строение, свойства, функции — урок. Биология, Общие биологические закономерности (9–11 класс).
Среди органических веществ клетки самыми разнообразными по свойствам и выполняемым функциям являются белки, или протеины. В белках, в отличии от углеводов и липидов, кроме углерода, кислорода и водорода содержится азот, а также могут присутствовать атомы серы, фосфора и железа.
Белки — это биополимеры, мономерами в которых служат аминокислоты. В образовании всего разнообразия белков участвует \(20\) α-аминокислот. Молекулы аминокислот имеют две функциональные группы: карбоксильную (кислотную) и аминогруппу (основную).
Рис. \(1\). Молекула аминокислоты
Аминогруппа и карбоксильная группа способны взаимодействовать между собой с отщеплением воды и образованием пептидной связи CO−NH. Пептидными связями молекулы аминокислот соединяются друг с другом в длинные цепи./protein-hemoglobin-58a222955f9b58819ca8f902.jpg) Число остатков аминокислот в цепи может составлять несколько сотен и даже тысяч. Такие большие молекулы называют макромолекулами.
Число остатков аминокислот в цепи может составлять несколько сотен и даже тысяч. Такие большие молекулы называют макромолекулами.
Структура белков
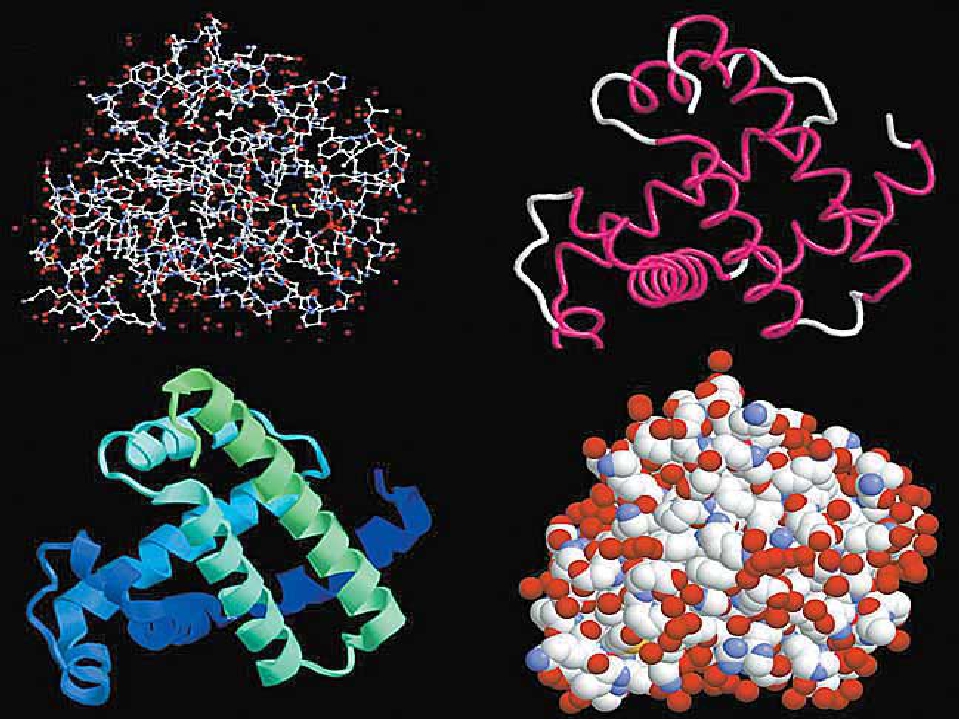
Порядок соединения аминокислот в макромолекуле белка называют первичной структурой. Для каждого типа белка эта структура уникальна. Она определяет структуры высших уровней, свойства белка и его функции.
Полипептидная цепь сворачивается в спираль за счёт образования водородных связей между группировками атомов −NH и −CO, расположенными на разных участках макромолекулы. Эту спираль называют вторичной структурой белка.
Третичная структура белка возникает при взаимодействии радикалов аминокислот, а также за счёт дисульфидных мостиков, водородных и ионных связей. Молекула белка принимает форму глобулы (шарика).
У некоторых белков формируется четвертичная структура. Она представляет собой комплекс нескольких макромолекул, имеющих третичную структуру. Четвертичную структуру удерживают непрочные ионные и водородные связи, а также гидрофобные взаимодействия.
Четвертичную структуру удерживают непрочные ионные и водородные связи, а также гидрофобные взаимодействия.
Рис. \(2\). Структуры белка
Белки могут соединяться с углеводами, жирами и нуклеиновыми кислотами с образованием комплексных соединений: гликопротеинов, липопротеинов, нуклеопротеинов.
Под действием внешних факторов: облучения, нагревания, некоторых химических веществ и др. — происходит нарушение пространственной структуры белковых молекул. Этот процесс называется денатурацией.
Сначала происходит разрушение четвертичной структуры, потом третичной и вторичной. Первичная структура при денатурации сохраняется, но белок утрачивает свои свойства и функции.
Денатурация в некоторых случаях обратима. Обратный процесс называется ренатурацией.
Рис. \(3\). Денатурация и ренатурация белка
Разрушение первичной структуры необратимо.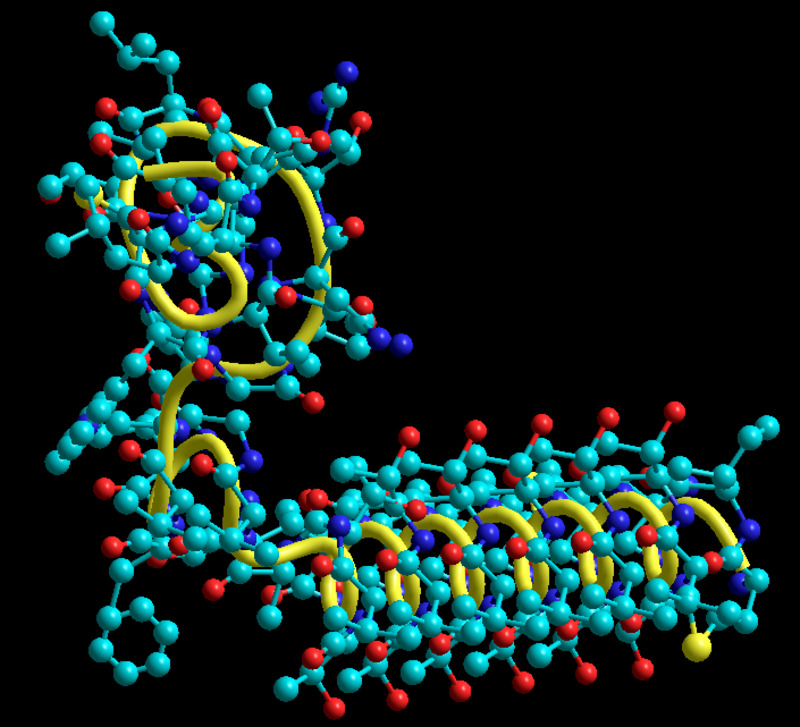 Оно происходит при гидролизе белка — макромолекулы распадаются на отдельные аминокислоты. Такой процесс идёт в органах пищеварения животных и в лизосомах клеток под действием гидролитических ферментов.
Оно происходит при гидролизе белка — макромолекулы распадаются на отдельные аминокислоты. Такой процесс идёт в органах пищеварения животных и в лизосомах клеток под действием гидролитических ферментов.
Функции белков
1. Важнейшей функцией белков является каталитическая, или ферментативная. Белки-ферменты участвуют во всех биохимических реакциях, протекающих в клетке, и повышают скорость этих реакций во много раз. Для каждой реакции существует особый фермент.
2. Белки выполняют структурную (строительную) функцию. Они входят в состав плазматических мембран, образуют соединительные ткани (эластин и коллаген), волосы и ногти (кератин).
Рис. \(4\). Структурные белки в плазматической мембране
3. Сигнальную функцию также осуществляют белки, встроенные в мембрану. Под действием внешних факторов эти белки изменяют третичную структуру, что отражается на функционировании клетки.
4. Транспортная функция белков проявляется в переносе ионов через клеточные мембраны, транспорте гемоглобином крови кислорода и углекислого газа, альбуминами плазмы — жирных кислот и т. д.
5. Двигательную функцию обеспечивают белки актин и миозин, способные сокращаться и растягиваться. Они приводят в движение реснички и жгутики одноклеточных организмов, сокращают мышцы у животных.
Рис. \(5\). Сократительные белки
6. Защитная функция обеспечивается антителами иммунной системы организма, белками системы свёртывании крови (фибриногеном, протомбином и др.).
7. Регуляторную функцию выполняют белки-гормоны (инсулин, тиреотропин, соматотропин, глюкагон и др.).
8. Энергетическую функцию белки выполняют после израсходования запасов углеводов и жиров. При полном расщеплении \(1\) г белка до конечных продуктов выделяется \(17,6\) кДж энергии.
Источники:
Рис. 1. Молекула аминокислоты. Автор: X-romix — собственная работа, Общественное достояние, https://commons.wikimedia.org/w/index.php?curid=10280776. 09.09.2021.
Рис. 2. Структуры белка. https://image.shutterstock.com/image-vector/protein-structure-primary-secondary-tertiary-600w-1474657079
Рис. 3. Денатурация и ренатурация белка. © ЯКласс.
Рис. 4. Структурные белки в плазматической мембран. https://shutterstock.puzzlepix.hu/kep/376416385е
Рис. 5. Сократительные белки. © ЯКласс.
Биологи определили, какие белки коронавируса отвечают за «взлом» клеточных систем — Наука
ТАСС, 14 сентября. Ученые создали атлас трехмерных моделей вирусных белков, которые образуются при заражении коронавирусом нового типа. Благодаря этому исследователи выделили пять ключевых молекул, отвечающих за «взлом» клеточных систем. Результаты их исследования опубликовал научный журнал Molecular Systems Biology.
«Благодаря трехмерным моделям белков мы получили критически важную информацию о структуре SARS-CoV-2, необходимую для создания лекарств и вакцин. В будущем такие модели помогут быстро выяснить, чем отличаются новые разновидности коронавируса от уже известных, то есть упростит борьбу с ними», – рассказал один из авторов статьи, профессор Института медицинских исследований Гарвана (Австралия) Шон О’Донохью.
В будущем такие модели помогут быстро выяснить, чем отличаются новые разновидности коронавируса от уже известных, то есть упростит борьбу с ними», – рассказал один из авторов статьи, профессор Института медицинских исследований Гарвана (Австралия) Шон О’Донохью.
Геном коронавируса нового типа представляет собой одноцепочную молекулу РНК, в которой находится около 30 тыс. «букв»-нуклеотидов. Они управляют производством всех белков оболочки SARS-CoV-2 и вспомогательных молекул, которые необходимы для размножения вируса в клетках людей и животных.
Помимо этого, коронавирус использует множество клеточных ферментов, чтобы копировать свой геном и упаковывать эти копии в новые вирусные частицы. Ученые достаточно давно пытаются понять, какие системы SARS-CoV-2 «взламывает» для этого, так как понимание подобного механизма позволит создать препараты, блокирующие цикл размножения вируса.
Уязвимые места коронавируса
О’Донохью и его коллеги уже много месяцев работают над этой проблемой в рамках масштабного проекта по составлению полного трехмерного «портрета» коронавируса. Для этого ученые подготовили точные модели всех 27 белков коронавируса и проанализировали свыше двух тысяч их возможных реакций с различными клеточными системами и ферментами.
Для этого ученые подготовили точные модели всех 27 белков коронавируса и проанализировали свыше двух тысяч их возможных реакций с различными клеточными системами и ферментами.
В результате исследователи обнаружили, что сразу три вирусных молекулы, NSP3, NSP13 и NSP16, имитируют по своей структуре клеточные белки, что помогает возбудителю COVID-19 избегать внимания врожденного иммунитета. Кроме того, исследователи выделили пять молекул, отвечающих за «взлом» различных клеточных систем, необходимых для формирования новых вирусных частиц.
«В дополнение к этому, мы выделили сразу восемь вирусных белков, которые взаимодействуют друг с другом и участвуют в сборке более сложных структур. Понимание того, как протекают эти реакции, помогло нам раскрыть многие неизвестные детали в процессе сборки новых копий вирусного генома», – пояснил О’Донохью.
Последующее изучение этих белковых молекул, как надеются исследователи, позволит им найти уязвимые точки в цикле размножения коронавируса и использовать эти сведения для создания лекарств и вакцин, атакующих данные механизмы. Подобные препараты потенциально будут более долговечными, так как они будут воздействовать на те части вируса, которые почти не меняются с течением времени.
Подобные препараты потенциально будут более долговечными, так как они будут воздействовать на те части вируса, которые почти не меняются с течением времени.
Лаборатория новых методов в биологии (№ 22) ИБП РАН
Заведующий лабораторией кандидат физико-математических наук Сергей Евгеньевич Пермяков
8 977 193 77 41
Фото лаборатории
Содержание
Направления деятельности
Группа исследования белков (руководитель — к.ф.м.н. С.Е. Пермяков)
- Исследование физико-химических и функциональных свойств металл-связывающих белков: α-лактальбумин, лизоцим, парвальбумин, кальмодулин, рековерин, S-100 и т.д. Металлопротеомика. Исследование роли катионов металлов в функционировании белков.
- Исследование механизмов функционирования металл-связывающих белков в клетке.
- Исследование нативно неупорядоченных белков. Белок-белковые взаимодействия.
- Разработка методов конструирования белковых структур с заданными свойствами.
- Разработка молекулярных основ новых методов диагностики и лечения онкологических заболеваний.
- Поиск и разработка новых перспективных экспериментальных методов для физико-химической биологии.
Картографическая группа (руководитель — В.И. Степанова)
- Разработка новых методов обработки карт различных поверхностей, а также сбор, обработка, систематизация, размножение и хранение графических, фотографических и картографических материалов.
Технические разработки
Состав лаборатории
- Пермяков Сергей Евгеньевич, к.ф.-м.н., и.о. зав. лаб. Новых методов в биологии
- Пермяков Евгений Анатольевич, д.б.н., проф., г.н.с.
- Денесюк Александр Ильич, д.ф.-м.н., г.н.с.
- Грищенко Валерий Мефодьевич, к.б.н., с.н.с.
- Емельяненко Виктор Иванович, к.ф.-м.н., c.н.с.
- Казаков Алексей Сергеевич, к.б.н., с.н.с.
- Авхачева Надежда Владимировна, к.
 б.н., с.н.с.
б.н., с.н.с. - Литус Екатерина Андреевна, к.м.н., с.н.с.
- Дерюшева Евгения Игоревна, к.ф.-м.н., с.н.с.
- Шевелева Марина Петровна, к.х.н., ст.н.с.
- Немашкалова Екатерина Леонидовна, к.б.н., с.н.с.
- Вологжанникова Алиса Андреевна, к.б.н., с.н.с.
- Соколов Андрей Сергеевич, к.б.н., с.н.с.
- Лаптева Юлия Сергеевна, к.б.н., с.н.с.
- Пермякова Мария Евгеньевна, н.с.
- Назипова Алия Анцаровна, н.с.
- Ломоносова Анна Викторовна, н.с.
- Растрыгина Виктория Александровна, н.с.
- Улитин Андрей Борисович, н.с.
- Боброва Лолита Алексеевна, м.н.с.
- Степанова Вера Игоревна, н.с., руководитель Группы картографии
- Баранов Игорь Павлович, инж.
- Клам Екатерина Евгеньевна, вед. констр.
- Платонова Татьяна Марьяновна, лаб.
Основные приборы и оборудование лаборатории
- Спектрополяриметр J-810, Jasco.
- Спектрофотометр Cary 100, Varian.
- Инфракрасный спектрометр с Фурье-преобразованием Nicolet-6700, Thermo Scientific.
- Хроматограф высокого давления Shimadzu
- Центрифуги Backman Coulter Avanti J-26XP; Hitachi Himac CP80WX; Eppendorf 5804R.
- Многоцветная система регистрации ПЦР в реальном времени IQ5 (BioRad).
- Гель-сканер Pharos FX Plus (BioRad).
- Система для измерения плазмонного резонанса ProteOn XPR (BioRad).
- Сканирующий калориметр Nano-DSC (AT Instruments).
- Изотермический титрационный калориметр Nano-ITC (AT Instruments).
- Оптический микроскоп (Kyence Biozero).
- Инкубатор CO2 (ИБП РАН).
- Дезинтегратор клеток (French press) (ИБП РАН).
- Дзета-сайзер Nano ZS (Malvern)
- Вискозиметр SV-10 (A&D Company)
- Генератор льда
- Стерилизатор
- Высокоскоростной сортировщик клеток MoFlo XDP, Beckman Coulter, USA
Гранты
- Грант SABIT (США) — 1993
- Грант CAST (США) — 1993
- Грант Сороса — 1994-1995
- Грант НАТО — 1994-1996
- Грант РФФИ — 1998
- Грант НАТО — 1999
- Грант РФФИ — 2001
- Грант INTAS — 2002
- Грант INTAS — 2004
- Грант РФФИ-Московской области — 2004
- Грант Программы Президиума РАН «Фундаментальные науки — медицине» — 2004
- Грант Программы Президиума РАН «Молекулярная и клеточная биология» — 2004-2008
- Грант CRDF — 2005
- Грант Фонда содействия отечественной науке — 2006
- Грант Федерального агенства по науке и инновациям — 2006
- Грант Программы Президиума РАН «Фундаментальные науки — медицине» — 2007
- Грант Президента РФ — 2007-2008
- Грант Программы Президиума РАН «Молекулярная и клеточная биология» — 2009-2013
- Грант Программы Президиума РАН «Фундаментальные науки — медицине» — 2012
- Грант РФФИ — 2012
- Грант Программы Президиума РАН «Фундаментальные науки — медицине» — 2013-2014
- Грант РФФИ — 2014
- Грант Программы Президиума РАН «Молекулярная и клеточная биология» — 2014
- Грант РФФИ-Московская область — 2014
- Лот НТП по приоритетному направлению «Науки о жизни» Министерства образования и науки — 2014
- Грант Российского научного фонда 2016-2018 гг.
- Грант Российского научного фонда — 2019-2021 гг.
- Грант РФФИ — 2019-2021 гг
- Грант Российского научного фонда — 2020-2023 гг.
Индекс цитирования руководителя и сотрудников лаборатории
Количество ссылок без самоцитирования и индекс Хирша за годы, начиная с 1975, согласно базе данных Google scholar:
Монографии
- Пермяков Е.А. Парвальбумин и родственные кальций-связывающие белки. Москва, «Наука», 1985, 191 с.
- Пермяков Е.А. Кальцийсвязывающие белки. Москва, Наука, 1993, 190 с.
- Permyakov, E.A. Luminescent Spectroscopy of Proteins. CRC Press, Boca Raton, Ann Arbor, London, Tokyo, 1993, 164 p.
- Пермяков Е.А. Метод собственной люминесценции белка. Москва, «Наука», 2003, 189 с.
- Степанов И.Н. Теория пластики рельефа и новые тематические карты. Москва, Наука, 2006, 229 с.
- Permyakov, E.A. a-Lactalbumin. Nova Science Publishers, New York, 2005, 136 p.

- Permyakov, E.A. Parvalbumin. Nova Science Publishers, New York, 2006, 196 p.
- Permyakov, E.A. Metalloproteomics. A John Wiley & Sons, Inc., Hoboken, New Jersey, 2009, 786 p.
- Permyakov, E.A., and Kretsinger, R.H. Calcium Binding Proteins. A John Wiley & Sons, Inc., Hoboken, New Jersey, 2011, 573 p.
- Пермяков Е.А. Металлсвязывающие белки: структура, свойства, функции. Научный мир, Москва, 2012, 544 с.
- Morozova, L.A., Berliner, L.J. and Permyakov, E.A. Calcium Binding Lysozymes. Nova Science Publishers, New York, 2012.
Сборники статей под редакцией В.Н. Уверского и Е.А. Пермякова
- Methods in Protein Structure and Stability Analysis. Part A. Luminescence Spectroscopy and Circular Dichroism. Nova Biomedical Books, New York, 2007, V.N. Uversky & E.A. Permyakov, eds.
- Methods in Protein Structure and Stability Analysis. Part B. Vibrational Spectroscopy. Nova Biomedical Books, New York, 2007, V.
 N. Uversky & E.A. Permyakov, eds.
N. Uversky & E.A. Permyakov, eds. - Methods in Protein Structure and Stability Analysis. Part С. Conformational Stability, Size, Shape and Surface of Protein Molecules. Nova Biomedical Books, New York, 2007, V.N. Uversky & E.A. Permyakov, eds.
- Methods in Protein Structure and Stability Analysis. Part С. NMR and EPR Spectroscopies, Mass-Spectrometry and Protein Imaging. Nova Biomedical Books, New York, 2008, V.N. Uversky & E.A. Permyakov, eds.
- Kaleidoscope of Structural Properties and Functions, Research Singpost, Kerala, India, 2003, Ed. V.N. Uversky.
- The Wiley Series in Protein and Peptide Science (Uversky V.N. series Ed.), John Wiley & Sons, Inc, Hoboken, New Jersey, USA. ISBN: 978-0-470-59169-7 (2012)
- Methods and Experimental Tools (Editors: Uversky V.N., Dunker A.K.), Humana Press, Totowa, NJ, USA. ISBN: 978-161-77-9926-6 (2012)
- Kretsinger R.H., Uversky V.N., Permyakov E.
 A. (eds) Encyclopedia of Metalloproteins. Springer, 2013, 2574 p. 1108 illus., 777 illus. in color. In 4 volumes.
A. (eds) Encyclopedia of Metalloproteins. Springer, 2013, 2574 p. 1108 illus., 777 illus. in color. In 4 volumes.
Авторские свидетельства
- Орешкин Е.Ф., Борисова М.А., Чубарова Г.С., Пермяков Е.А., Бурштейн Э.А. Способ определения максимально допустимой температуры нагрева мясного сырья высшего сорта. Авторское свидетельство N 1411666, 1988.
- Пермяков Е.А., Шныров В.Л., Дятчин И.В., Бурштейн Э.А. Способ определения концентрации кальция в растворе. Авторское свидетельство N 1580256, 1990.
- Ларин В.Т., Пермяков Е.А., Фрейдин А.А. Способ проведения групповых анализов биохимических образцов в жидкой форме. Патент на изобретение N2142638, 1999.
- Редикюльцев Ю.В., Ширшиков Н.В., Кудряшов В.К., Пермяков Е.А. Способ каскадно-проточного культивирования микроорганизмов. Патент на изобретение N2193594, 2002.
- WO 2015174978 A1 by Ilya Alexandrov, Khikmet Sadykov, Roman Mikhailov, Dina Abramovich, Sergei Permyakov, Tajib Mirzabekov, Alexey Repik, Vasily Ignatiev, Mikhail Samsonov, Yan Lavrovsky: “Antibodies against human receptor integrin alpha-4”; priority 2014/05/15
Защиты диссертаций
В лаборатории защитили диссертации:
- Дейкус Г.
 Ю., 1996 г., к.ф.м.н.
Ю., 1996 г., к.ф.м.н. - Вепринцев Д.Б., 1998 г., к.б.н.
- Уверский В.Н., 1998 г., д.ф.-м.н.
- Пермяков С.Е., 2001 г., к.ф.-м.н. — автореферат диссертации
- Морозова-Рош Л.А., 2002 г., д.ф.-м.н.
- Жадан А.П., 2003 г., к.б.н. — автореферат диссертации
- Бакунц А.Г., 2009 г., к.б.н. — автореферат диссертации
- Казанцев А.П., 2009 г., к.т.н.
- Князева (Немашкалова) Е.Л., 2010 г., к.б.н. — автореферат диссертации
- Сенин А.А., 2012 г., к.т.н.
- Лаптева Ю.С., 2012 г., к.б.н.
- Ломоносова А.В., 2017 г., к.б.н.
- Соколов А.С., 2018 г., к.б.н.
- Вологжанникова А.А., 2020 г., к.б.н.
- Хорн П.А., 2021 г., к.б.н.
Сотрудничество
Лаборатория сотрудничает со следующими научными организациями:
- Denver University, Denver, CO, USA
- University of South Florida, Tampa, FL, USA
- Abo Akademi University, Turku, Finland
- Umeo University, Umeo, Sweden
- Московский государственный университет, г.
 Москва
Москва - Филиал института биоорганической химии РАН, г. Пущино
- Институт теоретической и экспериментальной биофизики РАН, г. Пущино.
назад
Ученые нашли белок в костях динозавра возрастом 200 млн лет
- Хелен Бриггс
- Би-би-си
Автор фото, SPL
Подпись к фото,
Белки были найдены в костях древнего люфенгозавра
Внутри окаменелой кости динозавра, жившего, как полагают, примерно 195 миллионов лет назад, были найдены древние протеины.
Ранее имевшиеся в распоряжении ученых образцы относились к более позднему периоду, отстоящему от нас приблизительно на 100 миллионов лет.
Ученые также обнаружили следы минерала, который, вероятно, поступил из крови динозавра, жившего на заре Юрского периода.
Мягкие ткани обеспечивают новое понимание биологии динозавров и того, как они развивались.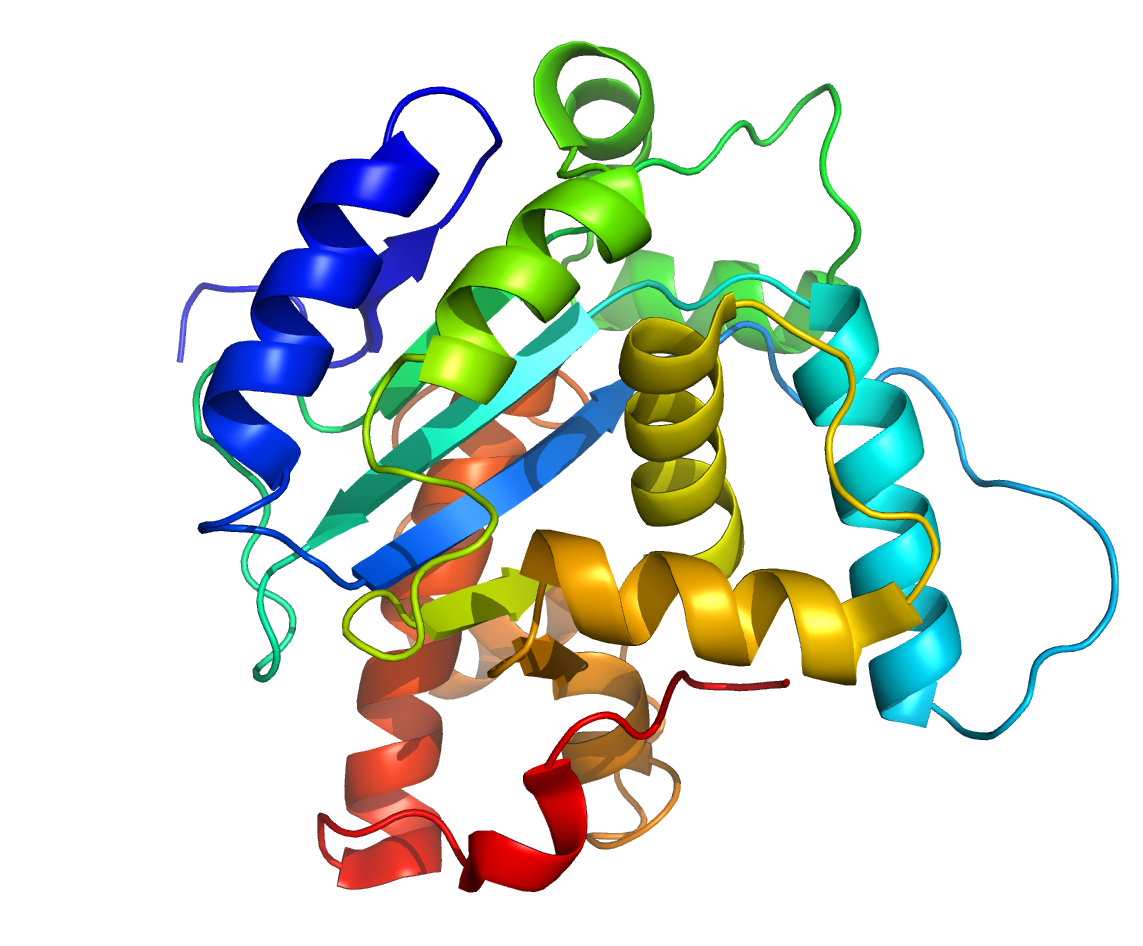 Части мягких тканей исключительно редко сохраняются в процессе фоссилизации, в ходе которого кости и зубы животных постепенно превращается в камень.
Части мягких тканей исключительно редко сохраняются в процессе фоссилизации, в ходе которого кости и зубы животных постепенно превращается в камень.
До сих пор считалось невозможным, чтобы белок мог сохраняться на протяжении столь длительного срока.
«Мы надеемся, что сможем узнать больше о биологии этих животных, — считает палеонтолог из канадского университета Торонто Миссиссога Роберт Рейсз. — Чем больше мы знаем об их мягких тканях, тем больше мы будем знать о них в целом. Мы сейчас больше изучаем сохранность материалов, которые были внутри живого организма, чем оттиски мягких тканей, оставленные им».
Автор фото, Robert Reisz
Подпись к фото,
Сохранившиеся фрагменты коллагена и богатых железом белков были обнаружены в стенках кровеносных сосудов, проходящих через реберные кости
Автор фото, Robert Reisz
Подпись к фото,
Ученые смогли исследовать химический состав реберной кости люфенгозавра, не вынимая ее содержимого и не повреждая его
Кровь динозавра
Ученые исследовали окаменелости люфенгозавра, найденные на Тайване, с помощью инфракрасной спектроскопии с синхротроном.
Сохранившиеся фрагменты коллагена и богатых железом белков были обнаружены в стенках кровеносных сосудов, проходящих через реберные кости.
Исследователи полагают, что остатки крови динозавра, возможно, сохраняли коллаген в течение почти 200 миллионов лет.
«Найти белки в ископаемом динозавре возрастом в 195 миллионов лет — это поразительное открытие, — сказал в интервью Би-би-си Стивен Брузатте из Эдинбургского университета, не участвовавший в данном исследовании. — Это звучит даже слишком хорошо, чтобы быть правдой. Но эта группа ученых использовала все доступные им методы, чтобы перепроверить свое открытие, так что, похоже, всё это действительно так».
По его словам, тот факт, что белки могут сохраняться так долго, позволяет сделать два важных вывода.
Сохранение микроскопических белков в течение почти 200 миллионов лет говорит о том, что нам еще предстоит многое узнать о процессе формирования окаменелостей.
Также находки, по всей видимости, подтверждают поступавшие ранее сообщения об обнаружении схожих белков в останках других динозавров, возраст которых оценивался примерно в 70 миллионов лет.
«Это открытие говорит нам, что, по всей вероятности, возможно сохранить мягкие микроскопические белки внутри костей динозавров на десятки или сотни миллионов лет, — добавил Брузатте. — И это открывает совершенно новые возможности для изучения динозавров».
Гигантское травоядное
Люфенгозавр принадлежит к группе динозавров, которые жили во время раннеюрского периода на юго-западе нынешнего Китая.
В ходе проводившихся там раскопок были найдены десятки скелетов травоядного ящера.
Нынешнее открытие сделано той же исследовательской группой, что ранее смогла обнаружить самых старых из когда-либо раскопанных эмбрионов динозавров.
Тайная жизнь белков коронавируса | Euronews
Ультрафиолетовые лазеры, масс-спектрометрия, ДНК, платформы для вакцин, белковые структуры. …. Европейские исследователи в поиске эффективного орудия борьбы с ковидом используют весь имеющийся арсенал знаний и инструментов. Глобальные усилия в области фундаментальных исследований, объединяющие, безусловно, вирусологов, но также физиков, химиков, генетиков программистов начнут, по мнению наблюдателей, приносить плоды. Посмотрим на прогресс научного сообщества ближе
…. Европейские исследователи в поиске эффективного орудия борьбы с ковидом используют весь имеющийся арсенал знаний и инструментов. Глобальные усилия в области фундаментальных исследований, объединяющие, безусловно, вирусологов, но также физиков, химиков, генетиков программистов начнут, по мнению наблюдателей, приносить плоды. Посмотрим на прогресс научного сообщества ближе
В Гамбурге исследователи (Heinrich Pette institute — Leibniz institute) спешат разгадать секрет структуры белков, позволяющих коронавирусам, включая новый подтип, так быстро размножаться в клетках человека. Чем лучше ученые поймут глубинную природу этих веществ, тем увереннее продвинутся на пути разработки вакцины или метопов лечения. Первый шаг — исследование процесса воспроизводства белков в клетках разного типа.
Вирусолог Борис Крихель отмечает: «Белки должны воспроизводиться в разных типах клеток, но некоторые сопротивляются нашим усилиям. Это немного сложно. Речь, по-видимому, идет о слишком крупных или модифицированных белках. Вот почему мы должны брать строго определенные виды клеток, которые представляют адекватный контекст для наших исследований».
Специалисты говорят, что их сегодняшние усилия имеют определяющее значение для поиска эффективного антидота.
«Миссия этих белков — в размножении вирусного генома, — уточняет Крихель. — Когда мы поймем, как функционируют различные элементы белка, из чего они состоят, мы получим бесценную информацию для разработки препаратов, блокирующих их активность».
Синтезированные белки специалисты анализируют с помощью секретного оружия — масс-спектрометра. Молекулярный биолог Кира Шамони рассказывает: «Я могу сначала использовать масс-спектрометр для замеров белка, а затем — для анализа вещества, которое, возможно, связывается с белком. То есть сначала я работаю только с белком, а затем — с соединением. С помощью масс-спектрометра я вижу, насколько увязаны эти элементы, насколько сильна эта связь. И это важно, например, для разработки вакцины или лекарств».
Обычно белки исследуют с помощью кристаллографии или с опорой на электронные микроскопы. Для ученых преимущества масс-спектрометрии очевидны.
Вирусолог Шарлотт Утрехт поясняет: «С помощью масс-спектрометрии мы можем одновременно увидеть различные состояния функционирования белка. Мы можем увидеть, был ли он модифицирован, изменилось ли его состояние из-за этой модификации. В рамках проекта мы пытаемся разработать масс-спектрометрию так, чтобы она не только дополнила методы исследования электронным микроскопом, другие структурные анализы, но и достигла аналогичного уровня разрешения детализации».
«Я могу использовать масс-спектрометр для оценки активного состояния белка, — добавляет Кира Шамони. — Могу видеть вещество в динамике. Другие инструменты структурной биологии не позволяют мне провести наблюдения такого типа».
Проект по изучению белковых структур коронавируса, патронируемый Евросоюзом, стартовал еще до пандемии, в 2018 году. На пике кризиса ученые активизировали работу; уже пройденный путь дал им много важнейших элементов, касающихся понимания механизмов размножения вируса.
«Коронавирусы — крепкий орешек для исследователей из-за особенностей их генетического материала, — добавляет Шарлотт Утрехт. — Мы начали проект с SARS-вирусов и родственных им представителей, а сегодня экстраполируем исследования на ковид-19. Надо сказать, что вирусы семейства очень схожи, так что перед нами фактически — те же задачи».
Проект входит в десятки программ ЕС, нацеленных на борьбу с пандемией. По словам Марии Габриэль, комиссара ЕС по исследованиям и инновациям, все они ориентированы на разработку вакцины, поиск методов в лечения и улучшение диагностики.
«Европа- это 7% населения планеты, при это мы обеспечиваем 20% мировых инвестиций в исследования и инновации и 21% научных работ высокого уровня, — напомнила комиссар Габриэль. — Мы должны продолжать поддерживать усилия Европейского исследовательского совета и так обеспечить рабочее состояние нашему исследовательскому мотору. Его приводит в действие любопытство, подталкивающее ученых предельно расширять границы знаний, чтобы предвидеть, что же нас ждет в будущем».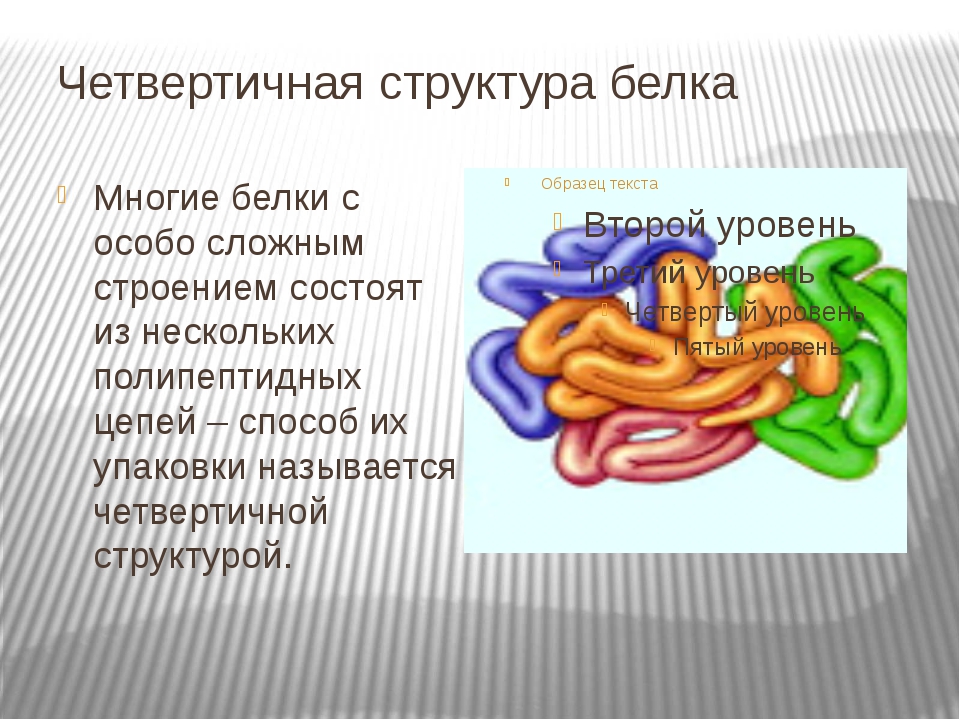
Потребности в науке и исследованиях сегодняшнего и завтрашнего дня будут учтены в готовящейся рамочной программе исследований и инноваций «Горизонт Европа», которая стартует в чрезвычайно сложном контексте. «У нас есть две основные задачи, — рассказывает Мария Габриэль. — Первая — продолжать инвестировать в исследования и инновации; пандемия показала, насколько это важно для поиска решений, которые позволят нашей экономике и обществу лучше подготовиться к следующим кризисам. Вторая задача — сотрудничество и координация. Мы все прекрасно понимаем: вирус не знает границ. Важно сохранить акцент на этом аспекте глобального сотрудничества и координации, чтобы доказать: исследования, патронируемые Евросоюзом, в конечном итоге служат общему благу».
Мы вновь в Гамбурге: исследователи говорят, что их методика не ограничивается коронавирусами или вирусами вообще, она может использоваться применительно к любому биологическому белковому комплексу. Вирусолог Шарлотт Утрехт приводит следующую аналогию: «Возьмем машину.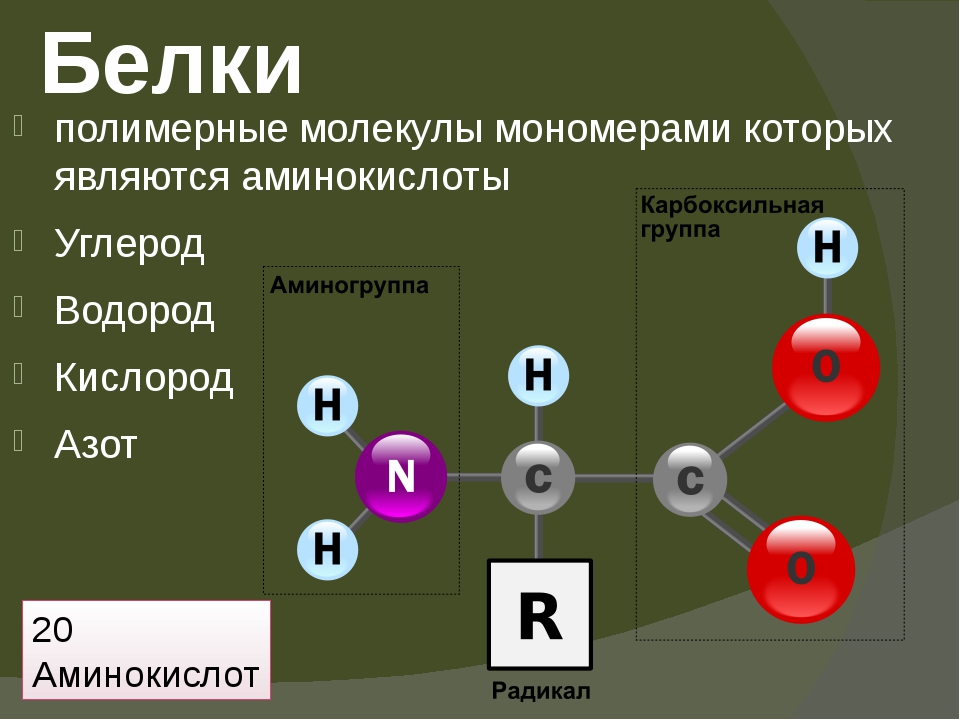 Если мы просто посмотрим на нее снаружи, просто послушаем ее урчание при работе, мы не узнаем, как именно она функционирует. Для этого нам придется рассмотреть двигатель, увидеть его в действии, возможно, разобрать, понять функцию отдельных компонентов. Именно это пытается делать структурная биология: наблюдая за отдельными белками или их соединениями, постигать общую картину, видеть весь клеточный механизм в работе».
Если мы просто посмотрим на нее снаружи, просто послушаем ее урчание при работе, мы не узнаем, как именно она функционирует. Для этого нам придется рассмотреть двигатель, увидеть его в действии, возможно, разобрать, понять функцию отдельных компонентов. Именно это пытается делать структурная биология: наблюдая за отдельными белками или их соединениями, постигать общую картину, видеть весь клеточный механизм в работе».
Исследователи говорят, что довольны имеющимися в их распоряжении технологиями, однако намерены идти дальше и максимально расширить границы знания. Их проект по наблюдению за белками в структуре клетки рассчитан до 2022 года.
3.4 Белки — Биология 2e
Цели обучения
К концу этого раздела вы сможете делать следующее:
- Описывать функции, которые белки выполняют в клетке и тканях
- Обсудить взаимосвязь между аминокислотами и белками
- Объясните четыре уровня белковой организации
- Опишите способы, которыми связаны форма и функция белка
Белки являются одними из наиболее распространенных органических молекул в живых системах и обладают самым разнообразным набором функций среди всех макромолекул.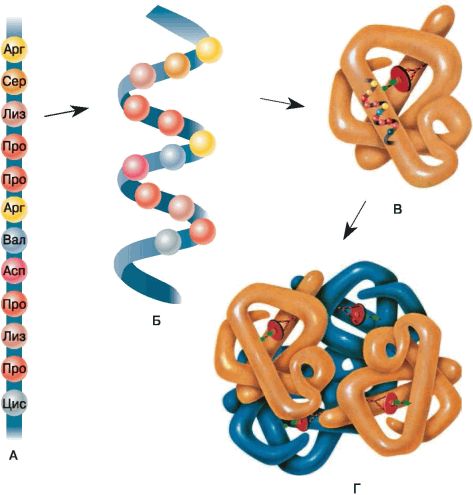 Белки могут быть структурными, регуляторными, сократительными или защитными. Они могут служить для транспортировки, хранения или перепонки; или они могут быть токсинами или ферментами. Каждая клетка в живой системе может содержать тысячи белков, каждый из которых выполняет уникальную функцию. Их структуры, как и их функции, сильно различаются. Однако все они представляют собой аминокислотные полимеры, расположенные в линейной последовательности.
Белки могут быть структурными, регуляторными, сократительными или защитными. Они могут служить для транспортировки, хранения или перепонки; или они могут быть токсинами или ферментами. Каждая клетка в живой системе может содержать тысячи белков, каждый из которых выполняет уникальную функцию. Их структуры, как и их функции, сильно различаются. Однако все они представляют собой аминокислотные полимеры, расположенные в линейной последовательности.
Типы и функции белков
Ферменты, которые вырабатывают живые клетки, являются катализаторами биохимических реакций (например, пищеварения) и обычно представляют собой сложные или конъюгированные белки.Каждый фермент специфичен для субстрата (реагента, который связывается с ферментом), на который он действует. Фермент может помочь в реакциях разложения, перегруппировки или синтеза. Мы называем ферменты, расщепляющие субстраты, катаболическими ферментами. Те, которые строят более сложные молекулы из своих субстратов, являются анаболическими ферментами, а ферменты, которые влияют на скорость реакции, являются каталитическими ферментами. Обратите внимание, что все ферменты увеличивают скорость реакции и, следовательно, являются органическими катализаторами. Примером фермента является амилаза слюны, которая гидролизует свою субстратную амилозу, компонент крахмала.
Обратите внимание, что все ферменты увеличивают скорость реакции и, следовательно, являются органическими катализаторами. Примером фермента является амилаза слюны, которая гидролизует свою субстратную амилозу, компонент крахмала.
Гормоны — это химические сигнальные молекулы, обычно небольшие белки или стероиды, секретируемые эндокринными клетками, которые контролируют или регулируют определенные физиологические процессы, включая рост, развитие, метаболизм и размножение. Например, инсулин — это белковый гормон, который помогает регулировать уровень глюкозы в крови. В таблице 3.1 перечислены основные типы и функции белков.
Типы и функции белков
| Тип | Примеры | Функции |
|---|---|---|
| Пищеварительные ферменты | Амилаза, липаза, пепсин, трипсин | Помощь в пище путем катаболизма питательных веществ в мономерные единицы |
| Транспорт | Гемоглобин, альбумин | Переносит вещества в крови или лимфе по всему телу |
| Структурный | Актин, тубулин, кератин | Создавать различные структуры, такие как цитоскелет |
| Гормоны | Инсулин, тироксин | Координировать деятельность различных систем организма |
| Defense Protect | Иммуноглобулины | тело от чужеродных патогенов |
| Сокращение | Актин, миозин | Эффект сокращения мышц |
| Хранение | Запасные белки бобовых, яичный белок (альбумин) | Обеспечивают питание на ранних этапах развития эмбриона и проростков |
Стол
3. 1
1
Белки имеют разную форму и молекулярную массу. Некоторые белки имеют шаровидную форму; тогда как другие имеют волокнистую природу. Например, гемоглобин — это глобулярный белок, а коллаген, находящийся в нашей коже, — это волокнистый белок. Форма белка имеет решающее значение для его функции, и многие различные типы химических связей поддерживают эту форму. Изменения температуры, pH и воздействие химикатов могут привести к необратимым изменениям формы белка, что приведет к потере функции или денатурации.Все белки содержат разные расположения одних и тех же 20 типов аминокислот. Недавно были открыты две редкие новые аминокислоты (селеноцистеин и пирролизин), и к этому списку могут быть добавлены новые открытия.
Аминокислоты
Аминокислоты — это мономеры, из которых состоят белки. Каждая аминокислота имеет одинаковую фундаментальную структуру, которая состоит из центрального атома углерода или альфа ( α ) углерода, связанного с аминогруппой (NH 2 ), карбоксильной группой (COOH) и с водородом. атом.Каждая аминокислота также имеет другой атом или группу атомов, связанных с центральным атомом, известную как группа R (рис. 3.22).
атом.Каждая аминокислота также имеет другой атом или группу атомов, связанных с центральным атомом, известную как группа R (рис. 3.22).
Фигура
3,22
Аминокислоты имеют центральный асимметричный углерод, к которому присоединены аминогруппа, карбоксильная группа, атом водорода и боковая цепь (R-группа).
Ученые используют название «аминокислота», потому что эти кислоты содержат как аминогруппу, так и карбоксильную кислотную группу в своей основной структуре. Как мы уже упоминали, в белках присутствует 20 распространенных аминокислот.Девять из них являются незаменимыми аминокислотами для человека, потому что человеческий организм не может их производить, и мы получаем их из своего рациона. Для каждой аминокислоты группа R (или боковая цепь) отличается (рис. 3.23).
Визуальное соединение
Визуальное соединение
Фигура
3,23
В белках обычно встречаются 20 общих аминокислот, каждая из которых имеет свою R-группу (вариантную группу), которая определяет его химическую природу.
Какие категории аминокислот вы ожидаете найти на поверхности растворимого белка, а какие — внутри? Какое распределение аминокислот вы ожидаете найти в белке, встроенном в липидный бислой?
Химическая природа боковой цепи определяет природу аминокислоты (то есть, является ли она кислотной, основной, полярной или неполярной).Например, аминокислота глицин имеет атом водорода в качестве группы R. Аминокислоты, такие как валин, метионин и аланин, неполярны или гидрофобны по природе, тогда как аминокислоты, такие как серин, треонин и цистеин, полярны и имеют гидрофильные боковые цепи. Боковые цепи лизина и аргинина заряжены положительно, поэтому эти аминокислоты также являются основными аминокислотами. Пролин имеет группу R, которая связана с аминогруппой, образуя кольцеобразную структуру. Пролин является исключением из стандартной структуры аминокислоты, поскольку его аминогруппа не отделена от боковой цепи (Рисунок 3.23).
Одна заглавная буква или трехбуквенное сокращение обозначают аминокислоты. Например, буква V или трехбуквенный символ val обозначают валин. Так же, как некоторые жирные кислоты необходимы для диеты, некоторые аминокислоты также необходимы. Эти незаменимые аминокислоты для человека включают изолейцин, лейцин и цистеин. Незаменимые аминокислоты относятся к тем, которые необходимы для создания белков в организме, но не к тем, которые организм производит. Какие аминокислоты являются незаменимыми, варьируется от организма к организму.
Например, буква V или трехбуквенный символ val обозначают валин. Так же, как некоторые жирные кислоты необходимы для диеты, некоторые аминокислоты также необходимы. Эти незаменимые аминокислоты для человека включают изолейцин, лейцин и цистеин. Незаменимые аминокислоты относятся к тем, которые необходимы для создания белков в организме, но не к тем, которые организм производит. Какие аминокислоты являются незаменимыми, варьируется от организма к организму.
Последовательность и количество аминокислот в конечном итоге определяют форму, размер и функцию белка. Ковалентная связь или пептидная связь присоединяется к каждой аминокислоте, образуя реакцию дегидратации. Карбоксильная группа одной аминокислоты и аминогруппа входящей аминокислоты объединяются, высвобождая молекулу воды. Полученная связь является пептидной связью (рис. 3.24).
Фигура
3,24
Образование пептидной связи — это реакция синтеза дегидратации. Карбоксильная группа одной аминокислоты связана с аминогруппой входящей аминокислоты. В процессе он высвобождает молекулу воды.
В процессе он высвобождает молекулу воды.
Продукты, образующиеся при таких связях, являются пептидами. Чем больше аминокислот присоединяется к этой растущей цепи, получающаяся цепь представляет собой полипептид. Каждый полипептид имеет свободную аминогруппу на одном конце. Этот конец N-конец или N-конец, а другой конец имеет свободную карбоксильную группу, а также C- или карбоксильный конец. Хотя термины полипептид и белок иногда используются взаимозаменяемо, полипептид технически представляет собой полимер аминокислот, тогда как термин белок используется для полипептида или полипептидов, которые объединились вместе, часто имеют связанные непептидные простетические группы, имеют различную форму. , и имеют уникальную функцию.После синтеза (трансляции) белков большинство белков модифицируются. Они известны как посттрансляционные модификации. Они могут подвергаться расщеплению, фосфорилированию или могут потребовать добавления других химических групп. Только после этих модификаций белок становится полностью функциональным.
Ссылка на обучение
Ссылка на обучение
Просмотрите этапы синтеза белка в этом интерактивном руководстве.
Связь эволюции
Связь эволюции
Эволюционное значение цитохрома c Цитохром c является важным компонентом цепи переноса электронов, частью клеточного дыхания, и обычно он располагается в клеточной органелле, митохондрии.Этот белок имеет простетическую группу гема, и центральный ион гема поочередно восстанавливается и окисляется во время переноса электрона. Поскольку роль этого важного белка в производстве клеточной энергии имеет решающее значение, за миллионы лет он очень мало изменился. Секвенирование белков показало, что существует значительная гомология аминокислотной последовательности цитохрома с среди различных видов. Другими словами, мы можем оценить эволюционное родство, измеряя сходства или различия между последовательностями ДНК или белков различных видов.
Ученые определили, что цитохром с человека содержит 104 аминокислоты. Для каждой молекулы цитохрома с из разных организмов, которые ученые секвенировали на сегодняшний день, 37 из этих аминокислот находятся в одном и том же положении во всех образцах цитохрома с. Это указывает на то, что, возможно, был общий предок. При сравнении последовательностей белков человека и шимпанзе ученые не обнаружили разницы в последовательностях. Когда исследователи сравнили последовательности человека и макаки-резуса, единственное различие было в одной аминокислоте.В другом сравнении секвенирование человека и дрожжей показывает разницу в 44-м положении.
Структура белка
Как мы обсуждали ранее, форма белка имеет решающее значение для его функции. Например, фермент может связываться со специфическим субстратом в активном центре. Если этот активный сайт изменяется из-за локальных изменений или изменений в общей структуре белка, фермент может быть неспособен связываться с субстратом. Чтобы понять, как белок приобретает свою окончательную форму или конформацию, нам необходимо понять четыре уровня структуры белка: первичный, вторичный, третичный и четвертичный.
Чтобы понять, как белок приобретает свою окончательную форму или конформацию, нам необходимо понять четыре уровня структуры белка: первичный, вторичный, третичный и четвертичный.
Первичная структура
Уникальная последовательность аминокислот в полипептидной цепи — это ее первичная структура. Например, гормон поджелудочной железы инсулин имеет две полипептидные цепи, А и В, и они связаны между собой дисульфидными связями. N-концевая аминокислота A-цепи представляет собой глицин; тогда как С-концевой аминокислотой является аспарагин (рис. 3.25). Аминокислотные последовательности в цепях A и B уникальны для инсулина.
Фигура
3,25
Инсулин бычьей сыворотки — это белковый гормон, состоящий из двух пептидных цепей: A (длиной 21 аминокислота) и B (длиной 30 аминокислот).В каждой цепи трехбуквенные сокращения, которые представляют названия аминокислот в порядке их присутствия, указывают на первичную структуру. Аминокислота цистеин (cys) имеет сульфгидрильную (SH) группу в качестве боковой цепи.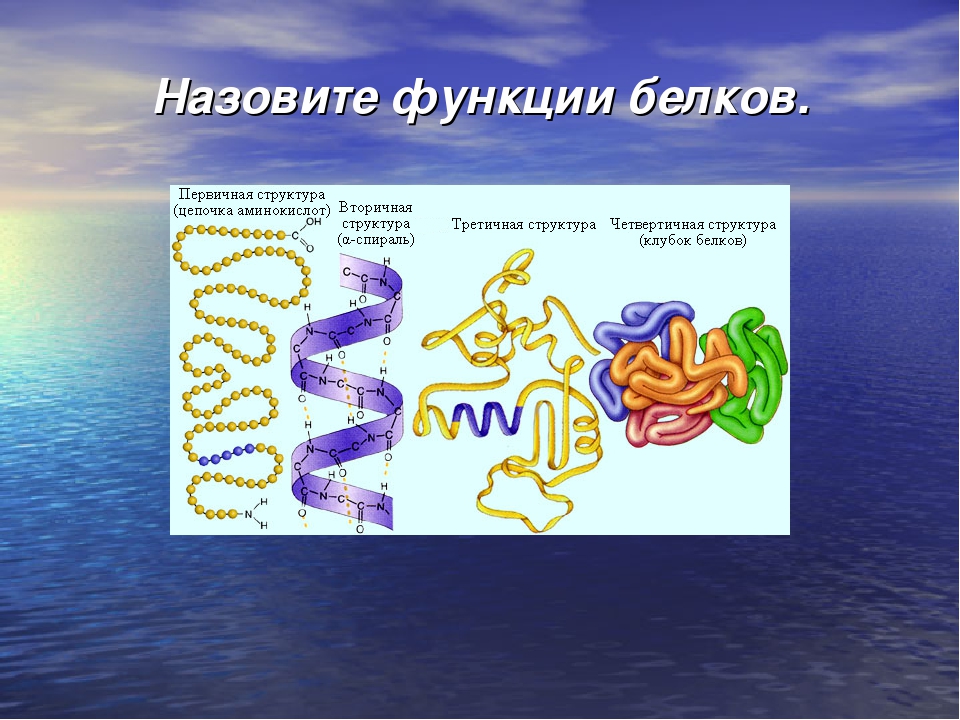 Две сульфгидрильные группы могут реагировать в присутствии кислорода с образованием дисульфидной (S-S) связи. Две дисульфидные связи соединяют цепи A и B вместе, а третья помогает цепи A свернуться в правильную форму. Обратите внимание, что все дисульфидные связи имеют одинаковую длину, но для ясности мы изобразили их разного размера.
Две сульфгидрильные группы могут реагировать в присутствии кислорода с образованием дисульфидной (S-S) связи. Две дисульфидные связи соединяют цепи A и B вместе, а третья помогает цепи A свернуться в правильную форму. Обратите внимание, что все дисульфидные связи имеют одинаковую длину, но для ясности мы изобразили их разного размера.
Ген, кодирующий белок, в конечном итоге определяет уникальную последовательность для каждого белка. Изменение нуклеотидной последовательности кодирующей области гена может привести к добавлению другой аминокислоты к растущей полипептидной цепи, вызывая изменение структуры и функции белка. При серповидно-клеточной анемии цепь гемоглобина β (небольшая часть которой показана на рис. 3.26) имеет единственную аминокислотную замену, вызывающую изменение структуры и функции белка. В частности, валин в цепи β заменяет глутаминовую аминокислоту.Примечательно то, что молекула гемоглобина состоит из двух альфа- и двух бета-цепей, каждая из которых состоит примерно из 150 аминокислот.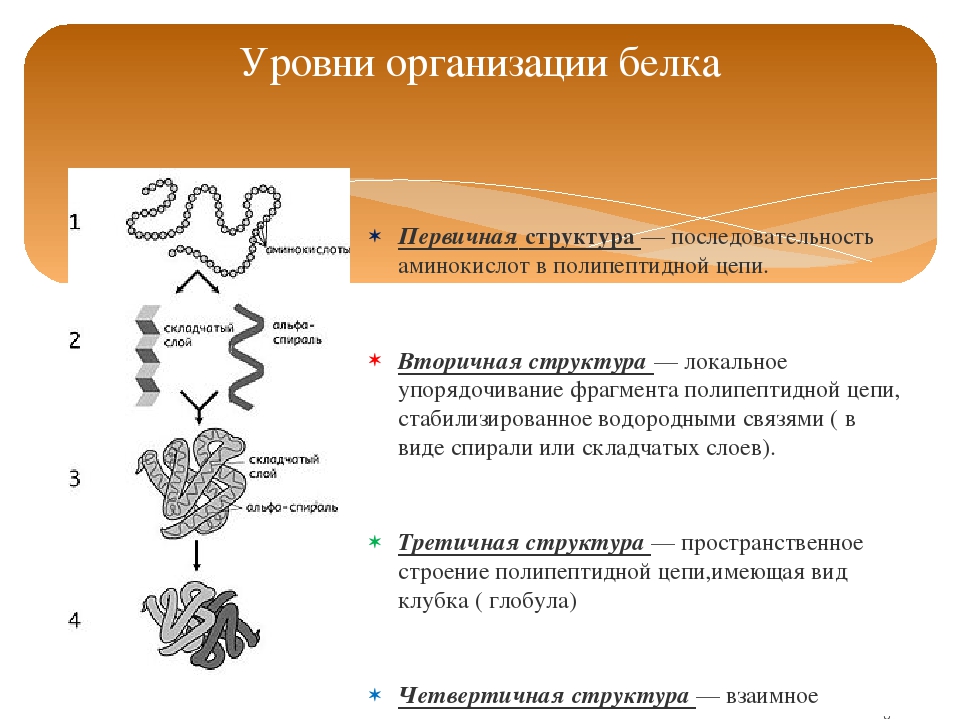 Таким образом, молекула содержит около 600 аминокислот. Структурное различие между нормальной молекулой гемоглобина и молекулой серповидноклеточных клеток — что резко снижает продолжительность жизни — заключается в одной аминокислоте из 600. Что еще более примечательно, так это то, что три нуклеотида каждый кодируют эти 600 аминокислот и одно изменение основания (точечная мутация), 1 из 1800 оснований вызывает мутацию.
Таким образом, молекула содержит около 600 аминокислот. Структурное различие между нормальной молекулой гемоглобина и молекулой серповидноклеточных клеток — что резко снижает продолжительность жизни — заключается в одной аминокислоте из 600. Что еще более примечательно, так это то, что три нуклеотида каждый кодируют эти 600 аминокислот и одно изменение основания (точечная мутация), 1 из 1800 оснований вызывает мутацию.
Фигура
3,26
Из-за этого изменения одной аминокислоты в цепи молекулы гемоглобина образуют длинные волокна, которые искажают двояковогнутую или
дискообразные эритроциты и заставляют их принимать форму полумесяца или «серпа», что закупоривает кровеносные сосуды (рис. 3.27). .его
3.4 • Белки 81
Бета (β) — цепь гемоглобина составляет 147 аминокислот в
длины, но с одной аминокислотной заменой в первичном
последовательность приводит к изменениям во вторичных, третичных и четвертичных
структуры и серповидноклеточная анемия.В нормальном гемоглобине
аминокислота в шестом положении — глутамат. В серповидноклетке
В серповидноклетке
глутамат гемоглобина заменяется валином.
Предоставлено: Рао, А., Тэг, А. Райан, К. и Флетчер, С. Департамент.
биологии Техасского университета A&M.
Из-за этого изменения одной аминокислоты в цепи молекулы гемоглобина образуют длинные волокна, которые искажают двояковогнутые или дискообразные эритроциты и заставляют их принимать серповидную или серповидную форму, которая закупоривает кровеносные сосуды (Рис. 3.27). Это может привести к множеству серьезных проблем со здоровьем, таких как одышка, головокружение, головные боли и боли в животе у людей, страдающих этим заболеванием.Уильям Уоррик Кардозо показал, что серповидно-клеточная анемия является наследственным заболеванием, а это означает, что разница в кодирующей области конкретного гена передается от родителей к детям. Как вы узнаете в отделе генетики, наследование таких признаков определяется комбинацией генов от обоих родителей, и эти очень небольшие различия могут оказывать значительное влияние на организмы.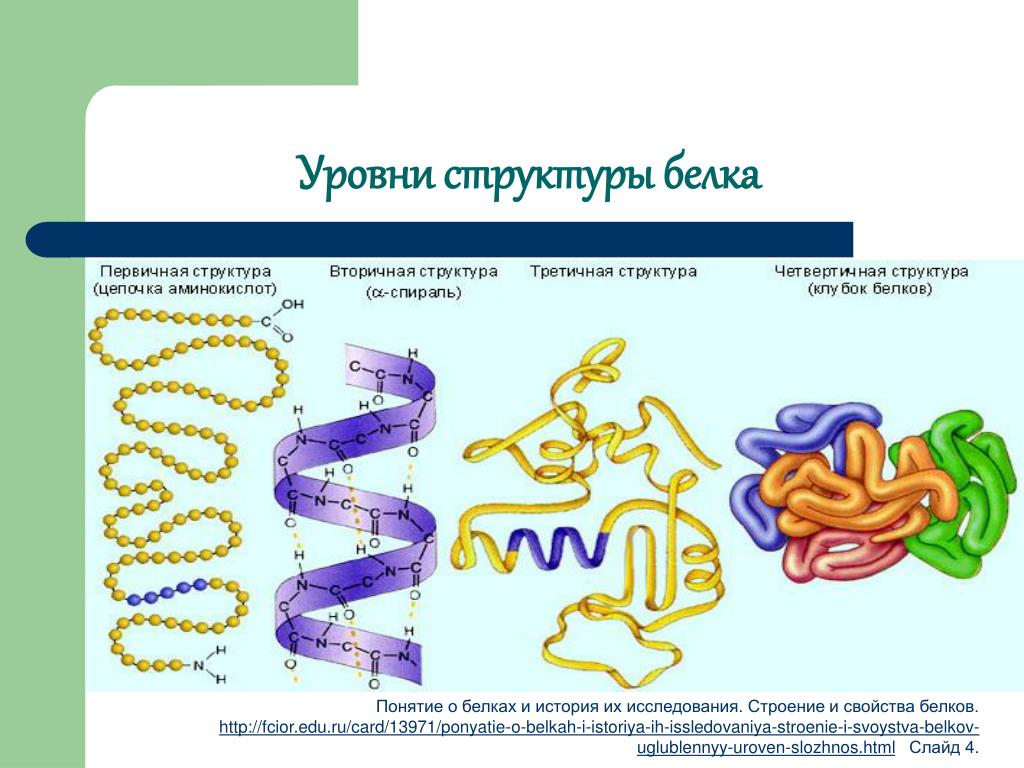
Фигура
3,27
В этом мазке крови, визуализированном при 535-кратном увеличении с помощью светлопольной микроскопии, серповидные клетки имеют форму полумесяца, в то время как нормальные клетки имеют форму диска.(кредит: модификация работы Эда Усмана; данные шкалы от Мэтта Рассела)
Вторичная структура
Локальное сворачивание полипептида в некоторых областях приводит к вторичной структуре белка. Наиболее распространены листовые структуры α, -спираль и β (рис. 3.28). Обе структуры удерживаются в форме водородными связями. Водородные связи образуются между атомом кислорода в карбонильной группе одной аминокислоты и другой аминокислотой, которая находится на четыре аминокислоты дальше по цепи.
Фигура
3,28
Спираль α и складчатый лист β представляют собой вторичные белковые структуры, образующиеся при образовании водородных связей между карбонильным кислородом и
аминоводород в пептидном скелете. Некоторые аминокислоты имеют склонность к образованию α-спирали, в то время как другие способствуют образованию β-складчатых листов. Черный =
Некоторые аминокислоты имеют склонность к образованию α-спирали, в то время как другие способствуют образованию β-складчатых листов. Черный =
углерод, белый = водород, синий = азот и красный = кислород. Предоставлено: Рао, А., Райан, К. Флетчер, С. и Тэг, А., Департамент биологии, Техас, A&M
Университет.
Каждый виток альфа-спирали содержит 3,6 аминокислотных остатка. Группы R полипептида (группы вариантов) выступают из -спиральной цепи . В листе с складками β водородные связи между атомами в основной цепи полипептидной цепи образуют «складки». Группы R прикреплены к атомам углерода и простираются выше и ниже складок складок. Гофрированные сегменты выстраиваются параллельно или антипараллельно друг другу, а водородные связи образуются между частично положительным атомом водорода в аминогруппе и частично отрицательным атомом кислорода в карбонильной группе пептидного остова.Спиральные α и складчатые листовые структуры β присутствуют в большинстве глобулярных и волокнистых белков и играют важную структурную роль.
Третичная структура
Уникальная трехмерная структура полипептида — это его третичная структура (рис. 3.29). Эта структура частично обусловлена химическими взаимодействиями в полипептидной цепи. В первую очередь, взаимодействия между группами R создают сложную трехмерную третичную структуру белка. Природа групп R в задействованных аминокислотах может противодействовать образованию водородных связей, которые мы описали для стандартных вторичных структур.Например, группы R с одинаковыми зарядами отталкиваются друг от друга, а группы с разными зарядами притягиваются друг к другу (ионные связи). Когда происходит сворачивание белка, гидрофобные группы R неполярных аминокислот лежат внутри белка; тогда как гидрофильные группы R расположены снаружи. Ученые также называют первые типы взаимодействия гидрофобными взаимодействиями. Взаимодействие между боковыми цепями цистеина образует дисульфидные связи в присутствии кислорода, единственной ковалентной связи, которая образуется во время сворачивания белка.
Фигура
3,29
Различные химические взаимодействия определяют третичную структуру белков. К ним относятся гидрофобные взаимодействия, ионные связи, водородные связи и дисульфидные связи.
Все эти взаимодействия, слабые и сильные, определяют окончательную трехмерную форму белка. Когда белок теряет свою трехмерную форму, он может больше не функционировать.
Четвертичная структура
В природе некоторые белки образуются из нескольких полипептидов или субъединиц, и взаимодействие этих субъединиц образует четвертичную структуру.Слабые взаимодействия между субъединицами помогают стабилизировать общую структуру. Например, инсулин (глобулярный белок) имеет комбинацию водородных и дисульфидных связей, которые заставляют его в основном слипаться в форму шара. Инсулин начинается как отдельный полипептид и теряет некоторые внутренние последовательности в присутствии посттрансляционной модификации после образования дисульфидных связей, которые удерживают вместе оставшиеся цепи. Шелк (волокнистый белок), однако, имеет складчатую листовую структуру β , которая является результатом водородной связи между различными цепями.
Шелк (волокнистый белок), однако, имеет складчатую листовую структуру β , которая является результатом водородной связи между различными цепями.
Рисунок 3.30 иллюстрирует четыре уровня структуры белка (первичный, вторичный, третичный и четвертичный).
Фигура
3,30
Обратите внимание на четыре уровня белковой структуры на этих иллюстрациях. Предоставлено: Рао, А. Райан, К. и Тэг, А. Департамент биологии, Техас.
Университет A&M.
Денатурация и сворачивание белков
Каждый белок имеет свою уникальную последовательность и форму, которые удерживаются химическими взаимодействиями. Если белок подвержен изменениям температуры, pH или воздействию химических веществ, структура белка может измениться, потеряв свою форму без потери своей первичной последовательности, что ученые называют денатурацией.Денатурация часто обратима, поскольку первичная структура полипептида сохраняется в процессе, если денатурирующий агент удаляется, позволяя белку возобновить свою функцию. Иногда денатурация необратима, что приводит к потере функции. Одним из примеров необратимой денатурации белка является жарка яйца. Белок альбумина в жидком яичном белке денатурирует при помещении на горячую сковороду. Не все белки денатурируют при высоких температурах. Например, бактерии, которые выживают в горячих источниках, содержат белки, которые функционируют при температурах, близких к температуре кипения.Желудок также очень кислый, имеет низкий pH и денатурирует белки как часть процесса пищеварения; однако пищеварительные ферменты желудка сохраняют свою активность в этих условиях.
Иногда денатурация необратима, что приводит к потере функции. Одним из примеров необратимой денатурации белка является жарка яйца. Белок альбумина в жидком яичном белке денатурирует при помещении на горячую сковороду. Не все белки денатурируют при высоких температурах. Например, бактерии, которые выживают в горячих источниках, содержат белки, которые функционируют при температурах, близких к температуре кипения.Желудок также очень кислый, имеет низкий pH и денатурирует белки как часть процесса пищеварения; однако пищеварительные ферменты желудка сохраняют свою активность в этих условиях.
Сворачивание белка имеет решающее значение для его функции. Первоначально ученые думали, что сами белки несут ответственность за процесс сворачивания. Только недавно исследователи обнаружили, что часто они получают помощь в процессе сворачивания от белков-помощников или шаперонов (или шаперонинов), которые связываются с целевым белком во время процесса сворачивания.Они действуют, предотвращая агрегацию полипептидов, которые составляют полную структуру белка, и они отделяются от белка, как только целевой белок сворачивается.
Ссылка на обучение
Ссылка на обучение
Чтобы получить дополнительную информацию о белках, просмотрите этот анимационный ролик под названием «Биомолекулы: белки».
белков — предметный тест GRE: биология
Если вы считаете, что контент, доступный через Веб-сайт (как определено в наших Условиях обслуживания), нарушает
или несколько ваших авторских прав, сообщите нам, отправив письменное уведомление («Уведомление о нарушении»), содержащее
в
информацию, описанную ниже, назначенному ниже агенту.Если репетиторы университета предпримут действия в ответ на
ан
Уведомление о нарушении, оно предпримет добросовестную попытку связаться со стороной, которая предоставила такой контент
средствами самого последнего адреса электронной почты, если таковой имеется, предоставленного такой стороной Varsity Tutors.
Ваше Уведомление о нарушении прав может быть отправлено стороне, предоставившей доступ к контенту, или третьим лицам, таким как
в качестве
ChillingEffects.org.
Обратите внимание, что вы будете нести ответственность за ущерб (включая расходы и гонорары адвокатам), если вы существенно
искажать информацию о том, что продукт или действие нарушает ваши авторские права.Таким образом, если вы не уверены, что контент находится
на Веб-сайте или по ссылке с него нарушает ваши авторские права, вам следует сначала обратиться к юристу.
Чтобы отправить уведомление, выполните следующие действия:
Вы должны включить следующее:
Физическая или электронная подпись правообладателя или лица, уполномоченного действовать от их имени;
Идентификация авторских прав, которые, как утверждается, были нарушены;
Описание характера и точного местонахождения контента, который, по вашему мнению, нарушает ваши авторские права, в \
достаточно подробностей, чтобы позволить репетиторам университетских школ найти и точно идентифицировать этот контент; например нам требуется
а
ссылка на конкретный вопрос (а не только на название вопроса), который содержит содержание и описание
к какой конкретной части вопроса — изображению, ссылке, тексту и т. д. — относится ваша жалоба;
Ваше имя, адрес, номер телефона и адрес электронной почты; а также
Ваше заявление: (а) вы добросовестно полагаете, что использование контента, который, по вашему мнению, нарушает
ваши авторские права не разрешены законом, владельцем авторских прав или его агентом; (б) что все
информация, содержащаяся в вашем Уведомлении о нарушении, является точной, и (c) под страхом наказания за лжесвидетельство, что вы
либо владелец авторских прав, либо лицо, уполномоченное действовать от их имени.
Отправьте жалобу нашему уполномоченному агенту по адресу:
Чарльз Кон
Varsity Tutors LLC
101 S. Hanley Rd, Suite 300
St. Louis, MO 63105
Или заполните форму ниже:
Как работают ДНК и белки
E ВСЕ НА Земля состоит из атомов, большинство из которых плотно прилегают друг к другу в форме минералов.Жизнь находит свое применение в минералах — спросите коралловый риф, — но ее суть заключается в атомах, расположенных как отдельные молекулы, и в способе их взаимодействия.
Послушайте эту историю
Ваш браузер не поддерживает элемент
Больше аудио и подкастов на iOS или Android.
Биологические молекулы различаются по-разному. Во-первых, они действительно могут быть очень большими. Простые неорганические молекулы, из которых состоит воздух и океаны, обычно содержат всего несколько атомов, а часто всего два или три.Многие биологические молекулы содержат тысячи. Некоторые из них содержат миллиарды. Эти молекулы не только большие, но и имеют четкую структуру. Более того, эти структуры можно воссоздать с точностью до атома.
Эти отчетливо похожие на жизнь качества проистекают из того факта, что у биологических молекул есть цели, переданные им в процессе эволюции. Например, жизни нужны молекулы, которые могут катализировать химические реакции, и молекулы, которые могут хранить и передавать генетическую информацию, необходимую для создания этих катализаторов.Этим требованиям удовлетворяют два вида больших молекул: белки, которые выполняют большую часть катализа, а также многое другое, и нуклеиновые кислоты, которые в основном хранят и передают информацию.
Нуклеиновые кислоты и белки являются линейными полимерами; длинные неразветвленные цепочки из похожих на вид компонентов, например бумажных цепочек на детской вечеринке или бусинок на ожерельях. В обоих случаях диапазон составляющих «мономеров» — звеньев бумажной цепочки или бусинок — ограничен. Нуклеиновые кислоты состоят всего из пяти различных мономеров, известных как нуклеотиды; белки обычно состоят из 20 различных разновидностей аминокислот.В обоих случаях сборка цепей происходит по одному звену за раз с использованием определенного типа химической реакции. Нуклеотиды связаны вместе с помощью так называемых сложноэфирных связей; белки, использующие так называемые пептидные связи.
Этот линейный модульный подход означает, что одно и то же оборудование может производить множество разных молекул. Все, что требуется, — это система, которая может катализировать добавление нового мономера к удлиняющейся цепи, способ сообщить этой системе, какой тип мономера добавить следующим, и определенная настойчивость.Типичный человеческий белок состоит примерно из 400 аминокислот; некоторые намного длиннее. Молекулы ДНК, одного из двух типов нуклеиновых кислот жизни, еще намного длиннее. Самая короткая молекула ДНК человека имеет длину около 17 000 нуклеотидов; самые длинные состоят из более 100 метров.
Порядок, в котором появляются эти нуклеотиды, определяет, какая информация хранится в ДНК . Порядок различных аминокислот определяет форму созданного из них белка, контролируя способ сворачивания цепи.Этот процесс может создавать огромное количество форм и возможностей, все из которых зависят только от порядка расположения аминокислот.
Тот факт, что и белки, и ДНК представляют собой упорядоченные модульные цепи, не только отражает легкость, с которой могут быть созданы такие молекулы. Это также то, что делает возможной самую важную вещь, которую каждый должен знать о молекулярной биологии. Порядок нуклеотидов в конкретных последовательностях ДНК — генах — определяет порядок аминокислот в конкретных белках.
В ДНК система, которая катализирует создание нового полимера, представляет собой механизм, называемый полимеразой ДНК , который состоит из ряда белковых субъединиц. Он получает инструкции относительно того, какой вид нуклеотида добавить следующим, из уже существующего фрагмента ДНК , используемого в качестве матрицы.
Четыре разных нуклеотида, используемые в ДНК , различаются по химическому «основанию», которое они несут; основаниями являются аденин ( A ), цитозин ( C ), гуанин ( G ) и тимин ( T ).Одно из открытий, которое привело Фрэнсиса Крика и Джеймса Уотсона к их модели двойной спирали ДНК в 1953 году, заключалось в том, что ДНК всегда содержит то же количество C , что и G, , и A , как T с.
Нуклеотид, несущий гуанин, может свободно соединяться друг с другом на другой цепи ДНК , несущей цитозин; нуклеотиды, несущие аденин и тимин, могут делать то же самое. Это основа двойной спирали, которая состоит из двух молекул ДНК , обернутых друг вокруг друга.Когда у одного есть тимин, у другого есть аденин, а где у одного есть гуанин, у другого есть цитозин; притяжение между этими парными основаниями удерживает две нити вместе. Это также объясняет, почему существует такое же количество G s, как и C s, и A s, как T s.
Конец статьи, в которой была раскрыта эта структура, может похвастаться одним из величайших недооцененных моментов за все время: «От нашего внимания не ускользнуло то, что конкретная пара, которую мы постулировали, сразу же предполагает возможный механизм копирования генетического материала.«Распакуйте двойную спираль, и каждая из двух нитей станет шаблоном для переделки другой. Когда полимераза встречает T на существующей цепи, она добавляет A , и наоборот; он меняет местами G s на C s аналогичным образом.
Копирование ДНК таким образом дает две двойные спирали, каждая из которых содержит одну и ту же последовательность пар оснований, то есть одинаковую информацию. Когда одна из новых двойных спиралей входит в яйцеклетку или сперматозоид, вся записанная на ней информация, «генетический материал» Уотсона и Крика, передается следующему поколению.
Более того, если сообщение изменится — возможно, из-за мутации, при которой случайный кусочек космического излучения превращает A в G — новую последовательность обычно можно скопировать так же хорошо, как и старую. . Тот факт, что степень воспроизводимости фрагмента ДНК не зависит от того, что в нем говорится, позволяет мутациям сохраняться достаточно долго, чтобы эволюция могла найти те, которые приносят пользу. Стивен Беннер, биохимик, суммирует это фундаментальное и жизненно важное свойство DNA с помощью аккуратного акронима COSMIC LOPER : «Способность к поиску пространства мутаций независимо от опасений по поводу потери свойств, необходимых для репликации».«Без способа хранения генома COSMIC — LOPER жизнь в чем-либо подобном ее земной форме не могла бы существовать.
Увеличение массы
Производство белков также требует системы, катализирующей добавление следующего мономера к удлиняющейся цепи, и способа узнать, какой мономер добавить следующим. На этот раз катализатор представляет собой сложный элемент молекулярного механизма, называемый рибосомой, а шпаргалка «какой-мономер-следующий» представляет собой отредактированную копию некоторой информации о последовательности, хранящейся в нуклеотидах генома.
Для этого процесса требуется посредник: РНК , нуклеиновая кислота, очень близкая к ДНК , но не образующая двойных спиралей и имеющая пятое основание, называемое урацилом ( U ), вместо ДНК . тимин. Сначала система, называемая полимеразой РНК , использует последовательность ДНК в качестве матрицы для создания части РНК таким же образом, как полимераза ДНК создает новую цепь при репликации. Затем этот транскрипт преобразуется в так называемый мессенджер РНК (m РНК ).
Это сообщение затем читается рибосомой. Каждая тройка букв в m RNA сообщает механизму трансляции, какую из различных разновидностей аминокислоты добавить следующей. Связь между этими различными триплетами нуклеотидов и аминокислотами, к которым они относятся, является генетическим кодом, поэтому триплеты называются кодонами.
Расшифровка РНК m для создания белка намного сложнее, чем просто сопоставление нового нуклеотида с существующим, как это делают полимеразы ДНК и РНК .В результате рибосома представляет собой гораздо более крупный и сложный элемент молекулярного механизма. В то время как полимераза DNA состоит только из белков, в рибосому также примешано некоторое количество RNA , а также используются другие небольшие части материала, RNA s, для распознавания кодонов и добавления соответствующих аминокислот. По мере того как цепь удлиняется, притяжение и отталкивание между ее различными аминокислотами приводят к тому, что она складывается в требуемую форму (хотя иногда помогают другие белки, называемые шаперонинами).
У человека геном содержит более 21000 последовательностей ДНК , которые описывают белки, и человеческие клетки обладают способностью редактировать РНК , созданную из некоторых из этих последовательностей, для получения ряда различных m RNA s, что позволяет им чтобы производить как минимум в четыре раза больше белков, а может, и в десять раз больше. У бактерии Escherichia coli , наиболее изученной в лабораториях, геном описывает всего 4285 различных белков. Но этого все еще достаточно, чтобы обеспечить все белки, используемые в рибосомах и различных полимеразах, чтобы катализировать все реакции, которые создают другие молекулы, необходимые бактерии, например те, которые составляют ее внешнюю поверхность, и расщеплять пищу. он использует, чтобы обеспечить энергию, которая движет всем остальным.
В успешно растущей кишечной палочке E. coli содержится около 3 млн индивидуальных белковых молекул, что составляет 55% сухой массы организма. Всего 300 000 молекул РНК — в основном РНК по количеству и в основном рибосомная РНК по весу, — которые составляют 20% сухой массы. Миллионы молекул, участвующих в создании мембран и клеточной стенки, которые определяют внешнюю поверхность клетки, составляют 15% сухой массы. Все остальное — совокупность молекул, участвующих в производстве энергии из пищи и ее хранении, компоненты, необходимые для создания более крупных молекул, различные другие губбины и сама ДНК — вместе составляют последние 10%.
Однако стоит помнить, что есть одна последняя жизненно важная молекула, и это та, которая присутствует в наибольшем количестве. Сухая масса клетки, измеренная в лаборатории, составляет всего треть от общей массы, которую она имеет при жизни. Остальные две трети — это старый добрый H 2 O, растворитель, в котором находится все остальное и который позволяет проводить большую часть необходимой химии. Большие сложные молекулы — уникальное и замечательное вещество жизни. Но жизни тоже нужна вода. ■
В этой серии об уровнях жизни
1 Большие молекулы биологии *
2 Клетки и как они питаются
3 Создание органов
4 История жизни
5 Что это вообще вид?
6 Поиск живых планет
Эта статья появилась в кратком разделе «Школы» печатного издания под заголовком «Цепи и реакции». Journal
В отличие от AMP, обилие и функциональная значимость малых белков у прокариот и эукариот в значительной степени игнорировались в прошлом и получили значительное внимание только в последние годы [[1, 21, 26, 35, 93-95]] .Они кодируются smORF [[21, 96-98]], альтернативными ORF (altORF) [[21, 99, 100]], восходящими ORF (uORF) [[21]] или РНК, которые изначально считались некодирующими. , такие как кольцевые РНК [[101, 102]]. Не включаются в растущее число мелких белков пептиды, полученные в результате протеолитического расщепления более крупных белков. Несмотря на их небольшой размер и полное отсутствие детектируемых мотивов, небольшие белки участвуют во многих клеточных процессах.
Малые белки, участвующие в стрессовой реакции и регуляции метаболизма
Поддержание клеточного гомеостаза зависит от скоординированной реакции на изменение условий окружающей среды.У эукариот накопление развернутых белков в эндоплазматическом ретикулуме запускает ответ развернутых белков (UPR) [[103]], который включает подавление синтеза белка и одновременную активацию шаперонов и протеаз. Недавно было показано, что отсутствие малого белка PIGBOS приводит к усилению UPR и апоптозу [[39]] (Рис. 2A). PIGBOS расположен на внешней митохондриальной мембране и взаимодействует с ER белком CLCC1, указывая тем самым, что сайты контакта ER-митохондрии являются критическими для UPR.Другой небольшой белок, регулирующий UPR, — это недавно идентифицированный FORCP, который кодируется тем, что считалось длинной некодирующей РНК (lncRNA) [[104]]. FORCP представляет собой ER-локализованный белок с двумя предполагаемыми трансмембранными доменами. При стрессе ER FORCP ингибирует базальную пролиферацию и индуцирует апоптоз [[104]]. Однако молекулярные механизмы, с помощью которых PIGBOS или FORCP регулируют апоптоз в ответ на стресс, неизвестны. Митохондриальный апоптотический путь дополнительно регулируется цитопротекторным гуманином, полученным из митохондрий.Гуманин был впервые обнаружен в нейронах пациентов, страдающих болезнью Альцгеймера [[105]]. Гуманин кодируется альтернативной ORF в митохондриальном гене 16S рРНК и транслируется как в митохондриальном матриксе, так и в цитозоле, приводя к продуктам из 21 или 24 аминокислот соответственно [[106]]. Гуманин подавляет апоптоз, предотвращая активацию Bax [[107, 108]], и, как таковой, действует как фактор выживания для клеток, подвергшихся стрессовым условиям. Мелкие белки митохондриального происхождения также участвуют в регуляции метаболической активности в стрессовых условиях.Это было показано для MOXI, белка длиной 56 аминокислот, который регулирует β-окисление в мышечных клетках [[109]]. MOXI — это кодируемый ядром белок, связанный с внутренней митохондриальной мембраной. MOXI взаимодействует с митохондриальным трифункциональным белком MTP, который катализирует заключительные стадии окисления длинноцепочечных жирных кислот. Отсутствие МОКСИ приводит к снижению работоспособности, указывая на то, что он необходим для координации β-окисления с потребностью в энергии. У Arabidopsis thaliana цветение контролируется транскрипционным регулятором CONSTANS в зависимости от длины дня.Однако продукция двух небольших белков, mP1a и mP1b, задерживает цветение за счет привязки CONSTANS к ингибирующему белку [[110]].
Маленькие белки участвуют в различных механизмах реакции на стресс. Маленькие белки, участвующие в различных механизмах реакции на стресс, показаны красным цветом. (A) У эукариот небольшие белки PIGBOS и FORCP влияют на UPR, что приводит к усилению регуляции протеаз и шаперонов и снижению синтеза белка, когда клетки сталкиваются со стрессом ER.PIGBOS ингибирует UPR и взаимодействует с ER-белком CLCC1. FORCP вызывает апоптоз в клетках колоректального рака. Небольшой митохондриальный белок MOXI регулирует β-окисление и физическую работоспособность, когда мышечные клетки проявляют высокую потребность в энергии. (B) У прокариот небольшие мембранные белки YohP и Lpt могут приводить к конденсации нуклеоидов с последующим подавлением синтеза белка в неблагоприятных условиях. Было показано, что стабилизирующие мембраны эффекты и гомеостаз железа зависят от небольшого мембранного белка MspA в S.aureus . Активация стрессосом опосредуется небольшим белком Prli42, который, как было показано, играет главную роль в активации SigB в Listeria . Эта активация приводит к изменению транскрипции ~ 150 различных генов.
У прокариот также идентифицируется все большее количество мелких белков, но только некоторым из них приписана биологическая функция [[1, 26, 111, 112]]. Модель E . База данных протеома coli в настоящее время перечисляет около 150 малых белков, но большинство малых белков, вероятно, еще не идентифицировано [[26, 95]].Интересно, что производство многих малых белков увеличивается, когда E. coli сталкивается со стрессовыми условиями, такими как температурный стресс или окислительный стресс, или когда клетки переключаются с богатой среды на минимальную среду или переходят в стационарную фазу [[93]]. Многие из этих индуцированных стрессом небольших белков расположены на внутренней мембране и в основном существуют в виде единственного трансмембранного домена с лишь несколькими остатками, открытыми для цитоплазмы или периплазмы [113]. Помимо гидрофобности, общего знаменателя многих малых мембранных белков E.coli — их положительный чистый заряд, приводящий к высоким значениям pI (например, YohP: 9,3; AzuC: 10,3; YkgR: 8,14), хотя существуют исключения (например, YshB: 4,35). Эти значения pI в целом немного ниже, чем значения pI AMP, а более низкое содержание лизина и аргинина, вероятно, предотвращает дестабилизацию мембраны этими нативными мембранными белками. Единственным исключением является амфифильный AzuC, для которого DBAASP предсказывает антимикробную активность. Гидрофобное ядро AzuC, вероятно, слишком короткое, чтобы полностью покрыть мембрану, и AzuC, вероятно, только периферически прикреплен к мембране [[113, 114]].Хотя избыточное представительство основных остатков также наблюдается в более крупных мембранных белках [[115]], положение положительно заряженных остатков на N- или C-концах в небольших мембранных белках допускает специфические взаимодействия с полярными головными группами фосфолипидов и определяет их топология в мембране [[113, 114]]. Положительные заряды также могут способствовать взаимодействию с другими белками, в частности, когда олигомеризация [[114]] генерирует высоко заряженные кластеры в мембране. Положительный суммарный заряд может дополнительно облегчить взаимодействие с нуклеиновыми кислотами, и это предположение подтверждается данными, показывающими, что экспрессия L.rhamnosus малый мембранный белок Lpt в E. coli приводит к конденсации нуклеоидов [[116]], что также наблюдается, когда эндогенный малый мембранный белок YohP избыточно продуцируется [[114]] (Fig. 2B). Однако неизвестно, наблюдается ли это также на физиологических уровнях YohP. Конденсирующая нуклеоид активность некоторых небольших белков у прокариот также согласуется с данными, демонстрирующими, что некоторые эукариотические AMP, такие как буфорин, проявляют гистоноподобную активность, которая инактивируется заменой положительных остатков [[7]].Конденсация нуклеоидов связана со снижением транскрипции и синтеза белка [[117]], что является консервативной стратегией предотвращения вызванного стрессом повреждения клеток [[118, 119]]. Таким образом, некоторые небольшие мембранные белки могут защищать клетки от стресса за счет снижения транскрипции за счет конденсации нуклеоидов.
Малые мембранные белки также могут формировать толщину и текучесть мембран [[74]], что является важным фактором, определяющим устойчивость к стрессу [[120]]. Было показано, что флюидизаторы мембран индуцируют сети шаперонов, и текучесть мембраны или мембранных доменов может служить в качестве молекулярного переключателя для процессов передачи сигналов стресса [[121]].Таким образом, один из способов действия малых мембранных белков может заключаться в том, что они усиливают цитоплазматическую мембрану как окончательный барьер против внешних стрессоров, как обсуждалось для малого мембранного белка MspA из S . aureus [[122]] (Рис. 2B). Напротив, мембрано-дестабилизирующий эффект наблюдается для некоторых малых белков, принадлежащих к системам токсин-антитоксин [[123]]. Эти системы состоят из стабильного токсина и нестабильного РНК или белкового антитоксина, который противодействует токсичности [[124]].Одним из примеров является система токсин-антитоксин tisB / istR1 . TisB представляет собой небольшой мембранный белок длиной 29 аминокислот, который находится под контролем SOS и вырабатывается в ответ на повреждение ДНК [[125, 126]]. Накопление TisB в мембране вызывает утолщение мембраны [[74]] и деполяризацию мембраны, что впоследствии снижает уровень клеточного АТФ [[127]]. Продукция TisB также связана с образованием клеток-персистеров [[125, 126]], метаболически в значительной степени неактивной субпопуляции высокострессоустойчивых клеток, которые являются основной причиной неэффективности лечения антибиотиками и хронических инфекций [[128]].
У грамположительных бактерий общий стрессовый ответ инициируется стрессосомой, большим цитозольным белковым комплексом более 1 МДа [[129]]. Стрессосома улавливает изменения во внеклеточной среде и активирует фактор транскрипции B (SigB), который, в свою очередь, регулирует более 150 генов в Listeria monocytogenes или B. subtilis [[130]]. Активация σ B зависит от небольшого мембранного белка Prli42, который, как предполагается, привязывает стрессосому к мембране и обеспечивает связь между ощущением стресса и активацией стрессосомы [[131]].
У Klebsiella pneumoniae , патогенной бактерии, вызывающей широкий спектр заболеваний человека, гипервирулентность связана с фенотипом гипермуковязкости (HMV). Недавно было показано, что 58-аминокислотный мембранный белок RmpD необходим для фенотипа HMV и консервативен среди гипервирулентных штаммов Klebsiella [[132]]. На основе предсказаний структуры и последовательности предполагается, что RmpD имеет α-спиральную структуру с топологией N из -C в в мембране.Положительно заряженный С-конец RmpD расположен в цитозоле и консервативен между различными штаммами Klebsiella . Вероятно, он обеспечивает стыковочный сайт для белок-белковых взаимодействий, которые опосредуют вирулентность, но это еще не подтверждено экспериментально [[132]].
Помимо их важности в регулировании реакции на стресс и патогенности, небольшие белки также играют важную роль в регулировании метаболической активности в зависимости от доступности питательных веществ.В фитопатогене Agrobacterium tumefaciens было продемонстрировано, что три небольших белка, богатых аргинином, содержащие домен с неизвестной функцией (DUF1127), контролируют экспрессию нескольких генов в поздней экспоненциальной и стационарной фазе роста. Делеция соответствующих генов приводит к усилению агрегации клеток и образованию биопленок. Подробный анализ показал наибольшее влияние на гены, имеющие отношение к поглощению фосфатов, гомеостазу глицина / серина и дыханию нитратов [[133]].У цианобактерий было обнаружено, что несколько небольших белков участвуют в регуляции метаболизма [[112]]. У Synechocystis sp. Небольшой растворимый белок AcnSP, как было показано, модулирует активность аконитазы и тем самым регулирует поток углерода в цикл TCA [[134]]. Более того, два небольших белка IF7 и IF17 (инактивирующий фактор) регулируют поглощение азота путем инактивации глутаминсинтетазы (GS) в Synechocystis в богатых азотом условиях [[135]]. В отличие от многих других бактерий, которые регулируют активность GS посредством обратимого аденилирования, активность GS у цианобактерий регулируется исключительно за счет количества IF7 и IF17 [[112]].Положительная регуляция активности GS небольшими белками наблюдалась в модельном организме архей Methanosarcina mazei ; здесь было показано, что 23 аминокислоты, содержащие небольшой белок sp26, стимулируют активность GS [[136]].
Активность или стабильность более 40% всех охарактеризованных белков зависят от металлов [[137, 138]], например, более 1000 белков человека содержат цинк-связывающие домены (эталонный протеом UniProt-Human; https: //www.uniprot .org / proteomes /), а небольшие белки играют решающую роль в поддержании гомеостаза металлов. В Escherichia coli они регулируют захват металлов, как показано для MgtS, который регулирует захват Mg 2+ АТФазой P-петли MgtA [[139]] или MntS, который регулирует внутриклеточную концентрацию Mn 2+ [ [140]]. В Salmonella typhimurium малый белок MgtR способствует деградации транспортеров Mg 2+ MgtA и MgtB с помощью мембраносвязанной протеазы FtsH, тогда как малый белок MgtU, по-видимому, защищает MgtB от деградации [[141]].Небольшие белки, такие как консервативные CopZ-подобные белки [[142, 143]], также служат металло-шаперонами и доставляют медь для биогенеза металлопротеинов, одновременно предотвращая индуцированную медью токсичность [[144]]. Продукция CopZ у некоторых видов индуцируется рибосомным сдвигом рамки во время трансляции мРНК copA [[99, 100]], которая кодирует Cu-экспортирующую АТФазу [[144]].
Общая тема, возникающая в результате многочисленных исследований малых белков у прокариот и эукариот, заключается в том, что они, по-видимому, осуществляют свою биологическую активность в основном за счет модуляции активности других белков или белковых комплексов.Хотя маленькие белки, как правило, слишком малы для выполнения ферментативной активности, их небольшой размер помогает им стыковаться с более крупными белками / белковыми комплексами. Более того, учитывая, что трансляция — довольно медленный процесс [[145]], их небольшой размер позволяет гораздо быстрее производить и быстро настраивать метаболизм в зависимости от различных условий роста. По существу, небольшие белки могут действовать как внутриклеточные сигнальные молекулы, аналогичные внеклеточным сигнальным молекулам, таким как цитокины, и они могут регулировать ферментативную активность посредством нековалентных взаимодействий.Это могло бы объяснить, почему многие мелкие белки активируются в стрессовых условиях или изменениях окружающей среды [[146]]. Например, на примере Salmonella было показано, что небольшой мембранный белок PmrR ингибирует ферментативную активность внутреннего мембранного белка LpxT, который отвечает за увеличение отрицательного заряда бактериального липополисахарида и, таким образом, позволяет бактериям воспринимать и модифицировать клетку. поверхность в ответ на окружающую среду [[147]].
Малые белки как дополнительные субъединицы и факторы сборки комплексов мембранных белков
Важность малых белков была первоначально признана в результате наблюдения, что многие белковые комплексы дыхательных цепей содержат небольшие белки в качестве дополнительных субъединиц [[12]].Одним из примеров является бычий комплекс I, который состоит из 45 различных субъединиц, 14 из которых являются небольшими однопроходными мембранными белками [[148]]. Другими примерами являются небольшие субъединицы мембранного белка терминальных оксидаз (комплекс IV) [[149]].
Имеющиеся данные предполагают, по крайней мере, две различные функции малых мембранных белков для комплексов мембранных белков: (а) обеспечение гидрофобной поверхности, которая способствует взаимодействиям спираль-спираль и впоследствии стабилизирует взаимодействия субъединиц в более крупных белковых комплексах, и (б) вставка кофактора в дыхательные комплексы.Стабилизирующий эффект KdpF на транспортер калия KdpFABC был задокументирован и объясняется локализацией KdpF между двумя субъединицами транспортера KdpA и KdpB [[150, 151]] (Fig. 3). Аналогичный стабилизирующий эффект наблюдался для небольшой субъединицы CcoQ cbb 3 -типа цитохромоксидазы [[15, 17]]. cbb 3 цитохромоксидаза типа демонстрирует пониженную сборку и активность в отсутствие CcoQ, что, по-видимому, в первую очередь препятствует стабильному связыванию субъединицы цитохрома c -типа CcoP с каталитическим ядром фермента [[17]] .Малые мембранные белки также стабилизируют дыхательные суперкомплексы [[152, 153]]; это было показано для CcoH, небольшой мембранной субъединицы cbb 3 -типа цитохромоксидазы [[16]], которая необходима для стабилизации суперкомплекса, состоящего из комплекса цитохрома bc 1 , мембраносвязанного c цитохром типа c y, и cbb 3 цитохромоксидаза типа [[154]] (рис. 3). В митохондриях кодируемый днРНК малый белок миторегулин связывает кардиолипин и способствует образованию суперкомплексов, обеспечивая молекулярную связь [[155]].Важно подчеркнуть, что небольшие мембранные белки не только приводят к структурной стабилизации белковых комплексов, но и напрямую влияют на их активность. Это было показано для AcrZ, который стабилизирует и аллостерически регулирует насос оттока нескольких лекарственных препаратов AcrAB [[156, 157]] (рис. 3). Другим примером является CydY, небольшая одномембранная субъединица хинолоксидаз типа bd , которая, как предполагается, регулирует активность фермента, блокируя доступ дикислорода к каталитическому сайту [[158]] (рис.3).
Малые мембранные белки как вспомогательные субъединицы комплексов мембранных белков. Кристаллические структуры комплексов мембранных белков с ассоциированными малыми субъединицами мембранных белков были получены из банка данных белков RCSB с использованием следующих кодов доступа: комплекс AcrBZ (PDB 4CDI), cbb 3 суперкомплекс цитохромоксидазы типа (PDB 6XKW), KdpFABC комплекса (PDB 5MRW) и хинолоксидазы типа bd (PDB 6RX4).Малая субъединица мембранного белка CcoQ цитохромоксидазы типа cbb 3 и периплазматический домен цитохрома c y не разрешены в текущей структуре. Связанные маленькие белки отображаются красным цветом.
Роль малых мембранных белков во встраивании кофакторов была продемонстрирована для CydX, другой малой мембранной субъединицы хинолоксидазы типа E. coli bd [[159]], и CcoS, фактора сборки для cbb 3 Цитохромоксидаза -типа [[19, 160]].Было показано, что CydX необходим для сборки каталитического сайта дигема в bd -оксидазе [[158, 161]], в то время как отсутствие CcoS приводит к образованию цитохрома типа cbb 3 . оксидаза, в которой отсутствует каталитический сайт гема b -Cu B [[18, 19]]. Несмотря на их очевидную схожую роль, CydX и CcoS не обнаруживают значительного сходства последовательностей, и они могут работать, увеличивая стабильность промежуточных соединений сборки и, таким образом, расширяя временное окно для вставки кофактора.Альтернативно, связываясь с белками, содержащими кофактор, они могут предотвращать диссоциацию нековалентно связанных кофакторов, таких как гем b [[18]]. Небольшие белки также обеспечивают ферредоксин-зависимый перенос электронов в фотосинтетический комплекс I у цианобактерий. Здесь положительно заряженный С-конец NdhS, вероятно, улавливает ферредоксин, который, в свою очередь, способствует переносу электронов на каталитическую субъединицу NdhI [[162]].
Последние достижения в структурной биологии мембранных белков — Курс и конференция
Обзор
EMBL стремится делиться научными достижениями и поддерживать научное взаимодействие на протяжении всей пандемии коронавируса.Мы рады сообщить, что конференция станет виртуальной, и приглашаем вас присоединиться к нам онлайн. Виртуальная конференция включает в себя живые выступления приглашенных спикеров, докладчиков с короткими выступлениями, цифровые стендовые доклады и сетевые возможности.
Обзор конференции
Интегральные мембранные белки (IMP) кодируются 20-30% генов в геноме и обеспечивают ключевые функции в клетке. Из-за их гидрофобной природы их структурно и биохимически сложно изучать.Традиционно ИМП экстрагировали с использованием детергентов, которые плохо имитируют липидную мембрану и, как было показано, денатурируют мембранные белки. Однако недавно разработанные подходы липид-нанодисков позволяют проводить структурные и функциональные исследования IMP в не содержащей детергентов, почти нативной мембранной среде. Поэтому центральной темой этого семинара является подчеркивание важности липидов в исследованиях МП и способов их сохранения.
Различные экспериментальные и вычислительные инструменты, в том числе новые стратегии выражения и стабилизации, были разработаны для преодоления проблем, создаваемых множеством MP.Эти технологии позволили получить впечатляющее представление о структуре, динамике и функциях мембранных белков за последние несколько лет. Эта конференция соберет вместе исследователей, разрабатывающих и применяющих эти новые методологии. Помимо форума для обмена последними разработками в этой области, конференция предоставляет отличные возможности для начинающих ученых в этой области изучить все аспекты различных инструментов для изучения структуры и функции мембранных белков.
Темы сессий
- Новые миметики для исследования МП
- Экспрессия и очистка МП
- Биофизика липидных бислоев
- Рентгеновские методы исследования структуры МП
- Связывающие белки для структурных и функциональных исследований МП
- Достижения крио-ЭМ для малых МП
- Крио-ЭМ и криотомография на больших МП-комплексах
- Масс-спектрометрия и одномолекулярный FRET на МП
- Вычислительные методы исследования структуры и динамики МП
Динамики
Основные докладчики
Динамики
Научные организаторы
Организаторы конференций
Мария Мерседес Бакадаре Гойтия
EMBL Heidelberg
Германия
Натали Шнайдер
EMBL Heidelberg
Германия
Программа
Есть что сказать? Напишите об этом в Твиттере! #EMBOMPs
- Виртуальная конференция включает приглашенных докладчиков и докладчиков с короткими выступлениями в прямом эфире с сессиями вопросов и ответов после каждого выступления, а также встречи со спикерами.
- Всем ведущим цифровых плакатов рекомендуется загружать записанные краткие выступления или комментарии со своим цифровым плакатом и планировать видеозвонки на время проведения стендовых сеансов.
- Информация о прямой трансляции и доступ к дискуссионной платформе и цифровым постерам будет предоставлена незадолго до начала мероприятия.
- Доступ к записанным выступлениям будет доступен в течение 2 недели после начала мероприятия.
Все время в программе ниже показано как время в Европа / Берлин.
Чтобы узнать эквивалентный часовой пояс в вашем регионе, введите Берлин, время и дату программы, а также ваш город в Конвертер часовых поясов.
День 1 — понедельник, 29 ноября 2021 г.
| Время | Спикер |
|---|---|
| 11:00 — 11:15 | Вступительное слово |
| Виртуальная сессия 1: Новые миметики для исследования МП Председатель: Сандро Келлер — Университет Граца, Австрия |
|
| 11:15 — 11:45 | Моющие средства с подвесками для структурного исследования мембранных белков Pil Seok Chae — Hanyang Университет, Южная Корея ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 11:45 — 12:00 | Включение SMALP в гигантские однослойные везикулы с использованием капельной микрофлюидики Jorik Waeters ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ПРЯМОГО ПОТОКА |
| 12:00 — 12:30 | CyclAPols: новое поколение амфиполов, сочетающих солюбилизирующие и стабилизирующие свойства, необходимые для исследований мембранных белков Manuela Zoonens — Французский национальный центр научных исследований , Франция ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА | Переход к встрече со спикерами из виртуальной сессии 1 |
| 12:35 — 12:55 | Встреча со спикерами из виртуальной сессии 1 : Пил Сеок Чае, Джорик Ветершут и Мануэла Зооненс |
| 12:55 — 13:35 | Перерыв на обед |
| 13:35 — 14:45 | Виртуальная сессия 1 (A — L) |
| 14:45 — 14:50 | Перерыв |
| Виртуальный сеанс 2: 904 30 Экспрессия и очистка MPs Председатель: Маркус Зигер — Университет Цюриха, Швейцария |
|
| 14:50 — 15:20 | Передовые методы производства высокоуровневого (мембранного) белка в клетках млекопитающих Джонатан Элегхерт — Французский национальный центр научных исследований, Франция ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 15:20 — 15:35 | Экспрессия и очистка липидных флиппаз Р4-АТФазы для функциональных и структурных исследований Гийом Ленуар — Институт интегративной биологии клетки (I2BC) — Университет Париж-Сакле, Франция ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ПОТОКА В РЕЖИМЕ ЖИВОГО ПОТОКА |
| 15:35 — 16:05 | Клетка- свободная экспрессия мембранных белков для структуры и функции Франк Бернхард — Франкфуртский университет, Германия |
| 16:05 — 16:50 | Основной докладчик: Выявление структуры и функции мембранных белков Стивен Слигар — Университет Иллинойса в Урбана-Шампейн, США 904 СПРОС ПОСЛЕ ЖИВОГО ПОТОКА |
| 16:50 — 17:05 | Перерыв |
| Virtual Сессия 3: Биофизика липидных бислоев Кристина Кафедра университета: из Штутгарта, Германия |
|
| 17:05 — 17:35 | Использование стабилизированных полимером нанодисков для определения структуры и функции мембранного белка Карен Эдлер — Университет Бата, Великобритания В НАЛИЧИИ СПРОС ПОСЛЕ ПРЯМОГО ПОТОКА |
| 17:35 — 17:50 | Evi Обоснование механизма «ловушки и переворота» в протон-зависимом переносчике липидов Камило Перес — Базельский университет, Швейцария ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 17:50 — 18:20 | Асимметрия мембраны: Почему мембрана белков может беспокоить Георг Пабст — Университет Граца, Австрия ДОСТУПНО ПО СПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| Переход для встречи со спикерами из Виртуальная сессия 2 и 3 |
|
| 18:25 — 18:45 | Встреча со спикерами из Виртуальная сессия 2 и 3 : Джонатан Элегхерт, Гийом Ленуар, Франк Бернхард, Стивен Слигар, Карен Эдлер, Камило Перес и Георг Пабст |
| 18:45 — 19:15 | Социальные сети программа: Смеситель Virtual Bar |
| 19:15 | Конечный день 1 |
Цифровые плакаты и предварительно записанные выступления, а также каналы для обсуждения будут доступны зарегистрированным участникам на протяжении всего виртуального собрания.
День 2 — вторник, 30 ноября 2021 г.
| Время | Спикер |
|---|---|
| 13:00 — 13:45 | Социальная программа: Virtual Speed Networking |
| Виртуальная сессия 4: Рентгеновские методы для структурных исследований МП Председатель: Томас Шнайдер — EMBL, Гамбург, Германия, |
|
| 14:00 — 14:30 | Структурные измерения от фемтосекунд до миллисекунд Биология в SLS и SwissFEL Jörg Standfuss — Институт Пауля Шеррера, Швейцария ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 14:30 — 14:45 | Анализ ингибирования захвата глицина структура человеческого GlyT1, определенная с помощью серийной синхротронной кристаллографии Azadeh Shahsavar — Орхусский университет, Денмар k ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 14:45 — 15:00 | Структурное исследование перенесенного белка внешней мембраны Bdellovibrio bacteriovorus Ребекка Парр — Университет 410, Великобритания ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 15:00 — 15:15 | Перерыв |
| Виртуальная сессия 5: Связывающие белки для структурных и функциональных исследований МП Председатель: Кристиан Löw — EMBL Hamburg, Германия |
|
| 15:15 — 15:45 | Конформационный захват мембранного белка с помощью sybody Markus Seeger — Цюрихский университет, Швейцария ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 15:45 — 16:00 | Разработка универсального нанотела y-связывающий Fab-модуль для крио-ЭМ-исследований мембранных белков на основе реперных точек Joël Bloch — ETH Zurich, Switzerland ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 16:00 — 16:30 | N-концевой трансмембранно-спиральный эпитопный тег для рентгеновской кристаллографии и электронной микроскопии малых мембранных белков Randy Stockbridge — Мичиганский университет, США ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 16:30 — 16 : 35 | Переход для встречи со спикерами из Виртуальная сессия 4 и 5 |
| 16:35 — 16:55 | Встреча со спикерами из Виртуальная сессия 4 и 5: Йорг Стандфус, Азаде Шахсавар, Ребекка Парр, Маркус Сигер, Джоэль Блох и Рэнди Стокбридж |
| 16:55 — 18:05 | Виртуальная сессия постеров 2 (M — Z) |
| 18:05 — 18:10 | Перерыв |
| Виртуальная сессия 6: Достижения крио-ЭМ для малых MPI I Председатель: Jörg Standfuss — Институт Пола Шерера, Швейцария |
|
| 18:10 — 18:55 | Основной докладчик: Роль динамики белка в активации рецепторов, связанных с G-белком Брайан Кобилка — Стэнфордский университет, США |
| 18:55 — 19:25 | MicroED: концепция, практика и будущие возможности Тамир Гонен — Калифорнийский университет, Лос-Анджелес, США |
| 19:25 — 19:40 | Анализ двухслойный механизм опрокидывания фолат-специфичного транспортера ECF с помощью cryo-EM Chancievan Thangaratnarajah — Университет Гронингена, Нидерланды ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ LI VE STREAM |
| 19:40 — 19:55 | Структурные снимки человеческого PepT1 и PepT2 раскрывают механистическое понимание субстрата и транспорта лекарств через эпителиальные мембраны Максим Киллер — EMBL Гамбург, Германия В НАЛИЧИИ СПРОС ПОСЛЕ ЖИВОГО ПОТОКА |
| 19:55 — 20:25 | Мониторинг липидов в крио-ЭМ структурах мембранного транспортера и каналов Christine Ziegler — Университет Регенсбурга, Германия В НАЛИЧИИ СПРОС ПОСЛЕ ЖИВОГО ПОТОКА |
| 20:25 — 20:35 | Перерыв |
| Виртуальная сессия 7: Развитие крио-ЭМ для малых MPs II Председатель: Кристина Паулино — Университет Гронингена, Нидерланды |
|
| 20:35 — 21:05 | Простой механизм чередующегося доступа в каналах мочевины / амида, обнаруженный крио-ЭМ высокого разрешения Цзянсен Цзян — Национальный институт сердца, легких и крови, США ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 21 : 05 — 21:20 | Активация рецептора меланокортина 4 — путь к лекарствам от ожирения Николас Хейдер — Шарите Берлин, Германия ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 21:20 — 21:25 | Переход, чтобы встретиться со спикерами из Виртуальная сессия 6 и 7 |
| 21:25 — 21:45 | Встреча со спикерами из виртуальной сессии 6 и 7: Брайан Кобилка, Тамир Гонен, Чансиван Тангаратнараджа, Максим Киллер, Кристин Зиглер, Цзянсен Цзян и Николас Хейдер |
| 21:45 — 22:15 | 9043 0 Социальная программа: Концерт в гостиной с ленивым мехом |
| 22:15 | Конечный день 2 |
Цифровые плакаты и предварительно записанные выступления, а также каналы для обсуждения будут доступны зарегистрированным участникам на протяжении всего виртуального собрания.
День 3 — среда, 1 декабря 2021 г.
| Время | Спикер |
|---|---|
| Виртуальная сессия 8: Крио-ЭМ и крио-томография на больших МП-комплексах Председатель: Панайотис Л. Кастритис — Университет Мартина Лютера Галле-Виттенберг, Германия |
|
| 14:00 — 14:30 | cryo-CLEM ямок, покрытых клатрином, в интактных клетках Ори Авиноам — Институт науки Вейцмана, Израиль ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 14:30 — 14:45 | Мелкомолекулярная модуляция Slo-канала дрозофилы, выявленная крио-ЭМ Tobias Raisch — Институт молекулярной физиологии Макса Планка, Германия |
| 14:45 — 15:00 | Фазовое разделение мембранных белков при сборке органелл и слиянии мембран Al exander von Appen — Общество Макса Планка, Германия ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 15:00 — 15:15 | Перерыв |
| Виртуальная сессия 9: Масс-спектрометрия и одиночные молекулы FRET на MP Председатель: Кристина Циглер — Университет Регенсбурга, Германия |
|
| 15:15 — 15:45 | Структурная динамика сворачивания мембран-белков из одномолекулярного FRET Георг Крайнер — Кембриджский университет, Великобритания ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 15:45 — 16:00 | Липосомы как носители мембранно-ассоциированных белков и пептидов для масс-спектрометрического анализа Мелисса Фрик — Междисциплинарный исследовательский центр HALOmem, Университет Мартина Лютера Галле-Виттенберг, Германия В НАЛИЧИИ ПО ТРЕБОВАНИЮ ПОСЛЕ ЖИВОГО ПОТОКА |
| 16:00 — 16:15 | Конформационная связь через бислой мембраны полноразмерного рецептора эпидермального фактора роста в нанодисках Shwetha Srinivasan — Массачусетский технологический институт , США ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ЖИВОГО ПОТОКА |
| 16:15 — 16:45 | Липидный обмен: соединяем точки Анн-Клод Гэвин — Женевский университет, Швейцария |
| 16 : 45 — 16:50 | Переход, чтобы встретиться со спикерами из Виртуальная сессия 8 и 9 |
| 16:50 — 17:10 | Познакомьтесь с докладчиками из виртуальной сессии 8 и 9: Ори Авиноам, Тобиас Райш, Александр фон Аппен, Георг Крайнер, Мелисса Фрик, Швета Сринивасан и Анн-Клод Гэвин |
| 1 7:10 — 17:40 | Перерыв |
| Виртуальная сессия 10: Вычислительные методы исследования структуры и динамики MP Председатель: Георг Крайнер — Кембриджский университет, Соединенное Королевство |
|
| 17:40 — 18:10 | Мультимасштабное моделирование молекулярной динамики сложных биомембран Кристина Плугацкова — Штутгартский университет, Германия |
| 18:10 — 18:25 | Практически законченная структурная модель каркаса комплекса ядерных пор человека Агнешка Обарска-Косинская — Институт биофизики Макса Планка, Германия В НАЛИЧИИ ПО ЗАПРОСУ ПО ЗАПРОСУ |
| 18:25 — 18:55 | Молекулярная ансамбль свободные энергетические ландшафты мембран и белков Jochen Hub — Saarland University, Германия ДОСТУПНО ПО ЗАПРОСУ ПОСЛЕ ПРЯМОГО ПОТОКА |
| 18:55 — 19:00 | Переход к встрече с докладчиками из виртуальной сессии 10 |
| 19:00 — 19:20 | Знакомство с докладчиками из Virtual Session 10 : Кристина Плугацкова, Агнешка Обарска-Косинская и Jochen Hub |
| 19:20 — 19:25 | Переход к панельной дискуссии |
| 19:25 — 20:10 | Панельная дискуссия : Текущие проблемы и будущее направление структурной биологии мембранных белков Председатели: Сандро Келлер — Университет Граца, Австрия Кристина Паулино — Университет Гронингена, Нидерланды |
| 20:10 — 20: 20 | Заключительные замечания |
| 20:20 | Конечный день 3 |
Цифровые плакаты и предварительно записанные выступления, а также каналы для обсуждения будут доступны зарегистрированным участникам на протяжении всего виртуального собрания.
Практическая информация
Регистрационный взнос и подача тезисов
Регистрационный взнос (включает доступ ко всем выступлениям, цифровым стендовым докладам и групповым онлайн-обсуждениям и помогает нам покрыть наши расходы на проведение мероприятия. Для получения дополнительной информации см. Страницу часто задаваемых вопросов):
| Академия | 190 евро |
| Аспирант | 140 евро |
| Промышленность | 240 евро |
| Персонал EMBL 94 | Доступ к внутренней сети |
| Аминокислоты — это мономеры, из которых состоят белки. Генерал структура аминокислоты как: , где NH 2 представляет собой аминогруппу, Реакция конденсации между
Функциональный белок может содержать один или несколько Роль водородных связей, ионная Белки выполняют множество функций во всех живых биуретовый тест на белки. Студенты должны быть |
AT f Студенты могли использовать и интерпретировать результаты биурета AT g Студенты могут использовать хроматографию с известными стандартными растворами, чтобы |
AlphaFold: решение грандиозной проблемы биологии 50-летней давности
Мы также увидели признаки того, что прогнозирование структуры белка может быть полезным в будущих усилиях по реагированию на пандемию, как один из многих инструментов, разработанных научным сообществом.Ранее в этом году мы предсказали несколько белковых структур вируса SARS-CoV-2, включая ORF3a, структура которого ранее была неизвестна. В CASP14 мы предсказали структуру другого белка коронавируса, ORF8. Впечатляюще быстрая работа экспериментаторов подтвердила структуры как ORF3a, так и ORF8. Несмотря на их сложный характер и наличие очень небольшого количества связанных последовательностей, мы достигли высокой степени точности обоих наших прогнозов по сравнению с их экспериментально определенными структурами.
Мы не только ускоряем понимание известных болезней, но и рады возможности этих методов исследовать сотни миллионов белков, для которых у нас в настоящее время нет моделей, — обширную территорию с неизвестной биологией. Поскольку ДНК определяет аминокислотные последовательности, которые составляют структуры белков, революция в геномике сделала возможным считывание последовательностей белков из естественного мира в массовом масштабе — с 180 миллионами последовательностей белков и подсчетом в базе данных Universal Protein (UniProt).Напротив, учитывая экспериментальную работу, необходимую для перехода от последовательности к структуре, только около 170 000 белковых структур находятся в банке данных о белках (PDB). Среди неопределенных белков могут быть некоторые с новыми интересными функциями, и — так же, как телескоп помогает нам заглянуть глубже в неизвестную вселенную — такие методы, как AlphaFold, могут помочь нам найти их.
Открывая новые возможности
AlphaFold — одно из наших самых значительных достижений на сегодняшний день, но, как и во всех других научных исследованиях, есть еще много вопросов, на которые нужно ответить.Не всякая структура, которую мы прогнозируем, будет идеальной. Еще многое предстоит узнать, в том числе о том, как несколько белков образуют комплексы, как они взаимодействуют с ДНК, РНК или небольшими молекулами, и как мы можем определить точное расположение всех боковых цепей аминокислот. В сотрудничестве с другими можно многое узнать о том, как лучше всего использовать эти научные открытия при разработке новых лекарств, о способах управления окружающей средой и многом другом.
Для всех нас, работающих над вычислительными методами и методами машинного обучения в науке, такие системы, как AlphaFold, демонстрируют потрясающий потенциал ИИ как инструмента для фундаментальных открытий.Подобно тому, как 50 лет назад Анфинсен поставил задачу, выходящую за пределы возможностей науки в то время, многие аспекты нашей Вселенной остаются неизвестными. Объявленный сегодня прогресс вселяет в нас уверенность в том, что ИИ станет одним из самых полезных инструментов человечества в расширении границ научных знаний, и мы с нетерпением ждем впереди многих лет упорной работы и открытий!
Пока мы не опубликуем статью об этой работе, пожалуйста, цитируйте:
Высокоточное прогнозирование структуры белка с использованием глубокого обучения
Джон Джампер, Ричард Эванс, Александр Прицель, Тим Грин, Майкл Фигурнов, Кэтрин Туньясувунакул, Олаф Роннебергер, Расс Бейтс, Огюстин Жидек, Алекс Бриджленд, Клеменс Мейер, Саймон А.А. Коль, Анна Потапенко, Эндрю Дж. Баллард, Эндрю Ромера, Бернардино -Паредес, Станислав Николов, Ришуб Джейн, Йонас Адлер, Тревор Бэк, Стиг Петерсен, Дэвид Рейман, Мартин Штайнеггер, Михалина Пачольска, Дэвид Сильвер, Ориол Виньялс, Эндрю У.