановая кислота) — алифатическая основная α-аминокислота. Оптически активна, существует в виде L- и D- изомеров. L-Аргинин входит в состав пептидов и белков, особенно высоко содержание аргинина в основных белках — гистонах и протаминах (до 85%).
Химические свойства
Цвиттер-ион аргинина
Аргинин является основной аминокислотой, несущей два основных центра: аминогруппу в α-положении и гуанидиновую в δ-положении. Гуанидиновая группа благодаря резонансной делокализации заряда при протонировании является сильно основной (pKa 12.48), находится в протонированной катионной форме при pH < 10 и способна образовывать множественные водородные связи. В слабощелочных и нейтральных растворах аргинин образует цвиттер-ион. Высокая основность аргинина и, соответственно, способность образовывать ионные связи с фосфатными группами ДНК, обуславливает образование нуклеопротеидов — комплексов гистон-ДНК хроматина и протамин-ДНК гетерохроматина сперматозоидов.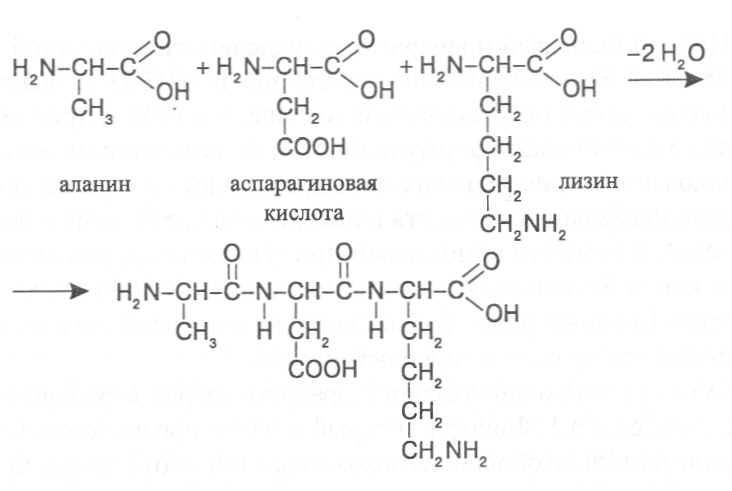
Биологические свойства
Аргинин — условно-незаменимая аминокислота. У взрослого и здорового человека аргинин вырабатывается организмом в достаточном количестве. В то же время, у детей и подростков, у пожилых и больных людей уровень синтеза аргинина часто недостаточен. Биосинтез аргинина осуществляется из цитруллина под действием аргининсукцинатсинтазы и аргининсукцинатлиазы.
Аргинин является одним из ключевых метаболитов в процессах азотистого обмена (орнитиновом цикле млекопитающих и рыб).
Аргинин является субстратом NO-синтаз в синтезе оксида азота NO, являющегося локальным тканевым гормоном с множественными эффектами — от провоспалительного до сосудистых эффектов и стимуляции ангиогенеза.
Содержание в продуктах питания
L-Аргинин встречается во многих продуктах питания, как животного, так и растительного происхождения. В таблице приведены данные по содержанию аргинина на 100 грамм продукта и указано содержание белка.
| Продукт | Белок | Аргинин | А/Б |
|---|---|---|---|
| Свинина сырая | 20,95 г | 1394 мг | 0 6,7% |
| Сырое куриное филе | 21,23 г | 1436 мг | 0 6,8% |
| Сырое филе лосося | 20,42 г | 1221 мг | 0 6,0% |
| Куриное яйцо | 12,57 г | 0 820 мг | 0 6,5% |
| Коровье молоко, 3,7% жирности | 0 3,28 г | 0 119 мг | 0 3,6% |
| Кедровые орехи | 13,69 г | 2413 мг | 17,6% |
| Грецкие орехи | 15,23 г | 2278 мг | 15,0% |
| Тыквенные семечки | 30,23 г | 5353 мг | 17,7% |
| Пшеничная мука г/п | 13,70 г | 0 642 мг | 0 4,7% |
| Кукурузная мука | 0 6,93 г | 0 345 мг | 0 5,0% |
| Рис нешлифованный | 0 7,94 г | 0 602 мг | 0 7,6% |
| Гречишный хлеб | 13,25 г | 0 982 мг | 0 7,4% |
| Горох сушеный | 24,55 г | 2188 мг | 0 8,9% |
Применение
Лекарственные средства
Аргинин присутствует в рецептуре гепатопротекторов, иммуномодуляторов, кардиологических препаратов, лекарственных препаратов для ожоговых больных, больных ВИЧ/СПИД, а также в рецептурах средств для парентерального питания в послеоперационный период. В последнее время лекарства с аргинином появились в геронтологии и онкологии.
В последнее время лекарства с аргинином появились в геронтологии и онкологии.
Проводится тестирование L-аргинина в качестве средства терапии инсультоподобных эпизодов при митохондриальном заболевании — синдроме MELAS.[1][2][3][4]
Пищевые добавки
Аргинин широко рекламируется как компонент БАД для бодибилдеров и спортсменов- тяжёлоатлетов с целью улучшения питания мышц, и пожилых людей для улучшения эректильной функции. Так же массово применяется аргинин и в пищевых добавках, рекламируемых «в целях стимулирования иммунитета». Эффективность аргинина в бодибилдинге опровергнута в нескольких исследованиях.[источник не указан 175 дней] Аргинин является донатором оксида азота, открытие биологических эффектов которого было удостоено Нобелевской премии в медицине.[5]
Примечания
- ↑ Koga Y, Akita Y, Junko N, Yatsuga S, Povalko N, Fukiyama R, Ishii M, Matsuishi T (June 2006).
 «Endothelial dysfunction in MELAS improved by l-arginine supplementation». Neurology 66 (11): 1766–9. DOI:10.1212/01.wnl.0000220197.36849.1e. PMID 16769961.
«Endothelial dysfunction in MELAS improved by l-arginine supplementation». Neurology 66 (11): 1766–9. DOI:10.1212/01.wnl.0000220197.36849.1e. PMID 16769961. - ↑ Koga Y (November 2008). «[L-arginine therapy on MELAS]» (Japanese). Rinsho Shinkeigaku 48 (11): 1010–2. PMID 19198147.
- ↑ Koga Y, Akita Y, Nishioka J, Yatsuga S, Povalko N, Katayama K, Matsuishi T (2007). «MELAS and L-arginine therapy». Mitochondrion 7 (1-2): 133–9. DOI:10.1016/j.mito.2006.11.006. PMID 17276739.
- ↑ Finsterer J (November 2009). «Management of mitochondrial stroke-like-episodes». Eur. J. Neurol. 16 (11): 1178–84. DOI:10.1111/j.1468-1331.2009.02789.x. PMID 19780807.
- ↑ Аргинин в медицине
Аминокислота L-аргинин для птиц | МЕГАМИКС
Кормовая компания Мегамикс
Контакты:
Адрес: ул. Б.Грузинская, д. 61, стр.2
123056
г. Москва
Москва
Телефон: (495) 123-34-45
Электронная почта: [email protected]
55.772386,37.584479
Адрес: п. Первомайский, промышленная зона
040706
Республика Казахстан, Алматинская обл.
Телефон: +7 (727) 299-39-99
Электронная почта: [email protected]
44.800584,78.1726
Адрес: ул.Городецкая 38А, офис 16
220125
Республика Беларусь, г. Минск
Телефон: +7 (017) 361-60-61, 361-60-62
Электронная почта: [email protected]
53.78897,27.977427
Адрес: Гипрозем 16
734067
Республика Таджикистан, г.Душанбе
Телефон: +9 (22) 372-31-08-63
Электронная почта: [email protected]
41.285265,69.309687
Адрес: ул. Фаргона йули, 23
100005
Республика Узбекистан, г.Ташкент
Телефон: +998 (71) 291-62-49
Электронная почта: info@vetteria. ru
ru
41.285265,69.309687
Адрес: ул.Добролюбова, 53/4 офис35
г. Ставрополь
Телефон: +7(8652)99-70-17
Электронная почта: [email protected]
45.037088,41.990607
Адрес: пер. Почтовый, д. 9
460000
г. Оренбург
Телефон: +7 (8442) 97-97-97 доб. 181
Электронная почта: [email protected]
51.760596,55.108337
Адрес: ул.Нальчикское шоссе,13
Ставропольский край, Пятигорск
Телефон: +7-926-029-79-00
Электронная почта: [email protected]
44.00935,43.104312
Адрес: Ракитянский р-он, ул. Пролетарская, д. 2А.
309310
Белгородская обл., п. Ракитное
Телефон: +7 (8442) 97- 97- 97 доб. 496
Электронная почта: [email protected]
50.834087,35.834156
Адрес: ул. Куйбышева, 1
Челябинская область, г. Коркино
Коркино
Телефон: +7 (8442) 97-97-97 доб. 491
Электронная почта: [email protected]
54.900808,61.396526
Адрес: ул. Дорожная, 5г
399540
Липецкая область, с. Тербуны
Телефон: +7 (8442) 97-97-97 доб.432
Электронная почта: [email protected]
52.123517,38.273675
Адрес: пос. Новофедоровское, д.Кузнецово, а/д «Украина», 60 км
108805
г. Москва
Телефон: +7 (495)122-23-70
Электронная почта: [email protected]
55.454195,36.949652
Адрес: пл. А.Невского, д. 2, БЦ Москва, оф. 1108
191167
г. Санкт-Петербург
Телефон: +7 (8442) 97-97-97 доб. 172
Электронная почта: [email protected]
59.924697,30.386157
Адрес: ул. Хрустальная, д. 107, оф.1
400123
г. Волгоград
Телефон: (8442) 97-97-97
Электронная почта: info@megamix. ru
ru
48.793832,44.534699
особенности применения для профилактики и лечения различных заболеваний
Аргинин — аминокислота, синтезирующаяся в организме. Пероральный аргинин использовался при различных состояниях, таких как гипертония, стенокардия, атеросклероз, мигрень и эректильная дисфункция. Считается, что его сосудорасширяющие свойства ответственны за положительный эффект. Аргинин также используется для улучшения заживления ран, улучшения иммунной функции и улучшения спортивных результатов. Некоторые исследования подтверждают использование аргинина при заболеваниях коронарных и периферических артерий (10) (11) (12) (13), но при длительном приеме добавок ухудшилось. ПОДКЛАДКА (14). Кроме того, пероральный прием добавок у пациентов с острым инфарктом миокарда не улучшил фракцию выброса или жесткость сосудов и может быть связан с более высокой смертностью (38). Мета-анализ добавок аргинина по маркерам сердечно-сосудистых заболеваний, ожирения или диабета также не дал положительных результатов, за исключением, возможно, избранной группы пациентов (39).
Менее масштабные исследования показывают, что добавление аргинина, глутамина и HMB может улучшить функцию эндотелия сосудов у пожилых людей (33), но добавление аргинина само по себе не улучшило кровоток или работоспособность мышц у пожилых женщин (34).
Наряду с витаминами-антиоксидантами аргинин снижал частоту преэклампсии у женщин с высоким риском (28), но добавки аргинина не улучшали артериальное давление или функцию почек у женщин с преэклампсией (15). Предварительные данные свидетельствуют о потенциальной пользе добавок аргинина при эректильной дисфункции (16) (40). Аргинин в сочетании с ибупрофеном может усилить обезболивание у пациентов с мигренью (17). Энтеральный аргинин снижает шок у пациентов с тяжелыми ожогами (4) и может быть полезен в качестве дополнительной терапии у пациентов с активным туберкулезом (29). У онкологических больных предварительные результаты смешаны для периоперационных энтеральных смесей, обогащенных аргинином для улучшения заживления ран (1) ( 41) и иммунной функции (27) (35) (42), но другие исследования показывают, что такие формулы могут уменьшить осложнения и сократить продолжительность пребывания в больнице (30) (36). Некоторые данные предполагают, что профилактическая добавка, содержащая аргинин, может снизить частоту синдрома кистей и стоп у пациентов с гепатоцеллюлярной карциномой, принимающих сорафениб (43).
Некоторые данные предполагают, что профилактическая добавка, содержащая аргинин, может снизить частоту синдрома кистей и стоп у пациентов с гепатоцеллюлярной карциномой, принимающих сорафениб (43).
Интересно, что лечение, основанное на депривации аргинина, также рассматривается как потенциальное лечение онкологических заболеваний (31) (32) (37).
Хотя метаанализ не выявил значительного влияния добавок аргинина на воспалительные биомаркеры, анализ подгрупп предполагает, что он может увеличить циркулирующий С-реактивный белок у онкологических больных, у пациентов старше 60 лет или с более высокими исходными уровнями СРБ, или при использовании энтеральных формул. (44). Необходимы дополнительные исследования, чтобы определить обстоятельства, при которых добавление аргинина может быть безопасным и эффективным.
Что такое L-аргинин? Польза, противопоказания, способы применения
Сейчас не только спортсменам,
но и людям, которые занимаются физическими упражнениями, посещают спортзал и
фитнес, ведут здоровый образ жизни, нужно часто, обильно и правильно питаться. Полноценное
Полноценное
питание поможет обеспечить организм всеми необходимыми питательными веществами.
Именно поэтому люди, которые занимаются спортом, интенсивными физическими нагрузками,
должны наполнять свой рацион не только фундаментальными макроэлементами, но и аминокислотами
в чистом виде. Предлагаем вместе с нами, командой Интернет-магазина ПРОТЕИН-СПОРТ разобраться в том, что
такое аргинин, зачем он нужен спортсменам, как принимать аргинин и кому он противопоказан. Для начала выясним, что же
это за вещество и почему его рекомендуют принимать физически активным людям.
L-аргинин что
это? Свойства аргинина
Начнём с определения.
Аргинин это одна из основных аминокислот, необходимых организму человека для азотистого
обмена. Благодаря ему клетки снабжаются питательными веществами, нормализуется
тонус кровеносных сосудов человека, из организма удаляются продукты распада
белков. Может ли организм человека сам синтезировать аргинин? Да, может. Но
когда речь идёт о спортсменах, которые тренируются систематически и интенсивно,
выработка этой аминокислоты организмом снижается. Тогда восполнить недостаток
Тогда восполнить недостаток
этого вещества в организме спортсмена лучше всего при помощи добавки.
На сегодняшний день научно
доказана польза аргинина, он используется в разных сферах:
- спорт
и фитнес; - медицина;
- диетология;
- косметология.
Нас интересует конкретно польза и побочные действия L-аргинина для спортсменов, а также людей, которые занимаются физическими нагрузками
разного уровня сложности.
Давайте посмотрим,
какими свойствами обладают добавки, в основе которых – аргинин:
- мышечные волокна обеспечиваются
кислородом; - рост мышечной массы стимулируется благодаря
интенсивному кровоснабжению; - происходит нормализация артериального
давления; - уменьшается риск образования тромбов;
- происходит ускоренная регенерация и восстановление
тканей; - улучшается выделительная функция почек;
- повышается иммунитет;
- улучшается потенция;
- улучшается память;
- происходит регуляция синтеза, а также
секреции многих гормонов, в том числе инсулина.
Таким образом Вы видите, сколько полезных функций
исполняет в нашем организме аргинин. Благодаря чему происходит интенсивное
кровоснабжение, которое способствует росту мышц? Если говорить простыми
словами, то в организме NO-синтазы (так называется группа ферментов) ускоряют процесс получения
оксида азота из аргинина. Это, в свою очередь, приводит в тонус артериальные
сосуды, стимулирует у спортсмена кровообращение, а также позволяет более
эффективно транспортировать к мышцам креатин. Это те основные свойства
аминокислоты, о которых должен знать каждый физически активный человек, который
заботится о своём здоровье.
Как
принимать Л-аргинин?
Производители
предлагают эту добавку в форме капсул, таблеток и порошка. Вы можете выбрать
тот вариант, который Вам будет удобнее принимать в течение суток. Как правильно принимать аргинин? Начинать желательно с минимального
количества – 1 г аргинина в сутки, постепенно можете увеличить дозу до 3-х
граммов этой аминокислоты. Суточная норма для
современных спортсменов – это 3 грамма аминокислоты, хотя некоторые спортсмены
увеличивают эту дозу до 5-6 граммов в сутки.
Часто задают вопрос: когда
принимать аргинин? Спортсменам рекомендуют принимать эту добавку за 30-60 минут
до начала тренировки, а также после пробуждения и перед сном. То есть,
оптимальный вариант приёма – 1-3 раза в сутки. Следуйте
инструкции по применению аргинина, указанной производителем.
Можно ли получить
аргинин из продуктов питания? Да, можно. На рисунке Вы можете увидеть основные
натуральные источники этой аминокислоты. Опять-таки, если Вы интенсивно
занимаетесь спортом, то этих продуктов питания будет недостаточно, и лучше
восполнить недостаток этой аминокислоты при помощи добавок от лучших
производителей.
Польза
аргинина и противопоказания
О пользе этой
аминокислоты мы уже говорили выше, она оказывает положительное воздействие и на
сердечно-сосудистую систему, и на мышечную систему, и на иммунитет, и на
выделительную систему каждого человека. Что касается противопоказаний
L-аргинина, то добавку нежелательно принимать беременным
и кормящим женщинам, детям. Может также наблюдаться индивидуальная
Может также наблюдаться индивидуальная
непереносимость такой добавки, тогда дозу можно уменьшить или попробовать
добавку от другого производителя, в крайнем случае – отказаться от её приёма.
Что
предлагает наш магазин?
В ассортименте нашего
Интернет-магазина PROTEIN—SPORT есть большой выбор аминокислот, в том числе и L-аргинина. Мы предлагаем только самые качественные аминокислоты от ведущих производителей:
- BioTech;
- Pro Nutrition;
- GNC;
- Form Labs;
- AllMax Nutrition;
- Olimp и др.
Кроме широкого
ассортимента спортивного питания, мы предлагаем также аксессуары, пояса и
перчатки, которые понадобятся при занятиях в спортивном зале, на фитнесе. Доставку
делаем по всей территории Украины. Наши сотрудники всегда готовы
проконсультировать по поводу выбора спортивного питания, а цены в магазине
смогут приятно Вас удивить!
Аргинин. Суточная норма. Недостаток аргинина
Аргинин (2-амино-5-гуанидинпентановая кислота L-Arginine) — это условно незаменимая аминокислота. Встречается в белковой пище, которую организм использует для выработки оксида азота. Это соединение является очень важным для жизни организма, которое участвует во всех обменных процессах.
Встречается в белковой пище, которую организм использует для выработки оксида азота. Это соединение является очень важным для жизни организма, которое участвует во всех обменных процессах.
Выработка окиси азота без аргинина невозможна. Аргинин частично может вырабатываться нашим организмом самостоятельно, но в недостаточном количестве. При малейшей патологии наш организм не успевает справляться с синтезом этой условно незаменимой аминокислоты. У детей, подростков, больных и пожилых людей синтез этой важной аминокислоты в организме незначителен или вовсе отсутствует. Поэтому обеспечить дополнительное поступление аргинина в организм можно с помощью пищи и специальных добавок. А также немаловажно знать, сколько должна составлять суточная потребность аргинина для человека.
Суточная потребность в аргинине
Согласно нормам, которые установили специалисты, суточная потребность в аргинине для взрослых людей будет составлять до 6,0 г, а для детей до 4,0 г. Что касается бодибилдеров, то для того, чтобы набрать мышечную массу, достаточно употреблять 3-9 г. в сутки, но начинать необходимо с минимальной дозы. При заболеваниях печени аргинин рекомендуется принимать в небольших дозах. Аргинин можно принимать также в комплексе с другими аминокислотами, например, такими как лизин, валин, глутамин.
в сутки, но начинать необходимо с минимальной дозы. При заболеваниях печени аргинин рекомендуется принимать в небольших дозах. Аргинин можно принимать также в комплексе с другими аминокислотами, например, такими как лизин, валин, глутамин.
В любом случае за более точной информацией по данному вопросу стоит обратиться к специалисту, чтобы избавить себя и своих близких от нежелательных последствий недостатка или избытка в организме аргинина.
Последствия недостатка аргинина в организме
При недостатке аргинина в нашем организме происходят серьезные изменения и последствия могут быть не очень приятными. При недостатке аргинина организм ослабляется в следствии чего возрастает риск развития тромбов, идет нарушение мозговой деятельности, раннее старение, ожирение, инфаркт, атеросклероз, гипертония, замедление роста организма и даже образование раковых опухолей тяжелых форм. Неприятными последствиями могут быть не только при недостатке аргинина в организме, но и при его избытке.
Последствия избытка аргинина в организме
При избытке аргинина в организме могут произойти проблемы с кожей, аллергические реакции, крапивница, раздражительность, агрессия, тошнота, диарея, снижение артериального давления. За всем эти просто необходимо следить и тогда вы испытаете на себе лишь полезные свойства аргинина без какого-либо негативного воздействия на организм.
Полезные свойства аргинина
Как уже было сказано выше, аргинин участвует во всех обменных процессах. Он нормализует их и без него не могут правильно функционировать нервная и иммунная системы. Еще следует обратить внимание на его участие в выработке гормонов и ферментов.
Особенное место этой аминокислоте отводится и в мужском организме. Мужской тестостерон также образуется благодаря аргинину. Данная условно незаменимая аминокислота увеличивает количество семенной жидкости, повышает сексуальную активность, продлевает половой акт. Поэтому при ежедневном употреблении аргинина можно избавиться от многих сексуальных проблем.
Аргинин зарекомендовал себя как элемент питания для спортсменов, бодибилдеров. Ведь эта аминокислота содействует развитию мышечной массы, вместе с тем снижая жиры в организме. Без аргинина не вырастут ни бицепсы, ни трицепсы.
А представительницы женского пола, своими отзывами отметили то, что после приема добавки у них улучшилось общее состояние и внешность. Так что те, кто действительно беспокоятся за свою кожу, должны знать то, что аргинин способен улучшать ее состояние, очищать и улучшать ее цвет. Также он придется по душе людям страдающим лишним весом. На женщин он воздействует так, что они становятся стройными, спортивными не приобретая лишней мускулатуры, при этом дополняя все это двигательной активностью и правильным рационом.
Результаты проведенных исследований с позиции медицины свидетельствуют об эффективности применения аргинина при таких состояниях как ишемическая болезнь сердца, хроническая сердечная недостаточность. Он регулирует сердечную деятельность, снижает уровень инсулина, предотвращает образование тромбов. Помимо этого аргинин очищает печень от ядов. Аргинин используют для лечения вируса иммунодефицита человека (ВИЧ) и синдрома приобретенного иммунного дефицита (СПИД). Воздействуя на вилочковую железу, он способен повышать уровень гранулярных лимфоцитов (NK) и Т-клеток в крови. Он замедляет развитие опухолей. Если включить аргинин в рацион питания во время лечения, он будет способствовать сохранению иммунных функций. Также не менее главным его достоинством является влияние на ускорение регенерации поврежденных тканей. В течение последних лет все эти доказательства воздействия аргинина на организм человека свидетельствуют о возможности действенного и безопасного применения аминокислоты при различной патологии.
Благодаря пользе, аминокислоту необходимо принимать тому, кто серьезно заботится о своей красоте и долголетии, так как от ее влияния зависит качественно здоровая и продолжительная жизнь. Но стоит учесть и тот момент, что аргинин имеет противопоказания и это в основном касается пищевых добавок, в которых он содержится.
Противопоказания аргинина
Как уже известно, эта аминокислота практически безопасна, она вырабатывается организмом и поступает к нам вместе с едой. Но дополнительно в качестве пищевых добавок ее не рекомендуется принимать людям с вирусными инфекциями и при индивидуальной непереносимости. Людям с шизофренией или иными психическими заболеваниями, беременным и кормящим женщинам также стоит воздержаться от употребления аргинина в виде пищевой добавки. С осторожностью его назначают при патологиях почек, отсутствии мочеиспускания (анурии). Перед началом курса не лишним будет посетить врача. Необходимо понимать, что прием аргинина принесет пользу в том случае, если его употреблять в разумных дозах.
Людям, которые страдают определенными болезнями, помимо приема добавок, не мешало бы увеличить потребление продуктов, содержащих эту важную для жизнедеятельности человека аминокислоту.
Продукты питания богатые аргинином
Самыми основными источниками аргинина являются продукты питания растительного и животного происхождения, в которых есть белок. В больших количествах его можно встретить в мясе, это относится к таким продуктам как свинина, говядина и куриное филе. Аргинин есть в молоке и твороге. Эти продукты, как и мясо должны присутствовать не только во взрослом, но и в детском рационе ежедневно. Семечки тыквы, кунжут, миндаль, грецкие орехи и кедровые орехи также являются источниками значительного количества аргинина. Присутствует аминокислота аргинин и в лососе, тунце, молоках рыб, улитках, креветках, а также в соевых бобах, сушеном горохе, буром рисе и куриных яйцах.
Многим любителям сладкого будет приятно узнать о том, что аминокислота аргинин присутствует в шоколаде, десертах и продуктах питания на основе желатина.
Не менее важно знать не только в каких продуктах питания есть аргинин, но и какое влияние оказывает процесс приготовления на его содержание.
Влияние процесса приготовления продуктов питания на содержание аргинина
Не стоит забывать о том, что в процессе приготовления продуктов питания количество аргинина меняется. Так, например, в сыром мясе аргинина приблизительно на 40% меньше, чем жареном виде, а в тушеном еще больше. В куриных сырых и вареных яйцах его количество одинаковое, но в жареном виде немного больше. Если это касается варки морепродуктов, то его количество возрастает на 10%, а при запекании значительно снижается.
L- Аргинин 1000 мг — состав, свойства, описание, применение
L-Аргинин — это аминокислота, которая является необходимой для того чтобы организм, а в частности именно мышечная система организма, были здоровыми и находились в тонусе. Она является незаменимой, особенно её нехватка ощущается у людей пожилого возраста.
Особенно важен этот препарат для спортсменов и людей, которые испытывают активные физические нагрузки. Поскольку эта аминокислота способствует расширению кровеносных сосудов, то это усиливает приток крови к мышцам. А это в свою очередь помогает увеличить выносливость. Происходит это за счет улучшения работы мышц.
L-Аргинин: состав и форма выпуска
В состав входит один активный компонент — L-аргинин 1000 мг.
Выпускается препарат в форме таблеток во флаконе зеленого цвета. В одной упаковке находится 50 штук.
L-Аргинин: свойства
Входящий в состав белка аргинин, является аминокислотой, которая так необходима для улучшения работы мышц. Без выработки аргинина значительно ухудшается общее самочувствие и снижается уровень здоровья. Чтобы избежать таких неприятных последствий необходимо принимать его извне. Особенно это становится актуальным для людей более старшего возраста, так как с возрастом уменьшается способность вырабатывать аргинин.
Кроме того L-аргинин способствует увеличению притока крови к мышцам, позволяя им активно работать и быть всегда в тонусе. Особенно важно это для людей, занимающихся активно спортом или переносящие большие физические нагрузки.
Также прием препарата положительно сказывается на половой системе мужчин, усиливая приток крови к половым органам и как следствие улучшая потенция и эректильную функцию.
L-Аргинин: показания к применению и противопоказания
Прием L-аргинина назначают при следующих ситуациях:
— сильные физические нагрузки, в том числе при занятиях спортом,
— ухудшения кровообращения в мышцах,
— заболевания печени (гепатит, цирроз),
— ожирение,
— стенокардия,
— гипертония,
— ВИЧ-заболевания,
— нарушение репродуктивной функции у мужчин.
Препарат не рекомендуется к применению при индивидуальной непереносимости препарата, а также при беременности и периоде кормления молоком рекомендуется посоветоваться со специалистом.
L-Аргинин: инструкция по применению
Препарат назначают по 1 таблетке 3 раза в день во время приема пищи. Длительность курса составляет 1 месяц.
L-Аргинин: цена и продажа
Купить БАД L-Аргинин Вы можете на нашем сайте, поместив товар в корзину и оформив заказ или же позвонив по телефону нашим менеджерам. Они проконсультируют по всем интересующим вопросам и помогут всё оформить.
Как только заказ будет оформлен и подтвержден, он будет передан в службу доставки и в самые кратчайшие сроки доедет к Вам.
Для регионов действует бесплатный номер 8 800 550 52 96
Взаимодействие с боковыми цепями аргинина и роль аргинина в качестве носителя заряда в чувствительных к напряжению ионных каналах
Harms, M. J. et al. Скрытый лизин, который титруется нормальным pK (a): роль конформационной гибкости на границе раздела белок-вода как детерминанта значений pK (a). Protein Sci. 17, 833–845 (2008).
CAS
PubMed
PubMed Central
Google ученый
Fitch, C. A., Platzer, G., Окон, М., Гарсия-Морено, Б. и Макинтош, Л. П. Аргинин: пересмотр его значения pK (a). Protein Sci. 24. С. 752–761 (2015).
CAS
PubMed
PubMed Central
Google ученый
Хармс, М. Дж., Шлессман, Дж. Л., Сью, Г. Р. и Гарсия-Морено, Б. Остатки аргинина во внутренних положениях в белке всегда заряжены. Proc. Natl. Акад. Sci. USA 108, 18954–18959 (2011).
ADS
CAS
PubMed
Google ученый
Li, L.Б., Воробьев И. и Аллен Т. В. Различные взаимодействия боковых цепей лизина и аргинина с липидными мембранами. J. Phys. Chem. В 117, 11906–11920 (2013).
CAS
PubMed
PubMed Central
Google ученый
Catterall, W. A. Датчики напряжения с ионным каналом: структура, функции и патофизиология. Нейрон 67, 915–928 (2010).
CAS
PubMed
PubMed Central
Google ученый
Тао, Х., Ли, А., Лимапичат, В., Догерти, Д. А., Маккиннон, Р. Стробирующий центр передачи заряда в датчиках напряжения. Science 328, 67–73 (2010).
ADS
CAS
PubMed
PubMed Central
Google ученый
Campos, F. V., Chanda, B., Roux, B. и Bezanilla, F. Два атомных ограничения однозначно позиционируют сегмент S4 относительно сегментов S1 и S2 в закрытом состоянии канала Shaker K. Proc. Natl. Акад. Sci. USA 104, 7904–7909 (2007).
ADS
CAS
PubMed
Google ученый
Li, Q. F. et al. Структурный механизм зависящего от напряжения стробирования в изолированной области измерения напряжения. Nat. Struct. Мол. Биол. 21. С. 244–252 (2014).
CAS
PubMed
PubMed Central
Google ученый
Аггарвал, С. К. и Маккиннон, Р. Вклад сегмента S4 в стробирующий заряд в канале Shaker K +.Нейрон 16, 1169–1177 (1996).
CAS
PubMed
Google ученый
Ахерн, К. А. и Хорн, Р. Сфокусированное электрическое поле на датчике напряжения калиевых каналов. Нейрон 48, 25–29 (2005).
CAS
PubMed
Google ученый
Pless, S.A. et al. Асимметричный функциональный вклад кислотных и ароматических боковых цепей в доменах датчиков напряжения натриевых каналов.J. Gen. Physiol. 143. С. 645–656 (2014).
CAS
PubMed
PubMed Central
Google ученый
Коленсо, К. К., Цао, Ю., Сешнс, Р. Б., Хэнкокс, Дж. К. и Демпси, К. Э. Перенос стробирующего заряда датчика напряжения в модели калиевого канала hERG. Биофиз. J. 107, L25 – L28 (2014).
CAS
PubMed
PubMed Central
Google ученый
Мейсон, П.Е., Нейлсон, Г. В., Демпси, К. Е., Барнс, А. С. и Крукшенк, Дж. М. Гидратная структура гуанидиния и тиоцианат-ионов: влияние на стабильность белка в водном растворе. Proc. Natl. Акад. Sci. USA 100, 4557–4561 (2003).
ADS
CAS
PubMed
Google ученый
Schwaiger, C. S., Bjelkmar, P., Hess, B. & Lindahl, E. Конформация 3 (10) -Helix облегчает переход сегмента S4 датчика напряжения в неактивное состояние.Биофиз. J. 100, 1446–1454 (2011).
ADS
CAS
PubMed
PubMed Central
Google ученый
Jensen, M.O. et al. Механизм стробирования напряжения в калиевых каналах. Science 336. С. 229–233 (2012).
ADS
CAS
PubMed
Google ученый
Delemotte, L., Tarek, M., Klein, M. L., Amaral, C. & Treptow, W. Промежуточные состояния Kv1.2 датчика напряжения из моделирования атомистической молекулярной динамики. Proc. Natl. Акад. Sci. USA 108, 6109–6114 (2011).
ADS
CAS
PubMed
Google ученый
Фрейтес, Дж. А., Шоу, Э. В., Уайт, С. Х. и Тобиас, Д. Дж. Микроскопическое происхождение флуктуаций тока затвора в датчике напряжения калиевого канала. Биофиз. J. 102, A44 – A46 (2012).
Google ученый
Танки, Н., Торнтон, Дж. М. и Гудфеллоу, Дж. М. Распределение воды вокруг аминокислотных остатков в белках. J. Mol. Биол. 202, 637–657 (1988).
CAS
PubMed
Google ученый
Галливан, Дж. П. и Догерти, Д. А. Катион-пи-взаимодействия в структурной биологии. Proc. Natl. Акад. Sci. USA 96, 9459–9464 (1999).
ADS
CAS
PubMed
Google ученый
Сингх, Дж.И Торнтон, Дж. М. Атлас взаимодействий белковых боковых цепей (IRL Press, Oxford, 1992).
Флокко, М. и Моубрей, С. Л. Взаимодействия аргинина и ароматических боковых цепей в белках при плоском укладывании. J. Mol. Биол. 235, 709–717 (1994).
CAS
PubMed
Google ученый
Дональд, Дж. Э., Кулп, Д. В. и ДеГрадо, В. Ф. Солевые мосты: геометрически определенные, моделируемые взаимодействия. Proteins 79, 898–915 (2011).
CAS
PubMed
PubMed Central
Google ученый
Такешита К. и др. Рентгеновская кристаллическая структура потенциалзависимого протонного канала. Nat. Struct. Мол. Биол. 21, 352 – U170 (2014).
CAS
PubMed
Google ученый
Имаи, Ю. Н., Иноуэ, Ю. и Ямамото, Ю. Склонности полярных и ароматических аминокислот в неканонических взаимодействиях: анализ несвязанных контактов комплексов белок-лиганд в кристаллических структурах.J. Med. Chem. 50, 1189–1196 (2007).
CAS
PubMed
Google ученый
Мейсон П. Э., Демпси К. Э., Нейлсон Г. У., Клайн С. Р. и Брэди Дж. У. Предпочтительные взаимодействия ионов гуанидина с ароматическими группами по сравнению с алифатическими группами. Варенье. Chem. Soc. 131, 16689–16696 (2009).
CAS
PubMed
PubMed Central
Google ученый
Чакрабарти, П.И Бхаттачарья, Р. Геометрия несвязанных взаимодействий с участием плоских групп в белках. Прог. Биофиз. Мол. Биол. 2007. Т. 95. С. 83–137.
CAS
PubMed
Google ученый
Демпси, К. Э., Пиггот, Т. Дж. И Мейсон, П. Е. Диссекция вкладов в денатурантную чувствительность белков. Биохимия 44, 775–781 (2005).
CAS
PubMed
Google ученый
Лим, W.K., Rosgen, J. & Englander, S. W. Мочевина, но не гуанидин, дестабилизирует белки, образуя водородные связи с пептидной группой. Proc. Natl. Акад. Sci. USA 106, 2595–2600 (2009).
ADS
CAS
PubMed
Google ученый
Рейф М. М., Хуненбергер П. Х. и Остенбринк К. Новые параметры взаимодействия заряженных боковых цепей аминокислот в силовом поле ГРОМОС. J. Chem. Теория вычисл. 8, 3705–3723 (2012).
CAS
PubMed
Google ученый
Дебец, К.Т., Гроненборн А. М. и Чонг Л. Т. Оценка прочности солевых мостиков: сравнение современных биомолекулярных силовых полей. J. Phys. Chem. В 118, 6561–6569 (2014).
CAS
PubMed
PubMed Central
Google ученый
Лакруа, Дж. Дж. И Безанилла, Ф. Управление переходом конечного стробирующего заряда с помощью гидрофобного остатка в сегменте S2 датчика напряжения канала K +. Proc. Natl. Акад. Sci. USA 108, 6444–6449 (2011).
ADS
CAS
PubMed
Google ученый
Cheng, Y. M. et al. Функциональное взаимодействие зарядов датчика напряжения с гидрофобной пробкой S2 в каналах hERG. J. Gen. Physiol. 142, 289–303 (2013).
CAS
PubMed
PubMed Central
Google ученый
Лакруа, Дж. Дж., Хайд, Х. К., Кампос, Ф. В. и Безанилла, Ф. Перемещение стробирующих зарядов через стробирующую пору в датчике напряжения Kv-канала.Proc. Natl. Акад. Sci. USA 111, E1950 – E1959 (2014).
CAS
PubMed
Google ученый
Эль-Дин, Т.М.Г., Шойер, Т. и Каттералл, В.А. Отслеживание движения S4 с помощью стробирования поровых токов в бактериальном натриевом канале NaChBac. J. Gen. Physiol. 144. С. 147–157 (2014).
Google ученый
Лонг, С. Б., Тао, X., Кэмпбелл, Э. Б. и Маккиннон, Р. Атомная структура потенциал-зависимого K + канала в липидной мембраноподобной среде.Nature 450, 376–382 (2007).
ADS
CAS
PubMed
Google ученый
Томбола, Ф., Патак, М. М. и Исакофф, Э. Ю. Аргинины, чувствительные к напряжению, в пермеате калиевого канала и закупоривают катион-селективные поры. Нейрон 45, 379–388 (2005).
CAS
PubMed
Google ученый
Miceli, F. et al. Чувствительный к напряжению домен каналов K (v) 7.2 как молекулярная мишень для мутаций, вызывающих эпилепсию, и противосудорожных средств.Передний. Pharmacol. 2, 2 (2011).
CAS
PubMed
PubMed Central
Google ученый
Мейсон П. Э., Брэди Дж. У., Нейлсон Г. У. и Демпси К. Э. Взаимодействие ионов гуанидиния с модельным пептидом. Биофиз. J. 93, L4 – L6 (2007).
Google ученый
Wang, G.L. & Dunbrack, R.L. PISCES: сервер отбора белковой последовательности. Биоинформатика 19, 1589–1591 (2003).
CAS
PubMed
Google ученый
Берман, Х. М. и др. Банк данных о белках. Nucleic Acids Res. 28. С. 235–242 (2000).
ADS
CAS
PubMed
PubMed Central
Google ученый
ван дер Уолт, С., Колберт, С. и Варокво, Г. Массив NumPy: структура для эффективных численных вычислений. Comput. Sci. Англ. 13, 22–30 (2011).
Google ученый
Система молекулярной графики PyMOL, версия 1.4.1 Schrödinger, LLC.
Коленсо, К.К., Сешнс, Р. J. Chem. Инф. Мод. 2013. Т. 53. С. 1358–1370.
CAS
Google ученый
Аргинин: не только протеин | Американский журнал клинического питания
РЕФЕРАТ
Аргинин, полусущественная или условно незаменимая аминокислота для человека, является одной из наиболее метаболически универсальных аминокислот и служит предшественником для синтеза мочевины, оксида азота, полиаминов, пролина, глутамата, креатина и агматина.Аргинин метаболизируется сложным и строго регулируемым набором путей, которые остаются не полностью изученными как на уровне всего тела, так и на клеточном уровне. Сложность метаболизма усугубляется тем фактом, что ограниченная доступность аргинина может избирательно влиять на экспрессию определенных генов, большинство из которых сами участвуют в некоторых аспектах метаболизма аргинина. В этом обзоре освещены отдельные аспекты метаболизма аргинина, включая области, в которых наши знания остаются фрагментарными и неполными.
ВВЕДЕНИЕ
Интерес к метаболизму аргинина больше не ограничивается биохимиками и диетологами, о чем свидетельствуют многочисленные веб-сайты и статьи в журналах о здоровье и фитнесе, рекламирующие преимущества добавок аргинина, обычно в отношении улучшения сексуальной функции, но также и в отношении улучшения иммунной функции. и общее состояние здоровья. Утверждения о благоприятных эффектах (часто экстраполированные далеко за пределы того, что можно сделать на основании опубликованных научных исследований) обычно основаны на том факте, что аргинин является предшественником оксида азота (NO), и иногда кажется, что это фактически единственный аспект метаболизма аргинина. о чем знают многие биомедицинские исследователи.Однако этот взгляд, ориентированный на NO, постепенно заменяется более широкой перспективой, поскольку все больше исследователей начинают осознавать сложность и важность метаболизма аргинина. Помимо своей роли в синтезе белков и NO, аргинин является предшественником по крайней мере 6 биологически важных соединений у млекопитающих (рис. 1) и, таким образом, является одной из наиболее метаболически универсальных аминокислот. Хотя эта универсальность предоставила плодородную почву для исследователей во многих областях биомедицинских исследований, она также создает значительные проблемы для разработки комплексного представления о метаболизме аргинина in vivo.
РИСУНОК 1.
Источники и метаболическая судьба аргинина. Путресцин, спермин и спермидин — это полиамины, указанные на этом рисунке. NO, оксид азота.
РИСУНОК 1.
Источники и метаболическая судьба аргинина. Путресцин, спермин и спермидин — это полиамины, указанные на этом рисунке. NO, оксид азота.
Прежде чем продолжить, необходимо вкратце рассказать о заголовке этой статьи. Он был назначен для этой презентации Верноном Янгом, но, к сожалению, у меня не было возможности узнать, что он, возможно, имел в виду с его помощью.Поэтому я позволил себе интерпретировать это как нечто вроде «Что мы знаем о роли аргинина, помимо его участия в структуре и функции белка, и насколько хорошо мы это знаем?» Соответственно, в этой статье будет кратко рассмотрен статус наших знаний по отдельным аспектам метаболизма аргинина и выделены области, в которых наши знания остаются фрагментарными и неполными. Из-за множества ферментов, участвующих в метаболизме аргинина, и множества различных комбинаций, которые экспрессируются в различных типах клеток, регуляция метаболизма аргинина — хотя и чрезвычайно важна — является слишком обширной темой, чтобы обсуждать ее в этом обзоре.Читателя отсылают к предыдущим обзорам, где можно найти обширные списки ссылок и особенности метаболизма аргинина, которые здесь не рассматриваются (1–10).
ИСТОЧНИКИ АРГИНИНА
Источниками свободного аргинина в организме являются диетический белок, эндогенный синтез и обмен белков организма (рис. 1). Около 40% пищевого аргинина катаболизируется в кишечнике, прежде чем он попадает в кровоток (8). Во время голодания ≈85% аргинина, поступающего в кровоток, происходит за счет белкового обмена, а оставшаяся часть — за счет синтеза de novo (8).На уровне всего организма большая часть синтеза аргинина de novo происходит в результате метаболического взаимодействия между тонким кишечником и почками в так называемой кишечно-почечной оси синтеза аргинина (8, 9). Величина эндогенного синтеза настолько велика, что аргинин не является незаменимой аминокислотой для здоровых взрослых. Однако эндогенный синтез аргинина не может полностью удовлетворить потребности младенцев и растущих детей или взрослых в условиях катаболического стресса или с дисфункцией тонкой кишки или почек; Таким образом, аргинин классифицируется как полуосновная или условно незаменимая аминокислота (5, 11).
Используя стабильные изотопы, Вернон Янг и его коллеги показали, что скорость эндогенного синтеза практически не зависит от потребления аргинина (12), что указывает на то, что гомеостаз аргинина достигается главным образом за счет регуляции катаболизма аргинина. Этот вывод подтверждается наблюдением, что циркулирующие концентрации аргинина значительно повышены у мышей, у которых снижена экспрессия аргиназы типа II (13).
De novo синтез аргинина из цитруллина происходит в основном в проксимальных канальцах почек (8).Однако эта способность не только обнаруживается во многих других типах клеток в той или иной степени (14–16), но также может сильно индуцироваться цитокинами и другими агентами, обычно в сочетании с повышенной экспрессией индуцибельной изоформы синтазы оксида азота (iNOS ) (16–18). В таких случаях часть цитруллина, продуцируемого NOS, может быть переработана в аргинин по пути, называемому циклом цитруллин-NO (8, 17).
Хотя это важно для полного понимания метаболизма аргинина, мы мало знаем о предпочтительных источниках (например, эндогенный синтез, обмен белка или захват из внеклеточной жидкости) аргинина (или других субстратов), используемых различными аргининами. метаболические ферменты в любом конкретном типе клеток.Существуют убедительные доказательства того, что аргинин, синтезируемый в цикле мочевины, недоступен для синтеза NO в печени (19) и что аргинин, импортируемый извне клетки, по-видимому, является предпочтительным субстратом для синтеза NO эндотелиальными клетками [явление, известное как парадокс аргинина ( 20, 21)], но эндогенно синтезированный аргинин может быть более важным для синтеза NO с высоким выходом активированными гладкомышечными клетками (22). Еще больше усложняют картину недавние доказательства того, что множественные пулы внутриклеточного аргинина существуют в эндотелиальных клетках, но не в некоторых других типах клеток (23).
МЕТАБОЛИЧЕСКИЕ СУДЬБЫ АРГИНИНА
В отличие от единственного фермента, который производит аргинин, 4 фермента используют аргинин в качестве субстрата: аргининдекарбоксилаза, аргинин: глицин-амидинотрансфераза, аргиназа и NOS (рис. 2). (Хотя аргинил-тРНК-синтетаза также использует аргинин в качестве субстрата, он не считается метаболическим ферментом и поэтому здесь не обсуждается.) Все ферменты, перечисленные на рисунке 2, были клонированы и, по крайней мере, частично охарактеризованы. Аргиназа и NOS выражаются в виде 2 или 3 изоферментов, соответственно, которые являются продуктами разных генов (3).Действие 4 наборов ферментов в конечном итоге приводит к производству 7 низкомолекулярных продуктов, изображенных на рисунке 1. В дополнение к аргинину, орнитин и цитруллин выделены на рисунке 2 не только потому, что они представляют ключевые точки ветвления в метаболизме аргинина, но и также потому, что они обычно количественно оцениваются в клинических анализах биологических жидкостей и, таким образом, могут дать ключ к разгадке нарушений метаболизма аргинина при заболевании. Термин указывает на используется сознательно, потому что из рисунка должно быть очевидно, что невозможно сделать окончательный вывод об изменениях активности любого из перечисленных здесь ферментов исключительно на основе изменений, например, концентраций аргинина в плазме крови. орнитин или цитруллин.
РИСУНОК 2.
Обзор метаболических путей аргинина. Показаны только ферменты, которые напрямую используют или производят аргинин, орнитин или цитруллин. Для ясности показаны не все реагенты и продукты. Ингибирование синтазы оксида азота (NOS) и аргиназы асимметричным диметиларгинином (ADMA) и N G -гидрокси-1-аргинином (NOHA), соответственно, обозначены пунктирными линиями и штрихом в круге. Аминокислотные остатки в белках указаны в скобках.ADC, аргининдекарбоксилаза; АГАТ, аргинин: глицинамидинотрансфераза; ARG, аргиназа; ASL, аргининосукцинатлиаза; АСС, аргининосукцинатсинтетаза; ДДАГ, диметиларгининдиметиламиногидролаза; Me 2 , диметил; NO, оксид азота; ОАТ, орнитинаминотрансфераза; ODC, орнитиндекарбоксилаза; OTC, орнитин-транскарбамилаза; P5C, 1-Δ 1 -пирролин-5-карбоксилат; PRMT, протеин-аргининметилтрансфераза. Примечание: P5C находится в химическом равновесии с l-глутамат-γ-полуальдегидом посредством спонтанной неферментативной реакции (не показано).Изменено с рисунка 1 ссылки 24 с разрешения Американского общества диетологии.
РИСУНОК 2.
Обзор метаболических путей аргинина. Показаны только ферменты, которые напрямую используют или производят аргинин, орнитин или цитруллин. Для ясности показаны не все реагенты и продукты. Ингибирование синтазы оксида азота (NOS) и аргиназы асимметричным диметиларгинином (ADMA) и N G -гидрокси-1-аргинином (NOHA), соответственно, обозначены пунктирными линиями и штрихом в круге.Аминокислотные остатки в белках указаны в скобках. ADC, аргининдекарбоксилаза; АГАТ, аргинин: глицинамидинотрансфераза; ARG, аргиназа; ASL, аргининосукцинатлиаза; АСС, аргининосукцинатсинтетаза; ДДАГ, диметиларгининдиметиламиногидролаза; Me 2 , диметил; NO, оксид азота; ОАТ, орнитинаминотрансфераза; ODC, орнитиндекарбоксилаза; OTC, орнитин-транскарбамилаза; P5C, 1-Δ 1 -пирролин-5-карбоксилат; PRMT, протеин-аргининметилтрансфераза. Примечание: P5C находится в химическом равновесии с l-глутамат-γ-полуальдегидом посредством спонтанной неферментативной реакции (не показано).Изменено с рисунка 1 ссылки 24 с разрешения Американского общества диетологии.
Сама сложность рисунка 2 может создать впечатление, что мы знаем о метаболизме аргинина in vivo больше, чем это есть на самом деле. Фактически, рисунок слишком упрощен, потому что он не указывает, какие ферменты экспрессируются как изоферменты, паттерны тканеспецифической экспрессии, субклеточная локализация, наличие различных меж- и внутриклеточных транспортных систем, какие ферменты регулируются, или поток субстратов. .Поскольку он представляет собой совокупность реакций, присутствующих в организме в целом, рисунок 2 не следует рассматривать как отражающий метаболизм аргинина, происходящий в каком-либо отдельном органе или типе клеток. Например, циклическое превращение орнитина в цитруллин, аргинин в орнитин представляет собой основные реакции цикла мочевины, которые в высокой степени выражены почти исключительно в перипортальных гепатоцитах. Однако превращение орнитина в цитруллин в аргинин также представляет собой путь чистого синтеза аргинина, который происходит в первую очередь в кишечно-почечной оси, отмеченной ранее.Таким образом, тканеспецифическая экспрессия различных подгрупп или изоформ ферментов метаболизма аргинина, которая приводит к множеству различных метаболических исходов, не очевидна только из рисунка.
Известны взаимодействия между некоторыми ферментами и метаболитами. Благодаря общему использованию аргинина в качестве субстрата, NOS и аргиназа могут взаимно ограничивать доступность субстрата (8). Было показано аргиназозависимое ограничение доступности субстрата для NOS (25), но не наоборот.Вместо этого N G -гидрокси-1-аргинин, промежуточный продукт в синтезе NO, также является мощным ингибитором аргиназы (26), и во время синтеза NO с высоким выходом он может накапливаться в достаточной степени, чтобы значительно ингибировать аргиназу (27). Наконец, при обмене белков, содержащих метилированные остатки аргинина, высвобождается асимметричный диметиларгинин (ADMA), который является мощным ингибитором ферментов NOS (28). Хотя эти взаимодействия можно легко продемонстрировать на моделях культивированных клеток, оценка таких эффектов in vivo, особенно в определенных анатомических участках, представляет собой значительные технические проблемы.
Многое из того, что известно о динамике метаболизма аргинина in vivo, особенно у людей, получено из элегантных исследований индикаторов, проведенных Верноном Янгом и его коллегами. Поскольку большая часть этой информации была представлена в другом месте (8), здесь будут отмечены лишь некоторые моменты. Скорость производства аргинина, его предшественников и производных сильно различается. Например, скорость синтеза мочевины и креатина у здоровых взрослых натощак, соответственно, ≈180 и 8 раз выше, чем у NO, что составляет 1 мкмоль · ч -1 · кг массы тела -1 (8).Скорость метаболизма аргинина различными путями значительно различается между младенцами и взрослыми (8), а также во время травм и болезней (29–32). Сравнимых данных относительно скорости синтеза агматина и полиаминов нет.
Ветвь метаболизма аргинина млекопитающих, о которой мы знаем меньше всего, связана с агматином, продуктом декарбоксилирования аргинина. Хотя его свойства как сигнального агента клетки изучаются фармакологами в течение некоторого времени (33), предполагаемая физиологическая роль агматина или его производных, например, в регуляции синтеза NO или полиамина (6, 34–36), еще не изучена. быть твердо установленным.Недавно сообщалось о клонировании аргининдекарбоксилазы (37), но существуют разногласия относительно того, является ли она ферментативно активной в тканях млекопитающих (38), несмотря на сообщения об обратном (39, 40). Агматиназа, фермент, превращающий агматин в путресцин, также был клонирован (41, 42), но анализ последовательности показывает, что он может не функционировать у всех животных, включая мышей (43). Взятые вместе, аргининдекарбоксилаза и агматиназа составляют путь синтеза полиаминов, который является альтернативой орнитиндекарбоксилазозависимому пути, но практически отсутствуют данные относительно количественного значения этого альтернативного пути.
Поскольку агматин является продуктом микробного метаболизма, также неясно, какое количество агматина, обнаруженного в тканях (44), является продуктом клеток млекопитающих или кишечных бактерий. Агматин можно найти в некоторых продуктах питания, особенно в продуктах ферментации, таких как сыры и некоторые спиртные напитки (45, 46), но маловероятно, что небольшие количества, обнаруженные в таких продуктах, вносят значительный вклад в общее количество агматина, присутствующего в организме. .
Хотя в этой статье не обсуждается роль аргинина и его посттрансляционных модификаций в функции белков, следует понимать, что эти модификации включают не только метилирование остатков аргинина (47), но также превращение в цитруллин (48) или орнитин ( 49) (Рисунок 2).Модифицированные остатки аргинина высвобождаются в виде свободных аминокислот во время белкового обмена, а повышенные концентрации хотя бы одной из них (ADMA) представляют собой фактор риска сердечно-сосудистых заболеваний (50), вероятно, из-за его ингибирования ферментов NOS (27), поскольку показано на рисунке 2. Поскольку гомоцистеин может ингибировать активность диметиларгининдиметиламиногидролазы (51), фермента, который разрушает ADMA (52), существует сложное взаимодействие между метаболизмом серосодержащих аминокислот и метаболизмом аргинина.На уровне всего тела доля свободного цитруллина и орнитина, получаемая в результате белкового обмена, вероятно, довольно мала. Однако, поскольку недавние исследования показывают, что множественные внутриклеточные пулы аргинина могут существовать по крайней мере в некоторых типах клеток (23), аргинин, полученный в результате белкового обмена, может, следовательно, представлять значительную долю по крайней мере одного из этих пулов в определенных клетках. Исследователи только недавно узнали, что обмен белков может высвобождать различные производные аргинина; таким образом, потенциальное влияние этого процесса на метаболизм аргинина еще предстоит оценить.
РЕГУЛИРОВАНИЕ ЭКСПРЕССИИ ГЕНОВ АРГИНИНОМ
Снижение доступности аргинина, которое может происходить во время катаболического стресса, может предпочтительно изменять экспрессию определенных белков сверх того, что можно было бы ожидать, просто как следствие изменений в скорости глобального синтеза белка. Неудивительно, что большинство пораженных белков сами участвуют в каком-либо аспекте метаболизма аргинина. В соответствии с текущими тенденциями в номенклатуре, мы можем рассматривать исследование избирательного воздействия аргинина на экспрессию генов как область «аргеномики», подмножества эффектов питательных веществ на экспрессию генов (т.е. нутригеномики).Можно считать, что аргеномика зародилась в начале 1960-х годов, когда Роберт Шимке показал, что активность аргининосукцинатсинтетазы и аргининосукцинатлиазы в некоторых линиях клеток млекопитающих подавляется аргинином и увеличивается при замене аргинина цитруллином (53, 54). Механизмы, лежащие в основе таких изменений, в то время не были известны, но позже было показано, что они включают регуляцию как на транскрипционном, так и на посттранскрипционном уровне гена аргининосукцинатсинтетазы (55, 56).Однако точные механизмы, посредством которых происходят изменения транскрипции, еще предстоит выяснить примерно через 40 лет.
Совсем недавно было показано, что на экспрессию нескольких других генов преимущественно влияет снижение доступности аргинина, которое может происходить в результате повышенной активности аргиназы. К ним относятся iNOS, переносчик катионных аминокислот 1 (CAT-1) и дзета-цепь рецептора Т-клеток. Интересно, что эффекты депривации аргинина не идентичны ни для этих генов, ни для задействованных механизмов.Депривация аргинина приводит к снижению экспрессии iNOS (57, 58) и дзета-цепи Т-клеточного рецептора (59, 60), но приводит к увеличению экспрессии CAT-1 (61–64). Эти изменения происходят за счет снижения эффективности трансляции матричной РНК (мРНК) iNOS (57), снижения стабильности белка iNOS (58), уменьшения периода полужизни мРНК, кодирующей дзета-цепь Т-клеточного рецептора (59), и увеличения как в эффективность трансляции мРНК CAT-1 (64) и транскрипции гена CAT-1 (62) (рис. 3).Измененная эффективность трансляции мРНК iNOS и CAT-1 является следствием повышенной активности киназы GCN2, что приводит к фосфорилированию (и снижению активности) фактора инициации эукариот 2α (57), но с противоположными результатами. В случае мРНК iNOS инициация трансляции может быть полностью отменена в этих условиях, но в случае мРНК CAT-1 ремоделирование структуры лидерной последовательности мРНК и усиление инициации трансляции на внутреннем сайте входа в рибосому приводят к увеличению CAT-1. -1 выражение.Некоторые из регуляторных эффектов депривации аргинина также могут быть специфичными для клеточного типа. Например, депривация аргинина приводит к снижению стабильности белка iNOS в макрофагах (58), но не в астроцитах (57). Более того, неопубликованные исследования в лаборатории автора показывают, что депривация аргинина также может снижать экспрессию iNOS на претрансляционном уровне в некоторых типах клеток. Поскольку молекулы, которые воспринимают изменения в концентрации аргинина, и пути передачи сигнала, которые связывают датчики с конечными эффектами, неизвестны, их идентификация представляет собой сложный аспект текущих исследований метаболизма аргинина.
РИСУНОК 3.
Влияние ограничения доступности аргинина на экспрессию генов. Блочные стрелки обозначают увеличение или уменьшение указанных функций. Пути, которые еще не охарактеризованы, обозначены пунктирными линиями и вопросительными знаками. CAT-1, переносчик катионных аминокислот 1; eIF2α, фактор инициации эукариот 2α; GCN2, общий контроль питания 2; iNOS, индуцибельная синтаза оксида азота; мРНК, информационная РНК; TCR, рецептор Т-клеток.
РИСУНОК 3.
Влияние ограничения доступности аргинина на экспрессию генов. Блочные стрелки обозначают увеличение или уменьшение указанных функций. Пути, которые еще не охарактеризованы, обозначены пунктирными линиями и вопросительными знаками. CAT-1, переносчик катионных аминокислот 1; eIF2α, фактор инициации эукариот 2α; GCN2, общий контроль питания 2; iNOS, индуцибельная синтаза оксида азота; мРНК, информационная РНК; TCR, рецептор Т-клеток.
ЗАКЛЮЧИТЕЛЬНЫЕ ЗАМЕЧАНИЯ
Очевидно, что наши знания об идентичности метаболитов аргинина и ферментов, которые используют или производят аргинин, значительно расширились за последние годы, и отдельные главы можно легко написать (или уже в некоторых случаях уже писали) для каждого из них. описанные здесь ферменты и метаболиты.В то же время также очевидно, что многое еще предстоит сделать, включая расширение наших знаний о динамическом состоянии метаболизма аргинина in vivo, особенно в пределах местных анатомических участков, а также характеристику ролей и регуляции многих метаболических ферментов аргинина. и транспортные системы в здоровье и болезнях. Это обнадеживающее положение не только для тех из нас, кто уже увлекся сложностями метаболизма этой замечательной аминокислоты, но и для тех, кто ищет новых проблем.
СММ провела обзор литературы и несла полную ответственность за подготовку рукописи. У автора не было конфликта интересов.
ССЫЛКИ
1.
Моррис
SM
мл.
Регуляция ферментов цикла мочевины и метаболизма аргинина
.
Annu Rev Nutr
2002
;
22
:
87
—
105
.2.
Моррис
SM
мл.
Последние достижения в метаболизме аргинина
.
Curr Opin Clin Nutr Metab Care
2004
;
7
:
45
—
51
.3.
Wu
G
,
Morris
SM
Jr. Метаболизм аргинина у млекопитающих. В:
Cynober
LA
, изд.
Метаболические и терапевтические аспекты аминокислот в лечебном питании.
Бока-Ратон, Флорида
:
CRC Press
,
2004
:
153
—
67
.4.
Метаболизм аргинина: энзимология, питание и клиническое значение. В: Morris SM Jr, Loscalzo J, Bier D, Souba WW, eds. Материалы симпозиума, посвященного памяти Вернона Р. Янга. 5–6 апреля 2004 г. Бермудские острова
.
J Nutr
2004
;
134
(доп.):
2741S
—
897S
. 5.
Флинн
NE
,
Meininger
CJ
,
Haynes
TE
,
Wu
G
.
Метаболические основы аргининового питания и фармакотерапии
.
Biomed Pharmacother
2002
;
56
:
427
—
38
.6.
Сатриано
Дж
.
Пути аргинина и воспалительная реакция: взаимодействие оксида азота и полиаминов: обзорная статья
.
аминокислоты
2004
;
26
:
321
—
9
.7.
Closs
EI
,
Simon
A
,
Vekony
N
,
Rotmann
A
.
Плазменные мембранные переносчики аргинина
.
J Nutr
2004
;
134
:
2752S
—
9S
.8.
Wu
G
,
Morris
SM
Jr.
Метаболизм аргинина: оксид азота и выше
.
Biochem J
1998
;
336
:
1
—
17
.9.
Рабье
D
,
Kamoun
P
.
Метаболизм цитруллина у человека
.
аминокислоты
1995
;
9
:
299
—
316
.10.
Curis
E
,
Nicolis
I
,
Moinard
C
и др.
Практически все о цитруллине у млекопитающих
.
аминокислоты
2005
;
29
:
177
—
205
.11.
Абумрад
NN
,
Barbul
A
. Использование аргинина в клинической практике.В:
Cynober
LA
, изд.
Метаболические и терапевтические аспекты аминокислот в лечебном питании.
Бока-Ратон, Флорида
:
CRC Press
,
2004
:
595
—
611
.12.
Castillo
L
,
Chapman
TE
,
Sanchez
M
и др.
Кинетика аргинина и цитруллина в плазме у взрослых, получавших адекватную диету без аргинина
.
Proc Natl Acad Sci U S A
1993
;
90
:
7749
—
53
. 13.
Shi
O
,
Morris
SM
Jr,
Zoghbi
H
,
Porter
CW
,
O’Brien
WE
.
Создание мышиной модели дефицита аргиназы II путем направленного разрушения гена аргиназы II
.
Mol Cell Biol
2001
;
21
:
811
—
3
.14.
Игл
H
.
Метаболизм аминокислот в культурах клеток млекопитающих
.
Science
1959
;
130
:
432
—
7
. 15.
Джексон
MJ
,
Beaudet
AL
,
O’Brien
WE
.
Ферменты цикла мочевины млекопитающих
.
Annu Rev Genet
1986
;
20
:
431
—
64
. 16.
Моррис
SM
мл. Синтез, метаболизм и транспорт аргинина: регуляторы синтеза оксида азота. In:
Laskin
JD
,
Laskin
DL
, ред.
Клеточная и молекулярная биология оксида азота.
Нью-Йорк, Нью-Йорк
:
Марсель Деккер, Инк
,
1999
:
57
—
85
. 17.
Моррис
SM
мл. Регулирование доступности аргинина и его влияние на синтез NO.В:
Игнарро
LJ
, изд.
Оксид азота. Биология и патобиология.
Сан-Диего, Калифорния
:
Academic Press
,
2000
:
187
—
97
0,18.
Мори
M
,
Gotoh
T
.
Регулирование производства оксида азота ферментами метаболизма аргинина
.
Biochem Biophys Res Commun
2000
;
275
:
715
—
9
.19.
Пастор
CM
,
Morris
SM
Jr,
Billiar
TR
.
Источники аргинина для индуцированного синтеза оксида азота в изолированной перфузированной печени
.
Am J Physiol
1995
;
269
:
G861
—
6
.20.
Forstermann
U
,
Closs
EI
,
Pollock
JS
и др.
Изоферменты синтазы оксида азота.Характеристика, очистка, молекулярное клонирование и функции
.
Гипертония
1994
;
23
:
1121
—
31
,21.
Курц
S
,
Харрисон
DG
.
Инсулин и аргининовый парадокс
.
J Clin Invest
1997
;
99
:
369
—
70
. 22.
Xie
L
,
Брутто
SS
.
Сверхэкспрессия аргининосукцинатсинтетазы в гладкомышечных клетках сосудов потенцирует индуцированную иммуностимуляторами продукцию NO
.
J Biol Chem
1997
;
272
:
16624
—
30
. 23.
Simon
A
,
Plies
L
,
Habermeier
A
,
Martine
U
,
Reining
M
,
Closs
000.
Роль нейтрального транспорта аминокислот и распада белка в субстрате синтазы оксида азота в эндотелиальных клетках человека
.
Circ Res
2003
;
93
:
813
—
20
. 24.
Моррис
SM
мл.
Ферменты метаболизма аргинина
.
J Nutr
2004
;
134
:
2743S
—
7S
. 25.
Tenu
J-P
,
Lepoivre
M
,
Moali
C
,
Brollo
M
,
Mansuy
D
,
Boucher 9 -0004.
Влияние нового ингибитора аргиназы N ω -гидрокси-нор-L-аргинин на активность NO-синтазы в макрофагах мыши
.
оксид азота
1999
;
3
:
427
—
38
,26.
Daghigh
F
,
Fukuto
JM
,
Ясень
DE
.
Ингибирование аргиназы печени крысы промежуточным звеном в биосинтезе NO, N G -гидрокси-L-аргинин: значение для регуляции биосинтеза оксида азота аргиназой
.
Biochem Biophys Res Commun
1994
;
202
:
174
—
80
0,27.
Buga
GM
,
Singh
R
,
Pervin
S
и др.
Активность аргиназы в эндотелиальных клетках: ингибирование N G -гидроксиаргинин во время производства оксида азота с высоким выходом
.
Am J Physiol
1996
;
271
:
h2988
—
98
.28.
Boger
RH
,
Vallance
P
,
Cooke
JP
.
Асимметричный диметиларгинин (ADMA): ключевой регулятор синтазы оксида азота
.
Atheroscler Suppl
2003
;
4
:
1
—
3
,29.
Аргаман
Z
,
Янг
VR
,
Новиски
N
и др.
Метаболизм аргинина и оксида азота у детей с сепсисом в критическом состоянии
.
Crit Care Med
2003
;
31
:
591
—
7
. 30.
Castillo
L
,
DeRojas-Walker
T
,
Yu
YM
и др.
Метаболизм аргинина в организме и синтез оксида азота у новорожденных со стойкой легочной гипертензией
.
Pediatr Res
1995
;
38
:
17
—
24
. 31.
Lau
T
,
Owen
W
,
Yu
YM
и др.
Метаболизм аргинина, цитруллина и оксида азота у пациентов с терминальной стадией почечной недостаточности
.
J Clin Invest
2000
;
105
:
1217
—
25
. 32.
Yu
YM
,
Ryan
CM
,
Castillo
L
и др.
Кинетика аргинина и орнитина у пациентов с тяжелыми ожогами: повышенная скорость выведения аргинина
.
Am J Physiol Endocrinol Metab
2001
;
280
:
E509
—
17
.33.
Raasch
W
,
Schafer
U
,
Chun
J
,
Dominiak
P
.
Биологическое значение агматина, эндогенного лиганда в сайтах связывания имидазолина
.
Br J Pharmacol
2001
;
133
:
755
—
80
. 34.
Grillo
MA
,
Colombatto
S
.
Метаболизм и функция в тканях животных агматина, биогенного амина, образованного из аргинина
.
аминокислоты
2004
;
26
:
3
—
8
,35.
Blantz
RC
,
Satriano
J
,
Gabbai
F
,
Kelly
C
.
Биологические эффекты метаболитов аргинина
.
Acta Physiol Scand
2000
;
168
:
21
—
5
,36.
Сатриано
Дж
,
Келли
CJ
,
Бланц
RC
.
Возрастающая роль агматина
.
Kidney Int
1999
;
56
:
1252
—
3
0,37.
Zhu
MY
,
Iyo
A
,
Piletz
JE
,
Regunathan
S
.
Экспрессия человеческой аргининдекарбоксилазы, фермента биосинтеза агматина
.
Biochim Biophys Acta
2004
;
1670
:
156
—
64
.38.
Coleman
CS
,
Hu
G
,
Pegg
AE
.
Биосинтез путресцина в тканях млекопитающих
.
Biochem J
2004
;
379
:
849
—
55
. 39.
Li
G
,
Regunathan
S
,
Reis
DJ
.
Агматин синтезируется митохондриальной аргининдекарбоксилазой в головном мозге крысы
.
Ann N Y Acad Sci
1995
;
763
:
325
—
9
.40.
Горынь
О
,
Луховый
Б
,
Лазаров
А
и др.
Биосинтез агматина в изолированных митохондриях и перфузированной печени крысы: исследования с N-меченным аргинином
.
Biochem J
2005
;
388
:
419
—
25
.41.
Mistry
SK
,
Burwell
TJ
,
Chambers
RM
и др.
Клонирование агматиназы человека. Альтернативный путь синтеза полиаминов, индуцированный в печени вирусом гепатита B
.
Am J Physiol
2002
;
282
:
G375
—
81
.42.
Iyer
RK
,
Kim
HK
,
Tsoa
RW
,
Grody
WW
,
Cederbaum
SD
.
Клонирование и характеристика агматиназы человека
.
Mol Genet Metab
2002
;
75
:
209
—
18
. 43.
Моррис
SM
Jr.
Агматиназы позвоночных: какую роль они играют в катаболизме агматина?
Ann N Y Acad Sci
2003
;
1009
:
30
—
3
. 44.
Raasch
W
,
Regunathan
S
,
Li
G
,
Reis
DJ
.
Агматин широко и неравномерно распределен в органах крыс
.
Ann N Y Acad Sci
1995
;
763
:
330
—
4
. 45.
Sayem-el-Daher
N
,
Simard
RE
,
L’Heureux
L
,
Roberge
AG
.
Определение моно-, ди- и полиаминов в пищевых продуктах с использованием одноколоночного аминокислотного автоанализатора
.
J Chromatogr
1983
;
256
:
313
—
21
.46.
Okamoto
A
,
Sugi
E
,
Koizumi
Y
,
Yanagida
F
,
Udaka
S
.
Содержание полиаминов в обычных пищевых продуктах и различных ферментированных продуктах
.
Biosci Biotechnol Biochem
1997
;
61
:
1582
—
4
. 47.
McBride
AE
,
Серебро
PA
.
Состояние arg: метилирование белка по аргинину достигает возраста
.
Cell
2001
;
106
:
5
—
8
. 48.
Vossenaar
ER
,
Zendman
AJ
,
van Venrooij
WJ
,
Pruijn
GJ
.
PAD, растущее семейство цитруллинирующих ферментов: гены, особенности и участие в заболевании
.
Bioessays
2003
;
25
:
1106
—
18
,49.
Продам
DR
,
Monnier
VM
.
Превращение аргинина в орнитин путем улучшенного гликирования в стареющих человеческих кристаллах коллагена и хрусталика
.
J Biol Chem
2004
;
279
:
54173
—
84
,50.
Богер
RH
.
Асимметричный диметиларгинин, эндогенный ингибитор синтазы оксида азота, объясняет «парадокс L-аргинина» и действует как новый фактор риска сердечно-сосудистых заболеваний
.
J Nutr
2004
;
134
:
2842S
—
7S
.51.
Stuhlinger
MC
,
Tsao
PS
,
Her
JH
,
Kimoto
M
,
Balint
RF
,
000 Cooke
000
Гомоцистеин нарушает путь синтазы оксида азота: роль асимметричного диметиларгинина
.
Тираж
2001
;
104
:
2569
—
75
,52.
Тран
CT
,
Leiper
JM
,
Валанс
P
.
Путь DDAH / ADMA / NOS
.
Atheroscler Suppl
2003
;
4
:
33
—
40
. 53.
Шимке
РТ
.
Репрессия ферментов биосинтеза аргинина в культуре тканей млекопитающих
.
Biochim Biophys Acta
1962
;
62
:
599
—
601
. 54.
Шимке
РТ
.
Ферменты метаболизма аргинина в культуре клеток млекопитающих.I. Репрессия аргининосукцинатсинтетазы и аргининосукциназы
.
J Biol Chem
1964
;
239
:
136
—
45
, 55.
Джексон
MJ
,
Allen
SJ
,
Beaudet
AL
,
O’Brien
WE
.
Метаболическая регуляция аргининосукцинатсинтетазы в культивируемых клетках человека
.
J Biol Chem
1988
;
263
:
16388
—
94
.56.
Jackson
MJ
,
O’Brien
WE
,
Beaudet
AL
.
Аргинин-опосредованная регуляция минигена аргининосукцинатсинтетазы в нормальных и устойчивых к канаванину клетках человека
.
Mol Cell Biol
1986
;
6
:
2257
—
61
. 57.
Lee
J
,
Ryu
H
,
Ferrante
RJ
,
Morris
SM
Jr,
Ratan
RR
.
Трансляционный контроль индуцибельной экспрессии синтазы оксида азота аргинином может объяснить аргининовый парадокс
.
Proc Natl Acad Sci U S A
2003
;
100
:
4843
—
8
,58.
Эль-Гаяр
S
,
Тюринг-Нахлер
H
,
Pfeilschifter
J
,
Rollinghoff
M
,
Bogdan
C
.
Трансляционный контроль индуцибельной синтазы оксида азота с помощью IL-13 и доступности аргинина в воспалительных макрофагах
.
J Immunol
2003
;
171
:
4561
—
8
, 59.
Rodriguez
PC
,
Zea
AH
,
Culotta
KS
,
Zabaleta
J
,
Ochoa
JB
a,
Ochoa
.
Регулирование экспрессии цепи CD3zeta Т-клеточного рецептора с помощью L-аргинина
.
J Biol Chem
2002
;
277
:
21123
—
9
.60.
Taheri
F
,
Ochoa
JB
,
Faghiri
Z
и др.
L-аргинин регулирует экспрессию дзета-цепи Т-клеточного рецептора (CD3zeta) в клетках Jurkat
.
Clin Cancer Res
2001
;
7
:
958с
—
65с
.61.
Аулак
KS
,
Mishra
R
,
Zhou
L
и др.
Посттранскрипционная регуляция транспортера аргинина Cat-1 по доступности аминокислот
.
J Biol Chem
1999
;
274
:
30424
—
32
0,62.
Fernandez
J
,
Lopez
AB
,
Wang
C
и др.
Транскрипционный контроль транспортера аргинина / лизина, cat-1, физиологическим стрессом
.
J Biol Chem
2003
;
278
:
50000
—
9
0,63.
Fernandez
J
,
Yaman
I
,
Merrick
WC
и др.
Регуляция внутренней трансляции, опосредованной сайтом входа в рибосомы, фосфорилированием эукариотического фактора инициации-2 альфа и трансляцией небольшой открытой рамки считывания, расположенной выше
.
J Biol Chem
2002
;
277
:
2050
—
8
0,64.
Fernandez
J
,
Yaman
I
,
Mishra
R
и др.
Внутренняя трансляция мРНК млекопитающих, опосредованная сайтом входа в рибосомы, регулируется доступностью аминокислоты
.
J Biol Chem
2001
;
276
:
12285
—
91
.
© 2006 Американское общество клинического питания
Исследование влияния поверхностного лизина на мутагенез аргинина на стабильность и структуру белка с использованием зеленого флуоресцентного белка
Abstract
Две положительно заряженные основные аминокислоты, аргинин и лизин, в основном подвергаются воздействию поверхности белка и играют важную роль в стабильности белка, формируя электростатические взаимодействия.В частности, гуанидиниевая группа аргинина допускает взаимодействия в трех возможных направлениях, что позволяет аргинину образовывать большее количество электростатических взаимодействий по сравнению с лизином. Более высокое значение pKa основного остатка в аргинине может также вызывать более стабильные ионные взаимодействия, чем лизин. В этой статье сообщается об исследовании, можно ли использовать преимущества аргинина по сравнению с лизином для повышения стабильности белка. Вариант зеленого флуоресцентного белка (GFP) был создан путем мутации максимально возможного количества остатков лизина на поверхности в аргинины при сохранении активности.Когда стабильность варианта была исследована в диапазоне денатурирующих условий, вариант был относительно более стабильным по сравнению с контрольным GFP в присутствии химических денатурантов, таких как мочевина, щелочной pH и ионные детергенты, но термостабильность белка не была измененный. Смоделированная структура варианта указывает на предполагаемые новые солевые мостики и взаимодействия водородных связей, которые помогают улучшить жесткость белка против различных химических денатурантов. Структурный анализ электростатических взаимодействий также подтвердил, что геометрические свойства гуанидиниевой группы в аргинине имеют такие эффекты.С другой стороны, измененные электростатические взаимодействия, вызванные мутагенезом поверхностных лизинов в аргинины, отрицательно влияют на укладку белка, что снижает продуктивность функциональной формы варианта. Эти результаты предполагают, что мутагенез поверхностного лизина до аргининов можно рассматривать как один из параметров в инженерии стабильности белка.
Образец цитирования: Sokalingam S, Raghunathan G, Soundrarajan N, Lee S-G (2012) Исследование влияния поверхностного лизина на мутагенез аргинина на стабильность и структуру белка с использованием зеленого флуоресцентного белка.PLoS ONE 7 (7):
e40410.
https://doi.org/10.1371/journal.pone.0040410
Редактор: Пермяков Евгений Александрович, Институт биологического приборостроения РАН, Российская Федерация
Поступила: 14 марта 2012 г .; Принято к печати: 6 июня 2012 г .; Опубликовано: 9 июля 2012 г.
Авторские права: © 2012 Sokalingam et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Это исследование было поддержано Программой фундаментальных наук через Национальный исследовательский фонд Кореи (NRF), финансируемого Министерством образования, науки и технологий (NRF-2011-0021222). Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили, что конкурирующих интересов не существует.
Введение
Устойчивость белка к нефизиологическим условиям, таким как высокие температуры, кислотный или щелочной pH и детергенты, является важной проблемой в промышленных процессах и с академической точки зрения.Повышение стабильности белка становится все более важным из-за растущего спроса на использование белков как частей биологических устройств, обрабатываемых и работающих в суровых условиях в дополнение к традиционным ферментативным процессам [1] — [3]. Глубокое понимание механизма сворачивания белка и структуры белка дало представление о стабильности белка, что позволило разработать ряд подходов к инженерии белка для улучшения стабильности белка [4] — [8]. Например, стабильность белка можно повысить путем введения большего количества внутридисульфидных связей, создания более гидрофобных ядер или изменения поверхностных зарядов [9] — [12].Тем не менее, необходимо определить дополнительные параметры, относящиеся к стабильности белка, поскольку стабильность белка является кооперативным феноменом различных взаимодействий в белке.
Аргинин и лизин — это положительно заряженные основные аминокислоты в физиологических условиях, которые в основном находятся на поверхности белка [13], [14]. Две аминокислоты на поверхности белка играют важную роль в стабильности белка, образуя ионные взаимодействия и водородные связи в белках, а также взаимодействуя с молекулами воды [15], [16].Хотя оба они функционируют как основные остатки, остаток аргинина обеспечивает структуру белка с большей стабильностью, чем лизин, благодаря своей геометрической структуре. Группа гуанидиния в аргинине допускает взаимодействия в трех возможных направлениях через три асимметричных атома азота (N ε , N η1 , N η2 ), тогда как основная функциональная группа лизина допускает только одно направление взаимодействия [ 17], [18]. Это позволяет аргинину образовывать большое количество электростатических взаимодействий, таких как солевые мостики и водородные связи по сравнению с лизином, что предположительно приводит к более сильным взаимодействиям, чем взаимодействия, генерируемые лизином.Эти эффекты были косвенно продемонстрированы путем сравнения количества солевых мостиков, генерируемых аргинином и лизином в ряде белков, а также путем анализа отношения аргинина к лизину и солевых мостиков в термофильных и мезофильных белках [19] — [23]. В дополнение к геометрическому эффекту, ионные взаимодействия, образованные аргинином, могут быть более стабильными, чем взаимодействия лизинов, особенно при щелочном pH, из-за более высокого pKa основного остатка в аргинине, чем у лизина [24].
Превосходство аргинина по биохимическим свойствам над лизином в отношении электростатических взаимодействий, описанных выше, поднимает следующий вопрос.Что произойдет с точки зрения стабильности и структуры белка, если остатки лизина на поверхности белка заменятся на аргинин? В частности, это исследование фокусируется на следующем: (1) может ли замена аргининов лизинами на поверхности белка вызывать больше электростатических взаимодействий в белке; (2) может ли такой мутагенез стабилизировать белок; и (3) какой тип стабильности можно улучшить мутагенезом. Ответив на эти вопросы, можно было бы не только более точно определить влияние аргинина и лизина на стабильность и структуру белка, но и определить, можно ли рассматривать такой мутагенез как способ стабилизации белка.
В этом исследовании была предпринята попытка ответить на поставленные выше вопросы путем мутации остатков лизина на поверхности белка в аргинин и изучения его влияния на стабильность белка и электростатические взаимодействия. В качестве модельной системы был использован зеленый флуоресцентный белок (GFP), белок с флуоресцентным активным центром, где сворачивание и разворачивание можно отслеживать по его спектральным свойствам [25]. GFP использовался в основном как репортерная система [26], но он также используется в биологических устройствах, таких как биочипы и биосенсоры [27], [28], которые требуют стабильности белков в суровых условиях.На первом этапе этого исследования поверхностные остатки лизина GFP были максимально мутированы в аргинины с учетом складчатости белка (рис. 1). На втором этапе стабильность варианта была исследована в ряде условий денатурации белка, таких как высокие температуры, мочевина, щелочной pH и детергенты. Влияние мутагенеза на электростатические взаимодействия было также исследовано путем анализа смоделированной структуры варианта. Наконец, обсуждается возможность того, что мутагенез поверхностного лизина до аргинина может быть подходом к инженерии стабильности белка.
Рисунок 1. Структурное представление вариантов GFP.
Трехмерное мультипликационное изображение GFP con и GFP14R, показывающее поверхностные лизины (синий) и поверхностные аргинины (красный) в виде палочек, которые были мутированы в GFP14R. Графическое изображение было создано с помощью Discovery Studio Visualizer от Accelrys Software Inc.
https://doi.org/10.1371/journal.pone.0040410.g001
Результаты
Анализ электростатических взаимодействий в GFP
Электростатические взаимодействия в GFP анализировали с использованием кристаллической структуры GFP дикого типа с PDB Id 1GFL [29].В таблице 1 перечислены солевые мостики и водородные связи, связанные с остатками лизина и аргинина в GFP. GFP содержит 20 аминокислот лизина и 6 аминокислот аргинина, что соответствует 8,4% и 2,5% от его общего аминокислотного состава, соответственно. Для 20 лизинов 1, 8 и 11 расположены в областях спирали, бета-цепи и петли, соответственно. Для 6 аргининов 4 и 2 расположены в областях бета-цепи и петли, соответственно. Более 25% боковых цепей подвергаются действию растворителя для 19 остатков лизина и 5 остатков аргинина, которые в данном исследовании считались открытыми остатками.Остатки лизина и аргинина вносят вклад в 9 и 4 взаимодействия соляного мостика соответственно. Остатки, такие как K45, K113, R109 и R122, образуют сложные солевые мостики, взаимодействуя более чем с одним противоположно заряженным остатком, тогда как остальные представляют собой простые солевые мостики, образуя ионное взаимодействие с одной аминокислотой. Среднее количество солевых мостиков на лизин и аргинин составило приблизительно 0,45 и 0,66 соответственно. Анализ взаимодействий водородных связей показал, что среднее количество взаимодействий водородных связей на лизин и аргинин составляет примерно 2 и 3, соответственно.Эти результаты предполагают, что, как и большинство других белков, большинство остатков лизина и аргинина подвергаются действию растворителя, а аргинин в GFP образует больше электростатических взаимодействий, чем лизины.
Мутагенез поверхностных лизинов в аргинины
GFP mut3.1b , GFP быстрого сворачивания с мутациями S65G и S72A [30], был использован в качестве целевого белка GFP и обозначен в этом отчете как GFP con . В первоначальном исследовании мутагенеза все 19 подвергшихся воздействию поверхности лизиновых аминокислот GFP con были мутированы в аргинины, что привело к образованию варианта, обозначенного GFP19R.Когда вариант экспрессировался в Escherichia coli при 37 ° C, белок был полностью неправильно свернут in vivo (фигура S1). Экспрессия GFP19R при более низких температурах была выполнена для повышения эффективности сворачивания варианта, но этот подход оказался неэффективным при производстве растворимого и свернутого белка (рис. S1).
Эти результаты предполагают, что некоторые остатки лизина в GFP важны для правильной укладки GFP. Предыдущее сообщение подтвердило важность образования сверхстабильного ядра с β-нитями 1, 3, 4, 5 и 6 в сворачивании GFP [31], что было использовано в качестве основы для преодоления проблемы неправильного сворачивания GFP19R.Поскольку пять лизинов (K41, K45, K107, K113, K126) в GFP участвовали в β-цепях образования сверхстабильного ядра, соответствующие пять остатков аргинина в GFP19R подверглись обратной мутации в лизины, которые образовали вариант GFP14R. GFP14R также был в основном неправильно свернут при экспрессии при 37 ° C (рисунок S1), но растворимая экспрессия и флуоресценция всей клетки были обнаружены при его производстве при более низких температурах, 25 ° C (рисунок S2), несмотря на гораздо более низкий уровень экспрессии, чем у GFP con . Это предполагает, что восстановление пяти лизинов, участвующих в формировании сверхстабильного ядра GFP, эффективно для небольшого повышения эффективности сворачивания GFP19R.Достаточное количество GFP14R могло быть достигнуто путем его экспрессии в течение более длительных периодов времени при 25 ° C, и белок был очищен для дальнейших исследований (рисунок S2). Очищенный GFP14R показал такие же спектры возбуждения и излучения, что и GFP con (рис. 2), а его удельная интенсивность флуоресценции была аналогична таковой у GFP con (рис. S3). Эти результаты предполагают, что 14 мутаций Arg, введенные в поверхностные остатки Lys, оказали существенное неблагоприятное влияние на эффективность укладки GFP, но не повлияли значительно на конечную структуру GFP из-за его флуоресцентной активности.Хотя уровень экспрессии варианта GFP14R был намного ниже, чем у GFP con , способность варианта сворачиваться в активную форму побудила нас провести дальнейшие исследования его стабильности.
Рис. 2. Спектральные свойства вариантов GFP.
A) Возбуждение и B) Спектр излучения GFPcon и GFP14R. Все амплитуды произвольно нормированы на максимальное значение 1,0. (au — условные единицы).
https://doi.org/10.1371 / journal.pone.0040410.g002
Влияние мутагенеза поверхностных лизинов на аргинины на стабильность GFP
GFP14R, полученный в результате вышеуказанного исследования, содержал 19 поверхностных аргининов и 5 лизинов, тогда как GFP con содержал 19 поверхностных лизинов и 5 аргининов. Это предполагает наличие заметного различия в соотношении поверхностного Arg к Lys между двумя белками. Чтобы изучить влияние такого различия на стабильность GFP, стабильность очищенного GFP14R и GFP con исследовали в четырех различных условиях денатурации белка, т.е.е. высокие температуры, добавление мочевины, щелочной pH и добавление моющего средства.
Сначала было исследовано влияние температуры на стабильность белков GFP. Температурно-зависимый анализ проводили путем инкубации GFP14R и GFP con в диапазоне температур в течение 30 минут и измерения их флуоресценции. Как показано на рисунке 3A, GFP14R и GFP con показали аналогичную устойчивость к температуре. Все они имели тенденцию постепенно терять свою флуоресценцию после 60 ° C и полностью теряли свою активность при 80 ° C.И GFP14R, и GFP con показали аналогичные модели, когда анализ, зависящий от времени, проводился при 70 ° C (рис. 3B). Их период полураспада при 70 ° C оценивается примерно в 8 минут. Эти результаты предполагают, что введенные мутации Arg в остатки Lys не повлияли на термостабильность GFP.
Рис. 3. Влияние температуры на стабильность вариантов GFP.
A) Образцы белка инкубировали при различных температурах в течение 30 минут и измеряли оставшуюся флуоресценцию.Флуоресценция в нулевой момент времени при соответствующих температурах была принята за 100%. B) Образцы белка инкубировали при 70 ° C в течение различных интервалов времени и измеряли оставшуюся флуоресценцию. Флуоресценцию в нулевой момент времени принимали за 100%. (Планка погрешностей — стандартное отклонение трех независимых экспериментов).
https://doi.org/10.1371/journal.pone.0040410.g003
Была протестирована стабильность варианта GFP14R против хаотропного денатуранта, мочевины. Очищенные образцы GFP14R и GFP con инкубировали с различными концентрациями мочевины от 4 M до 8 M при 50 ° C в течение 30 минут.Вариант GFP14R показал стабильность до 5 М мочевины, но начал терять свою активность от 6 М мочевины. Напротив, GFP con показал пониженную активность от 4 M мочевины (рис. 4A). Когда анализ, зависящий от времени, был выполнен с 6 М мочевины при 50 ° C, период полураспада GFP14R и GFP con был оценен приблизительно как 93 и 51 минута, соответственно (Рисунок 4B), что позволяет предположить, что стабильность На GFP против мочевины положительно влиял мутагенез поверхностных лизинов до аргининов.
Рис. 4. Влияние мочевины на стабильность вариантов GFP.
A) Образцы белка инкубировали при 50 ° C с различными концентрациями мочевины от 4 M до 8 M в течение 30 минут и измеряли оставшуюся флуоресценцию. Флуоресценцию в нулевой момент времени при соответствующих концентрациях мочевины приняли за 100%. B) Образцы белка инкубировали при 50 ° C в присутствии 6 M мочевины в течение различных интервалов времени и измеряли оставшуюся флуоресценцию. Флуоресценцию в нулевой момент времени принимали за 100%.(Планка погрешностей — стандартное отклонение трех независимых экспериментов).
https://doi.org/10.1371/journal.pone.0040410.g004
Значения pKa аргинина и лизина составляют 12,48 и 10,53 соответственно [32], что может приводить к различной стабильности ионных взаимодействий, связанных с аргинином. остатки и остатки лизина при щелочном pH, как описано во введении. Чтобы оценить, может ли такой эффект вызвать разницу в устойчивости белка к щелочному pH, GFP con и GFP14R инкубировали при определенном диапазоне pH и измеряли их флуоресценцию через 30 минут инкубации при 60 ° C.Эксперименты показали, что и GFP14R, и GFP con относительно стабильны до pH 12,0, но GFP14R показал более высокую стабильность, чем GFP con при pH 13,0 (рис. 5A). Анализ, зависящий от времени, при pH 13,0 и 60 ° C подтвердил, что мутант GFP14R имеет более высокую стабильность, чем контроль (рис. 5B). При pH 13,0 период полураспада GFP con и GFP14R составлял 19 и 34 минуты соответственно. Эти результаты предполагают, что введенные мутации стабилизировали белок GFP до щелочного pH, как и ожидалось.
Рис. 5. Влияние pH на стабильность вариантов GFP.
A) Образцы белка инкубировали при 60 ° C в течение 30 минут в буферных растворах с различным pH и измеряли оставшуюся флуоресценцию. Флуоресценция в нулевой момент времени в соответствующем буфере была принята за 100%. B) Образцы белка в буфере KCl с pH 13,0 инкубировали при 60 ° C в течение различных интервалов времени и измеряли оставшуюся флуоресценцию. Флуоресценцию в нулевой момент времени принимали за 100%. (Планка погрешностей — стандартное отклонение трех независимых экспериментов).
https://doi.org/10.1371/journal.pone.0040410.g005
Два анионных детергента (додецилсульфат натрия (SDS) и додецилбензолсульфонат натрия (SDBS)) и два катионных детергента (бромид цетилтриметиламмония (CTAB) и хлорид додецилтриметиламмония (DTAC)) были протестированы для изучения влияния детергента на стабильность варианта GFP. GFP con и GFP14R инкубировали с 1% соответствующих детергентов и проводили флуоресцентный анализ, зависящий от времени.Как показано на рисунке 6, хотя профили дезактивации различались в зависимости от добавленных детергентов, GFP14R всегда демонстрировал более высокую стабильность, чем GFP con . В таблице 2 приведен расчетный период полураспада (t 1/2 ) каждого варианта в присутствии ионных детергентов. Интересно, что эффект стабилизации был выше в анионных детергентах, чем в катионных детергентах. В частности, эффект был наиболее выражен в SDS. Стабильность GFP14R была дополнительно исследована в диапазоне концентраций SDS.Белки инкубировали с 1-5% SDS при 50 ° C в течение 30 минут и определяли их оставшуюся флуоресцентную активность. Как показано на рисунке 7, GFP con в основном инактивировался добавлением SDS при всех концентрациях, но GFP14R проявлял примерно 50% активности даже при 5% SDS. Эти результаты позволяют предположить, что вариант GFP14R более жесткий и устойчивый к детергентам по сравнению с GFP con .
Рис. 6. Влияние ионных детергентов на стабильность вариантов GFP.
Пробы белка инкубировали с 1% SDS (A), 1% SDBS (B), 1% CTAB (C) и 1% DTAC (D) при 50 ° C в течение различных интервалов времени и измеряли оставшуюся флуоресценцию. Флуоресценция в нулевой момент времени для соответствующего детергента была принята за 100%. (Планка погрешностей — стандартное отклонение трех независимых экспериментов).
https://doi.org/10.1371/journal.pone.0040410.g006
Анализ электростатических взаимодействий GFP14R по отношению к лизину и аргинину
Приведенные выше результаты подтвердили, что введение аргининов в 14 остатков Lys GFP увеличивало стабильность GFP против мочевины, щелочного pH и детергентов.Хотя на термостабильность GFP не влиял мутагенез, результаты предполагали возможность изменений во взаимодействиях, связанных с поверхностными основными остатками, в сторону стабилизации белка. Чтобы проверить возможность более непосредственно, изменения в электростатических взаимодействиях, вызванные мутациями, были исследованы путем анализа солевых мостиков и водородных связей в GFP14R. Ожидалось, что анализ также ответит на один из основных вопросов, описанных во введении: может ли замена аргининов лизинами на поверхности белка вызвать большее количество электростатических взаимодействий в белке?
Как упоминалось выше, сходные спектральные свойства GFP con и GFP14R (Рисунок 2) предполагают, что существенная структура GFP не может быть существенно изменена мутациями.Это побудило нас предсказать трехмерную структуру GFP14R методом минимизации энергии. Кристаллическая структура GFP с PDB Id 1GFL [29] использовалась как шаблонная структура при моделировании, что позволило нам предположить, что локальный минимум энергии моделируемого варианта GFP был его собственным состоянием. Структура GFP con также была смоделирована с использованием той же процедуры, что и для контроля. Модели in silico GFP14R и GFP con не показали значительных отклонений от структуры шаблона со среднеквадратичным отклонением приблизительно 0.1 Å. Молекулярно-динамическое моделирование смоделированных структур показало, что среднеквадратичное отклонение (среднеквадратичное отклонение) для GFP con и GFP14R оказалось в пределах 2,5 Å в течение 10-наносекундного моделирования (Рисунок S4), что подтвердило, что предсказанные модели были в значительной степени стабильный. При анализе электростатических взаимодействий двух смоделированных белков расстояние между N-O для электростатической пары, образующей сильное солевое мостиковое взаимодействие, было принято равным ≤4 Å, а водородная связь была определена с использованием критериев приблизительно 2.От 5 до 3,2 Å [16], [33].
В таблице 3 перечислены солевые мостики и водородные связи, связанные с 19 поверхностными остатками Lys в моделированном GFP con , а также измененные взаимодействия в моделированном GFP14R для 19 остатков. Процесс минимизации энергии при моделировании GFP con предсказал три новых взаимодействия солевого мостика (K79-E5, K131-D103 и K166-D180) и 12 новых водородных связей по сравнению с взаимодействиями в исходной структуре GFP, показанной в таблице 1. , но было потеряно 8 водородных связей, сохранив солевые мостики.Это привело к образованию 11 солевых мостиков и 30 водородных связей, связанных с 19 поверхностными лизинами GFP con . С другой стороны, в модели GFP14R было обнаружено, что 16 солевых мостиков и 39 водородных связей вовлечены в 19 основных остатков. Это предполагает, что мутагенез поверхностных 14 лизинов к аргининам в конечном итоге вызвал добавление еще 5 солевых мостиков и 9 водородных связей. Сравнение 11 солевых мостиков в GFP con и 16 солевых мостов в GFP14R показало, что 9 солевых мостиков GFP con сохранились, 2 солевых моста GFP con были потеряны и 7 солевых мостиков. были вновь образованы мутагенезом.Что касается водородных связей, 24 водородные связи GFP con были сохранены, 6 были потеряны и было образовано 15 новых водородных связей. Это ясно показывает, что введение аргининов в 14 поверхностных остатков лизина значительно изменило электростатические взаимодействия в GFP. Это также демонстрирует, что аргинин более благоприятен для образования солевых мостиков и водородных связей на поверхности белка, чем лизин. Повышенное количество электростатических взаимодействий GFP14R по сравнению с GFP con может быть одним из факторов, влияющих на улучшение стабильности, хотя точно определить, насколько добавленные солевые мостики и водородные связи, индуцированные мутагенезом, способствовали повышению стабильности GFP, сложно. для определения, потому что другие факторы, такие как электростатические взаимодействия между основными остатками и молекулами воды, также могут влиять на стабильность белка.
Рисунок 7. Влияние концентрации SDS на стабильность вариантов GFP.
Образцы белка инкубировали с различными концентрациями SDS при 50 ° C в течение 30 минут и измеряли оставшуюся флуоресценцию. Флуоресценция в нулевой момент времени при соответствующих концентрациях SDS была принята за 100%. (Планка погрешностей — стандартное отклонение трех независимых экспериментов).
https://doi.org/10.1371/journal.pone.0040410.g007
Структурный анализ вновь образованных солевых мостиков в GFP14R
Вышеупомянутое исследование электростатических взаимодействий в GFP14R показало, что добавление аргининов к остаткам лизина создает солевые мостики и водородные связи более благоприятно.Это коррелирует с различной геометрической структурой основных боковых цепей аргинина и лизина, описанной во введении. Вновь образованные 7 солевых мостиков при мутации лизина и аргинина были исследованы структурно с использованием их смоделированных структур, чтобы более точно определить разницу между лизином и аргинином в образовании электростатических взаимодействий.
В GFP con Lys79 образовывал солевой мостик с Glu5, тогда как аргинин в том же положении в GFP14R проявлял три различных взаимодействия (рис. 8A): два солевых мостика с Glu5 и Asp76 с использованием N η1 и N η2 . и водородную связь с использованием N ε с карбонильным кислородом основной цепи Tyr74.Это изменение боковой цепи в 79 -м положении также способствовало образованию в его основной цепи азота водородных связей с основными цепями Pro75 и Asp76. Lys209 и Lys52 из GFP con не могли вызвать какое-либо взаимодействие соляной мостик, даже несмотря на то, что вокруг них имелся соответствующий заряженный остаток, Asp216. С другой стороны, когда аргинин был введен в эти два положения, как Arg209, так и Arg52 образовали взаимодействия солевого мостика с Asp216 (Рисунок 8B). Потеря водородной связи между Arg209 и Tyr143 в GFP14R, которая присутствует в GFP con , может быть компенсирована новым солевым мостиком.Солевой мостик Lys131-Asp103 в GFP con был потерян в GFP14R, но Arg131 сформировал солевой мостик с Asp133 (фиг. 8C). Солевой мостик с Asp103 был утерян, потому что N ε Arg131 благоприятствовал водородной связи, тогда как N η1 и N η2 предпочитал солевой мостик с Asp133. Взаимодействие с Asp103 поддерживается за счет водородной связи с его основной цепью. Открытая поверхность Arg3 в GFP14R, расположенном на гибком N-конце белка, показала возможность нового взаимодействия путем создания солевого мостика с Glu6 (рис. 8D).Боковая цепь Lys140 в GFP con образовывала две водородные связи с основной цепью Asp133 и Gly134. С другой стороны, гуанидиновый фрагмент аргинина в этом положении в GFP14R образовал новый солевой мостик с Asp173 с использованием N η1 , сохраняя водородные связи с использованием N ε с Gly134 и N η2 с Asp133 (Рисунок 8E ).
Рис. 8. Структурное представление новых взаимодействий соляных мостиков.
Сравнение расстояний взаимодействия солевого мостика между GFP con (левая панель) и GFP14R (правая панель): (A) Lys / Arg79-Glu5, (B) Lys / Arg209-Asp216 и Lys / Arg52-Asp216, ( C) Lys / Arg131-Asp103, (D) Lys / Arg3-Glu6 и (E) Lys / Arg140-Asp173.Черные прямые и зеленые пунктирные линии обозначают взаимодействия соляной мостик и водородные связи, соответственно. Графическое изображение было создано с помощью Discovery Studio Visualizer от Accelrys Software Inc.
https://doi.org/10.1371/journal.pone.0040410.g008
Недавно предсказанные солевые мосты в GFP14R были ≤4 Å, тогда как они были ≥4 Å для GFP con в семи позициях. Шесть новых взаимодействий солевого мостика в GFP14R были сформированы в петлевых областях белка, за исключением солевого мостика K41-D36, где вариант не содержит мутации в K41.Аргинин принимает большее количество конформаций из-за своего размера, гибкости боковой цепи и большего количества атомов, способных нести заряды, чем лизин [16]. Предполагалось, что гибкость петли и геометрическое преимущество аргинина увеличивают вероятность большего количества электростатических взаимодействий в GFP14R.
Расстояния между атомами, участвующими в новых ионных парах GFP14R, были дополнительно проанализированы путем выполнения молекулярно-динамического моделирования при комнатной температуре для повышения точности вновь образованных солевых мостиков.Было обнаружено, что два (R52-D216 и R140-D173) солевых мостика, предсказанные с использованием смоделированной структуры GFP14R, имеют меньшее изменение расстояния во время моделирования 10 наносекунд, чем GFP con , в то время как расстояние между атомами для оставшейся соли- мосты оказались похожими на GFP con (Рисунок S5). Эти результаты динамического моделирования предполагают возможность того, что предсказанные солевые мосты с использованием статических смоделированных структур не были согласованными для всех случаев.
Обсуждение
Изучено влияние мутации поверхностного лизина GFP на аргинин на стабильность и структуру белка. Вариант GFP14R был получен заменой 14 поверхностных лизинов природного GFP на аргинин и сохранением 5 поверхностных лизинов, участвующих в формировании сверхстабильной сердцевинной структуры GFP. Вариант показал низкую скорость складывания (рис. S6), но его окончательная структура предположительно не изменилась. Тесты на стабильность варианта показали, что мутагенез поверхностного лизина до аргинина был эффективным в повышении стабильности GFP против мочевины, щелочного pH и детергентов, но не влиял на термостабильность GFP.Структурный анализ электростатических взаимодействий подтвердил не только то, что в результате мутаций возникли дополнительные солевые мостики и водородные связи, но и геометрические свойства группы гуанидиния в Arg. Эти результаты показали следующее: 1) введение аргининов на поверхностные остатки лизина может стабилизировать белок, но эффект может различаться в зависимости от условий денатурации; 2) мутагенез поверхностного лизина в аргинин может вызывать изменения электростатических взаимодействий дополнительным образом, что может быть фактором повышения стабильности; и 3) такой мутагенез может неблагоприятно повлиять на укладку белка.
Интересно, что термическая стабильность GFP не была изменена мутагенезом, тогда как стабильность против мочевины, щелочного pH и детергентов была улучшена. Одним из объяснений этого может быть различие в механизме термической денатурации и другой денатурации, опосредованной химическими веществами. В случае тепловой денатурации энергия, передаваемая белку, вызывает общее движение атомов в белке, что приводит к неспецифическому и одновременному нарушению различных взаимодействий для сворачивания белка.Следовательно, усиленные электростатические взаимодействия, вызванные мутагенезом, могут не так эффективно предотвращать термическую денатурацию в диапазоне обнаружения. Кроме того, взаимодействия, образующиеся на поверхности белка, как в этом случае, лишь незначительно влияют на термостабильность белка [34], что может еще больше усилить такой эффект. С другой стороны, мочевина, щелочной pH и ионные детергенты начинают денатурировать белки, взаимодействуя со специфическими атомами, участвующими в электростатических взаимодействиях в белке.Следовательно, лизины и аргинины, обычно участвующие в электростатических взаимодействиях, могут подвергаться более специфическому и чувствительному воздействию добавленных химических денатурантов. Конечно, для более точного понимания эффекта потребуются дальнейшие исследования.
В исследовании влияния pH GFP14R показал более высокую стабильность, чем GFP con при pH 13,0. Теоретическая оценка протонированных состояний основных остатков Arg и Lys на основе pKa их боковой цепи показала, что 20% боковых цепей аргинина протонированы при pH 13.0, тогда как большинство боковых цепей Lys депротонируются при этом pH (фигура S7). Это может привести к различию ионных взаимодействий на поверхности двух белков GFP и объяснить более низкую стабильность GFP con , чем GFP14R при щелочном pH. С другой стороны, GFP con был относительно стабилен при pH 22 (рис. 5A), несмотря на почти полное депротонирование основного остатка Lys при этом pH (рис. S7), что предполагает отсутствие прямой корреляции между ионными состояниями Теоретически оценены остатки Lys и стабильность GFP.Разница между pH окружающей среды на поверхности белка и pH в объеме растворителя может привести к такому результату. Кроме того, на гидрофильные аминокислоты, отличные от Arg и Lys, также может влиять pH, что также может вызывать такой эффект.
Исследование влияния ионного детергента на стабильность GFP показало, что отрицательно заряженные детергенты, такие как SDS, имели более заметный эффект, чем положительно заряженный детергент. Это можно объяснить, рассмотрев механизм разворачивания белка, вызванный детергентом.Хотя точный механизм неясен, было высказано предположение, что разворачивание белка ионными детергентами представляет собой комбинацию гидрофобных и электростатических взаимодействий [35] — [37]. Предыдущее исследование денатурации белка, индуцированной SDS, показало, что SDS связывается с полипептидной цепью вначале за счет гидрофобного взаимодействия с последующим отталкиванием заряда-заряда между мицеллами SDS и анионными боковыми цепями белка по мере увеличения концентрации SDS [38] — [40]. При поверхностном мутагенезе Lys в Arg частота аргинина была увеличена с 2.От 5% до 8,4% для GFP14R, в результате чего среда на поверхности белка становится все более положительно заряженной. Следовательно, отрицательно заряженная гидрофильная головка анионного детергента может более благоприятно связываться с этими положительно заряженными боковыми цепями аргинина в GFP14R, тем самым уменьшая возможность отталкивания заряда-заряда, необходимого для дальнейшей денатурации. Это может быть дополнительным фактором повышения стабильности GFP по отношению к анионным детергентам в дополнение к эффекту повышенных электростатических взаимодействий в белке, что может привести к более заметному эффекту для анионных детергентов.
При анализе электростатических взаимодействий GFP14R с использованием модели минимизированной энергии было предсказано, что 7 новых солевых мостиков были образованы путем добавления аргининов в 6 остатков лизина (K3, K52, K79, K131, K140 и K209) GFP. на . Чтобы оценить, насколько новообразованные солевые мостики способствуют стабильности GFP14R, был создан вариант GFP6R, имеющий аргинины вместо лизинов в этих шести положениях, и его стабильность сравнивали с GFPcon и GFP14R.Стабильность GFP6R тестировали в денатурирующих условиях, таких как 1% SDS и щелочной pH 13,0, условиях, в которых GFP14R показал преимущественно более высокую стабильность. Интересно, что стабильность GFP6R не была улучшена и аналогична контролю GFP (рисунок S8). Эти результаты означают, что предсказанные новые солевые мостики не являются эффективным стабилизирующим фактором для повышенной стабильности GFP14R. Согласно молекулярно-динамическому исследованию вариаций расстояния между ионными парами (рис. S5), существует вероятность того, что только два солевых мостика эффективны среди 6 солевых мостиков, что подтверждает этот экспериментальный результат.Мы предполагаем, что повышенная стабильность GFP14R была вызвана коллективными эффектами новых электростатических взаимодействий, включая взаимодействия аминокислоты с водой, а также взаимодействия аминокислоты с аминокислотами, генерируемые недавно введенными 14 аргининами. Для объяснения такого рода коллективных эффектов могут потребоваться дальнейшие подробные исследования.
Из исследования экспрессии GFP14R, белок GFP продуцировался в основном в нерастворимой форме, а продуктивность растворимой формы была низкой. Скорость сворачивания GFP14R была подтверждена как низкая по сравнению с GFP con (Рисунок S6).С другой стороны, GFP14R показал более высокую стабильность к химическим денатурантам и сопоставимую термостабильность по сравнению с GFP con . Эти результаты показывают, что мутации, внесенные на поверхность, отрицательно повлияли на сворачивание белка, но белок был стабильным после сворачивания. Это означает, что измененные электростатические взаимодействия, вызванные мутагенезом, стабилизировали GFP кинетически, а не термодинамически. Для более точного понимания механизма стабилизации могут потребоваться дальнейшие термодинамические и кинетические исследования.
Стратегия мутагенеза в настоящем исследовании все еще имеет ограничения, применимые непосредственно к стабилизации белка. Например, скорость сворачивания белка значительно снизилась из-за мутагенеза, что может вызвать проблему продукции целевого белка. Кроме того, в качестве целевого белка был протестирован только GFP. Эффект мутагенеза может различаться в зависимости от свойств целевых белков. Многие факторы, такие как исходная стабильность белка, структура белка, количество и расположение поверхностных остатков лизина, могут по-разному влиять на эффективность укладки и стабильность целевых белков в зависимости от мутагенеза.Следовательно, существует необходимость дальнейшего изучения этих эффектов с использованием других белков. Тем не менее, это исследование имеет смысл, поскольку влияние Lys и Arg на стабильность белка изучалось более непосредственно по сравнению с другими исследованиями. Это исследование подчеркивает возможность того, что мутагенез поверхностного лизина с образованием аргининов можно рассматривать как один из параметров в инженерии стабильности белка.
Материалы и методы
Материалы
Бактерия-хозяин E. coli штамм DH5α была использована для получения плазмидной ДНК.Штамм BL21 (DE3) E. coli использовали в качестве хозяина экспрессии для продукции рекомбинантных вариантов GFP. Реагенты для ПЦР и ДНК-лигаза Т4 были приобретены в New England Biolabs. Эндонуклеазы рестрикции были получены от Fermentas. IPTG (изопропил-D-тиогалактопиранозид), додецилсульфат натрия (SDS), додецилбензолсульфонат натрия (SDBS), бромид цетилтриметиламмония (CTAB), додецилтриметиламмонийхлорид (DTAC) и другие химические вещества были приобретены у Sigma Chemicals (St.Луис, Миссури, США). Аффинная колонка с Ni-NTA HisTrapHP была предоставлена компанией GE healthcare (Швеция). Вектор pET30b был получен от Novagen.
Конструирование плазмиды
Гены GFP con , GFP19R, GFP14R и GFP6R были синтезированы Genscript Corporation (Нью-Джерси, США). Синтезированные гены клонировали в сайты рестрикции NdeI и XhoI вектора pET30b с гексагистидиновой меткой на С-конце.
Экспрессия и очистка вариантов GFP
Варианты GFP, клонированные в вектор pET30b, трансформировали в штамм BL21 (DE3) для экспрессии.Белки экспрессировали при 37 ° C в течение пяти часов, если не указано иное, после индукции 1 мМ IPTG при O.D. 0,6–0,8. Лизат клеток был приготовлен из осадка клеток, соответствующего 1 мл 3 O.D. клетки с использованием набора для экстракции белков BugBuster (Novagen) и центрифугировали при 12000 об / мин в течение 30 минут для разделения растворимой и нерастворимой фракций для анализа SDS-PAGE. Вариант GFP14R экспрессировали при 25 ° C в течение 20 часов для увеличения выхода белка. Клетки собирали центрифугированием при 5000 об / мин в течение 15 минут и ресуспендировали в буфере PBS (фосфатно-солевой буфер — 137 мМ NaCl, 2.7 мМ KCl, 10 мМ Na 2 HPO 4 , 2 мМ KH 2 PO 4 и pH 7,4). Клетки разрушали с помощью French Press и центрифугировали при 20000 об / мин в течение 30 минут при 4 ° C для сбора растворимой фракции. Затем растворимые белки очищали с использованием колонки Ni-NTA объемом 5 мл, уравновешенной 50 мМ натрий-фосфатным буфером (50 мМ фосфата натрия, полученного из 1 М Na 2 HPO 4 и 1 М NaH 2 PO 4 , 300 мМ NaCl, pH 7,4), содержащий 10 мМ имидазол.Затем колонку промывали 5 объемами колонки 50 мМ натрий-фосфатного буфера, содержащего 50 мМ имидазола, и элюирование проводили, используя 50 мМ натрий-фосфатный буфер, содержащий 500 мМ имидазол. Фракции элюирования анализировали с помощью SDS-PAGE, фракции, обогащенные рекомбинантными белками, собирали и подвергали диализу буфером PBS, pH 7,4. Концентрации очищенных образцов белка определяли с использованием спектрофотометра UV / VIS при 280 нм с использованием коэффициента экстинкции, рассчитанного на основе последовательностей белка.
Измерение флуоресценции
Флуоресценцию целых клеток вариантов GFP измеряли с использованием счетчика Perkin Elmer / Wallac Victor 2 Multilabel (1420-011) с возбуждением и испусканием при 485 нм и 515 нм соответственно из щелей 5,0 нм после ресуспендирования осадка клеток в буфере PBS. Спектр возбуждения и испускания очищенных образцов белка 5 мкМ измеряли с использованием спектрофлуориметра Perkin Elmer LS-55 со щелями возбуждения / испускания 2,5 нм. Если не указано иное, флуоресценцию измеряли с образцами белка 5 мкМ с использованием 96-луночных планшетов с помощью счетчика Perkin Elmer / Wallac Victor 2 Multilabel Counter для всех анализов стабильности.
Термическая стабильность вариантов GFP
Температурно-зависимый анализ проводили путем инкубации образцов при температуре от 40 ° C до 100 ° C с интервалами 10 ° C в течение 30 минут, а флуоресценцию измеряли при комнатной температуре. Анализ, зависящий от времени, проводили путем инкубации образцов при 70 ° C в течение 30 минут с 10-минутными интервалами, а флуоресценцию измеряли при комнатной температуре.
Стабильность вариантов GFP при различных значениях pH
Пробы белка инкубировали с рядом буферов, содержащих pH 8.0 (50 мМ трис-HCl), pH 10,0 (50 мМ карбонат натрия), pH 12,0 и pH 13,0 (50 мМ KCl) при 60 ° C в течение 30 минут, и флуоресценцию измеряли при комнатной температуре. Кроме того, был проведен зависимый от времени анализ путем инкубации образцов с 50 мМ KCl (pH 13,0) при 60 ° C до 60 минут с 10-минутными интервалами, а флуоресценция измерялась при комнатной температуре.
Устойчивость вариантов GFP к воздействию мочевины и ионных детергентов
Для определения устойчивости вариантов к химическим денатурантам образцы инкубировали с 6 М мочевиной при 50 ° C в течение различных интервалов времени до 30 минут и измеряли флуоресценцию при комнатной температуре.Стабильность вариантов также измеряли в присутствии ионных детергентов, таких как додецилсульфат натрия (SDS) и додецилбензолсульфонат натрия (SDBS), которые являются анионными, и бромид цетилтриметиламмония (CTAB) и хлорид додецилтриметиламмония (DTAC), которые являются катионный. Для этого образцы инкубировали с 1% детергента при 50 ° C в течение различных интервалов времени до 60 минут. Кроме того, стабильность вариантов измеряли при различных концентрациях SDS, начиная от 1% до 5%, инкубируя образцы при 50 ° C в течение 30 минут.Во всех вышеупомянутых экспериментах pH образцов белка поддерживали на уровне 7,4 с помощью буфера PBS.
Рефолдинг вариантов GFP
Эффективность сворачивания вариантов GFP оценивали путем денатурирования образцов белка в суровых условиях и повторного укладывания после разбавления. Образцы белка с концентрацией 25 мкМ денатурировали путем инкубации при 95 ° C в течение 5 минут в буфере PBS, содержащем 8 М мочевину и 5 мМ дитиотреитол (DTT). Денатурированные образцы разбавляли до 100 раз с использованием буфера PBS, содержащего 5 мМ ДДТ, и флуоресценцию измеряли при комнатной температуре в течение 30 минут с интервалом времени 3 секунды с использованием спектрофлуориметра Perkin Elmer LS-55 (возбуждение 490 нм, испускание 511 нм, 2.Щель возбуждения / испускания 5 нм). Восстановленную флуоресценцию нормализовали к 1 путем деления на максимальное значение флуоресценции спектров.
Моделирование и моделирование молекулярной динамики вариантов GFP
Трехмерная структура GFP con и вариант моделировались с использованием структуры GFP с PDB Id 1GFL в качестве шаблона [29]. Модели были созданы с использованием протокола Build Homology Models из модуля Protein Modeling Discovery Studio 2.0 (Accelrys Software Inc., Сан-Диего, Калифорния, США). Минимизация энергии была выполнена с использованием протокола минимизации из модуля моделирования Discovery Studio 2.0 в соответствии с методом сопряженных градиентов после набора текста с автоматическим назначением силового поля Momany & Rone CHARMm и отсечки несвязанных взаимодействий, установленной на 10–12 Å. . В качестве растворителя использовались диэлектрики, зависящие от расстояния, с диэлектрической постоянной, установленной на 4, чтобы имитировать экранирующие эффекты растворителя на неочищенном уровне.Циклы минимизации энергии выполнялись для 3000 шагов до тех пор, пока допуск градиента RMS не стал ≤0,1 ккал · моль -1 Å -1 . Модели in silico вариантов GFP показали лишь небольшое отклонение от структуры шаблона со среднеквадратичным отклонением приблизительно 0,1 Å. Взаимодействие солевой мостик и водородная связь анализировали с помощью Discovery Studio 2.0 (Accelrys Software Inc., Сан-Диего, Калифорния, США). Предсказанные варианты GFP, сгенерированные выше путем молекулярного моделирования, были приняты в качестве стартовой структуры для выполнения молекулярно-динамического (МД) моделирования с использованием пакета моделирования МД GROMACS 4.5.5 [41]. С помощью силового поля GROMOS96 53a6 структура была центрирована в кубической элементарной ячейке на расстоянии 1,0 нм от края ящика и сольватирована с использованием модели воды SPC / E. Затем систему нейтрализовали Na + и Cl — в качестве противоионов и минимизировали энергию, используя алгоритм наискорейшего спуска для 2000 шагов. Система с минимизированной энергией была уравновешена с использованием моделирования с ограничением положения в ансамбле NVT (постоянное число частиц, объем и температура) в течение 100 пикосекунд для стабилизации температуры на уровне 300 K с помощью термостата Берендсена, за которым следует ансамбль NPT (постоянное число частиц, давления и температуры) в течение 100 пикосекунд, чтобы стабилизировать давление на 1.0 бар с коэффициентом сопряжения по давлению Парринелло-Рахмана. Наконец, произвольное моделирование МД было выполнено в течение 10 наносекунд с термостатом Берендсена 300 К и давлением 1,0 бар с коэффициентом связи давления Парринелло-Рахмана. Утилиты trjconv, g_rms, make_ndx и g_dist из GROMACS 4.5.5 использовались для анализа результатов MD.
Дополнительная информация
Рисунок S1.
Гель SDS-PAGE показывает профиль экспрессии белка. A) Дорожки 1 и 2 показывают растворимую фракцию GFP con и GFP19R соответственно, а дорожки 4 и 5 указывают нерастворимую фракцию GFP con и GFP19R, соответственно, индуцированную 1 мМ IPTG при 37 ° C для 5 часов.Дорожки 3 и 6 показывают растворимую и нерастворимую фракции GFP19R, соответственно, индуцированные 1 мМ IPTG при 25 ° C в течение 5 часов. B) Дорожка 1 и 2 указывают растворимую фракцию GFP , con и GFF14R, соответственно, а дорожка 3 указывает нерастворимую фракцию GFP14R, индуцированную 1 мМ IPTG при 37 ° C в течение 5 часов. Дорожка М — белковый маркер.
https://doi.org/10.1371/journal.pone.0040410.s001
(TIF)
Рисунок S2.
A) Гель SDS-PAGE показывает профиль экспрессии белка GFP con и GFP14R, индуцированный 1 мМ IPTG при 25 ° C в течение 5 часов.Дорожки 1 и 2 показывают растворимую фракцию GFP con и GFF14R, соответственно, а дорожки 3 и 4 указывают нерастворимую фракцию GFP con и GFF14R, соответственно. Дорожки 5 и 6 показывают очищенный белок GFP con и GFP14R соответственно. Дорожка М — белковый маркер. B) Флуоресценция целых клеток GFP con и GFP14R, нормализованная по оптической плотности при 600 нм. (au — условные единицы).
https://doi.org/10.1371/journal.pone.0040410.s002
(TIF)
Рисунок S6.
Денатурация и рефолдинг вариантов GFP. Эффективность сворачивания вариантов GFP измеряли путем денатурации в 8 М мочевине при 95 ° C с последующей ренатурацией путем разбавления при комнатной температуре. Нормированную флуоресценцию в произвольных единицах (а.е.) наносили на график зависимости от времени.
https://doi.org/10.1371/journal.pone.0040410.s006
(TIF)
Рисунок S7.
Состояния протонирования были оценены теоретически с использованием уравнения log ([AH] / [A — ]) = pKa-pH. Значения pKa боковой цепи лизина (pKa 10,53) и аргинина (pKa 12,48) использовали для оценки отношения [AH] к [A —] для каждого pH и преобразовывали в процент.
https://doi.org/10.1371/journal.pone.0040410.s007
(TIF)
Рисунок S8.
A) Стабильность вариантов GFP в присутствии 1% SDS при 50 ° C в течение 30 минут. Флуоресценция в нулевой момент времени в 1% SDS была принята за 100%. B) Стабильность вариантов GFP в присутствии 50 мМ буфера KCl pH 13.0 при 60 ° C в течение 30 минут. Флуоресценция в нулевой момент времени при pH 13,0 была принята за 100%. (Планка погрешностей — стандартное отклонение трех независимых экспериментов).
https://doi.org/10.1371/journal.pone.0040410.s008
(TIF)
Благодарности
Авторы хотели бы поблагодарить профессора Хёндона Юна, Школа биотехнологии, Университет Йоннам, Южная Корея, за предоставление флуоресцентного спектрофотометра.
Вклад авторов
Задумал и спроектировал эксперименты: SS S-GL.Проведены эксперименты: СС ГР НС. Проанализированы данные: SS S-GL GR. Предоставленные реагенты / материалы / инструменты анализа: S-GL. Написал статью: SS S-GL.
Список литературы
- 1.
Маррс Б., Делагрейв С., Мерфи Д. (1999) Новые подходы к открытию промышленных ферментов. Curr Opin Microbiol 2: 241–245. - 2.
Черри Дж. Р., Фиданцеф А. Л. (2003) Направленная эволюция промышленных ферментов: обновление. Curr Opin Biotechnol 14: 438–443. - 3.
Кирк О., Borchert TV, Fuglsang CC (2002) Промышленные применения ферментов.Curr Opin Biotechnol 13: 345–351. - 4.
Эйсинк В., Бьорк А., Гасейднес С., Сиреваг Р., Синстад Б. и др. (2004) Рациональная инженерия стабильности ферментов. J Biotechnol 113: 105–120. - 5.
Латтман Э. Э., Роуз Г. Д. (1993) Сворачивание белка — в чем вопрос? Proc Natl Acad Sci U S A 90: 439–441. - 6.
Baker D (2000) Удивительная простота сворачивания белка. Nature 405: 39–42. - 7.
Radford SE (2000) Сворачивание белка: достигнутый прогресс и перспективы на будущее.Тенденции Biochem Sci 25: 611–618. - 8.
Даггетт В., Фершт А.Р. (2003) Существует ли объединяющий механизм для сворачивания белков? Тенденции Biochem Sci 28: 18–25. - 9.
Тайгерстром А., Шварц Ф., Карлссон Г., Оквист М., Эльварес-Руа С. и др. (2004) Влияние новой дисульфидной связи и инженерных электростатических взаимодействий на термостабильность Азурина. Биохимия 43: 12563–12574. - 10.
Northey JGB, Di Nardo AA, Davidson AR (2002) Гидрофобная упаковка ядра в переходном состоянии сворачивания домена Sh4.Nat Struct Biol 9: 126–130. - 11.
Джексон С.Е., ЭльМасри Н., Фершт А.Р. (1993) Структура гидрофобного ядра в переходном состоянии для свертывания ингибитора химотрипсина 2: критический тест метода анализа белковой инженерии. Биохимия 32: 11270–11278. - 12.
Махатадзе Г.И., Лоладзе В.В., Грибенко А.В., Лопес М.М. (2004) Механизм термостабилизации в разработанном протеине холодового шока с оптимизированными поверхностными электростатическими взаимодействиями. J Mol Biol 336: 929–942. - 13.
Кумар С., Цай С.Дж., Нусинов Р. (2000) Факторы, повышающие термостабильность белка. Protein Eng 13: 179–191. - 14.
Yokota K, Satou K, Ohki S-y (2006) Сравнительный анализ термостабильности белков: различия в содержании и замене аминокислот на поверхности и в центральных областях термофильных и мезофильных белков. Наука и технология перспективных материалов 7: 255–262. - 15.
Стриклер С.С., Грибенко А.В., Грибенко А.В., Кейффер Т.Р., Томлинсон Дж. И др.(2006) Стабильность белка и поверхностная электростатика: заряженная взаимосвязь. Биохимия 45: 2761–2766. - 16.
Barlow DJ, Thornton JM (1983) Ионные пары в белках. J Mol Biol 168: 867–885. - 17.
C.l. Бордерс Дж., Бродвотер Дж. А., Бекени П. А., Лосось Дж. Э., Ли А. С. и др. (1994) Структурная роль аргинина в белках: множественные водородные связи с карбонильными атомами кислорода в основной цепи. Protein Sci 3: 541–548. - 18.
Дональд Дж. Э., Кулп Д. В., ДеГрадо В. Ф. (2011) Солевые мосты: геометрически определенные, проектируемые взаимодействия.Белки: Struct Funct Bioinform 79: 898–915. - 19.
Мусафия Б., Бюхнер В., Арад Д. (1995) Сложные солевые мостики в белках: статистический анализ структуры и функции. J Mol Biol 254: 761–770. - 20.
Matsutani M, Hirakawa H, Nishikura M, Soemphol W., Ali IAI, et al. (2011) Увеличение количества солевых мостиков на основе аргинина способствует термотолерантности термотолерантных уксуснокислых бактерий, Acetobacter tropicalis SKU1100. Biochem Biophys Res Commun 409: 120–124. - 21.
Кумар С., Нусинов Р. (1999) Стабильность солевого мостика в мономерных белках. J Mol Biol 293: 1241–1255. - 22.
Strub C, Alies C, Lougarre A, Ladurantie C, Czaplicki J, et al. (2004) Мутация экспонированных гидрофобных аминокислот на аргинин для повышения стабильности белка. BMC Biochem 5: 9. - 23.
Chan C-H, Yu T-H, Wong K-B (2011) Стабилизирующий солевой мостик повышает термостабильность белка за счет уменьшения изменения теплоемкости разворачивания.PLoS One 6: e21624. - 24.
Turunen O, vuorio M, Fenel F, Leisola M (2002) Разработка множества аргининов на поверхности Ser / Thr 1,4-β-ксиланазы II Trichoderma ressei endo- увеличивает термотолерантность и сдвигает оптимум pH в сторону щелочного pH . Protein Eng 15: 141–145. - 25.
Reid BG, Flynn GC (1997) Образование хромофора в зеленом флуоресцентном белке. Биохимия 36: 6786–6791. - 26.
Waldo GS, Standish BM, Berendzen J, Terwilliger TC (1999) Быстрый анализ сворачивания белка с использованием зеленого флуоресцентного белка.Нат Биотех 17: 691–695. - 27.
Rusmini F, Zhong Z, Feijen J (2007) Стратегии иммобилизации белков для белковых биочипов. Биомакромолекулы 8: 1775–1789. - 28.
Pavoor TV, Cho YK, Shusta EV (2009) Разработка биосенсоров на основе GFP, обладающих связывающими свойствами антител. Proc Natl Acad Sci U S A 106: 11895–11900. - 29.
Ян Ф, Мосс Л.Г., Филлипс Г.Н. (1996) Молекулярная структура зеленого флуоресцентного белка. Нат Биотех 14: 1246–1251. - 30.
P. Cormack B, H.Valdivia R, Falkow S (1996) оптимизированные для FACS мутанты зеленого флуоресцентного белка (GFP). Gene 173: 33–38. - 31.
Хуанг Дж-р, Сю С-ТД, Христодулу Дж., Джексон С.Е. (2008) Чрезвычайно медленное обменное ядро и кислотно-денатурированное состояние зеленого флуоресцентного белка. HFSP J 2: 378–387. - 32.
Бойл Дж. (2005) Принципы биохимии Ленингера (4-е изд.): Нельсон, Д., и Кокс, М. Биохимия и молекулярная биология, образование 33: 74–75. - 33.
Макдональд И.К., Торнтон Дж.М. (1994) Удовлетворение водородно-связывающего потенциала в белках. J Mol Biol 238: 777–793. - 34.
Такано К., Цучимори К., Ямагата Ю., Ютани К. (2000) Вклад солевых мостиков вблизи поверхности белка в конформационную стабильность. Биохимия 39: 12375–12381. - 35.
Тимашев С.Н. (2002) Гидратация белков, термодинамическое связывание и предпочтительная гидратация. Биохимия 41: 13473–13482. - 36.Gitlin I, Gudiksen KL, Whitesides GM (2006) Перацетилированная бычья карбоангидраза (BCA-Ac18) кинетически более стабильна, чем нативные BCA, по отношению к додецилсульфату натрия. J. Phys Chem B 110: 2372–2377.
- 37.
Otzen DE, Christiansen L, Schülein M (1999) Сравнительное исследование разворачивания эндоглюканазы Cel45 из Humicola insolens в денатуранте и поверхностно-активном веществе. Protein Sci 8: 1878–1887. - 38.
Бхуян А.К. (2010) О механизме денатурации белков, индуцированной SDS.Биополимеры 93: 186–199. - 39.
Нильсен М., Андерсен К., Вест П., Отцен Д. (2007) Развертывание белков β-листа в SDS. Biophys J 92: 3674–3685. - 40.
Xia K, Manning M, Hesham H, Lin Q, Bystroff C и др. (2007) Идентификация субпротеома кинетически стабильных белков с помощью диагонального 2D SDS / PAGE. Proc Natl Acad Sci U S A 104: 17329–17334. - 41.
Хесс Б., Кутцнер С., ван дер Споэль Д., Линдал Е. (2008) GROMACS 4: Алгоритмы для высокоэффективного, сбалансированного по нагрузке и масштабируемого молекулярного моделирования.J Chem Theory Comput 4: 435–447.
18.1: Свойства аминокислот
Цели обучения
- Для распознавания аминокислот и их классификации на основе характеристик их боковых цепей.
Белки всех живых существ, от бактерий до людей, состоят из одного и того же набора из 20 аминокислот, названных так потому, что каждая из них содержит аминогруппу, присоединенную к карбоновой кислоте. Аминокислоты в белках представляют собой α-аминокислоты, что означает, что аминогруппа присоединена к α-углероду карбоновой кислоты.Человек может синтезировать только половину необходимых аминокислот; остальные должны быть получены с пищей и известны как незаменимые аминокислоты. Однако две дополнительные аминокислоты были обнаружены в белках в ограниченных количествах: селеноцистеин был открыт в 1986 году, а пирролизин был открыт в 2002 году.
Аминокислоты представляют собой бесцветные нелетучие кристаллические вещества, плавящиеся и разлагающиеся при температуре выше 200 ° C. Эти температуры плавления больше похожи на температуры неорганических солей, чем у аминов или органических кислот, и указывают на то, что структуры аминокислот в твердом состоянии и в нейтральном растворе лучше всего представлены как имеющие как отрицательно заряженную группу, так и положительно заряженную группу.Такой вид известен как цвиттерион.
Классификация
Помимо амино и карбоксильных групп, аминокислоты имеют боковую цепь или группу R, присоединенную к α-углероду. Каждая аминокислота имеет уникальные характеристики, обусловленные размером, формой, растворимостью и ионизационными свойствами ее группы R. В результате боковые цепи аминокислот оказывают сильное влияние на структуру и биологическую активность белков. Хотя аминокислоты можно классифицировать по-разному, один общий подход состоит в том, чтобы классифицировать их в зависимости от того, является ли функциональная группа боковой цепи при нейтральном pH неполярной, полярной, но незаряженной, отрицательно заряженной или положительно заряженной.Структура и названия 20 аминокислот, их одно- и трехбуквенные сокращения, а также некоторые их отличительные особенности приведены в Таблице \ (\ PageIndex {1} \).
Первой выделенной аминокислотой в 1806 году был аспарагин. Он был получен из белка, содержащегося в соке спаржи (отсюда и название). Глицин, основная аминокислота, содержащаяся в желатине, был назван из-за его сладкого вкуса (по-гречески glykys , что означает «сладкий»). В некоторых случаях аминокислота, содержащаяся в белке, на самом деле является производным одной из 20 обычных аминокислот (одним из таких производных является гидроксипролин).Модификация происходит после того, как аминокислота собрана в белок.
Конфигурация
Обратите внимание в таблице \ (\ PageIndex {1} \), что глицин является единственной аминокислотой, у которой α-углерод , а не хиральный. Следовательно, за исключением глицина, аминокислоты теоретически могут существовать либо в D-, либо в L-энантиомерной форме и вращать плоско-поляризованный свет. Как и в случае с сахарами, химики использовали L-глицеральдегид в качестве эталонного соединения для определения абсолютной конфигурации аминокислот.Его структура очень напоминает структуру аминокислоты, за исключением того, что в последней аминогруппа занимает место группы ОН на хиральном углероде L-глицеральдегида, а карбоновая кислота заменяет альдегид. Современные стереохимические определения с использованием правил приоритета Кана-Ингольда-Прелога, повсеместно используемых в химии, показывают, что все встречающиеся в природе хиральные аминокислоты являются S, за исключением Cys, который представляет собой R.
Мы узнали, что все встречающиеся в природе сахара принадлежат к серии D.Поэтому интересно, что почти все известные растительные и животные белки полностью состоят из L-аминокислот. Однако некоторые бактерии содержат D-аминокислоты в своих клеточных стенках, а некоторые антибиотики (например, актиномицин D и грамицидины) содержат различные количества D-лейцина, D-фенилаланина и D-валина.
Сводка
Аминокислоты могут быть классифицированы на основе характеристик их отличительных боковых цепей как неполярные, полярные, но незаряженные, отрицательно заряженные или положительно заряженные.Аминокислоты, содержащиеся в белках, являются L-аминокислотами.
Упражнения по обзору концепции
-
Какова общая структура α-аминокислоты?
-
Определите аминокислоту, которая подходит под каждое описание.
- , также известный как аспартат
- почти такое же сильное основание, как гидроксид натрия
- не имеет хирального углерода
-
- аспарагиновая кислота
- аргинин
- глицин
-
Напишите боковую цепь каждой аминокислоты.
- серин
- аргинин
- фенилаланин
-
Напишите боковую цепь каждой аминокислоты.
- аспарагиновая кислота
- метионин
- валин
-
Изобразите структуру каждой аминокислоты.
- аланин
- цистеин
- гистидин
-
Изобразите структуру каждой аминокислоты.
- треонин
- глутаминовая кислота
- лейцин
-
Укажите аминокислоту, боковая цепь которой содержит (n)
- амидная функциональная группа.
- ароматическое кольцо.
- карбоксильная группа.
-
Укажите аминокислоту, боковая цепь которой содержит (n)
- Группа OH
- разветвленная цепь
- аминогруппа
-
- СН 2 ОН —
-
- аспарагин или глутамин
- фенилаланин, тирозин или триптофан
- аспарагиновая кислота или глутаминовая кислота
аминокислот
Введение:
Каждая аминокислота имеет по крайней мере одну аминную и одну кислотную функциональную группу.
группа, как следует из названия.Различные свойства являются результатом
вариации в строении различных R-групп. Группа R
часто называют боковой цепью аминокислоты. Аминокислоты
имеют специальные общеупотребительные имена, однако сокращенное трехбуквенное
поскольку имя используется большую часть времени. Вторая аббревиатура
, одна буква, используется в длинных белковых структурах.
таблица слева для структуры, названий и сокращений
20 аминокислот.
Существует четыре основных класса аминокислот.
определяется разными боковыми цепями: (1) неполярными и нейтральными,
(2) полярный и нейтральный, (3) кислый и полярный, (4) основной и полярный.
Принципы полярности:
Чем больше электроотрицательность разница между
атомов в связи, более полярная связь. Частичные отрицательные заряды
находятся на наиболее электроотрицательных атомах, остальные частично
положительный.Проверьте полярность функциональных
группы.
Неполярные боковые цепи:
Боковые цепи, содержащие чистых углеводородных алкильных групп
(алкановые ответвления) или ароматические (бензольные кольца) неполярные .
Примеры включают валин, аланин, лейцин, изолейцин, фенилаланин.
Число алкильных групп также влияет на полярность.
Чем больше присутствует алкильных групп, тем неполярнее аминокислота.
будет.Этот эффект делает валин более неполярным, чем аланин;
лейцин более неполярен, чем валин.
QUES. Перечислите все аминокислоты с неполярными боковыми цепями.
| Распределите следующие по возрастанию неполярность, т.е. 1 = наименее неполярный, 4 = наиболее неполярный. лей; phe; val; аля |
Ответ 1 = аля 2 = val 3 = лей 4 = phe Рейтинг основан на увеличивающиеся числа CHs. |
Полярные боковые цепи:
Боковые цепи, которые имеют различные функциональные группы, такие как кислот,
амиды, спирты и амины придадут более полярный
символ к аминокислоте. Рейтинг полярности будет зависеть
об относительном ранжировании полярности для различных функциональных групп
как определено в функциональных группах.
Кроме того, количество атомов углерода в алкане или
ароматическая часть боковой цепи должна рассматриваться вместе с
с функциональной группой.
Пример: аспарагиновая кислота более полярна, чем серин, потому что
кислотная функциональная группа более полярна, чем спиртовая группа.
Пример: серин более полярен, чем треонин, поскольку треонин
имеет на одну метильную группу больше, чем серин. Метильная группа дает
немного более неполярный характер треонину.
Пример: серин более полярен, чем тирозин, поскольку тирозин
имеет углеводородное бензольное кольцо.
QUES. Перечислите все аминокислоты по аббревиатуре, которые считаются
несколько полярными.
| Оцените следующие аминокислоты по возрастающая полярность. то есть 1 = более неполярный. ser; glu; жерех; лиз; ала; gln |
Ответ 1 = аля 2 = лиз 3 = сер 4 = глю 5 = жерех 6 = gln |
| Какая аминокислота наиболее нерастворима в воде: изолейцин или аланин? |
Ответ Изолейцин имеет больше CHs, поэтому больше нерастворим, чем ала. |
| Какая аминокислота наиболее растворима в воде: лиз или сер? |
Ответ Сер, алкогольная группа более полярен, чем амин в лиз. |
Свойства AA
Свойства AA
Этот список предоставляется в качестве учебного пособия, чтобы помочь
напоминают вам о свойствах боковых цепей / R-групп
двадцать аминокислот, которые присутствуют во вновь синтезированных белках.R-группы можно классифицировать по-разному, некоторые из
которые описаны ниже. Список запоминать не нужно,
но должен быть в состоянии предсказать свойства, исходя из ваших знаний о
структуры боковых цепей и основные принципы органического
химия. Чтобы узнать больше об аминокислотах, чтобы узнать, какие аминокислоты
кислота a сокращает однобуквенный код, и для просмотра его структуры щелкните
в теме!
- Все — G, A, V, L, I, P,
F, Y, W,
S, T, N, Q, C, M, D, E, H, K, R - Полярный / гидрофильный — N, Q, S, T, K, R, H, D, E, (C, Y) *
- Неполярный / гидрофобный — (G), A, V, L, I, P, Y, F, W, M, C
- H-соединение — C, W, N, Q, S, T, Y, K, R, H, D, E
- сера
содержащий — C, M - Заряжено при нейтральном pH Отрицательном / кислотном —
D, E, (C) - Заряжен при нейтральном pH Положительный / основной —
K, R, (H) - Ионизируемый — D, E, H, C, Y, K, R
- Ароматический — F, W, Y, (H, но без значительного УФ-поглощения)
- Алифатические — G, A, V, L, I, P
- Образует ковалентную поперечную связь (дисульфидная связь) —
C - Циклический — P
- Трудно запомнить однобуквенный код —
W, Y, K, N, Q, D, E
* Примечание: аминокислоты в скобках имеют указанный символ
в ограниченной степени.
| Домашняя страница MCB |
Домашняя страница UCD |
Авторские права © Charles S.
Гассер 2009, 2013.
Вот несколько глупых мнемоник, которые помогут запомнить некоторые аминокислоты или их
однобуквенные сокращения.
Фундаментальное свойство аргинина, выявленное посредством сольватации
Наша ДНК кодирует всего 20 аминокислот, которых достаточно, чтобы соединиться с десятками тысяч белков.Различия в их структуре и поведении приводят к изменчивости природы, но многие из их химических свойств было трудно изучить на самом фундаментальном уровне. Аргинин, аминокислота, имеющая более 100 изомеров, хрупкая и фрагментируется в газовой фазе. Достижения как в технике, так и в теории теперь позволили исследователям определить энергию ионизации аргинина, фундаментальную информацию, которая будет использоваться в технологиях химического зондирования для здоровья человека или восстановления окружающей среды.
Чтобы определить энергию ионизации, исследователи объединили метод, разработанный в Advanced Light Source (ALS), с новыми вычислительными методами, которые моделируют влияние молекул воды на аргинин в растворе. Вакуумная ультрафиолетовая (ВУФ) масс-спектрометрия и фотоэлектронная спектроскопия на Beamline 9.0 позволили получить представление о газовой фазе аргинина, испаряющемся из аэрозолей и при различных условиях pH. Комбинация экспериментов и теории позволила исследователям определить энергию ионизации аргинина в газовой фазе и проследить эффекты сольватации (добавления воды), такие как стабилизация молекулярного иона от фрагментации.
Расширение набора методов и теорий углубило наше фундаментальное понимание биомолекул; знание энергии ионизации аргинина имеет значение для окисления, восстановления и образования водородных связей в пептидах и белках. Эти знания могут быть распространены на другие мягкие рентгеновские исследования, относящиеся к кристаллографии и динамике белков. Кроме того, эти исследования дают информацию для разработки молекулярной электроники и датчиков, которые отслеживают процессы, имеющие отношение к здоровью и окружающей среде.
Аргинин — это аминокислота с более чем 100 изомерами, многие из которых стабильны при комнатной температуре. Из-за его тенденции к фрагментации в газовой фазе, аргинин было трудно изучать, но достижения в области техники и теории позволили исследователям анализировать его химические свойства с помощью вакуумной ультрафиолетовой (ВУФ) масс-спектрометрии и фотоэлектронной спектроскопии.
А. Баррозо, Б. Сю, А. О. Гунина, М. И. Якобс, К. Уилсон, О. Костко, М.