Молочная кислота
%D
%d.%M.%y
%h~:~%m
Молочную кислоту открыл шведский химик Карл Шееле. В 1780 году он выделил её из прокисшего молока в виде коричневого сиропа.
Молочная кислота — это á-гидроксикарбоновая кислота (á-гидроксипропановая кислота) с хиральным центром у второго атома углерода, и имеет, соответственно, два оптических изомера, однако чаще всего она используется в виде рацемата. Молочная кислота очень гигроскопична и обычно существует в виде водного растворов с концентрацией до мас. 90%. В связи с этим сложно установить температуру плавления, (в специализированной литературе указывается 18-33º градуса). Кроме того в таких растворах присутствует значительное количество лактоилмолочной кислоты и других олигомеров молочной кислоты.
Химические свойства весьма разнообразны.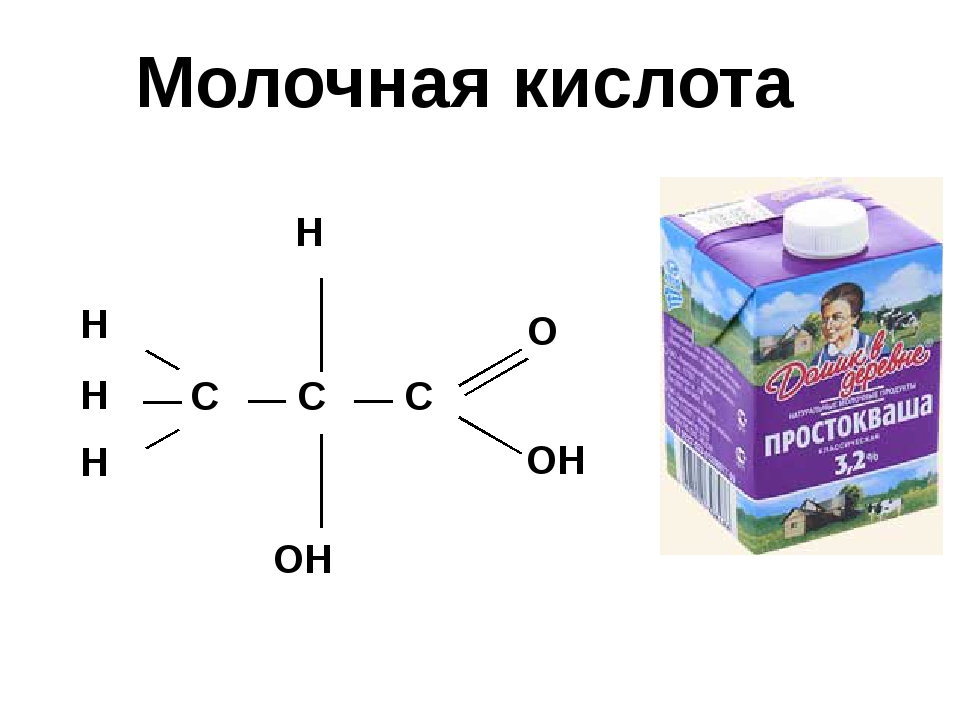 Благодаря наличию гидроксильной и карбоксильной групп в молекуле молочной кислоты, она может вступать в реакции, характерные как для спиртов, так и для кислот, что во многом объясняет ее высокую реакционную способность. Поэтому молочная кислота представляет большой интерес с точки зрения использования ее в качестве стартового соединения в органическом синтезе. Основные пути превращения молочной кислоты представлены на Схеме №1
Благодаря наличию гидроксильной и карбоксильной групп в молекуле молочной кислоты, она может вступать в реакции, характерные как для спиртов, так и для кислот, что во многом объясняет ее высокую реакционную способность. Поэтому молочная кислота представляет большой интерес с точки зрения использования ее в качестве стартового соединения в органическом синтезе. Основные пути превращения молочной кислоты представлены на Схеме №1
Схема №1 Пути превращения молочной кислоты.
С точки зрения промышленного применения продуктов, получаемых из молочной кислоты, на сегодняшний день наиболее востребована акриловая кислота, которая применяется в производстве полиакрилонитрильных волокон, а также эфиры молочной кислоты, использующиеся в качестве растворителей.
Однако наибольший интерес молочная кислота представляет как мономер для получения полилактида (PLA) (полимолочной кислоты) — биоразлагаемого пластика. Молочная кислота склонна к образованию эфиров дилактидов, дальнейшая полимеризация которых приводит к образованию полилактидов ( Схема №1). В результате образуется термопластичный полимер.
В результате образуется термопластичный полимер.
Полилактид используется в основном для получения биоразлагаемых упаковочных материалов, а также, благодаря высокой биосовместимости, широко применяется в медицинской практике: PLA является материалом, из которого изготавливаются одноразовые медицинские инструменты, биоразлагаемые хирургические нити, оболочки для лекарственных препаратов.
В связи с возрастающим спросом на биоразлагаемый пластик возрастает и спрос на производство молочной кислоты. Также молочная кислота и ее соли (лактаты) используют в пищевой промышленности, медицине, ветеринарии, а также для разнообразных технических целей.
Для использования в пищевых продуктах разрешены лактаты натрия (Е325), калия (Е326), кальция (Е327), аммония (Е328) и магния (Е329). Они применяются в производстве безалкогольных напитков, карамельных масс, кисломолочных продуктов. Как известно, молочная кислота сдерживает развитие гнилостных бактерий, поэтому она используется в качестве антисептика, в том числе и при хранении пищевых продуктов.
В промышленности молочную кислоту получают химическим (50%) и ферментативным (50%) способами в виде рацемата D и L — изомеров.
Химический способ получения молочной кислоты основан на стандартном циангидринном способе синтеза á-гидроксикислот, по реакции ацетальдегида с цианистым водородом, приводящей к образованию лактонитрила, кислотный гидролиз которого дает молочную кислоту ( Схему №2 ):
Схема №2 Циангидринный метод получения молочной кислоты
Основным недостатками данного способа получения молочной кислоты являются использование крайне токсичной синильной кислоты, двухстадийность процесса, а также необходимость тщательной отчистки готового продукта.
При получении молочной кислоты ферментативным путем в качестве исходного сырья используют сахара. В частности, в отечественном производстве сырьем служит смесь тростникового сахара-сырца, рафинадной патоки и свекловичной мелассы. Ферментация происходит анаэробным путем. Возбудителем молочнокислого брожения является в основном бактерия Lactobacillus Delbruckii, приводящая к образованию оптически недеятельной молочной кислоты. Следует отметить, что, несмотря на достаточно высокий выход молочной кислоты — 90%, ферментативный способ имеет ряд существенных недостатков, в частности, низкую производительность процесса и большое количество стадий отчистки, что, в свою очередь, увеличивает себестоимость продукта.
Возбудителем молочнокислого брожения является в основном бактерия Lactobacillus Delbruckii, приводящая к образованию оптически недеятельной молочной кислоты. Следует отметить, что, несмотря на достаточно высокий выход молочной кислоты — 90%, ферментативный способ имеет ряд существенных недостатков, в частности, низкую производительность процесса и большое количество стадий отчистки, что, в свою очередь, увеличивает себестоимость продукта.
Молочная кислота без молока! Самый интригующий ингредиент в составе веганских сыров. | VolkoMolko
Кислота молочная – именно про этот ингредиент в составе наших сыров мы слышим больше всего вопросов. Продукт веганский, а кислота — молочная! Что-то не сходится. Но не судите о продукте по названию, давайте лучше разберемся, чтобы знать наверняка, имеет ли этот ингредиент отношение к молочной промышленности.
Название «молочная» — это всего лишь дань способу, с помощью которого впервые выделили химическое вещество.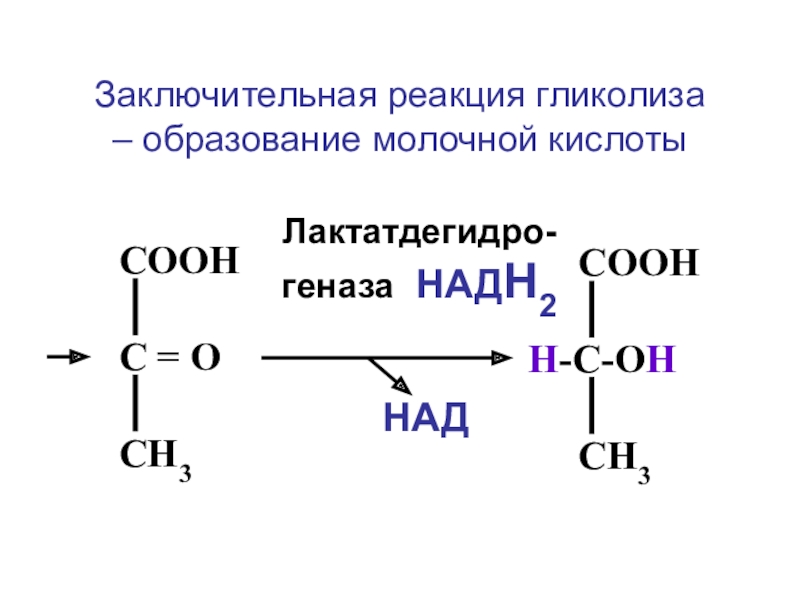 В современной промышленности молочную кислоту чаще всего получают без участия ингредиентов животного происхождения.
В современной промышленности молочную кислоту чаще всего получают без участия ингредиентов животного происхождения.
Кислота молочная или 2-гидроксипропионовая кислота — органическое соединение, представляющее собой прозрачную жидкость, с кислым вкусом, почти без запаха, по консистенции напоминающую сироп. На домашней кухне молочная кислота образуется при скисании молока, квашении капусты, солении овощей, созревании сыра.
А в промышленности ее получают получают двумя способами:
- ферментацией сахаросодержащих субстратов молочнокислыми микроорганизмами,
- или химическим синтезом (получение L-формы).
L-форма, как более экономичная и универсальная, используется все чаще. Именно ее мы закупаем для наших сыров.
В России молочная кислота одобрена для использования в пищевой промышленности в качестве регулятора кислотности и вкусовой добавки в следующих продуктах: нектары, джемы, желе, мармелады, масла, сыры, хлеб, макаронные изделия, пиво, квашеные, соленые овощи.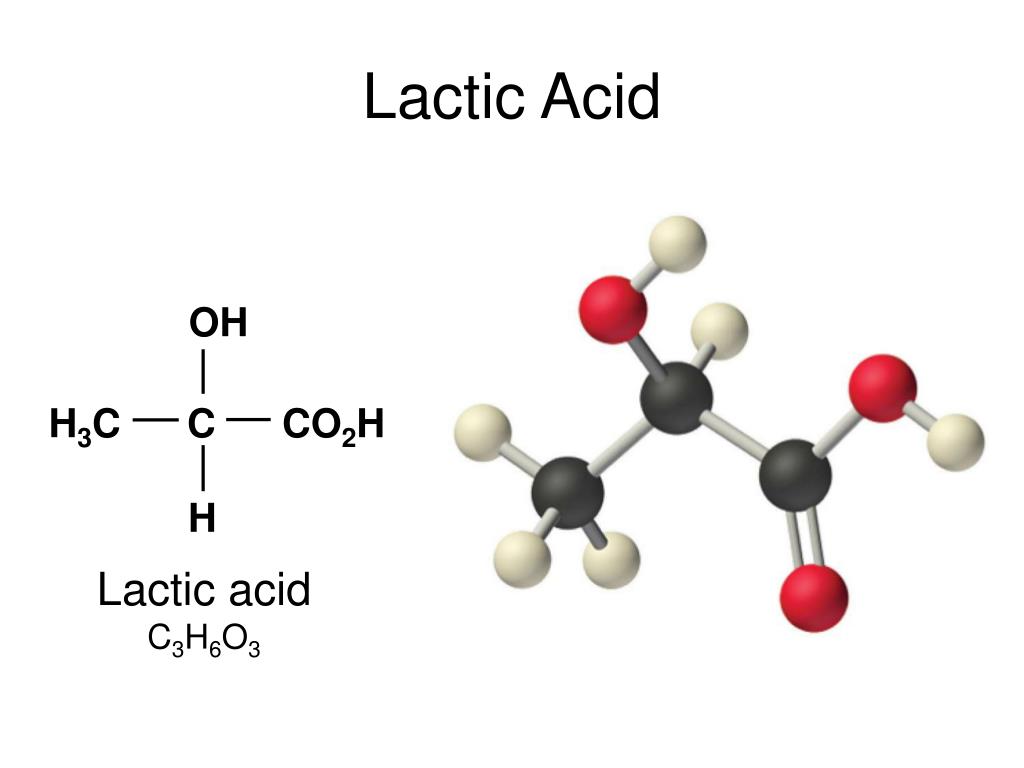 Кроме того, молочную кислоту разрешено использовать как консервант, но при этом ее расход очень высок, и использование чаще всего нерационально.
Кроме того, молочную кислоту разрешено использовать как консервант, но при этом ее расход очень высок, и использование чаще всего нерационально.
Важные и интересные факты о молочной кислоте:
- D-форма молочной кислоты образуется в мышцах человека при интенсивной физической нагрузке при распаде глюкозы (по одной из версий, боль в мышцах после тренировки обусловлена именно наличием молочной кислоты).
- Молочная кислота усваивается в организме с выделением энергии. Ее калорийность – 3 ккал/г.
- Она абсолютно безопасна для организма даже в больших количествах.
- В веганских продуктах используют только L-форму вещества, которая получена из сахаросодержащих растительных веществ с помощью химического синтеза.
- С помощью поликонденсации молочной кислоты можно получить биоразлагаемый пластик PLA. Его используют для пищевой упаковки и в медицине, для хирургических нитей.

Остались вопросы? Пишите в комментариях под статьей. Делитесь информацией с друзьями и не переставайте внимательно вчитываться в составы продуктов!
Молочная кислота — это… Что такое Молочная кислота?
Молочная кислота (лактат) — α-оксипропионовая (2-гидроксипропановая) кислота.
- tпл 25—26 °C оптически активная + или — форма.
- tпл 18 °C рацемическая форма.
Молочная кислота образуется при молочнокислом брожении сахаров, в частности в прокисшем молоке, при брожении вина и пива.
Была открыта шведским химиком Карлом Шееле в 1780 году.
В 1807 году Йенс Якоб Берцелиус выделил из мышц цинковую соль молочной кислоты.
Молочная кислота в организме человека и животных
Молочная кислота формируется при распаде глюкозы. Иногда называемая «кровяным сахаром», глюкоза является главным источником углеводов в нашем организме. Это основное топливо для мозга и нервной системы, так же как и для мышц во время физической нагрузки.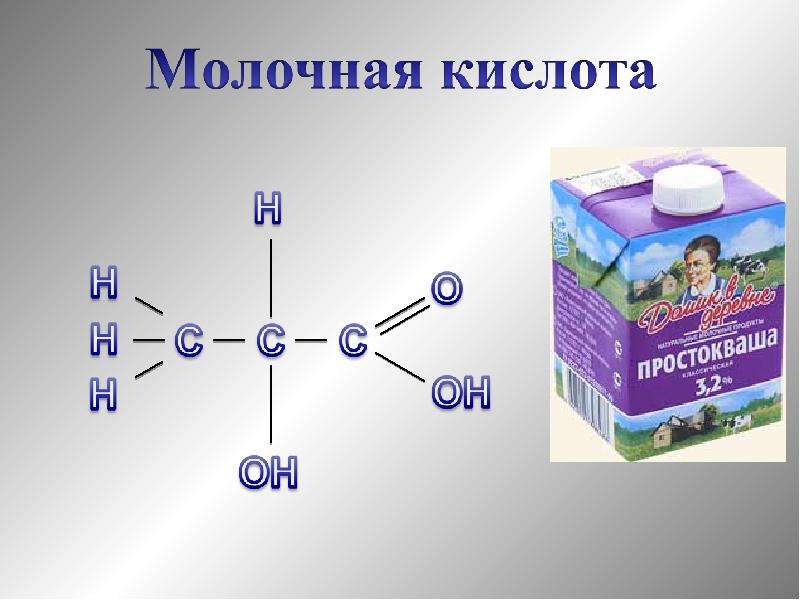 Когда расщепляется глюкоза, клетки производят АТФ (аденозина трифосфат), который обеспечивает энергией большинство химических реакций в организме. Уровень АТФ определяет, как быстро и как долго наши мышцы смогут сокращаться при физической нагрузке.
Когда расщепляется глюкоза, клетки производят АТФ (аденозина трифосфат), который обеспечивает энергией большинство химических реакций в организме. Уровень АТФ определяет, как быстро и как долго наши мышцы смогут сокращаться при физической нагрузке.
Производство молочной кислоты не требует присутствия кислорода, поэтому этот процесс часто называют «анаэробным метаболизмом» (см. Анаэробная тренировка). Многие считают, что мышцы производят молочную кислоту, когда недополучают кислород из крови. Другими словами, вы находитесь в анаэробном состоянии. Однако, учёные[1] утверждают, что молочная кислота образуется и в мышцах, получающих достаточно кислорода. Увеличение количества молочной кислоты в кровотоке свидетельствует лишь о том, что уровень её поступления превышает уровень удаления. Кислород не играет здесь существенной роли.
Зависимое от лактата производство АТФ очень незначительно, но имеет большую скорость. Это обстоятельство делает идеальным его использование в качестве топлива, когда нагрузка превышает 50 % от максимальной. При отдыхе и умеренной нагрузке организм предпочитает расщеплять жиры для получения энергии. При нагрузках в 50 % от максимума (порог интенсивности для большинства тренировочных программ) организм перестраивается на преимущественное потребление углеводов. Чем больше углеводов вы используете в качестве топлива, тем больше производство молочной кислоты.
При отдыхе и умеренной нагрузке организм предпочитает расщеплять жиры для получения энергии. При нагрузках в 50 % от максимума (порог интенсивности для большинства тренировочных программ) организм перестраивается на преимущественное потребление углеводов. Чем больше углеводов вы используете в качестве топлива, тем больше производство молочной кислоты.
Исследования показали, что у престарелых людей в головном мозге количество солей кислоты (лактатов) имеет повышенный уровень[2].
Регулятор обмена
Чтобы глюкоза могла проходить через клеточные мембраны, ей необходим инсулин. Молекула же молочной кислоты в два раза меньше молекулы глюкозы, и гормональная поддержка ей не нужна — она с лёгкостью сама проходит через клеточные мембраны.
Молочную кислоту можно обнаружить по следующим качественным реакциям:
- Взаимодействие с n-оксидифенилом и серной кислотой:
При осторожном нагревании молочной кислоты с концентрированной серной кислотой она вначале образует уксусный альдегид и муравьиную кислоту; последняя немедленно разлагается:
CH3CH(OH)COOH → CH3CHO + HCOOH (→ H2O + CO)
Уксусный альдегид взаимодействует с n-оксидифенилом, причём, по-видимому, происходит конденсация в o-положении к OH-группе с образованием 1,1-ди(оксидифенил)этана:
В растворе серной кислоты медленно окисляется в фиолетовый продукт неизвестного состава. Поэтому, как и при обнаружении гликолевой кислоты с помощью 2,7-диоксинафталина, в данном случае происходит взаимодействие альдегида с фенолом, при котором концентрированная серная кислота действует как конденсирующий агент и окислитель. Такую же цветную реакцию дают α-оксимасляная и пировиноградная кислоты.
Поэтому, как и при обнаружении гликолевой кислоты с помощью 2,7-диоксинафталина, в данном случае происходит взаимодействие альдегида с фенолом, при котором концентрированная серная кислота действует как конденсирующий агент и окислитель. Такую же цветную реакцию дают α-оксимасляная и пировиноградная кислоты.
Выполнение реакции: В сухой пробирке нагревают в течение 2 минут на водяной бане при 85 °C каплю исследуемого раствора с 1 мл концентрированной серной кислоты. После этого охлаждают под краном до 28 °C, добавляют небольшое количество твёрдого n-оксидифенила и, перемешав несколько раз, дают постоять 10-30 минут. Фиолетовое окрашивание появляется постепенно и через некоторое время становится более глубоким. Открываемый минимум: 1,5·10−6 г молочной кислоты.
- Взаимодействие с подкисленным серной кислотой раствором перманганата калия
Выполнение реакции: В пробирку прилить 1 мл молочной кислоты, а затем немного подкисленного серной кислотой раствора перманганата калия. Нагревать в течение 2 минут на слабом огне. Ощущается запах уксусной кислоты. С3Н6О3 + [O] = C3Н4O3 + H2O↑
Нагревать в течение 2 минут на слабом огне. Ощущается запах уксусной кислоты. С3Н6О3 + [O] = C3Н4O3 + H2O↑
Продуктом данной реакции может быть пировиноградная кислота С3Н4О3, которая тоже имеет запах уксусной кислоты.
С3Н6О3 + [O] = C3Н4O3 + H2O↑
Однако пировиноградная кислота при обычных условиях неустойчива и быстро окисляется до уксусной кислоты, поэтому реакция протекает согласно суммарному уравнению:
С3Н6О3 + 2[O] = CH3COOH↑ + CO2↑ + H2O↑
Применение и получение
В пищевой промышленности используется как консервант, пищевая добавка E270.
Получают молочную кислоту молочнокислым брожением глюкозы (ферментативная реакция):
C6H12O6 → 2CH3CH(OH)COOH + 21,8·104 Дж
См.
 также
также
Примечания
Ссылки
Молочная кислота
| Молочная кислота | |||
|---|---|---|---|
|
|
|||
| Систематическое наименование |
2-гидроксипропановая кислота | ||
| Хим. формула | CH3CH(OH)COOH | ||
| Рац. формула | C3H6O3 | ||
| Молярная масса | 90,08 г/моль | ||
| Плотность | 1,225 г/см³ | ||
| Температура | |||
| • плавления | 18 °C | ||
| • кипения | 122 °C | ||
| Константа диссоциации кислоты pKa | 3,86 (при 25 °C) | ||
| ГОСТ | ГОСТ 490-2006 ГОСТ 490-79 | ||
| Рег. номер CAS | 50-21-5 | ||
| PubChem | 612 | ||
| Рег. номер EINECS | 200-018-0 | ||
| SMILES | |||
| InChI |
1S/C3H6O3/c1-2(4)3(5)6/h3,4H,1h4,(H,5,6) JVTAAEKCZFNVCJ-UHFFFAOYSA-N |
||
| Кодекс Алиментариус | E270 | ||
| ChEBI | 78320 | ||
| ChemSpider | 592 | ||
| Сигнальное слово | Опасно | ||
| Пиктограммы СГС | |||
Приведены данные для стандартных условий (25 °C, 100 кПа), если не указано иное. |
|||
Молочная кислота (α-оксипропионовая, 2-гидроксипропановая кислота) CH3CH(OH)COOH — одноосновная карбоновая кислота с тремя атомами углерода, содержащая гидроксильную группу. Соли и эфиры молочной кислоты называются лактатами. Молочная кислота образуется при молочнокислом брожении сахаров и играет важную роль в метаболизме.
История
Молочную кислоту открыл шведский химик Карл Шееле. В 1780 году он выделил её из прокисшего молока в виде коричневого сиропа. Французский химик Анри Браконно обнаружил, что она образуется при молочнокислом брожении.
В 1807 году Йенс Якоб Берцелиус выделил из мышц цинковую соль молочной кислоты.
Физические свойства
Молочная кислота является простейшей хиральной карбоновой кислотой и может существовать в виде двух энантиомеров: L-(+)-молочной кислоты, D-(-)-молочной кислоты или их рацемической смеси — DL-молочной кислоты. Если в смеси энантиомеров один находится в избытке, его можно выделить дробными перекристаллизациями из смеси диэтилового эфира и диизопропилового эфира. Чистые энантиомеры имеют температуру плавления 52,7-52,8 °С.
Чистые энантиомеры имеют температуру плавления 52,7-52,8 °С.
Энантиомеры молочной кислоты: L-(-)-молочная кислота (слева) и D-(+)-молочная кислота (справа)
Молочная кислота очень гигроскопична и обычно существует в виде водного раствора с концентрацией до 90 мас. %. В связи с этим очень сложно установить её температуру плавления; литература приводит значения от 18 до 33 °С. Кроме того, в таких растворах присутствует значительное количество лактоилмолочной кислоты и других олигомеров молочной кислоты.
Молочная кислота растворима в воде, этаноле, диэтиловом эфире и других органических растворителях, смешивающихся с водой. Она практически нерастворима в бензоле и хлороформе.
Химические свойства
При взаимодействии с окислителями молочная кислота разлагается. При окислении кислородом воздуха или азотной кислотой в присутствии железа или меди она превращается в муравьиную кислоту, уксусную кислоту, щавелевую кислоту, уксусный альдегид, углекислый газ и пировиноградную кислоту. Молочную кислоту можно восстановить до пропионовой кислоты действием иодоводорода
Молочную кислоту можно восстановить до пропионовой кислоты действием иодоводорода
Промышленный интерес представляет дегидратация молочной кислоты до акриловой кислоты и её восстановление до пропиленгликоля.
Поскольку молочная кислота является одновременно карбоновой кислотой и спиртом, она вступает в межмолекулярную этерификацию, давая лактоилмолочную кислоту. При дальнейшей конденсации образуется лактид — циклический сложный эфир. Также при конденсации могут образовываться линейные полилактиды. Эти соединения являются примесями в молочной кислоте. Так, в 6,5%-й молочной кислоте содержится около 0,2 % лактоилмолочной кислоты, 88%-я молочная кислота содержит меньше 60 % свободной молочной кислоты, а 100%-я — только 32 %.
Получение
Ферментативное получение
В промышленности молочную кислоту получают либо ферментативным способом, либо синтетическим. Первый из них имеет преимущество, поскольку приводит к молочной кислоте с более высокой стереохимической чистотой, поэтому новые производства, открывавшиеся с 1995 года, пользуются этим способом.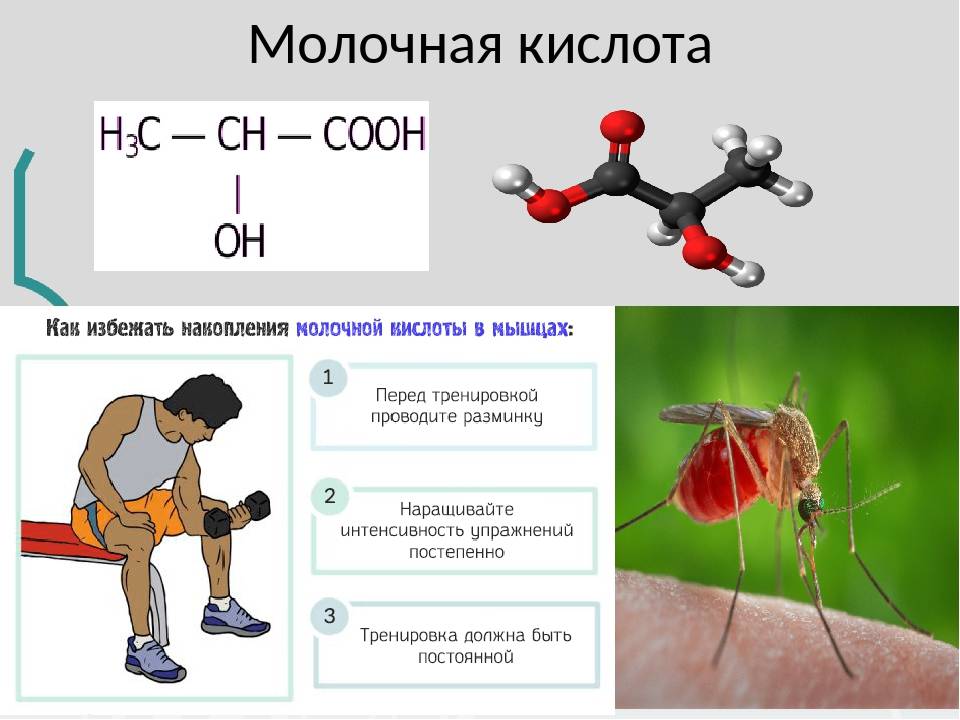
Для ферментативного производства молочной кислоты необходимо иметь углеводное сырьё, питательные вещества и соответствующие микроорганизмы. В качестве сырья используют глюкозу, кукурузные сиропы, мелассу, сок сахарной свёклы, сыворотку и крахмал. Питательными веществами являются пептиды и аминокислоты, фосфаты, соли аммония и витамины. Здесь находят применение дрожжевой экстракт, кукурузный ликёр, кукурузная глютеновая мука, солодовые ростки, соевый или мясной пептон. Молочную кислоту из углеводов вырабатывают культуры Lactobacillus, Bacillus и Rhizopus. Последняя из них является грибной и не требует сложных источников азота, хотя и даёт более низкий выход, чем бактериальные культуры.
Во время процесса вырабатывается кислота, поэтому необходимо поддерживать pH в области 5,0–6,5: для этого используют гидроксид кальция, карбонат кальция, аммиак и гидроксид натрия. Из-за этого в процессе ферментации образуются соответствующие соли молочной кислоты. Разработка новых подходов к ферментации связана с выведением бактерий, способных работать в области низких pH: это позволило бы получать саму молочную кислоту, а не её соли, снизило бы затраты на подщелачивающие реагенты и на серную кислоту, используемую для выделения молочной кислоты из солей.
После ферментации молочную кислоту подвергают очистке. Микроорганизмы отделяют флокуляцией в щелочной среде либо ультрафильтрацией. Образовавшиеся соли молочной кислоты переводят в саму кислоту под действием серной кислоты. При этом образуется также малополезный сульфат кальция. Ведётся поиск других методов очистки, при которых образование это побочной соли не происходило бы. Для некоторых пищевых целей смесь пропускают через активированный уголь и ионообменную колонну.
Для фармацевтической промышленности и получения полимеров необходима более глубокая очистка. Интерес представляет экстракция молочной кислоты в органическую фазу длинноцепными третичными аминами, а затем обратная экстракция в воду. Этот метод позволяет эффективно очистить продукт от остаточных углеводов и белков. Также возможна перегонка молочной кислоты, если предпринимаются меры против её олигомеризации.
Выход молочной кислоты в процессе ферментации составляет 85–95 %. Побочными продуктами являются муравьиная и уксусная кислота.
Синтетическое получение
С 1960-х гг. рацемическую молочную кислоту производят в промышленности также синтетически. Подход основан на реакции ацетальдегида с циановодородом и последующем гидролизе образовавшегося лактонитрила. Недостатком синтетического подхода является то, что получаемая молочная кислота является рацемической, а подходящий и дешёвый хиральный катализатор пока не найден. Последним крупным производителем синтетической молочной кислоты является фирма «Musashino».
Экономические аспекты
Годовая потребность в молочной кислоте составляет примерно 450 тыс. тонн. Крупнейшим производителем молочной кислоты является компания NatureWorks LLC: её завод, размещённый в штате Небраска (США), имеет приблизительную мощность 180 тыс. тонн в год. Это в 3-4 раза больше, чем у следующего завода по порядку. Рынок молочной кислоты очень чувствителен к потребности в полилактиде, поскольку его производство является либо вскоре станет крупнейшим направлением использования молочной кислоты.
Молочная кислота в организме человека и животных
Молочная кислота формируется при распаде глюкозы. Иногда называемая «кровяным сахаром» глюкоза является главным источником углеводов в нашем организме. Это основной источник энергии для мозга и нервной системы, так же как и для мышц во время физической нагрузки. Когда расщепляется глюкоза, клетки производят АТФ (аденозинтрифосфат), который обеспечивает энергией большинство химических реакций в организме. Уровень АТФ определяет, как быстро и как долго мышцы смогут сокращаться при физической нагрузке.
Производство молочной кислоты не требует присутствия кислорода, поэтому этот процесс её синтеза часто называют «анаэробным метаболизмом» (см. Анаэробная тренировка). Ранее считалось, что мышцы производят молочную кислоту при нехватке кислорода в крови. Другими словами, организм находится в анаэробном состоянии. Однако современные исследования показывают, что молочная кислота образуется и в мышцах, получающих достаточно кислорода. Увеличение количества молочной кислоты в кровотоке свидетельствует лишь о том, что уровень её поступления превышает уровень удаления. Резкое увеличение (в 2—3 раза) уровня лактата в сыворотке крови наблюдается при тяжёлых расстройствах кровообращения, таких как геморрагический шок, острая левожелудочковая недостаточность и др., когда одновременно страдает и поступление кислорода в ткани, и печёночный кровоток.
Увеличение количества молочной кислоты в кровотоке свидетельствует лишь о том, что уровень её поступления превышает уровень удаления. Резкое увеличение (в 2—3 раза) уровня лактата в сыворотке крови наблюдается при тяжёлых расстройствах кровообращения, таких как геморрагический шок, острая левожелудочковая недостаточность и др., когда одновременно страдает и поступление кислорода в ткани, и печёночный кровоток.
Зависимое от лактата производство АТФ очень незначительно, но имеет большую скорость. Это обстоятельство делает идеальным его использование в качестве источника энергии, когда нагрузка превышает 50 % от максимальной. При отдыхе и умеренной нагрузке организм предпочитает расщеплять жиры для получения энергии. При нагрузках в 50 % от максимума (порог интенсивности для большинства тренировочных программ) организм перестраивается на преимущественное потребление углеводов. Чем больше углеводов человек использует в качестве топлива, тем больше производство молочной кислоты.
Исследования показали, что у престарелых людей в головном мозге количество солей кислоты (лактатов) имеет повышенный уровень.
Регулятор обмена
Чтобы глюкоза могла проходить через клеточные мембраны, ей необходим инсулин. Молекула же молочной кислоты в два раза меньше молекулы глюкозы, и гормональная поддержка ей не нужна — она с лёгкостью сама проходит через клеточные мембраны.
Анализ
Количественный анализ молочной кислоты обычно проводят титрованием гидроксидом натрия в присутствии фенолфталеина. Этот способ не подходит для концентрированных растворов, потому что в них молочная кислота частично находится в виде лактоилмолочной кислоты. Если такой раствор нужно оттитровать, используют приём обратного титрования: молочную кислоту обрабатывают избытком щёлочи (лактоилмолочная кислота при этом гидролизуется), а затем остаток щёлочи оттитровывают соляной кислотой.
В промышленности и исследовательских лабораториях молочную кислоту анализируют методом ВЭЖХ. Содержание энантиомерных форм в молочной кислоте можно определить ферментативным методом либо ВЭЖХ на хиральной колонке. Такие колонки, как и образцы высокочистых энантиомеров молочной кислоты коммерчески доступны.
Молочную кислоту можно обнаружить по следующим качественным реакциям:
- Взаимодействие с n-оксидифенилом и серной кислотой:
При осторожном нагревании молочной кислоты с концентрированной серной кислотой она вначале образует уксусный альдегид и муравьиную кислоту; последняя немедленно разлагается:
CH3CH(OH)COOH → CH3CHO + HCOOH (→ H2O + CO)
Уксусный альдегид взаимодействует с n-оксидифенилом, причём, по-видимому, происходит конденсация в o-положении к OH-группе с образованием 1,1-ди(оксидифенил)этана:
В растворе серной кислоты медленно окисляется в фиолетовый продукт неизвестного состава. Поэтому, как и при обнаружении гликолевой кислоты с помощью 2,7-диоксинафталина, в данном случае происходит взаимодействие альдегида с фенолом, при котором концентрированная серная кислота действует как конденсирующий агент и окислитель. Такую же цветную реакцию дают α-оксимасляная и пировиноградная кислоты.
Выполнение реакции: В сухой пробирке нагревают в течение 2 минут на водяной бане при 85 °C каплю исследуемого раствора с 1 мл концентрированной серной кислоты. После этого охлаждают под краном до 28 °C, добавляют небольшое количество твёрдого n-оксидифенила и, перемешав несколько раз, дают постоять 10-30 минут. Фиолетовое окрашивание появляется постепенно и через некоторое время становится более глубоким. Открываемый минимум: 1,5⋅10−6 г молочной кислоты.
- Взаимодействие с подкисленным серной кислотой раствором перманганата калия
Выполнение реакции: В пробирку прилить 1 мл молочной кислоты, а затем немного подкисленного серной кислотой раствора перманганата калия. Нагревать в течение 2 минут на слабом огне. Ощущается запах уксусной кислоты. C3H6O3 + [O] = C3H4O3 + H2O↑ Продуктом данной реакции может быть пировиноградная кислота C3H4O3, которая тоже имеет запах уксусной кислоты. C 3H6O3 + [O] = C3H4O3 + H2O↑ Однако пировиноградная кислота при обычных условиях неустойчива и быстро окисляется до уксусной кислоты, поэтому реакция протекает согласно суммарному уравнению: C 3H6O3 + 2[O] = CH3COOH + CO2↑ + H2O
- Взаимодействие с фенолятом железа
Описание реакции: Эта реакция называется реакцией Уффельмана и используется, например, в клинической медицине для определения присутствия молочной кислоты в желудочном соке, открыта Юлиусом Уффельманом в 1880-х гг. Для проведения реакции нужно растворить одну каплю хлорида железа и 0,4 грамма фенола в 50 см3 воды. Затем добавить тестируемую жидкость, если в ней есть молочная кислота, то синий цвет раствора сменится жёлтым, поскольку образуется лактат железа.
Применение
Молочная кислота используется преимущественно в трёх сферах: пищевой промышленности, производстве полимеров и других промышленных целях.
В пищевой промышленности молочная кислота используется как консервант и подкислитель. Благодаря тому, что её соли хорошо растворимы в воде, их также можно использовать в тех продуктах, где важно значение pH. Молочная кислота и её соли используются в напитках, конфетах, мясных изделиях и соусах. Лактат кальция добавляется в продукты как источник кальция.
В полимерной промышленности из молочной кислоты получают полилактид. Производится он из лактида полимеризацией с раскрытием цикла. Сам же лактид получают конденсацией молочной кислоты.
Также молочная кислота используется в покрытии металлов, косметике, текстильной и кожевенной промышленности. Её эфиры находят применение в производстве красок и чернил, электроники и чистке металлов.
Безопасность
Молочная кислота — это органическая карбоновая кислота, которая присутствует во многих организмах и является безопасной для окружающей среды. Увеличение производства молочной кислоты и, соответственно, полилактида оказывает положительное влияние тем, что вытесняет использование полимеров, производимых из нефти, снижает выбросы углекислого газа и предоставляет более широкие возможности по утилизации отходов.
Молочная кислота также используется в пищевой промышленности и не является токсичной. Тем не менее при попадании в глаза или на повреждённую кожу она вызывает раздражение. Полулетальная доза для крыс при оральном приёме составляет 3,73 г/кг.
Молочная кислота в косметике: что это, как работает, зачем нам надо и как использовать?
Молочная, или лактановая, кислота (Lactic acid) – это альфа-гидрокислота (АНА), родственная коже и входящая в состав ее водно-липидного слоя. Она обладает выраженным увлажняющим эффектом благодаря своей способности связывать молекулы воды. Также молочная кислота естественным образом повышает эластичность и упругость кожи. Когда количество молочной кислоты в клетках уменьшается, лицо выглядит заметно более сухим и уставшим.
Что нужно знать о молочной кислоте?
pexels.com
Молочная кислота уже присутствует в нашем организме – она формируется при распаде глюкозы, и ее часто называют «кровяным сахаром». Косметологи относят этот компонент к фруктовым кислотам и с удовольствием используют в косметологии, ведь молочная кислота идеально подходит для ухода за кожей любого типа. В процедурах средства с молочной кислотой чаще включают в различные пилинги, уходы за волосами, а также в комплексы для рук и ногтей.
Молочная кислота ослабляет и разрушает связи между омертвевшими клетками, благодаря чему во время процедуры кожа обновляется естественным путем. Также она обладает выраженным отбеливающим действием, которое проявляется за счет удаления верхнего слоя кожи и уникальной способности кислоты равномерно распределять меланин и замедлять синтез фермента, повышающего выработку пигмента. Эффект гладкого и ровного лица – тоже про молочную кислоту. Во время процедуры она проникает глубоко в поры, нормализует работу сальных желез и предотвращает дальнейшее скопление загрязнений. Как результат – такой уход предупреждает появление черных точек и акне.
Молочная кислота рекомендована и для ухода за сухой увядающей кожей, и для нормализации состояния жирного и проблемного типа. Спектр ее действия достаточно широк, ведь ее используют и для активного увлажнения кожи, профилактики старения и омоложения, для очищения кожи от омертвевших клеток, для нормализации работы сальных желез и сужения пор, для терапевтического ухода при угревой сыпи, для устранения гиперпигментации и ускорения процессов регенерации, а также для повышения эффективности другой уходовой косметики.
1 — тоник с молочной кислотой Medik8, 2 — ультра выравнивающий лосьон для тела Ultra Retexturising Body Complex, Ultraceuticals, 3 — пилинг с молочной кислотой PIXI, 4 — обновляющая сыворотка для тела с молочной кислотой REN Skincare
При регулярном использовании молочной кислоты кожа становится более гладкой, увлажненной, ее рельеф выравнивается, морщины становятся менее выраженными, а воспаления, расширенные поры и черные точки исчезают.
Где искать молочную кислоту?
Сыворотки с 5–10% молочной кислоты отлично подойдут для домашнего ухода. Маленький процент молочной кислоты обеспечивает увлажнение и выравнивание рельефа кожи, слегка отшелушивает и смягчает.
Также сыворотки на основе молочной кислоты эффективно увлажняют кожу, выравнивают ее рельеф, осветляют и делают морщины, рубцы, следы от воспалений менее заметными. С таким уходом кожа почти мгновенно из уставшей превращается в отдохнувшую.
Включить средство с молочной кислотой лучше в вечерний уход – наносить нужно несколько капель сыворотки сразу после умывания на сухое лицо. Если кожа только привыкает к кислотам, первые несколько раз лучше использовать сыворотку как маску и держать 30–40 минут, после чего смыть водой и нанести привычный уход. Когда кожа привыкнет к воздействию молочной кислоты, схему использования можно изменить: держать сыворотку на коже 10–15 минут, не смывать и по истечении этого времени сразу нанести обычный ночной крем. Использовать средства с молочной кислотой каждый день нельзя – двух–трех раз в неделю будет достаточно. При распределении сыворотки по лицу важно избегать контакта средства с глазами. Не стоит забывать о необходимости утром использовать солнцезащитные средства – это правило работает со всеми кислотами.
1 — пилинг с молочной кислотой Lactica Exfoliate 10%, ARAVIA Professional, 2 — отшелушивающая сыворотка с молочной и гиалуроновой кислотами Mixit, 3 — увлажняющая отшелушивающая сыворотка с молочной кислотой от блогера Адэль Organic Kitchen, 4 — ежедневная увлажняющая сыворотка-пилинг для лица с 5% молочной и 3D гиалуроновой кислотой, FACT
Встретить молочную кислоту можно и в больших концентрациях, например, 40% или 90% – такие средства используются для пилингов. Наносить их в чистом виде на кожу нельзя. Важно знать, с чем развести концентрированную молочную кислоту, как распределить, сколько держать и как ухаживать за кожей после процедуры. Все эти вопросы лучше оставить профессиональному косметологу и не экспериментировать с концентрированными средствами в домашних условиях. Самостоятельно можно попробовать делать готовые пилинги с содержанием молочной кислоты около 15–20%.
Несмотря на то что молочная кислота рекомендована для всех типов кожи, включая чувствительную и сухую, у нее есть свои противопоказания: использовать кислоту стоит с осторожностью при склонности к реактивности кожи и аллергиях на косметические средства, а также при куперозе и воспалительных кожных заболеваниях, например, розацеа. Не следует использовать молочную кислоту вокруг глаз, и важно избегать ее попадания на слизистые оболочки.
применение в косметике и косметологии, свойства, польза для кожи
Польза молочной кислоты для кожи лица
Молочная кислота в косметике — спасение для тех, кто привык к курсам пилингов и не хочет делать перерыв даже летом. Но прежде предлагаем разобраться, что такое молочная кислота и в чем ее польза для лица.
Молочная кислота образуется при молочнокислом брожении сахаров и в нашем организме присутствует естественным образом. Ее выделяют мышцы при активной работе. Вспомните, как болит все тело наутро после тяжелой тренировки. Все это результат выработки молочной кислоты. Содержится она и в коже — входит в состав натурального увлажняющего фактора. Если количество молочной кислоты по каким-то причинам уменьшается, кожа тут же становится сухой. Кстати, в чем еще кроются причины сухости кожи, можно узнать из нашего подкаста.
Но вернемся к молочной кислоте и перечислим ее основные свойства:
-
оказывает легкое отшелушивающее действие;
-
увлажняет;
-
выравнивает и осветляет кожу;
-
успокаивает;
-
уменьшает выраженность несовершенств.
Хотите узнать, за что вам будет благодарна кожа? Ответьте на вопросы нашего теста.
1 / 5
Как выглядит ваш вечерний уход за собой?
2 / 5
Вас можно назвать человеком, который следует модным трендам, например, Slow Age или Clean Beauty?
3 / 5
Во сколько вы обычно ложитесь спать?
4 / 5
Вы курите?
5 / 5
Оцените, насколько правильно вы питаетесь, по 10-балльной шкале.
По сравнению с другими AHA-кислотами молочная считается одной из самых деликатных, но от этого она не менее эффективна. Благодаря молочной кислоте в коже запускаются следующие процессы:
«Кислотные пилинги нежелательны для обладателей тонкой, сухой, обезвоженной кожи, но что касается молочной кислоты, то она действует хоть и мягко, но крайне эффективно. Кроме того, пилинг с молочной кислотой можно делать даже летом. При условии, конечно, что после него кожа будет надежна защищена кремом с солнцезащитным фактором не менее 50».
-
усиливается синтез коллагена и эластина;
-
клетки получают импульс к обновлению;
-
благодаря пилингу на основе молочной кислоты межклеточное пространство наполняется влагой;
-
укрепляется защитный барьер кожи.
Вернуться к оглавлению
Косметика с кислотами: обзор лучших средств
В нашей подборке лучшие, по мнению редакции, средства с АНА-кислотами. В том числе и с молочной. Важно, что этот ингредиент подходит даже для чувствительной кожи.
Маска-пилинг Re-Plasty High Definition Peel, Helena Rubinstein
Мочевина и молочная кислота в составе этой плотной и прозрачной желейной маски способствуют восстановлению и очищению кожи всего за пять минут.
Минеральная маска-пилинг «Двойное сияние», Vichy
Фруктовые кислоты и частицы вулканического происхождения мягко отшелушивают и выравнивают текстуру кожи, улучшая цвет лица.
Сыворотка для кожи лица Blemish & Age Defense, SkinCeuticals
Формула, содержащая диоевую, гликолевую, салициловую и лимонную кислоты, направленно действует на выравнивание рельефа кожи и осветление пигментации.
Вернуться к оглавлению
Можно ли пользоваться молочной кислотой в домашних условиях
Домашние маски из кисломолочных продуктов хорошо отбеливают и увлажняют кожу. © Getty Images
Применяется молочная кислота для лица и в домашних условиях. При желании каждая из нас может стать сама себе косметологом. Но в случае пилингов этого делать не стоит, если только под рукой нет готового средства на основе молочной кислоты. Готовить же такой пилинг самостоятельно — дело рискованное.
Что касается масок домашнего приготовления, тут трудно возразить. Маски на основе молока и кисломолочных продуктов вполне безобидны, при условии, что вы используете свежие продукты. Вот парочка рецептов.
Маска с виноградным соком и молоком
Вам понадобятся по половине стакана виноградного (свежевыжатого) сока и молока.
- 1
Смешайте оба ингредиента, смочите марлевую салфетку и приложите компресс к лицу на 15 минут.
- 2
Затем умойтесь теплой водой и нанесите питательный или увлажняющий крем.
Маска питательная со сметаной
- 1
Возьмите по 1 столовой ложке дрожжей и жирной сметаны, а также по чайной ложке льняного масла и меда.
- 2
Разотрите дрожжи со сметаной, добавьте льняное масло и мед.
- 3
Поставьте миску со смесью в горячую воду и размешайте. Надо, чтобы дрожжи поднялись.
- 4
Теплую смесь нанесите на лицо на 10 минут.
- 5
Смойте прохладной или теплой водой.
Только не забывайте, что мед — один из сильнейших аллергенов, убедитесь, что у вас нет на него нежелательной реакции.
Вернуться к оглавлению
Пилинг с молочной кислотой: применение в косметологии
Если речь идет о профессиональном молочном пилинге, то при всей мягкости действия молочная кислота в косметологии все же несколько повышает чувствительность кожи к ультрафиолету. Поэтому после использования средств с концентрацией кислоты выше 10% рекомендуется наносить солнцезащитный крем с максимальным уровнем защиты (SPF 50+).
«Что касается салонного пилинга молочной кислотой, стоит понимать, что это курсовая процедура. Вряд ли после первого сеанса вы увидите впечатляющий результат. Продолжительность курса зависит от состава для нанесения и от состояния кожи».
Курсы пилингов рекомендуется повторять1-2 раза в год, исключая время жары и морозов.
Между процедурами стоит сосредоточиться на тщательном увлажнении и защите от ультрафиолета. Уход на основе термальной воды, гиалуроновой кислоты, витаминов В5 и В3 поможет уменьшить дискомфорт после процедуры.
Вернуться к оглавлению
Молочная кислота
Определение «Молочная кислота» в Большой Советской Энциклопедии
Молочная кислота, a-оксипропионовая кислота, CH3CH(OH)COOH, одноосновная оксикарбоновая кислота; важный промежуточный продукт обмена веществ у животных, растений и микроорганизмов. Молочная кислота — бесцветные кристаллы, легкорастворимые в воде; существует в двух оптически активных ( — )- и (+)-формах (tпл 25—26 °С) и в виде неактивной рацемической (±)-формы (tпл 18 °С). ( — )- и (+)-Молочная кислота рацемизуются при 130—150 °С; аналогично карбоновым кислотам образуют соли (лактаты) и эфиры. Молочная кислота образуется при брожении сахаристых веществ под влиянием различных бактерий молочнокислого брожения, например при скисании молока, квашении капусты, солении овощей, созревании сыра, силосовании кормов. При этом в зависимости от природы бактерий и сахара образуется либо рацемат, либо одна из оптически активных форм. В промышленности из тростникового сахара, декстрозы и мальтозы при действии Bacillus Delbrucki получают (±)-Молочная кислота (т. н. Молочная кислота брожения), которую используют в дублении для декальцинирования кож, в крашении — для восстановления хроматов при хромовом травлении. Соли Молочная кислота применяют в качестве протрав при крашении и печатании, эфиры — в качестве растворителей лаков.
В животных тканях (главным образом в мышцах) (+)-Молочная кислота — конечный продукт ферментативного анаэробного (без участия O2) расщепления углеводов — гликолиза. В отсутствие O2 количество образующейся в мышце Молочная кислота эквивалентно количеству исчезнувшего гликогена (мышечное утомление связано с истощением запасов гликогена и накоплением Молочная кислота). При наличии O2 около 20 % Молочная кислота окисляется до CO2 и H2O, а освобождающаяся при этом энергия используется для превращения остальной Молочная кислота в гликоген (см. Кори цикл); утомление при этом наступает медленнее. Обратимое восстановление пировиноградной кислоты (пирувата) в Молочная кислота (лактат) на последнем этапе гликолиза протекает под действием лактатдегидрогеназы. В растениях и микроорганизмах Молочная кислота образуется при восстановлении пирувата.
В. Н. Фросин.
Статья про «Молочная кислота» в Большой Советской Энциклопедии была прочитана 290 раз
Молочная кислота — обзор
3.17.1 Введение
Молочная кислота (2-гидроксипропионовая кислота) входит в число крупных химических веществ, производимых микробиологически, с годовым мировым объемом производства в пределах 370 000 тонн. Молочно-кислотная ферментация — одна из старейших промышленных ферментаций, промышленное производство которой путем ферментации началось в 1880-х годах. Семьдесят пять процентов текущего мирового производства молочной кислоты приходится на ферментационные предприятия Galactic, PURAC Corporation, Cargill Incorporated, Archer Daniels Midland Company и совместных предприятий, созданных этими компаниями [1].Исторически сложилось так, что молочная кислота в основном использовалась в пищевых продуктах для подкисления и консервирования, и ей был присвоен статус GRAS (общепризнанный как безопасный) FDA. Молочная кислота также находит применение при дублении кожи, косметике, фармацевтике, а также в различных других нишах [2–4]. Мировое производство молочной кислоты увеличилось в 10 раз за последнее десятилетие, в значительной степени из-за увеличения спроса на экологически чистые продукты, полученные из молочной кислоты, включая этиллактат и полимолочную кислоту (PLA) [3].Этилактат можно использовать в различных зеленых растворителях, и, хотя его низкая токсичность для человека по сравнению с углеводородными альтернативами является привлекательной, цена указывается как основная причина его ограниченного использования на рынке [1]. PLA — это полимер, который считается экологически чистой альтернативой пластмассам, полученным из нефти, благодаря его способности к биологическому разложению и уменьшенному углеродному следу [5]. Продукты PLA представлены на рынке в широком диапазоне применений, включая упаковку, волокна и пенопласт [1, 5, 6]. Основным производителем PLA в мире является компания NatureWorks LLC, которая в настоящее время полностью принадлежит Cargill Incorporated.Основные затраты при производстве PLA и этиллактата — это стоимость сырья, то есть молочной кислоты. Ключевыми параметрами, определяющими стоимость молочной кислоты, являются скорость, титр и выход, как при ферментации, так и при последующих операциях по извлечению продукта [3]. Кроме того, на производство молочной кислоты приходится значительная часть потребляемой энергии и выбросов парниковых газов (ПГ) в продуктах, полученных из молочной кислоты. Эти углеродные затраты могут иметь большое значение для маркетинга и жизнеспособности экологически чистых продуктов [6].
Как обсуждалось ранее, производство молочной кислоты происходит на протяжении более 100 лет, с незначительными изменениями условий или организмов-хозяев [2]. Молочная кислота производится путем ферментации, традиционно осуществляемой бактериями, принадлежащими к родам Lactobacillus, Lactococcus, Streptococcus, Bacillus и Enterococcus [2,3]. Для недавних применений молочной кислоты в качестве промежуточного экологически чистого химического вещества, например, для PLA, стоимость производства традиционным способом слишком высока.Смета расходов предполагает, что для обеспечения коммерческой жизнеспособности общие затраты на производство молочной кислоты должны быть на уровне или ниже 1,0 доллара за килограмм молочной кислоты. В результате производственный штамм для промышленной молочной кислоты должен соответствовать следующим критериям: производство> 100 г на л -1 молочной кислоты с выходами, близкими к теоретическим (0,9 г молочной кислоты на грамм декстрозы), высокая хиральная чистота молочной кислоты. произведено (> 99%) с тарифами, расходами на носители и восстановлением, способными удовлетворить вышеуказанные целевые затраты [3]. Снижение себестоимости производства может расширить рынок как молочной кислоты, так и ее зеленых производных.
Основные затраты, связанные с ферментацией, — это питательные вещества и сахара, необходимые для роста клеток и производства молочной кислоты, а также последующие процессы восстановления и очистки [7]. В дополнение к источнику сахара для традиционных бактериальных молочнокислых ферментаций обычно требуется источник органического азота (например, дрожжевой экстракт или кукурузный настой) вместе с добавками витамина B. Кроме того, эти ферментации требуют, чтобы pH поддерживался в диапазоне 5-7, что значительно выше p K a молочной кислоты [2].Поддержание pH в этом диапазоне требует нейтрализации молочной кислоты во время ферментации с последующими дорогостоящими последующими стадиями или подкислением для регенерации свободной молочной кислоты. Это значительно увеличивает стоимость брожения.
В 2008 году компания Cargill внедрила новую для мира технологию ферментации с использованием генетически модифицированных дрожжей, способных производить молочную кислоту с промышленно приемлемыми скоростями, титрами и урожайностью при значениях pH ≤ 3,0, что значительно ниже p K a молочной кислоты.Процесс ферментации с низким pH приводит к повышению качества продукта и последующей обработки, снижению использования химических веществ и затрат на питательные вещества, а также к снижению выбросов парниковых газов, связанных с производством молочной кислоты путем ферментации, на 35%. Кроме того, возможность потери продукта из-за атак бактериофагов и микробного загрязнения, которые могут возникнуть при традиционном бактериальном процессе, устраняется или значительно снижается при использовании дрожжевого процесса с низким pH. Такая повышенная надежность процесса способствует снижению общих затрат на производство молочной кислоты и впоследствии способствует росту рынка молочной кислоты и ее производных.
Ожидается, что будущие достижения в области дрожжевого процесса с низким pH еще больше снизят стоимость производства молочной кислоты за счет снижения стоимости источника углерода, ферментированного до молочной кислоты. Для достижения этого необходимо дальнейшее развитие дрожжей с низким pH для эффективного сбраживания недорогих источников углерода до высвобождения молочной кислоты. Анализ жизненного цикла показал, что за счет использования целлюлозного сырья, полученного из биомассы, и использования энергии ветра для производства молочной кислоты и PLA, общие выбросы ПГ могут быть рассчитаны как чистые отрицательные [6] ( Рисунок 1 ) .
Рисунок 1. Производство PLA из молочной кислоты.
Молочная кислота — обзор
8.3.1.1 Ферментация
Молочная ферментация была изучена как альтернативный метод извлечения хитина, производства гидролизатов белка и молочной кислоты, а также для экстракции пигментов из отходов ракообразных [51 –54]. Было четко установлено, что производство кислоты является ключевым фактором ферментации для подавления патогенов и микроорганизмов, вызывающих порчу, а также для удаления минералов из отходов ракообразных, а также pH влияет на активность протеаз, ответственных за гидролиз белка из этих раковин [55,56] .Таким образом, СВ креветок ( L. vannamei) гепатопанкреаса достигается солюбилизацией минералов органической кислотой из лактобацилл, тогда как СП приписывается пищеварительным и микробным протеазам, продуцируемым во время ферментации. В связи с этим Пачеко и соавт. [56] сообщили, что максимальное удаление белка было определено при оптимальном росте молочнокислых бактерий. Это продемонстрировало ключевую роль бактериальных протеаз для экстракции хитина во время молочнокислой ферментации отходов креветок с использованием гомофермента Lactobacillus plantarum в качестве закваски.Протеолитическая активность молочнокислых бактерий может быть ограничена, поскольку она осуществляется протеиназой клеточной оболочки и пептидазами. Однако их вклад важен, поскольку они смогли удалить 56% белка из отходов крабов в стерильном SmF Lactobacillus для исследования одновременного производства хитина и молочной кислоты [54].
Полное удаление белка из панциря ракообразных в процессе очистки хитина является труднодостижимым, поэтому было проведено несколько исследований для достижения высокого уровня удаления белка.Некоторые из этих исследований включали сочетание микроорганизмов более чем на одной стадии процесса. В связи с этим процесс глубокой ферментации молочной кислоты (SmF) был расширен с 0,25 до 300 л для очистки хитина из панциря креветок ( Crangon crangon ). В процессе использовали анаэробную, протеолитическую обогащенную культуру с дефицитом хитиназы из мясного фарша для депротеинизации и смешанную культуру LAB из биойогурта для декальцификации. Удаление белка в течение всего процесса в течение 40 часов показало эффективность 89% –91%, в то время как декальцификация молочнокислыми бактериями составила 85% и 90% в течение следующих 40 часов.Эти авторы сообщили, что хитозан, полученный путем деацетилирования хитина биологическим способом, имел более высокую вязкость, чем вязкость, определенная с хитозаном, полученным химическим путем [57]. Это согласуется с предыдущим отчетом Pacheco et al. [23], в которых метод твердофазной ферментации молочной кислоты (SSF) позволил избежать чрезмерной деполимеризации и потери кристалличности во время производства хитозана методом деацетилирования замораживанием-насосом-оттаиванием.
Другие сообщенные комбинации включают использование бактерий и грибов, таких как Kurthia gibsonii и Aspergillus flavus , которые были выделены из ферментированного молока и хлеба, соответственно.В этом исследовании хитин был успешно экстрагирован из панциря креветок, Fenneropenaeus semisulcatus и Fenneropenaeus indicus , с использованием двухэтапного процесса SmF с использованием 3-дневной бактериальной культуры бульона для DM в течение 24 часов, а затем 5 дней культивирование грибного бульона в течение 72 ч для ДП. Выход хитина, достигаемый в наилучших условиях для микробной обработки, был ниже, чем при химическом методе [58]. Комбинация грибов и бактерий также изучалась для экстракции хитина из панцирей креветок в двухстадийной SSF с помощью Lactobacillus brevis и Rhizopus oligosporus [59].Авторы заявили об извлечении безбелкового хитина с помощью микроорганизмов GRAS (общепризнанных как безопасных). При этом высвобожденные гидролизаты белка (120,56 мг белка / г) имели диапазон молекулярной массы (M W ) от 25 × 10 3 до 11 × 10 3 Да. Наибольшая концентрация астаксантина, извлеченного из жидкости, составила 8,78 мкг / г. Белковые гидролизаты и астаксантин показали активность по улавливанию радикалов в анализе DPPH с IC 50 1,13 ± 0,03 мг / г и 2.02 ± 0,01 мкг / г соответственно. Очищенный хитин имел молекулярную массу 1313 кДа, сохраняя высокий кристаллический индекс (I CR 87,5%) и степень ацетилирования 93,67% [59]. Хитины, полученные путем последовательной ферментации молочнокислых бактерий и грибов из отходов креветок, имели более высокую молекулярную массу и кристалличность, чем коммерческие хитины [59]. Эти характеристики могут улучшить свойства нановолокон, полученных из хитинов, экстрагированных с использованием L. brevi s с и без дополнительных инокуляций R.олигоспор . Нановолокна, полученные с использованием этого биологического хитина, показали значительно более высокий модуль Юнга, чем у коммерческого продукта. Метод экстракции оказывает существенное влияние на механические свойства получаемых нановолокон, что является важной характеристикой полимерных армирующих материалов [60].
Bacillus licheniformis продуцировал протеазы при выращивании в среде, содержащей порошок отходов креветок в качестве единственного источника углерода и азота. Процент удаления белка после 3 ч гидролиза при 60 ° C и соотношении фермент / субстрат 5 Ед / мг белка составил около 81% [61].
Ghorbel-Bellaaj et al., [62] изучали экстракцию хитина с помощью продуцирующих протеаз Bacillus pumilus , Bacillus mojavencis , B. . Достигнутая DP составляла более 80% для всех протестированных штаммов и продуцировала гидролизаты белка отходов креветок с активностью по улавливанию радикалов. Однако DM был относительно низким, 67%, но DM улучшился при добавлении глюкозы в среду.
Другая бактерия, часто используемая для продуцирования протеазы, — это Serratia marcescens , и наряду с последовательной обработкой L. plantarum дала лучший выход хитина (82,56%) из биомассы панциря омара с общим DP 87,19% и общим СМ 89,59% [63].
Молочная кислота | Encyclopedia.com
ОБЗОР
Молочная кислота (LAK-tik AS-id) — это бесцветная сиропообразная жидкость без запаха, которая существует в двух изомерных формах: D-молочная кислота и L-молочная кислота.Изомеры — это две или несколько форм химического соединения с одинаковой молекулярной формулой, но с разными структурными формулами и разными химическими и физическими свойствами. Форма D вырабатывается в ходе метаболических реакций, протекающих в мышечной ткани, а форма L вырабатывается дрожжевыми клетками. Синтетическое производство молочной кислоты приводит к продукту, состоящему из равных количеств форм D и L, смеси, известной как рацемическая смесь.
ОСНОВНЫЕ ФАКТЫ
ДРУГИЕ НАИМЕНОВАНИЯ:
2-гидроксипропановая кислота; ± -гидроксипропановая кислота; молочная кислота
ФОРМУЛА:
CH 3 CHOHCOOH
ЭЛЕМЕНТЫ:
Углерод, водород, кислород
ТИП СОЕДИНЕНИЯ:
Карбоновая кислота (органическая)
СОСТОЯНИЕ:
Жидкость
90.08 г / моль
ТОЧКА ПЛАВЛЕНИЯ:
16,8 ° C (62,2 ° F)
ТОЧКА КИПЕНИЯ:
Не применимо; разлагается при нагревании
РАСТВОРИМОСТЬ:
Хорошо растворим в воде и этаноле; мало растворим в эфире
Молочная кислота была впервые открыта в 1780 году шведским химиком Карлом Вильгельмом Шееле (1742–1786), который назвал свое открытие «молочной кислотой». Две изомерные формы кислоты были впервые идентифицированы в 1863 году немецким химиком Йоханнесом Вислиценусом (1835–1902), а это соединение было впервые коммерчески произведено в 1881 году американским химиком Чарльзом Э.Эйвери. Эйвери запатентовал свое изобретение в 1885 году и построил завод по производству молочной кислоты в Литтлтоне, штат Массачусетс.
Около 30 миллионов килограммов (72 миллионов фунтов) молочной кислоты ежегодно производится в Соединенных Штатах. Самый распространенный метод производства — это ферментация глюкозы дрожжами.
КАК ЭТО ПРОИЗВОДИТСЯ
В мышечных клетках молочная кислота является продуктом анаэробного дыхания, процесса, при котором глюкоза окисляется в отсутствие кислорода для производства энергии, необходимой клеткам.Хотя некоторое количество молочной кислоты всегда вырабатывается в мышечных клетках в очень низких концентрациях, она имеет тенденцию накапливаться во время упражнений, когда клетки не получают достаточного количества кислорода для нормального метаболизма кислорода. Молочная кислота, вырабатываемая во время упражнений, остается в организме лишь на короткое время, иногда менее чем за тридцать минут. Он метаболизируется в мышечных клетках, где он был произведен, что приводит к выработке энергии, углекислого газа, воды и других продуктов.
Молочная кислота также производится дрожжами в процессе ферментации.Ферментация — это процесс, при котором дрожжевые клетки превращают глюкозу в спирт и углекислый газ. В дрожжевых клетках при брожении используется почти тот же фермент, что и в мышечных клетках при анаэробном дыхании. Фермент мышечной клетки и дрожжевой фермент различаются только ориентацией одной группы атомов, что приводит к образованию D-изомера в одном случае и L-изомера в другом.
Синтетический процесс производства молочной кислоты был впервые внедрен в 1963 году. Этот процесс начинается с добавления цианистого водорода (HCN) к ацетальдегиду (этанал; CH 3 CHO), что приводит к образованию лактонитрила (CH 3 CH 2 OCN).Затем лактонитрил гидролизуют с использованием сильной кислоты, такой как серная кислота, в качестве катализатора, чтобы получить молочную кислоту.
Интересные факты
- На протяжении большей части столетия спортсмены и физиологи считали молочную кислоту основной причиной усталости во время упражнений высокой интенсивности. Однако ученые узнали, что молочная кислота действительно помогает предотвратить мышечную усталость. Мышечная болезненность, которую раньше считали вызванной молочной кислотой, с большей вероятностью является результатом повреждения мышечных клеток, вызванного чрезмерным употреблением.
- Молочная кислота, присутствующая на коже, привлекает комаров.
- Молочная кислота в организме существует в ионной форме, известной как лактат.
ОБЫЧНОЕ ИСПОЛЬЗОВАНИЕ И ПОТЕНЦИАЛЬНАЯ ОПАСНОСТЬ
Молочная кислота в основном используется в Соединенных Штатах в качестве пищевой добавки, где она действует как подкислитель и вкусовая добавка. Подкислитель — это соединение, которое обеспечивает кислую среду для пищевых продуктов, как в случае с йогуртом, пахтой, квашеной капустой, зелеными оливками, солеными огурцами и другими кислыми продуктами.В качестве вкусовой добавки он придает еде и напиткам терпкий или острый вкус, а также действует как консервант, предохраняющий их от порчи. Молочная кислота также имеет ряд важных промышленных применений, наиболее важным из которых является производство других органических химикатов, особенно этиллактата, акриловой кислоты, пропиленгликоля и полимера, известного как полиактид. Полиактид используется в производстве пластиковой пленки, волокна, упаковочного материала и наполнителей. Другие коммерческие и промышленные применения молочной кислоты включают:
- В качестве протравы при крашении;
- В качестве растворителя нерастворимых в воде красителей;
- Для обработки шкур животных при изготовлении кожаных изделий;
- В качестве катализатора при производстве некоторых видов пластмасс; и
- В качестве добавки в гальванических ваннах.
Слова, которые нужно знать
- КАТАЛИЗАТОР
- Материал, который увеличивает скорость химической реакции без каких-либо изменений в своей химической структуре.
- ЭЛЕКТРИЧЕСКОЕ ПОКРЫТИЕ
- Нанесение слоя никеля, серебра или золота на металл другого типа с помощью электрического тока.
- Брожение
- Процесс, с помощью которого дрожжи превращают глюкозу в спирт и диоксид углерода.
- ГИДРОЛИЗ
- Процесс, при котором соединение реагирует с водой с образованием двух новых соединений.
- ISOMER
- Одна из двух или более форм химического соединения с одинаковой молекулярной формулой, но разными структурными формулами и разными химическими и физическими свойствами.
- МЕТАБОЛИЗМ
- Процесс, который включает в себя все химические реакции, происходящие в клетках, посредством которых расщепляются жиры, углеводы и другие соединения с образованием энергии и соединений, необходимых для создания новых клеток и тканей.
- MORDANT
- Вещество, используемое при крашении и печати, которое химически вступает в реакцию как с красителем, так и с окрашиваемым материалом, помогая удерживать краситель на материале надолго.
- СИНТЕЗ
- Химическая реакция, в которой некоторый желаемый химический продукт образуется из простых исходных химикатов или реагентов.
Молочная кислота в нормальных концентрациях не представляет опасности для здоровья или безопасности людей или других животных. Однако одним из последствий для здоровья, связанных с молочной кислотой, является состояние, известное как подагра, тип артрита, который вызывает сильную боль в суставах. Подагра вызвана накоплением мочевой кислоты в крови. Поскольку молочная кислота блокирует выведение мочевой кислоты из организма, у людей с избыточным накоплением молочной кислоты, обычно вызванным высоким потреблением алкоголя, может развиться избыток кристаллов мочевой кислоты в крови и суставах, что приводит к подагре.
ДОПОЛНИТЕЛЬНАЯ ИНФОРМАЦИЯ
«Дыхание клеток». SparkNotes. http://www.sparknotes.com/testprep/books/sat2/biology/chapter6section1.rhtml (по состоянию на 14 октября 2005 г.).
Дрейк, Джефф. «Лактатный челнок. Вопреки тому, что вы слышали, молочная кислота — ваш друг». Bicycling (август 1992 года): 36.
Фрил, Джоэл. «Все спортсмены: плохой рэп с молочной кислотой». Электронные советы Ultrafit для спортсменов на выносливость. Октябрь 2004 г. 7, № 10. http://www.ultrafit.com/newsletter/october04.html # Joe (доступ 14 октября 2005 г.).
«Молочная кислота». Дж. Т. Бейкер. http://www.jtbaker.com/msds/englishhtml/l0522.htm (по состоянию на 14 октября 2005 г.).
Роджерс, Палмер, Цзян-Шин Чен и Мэри Джо Зидвик. Производство органических кислот и растворителей, часть I: уксусная, молочная, глюконовая, янтарная и полигидроксиалкановая кислоты . Раздел 2: Молочная кислота. Доступно в Интернете по адресу http://141.150.157.117:8080/prokPUB/chaphtm/306/04_00.htm (по состоянию на 14 октября 2005 г.).
Границы | Вклад молочнокислых бактерий в сложность кишечной микробиоты: свет и тень
Краткая история молочнокислых бактерий
Молочнокислые бактерии (LAB) включают большое количество родов бактерий, среди которых наиболее известны лактобациллы, лактококки, энтерококки, стрептококки, лейконостоки и педиококки.Эти роды различаются по морфологии, pH и солеустойчивости, температурному оптимуму, средам обитания и патогенному потенциалу. В настоящее время очень сложно провести четкую границу между полезными и вирулентными видами, поскольку некоторые проблемные признаки больше связаны с штаммом, чем с видами. Однако лактобациллы и лактококки считаются GRAS (обычно считаются безопасными).
ЛАБ были одними из первых живых организмов на Земле: они появились около трех миллиардов лет назад, в переходный период от анаэробиоза к аэробиозу.По-видимому, они хорошо приспособлены как к анаэробным, так и к аэробным условиям жизни, поскольку несут все необходимые белки для дыхания и несколько ферментов, участвующих в ферментативных путях. Тем не менее, в ходе эволюции они так и не приобрели способности биосинтезировать гем, важный компонент цитохромов, поэтому они могут перейти на респираторный метаболизм только с добавлением экзогенного гемина. В случае патогенных стрептококков они могут вычитать гем из гемоглобина хозяина с помощью гемолизина, белка, который считается фактором вирулентности.Следовательно, большинство видов LAB вынуждены жить за счет ферментационного метаболизма с более низким выходом энергии (Carr et al., 2002).
Кроме того, неспособность LAB синтезировать молекулы гема означает, что они не могут получить функциональные гемовые ферменты, такие как пероксидазы и каталазы, которые являются наиболее эффективными белками, участвующими в захвате гидроксильных радикалов. Это приводит к снижению устойчивости к токсичности O 2 . Несмотря на это, LAB могут переносить кислород и относятся к микроаэрофильным организмам (Брюханов, Нетрусов, 2007).
Эти бактерии разработали некоторые стратегии, чтобы заменить гемовые ферменты эффективными антиреактивными формами кислорода (АФК), например, использование металлов в качестве поглотителей радикалов: цинк представляет собой ионную ловушку для кислородных радикалов (Salvatore et al., 2007) марганец может действовать как система, подобная супероксиддисмутазе (De Angelis and Gobbetti, 1999), в то время как селен действует в белках, несущих селеноцистеин (Calomme et al., 1995). Известно, что виды LAB способны накапливать высокие внутриклеточные количества марганца (до 25 мМ), селена и цинка.
В процессе эволюции, из-за низкого прироста энергии в результате ферментации и тяжелых условий жизни в «окислительном» мире (таком, как тот, в котором мы сейчас живем), LAB были вынуждены специализировать свой метаболизм на защите от стресса, а не на приобретении стратегические биосинтетические способности. Поэтому они развили отношения симбионтов / паразитов с растениями и животными, которые могут поставлять витамины, белки и аминокислоты. Стоит отметить, что LAB приобрели способность распознавать несколько сахаров, таких как, например, ксилозу, целлобиозу, рибозу, арабинозу, глюкозу и фруктозу, прежде чем они развили способность ферментировать лактозу до лактата, что стало возможным только после того, как млекопитающие экспансия на Земле.Поэтому они сначала колонизировали экологические ниши фруктов и овощей, а затем сыр, вино и особенно молоко, которые составляют их среду обитания, богатую лактозой (Carr et al., 2002).
Возможно, что в раннем периоде жизни катаболизм сахара не был таким преобладающим, как в нынешнем метаболизме LAB, и что параллельные пути, такие как декарбоксилирование кислот и аминокислот и деструкция аргинина (ADI), могли быть одновременно активированы для получения энергии в плохих средах, таких как в качестве продуктов лизиса клеток (рибоза, жирные кислоты, аминокислоты) (Konings, 2006).
Однако комменсальные LAB, живущие как в кишечнике, так и в других экологических нишах слизистых оболочек, хотя и питаются обильными питательными веществами, по-прежнему ведут стрессовую жизнь и часто вынуждены справляться с антагонистическими факторами хозяина, а также с дрожжами или бактериями, живущими в одной среде обитания. Эти суровые условия позволили развить интересные метаболические особенности и перекрестные помехи.
Лаборатория биохимии
Большинство биохимических путей LAB до сих пор полностью выяснены из-за стратегической важности этих бактерий в пищевой промышленности как в качестве заквасок, так и в качестве агентов биоконтроля для порчи пищевых продуктов.Недавний интерес к LAB как установленным пробиотическим агентам дополнил информацию о метаболизме LAB и его роли в контексте экосистемы кишечника.
LAB Energy Metabolism
Энергетический метаболизм
LAB в основном основан на молочной ферментации, ADI, кислотном и аминокислотном декарбоксилировании. В то время как ферментация сахара играет истинную энергетическую роль, другие пути могут решать другие и более сложные функции. Молочная ферментация ценится как в кишечных, так и в промышленных ЛАБ, поскольку ее конечный продукт, лактат, вызывая подкисление, обеспечивает контроль над менее дружелюбными бактериями.
Сахарное брожение
Гомолактическая ферментация. Гомолактическая ферментация всегда происходит в результате гликолиза. Облигатные гомоферментативные LAB (педиококки, лактококки, стрептококки и некоторые лактобациллы) продуцируют 100% молочную кислоту посредством пути Эмбдена-Майероффа из различных гексоз, которые интернализуются с помощью определенных мембранных переносчиков, таких как пермеазы и симпортеры, а затем изомеризуются в глюкозу или фруктоза. Галактоза представляет собой исключение, поскольку она может быть либо изомеризована в глюкозу посредством реакции Лелуара, опосредованной высокоэнергетическими соединениями (UDPGlu-UDPGal), либо перейти к гликолизу через D-тагатозо-6-фосфатный путь, образуя диидроксиацетонфосфат, следовательно, фосфат глицеральдегида (Рисунок 1). .
Рисунок 1. Метаболизм галактозы в молочнокислых бактериях .
Когда присутствует дисахарид, такой как лактоза или сахароза, он может гидролизоваться до двух моносахаридов во внешней среде или, скорее, поглощаться как дисахарид, а затем гидролизоваться внутри клетки. Из каждой экзозы в конце гликолитического процесса образуются два моля молочной кислоты в результате восстановления пирувата до лактата посредством повторного окисления НАДН до НАД + .Молочная кислота может быть в оптической форме D или L, в зависимости от генетических детерминант каждого вида, кодирующей либо D-лактат (D-LDH), либо L-лактат (L-LDH) дегидрогеназу, соответственно. Некоторые виды могут продуцировать как D, так и L лактат в результате активности рацемазы или из-за наличия генетических детерминант для обеих изоформ ЛДГ. В этом случае вторая генетическая детерминанта может быть получена путем горизонтального переноса генов. Таким образом, наиболее распространенный катаболический путь, то есть превращение дисахарида лактозы в лактат, генерирует четыре моля молочной кислоты и четыре моля АТФ.При этом газ не образуется. Этот низкий выигрыш энергии иногда может быть улучшен симпортом протон-субстрат, то есть экскрецией молочной кислоты, созданием протонного градиента: поскольку эта система является электрогенной, она может увеличить выход энергии LAB.
Факультативные гомоферментеры могут направлять часть пировиноградной кислоты, которая образуется при гликолизе, на производство формиата, ацетата и этанола. Пируват-формиат-лиаза может превращать пируват (C3) в формиат (C1) и ацетилCoA (C2). Последний может подвергаться трансферазной реакции в ацетилфосфат, а затем превращаться в ацетат, что приводит к синтезу АТФ.Ацетат может либо накапливаться в среде для выращивания, либо, альтернативно, восстанавливаться до этанола с помощью ацетальдегида с потреблением НАДН, в зависимости от pH, и снижением доступности коферментов пиридина. Этот путь позволяет получить один дополнительный моль АТФ, но при этом вырабатывается меньше молочной кислоты. Поскольку формиат может декарбоксилироваться / окисляться, дополнительный моль CO 2 может быть произведен этим путем (газообразующие бактерии).
Гетеролактическая ферментация. Гетеролактическая ферментация касается LAB, в которых отсутствует гликолитический фермент фруктозо-1,6-бисфосфатальдолаза ( Leuconostoc , Oenococcus и некоторые виды Lactobacillus ), поэтому они не могут метаболизировать гексозы по пути Эмбдена-Майероффа.Следовательно, они используют пентозофосфатный путь с транскетолазной реакцией, соединяющей гликолиз с трехуглеродным метаболитом глицеральдегид-3-фосфатом. Оставшаяся единица C2, ацетилфосфат, затем превращается в этанол или в ацетат (последняя реакция, приводящая к дополнительному мольному увеличению АТФ), в зависимости от соотношения NAD + / NADH, как ранее описано для факультативных гомоферментеров. Тем не менее, производство ацетата происходит не так часто из-за необходимости повторного окисления НАДН.Энергетический выход транскетолазного пути ниже, чем при гомолактической ферментации, дающей только 1 моль АТФ, 0,5 моль лактата и 0,5 моль этанола или ацетата на моль израсходованной гексозы, плюс один моль CO 2 преобразование гексозы / пентозы (декарбоксилированием). Итак, гетероферментативный путь — добыча газа.
Все гетероферментативные LAB также могут разлагать пентозы, в то время как не все гомоферментеры также разлагают пентозу.
Выведение аргинина
Помимо фосфорилирования уровня сахара, LAB может получать энергию от ADI до цитруллина и от расщепления цитруллина до орнитина и карбамоилфосфата.Последний затем превращается в АТФ и диоксид углерода или, скорее, используется для биосинтеза пиримидина. Этот путь также генерирует два моля аммиака (один от перехода аргинин-цитруллин, второй от гидролиза карбамоилфосфата), полезного для компенсации кислотности, вызванной катаболизмом сахара в молочную кислоту, уксусную кислоту и муравьиную кислоту, как в гомо-, так и в гетероциклических соединениях. -условия заквашивания (De Angelis et al., 2002).
С другой стороны, орнитин обменивается на внеклеточный аргинин в антипортовой системе орнитин / аргинин, обеспечивая непрерывность реакций.В вине были выбраны LAB, обладающие этим путем, поскольку винная среда все еще богата аргинином после дрожжевого спиртового брожения. Хотя путь ADI чем-то похож на инвертированный цикл мочевины, с эволюционной точки зрения эти пути вообще не связаны (Liu et al., 1996).
Декарбоксилирование кислот и аминокислот
Помимо вторичного участия в буферизации pH, путь аргининдезиминазы в основном представляет собой путь генерации АТФ, полезный для дополнения фосфорилирования на уровне субстрата.Напротив, декарбоксилирование кислот и аминокислот решает более сложные роли, и их эволюционная история заслуживает внимания. Декарбоксилирование сочетается с электрогенными антипортовыми системами, генерирующими протонный градиент через цитоплазматическую мембрану, и вся система поддерживает двойную роль: контроль кислотности и генерацию протонной движущей силы (PMF), то есть дополнительной энергии в организмах, неспособных к дыханию. В случае кислотного декарбоксилирования реакция касается дикарбоновых и трикарбоновых кислот, таких как малат и цитрат, и продукты реакции не являются информационными молекулами.
Было высказано предположение, что на ранних этапах эволюции активность кислой декарбоксилазы была направлена в основном на борьбу с подкислением. Позже механизмы сохранения свободной энергии были приобретены путем сочетания декарбоксилирования с электрогенными антипортовыми системами, что позволило получить более сложные клеточные реакции, такие как выработка метаболической энергии и лучшая общая устойчивость к кислотному стрессу. Считается, что эти пути эволюционировали в результате горизонтального переноса генов, что позволило генам антипортерных белков располагаться в том же опероне, что и гены декарбоксилазы, таким образом подвергаясь тому же транскрипционному контролю (Макарова и Кунин, 2007). С другой стороны, геномы LAB, как известно, кодируют очень изменчивое количество генов, которое предполагает, что в процессе эволюции приобретение / потеря генов сыграло решающую роль в определении нынешней ситуации.Размер генома LAB варьируется от 1,8 МБ для L. gasseri до 3,3 МБ для L. plantarum , а также содержание G + C значительно варьируется (от 34 до 46%) (Siezen et al., 2004), что свидетельствует о частых встречах. события рекомбинации. Кроме того, присутствие растворимых кислых декарбоксилаз и мембраносвязанных обменников предшественников / продуктов в LAB (Lucas et al., 2005) представляет собой промежуточное событие между простым декарбоксилированием, происходящим в цитоплазме строго анаэробов (например, Clostridia и Fusobacterium ). ), где свободная энергия не сохраняется, а связанные с мембраной комплексы (включая декарбоксилазы и переносчики), обнаруженные у гамма-протеобактерий, у которых происходит активная экструзия натрия (Макарова и др., 2006).
Что касается декарбоксилаз аминокислот, их физиологические роли аналогичны функциям кислых декарбоксилаз (Molenaar et al., 1993). Тем не менее, были обнаружены некоторые дополнительные функции, такие как взаимодействие бактерий и бактерий и коммуникация между хозяином и симбионтом, которые являются информационными молекулами продуктов. Глутамат можно декарбоксилировать до гамма-амино-бутирата (ГАМК), гистидина до гистамина, фенилаланина до β-фенилэтиламина, тирозина до тирамина, триптофана до триптамина, орнитина до путресцина, лизина до кадаверина (Konings, 2006).
Декарбоксилирование аминокислот катализируется PLP или зависимыми от пировоил ферментами, которые могут быть растворимыми (Pessione et al., 2005) или располагаться на мембране. В Enterococcus был описан мембраносвязанный тирозин и фенилаланиндекарбоксилаза (Pessione et al., 2009): это клеточное расположение может способствовать лучшему взаимодействию между ферментом декарбоксилазы и белком-переносчиком. Это может стать дальнейшим эволюционным шагом к более эффективной системе получения энергии и противодействия кислотному стрессу у менее устойчивых к кислоте представителей рода, чем у других LAB (Рисунок 2).
Рис. 2. Увеличенное изображение протеомных карт обогащенной мембраной белковой фракции продуцента тирамина и бета-фенилэтиламина Enterococcus faecalis , выращенных в отсутствие (A) или в присутствии (B) тирозина. Кружком обозначен фермент TDC (тирозин / фенилаланин декарбоксилаза).
Некоторые производимые амины представляют собой биоактивные молекулы (гистамин, тирамин, триптамин, бета-фенилэтиламин, ГАМК), действующие на уровне сосудистой или центральной нервной системы человека-хозяина (Moreno-Arribas et al., 2003), в то время как другие являются информационными посланниками для микробного взаимодействия и для контроля стресса (по крайней мере, для путресцина у некоторых видов бактерий). Все они могут подвергаться дальнейшим биопревращениям под действием моноамино- и диаминооксидаз. Таким образом, эти пути можно рассматривать как «пути бегства» во время истощения запасов сахара или чрезмерного подкисления (т.е. стресс стационарной фазы), но также как средства взаимодействия с внешней средой. Следует учитывать, что большинство геномов LAB несут гены как ферментов пути ADI, так и декарбоксилаз аминокислот: часто эти два пути реципрокно регулируются (Lactobacillus , Lactococcus ) (Lamberti et al., 2011a, b), но иногда они не конкурируют друг с другом и используются параллельно ( Enterococcus ), что подчеркивает их сходные, но разные физиологические функции. Протеомные исследования могут добавить полезную информацию о физиологических и средовых модуляциях, действующих на эти основные пути. С человеческой точки зрения, в то время как кислотные декарбоксилирования ценятся, потому что они снижают кислотность (например, в вине), декарбоксилирование аминокислот и ADI рассматриваются как проблемные, поскольку они могут приводить к образованию аммиака и нежелательных биоактивных аминов, таких как гистамин. , то есть медиатор аллергии или тирамин, который участвует в гипертонии и более тяжелых синдромах, таких как кровоизлияние в мозг (Millichap and Yee, 2003).И наоборот, ГАМК и бета-фенилэтиламин являются важными молекулами, поскольку они могут контролировать аппетит, настроение и активность гладких мышц, что будет лучше объяснено в следующих параграфах (Inoue et al., 2003).
LAB протеолитическая система
Выбор использования аминокислот не только в качестве источника азота, но и для энергетического метаболизма требует высокой доступности аминокислот, и по этой причине LAB разработали сложные протеолитические и пептидолитические системы. Они включают три компонента: (1) внеклеточные или мембраносвязанные протеазы (в основном PrtP и CEP), гидролизующие белки в олигопептиды; (2) мембранные транспортные белки (Opp, Dtp и Dpp АТФ-связывающие кассетные транспортеры), катализирующие внутриклеточное поглощение олигопептидов, ди / трипептидов и дипептидов, соответственно; (3) внутриклеточные пептидазы, которые расщепляют пептиды на свободные аминокислоты (Konings, 2002).
У лактококков протеазы преимущественно закреплены за клеточной оболочкой (протеазы CEP), за исключением Lactococcus lactis subsp. cremoris , который способен секретировать протеиназы. У Lactobacilli некоторые из них имеют внеклеточную локализацию ( Lb. lactis и Lb. helveticus ), в то время как другие связаны с клеточной стенкой ( Lb. delbruekii ssp bulgaricus ). Большинство протеолитических ферментов LAB синтезируются как пре-проферменты, сигнальный пептид которых расщепляется при транслокации мембраны: полученная мембраносвязанная пропротеиназа подвергается автокаталитическому расщеплению для получения своей зрелой, активной конформации.Эти протеазы представляют собой сериновые протеиназы с высокой молекулярной массой и оптимумом pH 5,5-6,5, которые совместимы со средой, в которой живут LAB.
Однако могут присутствовать и другие системы, обеспечивающие доступность свободных аминокислот на внешней стороне цитоплазматической мембраны. Фактически, некоторые пептидазы были обнаружены на внутренней стороне цитоплазматической мембраны путем распознавания антител (PepX в Lactococci) или предположительно были связаны с анионо-специфической пермеазой, хотя они были предсказаны цитозолом на основе данных последовательностей генома. .Например, PepA не несет сигнального пептида или истинного гидрофобного домена, который мог бы оправдать его расположение на мембране. Тем не менее, его высокая субстратная специфичность в высвобождении N-концевых кислотных остатков из пептидов может объяснить его роль в поставке свободного глутамата для антипортера пути глутамат-ГАМК на мембранном уровне. В E. faecalis аналогичную функцию может выполнять ClpP: эта протеаза способна гидролизовать тетра- и трипептиды, содержащие сайты узнавания тирозин-триптофан или тирозин-лейцин (Katayama et al., 1988). Это может привести к доступности свободного тирозина для пути декарбоксилирования тирозина-тирамина.
Важную роль, которую выполняют как высвобождаемые, так и связанные с клеточной стенкой LAB протеиназы в кишечнике человека, является переваривание не полностью гидролизованных белков. Они могут укорачивать длинные и средние пептиды до более мелких. Эту роль также могут выполнять внутриклеточные пептидазы, высвобождаемые автолизом LAB-клеток. Поскольку протеолитическая система LAB получила развитие в экологической нише молока, она особенно подходит для гидролиза молочного белка.Помимо их пищеварительно-ассимиляционной функции, эти ферменты привлекли большое внимание, потому что они способны генерировать биоактивные пептиды как из казеинов, так и из белков молочной сыворотки (Law and Haandrikman, 1997). Дневные белки перевариваются в желудке и тонком кишечнике человека эндогенными протеолитическими ферментами. Это пищеварение дополняется протеазами, поставляемыми микрофлорой кишечника. Казеин состоит из четырех основных белков: альфа s1 казеина, альфа s2 казеина, бета-казеина и k-казеина, различающихся аминокислотной последовательностью, фосфорилированием и степенью гликозилирования, индексом гидропатичности.Гидролиз всех этих компонентов, соотношение которых составляет примерно 38: 11: 38: 13, может привести к образованию биоактивных молекул, действующих на разных биологических уровнях. Точно так же гидролитическое расщепление белков молочной сыворотки (альфа-лактоальбумин, бета-лактоглобулин, лактоферрин и иммуноглобулины) также может генерировать биоактивные пептиды. Протеолитические ферменты, которые высвобождаются LAB, были проанализированы с помощью 2DE-MS и оказались очень разными у разных видов и штаммов LAB, что привело к разному пулу биоактивных пептидов.Однако их активность, хотя и очень высокая при нормальном рН кишечника (6,5–7,0), оказалась различной в отношении альфа-1 и бета-казеина (Hébert et al., 1999). Полученные пептиды обычно стабильны, хотя необходимы дополнительные исследования для оценки их реальной продолжительности жизни in vivo . Наиболее интересные биоактивные пептиды, являющиеся результатом протеолитической активности LAB, участвуют в регуляции иммунной системы, сердечно-сосудистой системы и центральной нервной системы, пищевых добавках (например, предположение о наличии металлов) и антимикробных функциях (соединения, подобные антибиотикам).Иногда каждый отдельный пептид может выполнять две или несколько различных физиологических функций (Clare and Swaisgood, 2000).
Лаборатория биосинтетических возможностей
Несмотря на свои ограниченные биосинтетические способности (особенно в отношении синтеза аминокислот и витаминов), LAB может продуцировать представляющие интерес молекулы, среди которых наиболее интересными являются экзополисахариды (EPS) и фруктоолигосахариды (FOS), короткоцепочечные жирные кислоты (SCFA), конъюгированная линолевая кислота. кислоты (CLA) и селенопротеины (рис. 3).
Рис. 3. Молекулы, влияющие на нутрицевтический и пробиотический потенциал молочнокислых бактерий .
EPS и FOS
Как и большинство бактерий, LAB может синтезировать структурные полисахариды клеточной стенки (PS), такие как пептидогликан и липотейхоевые кислоты, а также внеклеточные полимеры. Последние включают как капсульный PS, где PS ковалентно связан с поверхностью клетки, так и EPS, которые образуют слабо связанный слой слизи, который также может секретироваться в окружающую среду.
EPS можно разделить на гомополисахариды (Homo-EPS) и гетерополисахариды (Hetero-EPS). Гомо-EPS состоит из остатков D-глюкозы (глюканы) или D-фруктозы (фруктаны) с разными типами связи и степенью разветвления. Гетеро-EPS сконструированы из нескольких копий олигосахарида и имеют небольшое структурное сходство друг с другом: глюкоза, галактоза, ксилоза, манноза, арабиноза и рамноза являются наиболее представленными сахарами, но также могут иногда присутствовать аминосахары и полиолы. как глюкуроновая кислота.Они часто сильно разветвлены с разными типами связывания. С биохимической точки зрения синтез Hetero-EPS — энергоемкий процесс, требующий четырех отдельных реакций: (1) интернализация сахара; (2) синтез предшественников сахарных нуклеотидов, обеспечивающих энергию для реакции полимеризации; (3) сборка повторяющейся единицы моносахарида за счет комбинированного действия различных типов гликозилтрансфераз, расположенных на уровне цитоплазматической мембраны; (4) экспорт EPS.Глюкозо-6-фосфат является ключевым метаболитом, который может быть направлен с гликолитического катаболического пути на синтез UDP-глюкозы и dTDP-глюкозы, но также путь Лелуара, при его наличии, может быть использован для преобразования галактозы в UDPGal. и UDPGlu. В мезофильных LAB ферменты, необходимые для поглощения сахара и для синтеза сахарных нуклеотидов, кодируются хромосомами, тогда как генетические детерминанты для EPS специфических ферментов локализуются в плазмидах (Laws et al., 2001). Напротив, для синтеза гомо-ЭПС требуется только одна внеклеточная гликозилтрансфераза, а сахароза используется в качестве субстрата вместо активированных сахаров.Таким образом, этот фермент может синтезировать альфа-глюканы (декстран, реутеран, мутан) или бета-фруктаны (леваны и инулин). Низкомолекулярные олигосахариды известны как ФОС и глюкоолигосахариды (рис. 4).
Рис. 4. Гомополисахариды, биосинтезируемые молочнокислыми бактериями .
На основании идентификации ящиков связывания сахарозы в геномных последовательностях LAB было идентифицировано несколько предполагаемых генов, кодирующих как глюкансахаразы, так и фруктансахаразы (van Hijum et al., 2006).
Как правило, LAB направляют небольшой процент своих сахарных субстратов на биосинтез EPS, физиологическая роль которого, вероятно, разнообразна и сложна: благодаря своим высоким водосвязывающим свойствам EPS представляют собой фактор защиты от высыхания и осмотического стресса (Kumar et al. ., 2007). Предполагалась также их роль в защите бактериальной клетки от фаговой атаки (Moineau et al., 1996), но более поздние эксперименты не смогли выявить значительный фенотип устойчивости к фагам у штаммов-продуцентов EPS (Deveau et al., 2002). То же самое верно для первой гипотетической роли внеклеточной энергии / запаса углерода, поскольку у большинства видов продуцентов отсутствует начало генов, необходимых для их собственной деградации EPS (Badel et al., 2011). Следовательно, этот резерв может быть полезен только в случае синтрофической / симбиотической жизни с другими бактериями. Фактически, EPS биосинтезируются большинством видов LAB под контролем кворума, и они связаны с образованием биопленок и адгезией к твердым поверхностям. В биопленке EPS также играет решающую роль в связывании основных катионов (Kumar et al., 2007). В этом контексте роль EPS как резерва углерода имеет смысл с учетом всего населения. Однако их основная биологическая функция in vivo заключается в защите бактериальных клеток от токсичных соединений (этанола, диоксида серы и токсичных ионов металлов), от антибиотиков и от иммунной системы хозяина, особенно от фагоцитоза. Вторая важная роль связана с адгезией к эукариотическим клеткам (растениям, поверхности зубов), распознаванием клеток, через связывание с лектинами (Cerning, 1990) и модуляцией иммунной системы.Krinos с соавторами (2001) продемонстрировали, что, как и капсульный PS, EPS также может влиять на поверхностную антигенность различных штаммов (разные EPS, приводящие к различным комбинациям), в конечном итоге приводя к сохранению или устранению определенных штаммов в экологической нише кишечника.
В последнее десятилетие фруктоолигосахариды (FOS = инулин и леваны) вызывают растущий интерес в качестве пребиотиков, то есть соединений, поддерживающих рост пробиотических организмов. Пребиотики — это молекулы, которые не метаболизируются человеком, но способствуют как адгезии, так и сохранению пробиотических бактерий в кишечной среде.Продукция инулина наблюдалась в Lactobacillus reuteri , и все инулосукразы имеют общий С-концевой мотив (LPXTG) для закрепления на клеточной стенке. Отображение этих ферментов на поверхности приводит к усилению адгезии бактерий к клеткам тканей человека, что способствует поддержанию здоровья слизистой мочеполовой системы. Таким образом, присутствие ферментов фруктансахаразы в комменсальных LAB может решить две различные проблемы: с одной стороны, способствуя колонизации и сохранению тканей, а с другой стороны, снабжению пребиотиками, изготовленными самим собой (например.g., fructans) комменсальную флору.
SCFA
Хотя SCFA образуются не в результате анаболической реакции, а скорее как конечные катаболиты энергетического метаболизма, их важность во взаимодействии бактерий-хозяев вызывает такой растущий интерес, что будет рассмотрено в этом параграфе. Наиболее изученными SCFA являются бутират, пропионат и ацетат. Lactobacilli может продуцировать SCFA путем ферментации пирувата, который образуется во время гликолитического пути, а также фосфокетолазного пути в условиях гетероферментации.Ацетат может способствовать подкислению окружающей среды, и это ценная особенность, поскольку LAB могут конкурировать с другими менее ацидофильными организмами (большинством патогенных видов бактерий), присутствующими в той же экосистеме. In vivo ацетат поступает в периферическое кровообращение и метаболизируется мышцами и другими тканями, в то время как пропионат поглощается печенью. Обе молекулы могут модулировать метаболизм сахара после стандартного приема пищи, снижая гликемию и улучшая чувствительность к инсулину. Утверждается, что бутират оказывает пагубное влияние на массу тела и ожирение, поскольку представляет собой дополнительный, легкодоступный источник энергии, который не вводится напрямую с диетой, но присутствует в результате метаболизма комменсальных бактерий (ферментация неперевариваемого PS в толстой кишки) (Turnbaugh et al., 2006). Как только SCFA всасывается в толстом кишечнике, он не только превращается в жир в печени, но и положительно регулирует гены хозяина, способствующие липогенезу и хранению жира в адипоцитах (DiBaise et al., 2008). Однако бутират легко абсорбируется колоноцитами, где он контролирует неконтролируемую пролиферацию, а также стимулирует дифференцировку, тем самым способствуя переключению с неопластических фенотипов на нормальные. Известно, что бутират по-разному влияет на нормальные колоноциты и опухолевые клетки толстой кишки.На первом он оказывает трофическое действие, индуцируя рост и пролиферацию, на втором он скорее вызывает остановку клеточного цикла, дифференцировку и апоптоз (Iacomino et al., 2001). Селективность бутирата в отношении двух разных популяций клеток считается хорошим оружием для лечения рака. Причина различных эффектов бутирата, вероятно, связана с его разными концентрациями в двух типах клеток: нормальные колоноциты быстро утилизируют (посредством митохондриального бета-окисления) бутират в качестве основного источника углерода / энергии, таким образом поддерживая синтез и пролиферацию АТФ.Остающиеся низкие уровни не могут подавлять развитие клеточного цикла или вызывать апоптоз (Boosalis et al., 2001). Напротив, клетки колоректальной опухоли из-за перехода к анаэробному метаболизму демонстрируют нарушенную способность окислительно катаболизировать бутират. Следовательно, в этом типе неопластических клеток его внутриклеточная концентрация выше. Этот высокий уровень бутирата может составлять критический порог, ответственный за активацию сигналов, которые вызывают остановку клеточного цикла, дифференцировку и апоптоз (Lupton, 2004).Влияние на остановку клеточного цикла частично связано с ингибированием специфической гистондеацетилазы (Davie, 2003), и это действие также может быть получено с помощью пропионата, но не ацетата. Некоторые авторы также предположили, что бутират и пропионат действуют синергетически (Minucci and Pelicci, 2006). Более того, сравнительные транскриптомные и протеомные исследования обработанных бутиратом и контрольных колоноцитов подчеркнули влияние бутирата на компоненты внеклеточного матрикса, которые важны для межклеточных взаимодействий, а также на ангиогенетический процесс, контролирующий уровень экспрессии VEGF (фактора роста эндотелия сосудов) и HIF-1 (фактор, индуцируемый гипоксией-1) (Pellizzaro et al., 2002). Наконец, бутират может контролировать окислительный и метаболический стресс на молекулярном уровне, усиливая восстановительные реакции (Sauer et al., 2007), и он может ингибировать синтез провоспалительных (IL12, TNF, гамма-IF) цитокинов, что также способствует развитию опухоли. контроль (Inan et al., 2000).
Разумно заключить, что бутират может способствовать здоровью хозяина при наличии неопластического риска, в то время как он является проблемным метаболитом в случае ожирения и нарушений обмена веществ.
CLA
LAB может также биосинтезировать CLA, которые являются позиционными и геометрическими изомерами цис9, цис12 окта-дека-диеновой кислоты (линолевая кислота). Пока что лучшие штаммы-продуценты принадлежат к родам Lactobacillus и Streptococcus . Наиболее часто встречающиеся изомеры линолевой кислоты имеют участки ненасыщенности в положениях 9, 11 или 10, 12, а именно Δ транс 9, цис 11 окта декадиеновая кислота и, в меньших количествах, Δ цис 10, транс 12 линолевая кислота.
CLA может оказывать положительное влияние на воспаление, рак (индукция апоптоза), метаболические нарушения (инсулинорезистентность, контроль массы тела) и сердечно-сосудистые заболевания.Тем не менее, наблюдались некоторые пагубные воздействия на здоровье млекопитающих, такие как стимулирующее опухоль действие или пагубное влияние на метаболизм глюкозы и липидный профиль. Есть данные, свидетельствующие о том, что разные изомеры CLA могут оказывать различное действие, наиболее опасным из которых является транс-10, цис-12-изомер. Кроме того, концентрации CLA играют роль в балансе благоприятных или вредных эффектов (Rose, 1997): антиканцерогенные эффекты наблюдаются при дозировке CLA 0,5–1% (вес: вес) от общей диеты, и исследования показали, что люди обычно выделяют 20 мг линолевой кислоты в день.Затем этот субстрат может быть доступен для бактерий, продуцирующих CLA, в кишечнике. Более высокое потребление линолевой кислоты с пищей может означать риски (Ewaschuk et al., 2006). Все эти соображения предполагают, что необходимы дальнейшие исследования, чтобы лучше охарактеризовать LAB, продуцирующую CLA, и продукты их метаболизма.
LAB биосинтез селенопротеинов
Способность производить ферменты, фиксирующие металлы, является еще одной важной особенностью LAB, которая делает их интересными комменсальными организмами. Фактически, несколько видов Lactobacillus способны внутриклеточно фиксировать селенит натрия (Рисунок 5) в селеноцистеины и селенометионины, обеспечивая тем самым более биодоступную форму этого металла, который обычно плохо адсорбируется человеческими клетками в неорганической форме (Calomme et al. al., 1995).
Рис. 5. Гранулы неорганического селена, отображаемые с помощью ПЭМ на поверхности фиксирующего селен штамма Lactobacillus reuteri .
Селеноцистеин определяется как 21-я аминокислота, кодируется кодоном UGA и обладает собственной тРНК, которая использует серин в качестве промежуточного звена (т-РНК SerSec). Селенометионин неспецифично включается в белки вместо метионина, и он также важен для реакций транс-селенации.Было продемонстрировано, что некоторые бактериальные ферменты содержат селеноцистеины в своем активном центре, и все они относятся к классу оксидоредуктаз: среди них глутатионпероксидаза, фермент, имеющий решающее значение для контроля окислительного стресса и всех связанных заболеваний как в бактериях, так и в эукариотических клетках. Lactobacillus reuteri , например, экспрессирует селеноцистеинлиазу (Lamberti et al., 2011a, b), PLP-зависимый фермент, играющий ключевую роль в биосинтезе новых селенопротеинов, поставляющий оптимальный субстрат (H 2 Se ) до селенфосфатсинтетазы, которые катализируют выработку «активированной» формы селена, полезной для селеноцистеинсинтетазы для производства L-селеноцистеинил-тРНК (Lacourciere and Stadtman, 1998).LAB также может действовать как хелатирующие агенты для других важных для нутрицевтики металлов на уровне толстой кишки. Исследования, проведенные на различных прокариотах, привели к характеристике многих механизмов «канализации» ионов цинка (Znll), включая цинк-связывающие, импортирующие цинк белки и системы экспорта цинка (Blencowe and Morby, 2003). Цинксодержащие и содержащие цинк LAB могут, таким образом, действовать как иммуномодуляторы, полезные для борьбы с вирусным гастроэнтеритом (Salvatore et al., 2007).
LAB Экология
Вклад LAB в общую экосистему кишечника был выяснен только недавно.Традиционные методы культивирования не смогли пролить свет на фактические микробные популяции, обитающие в кишечнике взрослых людей, но применение независимых от культуры методов, таких как ПЦР в сочетании с DGGE (денатурирующий градиентный гель-электрофорез), привело к более детальному изучению того, что происходит на самом деле. на уровне кишечника. Прежде всего, важно различать резидентные и транзиторные бактерии: Vaughan и соавт. (2002) сообщили о списке из видов Lactobacillus, , постоянно колонизирующих кишечник: L.acidophilus , L. brevis , L. casei , L. crispatus , L. delbrueckii , L. fermentum , L. fructivorans , L. gasseri , L. paracasei , L. paracasei , L. plantarum , L. rhamnosus , L. ruminis , L. sakei , L. salivarius , L. vaginalis . Стоит отметить, что LAB составляют 0,01–1,8% от всего бактериального сообщества, живущего в кишечнике человека (Louis et al., 2007).
Взаимодействие ЛАБ с кишечной микробиотой и антибактериальный потенциал ЛАБ
Как антагонизм, так и сотрудничество способствуют взаимным отношениям между LAB и другими кишечными микроорганизмами. Формирование биопленок, генетический обмен, опосредованный фагами и феромонами, и синтрофии являются хорошо отработанными стратегиями взаимодействия с другими кишечными микроорганизмами. Тем не менее антагонистическое поведение превалирует.
Пищевая, ферментативная и метаболическая конкуренция
Ферменты, нацеленные на клеточную стенку, такие как N-ацетилмурамидаза и N-ацетилмурамоил-L-аланинамидаза, являются белками, участвующими в обновлении пептидогликана, действующими как общие факторы лизиса клеточной стенки (Salazar and Asenjo, 2007).Их часто производят для борьбы с популяциями бактерий, занимающих одну и ту же экологическую нишу. Точно так же хитиназа, белок, гидролизующий важный компонент клеточных стенок дрожжей и грибов, то есть хитин, был обнаружен в экзопротеоме LAB (Genovese, неопубликованные результаты).
Тем не менее, конкуренция питательных веществ является наиболее известным как специфическое вмешательство, общее для всех микробных популяций, созданное LAB. Поскольку LAB являются преимущественно сахаролитическими, но также и утилизаторами аминокислот, они могут вычитать питательные вещества как для сахаролитических, так и для протеолитических видов, обитающих в кишечной среде.Кроме того, их специфический ферментативный метаболизм, генерирующий кислоты, представляет собой особую метаболическую конкуренцию за борьбу с другими эндогенными или экзогенными видами микробов вместе с диоксидом углерода, этанолом, перекисью водорода (продуцируемой и быстро устраняемой активностью НАДН-оксидазы). Прямое противомикробное действие органических кислот (молочная, уксусная и пропионовая кислоты) обусловлено их вмешательством в мембранный потенциал цитоплазмы бактерий и ингибированием активного транспорта, в то время как диоксид углерода (образующийся во время гетеролактической ферментации LAB) и перекись водорода предотвращают рост некоторых бактерии, создавая неблагоприятную среду.
Стоит отметить, что упомянутая протеолитическая активность LAB в отношении казеинов также может приводить к появлению антимикробных пептидов, таких как k-казецидин, пентапептид (Phe-Phe-Ser-Asp-Lys), полученный путем гидролиза k-казеина, и исрацидин, олигопептид произошел из казеина альфа s1. Обе молекулы оказались очень активными в отношении Staphylococus aureus , причем последний также стимулирует фагоцитоз. Искусственное удаление C-конца Lys и N-конца Phe из k-казецидина сильно снижает исходную активность природного пентапептида (Matin and Otani, 2002). L.acidophylus, L.plantarum, L.helveticus, L.rhamnosus и Lactococcus lactis также продуцируют другие пептиды, производные от казеина альфа s1, которые имеют совершенно другую аминокислотную последовательность. Последний оказался активным против Enterobacter sakazakii , грамотрицательного вида, вызывающего инфекции у новорожденных (Hayes et al., 2006).
Производство бактериоцинов
Многообещающей особенностью LAB является производство интерференционных молекул, бактериоцинов (Montalbán-López et al., 2011). Бактериоцины — это белковые молекулы, которые синтезируются на уровне рибосом (а не как вторичные метаболиты), которые могут мешать росту большинства бактерий. Они обладают бактерицидным действием и избирательны по отношению к прокариотам. Бактериоцины продуцируются в конце экспоненциальной фазы роста, и их спектр действия может варьироваться в зависимости от вида-продуцента. Штаммы-продуценты невосприимчивы к своим собственным бактериоцинам, потому что они обладают генами, которые кодируют механизмы иммунитета, которые позволяют различать «я» и «чужое».Защита может быть обеспечена специальным иммунным белком и / или специальной системой ABC-транспортера, которая перекачивает бактериоцин за пределы мембраны продуцента (Draper et al., 2009).
LAB особенно продуктивны в производстве бактериоцинов и могут биосинтезировать различные типы антагонистических молекул. Лантибиотики, названные так потому, что они содержат посттрансляционно модифицированные аминокислоты, такие как лантионин (два аланина, связанные серой), β-метил-лантионин, дегидроаланин и дегидробутирин, представляют собой небольшие пептиды (длиной 19–38 аминокислот), в основном активные против грамположительных бактерий (рис. 6).
Рис. 6. Общая структура лантибиотика: ковалентно связанные аланины и посттрансляционно модифицированные аминокислоты являются отличительными чертами этих антимикробных пептидов .
Их можно разделить на два подкласса в зависимости от их структуры и механизма действия: удлиненные амфифильные катионные лантибиотики (низин, лактококцин и педиоцин) активны за счет образования пор, что приводит к диссипации мембранного потенциала и оттоку жидкости. малых метаболитов из клеток-мишеней.Напротив, глобулярные лантибиотики (например, мерсадицин) действуют посредством ингибирования ферментов, препятствуя биосинтезу клеточной стенки. Бактериоцины класса II представляют собой небольшие (<10 кДа) термостойкие пептиды, не претерпевающие обширных посттрансляционных модификаций, хотя они могут содержать D-аминокислоты. Самый известный бактериоцин класса II, так называемый педиоцин-подобный, обладает узкой, но очень специфической активностью против пищевого патогена Listeria monocytogenes (Kazazic et al., 2002).Наконец, бактериолизины представляют собой большие термолабильные антимикробные белки, вызывающие лизис чувствительных клеток, катализируя гидролиз клеточной стенки. Их структура содержит каталитический N-концевой домен, который проявляет гомологию с эндопептидазами, и С-конец, который представляет сайт узнавания-мишени (Lai et al., 2002).
Все эти природные антимикробные соединения являются наиболее интересными конечными продуктами метаболизма антагонистов, синтезируемыми LAB. Исследования последних лет были сосредоточены на изучении их изоляции, структуры и механизма действия (Millette et al., 2007).
Взаимодействие лаборатории с человеком-хозяином
Колонизация LAB и диета
LAB, особенно Lactobacilli, колонизируют новорожденного, как только он находится на грудном вскармливании. Напротив, новорожденные на искусственном вскармливании обычно являются хозяевами различных видов бактерий, таких как Clostridia и Bacteroides. Присутствие LAB, которые в основном являются продуцентами сахаров, ацидогенов и бактериоцинов, действует как селективный агент, защищающий кишечную среду от гнилостных и сульфатредуцирующих бактерий, которые, как известно, участвуют в производстве газа, тиолов, фенолов, индола, гистолезивных протеаз. и канцерогены (Holzapfel et al., 1998). В то время как ферментация сахара в основном происходит в проксимальном отделе толстой кишки (pH 5–6), гниение белков и пептидов типично для дистального отдела толстой кишки (pH 7). Присутствие LAB максимально в течение срока и оказывается абсолютно полезным для здоровья и благополучия хозяина, но для поддержания необходим особый режим питания.
На самом деле хорошо известно, что диета играет решающую роль в выборе микробных популяций в сложной и динамичной экосистеме кишечных бактерий человека.Пища, содержащая LAB, такая как йогурт и сыр, может обогатить популяции кишечных LAB, поскольку эти бактерии могут легко выжить, попав в желудочный транзит. Фактически, LAB являются естественными ацидофильными организмами (оптимум роста pH 3,5–6,5), и они могут дополнительно улучшить свою устойчивость к кислому pH, установив стратегии, которые производят щелочные метаболиты (упомянутый путь ADI и пути декарбоксилирования аминокислот). Кроме того, поскольку человеческое или искусственное молоко может выделять LAB или Clostridia у новорожденных, аналогично, диета, богатая «мясным жиром», способствует выделению и увеличению Clostridia у взрослых, в то время как диета с неметаболизируемым сахаром (особенно фруктозой, инулином) — и питательные вещества, богатые целлюлозой, которые не попадают в организм человека при регулярном переваривании) в пользу LAB.Эпидемиологическая связь между диетой, богатой «мясом и жиром», и раком толстой кишки (Hill, 1975) частично может быть объяснена способностью Clostridia метаболизировать желчные кислоты до канцерогенных молекул, ответственных за неопластическую трансформацию слизистой оболочки кишечника (Murray et al., 1980). Таким образом, роль LAB не только ограничивается предотвращением колонизации экзогенными бактериями, но также и борьбой с токсинами и мутагенными молекулами эндогенного происхождения.
ЛАБОРАТОРНАЯ ведущая беседа и стресс-шапероны
Недавно было установлено, что «симбиоз» может запускать адаптивные реакции как у бактерий, так и у хозяина, которые выявляются с помощью транскриптомных и протеомных исследований (взаимодействие хозяина и микробиоты) и касаются активации конкретных метаболических путей, стимуляции / депрессии иммунной системы и стресса. ответы.
Со стороны хозяина активация важных генов, участвующих в клеточных сигнальных путях и иммуномодуляции, наблюдалась в эксперименте in vitro с использованием эпителиальных клеток CaCo-2, подвергшихся воздействию Lactobacillus acidophilus , естественного обитателя тонкой кишки человека. Использование других LAB пищевого происхождения не может вызвать такой же биологический ответ, демонстрируя тем самым, что длительный симбиоз выбрал специфические генетические признаки для взаимодействия хозяин-микроб (O’Flaherty and Klaenhammer, 2011).Кроме того, важность гликома клеточной стенки LAB в индукции цитокинов в эпителиальных клетках кишечника была доказана с использованием нокаута мутантов по способности гликозилирования белков. Эти мутанты не только неспособны стимулировать цитокины, но они также неспособны прикрепляться к клеткам CaCo 2 и обнаруживают значительное нарушение устойчивости в среде кишечника мышей (Tytgat et al., 2011).
С бактериальной стороны описаны адаптационные реакции, способствующие сохранению устойчивости организма-хозяина.В LAB генетические локусы, специфически индуцируемые транзитом через кишечник, участвуют в пищевой адаптации (гидролазы, переносчики ABC и системы PTS), а также в антистрессовых механизмах, таких как модификация мембран, гликозилирование поверхности и антиоксидантная защита (Lebeer et al. , 2011). Также была установлена продукция бактериального стрессового белка (усиленный синтез шаперонов GroEL, GroES, DnaK, Hrc, Clp, Cts) в ответ на факторы хозяина (Henderson et al., 2006). В настоящее время хорошо продемонстрировано, что, помимо помощи в правильной укладке белков, шапероны могут выполнять множество функций, таких как предотвращение денатурации, перегруппировка белков после окислительного и pH-стресса и выполнение целевого протеолиза необратимых денатурированных белков.Среди этих функций — адгезия к белкам внеклеточного матрикса тканей хозяина и стимуляция иммунной системы хозяина. Для выполнения этих дополнительных функций шапероны должны располагаться на поверхности или вне клетки.
Это явление известно как подработка, и к настоящему времени описано несколько подрабатывающих белков, как у эукарий, так и у бактерий, включая LAB. В их состав входят не только шапероны стресса (GroEL, DNAk), но также гликолитические ферменты и ферменты цикла TCA. Белки лунного света переносятся внеклеточно без какого-либо экспортного сигнала и без специфического мотива прикрепления к клеточной стенке или мембране: таким образом, поверхностная или внеклеточная локализация может быть связана с pH окружающей среды.Antikainen с соавторами (Antikainen et al., 2007a) продемонстрировали, что более высокий внеклеточный pH может вызывать высвобождение слабо связанных белков в Lactobacillus crispatus , но также может быть вовлечено обновление клеточной стенки, которое часто происходит во время фазы экспоненциального роста. в отрыве таких белков (Sánchez et al., 2008). Обычно открытые на поверхности и высвобождаемые подрабатывающие белки выполняют различные биологические функции, а именно адгезию (связывание с клеточной стенкой) и модуляцию иммунной системы (внеклеточные шапероны).
Адгезия LAB к тканям хозяина и иммуномодуляция
Адгезия. Взаимодействие бактерий и слизистой оболочки человека представляет собой динамическое равновесие, и несколько факторов могут изменить это равновесие. Кишечные движения и бактериальный хемотаксис вызывают слабую связь между бактериями и поверхностью эпителия кишечника, где преобладают электростатические и гидрофобные связи. На следующем этапе происходит более специфическое взаимодействие между молекулярными детерминантами на бактериальной поверхности и специфическими рецепторами слизистой оболочки кишечника.Конкретными мишенями для тканей человека являются коллаген I и IV типов, фибронектин и ламинин. В LAB хорошими эффекторами связывания являются PS, липотейхоевые кислоты, а также белки, такие как лектины и адгезины. Поэтому потенциал адгезии определенного штамма LAB имеет решающее значение для выживания и сохранения в кишечной среде.
Некоторые белки, расположенные на поверхности, могут действовать как адгезины. И шаперон GroEL, и фактор удлинения, EF-Tu, проявляют адгезионную способность по отношению к тканям человека у Lactobacillus johnsonii .EF-Tu представляет собой связывающий гуанозиновый нуклеотид белок, играющий центральную роль в синтезе белка (при экспрессии внутри клетки), но способный связывать фибронектин или муцины и эпителиальные клетки при воздействии на поверхность клетки (Granato et al., 2004). Триггерный фактор также связан с адгезией: было описано, что он экспонируется на поверхности Lactobacillus plantarum , а в L. reuteri NCIB11951, связывающий коллаген I белок, имеет высокую гомологию последовательности с E.триггерный фактор coli (Aleljung et al., 1994). Помимо этих белков, гликолитические ферменты также могут обладать адгезивными свойствами: GAPDH способен связывать фибронектин, плазмин и муцин у нескольких видов бактерий (Alvarez et al., 2003), и аналогичную роль может выполнять PGK (фосфоглицераткиназа), которая относится к внеклеточной локализации в Lactobacillus rhamnosus GG (Sánchez et al., 2008). Некоторые из этих белков будут рассмотрены в отдельной главе из-за их особого отношения к поведению также как рецепторы плазминогена.
Модуляция иммунной системы. Около 70% иммунной системы локализовано в желудочно-кишечном тракте как GALT (лимфоидная ткань, ассоциированная с кишечником). Между LAB и иммунной системой существует взаимная связь. С одной стороны, иммунная система выбирает виды LAB, которые будут приняты, с другой стороны, LAB может модулировать иммунологические функции. Наиболее очевидные эффекты LAB касаются увеличения соотношения между противовоспалительными (IL-10, β-TGF) и провоспалительными (IL-1 beta, IL-3, IL-4) цитокинами (Pessi et al., 2000) и отбор популяций Т-лимфоцитов (Karimi et al., 2009). Различные компоненты LAB, такие как тейхоевая и липотейхоевая кислоты, липопротеины и ЭПС, могут быть индукторами иммунного ответа (Weidenmaier and Peschel, 2008). Некоторые из упомянутых адгезивных белков также проявляют иммуномодулирующие свойства, когда они секретируются. Это особенно верно для шаперона GroEL, который, как было продемонстрировано, взаимодействует с макрофагами и стимулирует секрецию цитокинов. В Л.johnsonii GroEL также вызывает сильную агрегацию патогена Helicobacter pylori , способствуя снижению бактериальной нагрузки и, таким образом, облегчая удаление скопившихся патогенов слизью (Granato et al., 2004). У L. reuteri также была продемонстрирована строгая связь между адгезией и индукцией регуляторных Т-клеток (Smits et al., 2005). И последнее, но не менее важное: непрямая иммуностимулирующая активность может быть вызвана протеолитическими видами кишечных LAB через производство иммуномодулирующих пептидов из казеина.Эти молекулы в первую очередь усиливают пролиферацию лимфоцитов и фагоцитоз макрофагов, но они, вероятно, важны для регулирования развития иммунной системы у новорожденных, а также могут способствовать ослаблению аллергической реакции и развитию опухолей (Korhonen and Pihlanto, 2006).
Взаимодействие LAB с системой плазминоген-плазмин
Lactobacillus crispatus , Lactobacillus acidophilus, L. amylovorus, L. gallinarum, L. gasseri и Lactobacillus johnsonii проявляют способность связывать плазминоген на своей клеточной стенке (Hurmalainen et al., 2007). Было продемонстрировано, что несколько гликолитических ферментов, участвующих в адгезии (энолаза, GAPDH, фосфоглицераткиназа и фосфоглицератмутаза), гидролаза солей желчных кислот и стрессовый белок DnaK, действуют как рецепторы плазминогена (Pg). Все они имеют общую особенность — лизин на С-конце. Некоторые сообщения предполагают, что способность связывания плазминогена в LAB может представлять риск, потому что плазминоген может быть активирован до плазмина, тем самым запуская дальнейшие протеолитические каскады, приводящие к деградации внеклеточного матрикса кишечника.Тем не менее, LAB не обладают внутренним потенциалом активации плазминогена, типичным для патогенных видов. На самом деле патогенные бактерии, такие как Staphylococcus aureus и Streptococcus pneumoniae , могут превращать плазминоген в плазмин с помощью специфических факторов вирулентности (стафилокиназы и стрептокиназы соответственно) (Bergmann et al., 2003), которые до сих пор никогда не были обнаружены в геноме LAB. По мнению некоторых авторов (Antikainen et al., 2007b), LAB может использовать эндогенную систему хозяина (урокиназу и тканевые активаторы плазминогена) для запуска протеолиза, по мнению других (Sánchez et al., 2008). LAB может тушить плазминоген, вычитая его из патогенных бактерий. В настоящее время это все еще спорный вопрос, касающийся LAB, и необходимы дополнительные исследования, чтобы полностью пролить свет на этот аспект взаимодействия бактерии-хозяина.
LAB Доказанные полезные действия для здоровья человека
Наиболее важные полезные взаимодействия между кишечными LAB и людьми включают регуляцию метаболизма, инфекционный контроль и модуляцию воспаления / аллергии. Последние три эффекта так или иначе связаны с регуляторным действием иммунной системы со стороны LAB.
Метаболизм
Энергия / добыча полезных ископаемых
LAB может улучшить восстановление энергии из питательных веществ за счет разложения неметаболизируемых сахаров и снабжения хозяина собственной β-галактозидазой, тем самым минуя ферментативную непереносимость лактозы. Некоторые фосфопептиды казеина (CPP) демонстрируют способность удерживать в растворе ионы кальция даже при щелочном pH. Это приводит к увеличению абсорбции кальция, полезного при остеопорозе, но также к усиленному всасыванию железа, цинка и марганца, полезных в качестве кофакторов ферментов или простетических групп.Для выполнения этой функции необходимо, чтобы LAB переваривала казеин, особенно казеины альфа s1 и альфа s2, высвобождая CPP, фосфорилированный по остаткам Ser и Thr. Поскольку бета- и k-казеины бедны гидроксилированными аминокислотами, они менее подходят для фосфорилирования, а полученные из них пептиды менее активны в связывании металлов (Meisel, 1998).
Снижает уровень холестерина
При попадании в кишечник LAB SCFA может вызывать снижение синтеза холестерина в печени и перераспределение холестерина из крови в печень (Pereira and Gibson, 2002).Также деконъюгация желчных кислот с помощью LAB может играть аналогичную роль в снижении уровня холестерина в плазме. Деконъюгированные желчные кислоты плохо всасываются слизистой оболочкой кишечника и, следовательно, выводятся из организма. Как следствие, новый холестерин перемещается из крови в печень для синтеза de novo желчных кислот (St-Onge et al., 2000). В Lactobacillus reuteri биосинтезируется специфическая холоилглицингидролаза, катализирующая начальную реакцию шлюза для деконъюгации желчных кислот (Martoni et al., 2008).Способность снижать уровень холестерина также была продемонстрирована с помощью LAB-продуцируемых ЭПС (Pigeon et al., 2002). Таким образом, ожидается положительный эффект при сердечно-сосудистых патологиях и сердечных заболеваниях.
Инфекция и иммуномодуляция
LAB давно известны как способные контролировать инфекции из-за их прямого антибактериального действия (опосредованного бактериоцинами и продуцированием кислоты) и их непрямого (иммунологически опосредованного) действия против вирусов. Роль Lactobacilli в предотвращении диареи путешественников, росте Helicobacter pylori и токсина, продуцирующего E.coli была продемонстрирована в исследованиях in vitro и in vivo . Lactobacillus rhamnosus GG также успешно применялся для профилактики колита C. difficile , а также для лечения атопической экземы посредством модуляции интерлейкина-10 (Pessi et al., 2000), а Lactobacillus reuteri может контролировать IgE-опосредованные аллергии, действующие на уровне регуляторных Т-клеток (Karimi et al., 2009). Сообщалось также, что противовоспалительная (Kitazawa et al., 1998) и противоопухолевая активность обусловлены LAB-продуцируемыми EPS.Rhamnose Hetero-EPS оказался эффективным против язвы желудка (Badel et al., 2011), и была продемонстрирована стимуляция TNF (DeVuyst et al., 2007). Эксперименты с использованием коктейлей из Lactobacillus и Bifidobacterium также выявили способность вызывать апоптоз карцином толстой кишки.
Информационные молекулы и окислительный стресс
Очень передовое свидетельство — возможность того, что LAB может контролировать как антиоксидантную защиту человека, так и производство информационных молекул, влияющих на настроение, кровяное давление и, в более общем плане, на ось кишечник-мозг.
Lactobacillus helveticus , Lactobacillus delbrueckii subsp. bulgaricus SS1 и Lactococcus lactis subsp. cremoris FT4 может модулировать кровяное давление, производя пептиды, ингибирующие ангиотензин-1-конвертирующий фермент (ингибиторы АПФ), из белков молока. Наиболее активными молекулами являются трипептиды, состоящие из Val-Pro-Pro и Ile-Pro-Pro, но и другие антигипертензивные пептиды на основе казеина, такие как альфа s1, казокинин-5 (Phe-Phe-Val-Ala-Pro), альфа s1. казокинин-6 (Thre-Thre-Met-Pro-Leu-Trp), бетаказокинин (Lys-Val-Leu-Pro-Val) и дипептид Tyr-Pro также оказались эффективными для контроля артериального давления (Yamamoto et al. al., 1999). Поскольку их продукция является результатом протеолитической активности некоторых штаммов LAB по сравнению с казеином, она зависит от доступности казеина в кишечнике и от протеолитического потенциала каждого отдельного штамма. Обычно пептидазы LAB за счет укорачивания поли / олигопептидной цепи вносят вклад в усиление потенциала против АПФ. Интересно, что конечные активные короткие пептиды устойчивы как к изменениям pH, так и к пищеварительным ферментам человека (Sipola et al., 2002; Gobbetti et al., 2004). Антитромботические пептиды, полученные из k-казеингликопептидов человека и крупного рогатого скота, также были обнаружены у пятидневных новорожденных после грудного вскармливания и кормления смесью соответственно (Chabance et al., 1995). Даже если эти молекулы могут образовываться в результате протеолиза человека в пище, протеолитическая активность LAB в отношении казеина наверняка может способствовать их высвобождению in vivo . Эти молекулы, называемые казоплателином и пептидом, ингибирующим тромбин, представляют собой олигопептиды длиной от 6 до 11 аминокислот, предотвращающие агрегацию активированных АДФ тромбоцитов и связывание человеческого фибриногена (лямбда-цепь) с рецептором на поверхности тромбоцитов. Более короткая молекула, казопиастрин, также проявляла ингибирующую активность в отношении связывания фибриногена (Jollès et al., 1986).
Наиболее интересными молекулами, продуцируемыми с помощью протеазной системы LAB в молочных продуктах, являются опиоидные пептиды (экзорфины) и пептиды-антагонисты опиоидов (казоксины). В центральной и периферической нервной системе млекопитающих есть рецепторы (K, дельта и мю) для эндогенно продуцируемых опиоидных пептидов (эндорфинов), которые обладают ингибирующей активностью в отношении фермента аденилатциклазы. Фрагменты белка с опиоидоподобной активностью были обнаружены в результате воздействия как pH / протеаз пищеварения человека, так и микробных протеаз на молекулы казеина.Эти пептиды состоят из 4–10 аминокислот и могут вызывать миорелаксантный эффект, воздействуя на рецепторы мю (бета-казоморфины) или дельта-рецепторы (пептид, производный от казеина альфа s1). N-концевой остаток, который имеет решающее значение для запуска биологической активности, представляет собой Tyr в бета-казоморфинах и Arg в пептиде, производном от альфа s1 казеина (Loukas et al., 1983). Тем не менее, две молекулы демонстрируют разный период полураспада из-за их разной структуры, и наличие нескольких остатков пролина (шесть) в бета-казоморфинах, по-видимому, является фактором, повышающим устойчивость пептидов к ферментативному перевариванию в пищеварительном тракте человека.Здесь они могут влиять на время прохождения через кишечник, водный баланс и усвоение аминокислот. После всасывания в кровь они достигают мозга и периферических рецепторов, где оказывают расслабляющее действие, вызывая сон и спокойствие (Chabance et al., 1998). Противоположным этому действию является антиоптиоидная активность, проявляемая пептидами, полученными в результате гидролиза k-казеина. Эти молекулы, называемые казоксинами, действуют как антагонисты оппиоидов над рецепторами как мю, так и k-типа. Их возможное физиологическое действие на человека заключается в противодействии опасному для жизни угнетению ЦНС и дыхательной системы (Chiba and Yoshikawa, 1986).Наконец, некоторые пептиды, полученные из казеина с помощью протеолизов LAB, могут выполнять смешанные функции, такие как морфиноподобные и иммуностимулирующие (Kayser and Meisel, 1996).
Некоторые виды Lactobacillus (Cho et al., 2007) и Lactococcus (Mazzoli et al., 2010) также способны продуцировать путем декарбоксилирования глутамата (см. Параграф об энергетическом метаболизме) ГАМК, наиболее распространенный нейротрансмиттер. в центральной нервной системе позвоночных, которая также действует как модуляторный эффектор на уровне кишечника.Хотя ГАМК не может обойти гематоэнцефалический барьер, она может действовать как расслабляющая молекула над гладкими мышцами кишечника, может снижать артериальное давление у пациентов с легкой гипертонией (Inoue et al., 2003), а также играет регулирующую и трофическую роль в поджелудочная железа (Erlander et al., 1991). Даже если не все упомянутые LAB являются стабильными обитателями кишечника человека, они могут быть приобретены с пищей и могут сохраняться в подходящих условиях.
Что касается антиоксидантного потенциала, о котором говорилось в предыдущем абзаце, некоторые штаммы LAB способны фиксировать неорганический селенит в селеноаминокислоты, что открывает возможность биосинтеза in vivo селенопротеинов , необходимых факторов для контроля окислительного стресса.Было продемонстрировано, что селен является важным микроэлементом с точки зрения питания с 1957 года (Schwarz and Foltz, 1957). Однако потребление селена с пищей широко варьируется в разных странах: от 7 мкг в день в Китае до 100–200 мкг в Канаде, США и Венесуэле (Rayman, 2008). Причина таких различий кроется в разных почвах, на которых выращиваются овощи и животные. Теперь очевидно, что содержание селена в пище напрямую связано с доступностью селена в почве, и там, где эта доступность недостаточна, возникает острый эндемический дефицит селена (Zhao et al., 2007). Кроме того, концентрации селена в сыворотке крови отражают содержание селена в пище, тем самым демонстрируя, что альтернативный эндогенный способ получения этого металла невозможен (Combs, 2001).
Даже если общепризнано, что дефицит селена (за исключением очень специфических и редких синдромов) сам по себе не вызывает болезни, тем не менее, этот металл, по-видимому, является ключевым питательным веществом для подсчета бактериальных и вирусных инфекций (Beck et al., 2003 ), для баланса гормонов щитовидной железы (Tinggi, 2008), для ограничения атерогенеза, связанного с окисленными ЛПНП, и последующего сердечного заболевания (Furman et al., 2004), а также для контроля возрастных расстройств, таких как окислительный статус, иммунодефицит, общее воспаление (Méplan, 2011) и рак простаты (Klein et al., 2003). Многие данные свидетельствуют о противораковом эффекте добавок селена, поскольку этот металл оказался эффективным инструментом для контроля риска повреждения ДНК, связанного с неопластической дегенерацией. Также наблюдалась обратная корреляция между уровнем селена в крови и смертностью от рака.
Рекомендуемое потребление селена с пищей для людей составляет 55 мкг в день (Alzate et al., 2008). Биосинтезированные белки селена Lactobacillus не только представляют собой более биодоступный диетический источник, но также существует корреляция между этой органической формой и концентрацией этого металла во внешней среде, что открывает интересные перспективы выделения металлов в кишечную среду с помощью LAB. последующий контроль окислительного стресса.
LAB Свет и тени
LAB, как и любой организм на земле, несовершенен. Описанные метаболические и биохимические особенности подчеркивают риск того, что они могут продуцировать токсичные метаболиты, такие как аммиак и биоактивные нежелательные амины, такие как гистамин и тирамин.Их способность продуцировать бутират может быть серьезно оценена при наличии метаболического синдрома, поскольку этот метаболит, хотя и обладает защитным действием против неопластической дегенерации, может увеличивать прирост калорий у человека, а также ингибировать липолитические процессы. Точно так же некоторые изомеры CLA, продуцируемые LAB, могут быть вредными, поэтому тщательное типирование отдельных штаммов кишечных LAB и их метаболитов может помочь избежать нежелательных эффектов. Что касается способности активировать плазмин-опосредованные протеолитические каскады с использованием эндогенных факторов, этот риск уравновешивается возможным подавляющим эффектом высвобождаемых рецепторов палсминогена на сам плазминоген.
Отдельный комментарий необходим для рода Enterococcus . В то время как некоторые виды, такие как E. faecium , в настоящее время считаются безопасными и даже используются в качестве заквасок в пищевой промышленности и в качестве пробиотиков как в пищевых, так и в фармацевтических препаратах, другие виды, такие как E. faecalis , могут вызывать проблемы по разным причинам. Следует помнить, что энтерококки более толерантны к соли, менее ацидофильны и менее ацидогены, чем другие LAB, таким образом, составляя группу per se .Во-первых, они от природы устойчивы ко многим противомикробным препаратам, а также несут в себе факторы трансмиссивной устойчивости к антибиотикам. Кроме того, их продукция феромонов очень высока, что увеличивает частоту рекомбинации. Второй риск связан с тем фактом, что большинство признаков патогенности, выражаемых энтерококками, не являются видами, а скорее связаны с штаммами. Среди них стоит упомянуть желатиназу, сериновые протеазы, антиген эндокардита и фактор агрегации. Еще одна проблема связана с окислительным катаболизмом стероидных молекул.Во время энтерогепатической циркуляции желчные кислоты биотрансформируются кишечной микробиотой в различные метаболиты, некоторые из которых являются канцерогенными, особенно C7-дегидрокси- и C7-кетопроизводные (Murray et al., 1980). Хотя дегидрирование, эпимеризация и дегидроксилирование в основном осуществляются Clostridia , также сообщалось, что некоторые штаммы Enterococcus faecalis обладают дегидроксилирующей активностью по C7. Эти реакции направлены не только на желчные кислоты или холестерин, но и на различные молекулы стероидов, присутствующие в кишечнике (Groh et al., 1993).
Всегда трудно установить раз и навсегда, когда польза превышает риск, особенно когда речь идет о бактериях, частота мутаций, генетическая рекомбинация и эволюция которых могут изменить сценарий за несколько лет. Всего несколько десятилетий назад E. coli считались безопасными, а Clostridia — высокопатогенными. В настоящее время доказано, что около штаммов E. coli вызывают смерть, а Clostridia даже были предложены в качестве пробиотиков (Cartman, 2011).Несомненно, «омический» подход может помочь нам сейчас и в ближайшем будущем пролить свет на некоторые спорные аспекты, все еще касающиеся биологии лабораторных лабораторных исследований, и на то, как условия окружающей среды могут влиять на их биологию. Тем не менее, учитывая отсутствие основных признаков бактериальной патогенности (токсины, потенциал инвазии) и всех полезных свойств, предоставляемых LAB в контексте экосистемы кишечника (закисление, пребиотический биосинтез FOS, восстановление лактозы, высвобождение бактериоцинов, положительные биоактивные молекулы производство, действующее на настроение и аппетит, модуляцию иммунной системы и организацию селена), мы можем сделать вывод, что эта группа бактерий не только хорошо приспособлена к жизни, соответствующей человеку, но также может помочь человеку-хозяину поддерживать его / ее состояние здоровья. и, возможно, для повышения его / ее работоспособности и долголетия.
Заявление о конфликте интересов
Автор заявляет, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Особая благодарность доктору Роберто Маццоли за полезные обсуждения и критическое чтение рукописи и доктору Алессандро Пессионе за управление фигурой.
Список литературы
Алельджунг, П., Шен, В., Розальска, Б., Hellman, U., Ljungh, A., and Wadström, T. (1994). Очистка коллагенсвязывающих белков Lactobacillus reuteri NCIB 1. Curr. Микробиол . 28, 231–236.
Pubmed Аннотация | Pubmed Полный текст
Альварес, Р. А., Блейлок, М. У., и Бейсман, Дж. Б. (2003). Локализованная на поверхности глицеральдегид-3-фосфатдегидрогеназа Mycoplasma genitalium связывает муцин. Мол. Микробиол . 48, 1417–1425.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Альзате, А., Фернандес-Фернандес, А., Перес-Конде, М. К., Гутьеррес, А. М., и Камара, К. (2008). Сравнение биотрансформации неорганического селена Lactobacillus и Saccharomyces в процессе молочной ферментации йогурта и кефира. J. Agric. Food Chem . 56, 8728–8736.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Антикайнен, Ю., Купаринен, В., Ляхтеенмяки, К., и Корхонен, Т. К. (2007a). pH-зависимая ассоциация енолазы и глицеральдегид-3-фосфатдегидрогеназы Lactobacillus crispatus с клеточной стенкой и липотейхоевой кислотой. Дж. Бактериол . 189, 4539–4543.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Antikainen, J., Kuparinen, V., Lahteenmaki, K., and Korhonen, T. K. (2007b). Энолазы грамположительных бактериальных патогенов и комменсальных лактобацилл имеют функциональное сходство по признакам, связанным с вирулентностью. FEMS Immunol. Med. Микробиол . 51, 526–534.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Бергманн, С., Вильд, Д., Дикманн, О., Франк, Р., Брахт, Д., Чхатвал, Г.С., и Хаммершмидт, С. (2003). Идентификация нового плазмин (оген) -связывающего мотива на поверхности показала альфа-енолазу Streptococcus pneumoniae . Мол. Микробиол . 49, 411–423.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Boosalis, M. S., Bandyopadhyay, R., Bresnick, E. H., Pace, B. S., Van DeMark, K., Zhang, B., Faller, D. V., and Perrine, S. P. (2001). Производные короткоцепочечных жирных кислот стимулируют пролиферацию клеток и вызывают активацию STAT-5. Кровь 97, 3259–3267.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Брюханов А. Л., Нетрусов А. И. (2007). Аэротолерантность строго анаэробных микроорганизмов и факторы защиты от окислительного стресса: обзор. Заявл. Biochem. Микробиол . 43, 567–582.
Pubmed Аннотация | Pubmed Полный текст
Шабанс, Б., Жоллес, П., Искьердо, К., Мазойе, Э., Франсуаль, К., Друэ, Л., и Фиат, А. М. (1995).Характеристика антитромботического пептида каппа-казеина в плазме новорожденных после приема молока. руб. Дж. Нутр . 73, 583–590.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Шабанс, Б., Марто, П., Рамбо, Ж. К., Мильоре-Самур, Д., Бойнар, М., Перротен, П., Гийе, Р., Жолл, П., и Фиат, А. М. (1998). Высвобождение казеинового пептида и его переход в кровь у людей при переваривании молока или йогурта. Biochimie 80, 155–165.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Чиба, Х. и Йошикава, М. (1986). «Биологически функциональные пептиды из пищевых белков», в Protein Tailoring for Food and Medical Use , ред. Р. Э. Фини и Дж. Р. Уитакер (Нью-Йорк, Нью-Йорк: Марсель Деккер), 123–153.
Чо, Ю. Р., Чанг, Дж. Ю., и Чанг, Х. С. (2007). Производство гамма-аминомасляной кислоты (ГАМК) с помощью Lactobacillus buchneri , выделенного из кимчи, и его нейропротекторное действие на нейрональные клетки. J. Microbiol. Биотехнология . 17, 104–109.
Pubmed Аннотация | Pubmed Полный текст
Де Ангелис, М., и Гоббетти, М. (1999). Lactobacillus sanfranciscensis CB1, марганец, кислород, супероксиддисмутаза и метаболизм. Заявл. Microbiol. Биотехнология . 51, 358–363.
Pubmed Аннотация | Pubmed Полный текст
Де Ангелис, М., Мариотти, Л., Росси, Дж., Сервили, М., Фокс, П. Ф., Роллан, Г., и Гоббетти, М. (2002). Катаболизм аргинина молочнокислыми бактериями закваски: очистка и характеристика ферментов пути аргининдеиминазы из Lactobacillus sanfranciscensis CB1. Заявл. Environ. Микробиол . 68, 6193–6201.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
ДеВуйст, Л., Де Вин, Ф., и Камерлинг, Дж. П. (2007). «Экзополисахиды из молочнокислых бактерий», в Comprehensive Glycoscience , Vol. 2, изд. Дж. П. Камерлинг (Оксфорд, Эльзевир), 477–518.
ДиБайз, Дж. К., Чжан, Х., Кроуэлл, М. Д., Краймальник-Браун, Р., Деккер, Г. А., и Риттманн, Б. Э. (2008). Микробиота кишечника и ее возможная связь с ожирением. Mayo Clin. Proc . 83, 460–469.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Дрейпер, Л. А., Грейнджер, К., Диган, Л. Х., Коттер, П. Д., Хилл, К. и Росс, Р. П. (2009). Перекрестный иммунитет и иммунная мимикрия как механизмы устойчивости к лантибиотику лактицину (3147). Мол. Микробиол . 71, 1043–1054.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Эрландер, М. Г., Тиллакаратне, Н. Дж., Фельдблюм, С., Патель, Н., и Тобин, А. Дж. (1991). Два гена кодируют разные глутаматдекарбоксилазы. Neuron 7, 91–100.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Еващук, Дж. Б., Уокер, Дж. У., Диас, Х., и Мэдсен, К. Л. (2006). Биопродукция конъюгированной линолевой кислоты пробиотическими бактериями происходит in vitro и in vivo у мышей. Дж. Нутрь . 136, 1483–1487.
Pubmed Аннотация | Pubmed Полный текст
Фурман, К., Rundlöf, A.K., Larigauderie, G., Jaye, M., Bricca, G., Copin, C., Kandoussi, A.M., Fruchart, J.C., Arnér, E.S, and Rouis, M. (2004). Тиоредоксинредуктаза 1 активируется в атеросклеротических бляшках: специфическая индукция промотора в макрофагах человека окисленными липопротеинами низкой плотности. Free Radic. Биол. Мед . 37, 71–85.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Гоббетти М., Минервини Ф. и Риццелло К. Г. (2004). Ангиотензин I превращает ферментно-ингибирующие и антимикробные биоактивные пептиды. Внутр. Дж. Дэйри Технол . 57, 173–188.
Гранато Д., Бергонцелли Г. Э., Придмор Р. Д., Марвин Л., Руве М. и Кортези-Теулаз И. Э. (2004). Связанный с клеточной поверхностью фактор удлинения Tu опосредует прикрепление Lactobacillus jonhsonii NCC533 (La1) к клеткам кишечника и муцинам человека. Заражение. Иммун . 72, 2160–2169.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Groh, H., Schade, K., and Hörhold-Schubert, C.(1993). Метаболизм стероидов с кишечными микроорганизмами. J. Basic Microbiol . 33, 59–72.
Pubmed Аннотация | Pubmed Полный текст
Хейс, М., Росс, Р. П., Фицджеральд, Г. Ф., Хилл, К. и Стэнтон, К. (2006). Полученные из казеина антимикробные пептиды, генерируемые Lactobacillus acidophilus DPC (6026). Заявл. Environ. Микробиол . 72, 2260–2264.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Эбер, Э. М., Рая, Р. Р., и де Джорджи, Г. С. (1999). Характеристика протеиназы клеточной оболочки из Lactobacillus helveticus . Biotechnol. Lett . 21, 831–834.
Хилл, М. Дж. (1975). Роль анаэробов толстой кишки в метаболизме желчных кислот и стероидов и их связь с раком толстой кишки. Рак 36 (Дополнение 6), 2387–2400.
Pubmed Аннотация | Pubmed Полный текст
Holzapfel, W.H., Haberer, P., Snel, J., Schillinger, U., and Huis in’t Veld, J.Х. (1998). Обзор кишечной флоры и пробиотиков. Внутр. J. Food Microbiol . 41, 85–101.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Хурмалайнен В., Эдельман С., Антикайнен Дж., Бауманн М., Лахтенмаки К. и Корхонен Т. К. (2007). Внеклеточные белки Lactobacillus crispatus усиливают активацию плазминогена человека. Микробиология 153, 1112–1122.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Якомино, Г., Текче, М. Ф., Гримальди, К., Тосто, М., и Руссо, Г. Л. (2001). Транскрипционный ответ линии клеток аденокарциномы толстой кишки человека на бутират натрия. Biochem. Биофиз. Res. Коммуна . 285, 1280–1289.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Инан, М. С., Расулпур, Р. Дж., Инь, Л., Хаббард, А. К., Розенберг, Д. В., и Джардина, К. (2000). Бутират короткоцепочечных жирных кислот просвета модулирует активность NF-kappaB в линии эпителиальных клеток толстой кишки человека. Гастроэнтерология 118, 724–734.
Pubmed Аннотация | Pubmed Полный текст
Иноуэ, К., Шираи, Т., Очиай, Х., Касао, М., Хаякава, К., Кимура, М., и Сансава, Х. (2003). Эффект снижения артериального давления нового кисломолочного продукта, содержащего гаммааминомасляную кислоту (ГАМК), при легкой гипертонии. Eur. J. Clin. Нутр . 57, 490–495.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Жоллес, П., Леви-Толедано, С., Фиат, А.М., Сория, К., Гиллессен, Д., Томайдис, А., Данн, Ф. У., и Кан, Дж. П. (1986). Аналогия между фибриногеном и казеином. Влияние ундекапептида, выделенного из каппа-казеина, на функцию тромбоцитов. Eur. J. Biochem . 158, 379–382.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Карими К., Инман М. Д., Биненшток Дж. И Форсайт П. (2009). Lactobacillus reuteri, индуцированные регуляторными Т-клетками, защищают от аллергической реакции дыхательных путей у мышей. Am. J. Respir. Крит. Care Med . 179, 186–193.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Катаяма Ю., Готтесман С., Памфри Дж., Рудикофф С., Кларк В. П. и Маурици М. Р. (1988). Двухкомпонентная АТФ-зависимая протеаза Clp Escherichia coli . Очистка, клонирование и мутационный анализ АТФ-связывающего компонента. J. Biol. Chem . 263, 15226–15236.
Pubmed Аннотация | Pubmed Полный текст
Казазич, М., Ниссен-Мейер, Дж., И Фимланд, Г. (2002). Мутационный анализ роли заряженных остатков в связывании с клеткой-мишенью, активности и специфичности педиоцин-подобного бактериоцина сакацина P. Microbiology 148, 2019–2027.
Pubmed Аннотация | Pubmed Полный текст
Китадзава, Х., Харата, Т., Уэмура, Дж., Сайто, Т., Канеко, Т., и Ито, Т. (1998). Потребность в фосфатных группах для митогенной активации лимфоцитов внеклеточным фосфополисахаридом из Lactobacillus delbrueckii ssp.bulgaricus . Внутр. J. Food Microbiol . 40, 169–175.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Кляйн, Э. А., Томпсон, И. М., Липпман, С. М., Гудман, П. Дж., Албейнс, Д., Тейлор, П. Р. и Колтман, К. (2003). ВЫБЕРИТЕ: исследование профилактики рака селеном и витамином Е. Урол. Онкол . 21, 59–65.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Корхонен, Х., Пихланто, А. (2006). Биоактивные пептиды: производство и функциональность. Внутр. Молочный J . 16, 945–960.
Кринос, К. М., Койн, М. Дж., Вейнахт, К. Г., Цианабос, А. О., Каспер, Д. Л., и Комсток, Л. Е. (2001). Обширное поверхностное разнообразие комменсальных микроорганизмов за счет множественных инверсий ДНК. Nature 414, 555–558.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Лай, А.С., Тран, С., и Симмондс, Р.С. (2002). Функциональная характеристика доменов, обнаруженных в литическом ферменте, продуцируемом Streptococcus equi subsp. zooepidemicus . FEMS Microbiol. Lett . 215, 133–138.
Pubmed Аннотация | Pubmed Полный текст
Ламберти, К., Манджапане, Э., Пессионе, А., Маццоли, Р., Джунта, К., и Пессионе, Э. (2011a). Протеомная характеристика пробиотика, метаболизирующего селен, Lactobacillus reuteri Lb2 BM для нутрицевтики. Proteomics 11, 2212–2221.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Ламберти, К., Пурротти, М., Маццоли, Р., Фаттори, П., Барелло, К., Коиссон, Дж. Д., Джунта, К., и Пессионе, Э. (2011b). Путь ADI и декарбоксилирование гистидина реципрокно регулируются в Lactobacillus hilgardii ISE 5211, протеомное свидетельство. Аминокислоты 41, 517–527.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Ло, Дж., И Хаандрикман, А. (1997). Протеолитические ферменты молочнокислых бактерий. Внутр. Молочный J . 7, 1–11.
Лебеер, С., Клас, Дж. Дж., Ванденплас, С., Верхувен, Т. Л. А., Титгат, Х., Шуфс, Г., Де Кеерсмэкер, С. К. Дж., И Вандерлейден, Дж. (2011). « In vivo технология эспрессии раскрывает новые идеи в кишечной адаптации Lactobacillus rhamnosus GG» в Тезисах на 10-м симпозиуме по молочнокислым бактериям , Эгмонд-ан-Зее, Нидерланды, 28 августа — 1 сентября 2011 , плакат D002.
Лю С., Причард Г. Г., Хардман М. Дж. И Пилоне Г.Дж. (1996). Катаболизм аргинина у винных молочнокислых бактерий: через путь аргининдеиминазы или путь аргиназа-уреаза? J. Appl. Бактериол . 81, 486–492.
Лукас, С., Варуча, Д., Зиудру, К., Стрити, Р. А., и Клее, В. А. (1983). Опиоидная активность и структура экзорфинов, происходящих из альфа-казеина. Биохимия 22, 4567–4573.
Pubmed Аннотация | Pubmed Полный текст
Лукас, П. М., Волкен, В. А., Клесс, О., Лолкема, Дж.С., и Лонво-Фунель, А. (2005). Путь продуцирования гистамина, кодируемый нестабильной плазмидой в Lactobacillus hilgardii (0006). Заявл. Environ. Микробиол . 71, 1417–1424.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Луптон, Дж. Р. (2004). Продукты микробной деградации влияют на риск рака толстой кишки: разногласия по поводу бутирата. Дж. Нутрь . 134, 479–482.
Pubmed Аннотация | Pubmed Полный текст
Макарова, К., Слесарев, А., Вольф, Ю., Сорокин, А., Миркин, Б., Кунин, Э., Павлов, А., Павлова, Н., Карамычев, В., Полушин, Н., Шахова, В. , Григорьев, И., Лу, Ю., Роксар, Д., Лукас, С., Хуанг, К., Гудштейн, Д.М., Хокинс, Т., Пленгвидья, В., Велкер, Д., Хьюз, Дж., Гох, Ю., Бенсон, А., Болдуин, К., Ли, Дж. Х., Диас-Муньис, И., Дости, Б., Смеянов, В., Вехтер, В., Баработе, Р., Лорка, Г. , Альтерманн, Э., Баррангу, Р., Ганесан, Б., Се, Й., Русторн, Х., Тамир, Д., Паркер, К., Брейд, Ф., Бродбент, Дж., Hutkins, R., O’Sullivan, D., Steele, J., Unlu, G., Saier, M., Klaenhammer, T., Richardson, P., Kozyavkin, S., Weimer, B., and Mills , Д. (2006). Сравнительная геномика молочнокислых бактерий. Proc. Natl. Акад. Sci. США . 103, 15611–15616.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Мартони К., Бхатена Дж., Урбанска А. М. и Пракаш С. (2008). Микроинкапсулированная гидролаза желчных солей, продуцирующая Lactobacillus reuteri для пероральной адресной доставки в желудочно-кишечный тракт. Заявл. Microbiol. Биотехнология . 81, 225–233.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Матин, М.А., Отани, Х. (2002). Цитотоксическая и антибактериальная активность химически синтезированного каппа-казецидина и его частичных пептидных фрагментов. Дж. Дайри Рес . 69, 329–334.
Pubmed Аннотация | Pubmed Полный текст
Маццоли, Р., Пессионе, Э., Дюфур, М., Ларут, В., Джуффрида, М. Г., Джунта, К., Кокаин-Буске, М., и Лубьер, П. (2010). Индуцированные глутаматом метаболические изменения в Lactococcus lactis NCDO 2118 во время продукции ГАМК: комбинированный транскриптомный и протеомный анализ. Аминокислоты 39, 727–737.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Мейзел, Х. (1998). Обзор пептидов, полученных из молочного белка. Внутр. Молочный J . 8, 363–373.
Миллетт М., Дюпон К., Аршамбо Д. и Лакруа М. (2007). Частичная характеристика бактериоцинов, продуцируемых человеческими изолятами Lactococcus lactis и Pediococccus acidilactici . J. Appl. Микробиол . 102, 274–282.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Мойно, С., Боркаев, М., Холлер, Б. Дж., Уокер, С. А., Кондо, Дж. К., и Ведамутху, Э. Р. (1996). Выделение и характеристика лактококковых бактериофагов из культивируемых растений пахты в США. J. Dairy Sci . 79, 2104–2111.
Моленаар Д., Босчер Дж. С., тен Бринк Б., Дриссен А. Дж. И Конингс В. Н. (1993). Генерация протонной движущей силы декарбоксилированием гистидина и электрогенным антипортом гистидина / гистамина у Lactobacillus buchneri . Дж. Бактериол . 175, 2864–2870.
Pubmed Аннотация | Pubmed Полный текст
Монтальбан-Лопес, М., Санчес-Идальго, М., Вальдивия, Э., Мартинес-Буэно, М., и Македа, М. (2011). Недостаточно ли используются бактериоцины? Новые приложения для старых противомикробных препаратов. Curr. Pharm. Биотехнология . 12, 1205–1220.
Pubmed Аннотация | Pubmed Полный текст
Морено-Аррибас М. В., Поло М. К., Йорганес Ф. и Муньос Р. (2003). Скрининг продукции биогенных аминов молочнокислыми бактериями, выделенными из виноградного сусла и вина. Внутр. J. Food Microbiol . 84, 117–123.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Мюррей, В. Р., Бэквуд, А., Троттер, Дж. М., Калман, К. С., и Маккей, К. (1980). Фекальные желчные кислоты и клостридии в этиологии колоректального рака. руб. J. Cancer 41, 923–928.
Pubmed Аннотация | Pubmed Полный текст
О’Флаэрти, С., Кленхаммер, Т. (2011). Анализ экспрессии генов эпителиальных клеток кишечника человека, подвергшихся воздействию молочнокислых бактерий. Тезисы к 10-му симпозиуму по молочнокислым бактериям , Эгмонд-ан-Зее, Нидерланды, 28 августа — сентябрь 1 2011 г., плакат C076.
Пеллиццаро К., Корадини Д. и Дайдон М. Г. (2002). Модуляция синтеза белков, связанных с ангиогенезом, бутиратом натрия в клеточной линии рака толстой кишки HT29. Канцерогенез 23, 735–740.
Pubmed Аннотация | Pubmed Полный текст
Pessione, E., Mazzoli, R., Giuffrida, M. G., Lamberti, C., Гарсия-Моруно, Э., Барелло, К., Конти, А., и Джунта, К. (2005). Протеомный подход к изучению молочнокислых бактерий, продуцирующих биогенные амины. Proteomics 5, 687–698.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Pessione, E., Pessione, A., Lamberti, C., Coïsson, D. J., Riedel, K., Mazzoli, R., Bonetta, S., Eberl, L., and Giunta, C. (2009). Первое свидетельство мембраносвязанной, продуцирующей тирамин и бета-фенилэтиламин тирозиндекарбоксилазы в Enterococcus faecalis : протеомное исследование с двумерным электрофорезом. Proteomics 9, 2695–2710.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Голубь, Р. М., Куэста, Э. П., и Гилиллианд, С. Э. (2002). Связывание свободных желчных кислот клетками бактерий закваски йогурта. J. Dairy Sci . 85, 2705–2710.
Pubmed Аннотация | Pubmed Полный текст
Роуз, Д. П. (1997). Влияние пищевых жирных кислот на рак груди и простаты: данные экспериментов in vitro, и исследований на животных. Am. J. Clin. Нутр . 66 (Прил. 6), 1513S – 1522S.
Pubmed Аннотация | Pubmed Полный текст
Сальваторе, С., Хаузер, Б., Деврекер, Т., Виейра, М. К., Луини, К., Арриго, С., Несполи, Л., и Ванденплас, Ю. (2007). Пробиотики и цинк при остром инфекционном гастроэнтерите у детей: эффективны ли они? Nutrition 23, 498–506.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Санчес Б., Брессольер П. и Урдачи М.С. (2008). Экспортируемые белки в пробиотических бактериях: адгезия к поверхности кишечника, иммуномодуляция хозяина и молекулярный перекрестный диалог с хозяином. FEMS Immunol. Med. Микробиол . 54, 1–17.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Зауэр, Дж., Рихтер, К. К., и Пул-Зобель, Б. Л. (2007). Физиологические концентрации бутирата благоприятно модулируют гены окислительного и метаболического стресса в первичных клетках толстой кишки человека. J. Nutr. Биохим .18, 736–745.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Шварц К. и Фольц К. М. (1957). Селен как неотъемлемая часть фактора 3 против диетического некроза печени. J. Am. Chem. Soc . 79, 3292–3293.
Pubmed Аннотация | Pubmed Полный текст
Зизен, Р. Дж., Ван Энкеворт, Ф. Х., Клеребезем, М., и Тойзинк, Б. (2004). Анализ геномных данных молочнокислых бактерий: влияние биоинформатики. Curr. Opin. Биотехнология .15, 105–115.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Сипола М., Финкенберг П., Вапаатало Х., Пихланто-Леппала А., Корхонен Х., Корпела Р. и Нурминен М. Л. (2002). Альфа-лакторфин и бета-лакторфин улучшают артериальную функцию у крыс со спонтанной гипертонией. Life Sci . 71, 1245–1253.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Смитс, Х. Х., Энгеринг, А., ван дер Клей, Д., де Йонг, Э.К., Шиппер, К., ван Капел, Т. М., Заат, Б. А., Язданбахш, М., Виренга, Э. А., ван Койк, Ю., и Капсенберг, М. Л. (2005). Селективные пробиотические бактерии индуцируют продуцирующие IL-10 регуляторные Т-клетки in vitro , модулируя функцию дендритных клеток посредством специфичной для дендритных клеток межклеточной адгезии, 3-захватывающей неинтегрин. J. Allergy Clin. Иммунол . 115, 1260–1267.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Сент-Онж, М.П., Фарнворт, Э. Р., и Джонс, П. Дж. (2000). Потребление ферментированных и неферментированных молочных продуктов: влияние на концентрацию холестерина и метаболизм. Am. J. Clin. Нутр . 71, 674–681.
Pubmed Аннотация | Pubmed Полный текст
Тернбо, П. Дж., Лей, Р. Э., Маховальд, М. А., Магрини, В., Мардис, Е. Р., и Гордон, Дж. И. (2006). Микробиом кишечника, связанный с ожирением, с повышенной способностью собирать энергию. Nature 444, 1027–1031.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Тытгат, Х.Л. П., Лебеер, С., Клаас, И., Скуфс, Г., Верхувен, Т. Л. А., Де Кеерсмакер, С. К. Дж., Маршал, К., и Вандерлейден, Дж. (2011). «Гликобиология LGG», в Тезисы на 10-м симпозиуме по молочнокислым бактериям , Эгмонд-ан-Зее, Нидерланды, 28 августа — 1 сентября 2011 г., SL 15.
ван Хиджум, С. А., Краль, С., Озимек, Л. К., Дийкхейзен, Л., и ван Гил-Шуттен, И. Г. Х. (2006). Взаимосвязь между структурой и функцией глюкансахаразы и ферментов фруктансахаразы из молочнокислых бактерий. Microbiol. Мол. Биол. Ред. . 70, 157–176.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Воан, Э. Э., де Фрис, М. К., Зетендаль, Э. Г., Каутер Бен Амор, Аккерман, А. Д. Л. и Де Вос, В. М. (2002). Кишечные LAB. Антони Ван Левенгук 82, 341–352.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Ямамото, Н., Маэно, М., и Такано, Т. (1999). Очистка и характеристика антигипертензивного пептида из йогуртоподобного продукта, ферментированного Lactobacillus helveticus CPN4. J. Dairy Sci . 82, 1388–1393.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
Чжао, Ф. Дж., Лопес-Беллидо, Ф. Дж., Грей, К. В., Уолли, В. Р., Кларк, Л. Дж., И МакГрат, С. П. (2007). Влияние уплотнения почвы и орошения на концентрацию селена и мышьяка в зернах пшеницы. Sci. Тотал Энвирон . 372, 433–439.
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст
D-Молочная кислота как метаболит: токсикология, диагностика и обнаружение
Существуют два энантиомера молочной кислоты.В то время как L-молочная кислота является обычным соединением метаболизма человека, D-молочная кислота вырабатывается некоторыми штаммами микроорганизмов или некоторыми менее важными метаболическими путями. В то время как L-молочная кислота является эндогенным соединением, D-молочная кислота является вредным энантиомером. Воздействие D-молочной кислоты может происходить различными путями, включая зараженные продукты питания и напитки, а также микробиоту во время некоторых патологических состояний, таких как синдром короткой кишки. Воздействие D-молочной кислоты невозможно диагностировать, поскольку обычные аналитические методы не подходят для различения двух энантиомеров.В этом обзоре представлены пути распространения D-молочной кислоты, патологические процессы, а также диагностические и аналитические методы, за которыми следуют рисунки и таблицы. Обобщается и обсуждается текущая литература.
1. Введение
Токсичность оптических изомеров — это особая проблема, которую нелегко изучить. Когда соединение существует в форме оптического изомера (энантиомера), часто один изомер безвреден или даже необходим для гомеостаза (например, L-аминокислоты), в то время как другой изомер токсичен и способен вмешиваться в важные пути в организме.С другой стороны, некоторые изомеры не вызывают серьезных проблем (например, сахара). Токсичность, зависящая от хиральности, известна для природных токсинов, лекарств и пестицидов [1–5]. Производство энантиопчистых препаратов — это особый вопрос, в котором предъявляются высокие требования к качеству изготовления и контроля. Производственные процессы должны иметь возможность отдавать предпочтение только одному изомеру или быть полностью селективным по отношению к нему. Этого можно достичь с помощью биокатализа [6], расширенного органического синтеза [7] и энантиоспецифического разделения [8].
Хотя неравный биологический эффект оптических изомеров известен относительно давно, его часто недооценивают, поскольку для различения изомеров необходимы сложные и дорогостоящие лабораторные тесты. Этот обзор посвящен D-молочной кислоте, которая по сравнению с ее L-изомерным аналогом проблематична для людей, а чрезмерное потребление может иметь фатальные последствия. Обзор актуальной литературы, обсуждение известных фактов и текущих исследований представлены в этой работе.
2. Сравнение D- и L-молочной кислоты
Молочная кислота существует в форме двух энантиомеров: D-молочной кислоты и L-молочной кислоты. В правильных химических терминах L-молочная кислота должна называться L (+) молочная кислота или S (+) молочная кислота, в то время как вариант D называется D (-) молочной кислотой, соответствующей R (-) молочной кислоте. Изомеры изображены на рисунке 1.
Молочная кислота существует в виде конъюгированного основного лактата (L-лактат в случае L-молочной кислоты и D-лактат в случае D-молочной кислоты) при физиологическом pH 7.4, но наличие сопряженного основания не влияет на хиральность, которая сохраняется в основном анионе. L-молочная кислота является естественным энантиомером человека и других высших форм жизни. Нормальный уровень L-лактата в крови человека находится в диапазоне от 0,5 до 1 ммоль / л, повышенный уровень выше нормального физиологического диапазона называется гиперлактатемией и может быть инициирован некоторыми патологическими процессами. Низкий уровень L-лактата называется гиполактатемией [9]; однако гиполактатемия — довольно редкое явление по сравнению с указанием на патологические состояния гиперлактатемией, что является важным открытием в клинической биохимии.Лактоацидоз возникает, когда концентрация L-лактата в плазме превышает 4 ммоль / л, а pH крови в этой ситуации может упасть ниже 7,35. Гиперлактатемия в легкой форме или в форме лактоацидоза может возникать по нескольким причинам, таким как сепсис, геморрагический шок, остановка сердца, травмы, отравления, ишемия, ожоги, диабетический кетоацидоз, некоторые виды рака и интенсивная мышечная активность [10–18]. .
Стандартный метаболизм L-лактата у большинства организмов опосредуется L-лактатдегидрогеназой (EC 1.1.1.27) в присутствии NAD + в качестве кофактора. Реакция по своему принципу аналогична окислению D-лактата D-лактатдегидрогеназой (EC 1.1.1.28). Фермент достаточно консервативен с точки зрения эволюции, поскольку его можно найти у эукариот, бактерий и даже архей. Пируват и НАДН являются продуктами реакции. Принцип окисления L-лактата до пирувата под действием L-лактатдегидрогеназы показан на рисунке 2. L-лактатдегидрогеназа участвует в основном метаболизме, тесно связанном с гликолизом и глюконеогенезом, и является важной частью цикла Кори у человека. и высших животных, но он также участвует в процессах ферментации [19–23].
По сравнению с L-молочной кислотой, D-молочная кислота не участвует в основных метаболических процессах большинства форм жизни. Ферменты, ответственные за метаболизм лактата, включая L-лактатдегидрогеназу, проявляют специфичность только к L-изомеру, и они не способны преобразовывать D-молочную кислоту. Только некоторые исключительные L-лактатдегидрогеназы, такие как фермент из Leuconostoc mesenteroides , способны участвовать в превращении D-лактата помимо стандартного метаболизма L-лактата [24].
3. Основные биохимические пути получения D-молочной кислоты
Различные виды бактерий способны одновременно продуцировать D-лактат или одновременно D- и L-лактаты. Многие из них участвуют в процессах ферментации, включая процессы, известные из биотехнологической промышленности. Род бактерий Lactobacillus продуцирует D, L и рацемическую смесь, род Pediococcus продуцирует либо чистый L, либо рацемическую смесь некоторого штамма, Leuconostoc и Oenococcus являются продуцентами D-изомера, а род Weissella продуцирует либо D-изомер, либо рацемическая смесь [25].Сконструированные штаммы бактерий могут служить для производства оптических изомеров. Асо и соавторы описали основанную на Lactococcus lactis биотехнологию для производства D-молочной кислоты [26].
Существует больше способов производства D-лактата микроорганизмами, грибами и другими. D-лактатдегидрогеназа (EC 1.1.1.28) — одна из них. Грибы Phytophthora undulata , Pythium debaryanum и Sapromyces elongatus [27] и бактерии Lactobacillus delbrueckii [28], Lactobacillusbulgaricus [28], Lactobacillus bulgaricus, 9, colseudo249, colsea, и Lactobacillus bulgaricus, 9, colse, , colse, , colse, 9, colse, , colse, 902 aeruginosa [30] являются организмами, известными своей экспрессией D-лактатдегидрогеназы.D-лактатдегидрогеназа — это фермент, который превращает D-лактат в пируват путем современного восстановления NAD + до NADH.
D-лактатдегидрогеназа (цитохром) — еще один фермент, превращающий D-лактат. Это фермент, известный под кодом EC 1.1.2.4. Он потребляет D-лактат и требует двух молекул феррицитохрома с, в то время как пируват и две молекулы ферроцитохрома с являются продуктами реакции. Активность этого фермента описана, например, на растении Arabidopsis thaliana [31], дрожжах Hansenula polymorpha [32] и дрожжах Saccharomyces cerevisiae [33].Участие D-лактатдегидрогеназы (цитохрома) в метаболизме человека широко не исследовалось. В исследовании описана активность фермента в митохондриях человека и его делеция у лиц с дефицитом фермента, а также обсуждена связь с появлением D-лактата в моче у лиц с дефицитом фермента [34].
D-лактатдегидрогеназа (цитохром с-553) представляет собой фермент, подобный D-лактатдегидрогеназе (цитохрому) или, возможно, в некоторых случаях с такой же биологической структурой.Это фермент, окисляющий D-лактат до пирувата с одновременным восстановлением двух молекул феррицитохрома с-553 до двух молекул ферроцитохрома с-553 за одну стадию. Этот фермент описан у бактерии Desulfovibrio vulgaris [35]. Подробная информация о ферменте не представлена в текущей литературе.
Глиоксалаза 3 — это фермент, принимающий D-лактат в качестве субстрата и участвующий в его детоксикации. Фермент также известен под названием D-лактатдегидратаза и код EC 4.2.1.130. Он катализирует реакцию, в которой D-лактат производит метилглиоксаль (2-оксопропаналь) и одну молекулу воды. Этот фермент был идентифицирован у человека [36]. Глиоксалазы 1 и 2 также являются ферментами, участвующими в соответствующей продукции конверсии D-лактата у людей [37, 38]. Подобно тому, как система глиоксалаз связывает D-лактат и метилглиоксаль, метилглиоксаль в дальнейшем участвует в формировании конечных продуктов гликирования, и, в более широком смысле, он играет роль в нейродегенеративных нарушениях окислительного стресса [39–42].Ферменты, участвующие в метаболизме лактата, сведены в Таблицу 1.
Отравление |
— D-D 902. Молочнокислый ацидоз
Производство молочных кислот является обычным делом в биотехнологиях пищевой промышленности, а производство некоторых видов продуктов питания и напитков обычно основано на производстве молочной кислоты или связано с ним. Можно привести примеры процессов, основанных на так называемых молочнокислых бактериях [43].Производство молока и молочных напитков может быть затруднено загрязнением D-молочной кислотой в зависимости от типа присутствия бактерий [44]. Существует сомнительное влияние пробиотиков на повышенный уровень D-лактата в кровообращении. Хотя некоторые работы указывают на потенциальные риски пробиотиков, другие не показывают доказательств увеличения D-лактата из-за пробиотиков, и окончательный вывод еще не сделан [45–48]. Более того, продукция D-лактата пробиотиками не связана со всеми штаммами, и использование штаммов, продуцирующих D-молочную кислоту, регулируется, особенно в продуктах для детей [25, 49].В других экспериментах было доказано защитное действие пробиотиков, и их применение было связано со снижением уровня D-лактата [50]. Производство D-лактата микробиотой в организме человека в нормальных условиях ограничено. Микробиота может перепроизводить D-лактат при определенных обстоятельствах, например, при синдроме короткой кишки и операции тощего шунтирования, которые дополнительно поддерживаются, когда пациент принимает пищу с высоким содержанием сахара [51, 52]. Синдром абдоминального компартмента, полиорганная недостаточность, связанная с накоплением жидкости в перитонеальном и забрюшинном пространствах, является еще одним фактором риска повышения концентрации D-лактата в крови [53].Присутствие D-лактата может быть следствием отравления тяжелыми металлами, несмотря на то, что точный механизм остается неизвестным. Это было точно описано на модели крыс линии Вистар, подвергшихся воздействию свинца [54]. Инфекционные заболевания и заболевания, сопровождающиеся воспалительными процессами, также могут быть причиной или быть связаны с повышенным уровнем D-лактата [55–58]. D-лактат также может служить маркером некоторых инфекций и сепсиса [59]. D-лактоацидоз образуется из-за перепроизводства D-лактата [60]. D-молочная кислота может содержаться в продуктах питания и напитках, приготовленных с помощью биотехнологических процессов, или может быть загрязнена микроорганизмами или иным образом; е.е., пиво может быть загрязнено D-молочной кислотой [61–64]. D-лактат вызывает различные патологические проявления в зависимости от дозы, индивидуальных условий и специфического метаболизма. D-лактат оказывает прямое нейротоксическое действие, которое не зависит от падения pH крови и не свойственно L-лактату, который был тщательно изучен на телят голштинской породы в качестве экспериментальной модели [65]. В случае ягнят, отравленных D-лактатом, были выявлены ацидоз, атаксическая походка и предпочтительное положение лежа, а также возможная сонливость, и симптомы можно было подавить с помощью бикарбоната натрия [66].Энцефалопатия также является частым синдромом отравления D-лактатом [67, 68]. Нейротоксический эффект D-лактата может проявляться эпизодической спутанностью сознания и гиперпноэ [69]. Общее замешательство, головокружение, головная боль, агрессивное поведение и потеря памяти — это другие симптомы отравления D-молочной кислотой [70]. Обзор типов воздействия D-молочной кислоты и воздействия с проявлениями показан на рисунке 3. Хотя D-лактат может вызывать осложнения для здоровья, он не является высокотоксичным соединением, поскольку ожидаемые средние летальные дозы довольно высоки.Уровень значения LD 50 на перорально отравленных крыс составляет около 4,5 г / кг.
Из-за проблем с окружающей средой, хорошей биосовместимости и применения в 3D-печати полимолочная кислота (синоним полилактид) становится очень предпочтительным полимером, который может разлагаться в природе [71–76]. Полилактические полимеры, представленные на современном рынке, обычно получают из дешевой рацемической смеси D- и L-молочных кислот, и ее можно химически назвать поли-DL-лактидом. Использование оптически чистых изомеров, из которых образуются поли-D-лактид и поли-L-лактид, сделало бы конечные пластмассовые изделия более дорогими.С другой стороны, наиболее коммерчески доступной молочной кислотой является энантиомер L, полученный в процессе ферментации, а другим важным источником является смесь, полученная химическим путем из ацетальдегида [77]. Возможен также гидролиз целлюлозы [78, 79]; существуют также протоколы производства D-лактата из метана [80]. Полимолочная кислота может быть разложена различными процессами, но гидролиз связанного сложного эфира является наиболее распространенной реакцией с химической точки зрения [81–83]; многие организмы, такие как бактерия Rhodopseudomonas palustris [84] и грибы Aspergillus niger и Candida cylindracea [85], являются примерами, которые обладают хорошей способностью гидролизовать полимолочную кислоту.В гидролизе полимолочной кислоты не участвует ни один фермент. Липазы [85], карбоксилэстеразы [86] и сериновые протеазы [87] были названы ферментами, способными гидролизовать полимолочную кислоту. Токсичность полимеров на современном рынке, как правило, низкая, и ее трудно измерить. Однако в некоторых работах были выявлены риски, связанные с материалами, доступными на текущем рынке, а биоразлагаемые материалы, такие как полимолочная кислота, оказались на удивление более опасными в модели in vitro [88]. Что касается полимолочной кислоты, следует учитывать высвобождение D-лактата из-за метаболизма; следовательно, продукты, приготовленные из поли-DL-лактида и поли-D-лактида, могут представлять определенный риск при проглатывании. 5. Диагностика и терапияВоздействие D-молочной кислоты можно диагностировать с помощью стандартной биохимии, где некоторые маркеры демонстрируют хорошее соотношение доза-ответ. Кроме того, можно напрямую измерить остаточный уровень D-лактата в кровотоке. Биохимические маркеры могут указывать на тип отравления органической кислотой, но нелегко сделать вывод, что D-молочная кислота является возбудителем отравления, до тех пор, пока не будут проведены дальнейшие аналитические тесты. Точный анализ D-лактата в кровотоке непрост, потому что L-лактат и D-лактат имеют одинаковые физические и химические свойства, и большинство аналитических методов не могут их различить.В то же время L-лактат будет мешать анализу, потому что физиологическая концентрация в крови или плазме крови довольно высока; его концентрация может достигать 2,5 ммоль / л, а в некоторых условиях может даже превышать 4 ммоль / л [89]. В случае легкой гиперлактатемии L-лактат может достигать 7 ммоль / л и 12 ммоль / л в умеренном случае, а у пациентов с тяжелой гиперлактатемией концентрация может превышать 12 ммоль / л [90]. Стандартные биохимические маркеры могут помочь в диагностике воздействия D-лактата и оценить присутствие D-лактата в кровотоке.D-молочная кислота количественно продуцирует D-лактат в физиологических условиях, что приводит к падению pH крови с возможным возникновением большого анионного промежутка и метаболического ацидоза. Влияние D-лактата на биохимические маркеры можно показать на примере 14-летнего мальчика, страдавшего синдромом короткой кишки [69]. У пациента была значительно более высокая концентрация D-лактата, чем L-лактата в плазме крови. В то время как L-лактат был представлен в концентрации 2,89 ммоль / л, D-лактат достигал 11.2 ммоль / л одновременно. Избыток основания был равен -19,1 ммоль / л, анионный разрыв — 28,2 ммоль / л, а pH крови упал до 7,23. В другом описанном случае синдрома короткой кишки концентрация D-лактата в крови достигла 8,9 ммоль / л, в то время как L-лактат был равен 1,4 ммоль / л, а pH упал до 7,30 с одновременным избытком основания -11,8 ммоль / л в девяти случаях. -летний мальчик [91]. В клиническом случае описана пятилетняя девочка с синдромом короткой кишки [92]. Во время госпитализации у нее были следующие показатели сыворотки крови: pH 7.16, бикарбонат 5,2 ммоль / л, избыток основания -20,2 ммоль / л, L-лактат 0,92 ммоль / л и D-лактат 8,19 ммоль / л. Во всех трех предыдущих описаниях случались рецидивы и снова появлялся ацидоз, несмотря на медицинское наблюдение. Обзор биохимических параметров из представленных отчетов о случаях представлен в таблице 2.
Конечно, на биохимические маркеры могут влиять другие патологии, как представлено в отчете Хейремана и его коллег о пациенте, который одновременно страдает уросепсисом. неконтролируемый сахарный диабет 2 типа, передозировка парацетамолом и операция желудочного шунтирования [93].Эта комбинация привела к нарушению функции почек, снижению сознания, гипервентиляции, диарее, рвоте и развитию метаболического ацидоза с высоким анионным разрывом. Ацидоз и изменение других биохимических маркеров могут быть инициированы другими химическими веществами аналогично тому, как описано здесь для D-молочной кислоты. Этиленгликоль и метанол можно назвать органическими соединениями, инициирующими развитие ацидоза, поскольку в результате метаболизма из них образуются органические кислоты — гликолевая кислота, глиоксиловая кислота и щавелевая кислота из этиленгликоля и муравьиная кислота из метанола [94–96]. Распознавание точного типа яда невозможно без дальнейшей идентификации с помощью химического анализа. Существуют подходящие ферментные анализы, биоанализы и биосенсоры, в которых используются ферменты, специфичные только для одного изомера [33, 97–99]. Электрохимический биосенсор с D-лактатдегидрогеназой из архей Candidatus caldiarchaeum subterraneum был сконструирован Satomura и соавторами, и он был пригоден для определения концентрации D-лактата в диапазоне 0,03–2.5 ммоль / л [97]. D-лактат оксидоредуктаза (цитохром) дрожжей Saccharomyces cerevisiae служила для создания амперометрического биосенсора с полной селективностью к D-лактату [98]. D-лактат можно легко измерить с помощью обычного спектрального анализа в сочетании с распознавательной способностью селективного фермента. D-лактатдегидрогеназа была выбрана в качестве инструмента для измерения D-лактата в плазме лабораторных крыс [99]. В этой работе фермент окислял D-лактат до пирувата с одновременным восстановлением NAD + до NADH.Созданный НАДН давал сигнал флуоресценции с возбуждением при 340 нм и испусканием при 491 нм. Стандартные инструментальные аналитические методы, такие как хроматография или масс-спектрометрия, обычно не подходят для различения оптических изомеров; однако можно сделать некоторые улучшения, чтобы сделать их выборочными. Динг с сотрудниками дериватизировали D- и L-молочные кислоты с помощью L-ментола, и следующая газовая хроматография (ГХ) с масс-спектрометрией (МС) была подходящей для их селективного анализа [100].В другой работе (S) (+) — 1- (2-пирролидинилметил) -пирролидин служил для дериватизации D- и L-молочной кислоты, а продукт дериватизации был проанализирован с помощью сверхпроизводительной жидкостной хроматографии с масс-спектрометрией [101]. Еще одна возможность для целей анализа — высокоэффективная жидкостная хроматография с хиральной колонкой. Это было, например, выполнено Генри и сотрудниками для определения D-лактата в моче, и они объединили высокоэффективную жидкостную хроматографию (ВЭЖХ) с МС и использовали хиральную колонку Astec Chirobiotic ™ R для достижения селективности по изомерам [102] .Обзор аналитических методов для D-молочной кислоты представлен в Таблице 3.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||