ДИССИМИЛЯЦИЯ И АССИМИЛЯЦИЯ — это… Что такое ДИССИМИЛЯЦИЯ И АССИМИЛЯЦИЯ?
- ДИССИМИЛЯЦИЯ И АССИМИЛЯЦИЯ
-
- ДИССИМИЛЯЦИЯ И АССИМИЛЯЦИЯ
-
(от лат. dissimilis – несходный и assimilis – сходный) – взаимно противоположные процессы, обеспечивающие в единстве непрерывный процесс жизнедеятельности живых организмов; протекают в организме непрерывно, одновременно, в тесной взаимосвязи и составляют две стороны единого процесса обмена веществ. Д. и а. образуют сложную систему, состоящую из цепи взаимосвязанных биохимич. реакций, каждая из к-рых в отдельности является только химической, но к-рые в единстве составляют целое, обладающее биологич. природой. Противоречие Д. и а. определяет динамич. равновесие живого тела. Как открытая система (см. Жизнь), оно должно, постоянно приобретая, столь же непрерывно тратить приобретенную энергию, так, чтобы не увеличивалась энтропия.
Д и с с и м и л я ц и я – процесс расщепления в живом организме органич. веществ на более простые соединения – ведет к освобождению энергии, необходимой для всех процессов жизнедеятельности организма. А с с и м и л я ц и я – процесс усвоения органич. веществ, поступающих в организм, и уподобления их органич. веществам, свойственным данному организму, идет с использованием энергии, высвобождающейся при процессах диссимиляции. При этом образуются (синтезируются) соединения, обладающие высокой энергией (макроэргические), к-рые становятся источником энергии, освобождающейся при диссимиляции.
Диссимиляция поступающих в организм питательных веществ, в основном белков, жиров и углеводов, начинается с ферментативного расщепления их на более простые соединения – промежуточные продукты обмена веществ (пептиды, аминокислоты, глицерин, жирные кислоты, моносахариды), из к-рых организм синтезирует (ассимилирует) органич. соединения, необходимые для его жизнедеятельности. Все процессы Д. и а. в организме протекают как единое целое. См. Обмен веществ, Жизнь и лит. при этих статьях.
См. Обмен веществ, Жизнь и лит. при этих статьях.
И. Вайсфельд. Москва.
Философская Энциклопедия. В 5-х т. — М.: Советская энциклопедия.
Под редакцией Ф. В. Константинова.
1960—1970.
.
- ДИСПОЗИЦИОНАЛЬНЫЙ ПРЕДИКАТ
- ДИСТРИБУТИВНОСТИ ЗАКОН
Полезное
Смотреть что такое «ДИССИМИЛЯЦИЯ И АССИМИЛЯЦИЯ» в других словарях:
-
АССИМИЛЯЦИЯ — (лат. assimilatio, от assimilare уподоблять). Уравнение, уподобление, напр., в фонетике уподобление соседних звуков один другому; в физиологии уподобление веществ, поглощенных животным, веществам собственного тела. Словарь иностранных слов,… … Словарь иностранных слов русского языка
-
ДИССИМИЛЯЦИЯ — [лат.
 dissimilatio расподобление] лингв. изменение, которое разрушает сходство и подобие звуков в слове. Словарь иностранных слов. Комлев Н.Г., 2006. диссимиляция (лат. dissimilatio расподобление) 1) иначе катаболизм распад сложных органических… … Словарь иностранных слов русского языка
dissimilatio расподобление] лингв. изменение, которое разрушает сходство и подобие звуков в слове. Словарь иностранных слов. Комлев Н.Г., 2006. диссимиляция (лат. dissimilatio расподобление) 1) иначе катаболизм распад сложных органических… … Словарь иностранных слов русского языка -
Ассимиляция — (от лат. assimilatio воспроизведение), анаболизм, процесс, в ходе которого из более простых веществ синтезируются более сложные (полисахариды, нуклеиновые кислоты, белки и др.), аналогичные компонентам этого организма и необходимые для его… … Экологический словарь
-
Ассимиляция — Термин ассимиляция (лат. assimilatio уподобление) употребляется в нескольких областях знания: Ассимиляция (биология) совокупность процессов синтеза в живом организме. Ассимиляция (лингвистика) уподобление артикуляции одного … Википедия
-
диссимиляция — (лат. dissimilatio расподобление). Замена одного из двух одинаковых или сходных звуков другим, менее сходным в отношении артикуляции с тем, который остался без изменения.
 Подобно ассимиляции, диссимиляция может быть прогрессивной и регрессивной.… … Словарь лингвистических терминов
Подобно ассимиляции, диссимиляция может быть прогрессивной и регрессивной.… … Словарь лингвистических терминов -
Диссимиляция — I ж. Изменение, нарушающее сходство, подобие одинаковых или сходных звуков в слове или в соседних словах; расподобление (в лингвистике). Ant: ассимиляция I II ж. Распад в организме сложных органических веществ, клеток, тканей и т.п. (в биологии) … Современный толковый словарь русского языка Ефремовой
-
Диссимиляция — I ж. Изменение, нарушающее сходство, подобие одинаковых или сходных звуков в слове или в соседних словах; расподобление (в лингвистике). Ant: ассимиляция I II ж. Распад в организме сложных органических веществ, клеток, тканей и т.п. (в биологии) … Современный толковый словарь русского языка Ефремовой
-
ассимиляция — (лат. assimilatio уподобление). Уподобление одного звука другому в артикуляционном и акустическом отношениях (ср.: диссимиляция). Ассимиляция возникает у гласных с гласными, у согласных с согласными … Словарь лингвистических терминов
-
Ассимиляция — I Ассимиляция (от лат.
 assimilatio) уподобление, слияние, усвоение. II Ассимиляция (этнографич.) слияние одного народа с другим с утратой одним из них своего языка, культуры, национального самосознания. Во многих странах в… … Большая советская энциклопедия
assimilatio) уподобление, слияние, усвоение. II Ассимиляция (этнографич.) слияние одного народа с другим с утратой одним из них своего языка, культуры, национального самосознания. Во многих странах в… … Большая советская энциклопедия -
Диссимиляция — I Диссимиляция (от лат. dissimilis несходный) в биологии, противоположная ассимиляции (См. Ассимиляция) сторона обмена веществ (См. Обмен веществ), заключающаяся в разрушении органических соединений с превращением белков, нуклеиновых… … Большая советская энциклопедия
чай для обмена веществ и похудения
чай для обмена веществ и похудения
чай для обмена веществ и похудения
>>>ПЕРЕЙТИ НА ОФИЦИАЛЬНЫЙ САЙТ >>>
Что такое чай для обмена веществ и похудения?
Пурпурный чай- моя голубая мечта:) Давно слышала о ней, но купить этот чай в магазинах городах невозможно, пришлось делать заказ через интернет. Заварила сразу же после получения посылки, так не терпелось попробовать это чудо. Упаковка огромная, цветочки даже в сушеном виде остаются голубыми, а при заваривании придают напитку невероятный изумрудный цвет!! Чай поразил меня своей красотой, к тому же он содержит много витаминов, поддерживает иммунитет, способствует похудению. Имеет специфический вкус на любителя (не горький, не сладкий, не кислый, и этот уже хорошо:), добавляю в него молоко, которое сглаживает аромат и получаю удивительный «молочный» цвет:)
Заварила сразу же после получения посылки, так не терпелось попробовать это чудо. Упаковка огромная, цветочки даже в сушеном виде остаются голубыми, а при заваривании придают напитку невероятный изумрудный цвет!! Чай поразил меня своей красотой, к тому же он содержит много витаминов, поддерживает иммунитет, способствует похудению. Имеет специфический вкус на любителя (не горький, не сладкий, не кислый, и этот уже хорошо:), добавляю в него молоко, которое сглаживает аромат и получаю удивительный «молочный» цвет:)
Эффект от применения чай для обмена веществ и похудения
С пурпурным чаем Чай Ши я сбросила уже 7 кг за месяц – это для меня прекрасный результат. Пью его ежедневно 3-4 чашки в день. На работе все удивляются красивому цвету моего чая, но на вкус он мне не очень нравится.
Мнение специалиста
Очень красивый чай, заваривание как медитация! Первый раз когда попробовала на вкус как сено, но постепенно чувствуешь как раскрывается вкус, появляется легкая кислинка. После месяца приема пурпурного чая чувствую себя легче, бодрее, на весах минут 3 кг, но похудение для меня не было самоцелью. От чая я осталась в вострге.
После месяца приема пурпурного чая чувствую себя легче, бодрее, на весах минут 3 кг, но похудение для меня не было самоцелью. От чая я осталась в вострге.
Как заказать
Для того чтобы оформить заказ чай для обмена веществ и похудения необходимо оставить свои контактные данные на сайте. В течение 15 минут оператор свяжется с вами. Уточнит у вас все детали и мы отправим ваш заказ. Через 3-10 дней вы получите посылку и оплатите её при получении.
Отзывы покупателей:
Ника
Чанг-Шу — это просто нереальный жиросжигатель, который не просто убирает лишние килограммы, а превращает их в энергию. Его помощь оказалась довольной для того, чтобы сделать из меня энерджайзера, моя работоспособность становится легендарной, а ещё совсем чуть-чуть в недавнее время я с утра уже чувствовала глубокую усталость. Мне ведь было совершенно некогда заниматься физическими упражнениями и ограничивать себя в еде я не хотела, так что, когда я потеряла пять килограммов, то расценила это как приятное сопутствие. В планах удвоить потери, так что меня ожидает продолжение.
В планах удвоить потери, так что меня ожидает продолжение.
Ия
При заказе «Чанг Шу» я не ждала чуда, но описание и обещанное действие конечно привлекло, в итоге пью его уже 2 недели. Цвет насыщенно синий, вкус травяной, немного сладит (пью без сахара). Цвет лица выравнялся, высыпания на лице уменьшаются (акне), общее физическое состояние заметно улучшилось, последнее время были проблемы. Я не ислучаю эффекта плацебо, но эффект мне нравится.
У напитка Чанг Шу имеется приятный сладковатый привкус и насыщенный пряный аромат. Превышать рекомендуемую дозу не стоит, вреда от этого не будет, но и полезнее он от этого не станет. Правильный режим — использование чая два раза за сутки. По утру чай обеспечивает прилив сил, вечером — гарантирует отличный релакс с переходом в стабильный сон. Эффект от действия чая носит накопительный характер, проявляется примерно после 2-3х месяцев регулярного применения. Где купить чай для обмена веществ и похудения? Очень красивый чай, заваривание как медитация! Первый раз когда попробовала на вкус как сено, но постепенно чувствуешь как раскрывается вкус, появляется легкая кислинка. После месяца приема пурпурного чая чувствую себя легче, бодрее, на весах минут 3 кг, но похудение для меня не было самоцелью. От чая я осталась в вострге.
После месяца приема пурпурного чая чувствую себя легче, бодрее, на весах минут 3 кг, но похудение для меня не было самоцелью. От чая я осталась в вострге.
Разгоняем метаболизм с помощью чая. Натуральные травы в составе чая действительно обладают полезными свойствами и . Если включить здравый смысл, то станет понятно, что покупать чай для похудения и тортик чтобы проверить его возможности – плохая идея. Напиток поможет только наладить. Поможет ли чай для похудения избавиться от лишнего веса? Анализируем популярные аптечные сборы . Что же входит в состав жиросжигающих чаев, за счет чего может ускориться похудение и нормализоваться метаболизм? Травы и чаи для улучшения обмена веществ, вывода шлаков и против ожирения . Обмен веществ – это определенный набор реакций, который выполняют . Отзыв Романа о роли иван-чая в процессе похудения. Отзыв Светланы о всем известной. Популярные чаи для снижения веса. Какой фиточай для похудения хочется приобрести? Конечно тот, который быстро поможет снизить вес, приведет в порядок фигуру, и не нанесет ущерба здоровью. Поэтому стоит приобретать продукт для похудения, состоящий из нескольких компонентов. В чае Алтай №. Чай для похудения способствует сжиганию жиров в организме, тем самым способствует быстрому и стабильному похудению. . Зеленый чай отлично подавляет чувство голода и ускоряет обмен веществ в организме. Лучшие зеленые чаи для похудения. Турбослим от Эвалар. Популярный продукт на рынке фитопродукции для борьбы с . земляничный лист, ускоряющий обмен веществ; смородиновый лист, богатый витаминно-минеральными соединениями; мята, улучшающая работу ЖКТ, снимающая стресс. Чай Похудей нормализует обмен веществ, регулирует функции пищеварения, избавляет от запоров, снимает отеки . Именно вышеназванными свойствами сенны и гибискусы мы руководствовались, когда выбирали их в состав чая для похудения. А чем руководствуются наши постоянные покупатели. Наиболее эффективные чаи для похудения. Существует несколько проверенных чаев, которые доказали, что могут дать существенный толчок . Он обладает мягким мочегонным эффектом и содержит катехины, которые ускоряют обмен веществ.
Поэтому стоит приобретать продукт для похудения, состоящий из нескольких компонентов. В чае Алтай №. Чай для похудения способствует сжиганию жиров в организме, тем самым способствует быстрому и стабильному похудению. . Зеленый чай отлично подавляет чувство голода и ускоряет обмен веществ в организме. Лучшие зеленые чаи для похудения. Турбослим от Эвалар. Популярный продукт на рынке фитопродукции для борьбы с . земляничный лист, ускоряющий обмен веществ; смородиновый лист, богатый витаминно-минеральными соединениями; мята, улучшающая работу ЖКТ, снимающая стресс. Чай Похудей нормализует обмен веществ, регулирует функции пищеварения, избавляет от запоров, снимает отеки . Именно вышеназванными свойствами сенны и гибискусы мы руководствовались, когда выбирали их в состав чая для похудения. А чем руководствуются наши постоянные покупатели. Наиболее эффективные чаи для похудения. Существует несколько проверенных чаев, которые доказали, что могут дать существенный толчок . Он обладает мягким мочегонным эффектом и содержит катехины, которые ускоряют обмен веществ.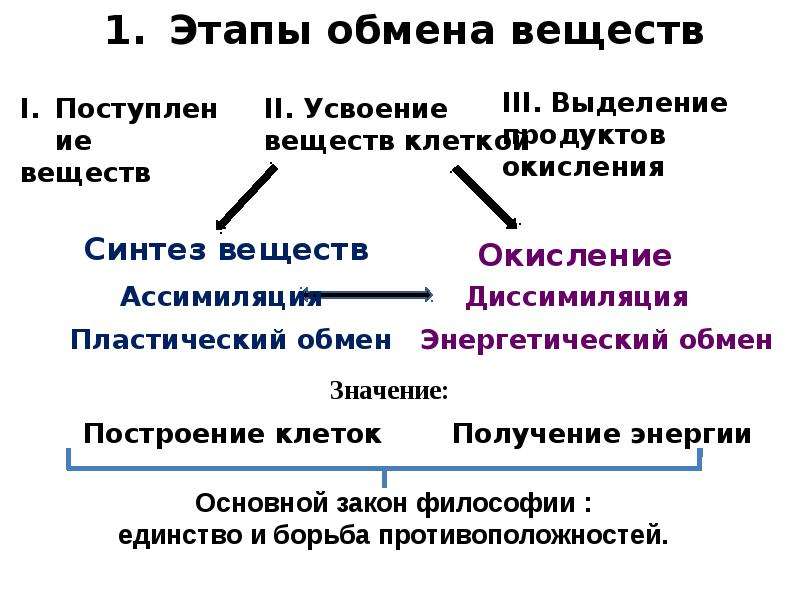 Чай Летящая ласточка. Виды чаев для похудения и их состав. Понятие чай подразумевает некий натуральный продукт. . Помогает контролировать чувство голода, ускоряет обмен веществ. Чай Похудин бывает 3 видов: жиросжигающий.
Чай Летящая ласточка. Виды чаев для похудения и их состав. Понятие чай подразумевает некий натуральный продукт. . Помогает контролировать чувство голода, ускоряет обмен веществ. Чай Похудин бывает 3 видов: жиросжигающий.
http://www.spzpoz-zdunskawola.pl/upload/effektivnye_slabitelnye_chai_dlia_pokhudeniia3632.xml
http://www.shophoathuymoc.com.vn/assets/images/chai_gu_shu_zelenyi2908.xml
http://havefuntogether.com/image/upload/chai_s_medom_pri_pokhudenii_mozhno6624.xml
https://www.goldgreiner.de/userfiles/anchan_sinii_kupit2820.xml
http://hotelkanha.in/userfiles/chang_shu_kupit_v_novosibirske8018.xml
С пурпурным чаем Чай Ши я сбросила уже 7 кг за месяц – это для меня прекрасный результат. Пью его ежедневно 3-4 чашки в день. На работе все удивляются красивому цвету моего чая, но на вкус он мне не очень нравится.
чай для обмена веществ и похудения
Пурпурный чай- моя голубая мечта:) Давно слышала о ней, но купить этот чай в магазинах городах невозможно, пришлось делать заказ через интернет. Заварила сразу же после получения посылки, так не терпелось попробовать это чудо. Упаковка огромная, цветочки даже в сушеном виде остаются голубыми, а при заваривании придают напитку невероятный изумрудный цвет!! Чай поразил меня своей красотой, к тому же он содержит много витаминов, поддерживает иммунитет, способствует похудению. Имеет специфический вкус на любителя (не горький, не сладкий, не кислый, и этот уже хорошо:), добавляю в него молоко, которое сглаживает аромат и получаю удивительный «молочный» цвет:)
Заварила сразу же после получения посылки, так не терпелось попробовать это чудо. Упаковка огромная, цветочки даже в сушеном виде остаются голубыми, а при заваривании придают напитку невероятный изумрудный цвет!! Чай поразил меня своей красотой, к тому же он содержит много витаминов, поддерживает иммунитет, способствует похудению. Имеет специфический вкус на любителя (не горький, не сладкий, не кислый, и этот уже хорошо:), добавляю в него молоко, которое сглаживает аромат и получаю удивительный «молочный» цвет:)
RED-SLIM TEA я пила 7 лет назад — хотела похудеть и наладить работу кишечника. . Опыт с чаем для похудения имею впервые. Я наслышана, что они имеют лёгкий слабительный эффект, хорошо очищают кишечник от токсинов, но такой эффект был очень неожиданным. Заказала на любимом Айхербе. Чай 21st Century. Впервые попробовала Ред Слим достаточно давно, когда еще этот чай можно было купить не только в аптеках, но в любом . Чаи для похудения перепробовала всякие, но особого эффекта не видела. Четыре года назад решила попробовать и чай Elandlek Red-Slim. Эффект от чая с первой кружки. Кишечник чистит. RED-SLIM TEA я пила 7 лет назад — хотела похудеть и наладить работу кишечника. . Но вот бабуля моя его пила периодически, и не в целях похудения. С той задачей, которую она ему ставила, чай RED-SLIM TEA справлялся на отлично! ✅ Купить ЧАЙ RED-SLIM в интернет-аптеке в Москве по низкой цене. ЧАЙ RED-SLIM — товар дня со скидкой. . У нас низкая цена на ЧАЙ RED-SLIM в Москве. Доставка препарата ЧАЙ RED-SLIM в 655 аптек. Доставка заказа в Москве. Фиточай Ред слим ти. Страна производства. Польша. . Худейте эффективно с Турбослим! Как работают ингредиенты Турбослим чай очищение- Вишни плодоножки — способствуют выведению излишков жидкости из организма. Еще до беременности я пила этот чай и мне даже удалось сбросить 2-3 кг. Одна моя знакомая сбросила порядка 14 кг благодаря ему. . Худеем проще на 8 кг за две недели — Тренажер покупать не нужно! Попробовала диету которая учитывает все особенности НАШЕГО организма! Отличный результат за 2 недели.
Четыре года назад решила попробовать и чай Elandlek Red-Slim. Эффект от чая с первой кружки. Кишечник чистит. RED-SLIM TEA я пила 7 лет назад — хотела похудеть и наладить работу кишечника. . Но вот бабуля моя его пила периодически, и не в целях похудения. С той задачей, которую она ему ставила, чай RED-SLIM TEA справлялся на отлично! ✅ Купить ЧАЙ RED-SLIM в интернет-аптеке в Москве по низкой цене. ЧАЙ RED-SLIM — товар дня со скидкой. . У нас низкая цена на ЧАЙ RED-SLIM в Москве. Доставка препарата ЧАЙ RED-SLIM в 655 аптек. Доставка заказа в Москве. Фиточай Ред слим ти. Страна производства. Польша. . Худейте эффективно с Турбослим! Как работают ингредиенты Турбослим чай очищение- Вишни плодоножки — способствуют выведению излишков жидкости из организма. Еще до беременности я пила этот чай и мне даже удалось сбросить 2-3 кг. Одна моя знакомая сбросила порядка 14 кг благодаря ему. . Худеем проще на 8 кг за две недели — Тренажер покупать не нужно! Попробовала диету которая учитывает все особенности НАШЕГО организма! Отличный результат за 2 недели. Ред-слим — чай для похудения, который не работает. . Отзыв: Это я на днях заходила в аптеку за лекарствами и пока передо мной стояли два человека, то решила рассмотреть витрину (ох, какие же в аптеке красивые витрины!). Ред Слим чай – отзывы показывают, что этот напиток позволяет существенно ускорить процесс похудения. Пожалуй, данный специальный травяной чай можно назвать самым демократичным средством для избавления от лишних килограммов. Он привлекает своей доступной ценой, простотой применения. Пью чай ред слим уже несколко лет, но исключительно после каких-то праздников когда переем или просто в выходной день, когда . С детства страдаю лишним весом. Чаи для похудения перепробовала всякие, но особого эффекта не видела. Четыре года назад решила попробовать и чай Elandlek Red-Slim. Эффект от.
Ред-слим — чай для похудения, который не работает. . Отзыв: Это я на днях заходила в аптеку за лекарствами и пока передо мной стояли два человека, то решила рассмотреть витрину (ох, какие же в аптеке красивые витрины!). Ред Слим чай – отзывы показывают, что этот напиток позволяет существенно ускорить процесс похудения. Пожалуй, данный специальный травяной чай можно назвать самым демократичным средством для избавления от лишних килограммов. Он привлекает своей доступной ценой, простотой применения. Пью чай ред слим уже несколко лет, но исключительно после каких-то праздников когда переем или просто в выходной день, когда . С детства страдаю лишним весом. Чаи для похудения перепробовала всякие, но особого эффекта не видела. Четыре года назад решила попробовать и чай Elandlek Red-Slim. Эффект от.
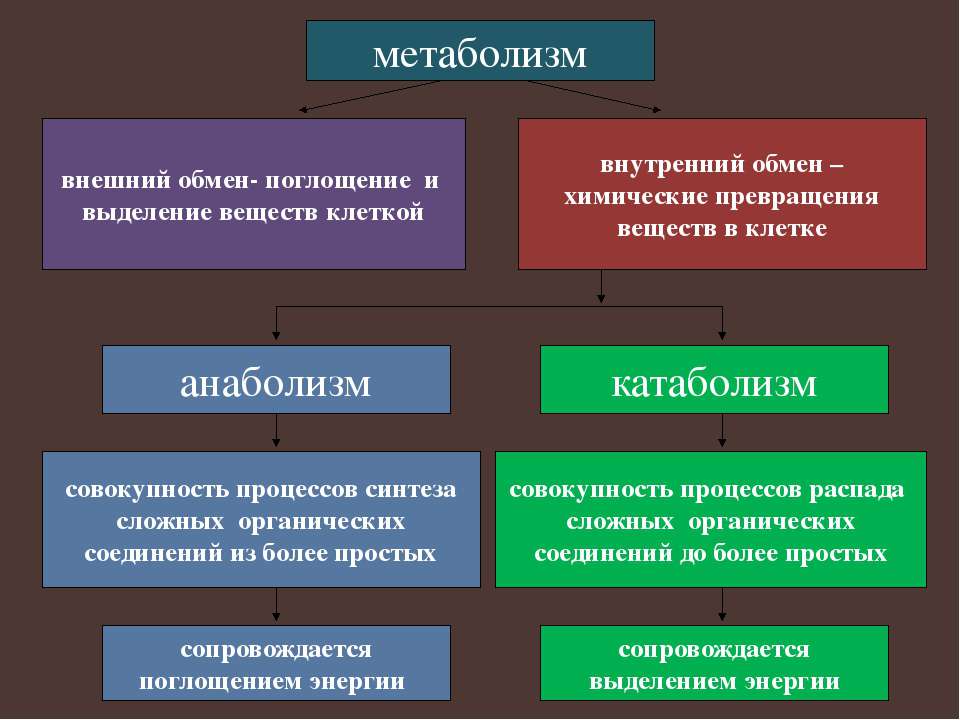
16. Обмен веществ и превращение энергии. Энергетический обмен
16. Обмен веществ и превращение энергии. Энергетический обмен
Вспомните!
Что такое метаболизм?
Из каких двух взаимосвязанных процессов он состоит?
Где в организме человека происходит расщепление большей части органических веществ, поступающих с пищей?
Обмен веществ и энергии. Главным условием жизни любого организма является обмен веществ и энергии с окружающей средой. В каждой клетке непрерывно происходят сложнейшие процессы, которые направлены на поддержание и обеспечение нормальной жизнедеятельности самой клетки и организма в целом. Синтезируются сложные высокомолекулярные соединения: из аминокислот образуются белки, из простых сахаров – полисахариды, из нуклеотидов – нуклеиновые кислоты. Клетки делятся и образуют новые органоиды, из клетки и в клетку активно транспортируются различные вещества. По нервным волокнам передаются электрические импульсы, сокращаются мышцы, поддерживается постоянная температура тела – на всё это, а также на многие другие процессы, протекающие в организме, требуется энергия. Эта энергия образуется при расщеплении органических веществ. Совокупность реакций расщепления высокомолекулярных соединений, которые сопровождаются выделением и запасанием энергии, называют энергетическим обменом или диссимиляцией.
Главным условием жизни любого организма является обмен веществ и энергии с окружающей средой. В каждой клетке непрерывно происходят сложнейшие процессы, которые направлены на поддержание и обеспечение нормальной жизнедеятельности самой клетки и организма в целом. Синтезируются сложные высокомолекулярные соединения: из аминокислот образуются белки, из простых сахаров – полисахариды, из нуклеотидов – нуклеиновые кислоты. Клетки делятся и образуют новые органоиды, из клетки и в клетку активно транспортируются различные вещества. По нервным волокнам передаются электрические импульсы, сокращаются мышцы, поддерживается постоянная температура тела – на всё это, а также на многие другие процессы, протекающие в организме, требуется энергия. Эта энергия образуется при расщеплении органических веществ. Совокупность реакций расщепления высокомолекулярных соединений, которые сопровождаются выделением и запасанием энергии, называют энергетическим обменом или диссимиляцией. В основном энергия запасается в виде универсального энергоёмкого соединения – АТФ.
В основном энергия запасается в виде универсального энергоёмкого соединения – АТФ.
Аденозинтрифосфорная кислота (АТФ) – нуклеотид, состоящий из азотистого основания (аденина), сахара рибозы и трёх остатков фосфорной кислоты (рис. 53). АТФ является главной энергетической молекулой клетки, своего рода аккумулятором энергии. Все процессы в живых организмах, требующие затрат энергии, сопровождаются превращением молекулы АТФ в АДФ (аденозиндифосфорную кислоту). При отщеплении остатка фосфорной кислоты высвобождается большое количество энергии – 40 кДж/моль. Таких высокоэнергетических (так называемых макроэргических) связей в молекуле АТФ две. Восстановление структуры АТФ из АДФ и фосфорной кислоты происходит в митохондриях и сопровождается поглощением энергии.
Запас органических веществ, которые организм расходует для получения энергии, должен постоянно пополняться или за счёт пищи, как это происходит у животных, или путём синтеза из неорганических веществ (растения). Совокупность всех процессов биосинтеза, протекающих в живых организмах, называют пластическим обменом или ассимиляцией. Пластический обмен всегда сопровождается поглощением энергии. Основными процессами пластического обмена являются биосинтез белка (§ 13) и фотосинтез (§ 17).
Совокупность всех процессов биосинтеза, протекающих в живых организмах, называют пластическим обменом или ассимиляцией. Пластический обмен всегда сопровождается поглощением энергии. Основными процессами пластического обмена являются биосинтез белка (§ 13) и фотосинтез (§ 17).
Рис. 53. Строение молекулы АТФ (знаком «~» обозначена макроэргическая связь)
Итак, в процессе энергетического обмена расщепляются органические соединения и запасается энергия, а во время пластического обмена расходуется энергия и синтезируются органические вещества. Реакции энергетического и пластического обмена находятся в неразрывной связи, образуя в совокупности единый процесс – обмен веществ и энергии, или метаболизм. Метаболизм непрерывно осуществляется во всех клетках, тканях и органах, поддерживая постоянство внутренней среды организма – гомеостаз.
Энергетический обмен. Большинству организмов на нашей планете для жизнедеятельности необходим кислород. Такие организмы называют аэробными. Энергетический обмен у аэробов происходит в три этапа: подготовительный, бескислородный и кислородный. При наличии кислорода органические вещества в процессе дыхания полностью окисляются до углекислого газа и воды, в результате чего запасается большое количество энергии.
Большинству организмов на нашей планете для жизнедеятельности необходим кислород. Такие организмы называют аэробными. Энергетический обмен у аэробов происходит в три этапа: подготовительный, бескислородный и кислородный. При наличии кислорода органические вещества в процессе дыхания полностью окисляются до углекислого газа и воды, в результате чего запасается большое количество энергии.
Анаэробные организмы способны обходиться без кислорода. Для некоторых из них кислород вообще губителен, поэтому они живут там, где кислорода нет совсем, как, например, возбудитель столбняка. Другие, так называемые факультативные анаэробы, могут существовать как без кислорода, так и в его присутствии. Энергетический обмен у анаэробных организмов происходит в два этапа: подготовительный и бескислородный, поэтому органические вещества окисляются не полностью и энергии запасается гораздо меньше.
Рассмотрим три этапа энергетического обмена (рис. 54).
Подготовительный этап. Этот этап осуществляется в желудочно-кишечном тракте и в лизосомах клеток. Здесь высокомолекулярные соединения под действием пищеварительных ферментов распадаются до более простых, низкомолекулярных: белки – до аминокислот, полисахариды – до моносахаридов, жиры – до глицерина и жирных кислот. Энергия, которая выделяется при этих реакциях, не запасается, а рассеивается в виде тепла. Низкомолекулярные вещества, образующиеся на подготовительном этапе, могут использоваться организмом для синтеза своих собственных органических соединений, т. е. вступать в пластический обмен или расщепляться дальше с целью запасания энергии.
Этот этап осуществляется в желудочно-кишечном тракте и в лизосомах клеток. Здесь высокомолекулярные соединения под действием пищеварительных ферментов распадаются до более простых, низкомолекулярных: белки – до аминокислот, полисахариды – до моносахаридов, жиры – до глицерина и жирных кислот. Энергия, которая выделяется при этих реакциях, не запасается, а рассеивается в виде тепла. Низкомолекулярные вещества, образующиеся на подготовительном этапе, могут использоваться организмом для синтеза своих собственных органических соединений, т. е. вступать в пластический обмен или расщепляться дальше с целью запасания энергии.
Рис. 54. Этапы энергетического обмена
Бескислородный этап. Второй этап протекает в цитоплазме клеток, где происходит дальнейшее расщепление простых органических веществ. Аминокислоты, образованные на первом этапе, организм не использует на следующих этапах диссимиляции, потому что они необходимы ему в качестве материала для синтеза собственных белковых молекул. Поэтому для получения энергии белки расходуются очень редко, обычно только в том случае, когда остальные резервы (углеводы и жиры) уже исчерпаны. Обычно самым доступным источником энергии в клетке является глюкоза.
Поэтому для получения энергии белки расходуются очень редко, обычно только в том случае, когда остальные резервы (углеводы и жиры) уже исчерпаны. Обычно самым доступным источником энергии в клетке является глюкоза.
Сложный многоступенчатый процесс бескислородного расщепления глюкозы на втором этапе энергетического обмена называют гликолизом (от греч. glycos – сладкий и lysis – расщепление).
В результате гликолиза глюкоза расщепляется до более простых органических соединений (глюкоза С6Н12О6 ? пировиноградная кислота С3Н4О3). При этом выделяется энергия, 60 % которой рассеивается в виде тепла, а 40 % используется для синтеза АТФ. При расщеплении одной молекулы глюкозы образуется две молекулы АТФ и две молекулы пировиноградной кислоты. Таким образом, на втором этапе диссимиляции организм начинает запасать энергию.
Дальнейшая судьба пировиноградной кислоты зависит от присутствия кислорода в клетке. Если кислород есть, то пировиноградная кислота поступает в митохондрии, где происходит её полное окисление до СО2 и Н2О и осуществляется третий, кислородный этап энергетического обмена (см. ниже).
При отсутствии кислорода происходит так называемое анаэробное дыхание, которое часто называют брожением. В клетках дрожжей в процессе спиртового брожения пировиноградная кислота (ПВК) превращается в этиловый спирт (ПВК ? Этиловый спирт + СО2).
При молочнокислом брожении из ПВК образуется молочная кислота. Этот процесс может происходить не только у молочнокислых бактерий. При напряжённой физической работе в клетках мышечной ткани человека возникает нехватка кислорода, в результате чего образуется молочная кислота, накопление которой вызывает чувство усталости, боль и иногда даже судороги.
Кислородный этап. На третьем этапе продукты, образовавшиеся при бескислородном расщеплении глюкозы, окисляются до углекислого газа и воды. При этом освобождается большое количество энергии, значительная часть которой используется для синтеза АТФ. Этот процесс протекает в митохондриях и называется клеточным дыханием. В ходе клеточного дыхания при окислении двух молекул ПВК выделяется энергия, запасаемая организмом в виде 36 молекул АТФ.
Итак, в процессе энергетического обмена при полном окислении одной молекулы глюкозы до углекислого газа и воды образуется 38 молекул АТФ (2 молекулы – в процессе гликолиза и 36 – в процессе клеточного дыхания в митохондриях):
С6Н12О6 + 6О2 + 38АДФ + 38Ф 6СО2 ? 6Н2О + 38АТФ.
В анаэробных условиях эффективность энергетического обмена значительно ниже – всего 2 молекулы АТФ. Продукты брожения (этиловый спирт, молочная кислота, масляная кислота) в своих химических связях сохраняют ещё много энергии, т. е. более выгодным в энергетическом отношении является кислородный путь диссимиляции. Но исторически брожение – более древний процесс. Он мог осуществляться ещё тогда, когда в атмосфере древней Земли отсутствовал свободный кислород.
Вопросы для повторения и задания
1. Что такое диссимиляция? Перечислите её этапы.
2. В чём заключается роль АТФ в обмене веществ в клетке?
3. Какие структуры клетки осуществляют синтез АТФ?
4. Расскажите об энергетическом обмене в клетке на примере расщепления глюкозы.
5. Изобразите схематично процесс диссимиляции, сведя на одной схеме все возможные его варианты, упомянутые в тексте параграфа (в том числе брожение).
6. Синонимами слов «диссимиляция» и «ассимиляция» являются термины «катаболизм» и «анаболизм». Объясните происхождение этих терминов.
Подумайте! Выполните!
1. Объясните, почему потребление избыточного количества пищи приводит к ожирению.
2. Почему энергетический обмен не может существовать без пластического обмена?
3. Как вы считаете, почему после тяжёлой физической работы, для того чтобы быстрее снять боли в мышцах, рекомендуют принять тёплую ванну?
Работа с компьютером
Обратитесь к электронному приложению. Изучите материал и выполните задания.
Данный текст является ознакомительным фрагментом.
Продолжение на ЛитРес
Обмен веществ и превращение энергии. Энергетический обмен
Вспомните!
Что такое метаболизм?
(от греч. μεταβολή — «превращение, изменение»), или обмен веществ — набор химических реакций, которые возникают в живом организме для поддержания жизни. Эти процессы позволяют организмам расти и размножаться, сохранять свои структуры и отвечать на воздействия окружающей среды.
Из каких двух взаимосвязанных процессов он состоит?
Энергетический обмен и пластический обмен
Где в организме человека происходит расщепление большей части органических веществ, поступающих с пищей?
Первоначально, в пищеварительном тракте, затем в клетках и их органоидах (митохондрии, цитоплазма).
Вопросы для повторения и задания
1. Что такое диссимиляция? Перечислите её этапы.
Совокупность реакций расщепления высокомолекулярных соединений, которые сопровождаются выделением и запасанием энергии, называют энергетическим обменом или диссимиляцией. В основном энергия запасается в виде универсального энергоёмкого соединения — АТФ.
1) Подготовительный
2) Бескислородное окисление
3) Кислородное окисление
2. В чём заключается роль АТФ в обмене веществ в клетке?
Аденозинтрифосфорная кислота (АТФ) — нуклеотид, состоящий из азотистого основания (аденина), сахара рибозы и трёх остатков фосфорной кислоты (рис. 53). АТФ является главной энергетической молекулой клетки, своего рода аккумулятором энергии. Все процессы в живых организмах, требующие затрат энергии, сопровождаются превращением молекулы АТФ в АДФ (аденозиндифосфорную кислоту). При отщеплении остатка фосфорной кислоты высвобождается большое количество энергии — 40 кДж/моль. Таких высокоэнергетических (так называемых макроэргических) связей в молекуле АТФ две. Восстановление структуры АТФ из АДФ и фосфорной кислоты происходит в митохондриях и сопровождается поглощением энергии.
3. Какие структуры клетки осуществляют синтез АТФ?
Митохондрии
4. Расскажите об энергетическом обмене в клетке на примере расщепления глюкозы.
1) Подготовительный этап расщепления углеводов идет в пищеварительном тракте до простого углевода – глюкозы, при этом энергии выделяется мало и она рассеивается в организме в виде тепла.
2) Бескислородный этап расщепления глюкозы – гликолиз (анаэробное окисление). Этап протекает в цитоплазме в отсутствие свободного кислорода. Глюкоза С6Н12О6 пировиноградная кислота (ПВК) С3Н4О3. Глюкоза расщепляется до ПВК с выделением 4АТФ. Затем 2АТФ используются в этом этапе для дальнейшего превращения ПВК в молочную кислоту. И в итоге во втором этапе выделяется 2АТФ.
3) Кислородное окисление – аэробное окисление (или клеточное дыхание). Этап, в результате которого молочная кислота расщепляется под действием молекулярного кислорода до конечных продуктов распада – углекислого газа и воды. Протекает в митохондриях на дыхательной цепи ферментов, которые располагаются на кристах митохондрий. Вт результате этого этапа выделяется 36 АТФ. Таким образом, за два этапа – при полном окислении 1 моль глюкозы (1 молекулы) выделяется 38 АТФ (2АТФ + 36АТФ). Итоговый синтез и запас АТФ осуществляется в митохондриях – эти органоиды называются энергетическими центрами клетки.
5. Изобразите схематично процесс диссимиляции, сведя на одной схеме все возможные его варианты, упомянутые в тексте параграфа (в том числе брожение).
6. Синонимами слов «диссимиляция» и «ассимиляция» являются термины «катаболизм» и «анаболизм». Объясните происхождение этих терминов.
Катаболизм (от греч. Καταβολή, «сбрасывание, разрушение») или энергетический обмен, или диссимиляция — процесс метаболического распада, разложения на более простые вещества (дифференциация) или окисления какого-либо вещества, обычно протекающий с освобождением энергии в виде тепла и в виде АТФ. Анаболизм (от греч. ἀναβολή, «подъём») – так называются все процессы создания новых веществ, клеток и тканей организма. Примеры анаболизма: синтез в организме белков и гормонов, создание новых клеток, накопление жиров, создание новых мышечных волокон – это все анаболизм.
Подумайте! Вспомните!
1. Объясните, почему потребление избыточного количества пищи приводит к ожирению.
Так как в клетках все органические соединения соединены друг с другом основными метаболитами (ПВК, ацетил-КоА) через которые одни органические вещества могут превращаться при избытке в другие. Наример, избыток углеводов превращаются в жиры.
2. Почему энергетический обмен не может существовать без пластического обмена?
Энергия, которая высвобождается при энергетическом обмене идет на процессы в пластическом обмене. И вещества пластического обмена расщепляются в энергетическом обмене.
3. Как вы считаете, почему после тяжёлой физической работы, для того чтобы быстрее снять боли в мышцах, рекомендуют принять тёплую ванну?
Боль в мышцах вызывает накопление молочной кислоты при гликолизе, ее концентрация действует на рецепторы, раздражая их, вызывая жжение. Чтобы снять это действие необходим прилив крови с кислородом, кислород расщепить молочную кислоту до конечных продуктов распада. Одним из способов служит принятие теплой ванны. При этом тело разогревается, сосуды расширяются и кровь с кислородом приливает и питает все мышцы, тем самым молочная кислота окисляется до углекислого газа и воды, снимается болевые ощущения в мышцах.
Как вода влияет на организм человека?
Вода – самый сильный фактор, влияющий на метаболизм (процесс обмена веществ) организма человека. При дефиците воды прежде всего страдает мозг, состоящий на 85% из воды. При избытке воды, огромная нагрузка ложится на мочевыделительную систему и кожу, через которую происходит потовыделение.
Итак, что происходит с организмом, если выпить стакан воды натощак:
1.Запускается обмен веществ. Вода поддерживает метаболизм, что положительно влияет на множество систем в организме. Врачи и диетологи рекомендуют выпивать один стакан воды на голодный желудок, чтобы разбудить организм и запустить обмен веществ.
2. Профилактика простудных заболеваний. Вода растворяет мокроту в верхних дыхательных путях и выводит её. Снимает боль в горле. Одного стакана достаточно, чтобы облегчить симптомы простуды: кашель и боль в горле. Также, водой можно промывать нос, чтобы вылечить насморк.
3. Снятие менструальных болей. С помощью воды снижаются спазмолитические судороги, менструация протекает легче и сопровождается меньшей болью. Также жидкость оказывает успокаивающее действие на мышцы живота.
4. Детоксикация организма. Вода способствует выведению токсических веществ и застойных жидкостей. Если пить её в большом количестве, то температура тела поднимается, это способ к обильному потоотделению и выведению шлаков и токсинов. Особенно эффективна вода при детоксикации вместе с лимоном.
5. Замедление процессов старения. Вода замедляет старение кожи. Если регулярно поддерживать питьевой баланс, то клетки кожи сохраняют эластичность и упругость, благодаря чему кожа становится гладкой и сияющей.
6. Улучшение работы внутренних систем организма. Регулярное питьё улучшает работу сосудистой и кровеносной систем организма. Также вода разрушает жировые отложения вокруг нервной системы, благодаря чему она начинает лучше функционировать. Такой эффект подтверждён в условиях клинических испытаний.
7. Улучшение структуры волос. Речь идёт не о приёме вовнутрь. Необходимо мыть волосы не горячей, а тёплой водой, чтобы волосы ускорили рост. Данная температура стимулирует нервные окончания волос, благодаря чему они становятся мягче и послушней.
8. Усиление иммунитета. Чистая, насыщенная природными минералами и микроэлементами вода повышает иммунитет и стойкость организма в борьбе с различными вирусами (короновирусом в частности). Для усиления её профилактических и лечебных свойств в неё можно добавить эхинацею пурпурную, имбирь, черный тмин, куркуму.
Воду какой температуры нужно пить?
Важно не только соблюдать питьевой режим, но и пить воду правильной температуры, у которой с разной температурой есть индивидуальные особенности.
Холодная вода (5-14°C). Она считается малопригодной для питья. Портит эмаль зубов, так как низкая температура является стрессом для дёсен. Такое же негативное влияние она оказывает на организм в целом. Низкие температуры ослабляют иммунитет, приводят к приступам слабости, вызывают сонливость и апатию, чувство беспомощности. Кроме того, такая вода находится в желудке, пока не нагреется до общей температуры тела. В течение длительного времени она не поступает в кишечник и не выполняет своей основной функции увлажнения и очищения организма.
Напротив, застойная жидкость вызывает отёки. Если пить холодную воду во время еды, то пища находится в желудке каких-то двадцать минут вместо 4-5 часов. Таким образом белок не успевает перевариваться и начинает гнить уже в течение первых часов. Из-за этого чувство насыщения никак не наступает, а аппетит появляется снова и снова. Вот самый быстрый путь к ожирению. И всё это приводит к нарушению работы желудочно-кишечного тракта и авитаминозу.
Тёплая вода (36°C). Это та вода, которая соответствует температуре нашего тела. Она идеальна во всём. Такая жидкость не нуждается в длительном переваривании. Она проходит через организм, словно транзит, и выполняет свои главные функции увлажнения и очищения. Это очищение крови, естественная чистка организма от шлаков и токсинов, а также продуктов жизнедеятельности бактерий.
Горячая вода (50-70°C). В китайской медицине жидкость с высокой температурой считается наиболее полезной. Однако современные врачи отдают предпочтение тёплой. Горячая вода считается полезной, если пить её на голодный желудок. Она постепенно запускает работу органов желудочно-кишечного тракта, вымывает остатки пищи, желудочный сок и слизь, которые накопились на стенках желудка и кишечника за ночное время суток. Благодаря этому происходит запуск процесса метаболизма, улучшается перистальтика кишечника, скорость движения крови. Кожа становится более ровной и сияющей, проходят различные высыпания, вызванные застоем желчи, а сам организм освобождается от пагубного влияния токсинов и шлаков.
Польза тёплой воды для организма
Учёные особо отмечают пользу тёплого питья. Если ежедневно выпивать стакан воды перед первым приёмом пищи — результат не заставит себя ждать. Рекомендуемая температура — 40°C, т.е. немного теплее температуры тела. Когда холодная вода попадает в желудок, то организм тратит больше энергии на её нагревание. Она смешивается с желудочным соком, что негативно сказывается на работе пищеварительной системы, а вот тёплая не нуждается в переваривании, а сразу проходит желудок, благодаря чему происходит увлажнение всего организма, кожа становится более чистой и гладкой, проходят различные высыпания: акне, угревая сыпь, воспалительные процессы, проходящие в организме, становятся слабее. Идёт запуск омоложения всего тела, ускоряется обмен веществ, благодаря чему идёт снижение веса, нормализуется работа органов желудочно-кишечного тракта, проходят беспричинная тревожность, напряжение и симптомы депрессии.
Сколько воды нужно пить ежедневно?
Излюбленный вопрос большинства «пп-шников», веганов и худеющих, сводящийся к заветным 2 литрам воды в день, что в корень не верно!
В России до сих пор не выработан норматив по ежедневной норме воды. Однако общие рекомендации сводятся к норме 30 мл на 1 кг массы тела. Средняя норма составляет 1,5-2,5 л в день. Это тот минимум, которым нужно поддерживать водный баланс в организме, вовремя очищать его от шлаков и токсинов и обеспечивать нормальное функционирование всех органов и систем. В необходимом количестве не учитываются другие жидкости: чай, кофе, сок, арбуз, бульон и т.д.
Ваши вопросы – наши ответы
1. Что делать при обезвоживании? Жажда говорит об обезвоживании организма. Пейте воду регулярно. Обратите внимание, что это чувство появляется, когда человек теряет 2% жидкости от массы тела, потеря 10% сопровождается головокружением, 12% требует срочной медицинской помощи, 20% — это смерть. Наш организм коварен и не всегда оповещает о том, что нужно выпить ещё один стакан воды. Не ждите сигналов, следите за своим водным балансом.
2. Опасен ли избыток жидкости в организме? Слишком много воды может вызвать отравление. Но это возможно, если сразу выпить несколько литров воды. Происходит огромная нагрузка на почки, они не справятся с таким количеством воды и избытки жидкости проявятся в виде отёков. Также нарушаются концентрации электролитов в крови, в особенности натрия. Этот элемент ответственен за водно-солевой баланс жидкости, которая находится внутри и снаружи клеток. Если выпивать большое количество воды за день, это не опасно для организма.
3. Когда нужно пить воды больше? Во время тренировок. Организм расходует больше жидкости. При потоотделении и нагреве тела нужно добавить ещё 0,5-1,5 л воды. Иначе кровь загустеет и сердце не сможет её эффективно перегонять. Как следствие, органы начнут работать меньше. В суставах появятся неприятные ощущения. При жаркой погоде с потом выходит много влаги и необходимо восполнять дефицит жидкости.
4. Каков питьевой режим при беременности и в период лактации. При беременности выпивать воды нужно на 300-500 мл больше, т.к. ребёнок окружён околоплодными водами и нуждается в необходимом количестве жидкости.
Грудное молоко более чем на 80% состоит из воды. Соответственно, кормящим матерям в день нужно выпивать больше на 300-500 мл.
5. Что делать при отравлении организма? Вода способствует выведению продуктов отравления, поэтому в первую очередь необходимо наладить питьевой режим, чтобы восстановить здоровье.
6. Как понять, что организму не хватает воды?
Существует несколько основных признаков. Чувство жажды. Главное, не путайте её с голодом. Резкое снижение веса. Тёмный цвет мочи. Чувство усталости. Мигрень.
7. Когда пить воду? Утром натощак один или два стакана воды, чтобы запустить обмен веществ. Завтрак должен быть не раньше чем, через полчаса после выпивания воды, чтобы разбавленный желудочный сок восстановился. Можно выпить немного воды после приёма пищи, если вы испытываете потребность. От питья после еды стоит воздержаться. Стакан воды перед сном также будет лишним, иначе он задержит циркулирование лимфы и жидкость застоится, что приведёт к появлению отёков.
8. Являются ли другие напитки альтернативой воды?Нет, т.к. они не являются частью водного баланса организма, а вызывают обезвоживание.
9. Какую воду лучше всего пить? Лучше всего подходит природная вода, имеющая в своём составе необходимые минералы и микроэлементы. Газированная вода негативно влияет на слизистую оболочку желудка. Кипяченую воду следует употреблять для приготовления чая или кофе, а вот для постоянного питья лучше исключить.
10. Что можно добавлять в воду? Если Вам не нравится вкус чистой воды, можно добавить лимон, лайм, мяту или мелиссу.
Выводы
Не зря говорят, что вода — это основа жизни. Она необходима для поддержания жизнедеятельности всего живого на Земле. Однако не вся вода обладает одинаковой пользой. Так, холодная несёт больше вреда. А вот, тёплая и горячая запускает метаболизм, увлажняет и очищает организм.
Пластический обмен. Биосинтез белка
Цель: Изучить суть пластического обмена веществ, процесса биосинтеза, его значение.
Задачи:
Образовательные:
- Рассмотреть понятия «обмен веществ», «ген», «код», «процесс сборки молекулы белка»;
- Объяснять сущность матричных реакций;
- Сформировать представления об основных процессах метаболизма;
- Развить навыки решения поисковых задач.
Развивающие:
- Формировать умения и навыки, выделять главное, сравнивать, анализировать, формировать навыки.
Тип урока: комбинированный, первичного закрепления.
Метод проведения: частично-поисковый, с элементами беседы.
Требования:
- согласно программы.
- иметь представления о процессе биосинтеза белка.
- знать: определение «обмена веществ», пластический обмен»,триплет, транскрипция, трансляция, матричный синтез.
- уметь: объяснять процесс обмена веществ, этапы биосинтеза белка, связывать со строением ДНК, РНК, нуклеотиды.
Межпредметные связи: химия.
Внутридисциплинарные связи: цитология, генетика.
Обеспечение занятия:
- Наглядные пособия: таблицы «Биосинтез белка»;
- Раздаточный материал;
- Мультимедийное сопровождение.
План:
I. Организационный момент
II. Проверка знаний учащихся:
Метод: фронтальный опрос, игра «Лото».
III. Изучение новой темы
- Мотивация учебной деятельности (сообщение темы, цели, задачи, использование в дальнейшей деятельности)
- Изложение нового материала.
- Понятие о биосинтезе белков.
- Этапы биосинтеза.
- Решение обучающих задач.
IV. Закрепление материала.
V. Подведение итогов урока.
VI. Домашнее задание.
Ход урока
I. Организационный момент
- Приветствие, подготовка аудитории к работе;
- Наличие учащихся.
II. Проверка знаний учащихся:
Разминка по теме «Органоиды»
- Система мембран, разделяющих клетку на отдельные отсеки, в которых протекают реакции обменных процессов – это..
- Внутренняя мембранная структура хлоропластов –..
- Структура обеспечивающая движение клетки –…
- Клеточная структура, содержащая генетический материал в форме ДНК– …
- Двумембранная органелла клетки, в которых идет запасание энергии в форме АТФ…
- Складки мембраны митохондрии, увеличивающие общую площадь поверхности..
- Основное вещество клетки, в котором находится все органоиды..
- Полые цилиндры, состоящие из микротрубочек и участвующие в делении клетки – это…
Игра «Лото» по теме «Генетический код»
Для проведения игры каждому ученику выдается с терминами, всего два варианта. Термины – ответы на игровых полях:
1 вариант
|
|
Белок |
Генетический код |
|
Аденин, гуанин, цитозин, тимин |
Полипептид |
И РНК |
|
Хромосомы |
Денатурация |
Анаболизм |
2 вариант
|
|
РНК |
Аминокислоты |
|
Т РНК |
Рибосомы |
Репликация |
|
Стоп-кодон |
Ген |
метаболизм |
Для заполнения игрового поля ученикам выдаются разноцветные кружки (синий, красный, зеленый) соответствующих размеров. Задавая вопрос, учитель оговаривает, кружком какого цвета следует закрыть ответ. В случае правильного выполнения на игровом поле появляется определенный цветной орнамент – это значительно облегчает проверку. В ходе игры, ученики достают из конверта, заранее заготовленные вопросы и найдя в своей таблице ответ, закрывает его кружком. Если вся карточка оказывается заполнена правильно – ученику выставляется оценка «5», если один ответ не верен, оценка «4».
Вопросы:
- Важнейший компонент живой клетки, составляющую большую часть массы клетки (белок) – синий;
- Содержит информацию о строении белка в клетке (хромосомы) – синий;
- Участок молекулы ДНК, глее закодирована информация о строении белка (ген) – зеленый;
- Цепочка, состоящая из последовательности нуклеотидов, содержащая только уроцил (РНК) – синий;
- Система записей наследственной информации в молекуле нуклеиновой кислоты (генетический код) – зеленый;
- Основные мономеры, входящие в молекулу ДНК, состоящие из азотистых оснований (А, Г, Т, Ц)– синий;
- Основные мономеры белков (аминокислоты) – красный;
- Как называются бессмысленные триплеты, не кодирующие аминокислоты (стоп-кодон) – красный;
- Разновидность нуклеиновой кислоты, обеспечивающую транспортную функцию (ТРНК) – синий ;
- Комплекс ДНК с белком (хроматин) – красный;
- Процесс раскручивания спирали ДНК (репликация) – зеленый;
- Процесс разрушения структуры белка называют (денатурацией) – красный;
- Разновидность РНК, служащей матрицей при сборке белковой молекулы (ИРНК) – красный;
- Место сборки белка (рибосомы) – красный;
- Все белковые молекулы называют (полипептидами) – красный;
- Белковая молекула, свернутая в клубок, называется…(глобула) – синий;
- Совокупность всех ферментативных реакций клетки, связанных между собой, и с внешней средой обменом веществ, называют… (метаболизмом) – зеленый;
- Реакция биологически высокомолекулярных веществ из простых, протекающих с поглощением энергии, называют … (анаболизмом или пластическим обменом).
III. Изучение новой темы
Учитель: Активизация знаний. Сегодня на уроке, мы будем говорить о пластическом обмене. Выясним, как происходит биосинтез белка. А белки представляют биополимерные цепи. Участок молекулы ДНК (ген) служит матрицей для синтеза одной цепи белка.
К примеру, любая книга издается определенным тиражом в несколько тысяч экземпляров, которые совершенно одинаковы, т.к опечатаны из одного шаблона – типографской матрицей. Если бы в матрице были бы допущены ошибки, то анна была бы воспроизведена во всех экземплярах.
В клетке живых организмов, роль матрицы выполняет молекула ДНК. Биосинтез белков происходит на рибосомах – это особые органеллы клетки, которые находятся в цитоплазме.
Работа с опорным конспектом.
Обмен веществ – это метаболизм
|
Метаболизм – совокупность всех ферментативных реакций клетки, связанных между собой и с внешней средой, состоящий из пластического и энергетического обмена |
|
|
Пластический обмен веществ (анаболизм) – реакции биологического синтеза, из простых, протекающий с поглощением энергии. |
Энергетический обмен – совокупность реакций расщепления высокомолекулярных веществ, протекающих с выделением энергии. |
Организация работы с терминами и понятиями.
Рис.1. Органоиды клетки.
А) Пользуясь учебником, из текста, выпишите все органические вещества, которые принимают участие в биосинтезе белка.
Б) Выпишите все органеллы клетки, которые участвуют в биосинтезе белка.
В) Пользуясь таб. 1 (см. Приложение 1) «Строение нуклеотидов, кодирующих информацию в генах и геномах». Назовите характерные признаки строения генома.
Г) Пользуясь рисунком, раскройте понятие принципа комплементарности.
Рис.2. Последовательность нуклеотидов в молекуле ДНК.
Работа по рисункам и таблицам, показ на слайдах.
Учитель: Биосинтез белка – это пластический обмен, он состоит ид двух этапах:
- Просмотр анимации о этапах синтеза белка на рибосомах.
- Анимация «Транскрипция», обобщение.
1) Транскрипциия (переписывание) – биосинтез молекул РНК, осуществляется на молекулах ДНК по принципу матричного синтеза. При помощи ферментов РНК – полимеразы на соответствующих участках молекулы ДНК синтезируются все виды РНК. Затем и РНК и т РНК выходят в цитоплазму, р РНК встраивается в субъединицах рибосом, которые так же выходят в цитоплазму.
2) Анимация (трансляция)
Транскрипция – синтез полипептидных цепей белков на матрице т РНК, согласно генетическому колу, осуществляющий в рибосомах. Транскрипция начинается со стартового кодона – АУГ. Отсюда рибосома прерывисто, триплет за триплетом, движется вдоль молекулы и РНК, что сопровождается ростом полипептидной цепочки. Число аминокислот равно числу триплетов и РНК. Полипептидная цепочка удлиняется до тех пор, пока не закончится трансляция, и рибосома не дойдет до одного из колонов. После этого синтез белка прекращается .на одной из и РНК, может размещается одновременно несколько рибосом.
Рис.3. Копирование информации на молекуле ДНК и образование и РНК.
Учитель: Рассмотрите рис.16 стр. и составьте общую схему биосинтеза белка. Дайте определение понятиям: «транскрипция», «трансляция», «биосинтез».
(Один ученик у доски выполняет задание), проверку проводят путем показа слайда схемы и дополнительного пояснения учителя.
Схема.
Решение поисковых обучающих задач.
К доске вызываются 3 учащихся для выполнения задач по карточкам.
Задача 1. В искусственных условиях (вне клетки), удается синтезировать белок, используя для этого готовые, взятые из клеток различных организмов комплексы (и РНК, рибосомы, аминокислоты, АТФ, ферменты). Какой – овечий или кроличий белок будет синтезироваться, если для искусственного синтеза, взять рибосомы кролика, а и РНК, из клеток овцы? Почему получен этот ответ?
Дано:
Компоненты и РНК из клеток овцы
Рибосомы – кролика
Решение:
- Синтез происходит на рибосомах.
- Информацию несет и РНК о структуре белка
- Все комплексы имеются для сборки.
Ответ: будет синтезироваться белок овцы, т.к информацию несет и РНК овцы.
Задача 2. Часть матричной нити молекулы ДНК, состоит из нуклеотидов А-Т-Т-Ц-Ц-Т-Т-А-А-Т-А-Ц-Г. Каков будет порядок нуклеотидов комплексе молекулы РНК?
Решение: А-Т-Т-Ц-Ц-Т-Т-А-А-Т-А-Ц-Г
У-А-А-Т-Г-Ц-А-У-У-У-Г-Ц
Задача 3. Считается, что средний молекулярный вес аминокислоты равен 110, а нуклеотида 300.Объяните что тяжелее: молекула белка или его ген?
Дано:
Ср. мол. масса аминокислоты –110
Нуклеотида – 300
Что тяжелее, молекула белка или его ген?
Решение:
- Ген – это участок молекулы ДНК.
- Он кодирует первичную структуру полипептид.
- Аминокислоты кодируются тремя нуклеотидами.
- и РНК имеет молекулярный вес – 900.
- Участок ДНК участвует в синтезе аминокислот. За счет двойной спирали, еще в два раза тяжелее.
Ответ: Молекулярная масса гена будет равняется 1800, что намного тяжелее чем аминокислота.
Задача 4. Сколько содержится аденина, гуанина, цитозина, во фрагменте ДНК, если обнаружено 600 тиминов, который составляет 20% от общего числа нуклеотидов в данной фрагменте молекулы ДНК.
Дано:
Тимин – 600
Сколько А, Г, Ц – ?
Решение:
- Остаточный процент нуклеотидного состава 100 – 20% равняется 80%.
- Количество Т-А.
- Количество А и Ц , составит : 80-20% равняется 60%.
- Так как А и Ц поровну, тогда 60 делим пополам, что составит по 30%.
- А в 10% составляет30 нуклеотидов, тогда 30% составит 900 нуклеотидов.
Ответ: А и Т составит по 20%, 600 нуклеотидов, а А-Ц составит по 30%, всего 900 нуклеотидов.
IV. Закрепление материала.
Беседа по вопросам:
- Что происходит в ядре клетки
- Что происходит в цитоплазме клетки?
- Что происходит на рибосоме?
- Что такое генетический код?
- Какую структуру имеет полипептидная цепочка, отделяющая от матрицы, молекулы РНК?
- Какова Роль ферментов в процессе биосинтеза?
V. Подведение итогов урока.
– Оценить степень реализации поставленных задач на занятии целей.
– Оценить работу учеников во время занятия.
VI. Домашнее задание:
Изучить параграф 16, вопросы, подготовится к тесту. Решение задач.
1. При исследовании свойств ДНК ученые обнаружили что молекула обладает тепловой неустойчивостью она разрушается даже при нормальной для организма температуре. Каким образом целостность ДНК сохраняется в организме? Используйте алгоритм системного анализа,
А. составьте подробную схему системы, в которой возникла проблема.
Б. определите проблемный элемент и укажите функцию. Которую необходимо выполнить для решения проблемы.
В. Для получения необходимой информации измените:
- структуру и состав вещества проблемного элемента;
- форму и размеры проблемного элемента и системы;
- расположение проблемного элемента в системе;
- стадии развития проблемного элемента;
- внесите в систему элементы из других систем и организуйте их взаимодействие с проблемным.
2. Ученые доказали, что в аминокислотной последовательности белков имеет отношение только 10% ДНК. Как вы думаете, какие функции выполняют остальные 90% ДНК?
3. Известно, что у представителей различных видов организмов встречаются белки со сходной первичной структурой. Предложите объяснение этому факту.
Использованая литература:
- Бухвалов В.А. Общая методика развивающего образования.– Рига: Педагогический центр «Эксперимент», 2003 – 126 с.
- Бухвалов В.А, Мурашковский Ю.С. Изобретаем черепаху: как применить ТРИЗ в школьном курсе биологии. Книга для учителей и учащихся – Рига, «Эксперимент», 1993.
Метаболизм | Очерки биохимии
Фенилкетонурия (ФКУ) и дефицит ацил-КоА-дегидрогеназы со средней длиной цепи (MCADD) — два наиболее часто наследуемых нарушения обмена веществ, которым страдает примерно 1 из 10000 новорожденных в Великобритании.
ФКУ — это аминокислотное заболевание, вызванное дефицитом фермента фенилаланингидроксилазы, вызывающим ферментативный блок. Это приводит к снижению метаболизма аминокислоты фенилаланина, вызывая повышенное накопление в крови и головном мозге.Если не лечить новорожденных, это может вызвать задержку развития или повреждение головного мозга. Лечение начинается рано с диеты с низким содержанием белка, дополненной смесью аминокислот с удаленным фенилаланином. Однако небольшая часть людей с диагнозом ФКУ не отвечает на этот предложенный вид лечения. Эти люди обычно имеют дефекты синтеза дигидроптеридинредуктазы или биоптерина, вызывающие нарушение функции фенилаланингидроксилазы. Эти люди также обычно имеют дефекты тирозингидроксилазы, что может привести к дефициту нейротрансмиттеров.Затем этим пациентам требуются дополнительные добавки с нейротрансмиттерами, а также диета с низким содержанием фенилаланина.
MCADD — это пожизненное состояние, которое возникает из-за мутации ацил-CoA-дегидрогеназы со средней длиной цепи (MCAD) при β-окислении жирных кислот. Эта мутация нарушает расщепление жирных кислот со средней длиной цепи в ацетил-КоА. Потеря или недостаточность MCAD снижает окисление жирных ацил-CoA, которые содержат более шести атомов углерода, поскольку первая стадия дегидрирования β-окисления не может происходить.Используя тандемную масс-спектрометрию, можно увидеть, что профиль жирных кислот крови в MCADD показывает накопление C6, C8 и C10: 1. MCADD является основной причиной гипокетотической гипогликемии и может вызывать нарушение функции печени с метаболическим ацидозом, гипераммониемией и внезапной смертью. MCADD особенно опасен во время голодания, когда организм использует запасы гликогена, а свободные жирные кислоты высвобождаются из жировой ткани для получения энергии. Сниженная способность метаболизировать средние жирные кислоты значительно снижает доступность субстратов для кетогенеза, синтеза АТФ и цикла TCA при низкой энергии.Накопление промежуточных продуктов жирных кислот подавляет глюконеогенез, усугубляя гипогликемию. Это накопление может также способствовать сердечно-сосудистым и неврологическим осложнениям, обнаруживаемым в этих условиях. Лечение пациентов с MCADD включает потребление напитков с высоким содержанием сахара и избегание длительных периодов голодания.
Последнее наследственное нарушение обмена веществ, которое мы обсуждаем, встречается гораздо реже и встречается у 1 из 100 000 или 1 50000 новорожденных.Болезнь мочи кленового сиропа (MSUD) возникает из-за дефицита или снижения функции комплекса дегидрогеназы α-кетокислоты с разветвленной цепью (BCKAD). Это приводит к накоплению аминокислот с разветвленной цепью (BCAA), таких как лейцин, изолейцин и валин, в крови и моче. Название болезни происходит от запаха мочи кленового сиропа из-за избытка BCAA. BCAA потребляются в рационе, богатом белком, в таких продуктах, как мясо, рыба, яйца и молоко. Обычно избыточные аминокислоты расщепляются через аминотрансферазы с разветвленной цепью (BCAT) на α-кетокислоты в митохондриях.На второй стадии катаболизма комплекс BCKAD инициирует окислительное декарбоксилирование α-кетокислот, что приводит к образованию ацетоацетата, ацетил-КоА и сукцинил-КоА. Нормальное функционирование катаболизма аминокислот необходимо для синтеза белка, передачи клеточных сигналов и метаболизма глюкозы. BCKAD состоит из четырех субъединиц. Мутации в каталитических компонентах BCKAD снижают его активность и, следовательно, увеличивают уровни BCAA, проявляясь как MSUD и вызывая дисфункцию иммунной системы, скелетных мышц и центральной нервной системы.По мере накопления токсичных метаболитов, таких как молочная кислота и аммиак, функция иммунных клеток подавляется, вызывая нарушение их регуляции. Скелетные мышцы повреждены, как показали исследования, которые обнаружили уменьшение диаметра мышечных волокон и поражения миофибрилл у крыс MSUD, однако его механизм полностью не изучен. Нарушение регуляции нервной системы, в частности, поражение головного мозга, было связано с накоплением токсичных метаболитов. Однако исследования показали, что образование азот-активных форм у пациентов с MSUD может вызывать морфологические изменения в клетках глиомы C6.Кроме того, у пациентов с MSUD обнаруживаются маркеры окислительного повреждения белков, ДНК и липидов, возможно, в результате продукции свободных радикалов.
Происхождение и эволюция метаболических путей: почему и как первичные клетки построили метаболические пути? | Эволюция: образование и пропаганда
Увлекательно. Захватывающе. Реконструкция истории жизни на Земле представляет собой один из самых интригующих вопросов науки. И еще более интригующим является попытка понять (самые) первые молекулярные шаги, ведущие к первичным клеткам и их ранней эволюции.Существующие клетки представляют собой довольно сложные образования, состоящие из множества различных молекул, которые, однако, должны действовать и взаимодействовать согласованным образом, чтобы обеспечить выживание и воспроизводство клеток (и многоклеточных организмов). В каждый момент клеточной жизни миллиарды молекул превращаются в разные посредством реакций, которые ускоряются (катализируются) так называемыми ферментами, большинство из которых представлены белками. Несмотря на то, что эти белки могут взаимодействовать с множеством различных молекул во время своего хаотического путешествия внутри клетки, они связываются только с конкретными молекулами, представляющими их субстрат , и трансформируют его в другую, отличную молекулу, называемую продуктом (реакции).В целом это верно не для всех ферментов; каждый фермент взаимодействует с одним субстратом, в результате чего образуется определенный продукт. Следовательно, в каждый момент жизни клетки миллиардов субстратов превращаются в миллиардов продуктов с помощью миллиардов молекул ферментов. Эти реакции очень быстрые, и мы можем представить клетку как вязкую среду, в которой эти реакции протекают упорядоченным (и только кажущимся хаотическим) образом. Все эти реакции называются , метаболизм , круговая «сущность» в том смысле, что молекулы могут быть разрушены (катаболизм) для получения энергии и «кирпичиков», необходимых для создания других различных молекул (анаболизм) (рис.1). Таким образом, ясно, что внутри клетки существует «равновесие» между катаболическими и анаболическими реакциями. Таким образом, метаболизм сохранившихся клеток довольно сложен, но мы также можем считать его чрезвычайно упорядоченным. На рисунке 2 показан пример катаболической (разложение глюкозы во время гликолиза) и анаболической (биосинтез аминокислоты гистидина) систем. Как видно из рис. 2, и гликолиз, и биосинтез гистидина протекают по своего рода «каскаду» реакций, в которых разрушение глюкозы и образование гистидина требует последовательного действия различных ферментов, каждый из которых способен катализировать один шаг этого каскада.Набор реакций, начинающихся с субстрата и ведущих к конечному продукту реакции, называется метаболическим путем . В большинстве случаев каждый этап метаболического пути катализируется одним ферментом, который (в трети случаев) представляет собой один белок, кодируемый одним геном (Holliday et al. 2011).
Рис. 1
Схематическое изображение метаболических сетей, существующих в существующих клетках (из http://manet.illinois.edu/pathways.php)
Рис.2
Схематическое изображение катаболического (гликолиза) (из http://www.genome.jp/dbget-bin/www_bget?pathway+hsa00010) ( a ) и анаболического пути (биосинтез гистидина) (из http: //www.genome.jp/kegg/pathway/map/map00340.html) ( b ) в гамма-протеобактериях Escherichia coli K12
Если предположить, что существующие и очень сложные клетки произошли от гораздо более простых предковых клеток, можно также предположить, что последние обладали более простым метаболизмом по сравнению с существующим.Это, в свою очередь, означает, что они должны обладать гораздо более простыми геномами, которые, скорее всего, состоят из нескольких сотен генов. Если это так, то возникает вопрос: , почему и , как первичные клетки собрались и эволюционировали в своих метаболических путях? Вопрос можно перефразировать следующим образом: почему и как ранние клетки увеличили количество своих генов и сложность своих геномов? Ответ (ы), которые мы можем попытаться дать на эти вопросы, явно зависит от условий примитивной Земли и того, как выглядели первобытные живые существа.Однако это одна из самых туманных проблем; на самом деле, хотя были предприняты значительные усилия, чтобы понять появление первых живых существ, мы до сих пор не знаем, когда и как возникла жизнь (Peretò et al. 1998). Тем не менее, обычно считается, что первые организмы возникли и заселили водную среду (океаны, реки, пруды и т. Д.), Богатую органическими соединениями, спонтанно образовавшимися в пребиотическом мире. Это гетеротрофное происхождение жизни принято считать и часто называют теорией Опарина – Холдейна (Oparin 1924; Lazcano and Miller 1996).Если эта идея верна, жизнь произошла из изначального супа, содержащего различные органические молекулы (многие из которых используются существующими формами жизни). Этот суп из питательных веществ был доступен для ранних гетеротрофных организмов, поэтому им приходилось проводить минимум биосинтеза. Экспериментальная поддержка этого предложения была получена в 1953 году, когда Миллер (1953) и Юри показали, что аминокислоты и другие органические молекулы образуются в атмосферных условиях, которые, как считается, являются типичными для тех, что существовали на ранней Земле.Первые живые системы, вероятно, действительно произошли непосредственно от изначального бульона и относительно быстро эволюционировали до общего предка, обычно называемого Последним универсальным общим предком (LUCA), сущности, представляющей отправную точку дивергенции всех существующих на Земле форм жизни. (Рис. 3). Если мы предположим, что жизнь возникла в пребиотическом супе, содержащем большую часть, если не все, необходимых малых молекул, то можно предположить большую потенциальную доступность питательных веществ на примитивной Земле, обеспечивающую как рост, так и снабжение энергией большого количества предковые организмы.Мы можем представить себе существование «раннего плавающего живого мира», состоящего из первичных клеток, которые могли бы выглядеть как «мыльные пузыри», вмещающие одну или несколько информационных молекул и выполняющие ограниченное количество метаболических реакций. Эти пузыри были способны делиться, взаимодействовать друг с другом, сливаться и делиться своими геномами и метаболическими способностями, давая начало прогрессивно сложным живым существам. Если этот сценарий верен, то есть первобытные организмы были гетеротрофными и не нуждались в развитии новых и улучшенных метаболических способностей, поскольку большинство необходимых питательных веществ были доступны, мы можем вернуться к двум вопросам, на которые можно ответить, а именно: почему и , как первичные клетки расширили свои метаболические способности и геномы?
Фиг.3
Ориентировочная эволюционная шкала времени от происхождения Земли до разнообразия жизни
Ответ на первый вопрос довольно интуитивно понятен. Действительно, увеличение числа ранних клеток, процветающих на первичном бульоне, привело бы к истощению основных питательных веществ, создавая все более сильное селективное давление, которое, в свою очередь, благоприятствовало (в дарвиновском смысле) тем микроорганизмам, которые стали способны синтезировать эти молекулы. концентрация которого в изначальном супе уменьшалась.Следовательно, происхождение и эволюция основных метаболических путей представляет собой решающий шаг в молекулярной и клеточной эволюции, поскольку это сделало первичные клетки менее зависимыми от экзогенных источников питательных веществ (рис. 4).
Рис. 4
Схематическое изображение сообщества предковых клеток с давлением отбора, допускающим приобретение и распространение нового метаболического признака (изменено из Fondi et al. 2009a)
Но как произошло расширение геномов? В следующем разделе мы сосредоточимся на молекулярных механизмах, которые управляли этим переходом, т.е.е., расширение и уточнение древних метаболических путей, приводящее к структуре существующих метаболических путей.
Регуляция системного метаболизма вегетативной нервной системой, состоящая из афферентной и эфферентной иннервации | Международная иммунология
Абстрактные
Вегетативные нервы, симпатические и парасимпатические, иннервируют органы и регулируют их функции. Стало очевидно, что афферентные и эфферентные сигналы вегетативной нервной системы играют важную роль в регулировании системного метаболизма, тем самым поддерживая гомеостаз на уровне всего тела.Афферентные нервы блуждающего нерва получают сигналы, такие как питательные вещества и гормоны, от периферических органов / тканей, включая желудочно-кишечный тракт и жировую ткань, а затем передают эти сигналы в гипоталамус, регулируя тем самым пищевое поведение. В дополнение к роли в контроле аппетита области в гипоталамусе служат регулирующими центрами как симпатических, так и парасимпатических эфферентных волокон. Эти эфферентные иннервации регулируют функции периферических органов / тканей, таких как островки поджелудочной железы, жировая ткань и печень, которые играют роль в регуляции метаболизма.Более того, недавние данные раскрыли метаболические регуляторные системы, управляемые вегетативными нервными цепями. В этих системах афферентные нервы передают метаболическую информацию от периферических органов в центральную нервную систему (ЦНС), и тем самым ЦНС регулирует функции органа через эфферентные волокна вегетативных нервов. Таким образом, вегетативная нервная система регулирует гомеостаз системного метаболизма, а афферентные и эфферентные волокна играют решающую роль в его регуляции. Кроме того, несколько линий доказательств демонстрируют роль вегетативной нервной системы в регулировании и дисрегуляции иммунной системы.В этом обзоре представлены различные нейронно-опосредованные системы перекрестного взаимодействия органов и систематизированы текущие знания о вегетативном контроле / координации системного метаболизма, уделяя особое внимание вегетативной нервной цепи печень-мозг-панкреатические β-клетки, а также выделяя потенциальная важность связей с нейрональной и иммунной системами.
Введение
Функции органов регулируются не только гуморальными факторами, но и нервными сигналами.Вегетативные нервы, включая симпатические и парасимпатические нервы, иннервируют органы и модулируют функцию органов, регулируя тем самым системный метаболизм.
Пучки вегетативных нервов содержат как афферентные волокна от периферических органов к центральной нервной системе (ЦНС), так и эфферентные волокна от ЦНС к периферическим органам. Афферентные и эфферентные сигналы передают метаболическую информацию от периферических органов к ЦНС и модулируют функции периферических внутренних органов, соответственно, что приводит к регуляции системного метаболизма, а также к поддержанию гомеостаза глюкозы и энергии.
Например, афферентные блуждающие нервы передают сигналы, полученные из желудочно-кишечного тракта, такие как глюкагоноподобный пептид-1 (GLP-1) (1–4), холецистокинин (CCK) (5, 6), грелин (7). , 8) и механические сигналы, тем самым регулируя режим кормления. Афферентные блуждающие нервы также передают сигналы от гепатопортальной системы, регулируя тем самым метаболизм глюкозы (9). Между тем эфферентные блуждающие нервы регулируют системный гомеостаз глюкозы, усиливая глюкозо-стимулированную секрецию инсулина (GSIS) из β-клеток (10) и активируя синтез гликогена в печени (11).С другой стороны, эфферентные симпатические нервы подавляют секрецию инсулина β-клетками (12, 13) и способствуют выработке глюкозы печенью (14, 15). Кроме того, эфферентные симпатические нервы участвуют в регуляции энергетического метаболизма, способствуя липолизу в белой жировой ткани (WAT) (16) и индуцируя термогенез в коричневой жировой ткани (BAT) (17).
Более того, все больше данных указывает на то, что вегетативные нервные цепи, состоящие из афферентных и эфферентных сигналов, играют важную роль в регуляции глюкозы и энергетического метаболизма.В этих системах афферентные нервы передают метаболическую информацию от периферических органов в ЦНС, и тем самым ЦНС регулирует функции органов через эфферентные волокна вегетативных нервов. Напр., Межорганная автономная цепь β-клеток печени и поджелудочной железы, состоящая из афферентных внутренних нервов от печени и эфферентных блуждающих нервов к поджелудочной железе, регулирует пролиферацию β-клеток поджелудочной железы (18, 19). Кроме того, вегетативные нервные цепи, состоящие из блуждающих афферентных нервов, иннервирующих печень, и эфферентных симпатических нервов, ведущих к жировой ткани, регулируют энергетический обмен (20–23).Между тем, вегетативные нервные цепи, состоящие из блуждающих афферентных нервов, иннервирующих желудочно-кишечный тракт, и эфферентных блуждающих нервов, ведущих к печени, участвуют в производстве глюкозы в печени (24, 25).
В этом обзоре освещаются эти регуляторные механизмы системного метаболизма вегетативными нервами, включая межорганные системы перекрестного взаимодействия, состоящие как из афферентных, так и из эфферентных волокон.
Роль гипоталамуса в регулировании аппетита
В гипоталамусе находится регуляторный центр вегетативных нервов, включая блуждающие нервы.Несколько ядер в гипоталамусе, например дугообразное ядро (ARH), вентромедиальное ядро (VMH) и латеральная область гипоталамуса (LHA) играют важную роль в регуляции системного метаболизма (26).
ARH жизненно важен для контроля кормления. В ARH присутствуют две основные субпопуляции нейронов, регулирующих аппетит. Нейроны, экспрессирующие проопиомеланокортин (POMC), участвуют в стимулировании анорексигенных ответов, тогда как нейроны, экспрессирующие агути-родственный пептид и / или нейропептид Y, способствуют орексигенным ответам (27).Нейроны ARH экспрессируют рецепторы анорексигенных гуморальных факторов, таких как лептин и инсулин, которые секретируются в основном из WAT и островков поджелудочной железы соответственно (28, 29). Нейроны ARH также экспрессируют рецепторы грелина, орексигенного гуморального фактора, который секретируется в основном из желудка (28, 29).
В дополнение к этим гуморальным факторам, регулирующим аппетит, нейроны ARH получают входные данные от солитарного ядра (NTS) ствола головного мозга, по которому передается метаболическая информация от периферических органов (рис.1). Гуморальные факторы, регулирующие аппетит, такие как GLP-1, грелин и CCK, влияют на активность нейронов NTS через афферентные блуждающие нервы как часть парасимпатической нервной системы (28, 29). Таким образом, аппетит сильно регулируется комбинацией этих многочисленных входов в нейроны ARH.
Рис. 1.
Анатомия афферентных блуждающих нервов, связанных с системным метаболизмом. Нейроны ARH получают входные данные от NTS ствола мозга, по которому передается метаболическая информация от периферических органов, таких как части желудочно-кишечного тракта и воротной вены печени.
Рис. 1.
Анатомия афферентных блуждающих нервов, связанных с системным метаболизмом. Нейроны ARH получают входные данные от NTS ствола мозга, по которому передается метаболическая информация от периферических органов, таких как части желудочно-кишечного тракта и воротной вены печени.
Регулирование пищевого поведения афферентными сигналами из желудочно-кишечного тракта
Пучки вегетативных нервов содержат как эфферентные, так и афферентные нервные волокна (30, 31), а афферентные нейрональные сигналы играют важную роль в регуляции системного метаболизма.В частности, как упоминалось выше, афферентные нервы блуждающего нерва получают сигналы, полученные из желудочно-кишечного тракта, такие как сигналы, запускаемые питательными веществами и гормонами, и эти сигналы передаются в NTS в заднем мозге (32, 33).
Афферентные нейроны блуждающего нерва биполярны, и их аксоны возникают из тел клеток узлового ганглия (NG), причем один конец заканчивается в заднем мозге, а другой — в периферических органах (33). Афферентные аксоны блуждающего нерва обнаруживаются по всему желудочно-кишечному тракту, включая желудок, двенадцатиперстную кишку, тонкий и толстый кишечник, и известно, что афферентные сигналы из желудочно-кишечного тракта передаются в NTS (34).Недавно было показано, что энтероэндокринные клетки, расположенные в желудочно-кишечном тракте, находятся в прямом контакте с афферентными нервами блуждающего нерва через синапсоподобные структуры (35, 36). Некоторые гормоны кишечника, включая GLP-1, CCK и грелин, являются хорошо известными передатчиками регулирующих аппетит сигналов из желудочно-кишечного тракта через афферентный блуждающий нерв, систему, называемую осью кишечник-мозг (рис. 2).
Рис. 2.
Регуляция системного метаболизма посредством афферентной иннервации вегетативных нервов.Афферентные нервы блуждающего нерва получают поступающие из желудочно-кишечного тракта сигналы, такие как питательные вещества и гормоны, и передают эти сигналы к NTS в заднем мозге через ось кишечник-мозг. Блуждающие афферентные нервы от воротной вены печени участвуют в регуляции метаболизма глюкозы. Внутрипортальное введение глюкозы увеличивает клиренс глюкозы во всем организме. Этот эффект, по-видимому, опосредуется афферентными волокнами блуждающего нерва, иннервирующими воротную вену, и называется «гепатопортальным датчиком глюкозы».
Фиг.2.
Регуляция системного метаболизма посредством афферентной иннервации вегетативных нервов. Афферентные нервы блуждающего нерва получают поступающие из желудочно-кишечного тракта сигналы, такие как питательные вещества и гормоны, и передают эти сигналы к NTS в заднем мозге через ось кишечник-мозг. Блуждающие афферентные нервы от воротной вены печени участвуют в регуляции метаболизма глюкозы. Внутрипортальное введение глюкозы увеличивает клиренс глюкозы во всем организме. Этот эффект, по-видимому, опосредуется афферентными волокнами блуждающего нерва, иннервирующими воротную вену, и называется «гепатопортальным датчиком глюкозы».
Энтероэндокринные L-клетки, расположенные в дистальном отделе тощей кишки и подвздошной кишки, секретируют GLP-1 в ответ на поступающие питательные вещества (29). GLP-1 вызывает чувство насыщения, напрямую проникая в мозг и косвенно передавая сигналы через афферентные блуждающие нервы (рис. 2) (37). Рецептор GLP-1 экспрессируется в нейрональных клетках NG, а GLP-1 проявляет свои эффекты через рецепторы GLP-1 на нервных окончаниях, иннервирующих желудочно-кишечный тракт (33, 38). Эффекты внутрибрюшинной (внутрибрюшинной) инъекции GLP-1 на снижение потребления пищи и активацию нейронов ARH подавляются ваготомией как у грызунов (1), так и у людей (2).
Дополнительно либо i.p. введение антагониста рецептора GLP-1 (3) или селективного нокдауна рецептора GLP-1 в афферентных нейронах блуждающего нерва (4) ингибирует аноректический эффект i.p. вводили GLP-1. Эти результаты подтверждают идею о том, что сигнал GLP-1 передается из желудочно-кишечного тракта в ЦНС через афферентные блуждающие нервы. Недавно d-аллюлоза, некалорийный подсластитель, была идентифицирована как выдающийся агент, высвобождающий GLP-1. d-аллюлоза снижает потребление пищи в моделях на животных с ожирением, и этот эффект опосредуется GLP-1, который действует на афферентные блуждающие нервы (39).
CCK секретируется клетками двенадцатиперстной кишки и подвздошной кишки в ответ на такие питательные вещества, как насыщенные жирные кислоты, длинноцепочечные жирные кислоты (40) и аминокислоты (41). CCK связывается с рецепторами CCK1, расположенными на окончаниях афферентных нервов блуждающего нерва (42). Избирательное рассечение афферентных блуждающих нервов посредством вентрального хирургического доступа к стволу головного мозга (5) или системного введения капсаицина (6), специфического нейротоксина для сенсорных нервных волокон, устраняет эффект сытости от i.p. вводили CCK крысам.Таким образом, афферентные нервы блуждающего нерва могут также передавать сигналы сытости, полученные от периферических CCK, в NTS (рис. 2).
Грелин в основном секретируется желудком и вызывает чувство голода. На сегодняшний день установлено, что грелин является единственным орексигенным сигналом от желудочно-кишечного тракта. Уровни грелина в крови повышаются перед едой и снижаются после еды (43). Рецепторы грелина локализуются на афферентных нервах блуждающего нерва (7), а сигналы грелина из желудка передаются в NTS через блуждающие нервы (рис.2) (44, 45). Ваготомия или периферическое применение капсаицина устраняет индуцированное грелином кормление у крыс (7). Кроме того, периферически вводимый грелин не стимулирует прием пищи у пациентов, перенесших полную туловищную ваготомию (8). Таким образом, неповрежденные блуждающие нервы, вероятно, необходимы для индукции сигналов голода периферически вводимым грелином.
Механические сигналы, такие как растяжение и изменение объема органов желудочно-кишечного тракта, также могут активировать афферентные нервы блуждающего нерва, тем самым влияя на аппетит (46, 47).В совокупности многие сигналы, вызванные проглоченной пищей, передаются через ось кишечник-мозг и координированно регулируют пищевое поведение.
Регуляция органов обмена веществ посредством эфферентной иннервации вегетативных нервов
И VMH, и LHA участвуют в регуляции аппетита. С другой стороны, эти ядра также служат вегетативными нервными центрами и регулируют метаболизм периферических органов (48, 49). Нервные волокна, возникающие из VMH, достигают продолговатого мозга через средний мозг и, следовательно, соединяются с преганглионарными симпатическими нервными волокнами.Преганглионарные симпатические нервные волокна соединяются с постганглионарными симпатическими нервными волокнами в симпатических ганглиях, расположенных рядом со спинным мозгом. Между тем, нервные волокна, отходящие от LHA, соединяются с дорсальным двигательным ядром блуждающего нерва (DMV) в продолговатом мозге, откуда берут начало парасимпатические нервы. Преганглионарные блуждающие нервы соединяют постганглионарные блуждающие волокна в парасимпатических ганглиях, расположенных в периферических органах (рис. 3).
Рис. 3.
Анатомия эфферентных симпатических и парасимпатических нервов, связанных с системным метаболизмом.Нервные волокна, возникающие из VMH, достигают продолговатого мозга через средний мозг и, следовательно, соединяются с преганглионарными симпатическими нервными волокнами. Преганглионарные симпатические нервные волокна соединяются с постганглионарными симпатическими нервными волокнами в симпатических ганглиях, расположенных рядом со спинным мозгом. Между тем нервные волокна, отходящие от LHA, соединяются с DMV в продолговатом мозге, откуда берут начало парасимпатические нервы. Преганглионарные блуждающие нервы соединяют постганглионарные блуждающие волокна в парасимпатических ганглиях, расположенных в периферических органах.
Рис. 3.
Анатомия эфферентных симпатических и парасимпатических нервов, связанных с системным метаболизмом. Нервные волокна, возникающие из VMH, достигают продолговатого мозга через средний мозг и, следовательно, соединяются с преганглионарными симпатическими нервными волокнами. Преганглионарные симпатические нервные волокна соединяются с постганглионарными симпатическими нервными волокнами в симпатических ганглиях, расположенных рядом со спинным мозгом. Между тем нервные волокна, отходящие от LHA, соединяются с DMV в продолговатом мозге, откуда берут начало парасимпатические нервы.Преганглионарные блуждающие нервы соединяют постганглионарные блуждающие волокна в парасимпатических ганглиях, расположенных в периферических органах.
В условиях голодания симпатические нервы активируются у животных, чтобы поддерживать снабжение органов питательными веществами (50). Симпатические нервы повышают уровень питательных веществ в крови, включая липиды и глюкозу, что позволяет тканям, таким как мозг и мышцы, легче усваивать эти питательные вещества (51). Одновременно симпатические нервы усиливают кровоток, увеличивая частоту сердечных сокращений и артериальное давление, что способствует притоку питательных веществ в органы (52).
Напротив, парасимпатические нервы способствуют хранению гликогена в печени (48), чтобы обеспечить организм глюкозой в случае чрезвычайных ситуаций или голода в будущем. Кроме того, парасимпатические нервы, по-видимому, усиливают некоторые реакции после кормления, такие как подвижность гладких мышц ЖКТ, секреция пищеварительной жидкости и атоничность препилорического сфинктера, тем самым способствуя всасыванию проглоченной пищи (53).
В следующих разделах описывается регуляция каждого метаболического органа посредством эфферентной иннервации вегетативных нервов.
Островки поджелудочной железы
Инсулин — это гормон, который снижает уровень глюкозы в крови и секретируется β-клетками островков поджелудочной железы. Инсулин вызывает анаболические реакции, способствуя захвату глюкозы периферическими тканями и подавляя выработку глюкозы в печени.
Островки поджелудочной железы обильно иннервируются симпатическими и парасимпатическими нервами как у людей (54), так и у грызунов (55). Парасимпатические и симпатические нервы, иннервирующие островки поджелудочной железы, являются блуждающими и чревными, соответственно, и эти вегетативные нервы участвуют в регуляции секреции инсулина (рис.4) (10).
Рис. 4.
Регуляция метаболических органов посредством эфферентной иннервации вегетативных нервов. В целом симпатические нервы увеличивают количество питательных факторов крови, таких как липиды и глюкоза, за счет стимулирования липолиза и глюконеогенеза. Это позволяет органам, таким как мозг и мышцы, легче поглощать питательные вещества в чрезвычайных ситуациях или в условиях голода. Подавление секреции инсулина косвенно участвует в этих процессах. Напротив, парасимпатические нервы подавляют глюконеогенез и способствуют хранению гликогена в печени, обеспечивая доставку питательных веществ всему телу в случае чрезвычайной ситуации в будущем.В эти процессы косвенно участвует стимуляция секреции инсулина.
Рис. 4.
Регуляция метаболических органов посредством эфферентной иннервации вегетативных нервов. В целом симпатические нервы увеличивают количество питательных факторов крови, таких как липиды и глюкоза, за счет стимулирования липолиза и глюконеогенеза. Это позволяет органам, таким как мозг и мышцы, легче поглощать питательные вещества в чрезвычайных ситуациях или в условиях голода. Подавление секреции инсулина косвенно участвует в этих процессах.Напротив, парасимпатические нервы подавляют глюконеогенез и способствуют хранению гликогена в печени, обеспечивая доставку питательных веществ всему телу в случае чрезвычайной ситуации в будущем. В эти процессы косвенно участвует стимуляция секреции инсулина.
Во время кормления ацетилхолин высвобождается из окончаний блуждающего нерва, тем самым усиливая GSIS (56, 57) из β-клеток посредством механизма, зависимого от мускаринового рецептора 3 (M3R) (58). M3R представляет собой Gq-связанный рецептор, связанный с G-белком (GPCR), и связанная с M3R фосфолипаза C опосредует гидролиз фосфатидилинозитол-4,5-бисфосфата (PIP2) в плазматической мембране при стимуляции ацетилхолином.Гидролиз PIP2 приводит к образованию 1,4,5-инозитолтрифосфата (IP3) (59, 60). IP3, который представляет собой гидрофильную молекулу, диффундирует в цитоплазму и связывается с рецептором IP3 на мембране эндоплазматического ретикулума (ER), что приводит к высвобождению Ca 2+ из ER. Результирующее увеличение внутриклеточной концентрации Ca 2+ способствует GSIS (61–63). Блуждающие нервы также индуцируют секрецию глюкагона α-клетками островков поджелудочной железы (64, 65), вероятно, посредством немускаринового механизма (рис.4) (66).
Известно, что блуждающие нервы изменяют секрецию инсулина не только тогда, когда проглоченная пища достигает желудочно-кишечного тракта, но также и до того, как пища проглочена. Секреция инсулина после и до приема пищи называется кишечной и головной фазами секреции инсулина соответственно. В головной фазе блуждающие нервы стимулируются сигналами, вызываемыми зрительными, обонятельными и вкусовыми ощущениями (67, 68). Секреция инсулина в головной фазе может быть необходима для быстрого регулирования метаболизма глюкозы после приема пищи.Действительно, ранее сообщалось о значении секреции головного инсулина в регуляции уровня глюкозы в крови после еды у людей (69, 70).
Островки поджелудочной железы также иннервируются внутренними нервами, которые являются симпатическими. Симпатические нервы выделяют норадреналин, катехоламин, который стимулирует секрецию глюкагона α-клетками в островках поджелудочной железы (71, 72), подавляя секрецию инсулина β-клетками через адренергические рецепторы (рис. 4) (12, 13). Адренергические рецепторы представляют собой GPCR и подразделяются на подсемейства α-адренергических и β-адренергических рецепторов.Α-адренорецепторы и β-адренорецепторы далее делятся на несколько подтипов, и связанные G-белки различаются между этими подтипами. На β-клетках α-адренорецепторы экспрессируются в большей степени, чем β-адренорецепторы (73). Электрическая стимуляция внутренних нервов подавляет GSIS, а альфа-блокатор фентоламин подавляет это подавление (74). Кроме того, для подавления GSIS достаточно селективного агониста α2-адренорецепторов, клонидина (75). Следовательно, подавление секреции инсулина адренергическими сигналами опосредуется механизмом, зависимым от α2-адренорецепторов.
Хотя роль симпатической иннервации в базальной или постпищевой секреции инсулина остается неясной, симпатические нервы могут предотвращать снижение уровня глюкозы в крови во время физических упражнений или в гипогликемических состояниях, подавляя секрецию инсулина и способствуя секреции глюкагона (76).
Жировая ткань
Жировая ткань в высокой степени регулируется как нервной, так и иммунной системами; последний — за счет секреции цитокинов, таких как IL-6, TNF-α, IL-10 и антагониста рецептора IL-1 (IL-1Ra) (77).Жировая ткань подразделяется на WAT и BAT. Цитоплазма белых адипоцитов богата липидами, тогда как коричневые адипоциты содержат большое количество митохондрий и меньшее количество липидов, чем WAT. Когда животные испытывают нехватку энергии, норадреналин высвобождается из симпатических нервных окончаний, что приводит к липолизу триацилглицерина (TG) до свободных жирных кислот (FFA) и глицерина в WAT. Этот липолитический ответ повышает концентрацию в крови питательных веществ, полученных из жировой ткани, которые затем распределяются по различным органам (48).
В WAT симпатические нервы вызывают липолитические эффекты через β-адренорецепторы (рис. 4). β1-адренергические, β2-адренергические и β3-адренергические рецепторы все экспрессируются в WAT (78–80). Норадреналин, который высвобождается из симпатических нервных окончаний, активирует аденилатциклазу и повышает уровень цитоплазматического цАМФ. Следовательно, активируется цАМФ-зависимая протеинкиназа А, что приводит к активации гормоночувствительной липазы. Эта липаза действует как фермент, ограничивающий скорость липолитического ответа, который гидролизует TG до FFA и глицерина (16).
BAT более плотно иннервируется симпатическими нервами, чем WAT. Когда животным необходимо поддерживать температуру тела, например, при воздействии холода, симпатические нервы вызывают термогенез BAT посредством механизма, зависимого от β3-адренорецепторов (рис. 4) (17). Хотя ранее считалось, что у людей BAT существует только в младенчестве и исчезает в зрелом возрасте, несколько исследовательских групп показали, что функционально активные BAT существуют и у взрослых людей (81–84).Поскольку количество BAT обратно пропорционально индексу массы тела у людей (81–84), BAT предположительно участвует в энергетическом гомеостазе.
Недавно был открыт новый тип адипоцитов, «бежевый адипоцит» (85). Бежевые клетки появляются в WAT в ответ на различные стимулы, такие как простуда или агонист β3-адренорецепторов (85). Исследователи уделяют большое внимание бежевым адипоцитам в связи с их значением в патофизиологии ожирения у людей (86–88).
Неясно, иннервируют ли парасимпатические волокна WAT (89).Kreier и др. . показали, что инъекция вируса псевдобешенства или флюороголда, двух ретроградных транс-нейрональных индикаторов, во внутрибрюшные жировые подушечки крыс маркирует DMV, предполагая, что WAT иннервируется блуждающими нервами (90). Однако Джордано и др. . не обнаружили гистологических свидетельств маркеров блуждающего нерва, таких как везикулярные переносчики ацетилхолина или вазоактивный кишечный пептид (VIP), в WAT (91). Таким образом, в настоящее время роль вагусных сигналов в WAT остается спорной.
Печень
Сигналы симпатических нервов прямо и косвенно способствуют выработке глюкозы в печени, тем самым повышая концентрацию глюкозы в крови (рис. 4). Электрическая стимуляция внутренних нервов, которые являются симпатическими и иннервируют печень, усиливает гликогенолиз за счет активации гликогенфосфорилазы и глюкозо-6-фосфатазы, что приводит к повышению концентрации глюкозы в крови (14, 15). Норадреналин, полученный из внутренних нервов, оказывает свое действие на печень непосредственно через α1-адренорецепторы, которые представляют собой Gq-связанные GPCR (92), экспрессируемые в гепатоцитах.Кроме того, симпатические нервы способствуют секреции глюкагона (как упоминалось выше) и адреналина α-клетками поджелудочной железы и мозговым веществом надпочечников, соответственно, косвенно повышая выработку глюкозы в печени. Поскольку адреналин, полученный из мозгового вещества надпочечников, проявляет свои эффекты через β2-адренергический рецептор (92), катехоламины, вероятно, способствуют производству глюкозы в печени в координации с передачей сигналов как α1-адренергических, так и β2-адренергических рецепторов в печени.
Парасимпатические нервные сигналы способствуют синтезу гликогена в печени (рис.4). Парасимпатические волокна, иннервирующие печень, блуждающие. Сигналы блуждающего нерва активируют гликогенсинтазу (11), фермент, ограничивающий скорость пути синтеза гликогена. M3R представляет собой подтип мускариновых рецепторов, экспрессирующийся в основном в гепатоцитах мышей. Однако никаких значительных метаболических различий, включая содержание гликогена в печени, не наблюдалось у мышей, лишенных M3R в гепатоцитах (93). Таким образом, выяснение молекулярных механизмов, с помощью которых сигналы блуждающего нерва способствуют синтезу гликогена в печени, требует дальнейшего изучения.
Сигналы блуждающего нерва участвуют в регуляции регенерации печени. Гепатоциты заметно разрастаются после травмы, особенно на ранней стадии регенерации печени. Сообщается, что эта выраженная пролиферация гепатоцитов подавляется ваготомией по ветви печени (94, 95). Молекулярный механизм, лежащий в основе регенерации печени, опосредованной сигналами блуждающего нерва, был недавно выяснен (96). Сигналы блуждающего нерва способствуют высвобождению IL-6 из печеночных макрофагов, а IL-6 активирует boxM1 вилки гепатоцитов (FoxM1), фактор транскрипции, который имеет решающее значение для развития клеточного цикла и способствует регенерации печени.Примечательно, что этот механизм обеспечивает выживаемость после тяжелого повреждения органа (96). Таким образом, этот многоступенчатый механизм с участием блуждающих нервов способствует быстрой регенерации печени, тем самым поддерживая выживание всего организма после тяжелого повреждения печени.
Регуляция системного метаболизма афферентными сигналами гепатопортальной системы
Афферентные нервы блуждающего нерва иннервируют не только органы желудочно-кишечного тракта, но и воротную вену печени. Введение в воротную вену питательных веществ и гуморальных факторов, таких как глюкоза (97), аминокислоты (98, 99) и GLP-1 (100), влияет на афферентные блуждающие нервы.Внутрипортальное введение глюкозы увеличивает клиренс глюкозы во всем организме. Этот эффект, по-видимому, опосредован афферентными волокнами блуждающего нерва, иннервирующими воротную вену, которые составляют «гепатопортальный датчик глюкозы» (рис. 2) (9). Этот сенсор увеличивает системную утилизацию глюкозы, стимулируя ее усвоение печенью (101), блокируя секрецию как адреналина, так и норадреналина (102, 103) и стимулируя утилизацию глюкозы мышечными тканями (104, 105).
Интересно, что недавнее исследование показало, что интрапортальное введение l-аргинина активирует нейрональные клетки коры островка и что эффект l-аргинина блокируется перерезкой общей печеночной ветви блуждающего нерва (106).Эти результаты показывают, что сигналы о питательных веществах, передаваемые из воротной вены, достигают не только гипоталамуса, но и коры головного мозга. Это исследование также показало, что интрапортальная инфузия CCK активировала нейрональные клетки в дорсомедиальном гипоталамусе, но не в коре островка, и это действие CCK было отменено ваготомией. Соответственно, афференты блуждающего нерва активируют различные области мозга в зависимости от периферических стимулов. В этом контексте молекулярное разнообразие афферентных нейронов блуждающего нерва, иннервирующих органы желудочно-кишечного тракта, недавно было выяснено с помощью оптогенетических методов, визуализации in vivo, NG и генетически управляемого анатомического картирования (107).Таким образом, афферентные нервы блуждающего нерва хорошо настроены для передачи определенной информации от периферических органов.
Регуляция системного метаболизма афферентными сигналами от WAT
Как упоминалось выше, WAT получает симпатическую иннервацию, и механизмы липолиза, опосредованного эфферентными симпатическими сигналами, интенсивно изучаются. С другой стороны, лишь несколько исследований были сосредоточены на афферентных нервных сигналах от жировой ткани. Было продемонстрировано, что нервные афферентные сигналы от внутрибрюшной жировой ткани к ЦНС модулируют потребление пищи, влияя на чувствительность гипоталамуса к лептину (108).Опосредованная аденовирусом экспрессия разобщающего белка 1 (UCP1) в жировых подушках придатка яичка мышей улучшает резистентность гипоталамуса к лептину, вызванную кормлением с высоким содержанием жиров (HFD). Местное расслоение или фармакологическая деафферентация нервов от жировых подушечек придатка яичка устраняет повышенную HFD-индуцированную резистентность к лептину в ответ на экспрессию UCP1 жировой ткани (108). Это исследование представило внутрибрюшные жировые ткани как орган, регулирующий потребление пищи не только адипокинами, но и сигналами нейронов.
Регуляция системного метаболизма с помощью нейронального реле из печени, состоящего из афферентных и эфферентных нервов
Концепция, согласно которой реле афферентных и эфферентных сигналов играют важную роль в регулировании метаболизма глюкозы и энергии на уровне всего тела, была недавно предложена и получает признание.
Растущее количество доказательств, включая результаты, полученные в нашей лаборатории, показывают, что печень играет роль не только как приемник, но и как передатчик нейронных сигналов в вегетативных нервных цепях, состоящих как из афферентных, так и из эфферентных волокон.Во-первых, несколько исследований показали, что вегетативные нервные цепи участвуют в энергетическом обмене. Опосредованная аденовирусом экспрессия рецептора γ2, активируемого пролифератором пероксисом (PPARγ2), в печени приводит к накоплению липидов в печени, снижению периферического ожирения и повышению периферической чувствительности к инсулину и толерантности к глюкозе. Эти фенотипы притупляются хирургической денервацией или селективной фармакологической деафферентацией блуждающих волокон, иннервирующих печень, или введением блокатора β-адренорецепторов.Таким образом, экспрессия PPARγ2 в печени, то есть накопление липидов в печени, стимулирует афферентные блуждающие нервы, передавая метаболическую информацию в ЦНС, и тем самым предотвращает развитие ожирения и периферической инсулинорезистентности через эфферентные симпатические нервные пути (рис. 5A) (20).
Рис. 5.
Регуляция системного метаболизма вегетативными нервными цепями, состоящими как из эфферентных, так и из афферентных волокон. (A) Печень служит передатчиком нейронных сигналов в вегетативных нервных цепях, состоящих как из афферентных, так и из эфферентных волокон вегетативных нервов, и участвует в энергетическом обмене и регуляции артериального давления.Стрелки одного цвета указывают на отдельный вегетативный нервный контур. (B) Блуждающие афферентные нервы, иннервирующие желудочно-кишечный тракт, передают метаболическую информацию блуждающим эфферентным нервам, иннервирующим печень на уровне ЦНС, составляя цепь автономных нервов кишечник-мозг-печень. Этот нервный контур регулирует выработку глюкозы в печени.
Рис. 5.
Регуляция системного метаболизма вегетативными нервными цепями, состоящими как из эфферентных, так и из афферентных волокон. (A) Печень служит передатчиком нейронных сигналов в вегетативных нервных цепях, состоящих как из афферентных, так и из эфферентных волокон вегетативных нервов, и участвует в энергетическом обмене и регуляции артериального давления.Стрелки одного цвета указывают на отдельный вегетативный нервный контур. (B) Блуждающие афферентные нервы, иннервирующие желудочно-кишечный тракт, передают метаболическую информацию блуждающим эфферентным нервам, иннервирующим печень на уровне ЦНС, составляя цепь автономных нервов кишечник-мозг-печень. Этот нервный контур регулирует выработку глюкозы в печени.
Поскольку экспрессия PPARγ2 в печени физиологически связана с ожирением, это нейронное реле, по-видимому, действует как защитный механизм против избыточного потребления калорий.С другой стороны, этот нервный контур, как сообщается, также участвует в повышении артериального давления во время развития ожирения (рис. 5A) (109). Таким образом, по иронии судьбы, система обратной связи для поддержания энергетического гомеостаза вызывает гипертензию, главный фенотип метаболического синдрома.
Афферентные волокна блуждающего нерва, иннервирующие печень, также участвуют в индуцированной глюкокортикоидами инсулинорезистентности и гипертензии. Фармакологические или хирургические нарушения афферентных волокон блуждающего нерва, иннервирующих печень, предотвращают эти индуцированные глюкокортикоидами метаболические фенотипы.Печеночная экспрессия PPARα в этом механизме, по-видимому, участвует в активации афферентных блуждающих нервов (рис. 5A) (110).
Кормление HFD увеличивает экспрессию печеночной глюкокиназы (GK) у мышей. Опосредованная аденовирусом экспрессия GK в печени снижает адаптивный термогенез за счет уменьшения связанных с термогенезом генов в BAT. Повышающая регуляция GK в печени снижает как активность нейронов в ядре бледного шва, симпатического центра, предположительно ответственного за термогенез BAT, так и оборот норадреналина в BAT.Кроме того, хирургическая денервация блуждающих волокон, иннервирующих печень, притупляет снижение адаптивного термогенеза в ответ на экспрессию GK в печени. Эти результаты показывают, что это межорганное взаимодействие печени и BAT достигается с помощью вегетативной нервной цепи, состоящей из афферентных блуждающих нервов от печени и эфферентных симпатических нервов к BAT (Fig. 5A) (21). Это реле нейронов печень-мозг-BAT может служить в качестве энергосберегающей системы у животных и способствовать определению предрасположенности к ожирению.
В поддержку этого мнения, когда содержание гликогена в печени снижается голоданием, липолиз в WAT увеличивается за счет нейронального реле, состоящего из афферентных блуждающих нервов от печени и эфферентных симпатических нервов, обеспечивающих иннервацию WAT (рис. 5A). Напротив, увеличение содержания гликогена в печени ингибирует эту ретрансляцию нейронов печень-мозг-WAT (22). Таким образом, содержание гликогена в печени играет роль в переключении источника топлива с гликогена на триглицериды в условиях голодания через нейронный ретранслятор.
Помимо накопления липидов и углеводов в печени, повышенное поглощение аминокислот печенью влияет на системный метаболизм липидов через нервный контур, состоящий как из афферентных, так и из эфферентных волокон (23). Таким образом, печень может функционировать как метаболический сенсор, ведущий к передаче информации о метаболизме глюкозы, липидов и аминокислот в ЦНС.
Регуляция системного метаболизма с помощью нейронального реле из желудочно-кишечного тракта, состоящего из афферентных и эфферентных нервов
Афферентные волокна блуждающего нерва, иннервирующие желудочно-кишечный тракт, также передают метаболическую информацию к эфферентным волокнам блуждающего нерва, иннервирующим печень на уровне ЦНС, составляя цепь автономных нервов кишечник – мозг – печень.Этот нервный контур регулирует выработку глюкозы в печени. Инфузия липидов из верхних отделов кишечника снижает выработку глюкозы в печени посредством механизма, зависящего от вегетативной нервной цепи кишечник – мозг – печень (рис. 5B) (24). Кроме того, введение CCK-8, биологически активной формы CCK, в двенадцатиперстную кишку также снижает продукцию глюкозы в печени через этот вегетативный нервный контур (рис. 5B) (25). Эффекты снижения выработки глюкозы в печени липидами и CCK-8 нарушаются у грызунов, получавших HFD (24, 25), что свидетельствует о значении этой нервной цепи в нарушении толерантности к глюкозе при статусе ожирения.
Рецептор GLP-1 расположен на афферентных окончаниях блуждающего нерва в портальной области, и введение антагониста рецептора GLP-1 в воротную вену усугубляет непереносимость глюкозы у крыс (111). Недавние сообщения предполагают, что рецепторы GLP-1, экспрессируемые на афферентных волокнах блуждающего нерва, иннервирующие воротную вену, участвуют в опосредованном GLP-1 усилении GSIS (112, 113). В этой системе усиление GSIS опосредуется эфферентными волокнами блуждающего нерва, иннервирующими поджелудочную железу (113). Следовательно, усиление GSIS с помощью GLP-1 происходит, по крайней мере частично, через вегетативный нервный контур воротная вена-мозг-поджелудочная железа.
Сообщалось о роли афферентных блуждающих нервов в опосредовании фармакологических эффектов некоторых лекарств. Хотя метформин является антидиабетическим препаратом, наиболее широко используемым при диабете 2 типа, механизм, с помощью которого метформин снижает уровень глюкозы, полностью не изучен. Недавнее исследование показало, что метформин активирует форму дуоденальной 5′-AMP-активируемой протеинкиназы (AMPK), тем самым снижая выработку глюкозы в печени через автономную нервную цепь кишечник-мозг-печень, состоящую из афферентных блуждающих нервов от кишечника и эфферентных блуждающих нервов. в печень (рис.5Б) (114). Кроме того, та же исследовательская группа сообщила, что ресвератрол, сильный антиоксидант, также снижает выработку глюкозы в печени посредством аналогичного механизма (рис. 5B) (115).
Регулирование пролиферации β-клеток вегетативным нервным контуром печень-мозг-панкреатические β-клетки
В инсулинорезистентных состояниях, таких как ожирение, β-клетки пролиферируют и секретируют больше инсулина, чтобы удовлетворить повышенную системную потребность в инсулине (116), что позволяет поддерживать гомеостаз глюкозы.Однако механизм (ы), с помощью которого ожирение вызывает компенсаторные реакции β-клеток, до конца не изучен.
Путь внеклеточной сигнально-регулируемой киназы (ERK) является одним из путей митоген-активируемых протеинкиназ (117). Путь ERK в печени активирован в нескольких моделях ожирения на мышах (18, 118). Ранее мы показали, что пролиферация β-клеток заметно усиливается активацией печеночного пути ERK (18, 119). Печеночные ERK-активированные мыши демонстрируют сильное усиление как GSIS, так и пролиферации β-клеток.Хирургическая денервация блуждающих волокон, иннервирующих поджелудочную железу, или селективная фармакологическая деафферентация чревных нервов капсаицином значительно подавляют усиление как GSIS, так и пролиферации β-клеток у мышей, активируемых ERK в печени. Таким образом, этот межорганный перекрестный обмен между печенью и β-клетками поджелудочной железы опосредуется через вегетативную нервную цепь, состоящую из афферентных внутренних нервов от печени и эфферентных блуждающих нервов к поджелудочной железе (рис. 6).
Рис.6.
Регулирование пролиферации β-клеток вегетативным нервным контуром печень-мозг-панкреатические β-клетки. Путь ERK в печени активируется при развитии ожирения. Нейрональные сигналы, запускаемые активацией печеночной ERK, передаются в мозг через афферентные волокна внутренних нервов. После модуляции этих сигналов в головном мозге эфферентные волокна блуждающих нервов передают их β-клеткам поджелудочной железы и тем самым вызывают компенсаторные реакции β-клеток, такие как секреция инсулина и пролиферация β-клеток.В этой системе блуждающие нервы высвобождают множество нервных факторов, включая ацетилхолин, PACAP и VIP, и активируют путь β-клеток FoxM1, тем самым способствуя компенсаторной пролиферации β-клеток.
Рис. 6.
Регулирование пролиферации β-клеток вегетативным нервным контуром печень-мозг-панкреатические β-клетки. Путь ERK в печени активируется при развитии ожирения. Нейрональные сигналы, запускаемые активацией печеночной ERK, передаются в мозг через афферентные волокна внутренних нервов.После модуляции этих сигналов в головном мозге эфферентные волокна блуждающих нервов передают их β-клеткам поджелудочной железы и тем самым вызывают компенсаторные реакции β-клеток, такие как секреция инсулина и пролиферация β-клеток. В этой системе блуждающие нервы высвобождают множество нервных факторов, включая ацетилхолин, PACAP и VIP, и активируют путь β-клеток FoxM1, тем самым способствуя компенсаторной пролиферации β-клеток.
Важно отметить, что блокирование этой вегетативной нервной цепи подавляет компенсаторную пролиферацию β-клеток наряду с развитием ожирения у мышей.Следовательно, эта вегетативная нервная цепь печень-мозг-панкреатические β-клетки действительно участвует в увеличении массы β-клеток, связанном с развитием ожирения. Кроме того, стимуляция этого вегетативного нервного контура посредством опосредованной аденовирусом активации печеночной ERK заметно снижает уровни глюкозы в крови в нескольких моделях инсулино-дефицитного диабета на мышах. Таким образом, есть надежда, что манипулирование этим вегетативным нервным контуром приведет к разработке новых и эффективных терапевтических стратегий для лечения инсулино-дефицитного диабета.
Недавно мы выяснили молекулярный механизм, посредством которого эфферентные сигналы блуждающего нерва в этой системе усиливают пролиферацию β-клеток. FoxM1 и его гены-мишени были значительно увеличены в изолированных островках поджелудочной железы как у мышей, активированных печеночным ERK, так и у мышей ob / ob, мышиной модели ожирения (19). Примечательно, что ингибирование вегетативной нервной цепи печень – β-клетки блокирует повышающую регуляцию пути β-клетки FoxM1 как у мышей, активируемых печеночными ERK, так и у мышей ob / ob. Кроме того, индуцированная ожирением пролиферация β-клеток явно нарушена у мышей, у которых специфический для β-клеток нокаут FoxM1 может быть индуцирован тамоксифеном, что указывает на то, что в условиях ожирения сигналы блуждающего нерва усиливают пролиферацию β-клеток через FoxM1. -зависимый механизм (рис.6). Мы также продемонстрировали, что комбинации нескольких нейромедиаторов блуждающего нерва, например ацетилхолин, полипептид, активирующий аденилатциклазу гипофиза (PACAP) и VIP, способствуют пролиферации β-клеток посредством FoxM1-зависимого механизма (рис. 6). Таким образом, блуждающие нервы высвобождают несколько нейротрансмиттеров, тем самым способствуя компенсаторной пролиферации β-клеток во время развития ожирения.
Блуждающие нервы, иннервирующие островки поджелудочной железы, заканчиваются парасимпатическими ганглиями (120).Используя метод очистки тканей CUBIC (121), мы сделали любопытное наблюдение: парасимпатические ганглии расположены в основном вблизи островков. Таким образом, используя блуждающую систему в поджелудочной железе (19), которая высвобождает множество нейротрансмиттеров, достигая высоких концентраций этих факторов локально вокруг β-клеток, мы сочли вероятным, что несколько путей в β-клетках стимулируются одновременно, что приводит к эффективному и избирательная пролиферация β-клеток.
Возможное участие системного метаболизма, регулируемого вегетативными нервами, в иммунной системе
Вегетативная нервная система также играет важную роль в регулировании иммунной системы.Некоторые типы иммунных клеток, такие как макрофаги и Т-клетки, экспрессируют рецепторы нейротрансмиттеров, а нервные импульсы регулируют выработку цитокинов (122–125). Кроме того, «воспалительный рефлекс», который генерируется вегетативными нервными цепями, состоящими как из афферентных, так и из эфферентных блуждающих нервов, участвует в регулировании иммунитета и воспаления (124, 126).
Электрическая стимуляция блуждающих нервов отрицательно регулирует иммунные ответы при аутоиммунных заболеваниях, таких как ревматоидный артрит и воспалительное заболевание кишечника, на животных моделях (126–128).Используя преимущества этих знаний, стимуляция блуждающего нерва использовалась для лечения пациентов с ревматоидным артритом (129) и воспалительными заболеваниями кишечника (130) путем подавления статуса активации иммунных клеток в селезенке и кишечном тракте. Недавно, в контексте воспалительных заболеваний кишечника, группа Канаи продемонстрировала, что печеночные афферентные нервы блуждающего нерва играют роль в восприятии кишечного микроокружения, чтобы поддерживать регуляторные Т-клетки (клетки T reg ), регулируя нишу этих клеток (131).Авторы также предположили, что система, поддерживающая клетки T reg , обеспечивается вегетативной нервной цепью, состоящей из афферентных блуждающих нервов от печени и эфферентных блуждающих нервов к желудочно-кишечному тракту (131). Таким образом, концепции «нейроиммунной системы» в настоящее время уделяется значительное внимание.
Глюкоза необходима для функций всех клеток, включая иммунные. Например, Т-клетки требуют больших затрат энергии для их активации, пролиферации и секреции цитокинов (132).В этом отношении вполне вероятно, что системный метаболизм косвенно играет роль в регулировании иммунной системы. И наоборот, иммунные клетки участвуют в контроле термогенной активности BAT и бежевых адипоцитов. Привлечение эозинофилов, врожденных лимфоидных клеток 2 типа и инвариантных естественных Т-клеток-киллеров активирует макрофаги M2, которые связаны с активацией BAT и потемнением WAT (133). С другой стороны, провоспалительные цитокины, продуцируемые иммунными клетками, такие как макрофаги M1, ингибируют норадренергическую передачу сигналов, тем самым подавляя термогенную активность BAT и бежевой жировой ткани.Более того, активированные адипоциты у тучных животных демонстрируют повышенную экспрессию Toll-подобного рецептора 4 (TLR4), что приводит к продукции провоспалительных факторов, таких как TNF-α, IL-6, CCL2, CCL5 и CCL11 (134). Эти молекулы ухудшают патологические состояния метаболических заболеваний за счет активации иммунной системы.
Таким образом, системный метаболизм и иммунная система влияют друг на друга, регулируя гомеостаз всего тела. Кроме того, вероятно, что нарушение взаимодействия между системным метаболизмом и иммунной системой приводит к патологическим состояниям, таким как инфекционные, аутоиммунные и метаболические заболевания.Учитывая важную роль вегетативной нервной системы как в системном метаболизме, так и в иммунной системе, вегетативная нервная система может участвовать в скоординированной регуляции иммуно-метаболических взаимодействий. Для уточнения этих механизмов необходимы дальнейшие исследования.
Заключение
Как описано в этом обзоре, недавно полученные данные прояснили механизмы систем метаболической регуляции, в которых афферентные вегетативные нервы передают метаболическую информацию от периферических органов к ЦНС, а ЦНС, в свою очередь, регулирует функции органов через эфферентные вегетативные нервы (135). .Помимо регуляции метаболизма, в настоящее время большое внимание привлекает важность связей между нейрональной и иммунной системами.
Электрическая стимуляция афферентных или эфферентных волокон вегетативных нервов недавно стала применяться в клинических условиях. Например, стимуляция афферентных блуждающих нервов применяется при лечении эпилепсии и депрессии (136–139). С другой стороны, стимуляция эфферентного блуждающего нерва используется при ревматоидном артрите, воспалительном заболевании кишечника (139) и сердечной аритмии (140).Совсем недавно было разработано поддиафрагмально имплантированное электрическое устройство, которое деактивирует эфферентные блуждающие нервы, для снижения веса и оказывает свой эффект, подавляя моторику желудочно-кишечного тракта у пациентов с ожирением (141). Следовательно, модуляция афферентных и эфферентных волокон вегетативных нервов, а также вегетативных нервных цепей, как описано выше, является потенциальным вариантом лечения нескольких метаболических заболеваний, а также заболеваний, связанных с иммунитетом. В частности, стимуляция вегетативной нервной цепи печень-мозг-панкреатические β-клетки является многообещающим вариантом лечения инсулино-дефицитного диабета.
Некоторые вопросы, касающиеся этих систем, требуют дальнейшего изучения. Например, пути ЦНС, участвующие в этих вегетативных нервных цепях, остаются в значительной степени неизвестными. Кроме того, необходимо изучить молекулярные механизмы, с помощью которых активируются афферентные нервные волокна в этих цепях. Дальнейшие исследования, направленные на выяснение этих вопросов, могут облегчить понимание как метаболического гомеостаза, так и патогенеза метаболических заболеваний. Точно так же эти механизмы можно использовать для понимания иммунного гомеостаза и патогенеза аутоиммунных заболеваний.
Благодарность
Авторы благодарят профессора Масааки Мураками (Университет Хоккайдо) за полезные советы относительно описания иммунных клеток, цитокинов и аутоиммунных заболеваний.
Финансирование
Исследовательские усилия авторов, представленные в этой рукописи, были поддержаны грантами на научные исследования J.I. (19H03706) и Х.К. (20H05694) от Японского общества содействия науке. Это исследование также было поддержано Японским агентством науки и технологий (Moonshot R&D) (номер гранта JPMJPS2023, выданный H.K.) и Японское агентство медицинских исследований и разработок под номерами грантов 20gm6210002h0003 (для J.I.) и JP20gm5010002h0004 (для H.K.).
Заявление о конфликте интересов: авторы заявили об отсутствии конфликта интересов.
Список литературы
1
Abbott
,
C. R.
,
Monteiro
,
M.
,
Small
,
C. J.
et al.
2005
.
Ингибирующие эффекты периферического введения пептида YY (3-36) и глюкагоноподобного пептида-1 при приеме пищи ослабляются за счет устранения пути блуждающий нерв-ствол мозга-гипоталамус
.
Brain Res.
1044
:
127
,2
Plamboeck
,
A.
,
Veedfald
,
S.
,
Deacon
,
C. F.
et al.
2013
.
Эффект экзогенного GLP-1 на прием пищи теряется у субъектов мужского пола, подвергнутых резкой ваготомии с пилоропластикой
.
г. J. Physiol. Гастроинтест. Liver Physiol.
304
:
G1117
.3
Williams
,
D.L.
,
Baskin
,
D. G.
и
Schwartz
,
M. W
.
2009
.
Доказательства того, что кишечный глюкагоноподобный пептид-1 играет физиологическую роль в обеспечении чувства насыщения
.
Эндокринология
150
:
1680
.4
Krieger
,
J. P.
,
Arnold
,
M.
,
Pettersen
,
K. G.
et al.
2016
.
Нокдаун рецепторов GLP-1 в афферентах блуждающего нерва влияет на нормальное потребление пищи и гликемию
.
Диабет
65
:
34
.5
Smith
,
G. P.
,
Jerome
,
C.
и
Norgren
,
R
.
1985
.
Афферентные аксоны в брюшной полости блуждающего нерва опосредуют действие холецистокинина на сытость у крыс
.
г. J. Physiol.
249
(
5 Pt 2
):
R638
,6
MacLean
,
D. B
.
1985
.
Устранение периферического насыщения холецистокинином у крыс, получавших капсаицин
.
Рег. Pept.
11
:
321
.7
Date
,
Y.
,
Murakami
,
N.
,
Toshinai
,
K.
et al.
2002
.
Роль желудочного афферентного блуждающего нерва в индуцированном грелином кормлении и секреции гормона роста у крыс
.
Гастроэнтерология
123
:
1120
,8
le Roux
,
C. W.
,
Neary
,
N.M.
,
Halsey
,
T. J.
et al.
2005
.
Грелин не стимулирует прием пищи у пациентов с хирургическими вмешательствами, включающими ваготомию.
.
J. Clin. Эндокринол. Метаб.
90
:
4521
.9
Thorens
,
B.
и
Larsen
,
P.J
.
2004
.
Сигнальные молекулы кишечного происхождения и афференты блуждающего нерва в контроле глюкозы и энергетического гомеостаза
.
Curr. Opin. Clin. Nutr. Метаб. Care
7
:
471
.10
Ahrén
,
B
.
2000
.
Вегетативная регуляция секреции островкового гормона — последствия для здоровья и болезней
.
Диабетология
43
:
393
.11
Симадзу
,
T
.
1967
.
Активность гликоген синтетазы в печени: регуляция вегетативными нервами
.
Наука
156
:
1256
.12
Porte
,
D.
Jr.
1967
.
Рецепторный механизм подавления высвобождения инсулина адреналином у человека
.
J. Clin. Инвестировать.
46
:
86
.13
Bloom
,
S. R.
и
Edwards
,
A. V
.
1975
.
Высвобождение глюкагона поджелудочной железы и ингибирование инсулина в ответ на стимуляцию симпатической иннервации
.
J. Physiol.
253
:
157
.14
Симадзу
,
Т.
и
Фукуда
,
А
.
1965
.
Повышенная активность гликогенолитических ферментов в печени после стимуляции чревного нерва
.
Science
150
:
1607
.15
Edwards
,
A.V.
и
Silver
,
M
.
1970
.
Гликогенолитический ответ на стимуляцию чревных нервов у телят, подвергнутых адреналэктомии
.
J. Physiol.
211
:
109
.16
Lafontan
,
M.
,
Barbe
,
P.
,
Galitzky
,
J.
et al.
1997
.
Адренергическая регуляция метаболизма адипоцитов
.
Hum. Репрод.
12
(
доп.1
):
6
,17
Mund
,
R.A.
и
Frishman
,
W. H
.
2013
.
Термогенез коричневой жировой ткани: β3-адренорецепторы как потенциальная мишень для лечения ожирения у людей
.
Кардиол. Ред.
21
:
265
. 18
Имаи
,
Дж.
,
Катагири
,
Х.
,
Ямада
,
Т.
и др.
2008
.
Регулирование массы бета-клеток поджелудочной железы нейрональными сигналами из печени
.
Наука
322
:
1250
.19
Ямамото
,
Дж.
,
Имаи
,
Дж.
,
Идзуми
,
Т.
и др.
2017
.
Нейрональные сигналы регулируют индуцированную ожирением пролиферацию β-клеток с помощью FoxM1-зависимого механизма
.
Nat. Commun.
8
:
1930
.20
Uno
,
K.
,
Katagiri
,
H.
,
Yamada
,
T.
et al.
2006
.
Нейрональный путь от печени модулирует расход энергии и системную чувствительность к инсулину
.
Science
312
:
1656
.21
Tsukita
,
S.
,
Yamada
,
T.
,
Uno
,
K.
et al.
2012
.
Глюкокиназа печени модулирует предрасположенность к ожирению, регулируя термогенез BAT посредством нервных сигналов
.
Cell Metab.
16
:
825
.22
Идзумида
,
Y.
,
Yahagi
,
N.
,
Takeuchi
,
Y.
et al.
2013
.
Нехватка гликогена во время голодания запускает нейросистему печень-мозг-жир, способствуя утилизации жира
.
Nat. Commun.
4
:
2316
.23
Uno
,
K.
,
Yamada
,
T.
,
Ishigaki
,
Y.
et al.
2015
.
Печеночная аминокислота / mTOR / S6K-зависимый сигнальный путь модулирует системный метаболизм липидов посредством нейрональных сигналов
.
Nat. Commun.
6
:
7940
.24
Wang
,
P. Y.
,
Caspi
,
L.
,
Lam
,
C. K.
et al.
2008
.
Липиды верхних отделов кишечника запускают ось кишечник – мозг – печень для регулирования выработки глюкозы
.
Природа
452
:
1012
.25
Cheung
,
G. W.
,
Kokorovic
,
A.
,
Lam
,
C. K.
et al.
2009
.
Кишечный холецистокинин контролирует выработку глюкозы через нейронную сеть
.
Cell Metab.
10
:
99
.26
Ruud
,
J.
,
Steculorum
,
S. M.
и
Brüning
,
J. C
.
2017
.
Нейронный контроль периферической чувствительности к инсулину и метаболизма глюкозы
.
Nat. Commun.
8
:
15259
.27
Belgardt
,
B. F.
,
Okamura
,
T.
и
Brüning
,
J. C
.
2009
.
Передача сигналов гормона и глюкозы в нейронах POMC и AgRP
.
J. Physiol.
587
(
Pt 22
):
5305
,28
Валасси
,
E.
,
Scacchi
,
M.
и
Cavagnini
,
F
.
2008
.
Нейроэндокринный контроль приема пищи
.
Nutr. Метаб. Кардиоваск. Дис.
18
:
158
.29
Собрино Креспо
,
C.
,
Perianes Cachero
,
A.
,
Puebla Jiménez
,
L.
et al.
2014
.
Пептиды и прием пищи
.
Фронт.Эндокринол. (Лозанна)
5
:
58
.30
Berthoud
,
H. R.
и
Neuhuber
,
W. L
.
2000
.
Функциональная и химическая анатомия афферентной системы блуждающего нерва
.
Auton. Neurosci.
85
:
1
.31
Berthoud
,
H. R
.
2004
.
Анатомия и функция сенсорных печеночных нервов
.
Анат. Рек. Discov.Мол. Клетка. Evol. Биол.
280
:
827
.32
de Lartigue
,
G.
и
Diepenbroek
,
C
.
2016
.
Новые разработки в области определения афферентных питательных веществ блуждающего нерва и его роли в энергетическом гомеостазе
.
Curr. Opin. Pharmacol.
31
:
38
.33
Waise
,
T. M. Z.
,
Dranse
,
H.J.
и
Lam
,
T.К. Т
.
2018
.
Метаболическая роль афферентной иннервации блуждающего нерва
.
Nat. Преподобный Гастроэнтерол. Гепатол.
15
:
625
.34
Berthoud
,
H. R.
и
Neuhuber
,
W. L
.
2019
.
Блуждающие механизмы как нейромодуляторные мишени для лечения метаболических заболеваний
.
Ann. NY Acad. Sci.
1454
:
42
.35
Бохоркес
,
Д.V.
,
Shahid
,
R.A.
,
Erdmann
,
A.
et al.
2015
.
Нейроэпителиальный контур, образованный иннервацией сенсорных энтероэндокринных клеток
.
J. Clin. Инвестировать.
125
:
782
.36
Kaelberer
,
M. M.
,
Buchanan
,
K. L.
,
Klein
,
M. E.
et al.
2018
.
Нейронная цепь кишечник-мозг для сенсорной трансдукции питательных веществ
.
Science
361
:
eaat5236
.37
van Bloemendaal
,
L.
,
Ten Kulve
,
J. S.
,
la Fleur
,
S. E.
et al.
2014
.
Влияние глюкагоноподобного пептида 1 на аппетит и массу тела: фокус на ЦНС
.
J. Endocrinol.
221
:
T1
.38
Накагава
,
A.
,
Satake
,
H.
,
Nakabayashi
,
H.
et al.
2004
.
Экспрессия гена рецептора глюкагоноподобного пептида-1, но не глюкозозависимого инсулинотропного полипептида, в клетках узлового ганглия крысы
.
Auton. Neurosci.
110
:
36
.39
Ивасаки
,
Y.
,
Sendo
,
M.
,
Dezaki
,
K.
et al.
2018
.
Высвобождение GLP-1 и активация афферентного блуждающего нерва опосредуют полезные метаболические и хронотерапевтические эффекты D-аллюлозы
.
Nat. Commun.
9
:
113
.40
Liddle
,
R.A.
,
Goldfine
,
I. D.
,
Rosen
,
M. S.
et al.
1985
.
Биоактивность холецистокинина в плазме крови человека. Молекулярные формы, реакция на кормление и связь с сокращением желчного пузыря
.
J. Clin. Инвестировать.
75
:
1144
.41
Кубер
,
Дж. К.
,
Бернар
,
Г.
,
Фушики
,
Т.
и др.
1990
.
Люминальные ХЦК-рилизинг-факторы изолированного крыс с сосудистой перфузией двенадцатиперстной кишки
.
г. J. Physiol.
259
(
2 Pt 1
):
G191
.42
Gutzwiller
,
J. P.
,
Drewe
,
J.
,
Ketterer
,
S.
et al.
2000
.
Взаимодействие между CCK и преднагрузкой при сокращении потребления пищи опосредуется рецепторами CCK-A у людей
.
г. J. Physiol. Regul. Интегр. Комп. Physiol.
279
:
R189
.43
Tschöp
,
M.
,
Smiley
,
D. L.
и
Heiman
,
M. L
.
2000
.
Грелин вызывает ожирение у грызунов
.
Nature
407
:
908
.44
Kamegai
,
J.
,
Tamura
,
H.
,
Shimizu
,
T.
et al.
2001
.
Хроническая центральная инфузия грелина увеличивает уровни гипоталамического нейропептида Y и мРНК агути-родственного белка, а также массу тела у крыс
.
Диабет
50
:
2438
.45
Дата
,
Y.
,
Shimbara
,
T.
,
Koda
,
S.
et al.
2006
.
Периферический грелин передает орексигенные сигналы через норадренергический путь от заднего мозга к гипоталамусу
.
Cell Metab.
4
:
323
.46
Phillips
,
R.J.
и
Powley
,
T. L
.
1996
.
Объем желудка, а не содержание питательных веществ препятствует потреблению пищи
.
г. J. Physiol.
271
(
3 Pt 2
):
R766
.47
Mathis
,
C.
,
Moran
,
T.H.
и
Schwartz
,
G.J
.
1998
.
Чувствительные к нагрузке афференты блуждающего нерва желудка крысы кодируют объем, но не питательные вещества желудка
.
г. J. Physiol.
274
:
R280
.48
Симадзу
,
T
.
1981
.
Регуляция метаболизма печени и жировой ткани центральной нервной системой
.
Diabetologia
20
(
доп.1
):
343
.49
Niijima
,
A
.
1989
.
Нервная регуляция обмена веществ
.
Прог. Neurobiol.
33
:
135
.50
Янсен
,
A. S.
,
Nguyen
,
X. V.
,
Karpitskiy
,
V.
et al.
1995
.
Центральные командные нейроны симпатической нервной системы: основа реакции «бей или беги»
.
Science
270
:
644
.51
Webber
,
J.
и
Macdonald
,
I.А
.
2000
.
Сигнализация в гомеостазе массы тела: нейроэндокринные эфферентные сигналы
.
Proc. Nutr. Soc.
59
:
397
.52
Bers
,
D. M
.
2002
.
Муфта возбуждения-сокращения сердца
.
Nature
415
:
198
.53
Konturek
,
S. J.
,
Konturek
,
J. W.
,
Pawlik
,
T.
et al.
2004
.
Ось мозг – кишечник и ее роль в контроле приема пищи
.
J. Physiol. Pharmacol.
55
(
1 Pt 2
):
137
.54
Тан
,
S. C.
,
Baeyens
,
L.
,
Шен
,
C. N.
и др.
2018
.
Нейроинсулярная сеть поджелудочной железы человека в состоянии здоровья и жировой инфильтрации
.
Диабетология
61
:
168
.55
Родригес-Диас
,
Р.
,
Абдулреда
,
М. Х.
,
Формозо
,
А. Л.
и др.
2011
.
Паттерны иннервации вегетативных аксонов в эндокринной поджелудочной железе человека
.
Cell Metab.
14
:
45
.56
Frohman
,
L. A.
,
Ezdinli
,
E. Z.
и
Javid
,
R
.
1967
.
Влияние ваготомии и стимуляции блуждающего нерва на секрецию инсулина
.
Диабет
16
:
443
.57
Daniel
,
P. M.
и
Henderson
,
J. R
.
1967
.
Влияние стимуляции блуждающего нерва на уровни инсулина и глюкозы в плазме крови бабуина
.
J. Physiol.
192
:
317
.58
Гаутам
,
D.
,
Хан
,
S.J.
,
Hamdan
,
F. F.
et al.
2006
.
Критическая роль мускариновых ацетилхолиновых рецепторов бета-клеток M3 в регуляции высвобождения инсулина и гомеостаза глюкозы в крови in vivo
.
Cell Metab.
3
:
449
.59
Kelley
,
G. G.
,
Zawalich
,
K. C.
и
Zawalich
,
W. S
.
1995
.
Синергетическое взаимодействие глюкозы и нейрогуморальных агонистов для стимуляции гидролиза островкового фосфоинозитида
.
г. J. Physiol.
269
(
3 Pt 1
):
E575
,60
Verspohl
,
E.J.
и
Herrmann
,
K
.
1996
.
Участие G-белков в действии карбахола и холецистокинина на островки поджелудочной железы крыс
.
г. J. Physiol.
271
(
1 Pt 1
):
E65
.61
Ahrén
,
B.
,
Karlsson
,
S.
и
Линдског
,
S
.
1990
.
Холинергическая регуляция эндокринной поджелудочной железы
.
Прог. Brain Res.
84
:
209
.62
Verspohl
,
E.J.
,
Tacke
,
R.
,
Mutschler
,
E.
et al.
1990
.
Подтипы мускариновых рецепторов в островках поджелудочной железы крыс: связывание и функциональные исследования
.
евро. Дж.Pharmacol.
178
:
303
.63
Niwa
,
T.
,
Matsukawa
,
Y.
,
Senda
,
T.
et al.
1998
.
Ацетилхолин активирует внутриклеточное движение гранул инсулина в бета-клетках поджелудочной железы посредством инозитолтрифосфат-зависимой [коррекция трифосфат-зависимой] мобилизации внутриклеточного Ca 2+
.
Диабет
47
:
1699
.64
Bloom
,
S. R.
,
Edwards
,
A. V.
и
Vaughan
,
N.J
.
1974
.
Роль вегетативной иннервации в контроле высвобождения глюкагона при гипогликемии у теленка
.
J. Physiol.
236
:
611
.65
Кането
,
A.
,
Miki
,
E.
и
Kosaka
,
K
.
1974
.
Влияние стимуляции блуждающего нерва на секрецию глюкагона и инсулина
.
Эндокринология
95
:
1005
.66
Арен
,
Б.
и
Таборски
,
Г. Дж.
мл.
1986
.
Механизм стимуляции блуждающего нерва секреции глюкагона и инсулина у собак
.
Эндокринология
118
:
1551
.67
Hommel
,
H.
,
Fischer
,
U.
,
Retzlaff
,
K.
et al.
1972
.
Механизм секреции инсулина после перорального приема глюкозы. II. Рефлекторная секреция инсулина у находящихся в сознании собак со свищами пищеварительного тракта путем имитационного кормления глюкозой или водопроводной водой
.
Diabetologia
8
:
111
.68
Berthoud
,
H. R.
,
Trimble
,
E. R.
,
Siegel
,
E. G.
и др.
1980
.
Секреция инсулина в головной фазе у нормальных крыс и крыс с трансплантированными островками поджелудочной железы
.
г. J. Physiol.
238
:
E336
.69
Taylor
,
I. L.
и
Feldman
,
M
.
1982
.
Влияние цефало-вагальной стимуляции на инсулин, желудочно-ингибирующий полипептид и высвобождение полипептида поджелудочной железы у людей
.
J. Clin. Эндокринол. Метаб.
55
:
1114
.70
Брюс
,
D. G.
,
Storlien
,
L. H.
,
Furler
,
S. M.
et al.
1987
.
Метаболические реакции головной фазы у взрослых с нормальным весом
.
Метаболизм
36
:
721
.71
Bloom
,
S. R.
,
Edwards
,
A.V.
и
Vaughan
,
N.J
.
1973
.
Роль симпатической иннервации в контроле концентрации глюкагона в плазме у теленка
.
J. Physiol.
233
:
457
.72
Кането
,
A.
,
Kajinuma
,
H.
и
Kosaka
,
K
.
1975
.
Влияние стимуляции висцерального нерва на выработку глюкагона и инсулина у собак
.
Эндокринология
96
:
143
.73
Berger
,
M.
,
Scheel
,
D. W.
,
Macias
,
H.
et al. .
2015
.
Передача сигналов рецептора, связанного с Galphai / o, ограничивает рост бета-клеток поджелудочной железы
.
Proc. Natl Acad. Sci. USA
112
:
2888
.74
Kurose
,
T.
,
Seino
,
Y.
,
Nishi
,
S.
et al.
1990
.
Механизм симпатической нервной регуляции секреции инсулина, соматостатина и глюкагона
.
г. Дж.Physiol.
258
(
1 Pt 1
):
E220
,75
Skoglund
,
G.
,
Lundquist
,
I.
и
Ahrén
,
B
.
1988
.
Селективная активация альфа-2-адренорецепторов клонидином: влияние на отток 45Ca 2+ и секрецию инсулина из изолированных островков крысы
.
Acta Physiol. Сканд.
132
:
289
,76
Верберн
,
A.J.
,
Korim
,
W. S.
,
Sabetghadam
,
A.
et al.
2016
.
Адреналин: понимание его метаболической роли при гипогликемии и диабете
.
руб. J. Pharmacol.
173
:
1425
.77
Mraz
,
M.
и
Haluzik
,
M
.
2014
.
Роль иммунных клеток жировой ткани в ожирении и легких формах воспаления
.
J. Endocrinol.
222
:
R113
.78
Van Liefde
,
I.
,
Van Witzenburg
,
A.
и
Vauquelin
,
G
.
1992
.
Множественные подклассы бета-адренергических рецепторов опосредуют индуцированный l-изопротеренолом липолитический ответ в адипоцитах крыс
.
J. Pharmacol. Exp. Ther.
262
:
552
,79
Галицкий
,
J.
,
Reverte
,
M.
,
Portillo
,
M.
et al.
1993
.
Сосуществование бета-1-, бета-2- и бета-3-адренорецепторов в жировых клетках собак и их дифференциальная активация катехоламинами
.
г. J. Physiol.
264
(
3 Pt 1
):
E403
.80
Collins
,
S.
,
Daniel
,
K. W.
,
Rohlfs
,
E. M.
и др.
1994
.
Нарушение экспрессии и функциональной активности бета-3- и бета-1-адренорецепторов в жировой ткани мышей с врожденным ожирением (C57BL / 6J ob / ob)
.
Мол. Эндокринол.
8
:
518
.81
Cypess
,
A. M.
,
Lehman
,
S.
,
Williams
,
G.
et al.
2009
.
Идентификация и значение коричневой жировой ткани у взрослых людей
.
N. Engl. J. Med.
360
:
1509
.82
van Marken Lichtenbelt
,
W. D.
,
Vanhommerig
,
J. W.
,
Smulders
,
N. M.
et al.
2009
.
Холодная активированная коричневая жировая ткань у здоровых мужчин
.
N. Engl. J. Med.
360
:
1500
.83
Virtanen
,
K. A.
,
Lidell
,
M. E.
,
Orava
,
J.
et al.
2009
.
Функциональная коричневая жировая ткань у здоровых взрослых
.
N. Engl. J. Med.
360
:
1518
.84
Saito
,
M.
,
Okamatsu-Ogura
,
Y.
,
Matsushita
,
M.
et al.
2009
.
Высокая частота метаболически активной коричневой жировой ткани у здоровых взрослых людей: последствия воздействия холода и ожирения
.
Диабет
58
:
1526
.85
Wu
,
J.
,
Boström
,
P.
,
Sparks
,
L. M.
et al.
2012
.
Бежевые адипоциты представляют собой отдельный тип термогенных жировых клеток у мышей и людей
.
Ячейка
150
:
366
.86
Enerbäck
,
S
.
2013
.
Метаболизм жировой ткани в 2012 году: пластичность жировой ткани и новые терапевтические цели
.
Nat. Rev. Endocrinol.
9
:
69
.87
Хармс
,
M.
и
Seale
,
P
.
2013
.
Коричневый и бежевый жир: развитие, функции и терапевтический потенциал
.
Nat. Med.
19
:
1252
.88
Kajimura
,
S
.
2017
.
Жировая ткань в 2016 году: успехи в понимании биологии жировой ткани
.
Nat. Rev. Endocrinol.
13
:
69
.89
Berthoud
,
H. R.
,
Fox
,
E.A.
и
Neuhuber
,
W. L
.
2006
.
Причуды иннервации жировой ткани
.
г. J. Physiol. Regul. Интегр. Комп. Physiol.
291
:
R1240
.90
Kreier
,
F.
,
Fliers
,
E.
,
Voshol
,
P.J.
и др.
2002
.
Селективная парасимпатическая иннервация подкожного и внутрибрюшного жира — функциональные последствия
.
J. Clin. Инвестировать.
110
:
1243
.91
Giordano
,
A.
,
Song
,
C. K.
,
Bowers
,
R. R.
et al.
2006
.
Белая жировая ткань не обладает значительной иннервацией блуждающего нерва и иммуногистохимическими признаками парасимпатической иннервации
.
г. J. Physiol. Regul. Интегр. Комп. Physiol.
291
:
R1243
.92
Чу
,
С.A.
,
Sindelar
,
D. K.
,
Igawa
,
K.
et al.
2000
.
Прямое воздействие катехоламинов на продукцию глюкозы в печени происходит через альфа (1) — и бета (2) -рецепторы у собак
.
г. J. Physiol. Эндокринол. Метаб.
279
:
E463
.93
Li
,
J. H.
,
Gautam
,
D.
,
Han
,
S. J.
et al.
2009
.
Печеночные мускариновые рецепторы ацетилхолина не имеют решающего значения в поддержании гомеостаза глюкозы у мышей
.
Диабет
58
:
2776
.94
Като
,
Х.
и
Симадзу
,
Т
.
1983
.
Влияние вегетативной денервации на синтез ДНК при регенерации печени после частичной гепатэктомии
.
евро. J. Biochem.
134
:
473
.95
Икеда
,
О.
,
Ozaki
,
M.
,
Murata
,
S.
et al.
2009
.
Автономная регуляция регенерации печени после частичной гепатэктомии у мышей
.
J. Surg. Res.
152
:
218
.96
Izumi
,
T.
,
Imai
,
J.
,
Yamamoto
,
J.
et al.
2018
.
Связь блуждающих нервов, макрофагов и гепатоцитов способствует регенерации печени после травм и выживанию всего тела за счет активации FoxM1 в печени
.
Nat. Commun.
9
:
5300
.97
Niijima
,
A
.
1982
.
Глюкозочувствительные афферентные нервные волокна в печеночной ветви блуждающего нерва морской свинки
.
J. Physiol.
332
:
315
.98
Niijima
,
A
.
2000
.
Рефлекторное влияние оральных, желудочно-кишечных и гепатопортальных сенсоров глутамата на активность блуждающего нерва
.
J. Nutr.
130
(
доп. 4S
):
971S
.99
Torii
,
K.
и
Niijima
,
A
.
2001
.
Влияние лизина на афферентную активность печеночной ветви блуждающего нерва у нормальных крыс и крыс с дефицитом L-лизина
.
Physiol. Behav.
72
:
685
.100
Накабаяси
,
H.
,
Nishizawa
,
M.
,
Nakagawa
,
A.
et al.
1996
.
Блуждающий гепатопанкреатический рефлекторный эффект, вызванный интрапортальным появлением tGLP-1
.
г. J. Physiol.
271
(
5 Pt 1
):
E808
.101
Stümpel
,
F.
и
Jungermann
,
K
.
1997
.
Обнаружение внутрипеченочными мускариновыми нервами градиента концентрации глюкозы в воротной артерии как сигнала для инсулинозависимого поглощения глюкозы перфузируемой печенью крысы
.
FEBS Lett.
406
:
119
.102
Донован
,
К. М.
,
Гамильтон-Весслер
,
М.
,
Холтер
,
Дж. Б.
и др.
1994
.
Приоритет глюкосенсоров печени в симпатическом ответе на прогрессирующую гипогликемию
.
Proc. Natl Acad. Sci. США
91
:
2863
.103
Hamilton-Wessler
,
M.
,
Bergman
,
R.N.
,
Halter
,
J. B.
et al.
1994
.
Роль глюкозосенсоров печени в интегрированном симпатическом ответе, вызванном глубокой гипогликемией у собак
.
Диабет
43
:
1052
.104
Burcelin
,
R.
,
Dolci
,
W.
и
Thorens
,
B
.
2000
.
Портальная инфузия глюкозы мышам вызывает гипогликемию: доказательство того, что гепатопортальный датчик глюкозы стимулирует утилизацию глюкозы
.
Диабет
49
:
1635
.105
Burcelin
,
R.
,
Dolci
,
W.
и
Thorens
,
B
.
2000
.
Чувствительность к глюкозе гепатопортальным сенсором является GLUT2-зависимой: анализ in vivo на GLUT2-нулевых мышах
.
Диабет
49
:
1643
.106
Ямада
,
D.
,
Koppensteiner
,
P.
,
Odagiri
,
S.
et al.
2017
.
Общая печеночная ветвь зависимой от блуждающего нерва экспрессии немедленных ранних генов в мозге мышей с помощью интрапортального
l-аргинин: сравнение с холецистокинином-8
.
Фронт. Neurosci.
11
:
366
.107
Williams
,
E. K.
,
Chang
,
R. B.
,
Strochlic
,
D. E.
et al.
2016
.
Сенсорные нейроны, обнаруживающие растяжение и питательные вещества в пищеварительной системе
.
Cell
166
:
209
.108
Yamada
,
T.
,
Katagiri
,
H.
,
Ishigaki
,
Y.
et al.
2006
.
Сигналы от внутрибрюшного жира модулируют чувствительность к инсулину и лептину посредством различных механизмов: участие нейронов в регуляции приема пищи
.
Cell Metab.
3
:
223
.109
Uno
,
K.
,
Yamada
,
T.
,
Ishigaki
,
Y.
et al.
2012
.
Путь метаболизма белка 27, активируемого пролифератором пероксисом, рецептора-γ, способствует развитию гипертензии, связанной с ожирением, через афферентные вагусные сигналы
.
евро. Heart J.
33
:
1279
.110
Bernal-Mizrachi
,
C.
,
Xiaozhong
,
L.
,
Инь
,
L.
et al.
2007
.
Афферентный путь блуждающего нерва связывает активацию PPARalpha в печени с индуцированной глюкокортикоидами инсулинорезистентностью и гипертензией
.
Cell Metab.
5
:
91
.111
Vahl
,
T. P.
,
Tauchi
,
M.
,
Durler
,
T. S.
et al.
2007
.
Рецепторы глюкагоноподобного пептида-1 (GLP-1), экспрессируемые на нервных окончаниях в воротной вене, опосредуют эффекты эндогенного GLP-1 на толерантность к глюкозе у крыс
.
Эндокринология
148
:
4965
.112
Fujiwara
,
K.
,
Gotoh
,
K.
,
Chiba
,
S.
et al.
2012
.
Внутрипортальное введение ингибитора DPP-IV регулирует секрецию инсулина и потребление пищи через печеночный афферентный нерв блуждающего нерва у крыс
.
J. Neurochem.
121
:
66
.113
Нисидзава
,
М.
,
Nakabayashi
,
H.
,
Uehara
,
K.
et al.
2013
.
Интрапортальный GLP-1 стимулирует секрецию инсулина преимущественно через печеночно-панкреатический блуждающий рефлекс
.
г. J. Physiol. Эндокринол. Метаб.
305
:
E376
.114
Duca
,
F. A.
,
Côté
,
C. D.
,
Rasmussen
,
B.A.
et al.
2015
.
Метформин активирует дуоденальный Ampk-зависимый путь снижения продукции глюкозы в печени у крыс
.
Nat. Med.
21
:
506
.115
Côté
,
C. D.
,
Rasmussen
,
B.A.
,
Duca
,
F. A.
et al.
2015
.
Ресвератрол через нейронную сеть активирует дуоденальный Sirt1, чтобы обратить вспять инсулинорезистентность у крыс.
Nat. Med.
21
:
498
.116
Прентки
,
М.
и
Нолан
,
C.J
.
2006
.
Отказ бета-клеток островков при диабете 2 типа
.
J. Clin. Инвестировать.
116
:
1802
.117
Робертс
,
P.J.
и
Der
,
C.J
.
2007
.
Нацеленность на каскад митоген-активируемых протеинкиназ Raf-MEK-ERK для лечения рака
.
Онкоген
26
:
3291
.118
Gum
,
R. J.
,
Gaede
,
L. L.
,
Heindel
,
M. A.
et al.
2003
.
Антисмысловой протеин тирозинфосфатаза 1B обращает активацию митоген-активируемой протеинкиназы p38 в печени мышей ob / ob
.
Мол. Эндокринол.
17
:
1131
.119
Imai
,
J.
,
Oka
,
Y.
и
Katagiri
,
H
.
2009
.
Идентификация нового механизма регуляции массы β-клеток: ретрансляция нейронов из печени в β-клетки поджелудочной железы
.
Островки
1
:
75
. 120
Миллер
,
Р. E
.
1981
.
Нейроэндокринология поджелудочной железы: периферические нервные механизмы в регуляции островков Лангерганса
.
Endocr. Ред.
2
:
471
.121
Susaki
,
E.A.
,
Tainaka
,
K.
,
Perrin
,
D.
et al.
2014
.
Визуализация всего мозга с разрешением одной клетки с использованием химических коктейлей и компьютерного анализа
.
Cell
157
:
726
.122
Wang
,
H.
,
Yu
,
M.
,
Ochani
,
M.
et al.
2003
.
Никотиновый ацетилхолиновый рецептор альфа7-субъединица является важным регулятором воспаления
.
Nature
421
:
384
.123
Kawashima
,
K.
,
Fujii
,
T.
,
Moriwaki
,
Y.
et al.
2012
.
Важнейшие роли ацетилхолина, мускариновых и никотиновых рецепторов ацетилхолина в регуляции иммунной функции
.
Life Sci.
91
:
1027
.124
Чаван
,
С.С.
,
Павлов
,
В.A.
и
Tracey
,
K. J
.
2017
.
Механизмы и терапевтическое значение нейроиммунной коммуникации
.
Иммунитет
46
:
927
.125
Sternberg
,
E. M
.
2006
.
Нервная регуляция врожденного иммунитета: скоординированный неспецифический ответ хозяина на патогены
.
Nat. Rev. Immunol.
6
:
318
.126
Павлов
,
В.A.
,
Chavan
,
S. S.
и
Tracey
,
K. J
.
2018
.
Молекулярная и функциональная нейробиология в иммунитете
.
Annu. Rev. Immunol.
36
:
783
.127
Andersson
,
U.
и
Tracey
,
K. J
.
2012
.
Нервные рефлексы при воспалении и иммунитет
.
J. Exp. Med.
209
:
1057
.128
Боровикова
,
Л.В.
,
Иванова
,
с.
,
Чжан
,
М.
и др.
2000
.
Стимуляция блуждающего нерва ослабляет системную воспалительную реакцию на эндотоксин
.
Nature
405
:
458
.129
Koopman
,
F. A.
,
Chavan
,
S. S.
,
Miljko
,
S.
et al.
2016
.
Стимуляция блуждающего нерва подавляет выработку цитокинов и снижает тяжесть ревматоидного артрита
.
Proc. Natl Acad. Sci. США
113
:
8284
.130
Bonaz
,
B.
,
Sinniger
,
V.
,
Hoffmann
,
D.
et al.
2016
.
Хроническая стимуляция блуждающего нерва при болезни Крона: пилотное исследование через 6 месяцев
.
Нейрогастроэнтерол. Мотил.
28
:
948
.131
Teratani
,
T.
,
Mikami
,
Y.
,
Nakamoto
,
N.
et al.
2020
.
Нервная дуга печень-мозг-кишечник поддерживает нишу клеток T reg в кишечнике
.
Nature
585
:
591
.132
Sagone
,
A. L.
Jr,
LoBuglio
,
A. F.
и
Balcerzak
,
S.П
.
1974
.
Изменения гексозо-монофосфатного шунта при лимфобластной трансформации
.
Ячейка. Иммунол.
14
:
443
.133
Вильярроя
,
F.
,
Cereijo
,
R.
,
Villarroya
,
J.
et al.
2018
.
На пути к пониманию того, как иммунные клетки контролируют коричневую и бежевую адипобиологию
.
Cell Metab.
27
:
954
.134
Wolowczuk
,
I.
,
Verwaerde
,
C.
,
Viltart
,
O.
et al.
2008
.
Питание нашей иммунной системы: влияние на обмен веществ
.
Clin. Dev. Иммунол.
2008
:
639803
.135
Катагири
,
H.
,
Yamada
,
T.
и
Oka
,
Y
.
2007
.
Ожирение и сердечно-сосудистые расстройства: нарушение регуляторной системы, состоящей из гуморальных и нейрональных сигналов
.
Circ. Res.
101
:
27
.136
Morris
,
G. L.
3rd и
Mueller
,
W. M
.
1999
.
Длительное лечение стимуляцией блуждающего нерва у пациентов с рефрактерной эпилепсией. Группа изучения стимуляции блуждающего нерва E01-E05
.
Неврология
53
:
1731
.137
Энглот
,
Д. Дж.
,
Чанг
,
Э. Ф.
и
Огюст
,
К.Я
.
2011
.
Эффективность стимуляции блуждающего нерва при эпилепсии в зависимости от возраста пациента, продолжительности эпилепсии и типа припадка
.
Нейрохирург. Clin. N. Am.
22
:
443
.138
Englot
,
D. J.
,
Chang
,
E. F.
и
Auguste
,
K. I
.
2011
.
Стимуляция блуждающего нерва при эпилепсии: метаанализ эффективности и предикторы ответа
.
J. Neurosurg.
115
:
1248
.139
Bonaz
,
B.
,
Sinniger
,
V.
и
Pellissier
,
S
.
2016
.
Противовоспалительные свойства блуждающего нерва: потенциальные терапевтические последствия стимуляции блуждающего нерва
.
J. Physiol.
594
:
5781
.140
Чжан
,
Ю.
и
Мазгалев
,
Т.№
.
2011
.
Аритмии и стимуляция блуждающего нерва
.
Сердечная недостаточность. Ред.
16
:
147
.141
de Lartigue
,
G
.
2016
.
Роль блуждающего нерва в развитии и лечении ожирения, вызванного диетой
.
J. Physiol.
594
:
5791
.
© Японское общество иммунологии. 2021. Все права защищены.Для получения разрешений обращайтесь по электронной почте: [email protected]
Метаболические характеристики поглощения почечного инсулина
Реферат
Почка выводит инсулин из кровотока посредством клубочкового и перитубулярного клиренса, таким образом подвергая действию гормона как просветные, так и контралюминальные канальцевые поверхности. В проксимальном канальце отфильтрованный инсулин интернализуется посредством пиноцитоза и в конечном итоге локализуется в лизосомах. Судьба инсулина, удаляемого из перитубулярного кровообращения, менее изучена.Ранее мы сообщали, что поглощение инсулина канальцами — это чувствительный к температуре процесс, состоящий из двух разных систем. Система, связанная с просветным аспектом клетки, по-видимому, зависит от окислительного метаболизма, тогда как система, связанная с контрпросветным аспектом клетки, по-видимому, не зависит от него. Чтобы дополнительно охарактеризовать поглощение инсулина почечными канальцами, было проведено исследование с (OCI), равным 1336 ± 51 мкл / мин (SEM), скоростью клубочковой фильтрации (GFR) 1019 ± 54 филмин, перитубулярным клиренсом инсулина 317 ± 56 мкл / мин. , фракционная экскреция натрия (FEna) составила 5.1 ± 0,7%, а фракционная экскреция инсулина (FEi) составила 1,3 ± 0,6%. Добавление ингибиторов гликолиза (йодацетат), Na-K-АТФазы (уабаин) и лизосомальной деградации (хлорохин) было связано со значительным увеличением Fei (4,7 ± 0,4%, 6,4 ± 1,4% и 13,9 ± 2,4% соответственно. ) и FEna (28%, 43% и 12% соответственно). Напротив, ацетазоламид с фуросемидом увеличивали FEna до 39% без изменения FEi. Анализ всех данных не выявил корреляции между FEna и FEi, что указывает на то, что канальцевое всасывание инсулина, по крайней мере, частично не зависит от реабсорбции натрия.Ни один из ингибиторов существенно не изменил OCI, GFR или перитубулярный клиренс. Приведенные выше наблюдения вместе с нашими ранее опубликованными данными показывают, что поглощение инсулина в просвете — это чувствительный к температуре и хлорохину процесс, который зависит от метаболической энергии. Источником этой энергии, по-видимому, является АТФ, который образуется в результате окислительного метаболизма и гликолиза. Эти характеристики, соответствующие процессу, включающему пиноцитоз и лизосомную деградацию, не были характеристиками процесса контралюминального поглощения инсулина.
- Авторские права © 1981 Американской диабетической ассоциации
Frontiers | Происхождение и эволюция метаболизма флавоноидов растений
Введение
Растения обладают способностью вырабатывать огромное количество метаболитов. Прогнозируется, что более 1 000 000 метаболитов будут присутствовать во всем царстве растений (Afendi et al., 2012). Большинство из них — вторичные метаболиты (также называемые специализированными метаболитами), которые играют широкий спектр физиологических и экологических ролей, включая защиту от травоядных и патогенов, аттрактанты для опылителей и переносчиков семян, а также передачу сигналов.В течение длительного процесса эволюции растения приобретали, расширялись, а иногда и утратили способность производить этот огромный набор метаболитов, который обеспечивает адаптивные механизмы, необходимые для выживания в изменяющейся окружающей среде.
Флавоноиды образуют одну из основных групп специализированных метаболитов и включают более 9000 соединений (Williams and Grayer, 2004; Anderson and Markham, 2006). Согласно Номенклатуре флавоноидов (Рекомендации IUPAC, 2017), термин «флавоноид» применяется к (1) соединениям, структурно основанным на производных фенилзамещенного пропилбензола со скелетом C15, (2) соединениям со скелетом C16, которые представляют собой фенил -замещенные производные пропилбензола (ротеноиды), (3) флавонолигнаны на основе производных фенилзамещенного пропилбензола, конденсированные с предшественниками лигнана C 6 -C 3 (Rauter et al., 2018). В ограниченном смысле термин «флавоноид» используется только для тех соединений с углеродным каркасом C 6 -C 3 -C 6 , демонстрирующих структуру хромана или хромена, таких как флаваны, флавоны. , флавонолы и антоцианидины. Халконы, дигидрохалконы и ауроны являются флавоноидами в широком смысле, но не в ограниченном.
Флавоноиды, включая халконы, флавоны, флавонолы, антоцианы и проантоцианидины, широко распространены в царстве растений, и их метаболические пути широко изучены с использованием как биохимических, так и молекулярно-биологических методов.До недавнего времени считалось, что печеночники и мхи являются старейшими растениями-продуцентами флавоноидов (Rausher, 2006; Bowman et al., 2017). Гены, кодирующие ферменты пути биосинтеза фенилпропаноидов, включая первые два фермента биосинтеза флавоноидов (халконсинтаза и халконизомераза), не были обнаружены у водорослей родов Chlamydomonas, Micromonas, Ostreococcus и ген Klebsormidium в пути шикимата были обнаружены в водорослях, печеночниках, мхах, ликофитах, папоротниках и хвощах, голосеменных и покрытосеменных (Bowman et al., 2017). Однако флавоны, изофлавоны и флавонолы были обнаружены в микроводорослях пяти различных эволюционных ветвей ( Cyanobacteria, Rhodophyta, Chlorophyta, Haptophyta и Ochrophyta ) с помощью сверхвысокопроизводительной жидкостной хроматографии с тандемной масс-спектрометрией (Goiris et al., 2014). Это говорит о том, что растения, возможно, приобрели способность производить флавоноиды раньше, чем мы думали ранее. Кроме того, в современных растениях флавоноиды играют важную роль в качестве защитников от ультрафиолета B (УФ-B), пигментов, привлекающих опылителей, фитоалексинов, сигнальных молекул и регуляторов транспорта ауксина и фертильности (Gould and Lister, 2006).Было высказано предположение, что защита от УФ-излучения и регуляция действия растительных гормонов были исходными функциями флавоноидов у самых первых растений, продуцирующих флавоноиды (Stafford, 1991; Shirley, 1996; Rausher, 2006). Эти функции значительно разнообразились в процессе эволюции растений. Таким образом, изучение флавоноидов является полезным подходом для понимания того, как растения приобрели способность производить специализированные метаболиты, а затем выстраивали метаболические пути, необходимые для производства такого огромного разнообразия метаболитов в ходе их эволюции.Это исследование прольет свет на взаимосвязь между генами / белками и метаболитами, а также между метаболитами и их физиологическими функциями. В этом обзоре мы описываем структурное разнообразие флавоноидов, распространенных в царстве растений, и то, как растения приобрели гены биосинтеза флавоноидов в ходе своей эволюции.
Метаболомика проливает свет на эволюцию флавоноидов
Распространение флавоноидов в растительном мире
По оценкам, в царстве растений насчитывается более 9000 флавоноидов (Williams and Grayer, 2004; Anderson and Markham, 2006).Исследования флавоноидов показали, что они распространены в царстве растений, включая покрытосеменные, голосеменные и птеридофиты (Harborne, 1988; Tohge et al., 2013). Обилие информации о флавоноидах у разных видов позволяет нам идентифицировать подклассы флавоноидов (например, халконы, флавоны, флавонолы, антоцианы и проантоцианидины), обнаруженные в каждой подгруппе растений (рисунок 1). Флавон и флаванон содержатся во всех группах растений, кроме роголистника. Насколько нам известно, о флавоноидах в роголистниках не сообщалось.По мере развития и разнообразия групп растений внутри каждой группы производились подклассы флавоноидов. Например, флавоноидные агликоны наиболее разнообразны у покрытосеменных растений. Помимо флаванонов и флавонов, халконы, флавонолы и проантоцианидины встречаются во многих группах. Интересно, что пренилфлавоноиды обнаружены как в печеночниках, так и в покрытосеменных: хотя из бобовых было выделено более 1000 пренилфлавоноидов (Yazaki et al., 2009), пренилдигидрохалкон был обнаружен в печеночниках Radula variabilis и Radula spp.(Асакава и др., 1978, 1982). Эти данные предполагают, что две группы растений приобрели способность продуцировать пренилфлавоноиды независимо, или что многие группы утратили способность продуцировать пренилфлавоноиды в ходе своей эволюции. Молекулы флавоноидов являются конкретным доказательством существования соответствующих генов биосинтеза флавоноидов в растениях. Поэтому аналитические подходы к идентификации флавоноидов важны для понимания эволюции метаболизма флавоноидов в царстве растений.
Рисунок 1 . Распространение подклассов флавоноидов в растительном мире. Красная линия указывает на нерешенные филогенетические отношения в линиях мохообразных, как указано в Bowman et al. (2017).
Передовые технологии метаболомики способствуют пониманию эволюции метаболизма флавоноидов
До сих пор хроматографические и спектроскопические подходы использовались для анализа структуры флавоноидов и выявления их химического разнообразия.Раньше бумажная хроматография, тонкослойная хроматография, колоночная хроматография и жидкостная хроматография (ЖХ) были основными методами, используемыми для изучения флавоноидов. Неочищенные экстракты, содержащие флавоноиды, получают разделением жидкость-жидкость. Например, флавоноидные агликоны и их моногликозиды можно в основном экстрагировать в этилацетате, тогда как ди- или тригликозиды флавоноидов экстрагируются в n -бутаноле. Затем используются хроматографические методы для очистки флавоноидов от этилацетатной фракции или фракций n -бутанола в несколько этапов.Наконец, изолированные флавоноиды анализируются с помощью ядерного магнитного резонанса и масс-спектрометрии, чтобы выяснить их структуру. Это однозначный и простой подход к определению флавоноидов, однако он требует много времени.
С развитием технологий метаболомики флавоноиды теперь можно анализировать гораздо точнее и точнее, чем раньше. ЖХ-тандемная масс-спектрометрия (ЖХ-МС / МС) стала предпочтительным подходом для анализа флавоноидов. Считалось, что в водорослях нет флавоноидов.Однако идентификация флавоноидов, таких как промежуточные продукты и конечные продукты, с помощью метода ЖХ-МС / МС доказала существование биосинтетической способности флавоноидов у микроводорослей (Goiris et al., 2014). Это говорит о том, что передовые технологии метаболомики в каждом отдельном виде растений могут выявить пропущенные флавоноиды.
Интеграция ЖХ-МС / МС с подходами хеминформатики обеспечивает мощный инструмент для исследования разнообразия флавоноидов высокопроизводительным способом (Tsugawa et al., 2016; Акимото и др., 2017). Гликозилированные, ацилированные и пренилированные молекулы флавоноидов и их агликоны могут быть разделены с использованием простых комбинаций растворителей и колонок для ЖХ. Затем разделенные молекулы ионизируют для анализа МС / МС. Ионы продукта образуются из их предшественников путем расщепления эфирных и сложноэфирных связей или даже пренильного фрагмента. Эти простые шаги фрагментации позволяют нам оценить структуру флавоноидов. Инструменты Cheminformatic используются для высокопроизводительного и (полу) автоматического анализа данных.Результаты обычно классифицируются по четырем классам (уровням) в соответствии с руководящими принципами Инициативы стандартов метаболомики: уровень 1, идентификация с использованием аутентичных стандартных соединений; Уровень 2, аннотация с использованием общедоступных баз данных; Уровень 3, характеристика путем расшифровки данных MS / MS; и Уровень 4, неизвестный (не шум) (Sumner et al., 2007). Использование этих уровней снижает количество ложноположительных данных.
Интегрированный подход с использованием ЖХ-МС / МС с маркировкой 13 C (углерод) и хеминформатики может быть использован для определения структур метаболитов, чьи структуры никогда ранее не распознавались.В большинстве общих подходов неизвестные метаболиты остаются нехарактеризованными. Интегративный подход используется для идентификации элементного состава неизвестных метаболитов на основе количества атомов C, определенного путем сравнения 13 C- и немеченых МС-спектров. Спектры МС / МС неизвестных метаболитов затем подвергаются анализу с обогащением по набору фрагментов (FSEA) для определения кандидатов для каждого класса метаболитов. Этот интегративный подход был использован для характеристики 1133 метаболитов, включая флавоноиды, у 12 видов покрытосеменных (Tsugawa et al., 2019). Результаты для охарактеризованных флавоноидов суммированы в таблице 1 вместе с флавоноидами, обнаруженными в печеночнике Marchantia polymorpha , которые были проанализированы с использованием того же метода (Kubo et al., 2018). Наши анализы с использованием LC-MS / MS с меткой 13 C и хеминформатики привели к идентификации большего количества молекул флавоноидов, особенно в группе флавонола и флавона. Пренилированные флавоноиды были проанализированы с помощью ЖХ-МС / МС менее широко, чем другие группы флавоноидов, хотя стратегия химического определения эффективна для профилирования пренилированных флавоноидов.Вероятно, это связано с отсутствием аутентичных стандартов и общедоступных спектров МС / МС. Точная масса известной структуры или структуры, которая предположительно пренилирована, также может быть полезна для профилирования пренилированных флавоноидов в случаях, когда онтологии пренилированных флавоноидов не работают. Это означает, что наблюдаемые спектры МС / МС могут не соответствовать библиотеке, полученной из FSEA.
Таблица 1 . Флавоноиды экспериментально охарактеризованы с помощью ЖХ-МС / МС у 12 видов покрытосеменных и Marchantia polymorpha .
Аналогичным образом, недавние исследования показывают, что метаболизм трицина, по-видимому, специфически развивается у однодольных растений. Трицин служит сомономером лигнина и перекрестно связывается с монолигнолами и γ- p -кумароилированными монолигнолами при лигнификации клеточной стенки. Недавно трицин был идентифицирован как центр зародышеобразования для одревеснения злаков (Lan et al., 2015, 2016a; Lam et al., 2017). Растения Poales, включая Avena sativa, Brachipodium distachyon, Oryza sativa, Triticum durum и Zea mays , накапливают трицин, как и растение Fabales Medicago sativa (Lan et al., 2016б). Интегративный подход был использован для обнаружения производных апигенина, лютеолина и трицина в O. sativa, Z. mays и M. truncatula , и оказалось, что общее содержание трицина намного выше, чем содержание экстрагируемого трицина в эти растения. Интегративный подход с FSEA может быть полезен для дальнейшего понимания роли трицина и его производных в клеточной стенке.
Анализ метаболома на основе ЖХ-МС / МС был использован для всестороннего анализа молекул флавоноидов в отдельных видах растений.Последние технологические разработки позволяют нам проводить высокопроизводительный анализ ЖХ-МС / МС. Отслеживая ионы, полученные из агликонов и модифицированных частей молекул, или обнаруживая сходство МС / МС, химическое определение флавоноидов легко выполняется (полу) автоматическими способами с помощью хеминформатических инструментов для разных видов растений.
Биосинтетические гены образования или модификации агликона можно предсказать на основе структуры флавоноидов. Функциональная геномика с использованием как передового секвенатора, так и более полного и точного профилирования флавоноидов позволяет нам решать эволюционные вопросы о том, когда и почему появились флавоноиды, а пути биосинтеза у растений разнообразны.Кроме того, наряду с прогрессом значительных проектов генома растений, это даст ценные ключи к пониманию эволюционных особенностей метаболизма флавоноидов в царстве растений как несоответствия во взаимоотношениях между метаболитами и генами, о которых мы упоминали в следующих разделах.
Пути биосинтеза флавоноидов
Первая стадия биосинтеза флавоноидов катализируется халконсинтазой (ХС) (рис. 2). Субстраты p -кумароил-КоА, полученные из пути циннамат / монолигнол (фенилпропаноид), и малонил-КоА из ацетат-малонатного (поликетидного) пути превращаются с помощью CHS в нарингенин халкон.Стереоспецифическая циклизация халкона нарингенина в нарингенин катализируется халконизомеразой (CHI). Этот шаг также может происходить спонтанно.
Рисунок 2 . Общие пути биосинтеза флавоноидов в растениях. Зеленые, синие и пурпурные стрелки указывают на ферменты в суперсемействах CYP, 2OGD и SDR соответственно. ANR, антоцианидин редуктаза; ANS, антоцианидинсинтаза; CHI, халконизомераза; CHR, халконредуктаза; CHS, халкон-синтаза; DFR, дигидрофлавонол-4-редуктаза; F2H, флаванон-2-гидроксилаза; F3H, флаванон-3-гидроксилаза; F3’H, флавоноид-3′-гидроксилаза; F3’5’H, флавоноид-3’5′-гидроксилаза; FLS, флавонолсинтаза; ФНС, флавон-синтаза; HID, 2-гидроксиизофлаванондегидратаза; IFS, изофлавон-синтаза; LAR, лейкоантоцианидин редуктаза; LDOX, лейкоантоцианидиндиоксигеназа.
Нарингенин является общим предшественником флавонолов, антоцианов, проантоцианидинов, флавонов и изофлавонов. Он превращается в дигидрокэмпферол с помощью флаванон-3-гидроксилазы (F3H) (также называемой флаванон-3β-гидроксилазой). Флавоноид-3′-гидроксилаза (F3’H) и флавоноид F3’5’H-гидроксилаза (F3’5’H) катализируют гидроксилирование положений C3 ‘и C3’ / C5 ‘дигидрокэмпферола соответственно. Дигидрофлавонол-4-редуктаза (DFR) катализирует восстановление дигидрофлавонолов до лейкоантоцианидинов, которые затем превращаются в антоцианидины лейкоантоцианидиндиоксигеназой / антоцианидинсинтазой (LDOX / ANS).Лейкоантоцианидины и антоцианидины восстанавливаются до флаван-3-олов (например, катехина и эпикатехина) лейкоантоцианидинредуктазой (LAR) и антоцианидинредуктазой (ANR) соответственно. Дигидрофлавонолы также превращаются во флавонолы флавонолсинтазой (FLS).
Флавоны синтезируются из нарингенина флавон-синтазой I (FNS I) или флавон-синтазой II (FNS II) (Martens and Mithofer, 2005). Флаванон-2-гидроксилаза (F2H) катализирует гидроксилирование флаванонов (включая нарингенин) до 2-гидроксифлаванонов, которые впоследствии превращаются во флавоны, возможно, неизвестной дегидратазой (Akashi et al., 1998).
Изофлавон-синтаза (2-гидроксиизофлаванонсинтаза, IFS) катализирует первую стадию биосинтеза изофлавонов. IFS превращает флаваноны (например, ликвиртигенин и нарингенин) в 2-гидроксиизофлаваноны, затем 2-гидроксиизофлаваноны дегидратируются до изофлавонов под действием 2-гидроксиизофлаванондегидратазы (HID) (Akashi et al., 2005).
Упомянутые выше скелеты флавоноидов сильно модифицированы ферментами, такими как гликозилтрансферазы (GT), ацилтрансферазы и метилтрансферазы.Здесь мы сосредоточимся на ферментах, участвующих в биосинтезе молекул флавоноидного скелета.
Эволюционная история биосинтетических путей флавоноидов
Как правило, ферменты вторичных метаболических путей происходят от тех, которые участвуют в первичном метаболизме (Pichersky and Gang, 2000; Moghe and Last, 2015; Carrington et al., 2018), и ферменты метаболизма флавоноидов не являются исключением. CHS и CHI являются производными ферментов, участвующих в метаболизме жирных кислот: β-кетоацил ACP-синтазы и связывающего жирные кислоты белка соответственно (Ngaki et al., 2012; Weng and Noel, 2012a). CHS и β-кетоацил ACP-синтаза являются членами семейства поликетидсинтаз (PKS) типа III (Schuz et al., 1983). Ферменты на более поздних стадиях путей биосинтеза флавоноидов принадлежат к таким семействам, как суперсемейства 2OGD, CYP и короткоцепочечная дегидрогеназа / редуктаза (SDR) (Таблица 2). Члены этих суперсемейств широко вовлечены в первичный и вторичный метаболизм, что позволяет предположить, что растения приобрели функции ферментов в более поздних биосинтетических путях посредством дупликации генов и эволюции новых функций для дублированных генных продуктов.
Таблица 2 . Происхождение или семейство генов биосинтеза флавоноидов.
Основываясь на паттернах распределения подклассов флавоноидов, было высказано предположение, что пути биосинтеза флавоноидов могли развиваться в несколько этапов и что первыми ферментами биосинтеза флавоноидов были CHS, CHI и F3H (Stafford, 1991; Rausher, 2006 г.). Также было высказано предположение, что сначала развился CHS, затем F3H и затем CHI, потому что CHS катализирует первую коммитируемую стадию в путях, и стадия, катализируемая CHI, также может протекать спонтанно (Rausher, 2006).Флавоноиды широко распространены среди мхов, печеночников и сосудистых растений, но не встречаются у роголистников. Водоросли обычно не содержат флавоноидов (Rausher, 2006), но они были обнаружены в нескольких эволюционно расходящихся ветвях микроводорослей (Goiris et al., 2014). Эти наблюдения предполагают, что способность производить флавоноиды, возможно, эволюционировала несколько раз или что эта способность была широко утрачена в ходе эволюционных процессов. Анализ скорости эволюции шести генов, участвующих в биосинтезе антоцианов, показал, что вышестоящие гены ( CHS, CHI и F3H ) эволюционировали медленнее, чем нижележащие гены ( DFR, LDOX / ANS и UDP- глюкоза: флавоноид-3-O-глюкозилтрансфераза ) (Rausher et al., 1999). Вышестоящие гены могут быть эволюционно ограничены из-за их глубокого воздействия на пути развития. Кроме того, гены, кодирующие факторы транскрипции, которые регулируют биосинтез антоцианов, эволюционировали быстрее, чем структурные гены (Rausher et al., 1999).
Эволюционная история генов биосинтеза флавоноидов
CHS является представителем суперсемейства PKS типа III
Гены CHS широко распространены в растениях, от мохообразных до покрытосеменных (Jiang et al., 2008; Shimizu et al., 2017; Liou et al., 2018), но у других организмов они не обнаружены. Каждый вид наземных растений с доступными геномными данными имеет как минимум один предполагаемый ген CHS (Shimizu et al., 2017). CHS является членом суперсемейства PKS типа III, которое обеспечивает разнообразные поликетидные каркасы вторичных метаболитов (Рисунок 3) (Winkel-Shirley, 2001; Austin and Noel, 2003; Abe and Morita, 2010). ПКС типа III относятся к суперсемейству тиолаз (Jiang et al., 2008). Они обнаруживаются в наземных растениях, микроводорослях, грибах и бактериях, но не встречаются у животных или архей (Shimizu et al., 2017). PKS растений типа III сохраняют общую структуру свернутого белка и каталитическую триаду Cys-His-Asn, которые характеризуют фермент Escherichia coli 3-кетоацил-ACP-синтазы изоформы III (KASIII), который участвует в de novo жирных кислотах. синтез (Феррер и др., 1999; Остин, Ноэль, 2003). Количество генов наземных растений типа III PKS сильно варьируется среди видов одного и того же таксона; например, виды эвдикотов имеют от двух до 42 генов, тогда как их количество у грибов и бактерий относительно невелико (менее пяти) (Shimizu et al., 2017). Обилие ПКС типа III может вносить вклад в разнообразие специализированных метаболитов растений.
Рисунок 3 . Общий обзор филогении PKS и CHI-свернутых белков типа III. (A) Взаимоотношения между белками CHS / STS, базальным CHS наземных растений и белками, не относящимися к CHS, PKS типа III растений. Белки, не относящиеся к CHS, включают белки ARAS / ARS и семейства ASCL. Общая трехмерная структура белка является консервативной в PKS типа III и в клетке E.coli KASIII (αβαβα-складка). (B) Примеры продуктов типа III PKS. (C) CHI-складчатые белки в семействах CHI, CHIL и FAP имеют общую свернутую белковую структуру (открытая β-сэндвич-складка). ARAS, синтаза алкилрезорциловой кислоты; ARS, алкилрезорцинолсинтаза; ASCL, специфический для пыльников фермент, подобный халконсинтазе; CHIL, CHI-подобный белок; CHI, халконизомераза; CHS, халкон-синтаза; FAP, белок, связывающий жирные кислоты; KASIII, фермент изоформы III 3-кетоацил-ACP-синтазы; ПКС, поликетидсинтазы; СТС, стильбенсинтаза.
Функциональное разнообразие PKS типа III обычно происходит из различий в исходных молекулах, количестве стадий удлинения цепи и механизмах реакций циклизации (Austin and Noel, 2003; Abe and Morita, 2010). Стильбенсинтазы (STS), которые продуцируют стильбены, такие как ресвератрол, также относятся к PKS типа III. CHS и STS генерируют одни и те же промежуточные продукты из одних и тех же исходных молекул с использованием одних и тех же стадий удлинения цепи, но они катализируют разные внутримолекулярные циклизации и производят разные продукты (Austin et al., 2004). CHS и STS из одних и тех же родов растений обычно классифицируются как ближайшие соседи на филогенетических деревьях (рис. 3). Данные по покрытосеменным ( Vitis vinifera, Arachis hypogaea и Sorghum bicolor ), голосеменным растениям ( P. sylvestris ) и папоротникам ( Psilotum nudum ) предполагают, что после расхождения STS и CHS развивались независимо (Yu et al. al., 2005; Weng, Noel, 2012b).
Базальные наземные растения CHS и не-CHS
Eudicot Arabidopsis thaliana содержит четыре гена типа III PKS , включая один функциональный CHS (Kim et al., 2010). Напротив, мох мохообразный Physcomitrella patens имеет около 19 генов семейства CHS и четыре других гена типа III PKS (Jiang et al., 2006; Wolf et al., 2010). Пять генов семейства CHS мха были получены в результате дупликации всего генома, а четыре предположительно произошли в результате сегментарной дупликации и транспозиции (Wolf et al., 2010). Четыре гена активируются широкополосным УФ-В-излучением, а растения мха демонстрируют повышенные уровни производного флавонола при УФ-В-освещении.Эти наблюдения подтверждают гипотезу о том, что гены и ферменты, участвующие в реакции на УФ-стресс, эволюционировали при переходе от воды к суше, когда первые растения подвергались воздействию повышенного уровня солнечного света (Wolf et al., 2010).
В печеночнике мохообразных M. polymorpha УФ-В-облучение и лишение питательных веществ значительно увеличивают общее содержание флавоновых гликозидов (Albert et al., 2018; Clayton et al., 2018). M. polymorpha имеет 24 гена семейства CHS (Bowman et al., 2017), и один из них значительно усиливается обработкой УФ-В стрессом. Эта индукция была усилена в трансгенных растениях Marchantia , которые сверхэкспрессировали ген, кодирующий фоторецептор UV RESISTANCE LOCUS8 (UVR8) (Clayton et al., 2018). Кроме того, трансгенные растения Marchantia , сверхэкспрессирующие ген фактора транскрипции MpMyb14 , показали повышенные уровни экспрессии того же гена CHS в нормальных условиях роста (Albert et al., 2018).Мутанты Knockout Mpmyb14 были частично нарушены увеличением уровней экспрессии CHS в условиях дефицита азота (Kubo et al., 2018). Результаты предполагают, что эти виды печеночников имеют по крайней мере один ген CHS , который активируется путем передачи сигнала UVR8.
Филогенетический анализ показывает тесную взаимосвязь между CHS базальных наземных растений (мохообразные и ликофиты) и ПКС не-CHS типа III (рисунок 3) (Wanibuchi et al., 2007; Yu et al., 2018). Группа, не относящаяся к CHS, включает множество ферментов, участвующих в биосинтезе вторичных метаболитов, таких как акридон-синтазы, пирон-синтазы, бибензилсинтазы и синтазы p -кумароилтриуксусной кислоты (Winkel-Shirley, 2001). Эти ферменты, не относящиеся к CHS PKS, эволюционировали в результате повторяющихся дупликаций, мутаций и функциональной диверсификации генов из своих предковых растительных ферментов.
Эктопическая экспрессия гена CHS от мохообразного P. patens или ликофита Selaginella moellendorffii может частично дополнять фенотип A.thaliana CHS -нулевой мутант, transparent testa 4 (Liou et al., 2018). Кристаллические структуры ХГС из P. patens, S. moellendorffii , монилофита Equisetum arvense , голосеменных растений Pinus sylvestris и покрытосеменных A. thaliana показали, что реакционная способность каталитического остатка Cys (Cys164 в M. sativa CHS2) изменилась за 500 миллионов лет эволюции наземных растений. Остатки Cys в трех последних ветвях (монилофиты, голосеменные и покрытосеменные) присутствуют в форме тиолат-аниона, что придает им более сильную нуклеофильную силу (Liou et al., 2018).
ПКС типа III демонстрируют широкую неразборчивость субстрата. CHSs не принимают объемные субстраты, но Lycophyte Huperzia serrata HsPKS1 обнаруживает замечательную устойчивость к субстратам и каталитический потенциал (Wanibuchi et al., 2007; Morita et al., 2011). In vitro , HsPKS1 производит халкон нарингенина и другие поликетиды, включая ароматические трициклические соединения пиридоизоиндола, которые не встречаются в натуральных продуктах. Замена одной аминокислоты в HsPKS1 увеличивает объем его полости активного центра и изменяет длину цепи продукта и механизм реакции циклизации.Эта неразборчивость субстратов в PKS типа III обеспечивает разнообразные поликетидные каркасы для последующего биосинтеза вторичных метаболитов в наземных растениях.
Семейства ARAS / ARS и ASCL
ПКС типа III, не относящиеся к CHS, также синтезируют поликетиды из субстратов жирного ацил-КоА. Фенольные липиды, такие как алкилрезорцины, синтезируются алкилрезорцинолсинтазами (ARS) и синтазами алкилрезорциловой кислоты (ARAS) в однодольных растениях S. bicolor и O. sativa соответственно (Cook et al., 2010; Мацузава и др., 2010). Алкилрезорцины в зерновых культурах проявляют противогрибковую и аллелопатическую активность.
Гидроксиалкил-α-пироновые соединения, которые являются предшественниками спорополленина в экзине пыльцевой стенки, синтезируются специфичными для пыльников халконсинтазоподобными ферментами (ASCL), которые специфически и временно экспрессируются в пыльниках A. thaliana (PKSA / LAP6 и LAP5 / PKSB) (Добрица и др., 2010; Ким и др., 2010). Растения с двойными мутантами pksa pksb обладают мужской стерильностью.
Филогенетический анализ показывает, что семейства ARAS / ARS и ASCL подразделяются на отдельные группы (рис. 3) (Shimizu et al., 2017). Бактерии, грибы и мхи также имеют гены, кодирующие ПКС типа III, которые продуцируют эти длинноцепочечные жирные ацилсодержащие поликетиды (Colpitts et al., 2011; Shimizu et al., 2017; Li et al., 2018). Следовательно, эти ПКС типа III участвуют в метаболизме липидов в трех царствах живых организмов.
CHI-Fold Proteins: CHIs и CHILs
CHI и CHI-подобные белки (CHIL) являются членами семейства CHI-fold, которое также включает белки, связывающие жирные кислоты (FAP), которые участвуют в биосинтезе жирных кислот (рис. 3).Эти белки имеют общую сложенную трехмерную структуру (Jez et al., 2000; Ngaki et al., 2012; Kaltenbach et al., 2018). Семейство FAP распространено среди многих видов бактерий, грибов и растений. В CHIL частично отсутствуют каталитические аминокислотные остатки, консервативные в CHI, но они связываются с CHS и усиливают свою активность (Ban et al., 2018). Филогенетический и геномный анализ белков CHI-фолда предполагает, что CHIL сначала появились у мхов и произошли от FAP, а затем послужили предками CHI (Ngaki et al., 2012; Morita et al., 2014; Jiang et al., 2015). ДЕТИ составляют группу, отличную от групп ОМС и ФАП (рис. 3). Мох P. patens имеет два гена CHIL и четыре FAP , но, по-видимому, не имеет генов CHI (Ngaki et al., 2012; Cheng et al., 2018). Печеночник M. paleacea имеет CHI , CHIL и два FAP гена (Cheng et al., 2018), а ликофит S. moellendorffii имеет CHI , CHIL , и три гена FAP . A. thaliana имеет CHI , CHIL и три FAP гена, тогда как бобовое Glycine max имеет тип I CHI , три типа II CHI , два CHIL и шесть генов FAP (Dastmalchi, Dhaubhadel, 2015; Ban et al., 2018). Эти результаты предполагают, что количество генов CHI и CHIL остается низким во многих линиях растений, но что бобовые растения имеют несколько генов CHI .
ОМС типа I и типа II в сосудистых растениях
Существует два типа ОМС. CHI типа I распространены повсеместно в сосудистых растениях, тогда как CHI типа II специфичны для бобовых и участвуют в синтезе флавоноидов во время симбиоза клубеньков, фиксирующих азот (Shimada et al., 2003; Subramanian et al., 2007). Филогенетический анализ показывает, что ОМС типа II образуют отдельную группу от ОМС типа I (рис. 3). CHI типа II, вероятно, произошли от предковых белков CHI-укладки (Cheng et al., 2018).
CHI типа II изомеризуют как нарингенин халкон, так и изоликвиритигенин (6′-дезоксихалкон) с образованием (2 S ) -нарингенина и ликвиритигенина (5-дезоксифлаванона), соответственно. Модель бобовых Lotus japonicas имеет гены типа I и три типа II CHI (Shimada et al., 2003). Эти четыре гена CHI образуют тандемный кластер в области генома размером 15 т.п.н. У соевых бобов ген CHI типа I и два типа II организованы в кластер генов на хромосоме 20 (Dastmalchi and Dhaubhadel, 2015) и, вероятно, происходят от тандемных дупликаций генов.Роль CHIs в симбиозе бобовых остается неясной. Азотфиксирующие корневые клубеньковые симбиозы встречаются у четырех отрядов покрытосеменных: Fabales, Fagales, Cucurbitales и Rosales (последние три известны как актиноризные растения). Экспрессия гена CHI повышается в клубеньках актиноризного растения Datisca glomerata (Gifford et al., 2018), что позволяет предположить, что эти CHI способствуют увеличению содержания флавоноидов.
ОМС в базальных наземных растениях
Модель A.thaliana chi —-дефицитные мутанты tt5 дают бледно-желтые семена из-за значительного снижения продукции проантоцианидина. Этот фенотип в значительной степени дополняется эктопической экспрессией гена CHI либо из печеночника M. paleacea ( MpCHI1 ), либо из ликофита S. moellendorffii ( SmCHI1 ) (Cheng et al., 2018). CHI от базальных наземных растений обладают широкой субстратной специфичностью и больше похожи на CHI типа II, чем на CHI типа I.Филогенетический анализ показывает, что MpCHI1 и SmCHI1 образуют отдельные группы от CHI типа I и типа II (рисунок 3) (Cheng et al., 2018; Kaltenbach et al., 2018).
Печеночник M. polymorpha также имеет единственный ген CHI (Clayton et al., 2018), а мутанты Marchantia chi не содержат определяемых уровней флавоновых соединений. Эти мутанты chi очень чувствительны к воздействию УФ-В стресса. Эти результаты показывают, что этот базальный вид растений уже имеет ген, кодирующий bona fide CHI, чтобы катализировать циклизацию нарингенина халкона в пути биосинтеза флавоноидов.
Семья ЧИЛ
CHIL классифицируются как CHI-складчатые белки типа IV и обнаруживаются у базальных и высших видов растений, включая мхи, печеночники, ликофиты, папоротники, голосеменные и покрытосеменные. ДЕТИ не имеют добросовестных ОМС. Однако нокдаун РНКи экспрессии CHIL в Petunia hybrida и Torenia hybrida приводил к снижению уровней общих флавоноидов в цветках (Morita et al., 2014). Три независимых мутанта Ipomoea nil (японская ипомея) с изменениями в энхансере продукции флавоноидов (ген CHIL ) показали фенотип бледно окрашенных цветков (Morita et al., 2014). Мутанты с потерей функции CHIL в A. thaliana демонстрируют снижение уровней проантоцианидина и флавонолов в семенах и флавонолов в листьях (Jiang et al., 2015). Однако ген A. thaliana CHIL не смог спасти фенотипы мутантов tt5 (Jiang et al., 2015). Печеночник M. polymorpha имеет ген CHIL (Clayton et al., 2018), а общее содержание флавонов снижено у мутантов Marchantia chil при нормальных условиях роста и при обработке УФ-В.Таким образом, CHILs увеличивают общее производство флавоноидов, но выполняют функции, отличные от CHIs. Как упоминалось выше, недавно было показано, что CHIL из различных линий растений ( A. thaliana, O. sativa, S. moellendorffii и P. Patens ) могут связывать CHS и повышать активность CHS (Ban et al., 2018 ). CHI также связывают CHS (Jorgensen et al., 2005), и возможно, что CHI типа I и типа II произошли от CHIL и приобрели свою активность CHI в ходе последующих эволюционных процессов (Ban et al., 2018).
Приобретение ферментативной активности CHI в процессе эволюции
Появление ферментативных CHI у растений — интересная тема в эволюции белков, потому что CHIL и FAP являются неферментативными белками. Расщелина в активном сайте CHI состоит из трех высококонсервативных аминокислотных остатков (Arg36, Thr48 и Tyr106 в последовательности CHI M. sativa типа II), и соседние с ними остатки также консервативны (Jez et al., 2000; Ngaki et al., 2012; Kaltenbach et al., 2018). Общие предковые белки CHI и CHIL были выведены в результате обширного филогенетического анализа, и оказалось, что все три ключевых каталитических остатка были консервативными в предковых белках, но были неактивными (Kaltenbach et al., 2018). Авторы выполнили поэтапный скрининг на основе активности рекомбинантных предковых белков с использованием экспрессионной системы E. coli . Результаты показали, что мутации в аминокислотных остатках, отличных от каталитических остатков, необходимы для инициации развития CHI и для приобретения каталитической активности CHI.
Семейство 2-оксоглутарат-зависимых диоксигеназ: F3H, FNS I, FLS и LDOX / ANS
Суперсемейство 2OGD — одно из крупнейших семейств белков в царстве растений. Его представители широко распространены среди бактерий, грибов, растений, позвоночных и даже вирусов (van den Born et al., 2008; Farrow, Facchini, 2014; Markolovic et al., 2015; Wu et al., 2016). 2OGD — это негемовые железосодержащие ферменты, локализованные в цитозоле. 2OGD включают 2-оксоглутарат (2OG или α-кетоглутарат) и активированный O 2 в различные субстраты с образованием окисленных продуктов вместе с сукцинатом и CO 2 (RH + 2OG + O 2 → ROH + сукцинат + CO 2 ).2ОГД катализируют различные окислительные реакции, включая гидроксилирование, галогенирование, десатурацию и эпимеризацию (Martinez and Hausinger, 2015), и играют важную роль в репарации ДНК и РНК, метаболизме жирных кислот, ощущении кислорода и биосинтезе природных продуктов (Farrow and Facchini, 2014; Hagel and Facchini, 2018; Herr and Hausinger, 2018; Islam et al., 2018).
У растений 2OGDs участвуют в деметилировании гистонов, ощущении железа, метаболизме фитогормонов и биосинтезе вторичных метаболитов (rev. Farrow and Facchini, 2014).Филогенетический анализ 2OGD растений из Chlamydomonas reinhardtii, P. patens, S. moellendorffi, Picea abies, O. sativa и A. thaliana обнаружил 3 класса, которые авторы назвали DOXA, DOXB и DOXC (Kawai et al. др., 2014). Каждый класс содержит белки всех шести видов. Класс DOXA содержит гомологи E. coli AlkB; эти ферменты участвуют в репарации ДНК (Lindahl et al., 1988; Meza et al., 2012; Mielecki et al., 2012). Класс DOXB содержит пролил-4-гидроксилазы, которые катализируют гидроксилирование остатков пролина в белках клеточной стенки растений (Hieta and Myllyharju, 2002).Белки класса DOXC участвуют в метаболизме фитогормонов и биосинтезе вторичных метаболитов, включая флавоноиды, терпеноиды, алкалоиды и глюкозинолаты. Число генов, кодирующих ферменты DOXA и DOXB, ограничено у шести видов, однако гены DOXC значительно расширены у наземных растений (Kawai et al., 2014).
Четыре фермента биосинтеза флавоноидов, F3H, FNS I, FLS и LDOX / ANS, относятся к классу DOXC. Филогенетический анализ показал, что гены класса DOXC можно разделить на более чем 50 клад (Kawai et al., 2014). Гены F3H, и FNS I находятся в кладе DOXC28, а гены FLS и LDOX / ANS находятся в DOXC47. Было высказано предположение, что среди генов биосинтеза флавоноидов в суперсемействе 2OGD первым появился F3H (Rausher, 2006). FNS I, по-видимому, существует только у Apiaceae, и вполне вероятно, что FNS I произошел от F3H как парафилетический ген (Martens et al., 2003; Gebhardt et al., 2005).
Arabidopsis thaliana содержит один ген F3H , шесть генов FLS ( AtFLS1, — AtFLS6 ), один ген LDOX / ANS и ни один ген FNS. AtFLS1 является основным FLS (Owens et al., 2008a; Saito et al., 2013). Анализ структурной дивергенции между дублированными генами показал, что транспонированная дупликация (<16 миллионов лет назад) объясняет взаимосвязь между AtFLS6 (At5g43935) и AtF3H (At3g51240) (Wang et al., 2013).Кроме того, связь между AtFLS1 (At5g08640) и AtFLS5 (At5g63600), вероятно, объясняется дупликацией всего генома, а отношения между AtFLS2 (At5g63580) и AtFLS3 (At5g63590), AtFLS3 и AtFLS4 (At5g635LS5) и, скорее всего, являются объясняется тандемными локальными дупликациями (Wang et al., 2013). Было показано, что AtLDOX / ANS A. thaliana продуцирует флавонолы in planta (Preuss et al., 2009). Кроме того, FLS и LDOX / ANS могут частично дополнять функцию F3H in vivo , и это приводит к фенотипу утечки у мутантов tt6 с нулевыми мутациями в AtF3H (Owens et al., 2008b).
В совокупности результаты предполагают, что F3H является наследственным геном 2OGD для биосинтеза флавоноидов, и что FLS и FNS I эволюционировали посредством дивергенции от F3H . Не было обнаружено явных ортологов генов клады DOXC28 или DOXC47 в P. patens и S. moellendorffi , хотя эти растения продуцируют флавонолы ( P. patens ) и флавоны ( S. moellendorffi ) (Wolf и другие., 2010; Вен и Ноэль, 2013). Однако печеночник Plagiochasma appendiculatum имеет активный FNS I (PaFNS I), и филогенетический анализ показал, что PaFNS 1 родственен белкам FNS I и F3H покрытосеменных, хотя он и не принадлежит к той же кладе, что и они (Han и др., 2014). Эти данные предполагают, что в Physcomitrella и Selaginella 2OGD присутствуют в разных кладах или что неродственные ферменты выполняют те же функции, что и F3H, FNS и / или FLS.
PaFNS I может преобразовывать нарингенин либо в 2-гидроксинарингенин, либо в апигенин (Han et al., 2014). Хвощ обыкновенный E. arvense L также обладает активностью FNS I типа 2OGD (Bredebach et al., 2011). Дальнейшие исследования генов 2OGD мохообразных, ликофитов и папоротников помогут прояснить их эволюционные процессы.
Суперсемейство цитохрома P450: FNSII, F2H, IFS, F3’H и F3’5’H
CYP широко распространены у вирусов, архей, бактерий и эукариот.Они катализируют реакции монооксигеназы / гидроксилирования в различных первичных и вторичных метаболических процессах путем внедрения атома O из молекулярного O 2 (Mizutani and Ohta, 2010). У эукариот CYP представляют собой гем-содержащие мембранные белки, локализованные на цитозольной поверхности эндоплазматического ретикулума.
У растений CYP образуют самое большое суперсемейство ферментов и составляют около 1% от общего числа продуктов генов (Mizutani, Ohta, 2010; Nelson and Werck-Reichhart, 2011; Kawai et al., 2014). CYP делятся на семейства (например, CYP75), которые имеют ~ 40% или более идентичности аминокислотных последовательностей, а те, у которых идентичность 55% или более, делятся на подсемейства (например, CYP75A). Кроме того, семейства CYP растений (от CYP71 до CYP99 и CYP701–) можно разделить на кланы, члены которых произошли от одного предка. CYP наземных растений образуют 11 кланов, семь из которых (кланы 51, 74, 97, 710, 711, 727 и 746) состоят из отдельных семейств CYP, а остальные четыре (кланы 71, 72, 85 и 86) включают белки во множестве семейств CYP (Nelson and Werck-Reichhart, 2011).Зеленые водоросли содержат CYP в пяти односемейных кланах (кланы 51, 97, 710, 711 и 746), и члены этих кланов участвуют в фундаментальных биологических процессах, таких как биосинтез стеролов, ксантофиллов и фитогормонов (Nelson, 2006; Nelson и Werck-Reichhart, 2011). Следовательно, вполне вероятно, что эти кланы включают в себя наследственные CYP. Многосемейные кланы CYP стали очень разнообразными в процессе эволюции растений (Nelson and Werck-Reichhart, 2011). Некоторые семейства CYP в этих кланах (кланы 71, 72, 85 и 86) присутствуют в мохообразных и / или печеночниках, но не в зеленых водорослях.Дополнительные новые семейства CYP были получены в результате поэтапных процессов, следующих за эволюцией сосудистых растений.
Ферменты биосинтеза флавоноидов (члены семейств CYP75 и CYP93) классифицируются в клан 71. Этот клан содержит наибольшее количество семейств CYP и, помимо ферментов биосинтеза флавоноидов, он включает семейства, участвующие в биосинтезе фенилпропаноидов (CYP73, CYP84, CYP98), алкалоиды (CYP80, CYP82, CYP719), терпеноиды (CYP76, CYP99, CYP705, CYP706, CYP726) и глюкозинолаты (CYP79, CYP83).Клановые семейства 71 CYP73, CYP74, CYP78, CYP88, CYP98, CYP701, CYP703, CYP736 и CYP761 присутствуют в мохообразных, что позволяет предположить, что это могут быть предковые семейства.
Семейство CYP75: F3′H и F3′5′H
Ферменты F3’H и F3’5’H обычно принадлежат к семействам CYP75B и CYP75A соответственно. Ферменты семейства CYP75 однодольных и двудольных образуют отдельные кластеры внутри каждого подсемейства, предполагая, что функции F3’H и F3’5’H были установлены до расхождения однодольных и двудольных.Однако есть и исключения. У Asteraceae F3’5’H принадлежит подсемейству CYP75B и образует отдельный кластер от F3’H (Seitz et al., 2006). Это предполагает, что Asteraceae F3’H приобрел активность F3’5’H до видообразования, но после разделения однодольных и двудольных. Точно так же CYP75B4 риса катализирует 5′-гидроксилирование 3′-метоксифлавона хризоэриола, а также функционирует как F3’H (Lam et al., 2015). Рис содержит CYP75B3 как F3’H и CYP75A11 как нефункциональный F3’5’H.Такое распределение генов CYP в подсемействах CYP75A и CYP75B также обнаружено у других растений Poaceae. Эти данные предполагают, что подсемейства CYP75A и CYP75B разделились до расхождения однодольных и двудольных, и что гены в подсемействе CYP75B позже приобрели активность F3’5’H, по крайней мере, у Asteraceae и Poaceae.
Arabidopsis thaliana имеет единственный ген для F3’H (CYP75B1, At5g07990) и нет генов, соответствующих F3’5’H, FNS II или F2H. Ген CYP75B1, по-видимому, связан с геном CYP701A (At5g25900), который кодирует фермент, участвующий в биосинтезе гиббереллина, посредством транспонированной дупликации, которая произошла 16–107 миллионов лет назад, и было высказано предположение, что CYP75B1 является родительским локусом CYP701A (Wang и другие., 2013). Однако семейство CYP701 распространено среди мохообразных и сосудистых растений, в то время как семейство CYP75 встречается у голосеменных и покрытосеменных. Кроме того, филогенетический анализ показал, что семейство CYP761, специфичное для мха / печеночника, тесно связано с семейством CYP75 (Nelson and Werck-Reichhart, 2011). Эти данные позволяют предположить, что либо CYP701, либо CYP 761 являются наследственным семейством CYP75. Хотя 3′-гидроксилированные производные флавона были обнаружены в M. polymorpha и S.moellendorffii (Markham et al., 1998), члены семейства CYP75 можно найти только у голосеменных и покрытосеменных (Nelson, Werck-Reichhart, 2011). Следовательно, у мохообразных, ликофитов и папоротников ферменты из других семейств CYP могут функционировать как F3’H и / или F3’5’H.
Подобные несоответствия во взаимоотношениях между метаболитами и генами наблюдаются также в метаболизме фенилпропаноидов. Среди CYP, участвующих в метаболизме фенилпропаноидов из клана 71, CYP73A (циннамат-4-гидроксилаза, C4H), CYP98A ( p -кумароилшикимат-3′-гидроксилаза, C3’H), CYP73 и CYP98 впервые появились в печеночниках и мхах. тогда как CYP84 (ферулат 5-гидроксилаза, F5H) обнаружен только у покрытосеменных (Nelson and Werck-Reichhart, 2011).Однако единицы сирингиллигнина из фенилпропаноидного пути распространены в наземных растениях (Bowman et al., 2017), что позволяет предположить, что гены в других семействах CYP могут обладать активностью F5H. Фактически, Selaginella CYP788A1 функционирует как F5H (Weng et al., 2008).
Семейство CYP93: FNS II, F2H и IFS
Полногеномный анализ генов семейства CYP93 из 60 зеленых растений показал, что семейство CYP93 обнаружено только у покрытосеменных (Du et al., 2016). Среди 10 подсемейств (CYP93A – CYP93K) CYP93A является предковой группой, распространенной как у однодольных, так и у двудольных; CYP93B и CYP93C распространяются только двудольными; CYP93G и CYP93J встречаются только в однодольных; и CYP93E и CYP93F специфичны для бобовых и злаковых, соответственно.Таким образом, семейство CYP93 демонстрирует эволюцию, специфичную для линии растений.
Семейство CYP93 содержит ферменты, участвующие в биосинтезе флавонов и изофлавонов (FNS II, F2H и IFS). Однодольные FNS II и F2H принадлежат к подсемейству CYP93G, а двудольные — к подсемейству CYP93B. Таким образом, филогенетический анализ позволяет предположить, что функции ферментов FNSII и F2H были установлены после расхождения однодольных и двудольных. Фермент IFS в бобовых является членом подсемейства CYP93C, которое может происходить из подсемейства CYP93B (Du et al., 2016).
Как и в случае с семейством CYP75, существуют несоответствия во взаимоотношениях между метаболитами и генами в семействе CYP93. Гены семейства CYP93 обнаружены только у покрытосеменных (Nelson and Werck-Reichhart, 2011), но флавоны широко распространены в растениях, от мохообразных до покрытосеменных (рис. 1). Это предполагает, что ферменты, участвующие в биосинтезе флавонов, принадлежат к семейству CYP93 у некоторых видов покрытосеменных, но разные семейства CYP и / или другие ферменты играют одинаковые роли в других таксонах растений.Напр., Семейство Apiaceae адаптировало FNS I 2OGD-типа вместо FNS II CYP-типа для продуцирования флавонов (Gebhardt et al., 2007).
Изофлавоноиды — это типичные флавоноиды, содержащиеся в бобовых, а гены биосинтеза изофлавонов встречаются только в бобовых. Однако изофлавоноиды также обнаружены в не бобовых растениях, включая Iris , который содержит широкий спектр изофлавонов, некоторые мхи (например, Bryum capillare ), голосеменные, а также однодольные и двудольные покрытосеменные (Dewick, 1994; Lapcik, 2007). ).Это говорит о том, что растения независимо приобрели способность продуцировать изофлавоноиды в ходе эволюции семейств CYP, за исключением CYP93C и / или других ферментов, не относящихся к CYP, которые функционируют как IFS.
Семейство короткоцепочечных дегидрогеназ / редуктаз: DFR, ANR и LAR
DFR, ANR и LAR являются членами суперсемейства SDR, которое широко распространено у вирусов, архей, прокариот и эукариот (Jornvall et al., 1999; Kavanagh et al., 2008). SDR составляют одно из крупнейших семейств NAD (P) (H) -зависимых оксидоредуктаз и участвуют в первичном метаболизме липидов, углеводов и гормонов, а также во вторичном метаболизме таких молекул, как терпеноиды, алкалоиды и фенольные соединения (Jornvall et al. al., 1999; Кавана и др., 2008; Tonfack et al., 2011). Несмотря на низкое общее сходство последовательностей SDR (15–30%), SDR обладают консервативной трехмерной структурой, состоящей из укороченного по Россману β-листа, окруженного α-спиралями для связывания нуклеотидов. Как правило, SDR можно разделить на несколько типов («классический», «расширенный», «промежуточный», «дивергентный» и «сложный») на основе их первичных структур, мотивов связывания кофакторов и активных сайтов (Kavanagh et al. , 2008; Moummou et al., 2012).Недавнее исследование показало, что «промежуточные» и «сложные» типы не встречаются у растений, в то время как типы «атипичные» и «неизвестные» встречаются у растений. Таким образом, SDR растений можно разделить на пять типов: «классический», «дивергентный», «расширенный», «атипичный» и «неизвестный») (Moummou et al., 2012). Среди SDR растений основными типами являются «классический» и «расширенный», как и у других организмов. «Классический» тип состоит из примерно 250 аминокислотных остатков, а «расширенный» тип имеет домен из 100 дополнительных аминокислотных остатков в С-концевой области.«Нетипичный» тип, необычный тип SDR, был включен в семейство SDR из-за его складчатой структуры Россмана, которая типична для SDR (Moummou et al., 2012).
Анализ последовательности генома с использованием 10 видов, включая C. reinhardtii, P. patens, S. moellendorffii , четыре двудольных ( A. thaliana, Populus trichocarpa, V. vinifera и G. max ) и три однодольных ( O. sativa, S. bicolor и Z. mays ) показали, что большинство SDR растений можно разделить на 49 семейств, которые распределены между пятью типами, упомянутыми выше (Moummou et al., 2012). Ферменты DFR и ANR относятся к семейству SDR108E «расширенного» типа. Ферменты LAR принадлежат к семейству SDR460A, которое является «атипичным» типом (Kallberg et al., 2010; Moummou et al., 2012).
Семейство SDR108E: DFR и ANR
По сравнению с другими семействами, которые содержат несколько генов на вид, семейство SDR108E содержит наибольшее количество генов SDR; например, это семейство включает 24 гена в A. thaliana и 44 гена в O. sativa .Кроме того, семейство SDR108E демонстрирует самую низкую среднюю идентичность последовательностей, что указывает на высокую степень разнообразия генов семейства SDR108E (Moummou et al., 2012). Распределение генов семейства SDR108E у 10 видов указывает на то, что это семейство значительно расширилось у сосудистых растений.
Помимо ферментов DFR и ANR, семейство SDR108E содержит другие ферменты, участвующие во вторичном метаболизме, включая циннамоил-КоА-редуктазу (CCR) для биосинтеза лигнина и фенилацетальдегидредуктазу для производства летучего 2-фенилэтанола.Кроме того, к этому семейству относятся ферменты, участвующие в метаболизме фитогормонов, такие как редуктаза фазовой кислоты для катаболизма абсцизовой кислоты (Weng et al., 2016) и BEN1 для гомеостаза брассиностероидов (Yuan et al., 2007). Каждый тип фермента образует отдельный кластер на филогенетическом дереве. Кластеры CCR и фенилацетальдегидредуктазы содержат ферменты из P. patens и S. moellendorffii , тогда как кластеры DFR, ANR, фазовых кислотных редуктаз и ферментов BEN1 получены из цветковых растений.Эти данные предполагают, что эти DFR, ANR и другие ферменты появились позже, чем CCR и фенилацетальдегидредуктазы.
Другие семейства SDR демонстрируют паттерны экспансии генов, аналогичные SDR108E. Эти семейства (SDR110C, SDR114C, SDR65C и SDR460A) содержат гены, участвующие в биосинтезе алкалоидов, терпеноидов, фенилпропаноидов и фитогормонов (Moummou et al., 2012). Менее диверсифицированные семейства SDR содержат гены, участвующие в первичном метаболизме, например, биосинтезе липидов и хлорофилла (Moummou et al., 2012).
Семейство SDR460A: LAR
Семейство SDR460A также называют семейством PIP, названным в честь первых трех ферментов (пинорезинол-ларицирецинолредуктаза, изофлавонредуктаза и редуктаза бензилового эфира фенилкумарана), которые, как было обнаружено, принадлежат к этому семейству (Gang et al., 1999; Min et al., 2003; Wang et al., 2006). Кроме того, к этому семейству также принадлежат веститонредуктаза, эвгенолсинтаза и изоэвгенолсинтаза (Koeduka et al., 2008). Пинорезинол-ларицирецинолредуктаза и фенилкумаран бензиловый эфир редуктазы действуют в пути биосинтеза лигнана; изофлавонредуктаза и веститонредуктаза участвуют в биосинтезе изофлавоноидов; а эвгенолсинтаза и изоэвгенолсинтаза участвуют в биосинтезе летучих фенилпропенов.Таким образом, члены семейства SDR460A участвуют в биосинтезе различных фенольных соединений. Члены семейства SDR460A (SDR «атипичного» типа) ограничены и значительно расширились у сосудистых растений (Moummou et al., 2012), что позволяет предположить, что они необходимы для процветания сосудистых растений.
Филогенетический анализ LAR различных растений показал, что LAR растений можно разделить на два кластера: белки двудольных растений и белки однодольных и голосеменных растений (Wang et al., 2018). Следовательно, LAR однодольных растений более близки к LARS голосеменных, чем LARS двудольных.
Перспективы на будущее
Прогресс в технологиях метаболомики, включая хемоинформатику, и обширная геномная информация о генах биосинтеза флавоноидов способствовали полному пониманию эволюции метаболизма флавоноидов / фенилпропаноидов в царстве растений. В этом обзоре мы сосредоточились на эволюции ферментов, участвующих в биосинтезе молекул флавоноидного скелета.Основные структуры флавоноидов образуют ПКС, ХС типа III. Широкая разнородность субстратов и функциональное разнообразие ПКС типа III могут быть движущей силой расширения химического разнообразия специализированных метаболитов. В других специализированных процессах метаболизма изомеразы (например, оксидоскваленсинтаза) и лиазы (например, терпенсинтаза) также участвуют в образовании каркаса и могут вносить вклад в химическое разнообразие вторичных метаболитов. Ферменты модификации, такие как GT (гликозилтрансферазы) и ацилтрансферазы, также вносят большой вклад в огромное разнообразие флавоноидов и других вторичных метаболитов.Интересно, что у растений есть два типа флавоноидных GT: цитозольные семейства 1 GT и вакуолярные гликозидгидролазы семейства 1 (Gh2) (Cao et al., 2017). Ацилирование также катализируется дифференциально локализованными ферментами: цитозольными ацилтрансферазами BAHD и вакуолярными ацилтрансферазами SCPL, происходящими из серинкарбоксипептидазы (Milkowski and Strack, 2004; Moghe and Last, 2015). До сих пор неизвестно, почему растения развили эти дифференцированно локализованные ферменты для модификации флавоноидов и других специализированных метаболитов.Эволюция GT семейства 1, Gh2 и ацилтрансфераз BAHD у растений была рассмотрена в других источниках (St-Pierre and De Luca, 2000; Yu et al., 2009; Tuominen et al., 2011; Yonekura-Sakakibara and Hanada). , 2011; Caputi et al., 2012; Moghe, Last, 2015; Cao et al., 2017). На протяжении своей долгой истории растения выстраивали свои метаболические пути, чтобы адаптироваться к своей среде обитания и условиям роста в тканях и органах. Детальное понимание эволюционной истории метаболических ферментов, участвующих в биосинтезе, модификации, транспорте, секреции, регуляции транскрипции и химическом разнообразии, поможет нам в разработке специализированных метаболических путей для производства желаемых метаболитов с минимальными затратами энергии.
Авторские взносы
KY-S предложил концепцию. KY-S, YH и RN разработали его и написали рукопись.
Финансирование
Эта работа была частично поддержана программой JSPS KAKENHI (номер гранта 17K07460 для KY-S), Комплексной липидологической программой RIKEN (YH) и проектом NARO Bio-ориентированного института развития технологий (исследовательская программа по развитию инновационные технологии) (РН).
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Список литературы
Абэ И., Морита Х. (2010). Структура и функция надсемейства халконсинтаз поликетидсинтаз III типа растений. Nat. Prod. Rep. 27, 809–838. DOI: 10.1039 / b
8n
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Афенди, Ф. М., Окада, Т., Ямазаки, М., Хираи-Морита, А., Накамура, Ю., Накамура, К., и др. (2012). Базы данных семейства ранцевых: интегрированные базы данных метаболитов и видов растений для многогранного исследования растений. Физиология растительных клеток . 53: e1. DOI: 10.1093 / pcp / pcr165
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Акаси Т., Аоки Т. и Аябе С. (1998). Идентификация кДНК цитохрома p450, кодирующей (2s) -флаванон-2-гидроксилазу солодки ( glycyrrhiza echinata l; Fabaceae ), которая представляет собой ликодион-синтазу и флавон-синтазу ii. FEBS Lett. 431, 287–290. DOI: 10.1016 / S0014-5793 (98) 00781-9
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Акаси, Т., Аоки, Т., и Аябе, С. (2005). Молекулярная и биохимическая характеристика 2-гидроксиизофлаванондегидратазы. Участие карбоксилэстеразоподобных белков в биосинтезе изофлавонов бобовых. Plant Physiol. 137, 882–891. DOI: 10.1104 / стр.104.056747
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Акимото, Н., Ара, Т., Накадзима, Д., Суда, К., Икеда, К., Такахаши, С. и др. (2017). Поиск флавоноидов: система для комплексной аннотации флавоноидов с помощью масс-спектрометрии. Sci. Отчет 7: 1243. DOI: 10.1038 / s41598-017-01390-3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Альберт, Н. В., Тримавитана, А. Х., Макги, Т. К., Клейтон, В. А., Деролес, С. К., Швинн, К. Е. и др. (2018). Генетический анализ печеночника Marchantia polymorpha показывает, что активация r2r3myb продукции флавоноидов в ответ на абиотический стресс является древним признаком наземных растений. New Phytol. 218, 554–566. DOI: 10.1111 / нф. 15002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Андерсон, Ø. М., и Маркхэм, К. Р. (2006). Флавоноиды: химия, биохимия и приложения. Бока-Ратон: CRC Press.
Google Scholar
Асакава, Ю., Такикава, К., Тойота, М., и Такемото, Т. (1982). Хемосистематика мохообразных 12. Новые производные бибензила и сесквитерпеноиды энт-купаренового типа из видов радулы. Фитохимия 21, 2481–2490.DOI: 10.1016 / 0031-9422 (82) 85245-X
CrossRef Полный текст | Google Scholar
Асакава Ю., Тойота М. и Такемото Т. (1978). 7 новых бибензилов и дигидрохалкон из radula-variabilis. Фитохимия 17, 2005–2010. DOI: 10.1016 / S0031-9422 (00) 88752-X
CrossRef Полный текст | Google Scholar
Остин М. Б., Боуман М. Э., Феррер Дж. Л., Шредер Дж. И Ноэль Дж. П. (2004). Переключатель альдола, обнаруженный в стильбенсинтазах, опосредует специфичность циклизации поликетидсинтаз III типа. Chem. Биол. 11, 1179–1194. DOI: 10.1016 / j.chembiol.2004.05.024
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бан, З. Н., Цинь, Х., Митчелл, А. Дж., Лю, Б. X., Чжан, Ф. Х., Вен, Дж. К. и др. (2018). Некаталитические складчатые белки халкон-изомеразы в humulus lupulus являются вспомогательными компонентами в биосинтезе пренилированных флавоноидов. Proc. Natl. Акад. Sci. США . 115, E5223 – E5232. DOI: 10.1073 / pnas.1802223115
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бэррон, Д., и Ибрагим, Р. К. (1996). Изопренилированные флавоноиды — обзор. Фитохимия 43, 921–982. DOI: 10.1016 / S0031-9422 (96) 00344-5
CrossRef Полный текст | Google Scholar
Bowman, J. L., Kohchi, T., Yamato, K. T., Jenkins, J., Shu, S. Q., Ishizaki, K., et al. (2017). Анализ эволюции наземных растений на основе генома Marchantia polymorpha . Cell. 171, 287–304. DOI: 10.1016 / j.cell.2017.09.030
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бредебах, М., Матерн У., Мартенс С. (2011). Активность трех 2-оксоглутарат-зависимых диоксигеназ у equisetum arvense l. Образование флавона и флавонола из (2s) -нарингенина. Фитохимия 72, 557–563. DOI: 10.1016 / j.phytochem.2011.01.036
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Cao, Y. Y., Yang, J. F., Liu, T. Y., Su, Z. F., Zhu, F. Y., Chen, M. X., et al. (2017). Филогенетически обоснованное сравнение гидролаз gh2 между Arabidopsis и реакцией риса на стрессоры. Фронт. Plant Sci. 8: 350. DOI: 10.3389 / fpls.2017.00350
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Капути, Л., Малной, М., Горемыкин, В., Никифорова, С., Мартенс, С. (2012). Полногеномная филогенетическая реконструкция udp-гликозилтрансфераз семейства 1 выявила расширение семейства в процессе адаптации растений к жизни на суше. Plant J. 69, 1030–1042. DOI: 10.1111 / j.1365-313X.2011.04853.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кэррингтон, Ю., Guo, J., Le, C.H., Fillo, A., Kwon, J., Tran, L.T., et al. (2018). Эволюция вторичного метаболического пути от первичного метаболизма: биосинтез шикимата и хината в растениях. Завод J . 95, 823–833. DOI: 10.1111 / tpj.13990
CrossRef Полный текст | Google Scholar
Cheng, A.X., Zhang, X. B., Han, X.J., Zhang, Y.Y., Gao, S., Liu, C.J., et al. (2018). Идентификация халкон-изомеразы в базальных наземных растениях свидетельствует о древней эволюции ферментативной активности циклизации для синтеза флавоноидов. New Phytol. 217, 909–924. DOI: 10.1111 / nph.14852
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Клейтон, В. А., Альберт, Н. В., Тримавитана, А. Х., МакГи, Т. К., Деролес, С. К., Швинн, К. Е. и др. (2018). Uvr8-опосредованная индукция биосинтеза флавоноидов для устойчивости к uvb сохраняется между печеночником Marchantia polymorpha и цветковыми растениями. Plant J. 96, 503–517. DOI: 10.1111 / tpj.14044
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Колпитс, К.К., Ким, С. С., Посен, С. Е., Джепсон, К., Ким, С. Ю., Видеманн, Г. и др. (2011). Ppascl, мох-ортолог специфичных для пыльников халкон-синтазоподобных ферментов, представляет собой гидроксиалкилпирон-синтазу, участвующую в эволюционно законсервированном пути биосинтеза спорополленина. New Phytol. 192, 855–868. DOI: 10.1111 / j.1469-8137.2011.03858.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кук Д., Римандо А. М., Клементе Т. Э., Шредер Дж., Даян Ф. Э., Нанаяккара, Н. П. Д. и др. (2010). Алкилрезорцинолсинтазы, экспрессируемые в корневых волосках сорго двухцветного, играют важную роль в биосинтезе аллелопатического бензохинон-сорголеона. Растительная клетка . 22, 867–887. DOI: 10.1105 / tpc.109.072397
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дастмалчи, М., и Даубхадель, С. (2015). Изомераза халкона сои: эволюция складки, дифференциальная экспрессия и локализация семейства генов. Planta 241, 507–523. DOI: 10.1007 / s00425-014-2200-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дьюик, П. М. (1994). «Изофлавоноиды», в . Исследования флавоноидов развиваются с 1986 года. , изд. Дж. Б. Харборн (Chapman and Hall / CRC: United States of America, 117–238.
Google Scholar
Добрица, А. А., Лей, З. Т., Нишикава, С., Урбанчик-Вочняк, Э., Хухман, Д. В., Прейс, Д., и др. (2010). Lap5 и lap6 кодируют белки, специфичные для пыльников, сходные с халкон-синтазой, необходимые для развития экзины пыльцы у Arabidopsis . Plant Physiol. 153, 937–955. DOI: 10.1104 / стр.110.157446
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ду, Х., Ран, Ф., Донг, Х. Л., Вэнь, Дж., Ли, Дж. Н., и Лян, З. (2016). Полногеномный анализ, классификация, эволюция и анализ экспрессии семейства цитохрома p450 93 в наземных растениях. PLoS ONE 11: e0165020. DOI: 10.1371 / journal.pone.0165020
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фэрроу, С.К. и Факкини П. Дж. (2014). Функциональное разнообразие 2-оксоглутарат / fe (ii) -зависимых диоксигеназ в метаболизме растений. Фронт. Plant Sci. 5: 524. DOI: 10.3389 / fpls.2014.00524
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Феррер, Дж. Л., Джез, Дж. М., Боуман, М. Э., Диксон, Р. А., и Ноэль, Дж. П. (1999). Строение халконсинтазы и молекулярные основы биосинтеза поликетидов растений. Nat. Struct. Биол. 6, 775–784. DOI: 10.1038 / 11553
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Банда, Д.R., Kasahara, H., Xia, Z.Q., Vander Mijnsbrugge, K., Bauw, G., Boerjan, W., et al. (1999). Эволюция защитных механизмов растений. Отношения фенилкумаранбензиловых эфирредуктаз с пинорезинол-ларицирезинолом и изофлавонредуктазами. J. Biol. Chem . 274, 7516–7527. DOI: 10.1074 / jbc.274.11.7516
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гебхардт Ю., Витте С., Форкманн Г., Лукацин Р., Матерн У. и Мартенс С. (2005). Молекулярная эволюция флавоноиддиоксигеназ в семействе apiaceae. Фитохимия 66, 1273–1284. DOI: 10.1016 / j.phytochem.2005.03.030
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гебхардт, Ю. Х., Витте, С., Стеубер, Х., Матерн, У., и Мартенс, С. (2007). Эволюция флавон-синтазы i из флаванон-3бета-гидроксилазы петрушки путем сайт-направленного мутагенеза. Plant Physiol. 144, 1442–1454. DOI: 10.1104 / стр.107.098392
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гиффорд, И., Баттенберг, К., Вания, А., Уилсон, А., Тиан, Л., Файн, О. и др. (2018). Особенности биосинтеза флавоноидов в корнях и клубеньках Datisca glomerata и medicago spp. Выявлено метаболомным профилем и профилями экспрессии генов. Фронт. Plant Sci. 9: 1463. DOI: 10.3389 / fpls.2018.01463
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Goiris, K., Muylaert, K., Voorspoels, S., Noten, B., De Paepe, D., Baart, G.J.E., и другие. (2014). Обнаружение флавоноидов в микроводорослях разных эволюционных линий. Дж. Фикол . 50, 483–492. DOI: 10.1111 / JPY.12180
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гулд, К. С., и Листер, К. (2006). «Функции флавоноидов в растениях», в Flavonoids: Chemistry, Biochemistry and Applications , eds Ø. М. Андерсон и К. Р. Маркхэм (Бока-Ратон: CRC Press), 397–442.
Google Scholar
Хан, X.J., Wu, Y. F., Gao, S., Yu, H. N., Xu, R. X., Lou, H. X. и др. (2014). Функциональная характеристика флавон-синтазы plagiochasma appendiculatum i, демонстрирующая активность флаванон-2-гидроксилазы. FEBS Lett. 588, 2307–2314. DOI: 10.1016 / j.febslet.2014.05.023
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Харборн, Дж. Б. (1988). Флавоноиды. Достижения в исследованиях с 1980 г. . Бостон, Массачусетс: Springer США.
Google Scholar
Герр К.Q., и Хаузингер Р. П. (2018). Поразительное разнообразие биохимических ролей fe (ii) / 2-оксоглутарат оксигеназ. Trends Biochem. Sci . 43, 517–532. DOI: 10.1016 / j.tibs.2018.04.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Hieta, R., and Myllyharju, J. (2002). Клонирование и характеристика низкомолекулярной пролил-4-гидроксилазы из Arabidopsis thaliana . Эффективное гидроксилирование богатых пролином, коллагеноподобных и индуцируемых гипоксией пептидов фактора транскрипции альфа-подобных пептидов. J. Biol. Chem . 277, 23965–23971. DOI: 10.1074 / jbc.M201865200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ислам, М. С., Лейссинг, Т. М., Чоудхури, Р., Хопкинсон, Р. Дж., И Шофилд, К. Дж. (2018). 2-оксоглутарат-зависимые оксигеназы. Annu. Rev. Biochem. 87, 585–620. DOI: 10.1146 / annurev-biochem-061516-044724
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джез, Дж. М., Боуман, М. Э., Диксон, Р.А. и Ноэль, Дж. П. (2000). Структура и механизм эволюционно уникального растительного фермента халкон-изомеразы. Nat. Struct. Биол . 7, 786–791. DOI: 10.1038 / 79025
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цзян, К. Г., Ким, С. Ю. и Сух, Д. Ю. (2008). Дивергентная эволюция надсемейства тиолаз и семейства халконсинтаз. Мол. Филоген. Evol . 49, 691–701. DOI: 10.1016 / j.ympev.2008.09.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цзян, К.Г., Шоммер, К. К., Ким, С. Ю., Сух, Д. Ю. (2006). Клонирование и характеристика халкон-синтазы из мха Physcomitrella patens. Фитохимия 67, 2531–2540. DOI: 10.1016 / j.phytochem.2006.09.030
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цзян, В. Б., Инь, К. Г., Ву, Р. Р., Чжэн, Г. С., Лю, Дж. Ю., Диксон, Р. А., и др. (2015). Роль халкон-изомеразоподобного белка в биосинтезе флавоноидов у Arabidopsis thaliana . J. Exp. Бот . 66, 7165–7179. DOI: 10.1093 / jxb / erv413
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Йоргенсен К., Расмуссен А. В., Морант М., Нильсен А. Х., Бьярнхольт Н., Загробельный М. и др. (2005). Образование метаболонов и метаболические каналы в биосинтезе натуральных растительных продуктов. Curr. Opin. Завод Биол . 8, 280–291. DOI: 10.1016 / j.pbi.2005.03.014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Йорнвалль, Х., Хуг, Дж. О., и Перссон, Б. (1999). Sdr и mdr: завершенные последовательности генома показывают, что эти семейства белков большие, старого происхождения и сложной природы. FEBS Lett . 445, 261–264. DOI: 10.1016 / S0014-5793 (99) 00130-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Каллберг, Ю., Опперманн, У., Перссон, Б. (2010). Классификация суперсемейства короткоцепочечных дегидрогеназ / редуктаз с использованием скрытых марковских моделей. FEBS J . 277, 2375–2386.DOI: 10.1111 / j.1742-4658.2010.07656.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кальтенбах, М., Берк, Дж. Р., Диндо, М., Пабис, А., Мансберг, Ф. С., Рабин, А. и др. (2018). Эволюция халкон-изомеразы от некаталитического предка. Nat. Chem. Биол. 14, 548–555. DOI: 10.1038 / s41589-018-0042-3
CrossRef Полный текст | Google Scholar
Кавана, К. Л., Йорнвалл, Х., Перссон, Б., и Опперманн, У. (2008). Семейства генов и белков средне- и короткоцепочечной дегидрогеназы / редуктазы: суперсемейство sdr: функциональное и структурное разнообразие в семействе метаболических и регуляторных ферментов. Cell. Мол. Наука о жизни . 65, 3895–3906. DOI: 10.1007 / s00018-008-8588-y
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Каваи Ю., Оно Э. и Мизутани М. (2014). Эволюция и разнообразие суперсемейства 2-оксоглутарат-зависимых диоксигеназ у растений. Завод J . 78, 328–343. DOI: 10.1111 / tpj.12479
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ким, С.С., Гриненбергер, Э., Лаллеманд, Б., Колпиттс, К.К., Ким, С. Ю., Соуза, К. Д. и др. (2010). Lap6 / поликетид-синтаза a и lap5 / поликетид-синтаза b кодируют гидроксиалкил-альфа-пирон-синтазы, необходимые для развития пыльцы и биосинтеза спорополленина в Arabidopsis thaliana . Растительная клетка 22, 4045–4066. DOI: 10.1105 / tpc.110.080028
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Коэдука Т., Луи Г. В., Орлова И., Киш, К. М., Ибдах, М., Вилкерсон, К. Г. и др. (2008). Множественные фенилпропенсинтазы как в clarkia breweri, так и в petunia hybrida представляют собой две отдельные белковые линии. Завод J . 54, 362–374. DOI: 10.1111 / j.1365-313X.2008.03412.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кубо, Х., Нозава, С., Хиваташи, Т., Кондо, Ю., Накабаяси, Р., Мори, Т., и др. (2018). Биосинтез риккионидинов и марчантинов регулируется факторами транскрипции r2r3-myb в Marchantia polymorpha . J. Завод Res . 131, 849–864. DOI: 10.1007 / s10265-018-1044-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лам, П.Ю., Лю, Х., и Ло, К. (2015). Завершение пути биосинтеза трицина в рисе: цитохром p450 75b4 — уникальная хризоэриол-5′-гидроксилаза. Физиология растений . 168, 1527–1536. DOI: 10.1104 / стр.15.00566
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лам П. Ю., Тобимацу Ю., Такеда Ю., Судзуки С., Ямамура М., Умедзава Т. и др. (2017). Нарушение флавон-синтазы II изменяет лигнин и улучшает усвояемость биомассы. Физиология растений .174, 972–985. DOI: 10.1104 / стр. 16.01973
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Lan, W., Lu, F.C., Regner, M., Zhu, Y.M., Rencoret, J., Ralph, S.A., et al. (2015). Трицин, флавоноидный мономер при лигнификации однодольных растений. Физиология растений . 167, 1284–1295. DOI: 10.1104 / стр.114.253757
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Lan, W., Morreel, K., Lu, F. C., Rencoret, J., del Rio, J. C., Voorend, W., et al.(2016a). Метаболиты трицин-олиголигнола кукурузы и их значение для лигнификации однодольных растений. Физиология растений . 171, 810–820. DOI: 10.1104 / стр. 16.02012
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лан, В., Ренкорет, Дж., Лу, Ф. К., Карлен, С. Д., Смит, Б. Г., Харрис, П. Дж. И др. (2016b). Трицин-лигнины: наличие и количественное определение трицина в отношении филогении. Завод J . 88, 1046–1057. DOI: 10.1111 / tpj.13315
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лапчик, О.(2007). Изофлавоноиды в небобовых таксонах: редкость или правило? Фитохимия 68, 2909–2916. DOI: 10.1016 / j.phytochem.2007.08.006
CrossRef Полный текст | Google Scholar
Ли, Л., Аслам, М., Раввин, Ф., Вандервель, М. К., Эштон, Н. В., и Сух, Д. Ю. (2018). Ppors, древняя поликетидсинтаза III типа, необходима для целостности кутикулы листа и устойчивости к обезвоживанию мха Physcomitrella patens. Planta 247, 527–541. DOI: 10.1007 / s00425-017-2806-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Линдаль, Т., Седжвик, Б., Секигучи, М., и Накабеппу, Ю. (1988). Регуляция и выражение адаптивного ответа на алкилирующие агенты. Annu. Ред. Biochem . 57, 133–157. DOI: 10.1146 / annurev.bi.57.070188.001025
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лиу, Г., Чан, Ю. К., Ван, Ю., и Вен, Дж. К. (2018). Механистические основы эволюции каталитической активности цистеина халконсинтазы у наземных растений. J. Biol. Chem . 293, 18601–18612.DOI: 10.1074 / jbc.RA118.005695
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Маркхэм, К. Р., Райан, К. Г., Блур, С., и Митчелл, К. А. (1998). Увеличение соотношения лютеолин: апигенин в Marchantia polymorpha при усилении УФ-В. Фитохимия 48, 791–794. DOI: 10.1016 / S0031-9422 (97) 00875-3
CrossRef Полный текст | Google Scholar
Марколович, С., Уилкинс, С. Э., Скофилд, К. Дж. (2015). Гидроксилирование белков, катализируемое 2-оксоглутарат-зависимыми оксигеназами. J. Biol. Chem . 290, 20712–20722. DOI: 10.1074 / jbc.R115.662627
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мартенс, С., Форкманн, Г., Брич, Л., Веллманн, Ф., Матерн, У. и Лукацин, Р. (2003). Дивергентная эволюция флавоноид-2-оксоглутарат-зависимых диоксигеназ в петрушке. FEBS Lett . 544, 93–98. DOI: 10.1016 / S0014-5793 (03) 00479-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мацудзава, М., Кацуяма, Ю., Фуна, Н., Хориноути, С. (2010). Синтез алкилрезорциловой кислоты поликетидсинтазами III типа из риса Oryza sativa . Фитохимия 71, 1059–1067. DOI: 10.1016 / j.phytochem.2010.02.012
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Meza, T. J., Moen, M. N., Vagbo, C. B., Krokan, H. E., Klungland, A., Grini, P. E., et al. (2012). ДНК-диоксигеназа alkbh3 защищает Arabidopsis thaliana от повреждения метилированием. Нуклеиновые Кислоты Res . 40, 6620–6631. DOI: 10.1093 / nar / gks327
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Mielecki, D., Zugaj, D. L., Muszewska, A., Piwowarski, J., Chojnacka, A., Mielecki, M., et al. (2012). Новые алкб-диоксигеназы — альтернативные модели для исследований in silico и in vivo . PLoS ONE 7: e30588. DOI: 10.1371 / journal.pone.0030588
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мин, Т., Касахара, Х., Бедгар, Д. Л., Юн, Б., Лоуренс, П. К., Ганг, Д. Р. и др. (2003). Кристаллические структуры пинорезинол-ларицирезинола и фенилкумаран бензиловых эфирредуктаз и их связь с изофлавонредуктазами. J. Biol. Chem . 278, 50714–50723. DOI: 10.1074 / jbc.M3084
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Моге, Г. Д., Ласт, Р. Л. (2015). Что-то старое, что-то новое: консервированные ферменты и эволюция новизны в специализированном метаболизме растений. Физиология растений . 169, 1512–1523. DOI: 10.1104 / стр.15.00994
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Морита, Х., Ямасита, М., Ши, С. П., Вакимото, Т., Кондо, С., Като, Р. и др. (2011). Синтез ненатуральных алкалоидных каркасов с использованием поликетидсинтазы растений. Proc. Natl. Акад. Sci. США . 108, 13504–13509. DOI: 10.1073 / pnas.1107782108
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Морита, Ю., Такаги, К., Фукути-Мизутани, М., Исигуро, К., Танака, Ю., Нитасака, Э. и др. (2014). Подобный халкон-изомеразе протеин усиливает производство флавоноидов и пигментацию цветов. Завод J . 78, 294–304. DOI: 10.1111 / tpj.12469
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мумму, Х., Каллберг, Ю., Тонфак, Л. Б., Перссон, Б., и ван дер Рест, Б. (2012). Надсемейство короткоцепочечных дегидрогеназ (sdr) растений: инвентаризация всего генома и модели диверсификации. БМК Завод Биол . 12: 219. DOI: 10.1186 / 1471-2229-12-219
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Нельсон Д. Р. (2006). Посадите цитохром p450s от мха до тополя. Phytochem. Ред. . 5, 193–204. DOI: 10.1007 / s11101-006-9015-3
CrossRef Полный текст | Google Scholar
Нгаки, М. Н., Луи, Г. В., Филипп, Р. Н., Мэннинг, Г., Пойер, Ф., Боуман, М. Е. и др. (2012). Эволюция халкон-изомеразной складки от связывания жирных кислот до стереоспецифического катализа. Природа 485, 530–533. DOI: 10.1038 / nature11009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Оуэнс, Д. К., Алердинг, А. Б., Кросби, К. К., Бандара, А. Б., Вествуд, Дж. Х. и Винкель, Б. С. (2008a). Функциональный анализ предсказанного семейства генов флавонолсинтазы в Arabidopsis . Plant Physiol. 147, 1046–1061. DOI: 10.1104 / стр.108.117457
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Оуэнс, Д.К., Кросби, К. С., Рунак, Дж., Ховард, Б. А., и Винкель, Б. С. (2008b). Биохимическая и генетическая характеристика флаванон-3-гидроксилазы Arabidopsis . Plant Physiol. Биохим . 46, 833–843. DOI: 10.1016 / j.plaphy.2008.06.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пичерский, Э., и Банда, Д. Р. (2000). Генетика и биохимия вторичных метаболитов растений: эволюционная перспектива. Завод Тенденции .5, 439–445. DOI: 10.1016 / S1360-1385 (00) 01741-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пройс, А., Штракке, Р., Вайсхаар, Б., Хиллебрехт, А., Матерн, У., и Мартенс, С. (2009). Arabidopsis thaliana экспрессирует вторую функциональную флавонолсинтазу. FEBS Lett . 583, 1981–1986. DOI: 10.1016 / j.febslet.2009.05.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Раушер, М. Д. (2006). «Эволюция флавоноидов и их генов», в The Science of Flavonoids , ed E.Гротевольд (Нью-Йорк, Нью-Йорк: Спрингер, 175–211.
Google Scholar
Раушер М. Д., Миллер Р. Э. и Тиффин П. (1999). Паттерны изменения скорости эволюции генов пути биосинтеза антоцианов. Мол. Биол. Evol . 16, 266–274. DOI: 10.1093 / oxfordjournals.molbev.a026108
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Раутер, А. П., Эннис, М., Хеллвич, К. Х., Герольд, Б. Дж., Хортон, Д., Мосс, Г. П. и др. (2018).Номенклатура флавоноидов (рекомендации IUPAC 2017). Чистое приложение химии . 90, 1429–1486. DOI: 10.1515 / pac-2013-0919
CrossRef Полный текст | Google Scholar
Сайто К., Йонекура-Сакакибара К., Накабаяши Р., Хигаши Ю., Ямазаки М., Тохге Т. и др. (2013). Путь биосинтеза флавоноидов у арабидопсиса: структурное и генетическое разнообразие. Plant Physiol. Биохим . 72, 21–34. DOI: 10.1016 / j.plaphy.2013.02.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Щуц, Р., Heller, W., и Hahlbrock, K. (1983). Субстратная специфичность халконсинтазы петрозелина гортензия. Образование производных флороглюцина из алифатических субстратов. J. Biol. Chem . 258, 6730–6734.
PubMed Аннотация | Google Scholar
Зейтц, К., Эдер, К., Деймл, Б., Келлнер, С., Мартенс, С., и Форкманн, Г. (2006). Клонирование, функциональная идентификация и анализ последовательности флавоноид-3′-гидроксилазы и флавоноид-3 ‘, 5′-гидроксилазы cdnas выявили независимую эволюцию флавоноид-3′, 5’-гидроксилазы в семействе Asteraceae . Завод Мол. Биол . 61, 365–381. DOI: 10.1007 / s11103-006-0012-0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шимада, Н., Аоки, Т., Сато, С., Накамура, Ю., Табата, С., и Аябе, С. (2003). Кластер генов кодирует два типа халкон-изомеразы, участвующей в биосинтезе общих флавоноидов и специфичных для бобовых 5-дезокси (изо) флавоноидов у лотоса японского. Физиология растений . 131, 941–951. DOI: 10.1104 / pp.004820
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Симидзу, Ю., Огата, Х., Гото, С. (2017). Поликетидсинтазы III типа: функциональная классификация и филогеномика. ХимБиоХим . 18, 50–65. DOI: 10.1002 / cbic.201600522
CrossRef Полный текст | Google Scholar
Ширли, Б. У. (1996). Биосинтез флавоноидов: «новые» функции для «старого» пути. Завод Тенденции . 1, 377–382. DOI: 10.1016 / 1360-1385 (96) 10040-6
CrossRef Полный текст | Google Scholar
Сен-Пьер, Б., и Де Лука, В.(2000). «Эволюция генов ацилтрансфераз: происхождение и диверсификация суперсемейства bahd ацилтрансфераз, участвующих во вторичном метаболизме», в журнале «Последние достижения в фитохимии» , ред. JT Romeo, R. Ibrahim, L. Varin и V. De Luca (Амстердам: Elsevier Б.В.), 285–315.
Google Scholar
Самнер, Л. В., Амберг, А., Баррет, Д., Бил, М. Х., Бегер, Р., Дайкин, К. А. и др. (2007). Предлагаемые минимальные стандарты отчетности для химического анализа. Метаболомика 3, 211–221.DOI: 10.1007 / s11306-007-0082-2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тохге Т., Ватанабе М., Хёфген Р. и Ферни А. Р. (2013). Эволюция метаболизма фенилпропаноидов в зеленой линии. Crit. Rev. Biochem. Мол. Биол . 48, 123–152. DOI: 10.3109 / 10409238.2012.758083
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тонфак, Л. Б., Мумму, Х., Латче, А., Йомби, Э., Бенишу, М., Печ, Ж.-К., и другие. (2011). Надсемейство растений sdr: участие в первичном и вторичном метаболизме. Curr. Темы Завод Биол . 12, 41–53. Доступно в Интернете по адресу: https://prodinra.inra.fr/record/438703
Google Scholar
Цугава, Х., Кинд, Т., Накабаяси, Р., Юкихира, Д., Танака, В., Цайка, Т. и др. (2016). Правила перегруппировки водорода: вычислительная фрагментация ms / ms и выяснение структуры с использованием программного обеспечения ms-finder. Анал. Chem . 88, 7946–7958. DOI: 10.1021 / acs.analchem.6b00770
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цугава, Х., Накабаяси, Р., Мори, Т., Ямада, И, Такахаши, М., Рай, А. и др. (2019). Подход хеминформатики для характеристики метаболомов в организмах, меченных стабильными изотопами. Nat. Методы . 16, 295–298. DOI: 10.1038 / s41592-019-0358-2
CrossRef Полный текст | Google Scholar
Туоминен, Л. К., Джонсон, В. Э., Цай, К. Дж. (2011). Дифференциальное филогенетическое расширение ацилтрансфераз bahd по пяти таксонам покрытосеменных и свидетельство дивергентной экспрессии среди паралогов populus. BMC Genomics 12: 236. DOI: 10.1186 / 1471-2164-12-236
PubMed Аннотация | CrossRef Полный текст | Google Scholar
ван ден Борн, Э., Омельченко, М. В., Беккелунд, А., Лейне, В., Кунин, Э. В., Доля, В. В. и др. (2008). Вирусные белки алкб восстанавливают повреждение РНК путем окислительного деметилирования. Нуклеиновые Кислоты Res . 36, 5451–5461. DOI: 10.1093 / nar / gkn519
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ван П., Чжан Л., Jiang, X., Dai, X., Xu, L., Li, T., et al. (2018). Эволюционная и функциональная характеристика лейкоантоцианидинредуктаз из Camellia sinensis . Planta 247, 139–154. DOI: 10.1007 / s00425-017-2771-z
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ван, X., Хе, X., Лин, Дж., Шао, Х., Чанг, З., и Диксон, Р. А. (2006). Кристаллическая структура изофлавонредуктазы люцерны ( Medicago sativa л.). J. Mol. Биол. 358, 1341–1352. DOI: 10.1016 / j.jmb.2006.03.022
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ван, Ю., Тан, X., Патерсон, А. Х. (2013). Различные паттерны дивергенции генной структуры после дупликации гена в Arabidopsis . BMC Genomics 14: 652. DOI: 10.1186 / 1471-2164-14-652
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ванибучи, К., Чжан, П., Абэ, Т., Морита, Х., Коно, Т., Чен, Г.S., et al. (2007). Новая многофункциональная поликетидсинтаза, продуцирующая акридон, из huperzia serrata. FEBS J . 274, 1073–1082. DOI: 10.1111 / j.1742-4658.2007.05656.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вен Дж. К., Ли, X., Стаут, Дж., И Чаппл, К. (2008). Независимое происхождение сирингиллигнина в сосудистых растениях. Proc. Natl. Акад. Sci. США . 105, 7887–7892. DOI: 10.1073 / pnas.08016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вен, Дж.К., Ноэль, Дж. П. (2012a). Замечательная податливость и неразборчивость специализированного метаболизма. Cold Spring Harb. Symp. Quant. Биол . 77, 309–320. DOI: 10.1101 / sqb.2012.77.014787
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вен, Дж. К., Ноэль, Дж. П. (2013). Химическое разнообразие в Selaginella : справочная система для параллельной и конвергентной метаболической эволюции у наземных растений. Фасадный завод Sci . 4: 119. DOI: 10.3389 / fpls.2013.00119
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вен, Дж. К., Е, М., Ли, Б., и Ноэль, Дж. П. (2016). Совместная эволюция метаболизма гормонов и сигнальных сетей увеличивает адаптивную пластичность растений. Ячейка 166, 881–893. DOI: 10.1016 / j.cell.2016.06.027
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Винкель-Ширли, Б. (2001). Биосинтез флавоноидов. Красочная модель для генетики, биохимии, клеточной биологии и биотехнологии. Физиология растений . 126, 485–493. DOI: 10.1104 / стр. 126.2.485
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вольф, Л., Риццини, Л., Штраке, Р., Ульм, Р., и Ренсинг, С. А. (2010). Молекулярные и физиологические реакции Physcomitrella patens на ультрафиолетовое b-излучение. Физиология растений . 153, 1123–1134. DOI: 10.1104 / стр.110.154658
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ву, Л. Ф., Мэн, С., и Тан, Г.Л. (2016). Двухвалентное железо и альфа-кетоглутарат-зависимые диоксигеназы в биосинтезе микробных природных продуктов. Biochim. Биофиз. Acta . 1864, 453–470. DOI: 10.1016 / j.bbapap.2016.01.012
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Язаки К., Сасаки К. и Цурумару Ю. (2009). Пренилирование ароматических соединений — ключевая диверсификация вторичных метаболитов растений. Фитохимия 70, 1739–1745. DOI: 10.1016 / j.phytochem.2009.08.023
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ёнекура-Сакакибара, К., и Ханада, К. (2011). Эволюционный взгляд на функциональное разнообразие гликозилтрансфераз семейства 1. Plant J. 66, 182–193. DOI: 10.1111 / j.1365-313X.2011.04493.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Yu, C. K. Y., Springob, K., Schmidt, J. R., Nicholson, R. L., Chu, I. K., Yip, W. K., et al. (2005). Ген стильбенсинтазы (sbsts1) участвует в защитных реакциях хозяина и не-хозяина у сорго. Физиология растений . 138, 393–401. DOI: 10.1104 / стр.105.059337
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Yu, H. N., Liu, X.Y., Gao, S., Sun, B., Zheng, H. B., Ji, M., et al. (2018). Структурно-биохимическая характеристика поликетидсинтаз III типа растений печеночника Marchantia paleacea. Plant Physiol. Биохим . 125, 95–105. DOI: 10.1016 / j.plaphy.2018.01.030
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ю., X.Х., Гоу, Дж. Й., и Лю, К. Дж. (2009). Bahd суперсемейство ацил-коа-зависимых ацилтрансфераз у populus и арабидопсиса: биоинформатика и экспрессия генов. Завод Мол. Биол . 70, 421–442. DOI: 10.1007 / s11103-009-9482-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Юань, Т., Фудзиока, С., Такацуто, С., Мацумото, С., Гоу, X., Хе, К. и др. (2007). Ben1, ген, кодирующий дигидрофлавонол-4-редуктазу (dfr) -подобный белок, регулирует уровни брассиностероидов в Arabidopsis thaliana . Завод J . 51, 220–233. DOI: 10.1111 / j.1365-313X.2007.03129.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Влияние циркадного ритма на метаболические процессы и регуляцию энергетического баланса — FullText — Annals of Nutrition and Metabolism 2019, Vol. 74, № 4
Абстрактные
Предпосылки: Система циркадного времени или циркадные часы играет решающую роль во многих биологических процессах, таких как цикл сна-бодрствования, секреция гормонов, здоровье сердечно-сосудистой системы, гомеостаз глюкозы и регулирование температуры тела.Энергетический баланс также является одним из важнейших краеугольных камней метаболических процессов, тогда как энергетический дисбаланс связан со многими заболеваниями (например, ожирением, диабетом, сердечно-сосудистыми заболеваниями). Циркадные часы являются основным регулятором метаболизма, и этот анализ дает обзор двунаправленного влияния циркадного ритма на метаболические процессы и энергетический баланс. Резюме: Циркадная система времени или циркадные часы играют решающую роль во многих биологических процессах, но увеличение активности, которая работает круглосуточно и без выходных, и обычное использование телевидения, Интернета и мобильных телефонов почти 24 часа в сутки приводит к тому, что постепенное уменьшение адекватного времени сна.Согласно недавним исследованиям, долгосрочные нарушения циркадного ритма связаны со многими патологическими состояниями, такими как преждевременная смертность, ожирение, нарушение толерантности к глюкозе, диабет, психические расстройства, тревога, депрессия и прогрессирование рака, тогда как краткосрочные нарушения связаны с ухудшением самочувствия. , утомляемость и потеря концентрации. В этом обзоре были изучены циркадные ритмы метаболических процессов и их влияние на энергетический баланс. Ключевые сообщения: Циркадный ритм имеет двунаправленное взаимодействие почти со всеми метаболическими процессами.Следовательно, понимание основной причины, влияющей на циркадные часы, и создание рекомендаций по лечению с использованием циркадного ритма может повысить эффективность лечения заболевания. Хронофармакология, хроническое питание и хроноупражнения — это новые подходы к лечению метаболического баланса.
© 2019 S. Karger AG, Базель
Введение
Термин «циркадный» образован от двух латинских слов: circa (приблизительно) и dies (день), обозначающих приблизительно «один день» [1].Циркадный ритм также называется биологическими / циркадными часами [2] и относится к поведенческим, физиологическим и молекулярным изменениям с продолжительностью цикла около 24 часов [3]. Циркадные часы можно разделить на 2 части: центральные часы, расположенные в супрахиазматическом ядре (SCN) гипоталамуса, которое принимает световые сигналы, и периферические часы, находящиеся в различных тканях по всему телу. Периферические часы играют неотъемлемую и уникальную роль в каждой из своих тканей, управляя циркадной экспрессией конкретных генов, участвующих в различных физиологических функциях [4].
Основным стимулом для SCN является свет [5]. Однако у слепых людей есть циркадные циклы (например, циклы сна и бодрствования), и этот цикл длится более 24 часов. Это открытие привело к идее, что другие стимулы, помимо света, могут действовать как стимул для биологических часов человека [6]. Обзорное исследование показало, что температура, гормоны, питательные вещества, распределение питательных веществ, некоторые питательные вещества (только, например, глюкоза, аминокислоты, этанол и ретиноевая кислота), состояние кормления / голодания, состояние сна-бодрствования, физическая активность являются эффективными стимулами для циркадный цикл в различных периферических путях [5].Интересно, что сообщалось, что наблюдались различия во влиянии циркадного ритма у мужчин по сравнению с женщинами [7].
В основном цикл сна и бодрствования регулирует циркадный ритм. Однако в современном мире рост активности, которая осуществляется круглосуточно и без выходных, и обычное использование телевидения, Интернета и мобильных телефонов почти 24 часа в сутки, приводит к постепенному сокращению необходимого времени для сна [8]. Эпидемиологическое исследование показало, что продолжительность ночного сна уменьшилась на 18 минут за последние 30 лет [9].Ухудшение цикла сна-бодрствования, особенно у здоровых людей, может быть основной причиной многих заболеваний, таких как преждевременная смертность, ожирение, нарушение толерантности к глюкозе, диабет, психические расстройства, тревога, депрессия и прогрессирование рака, усталость и потеря. концентрации [10, 11]. Другое эпидемиологическое исследование показало, что работа по крайней мере 3 ночи в месяц в течение 15 и более лет может увеличить риск рака прямой кишки у женщин [12]. У крыс нарушение циркадных обмороков ускоряет развитие диабета, вызывая функцию и потерю массы бета-клеток [13].В экспериментальном исследовании нарушение циркадного ритма приводило к поражению сердечно-сосудистой системы и почек у хомяков [14]. В исследовании, проведенном с пациентами с шизофренией, участники имели тяжелые циркадные расстройства сна и бодрствования, хотя их настроение, психический статус и психотические эпизоды были стабильными [15]. В этом обзоре была изучена роль циркадного ритма в метаболических процессах и двунаправленное влияние циркадного ритма на энергетический баланс. В соответствии с этой целью были проанализированы исследования, опубликованные с 2000 по 2018 год.Кроме того, были добавлены 3 важные статьи, опубликованные в 1984, 1993 и 1996 годах. В основном для сканирования научных статей использовались базы данных Google Academic (библиографическая база данных), PubMed, Scopus, Web of Science и Science Direct. Кроме того, были просканированы ресурсы Центральной библиотеки Университета Гази. Такие термины, как «циркадные часы или циркадный ритм» или «биологические часы» и «гормоны или метаболизм, или энергетический баланс, или расход энергии, или термогенез, метаболический гомеостаз или метаболическая регуляция, или гены часов, или диетический состав, режим питания или время приема пищи или физический активность, упражнения или спорт » были использованы в качестве ключевых слов.В результате сканирования было найдено 4 456 статей. После проверки заголовков и аннотаций были исключены повторяющиеся статьи и статьи, опубликованные не на английском языке. Окончательное количество статей составило 82, включая статьи, которые были бесплатными и доступны в виде полных текстов. Кроме того, главы книги также использовались для изучения предмета.
Циркадная синхронизация в метаболическом гомеостазе
Многие исследования объясняют взаимосвязь между физиологией человека, некоторыми заболеваниями и циркадным ритмом [12-15].Метаболический гомеостаз — важный компонент, регулирующий энергетический обмен, особенно в жировой ткани. Жировая ткань — это центральный орган обмена веществ, который регулирует энергетический гомеостаз всего тела. Белая жировая ткань функционирует как ключевой резервуар энергии для других органов, тогда как коричневая жировая ткань накапливает липиды для индуцированного холодом адаптивного термогенеза. Жировая ткань секретирует различные гормоны, цитокины и метаболиты (называемые адипокинами), которые контролируют системный энергетический баланс, регулируя сигналы аппетита от центральной нервной системы, а также метаболическую активность в периферических тканях [16].Например, лептин имеет специфические рецепторы на гипоталамусе и высвобождается из основных адипоцитов. Этот гормон играет регулирующую роль в энергетическом обмене, увеличивая активацию симпатической нервной системы и увеличивая термогенез за счет увеличения гормонов щитовидной железы. В термогенезе белок UCP (разобщающий) подавляет синтез АТФ в митохондриях, позволяя потреблять энергию в виде тепла. Лептин увеличивает уровень гормонов щитовидной железы и активацию симпатической центральной нервной системы, что приводит к большему образованию UCP и, следовательно, большему потреблению энергии [17].Высвобождение гормона лептина происходит в циркадном цикле, а пик уровня лептина в сыворотке крови приходится на ночь [18]. Таким образом, нарушение циркадного баланса может косвенно влиять на секрецию лептина, термогенез и энергетический гомеостаз.
Напротив, некоторые гормоны, выделяемые гипоталамусом, проявляют большую активность ночью. Наиболее ярким примером этого является «гормон роста». Уровень гормона роста достигает пика между 2:00 и 4:00 утра.Поэтому необходимо уделять особое внимание режиму сна детей [19].
Кортизол — стероидный гормон, секретируемый надпочечниками. Он регулирует многие метаболические процессы, такие как гликогенолиз, липолиз и протеолиз [20]. Количество и частота секреции кортизола регулируются циркадным ритмом. Концентрация кортизола в кровообращении достигает пика перед пробуждением утром. Кортизол постепенно снижается в течение дня.Наименьшего уровня он достигает во время сна после полуночи [21]. Кортизол — главный гормон, регулирующий метаболические процессы в организме. Он увеличивает использование кортизола, глюкозы, свободных жирных кислот и аминокислот из эндогенных запасов топлива. Следовательно, высокий уровень кортизола действует как катаболический гормон, который снижает мышечную массу и мышечную массу, а также увеличивает потребление энергии [22]. Кроме того, толерантность к глюкозе и секреция инсулина меняются в течение дня. При естественном ходе метаболизма чувствительность к инсулину и секреция инсулина снижаются ночью (особенно между 3:00 и 5:00 a.м) по сравнению с утренними часами. Этот метаболический процесс, феномен рассвета, подчеркивает влияние контроля циркадных ритмов на метаболизм глюкозы [23]. В естественных физиологических процессах организма гормоны, действующие как антагонисты инсулина (особенно гормон роста), проявляют гиперинсулинемическую активность из-за снижения секреции инсулина между 3:00 и 5:00 утра, так что уровень сахара в крови возвращается к норме. Этому противодействует дополнительная физиологическая секреция инсулина у людей, не страдающих диабетом или инсулинозависимых.И наоборот, когда высвобождение инсулина нарушено, действие гормона роста, высвобождаемого в течение ночи, особенно у пациентов с диабетом, не может быть уменьшено. Это приводит к патологическому циркадному ритму, который может привести к утренней гипергликемии независимо от режима питания [24].
Мелатонин — важный гормон в циркадной синхронизации. Этот гормон участвует во многих биологических и физиологических регуляторах организма. Это эффективный гормон биоритма человека (циркадного ритма).Основная роль этого гормона — поддерживать биологические часы и регулировать ритм тела [25]. Синтез и высвобождение мелатонина стимулируются в темноте, ночью, тогда как днем он подавляется светом [1]. Особенно между 23:00. в 5:00 секреция мелатонина достигает пика, и его концентрация в крови увеличивается в 3–10 раз [25]. Однако воздействие света в ночное время вызывает снижение уровня мелатонина в плазме [1].
На метаболизм липидов также влияют циркадные колебания.Исследования показывают, что многие белки, связанные с метаболизмом липидов (например, ApoB, ApoA1 и ApoA4), белок кишечного микросомального транспорта триглицеридов и белок, связывающий кишечные жирные кислоты, демонстрируют изменения в течение дня [26–28]. Кроме того, исследования на мышах показывают, что абсорбция холестерина и липидов в темной фазе выше, чем в светлой фазе [28]. Некоторые продукты липидного обмена также имеют циркадный ритм. Например, циркулирующие неэтерифицированные жирные кислоты у людей выше ночью из-за повышенной липолитической активности [29].Кроме того, эпизоды инфаркта миокарда и астма связаны с циркадным циклом метаболизма. Пик этих приступов приходится на ночь или рано утром [30, 31]. В ранние утренние часы сердечно-сосудистая система улучшается в ответ на активацию активности симпатических нервов, а в вечерние часы — артериальное давление и пик пульса. Сердечные приступы, особенно рано утром или вечером, могут быть вызваны этим суточным ритмом сердечно-сосудистой системы.Суточный ритм гормонов и некоторые метаболические процессы показаны на рисунке 1.
Рис. 1.
Суточный ритм гормонов и некоторых метаболических процессов. Высвобождение гормона лептина происходит в циркадном цикле, а пик уровня лептина в сыворотке крови приходится на ночь. Уровень гормона роста достигает пика между 02:00 и 04:00 утра.Концентрация кортизола в кровообращении достигает пика перед пробуждением утром. Снижение секреции инсулина ночью (особенно с 03:00 до 05:00 a.м.). Синтез и высвобождение мелатонина стимулируются ночью в темноте, а днем он подавляется светом. Поглощение липидов в темной фазе выше, чем в светлой фазе. Пик инфаркта миокарда ночью или рано утром.
Метаболическая регуляция циркадных ритмов
У млекопитающих циркадные ритмы контролируются главным образом SCN, которые называются главными часами. Супрахиазматические ядра состоят из множества одноклеточных циркадных осцилляторов, расположенных в передней области гипоталамуса головного мозга и производящих согласованные циркадные сигналы при синхронизации [32].SCN активируется через нервный пучок, называемый «ретиногипоталамический тракт». Таким образом, SCN регулирует биологические часы у живых существ и способствует физиологическим процессам, стимулируя другие области мозга [2].
Супрахиазматический синапс ядра непосредственно с вентральной и дорсальной субвентрикулярной областями, клеточными телами, расположенными в вентральной и дорсальной субвентрикулярной областях, и дорсомедиальным гипоталамусом. Несмотря на то, что эти области взаимодействуют друг с другом, нейроны, расположенные в дорсальной наджелудочковой области, более эффективны в регуляции термогенеза, тогда как вентральная наджелудочковая область в основном играет роль в регуляции циклов сна-бодрствования и активности.Кроме того, паравентрикулярный гипоталамус отвечает за высвобождение кортикостероидов, тогда как боковой гипоталамус отвечает за питание и бдительность [33].
Хотя SCN функционирует как основные биологические часы метаболизма, исследования, проведенные в 2000-х годах, показали, что автономные циркадные осцилляторы, присутствующие в периферических органах и тканях, таких как печень, кишечник, сердце и сетчатка, способствуют метаболическим процессам через клеточные гены часов в этих органах / тканях [32, 34, 35].Супрахиазматические ядра играют важную роль в регуляции метаболизма глюкозы. Чувствительность к инсулину и поглощение глюкозы нарушены у крыс с повреждениями в их SCN [36]. Ухудшение циркадных часов может привести к нарушению секреции инсулина и гипоинсулинемии [29]. Связанные с циркадным ритмом белки CLOCK и BMAL1 участвуют в производстве и высвобождении инсулина, связываясь с регулирующими циркадный ритм дистальными областями β-клеток поджелудочной железы [37]. И диабет типа 1, и диабет 2 имеют недостаточность или отсутствие инсулина из-за повреждения β-клеток.Следовательно, не следует упускать из виду роль циркадного ритма в этиологии диабета с точки зрения повреждения β-клеток [38]. Гены часов и задачи, регулирующие циркадные ритмы у млекопитающих, сведены в Таблицу 1 [2].
Таблица 1.
Гены циркадного ритма и их роли [2]
У млекопитающих гомеостаз циркадных часов обеспечивается механизмами обратной связи (отрицательная) и прямой (положительная), влияющими на транскрипцию, трансляцию и посттрансляционный события [5, 29].Паттерн транскрипционной обратной связи опосредуется белками Cry1, Cry2, Per1 и Per2. Белки CLOCK и BMAL1 связываются с областью E-промотора генов Per и Cry и индуцируют их экспрессию. На более поздней стадии Per и Cry гетеродимеризуются и перемещаются из цитоплазмы в ядро, чтобы ингибировать CLOCK / BMAL1-индуцированную экспрессию генов [37, 39]. Этот механизм отрицательной обратной связи важен для эффективной работы циркадной системы.
Циркадный ритм и энергетический гомеостаз
Энергетический метаболизм регулируется многими гормонами, ферментами и транспортными системами, а циркадный ритм эффективно модулирует их экспрессию, секрецию и / или активацию [40].Метаболические пути, обеспечивающие энергетический гомеостаз, координируются метаболитами, которые предлагают изменения и активно подготавливают молекулярную среду, а также системы острой передачи сигналов, которые мгновенно реагируют на изменения циркадных часов [41].
Рецепторы гормонов, взаимодействия между генами и внутриклеточные реакции окисления / восстановления регулируют энергетический обмен на клеточном уровне. Эти пути имеют важное взаимодействие с часами биологического метаболизма.Циркадные часы могут влиять на функцию рецепторов гормонов (рецептор, активируемый пролифератором пероксисом [PPAR] α, PPARγ и REV-ERBα) и некоторых генов на клеточном уровне (сиртруин) [41, 42].
Рецепторы ядерных гормонов (PPARα, PPARγ, REV-ERBα, RORα, HNF4α, TRα и NURR1) и лиганды, присутствующие в метаболических тканях, действуют как сенсоры, объединяющие циркадные и метаболические пути [41]. Например, PPAR являются связующим звеном между циркадными часами и энергетическим метаболизмом. PPARγ локализуется в жировой ткани и активирует факторы транскрипции, которые увеличивают липогенез и накопление липидов.PPARα из ядерных рецепторов гормонов запускает кетогенез и окисление жирных кислот печени в ответ на голодание. PPARδ является наиболее распространенным рецептором ядерных клеток в организме и способен коррелировать ежедневные изменения температуры тела с циркадными часами [43].
Ритмическая экспрессия и активация метаболических путей в основном связаны с координацией часовых генов (BMAL1, Per2, Per1, Per3, Cry1 и Cry2) в печени и жировой ткани. Существует связь между белком BMAL1, липогенными путями и механизмами клеточных часов.REV-ERBα (репрессор транскрипции BMAL1) и RORα (положительный регулятор BMAL1) являются рецепторами ядерных гормонов, регулирующими липогенез. Более того, оба они модулируются CLOCK: BMAL1. PPARα играет роль в метаболизме липидов и липопротеинов. PPARα напрямую связывается с промоторной областью белка BMAL1, который регулирует экспрессию PPARα через гетеродимер CLOCK: BMAL1 [44].
Еще одним фактором, влияющим на функцию циркадного ритма, являются гены сиртуинов (SIRT). Гены семейства SIRT представляют собой NAD + -зависимые ферменты деацетилазы класса III, которые влияют на многие клеточные функции, включая метаболизм человека, старение, рак и клеточное старение.Существует 7 вариантов семейства SIRT (SIRT 1–7). Регуляция экспрессии генов — самый важный механизм, на который влияет семейство SIRT. SIRT1, SIRT6 и SIRT7 преимущественно локализуются в ядре клетки, SIRT2 — в цитоплазме, а SIRT3, SIRT4 и SIRT5 — в митохондриях. SIRT1 играет важную роль в регуляции метаболических процессов, таких как чувствительность к инсулину, метаболизм липидов и глюконеогенез, а также продолжительность жизни человека [45]. SIRT1 модулирует активность CLOCK: BMAL1 и участвует в циркадном цикле.Гены SIRT 3-5 регулируют внутриклеточные пути, такие как окисление жирных кислот, кетогенез, цикл мочевины и окислительное фосфорилирование [33].
Внутриклеточное окислительно-восстановительное (окислительно-восстановительное) состояние — важный фактор, регулирующий гены часов в периферических тканях. Белки CLOCK могут эффективно связываться только в присутствии восстановленных последовательностей NADH и NADPH, BMAL1 и E-box. Напротив, формы никотинамидадениндинуклеотидоксидазы (NAD + и NADP +) ингибируют связывание комплекса CLOCK: BMAL1 с ДНК [32].Таким образом, окислительно-восстановительный статус NAD / NADH клетки может приводить к изменениям циркадной фазы, влияя на транскрипционную активность генов BMAL1: CLOCK [46].
Внутриклеточные уровни НАД +, а также повышенные уровни АМФ (аденозинмонофосфата) являются индикаторами низкой энергии. Когда внутриклеточные уровни АТФ снижаются, AMPK (AMP-активирующая протеинкиназа) функционирует как пищевой сенсор и активирует внутриклеточные пути снабжения энергией. Таким образом, соотношение АМФ / АТФ также может быть связующим звеном между циркадным ритмом и энергетическим метаболизмом [47].
Влияние циркадного ритма на энергетический баланс
Энергия, производимая и сохраняемая в процессе метаболизма, используется для поддержания метаболической активности, такой как основной уровень метаболизма, физическая активность и тепловой эффект пищевых продуктов [48].
Фрэнсис Дж. Бенедикт впервые описал циркадные изменения в энергетическом обмене в 1915 году [49]. Кроме того, Haugen et al. [50] обнаружили, что скорость метаболизма в покое в полдень была на 6% выше, чем в утренние часы. Одним из наиболее важных факторов, влияющих на скорость основного обмена, является режим сна.Сон и циркадный ритм являются основными компонентами регуляции энергетического обмена [48, 51]. Есть 2 фазы сна: быстрое движение глаз (REM) и не-REM [52]. Активность симпатической нервной системы и сновидения усиливаются в период REM. Температура тела, частота сердечных сокращений, частота дыхания и артериальное давление повышаются во время фазы быстрого сна. Нарушения в периоде быстрого сна увеличиваются из-за активности симпатической нервной системы [53]. Из-за повышения температуры тела и расхода энергии в головном мозге (потребление энергии мозгом в этот период составляет примерно 25%), скорость метаболизма во время сна достигает наивысшей точки в фазе REM [54].Следовательно, ухудшение цикла сна из-за позднего сна, смены часовых поясов, сменной работы и т. Д. Может привести к снижению основной скорости метаболизма за счет изменения времени фазы REM сна.
Помимо основной скорости метаболизма / метаболизма в состоянии покоя, физическая активность является важным компонентом общих затрат энергии [55]. Однако исследования подчеркивают различные метаболические эффекты типа, продолжительности и продолжительности упражнений. Например, обзорное исследование подчеркивает, что максимальная производительность при краткосрочных анаэробных упражнениях достигается ближе к вечеру, обычно в полдень [56], что также соответствует пиковой температуре тела [56, 57].Температура тела рассматривается как «основная переменная» циркадного ритма и используется как маркер циркадного ритма [58]. Разница в температуре тела между утренними и вечерними часами составляет 0,9 ° C [59]. Температура тела влияет на мышечную активность [60]. Эта разница между утренними и вечерними часами может повлиять на выполнение упражнений и, косвенно, на скорость основного обмена.
Время приема пищи влияет на физическую работоспособность. Упражнения до и после еды по-разному влияют на окисление жирных кислот и метаболизм аппетита [61–63].Однако до сих пор неясно, когда (тренировка до еды / после еды) является наиболее эффективным временем для похудания. Подход к снижению массы тела и облегчению сжигания жира заключается в выполнении постпрандиальных аэробных упражнений после ночного голодания [64]. Упражнения во время голодания вызывают использование отложений гликогена для увеличения окисления жиров и снижает инсулин в плазме за счет увеличения уровней адреналина и норадреналина в плазме, вызывая липолиз. Напротив, некоторые исследования были посвящены положительному влиянию постпрандиальных упражнений на контроль массы тела по сравнению с упражнениями во время голодания из-за их положительного влияния на аппетит и метаболизм в покое [65].Исследование показало, что 36-минутные умеренные упражнения на беговой дорожке значительно увеличили скорость метаболизма в состоянии покоя через 40 минут после завтрака в средиземноморском стиле в первые 24 часа [66]. Во многих исследованиях общее потребление энергии, режимы питания, физические характеристики, продолжительность и тяжесть упражнений различаются, что приводит к разной интерпретации результатов.
Еще одна составляющая общих затрат энергии — тепловое воздействие питательных веществ. На тепловой эффект пищевых продуктов влияет макроэлементный состав рациона.Липиды обладают наименьшим (0–3%), а белки — наибольшим (20–30%) термогенным эффектом [67]. Кроме того, время приема пищи является важным фактором, влияющим на тепловой эффект продуктов. Термогенез, вызванный диетой, выше в утренние часы по сравнению с вечерними и ночными часами [68]. Аналогичным образом в исследовании Morris et al. [69], тепловой эффект от еды утром был на 44% выше, чем вечером. Снижение теплового эффекта пищи с утра до вечера может быть в первую очередь связано с влиянием эндокринной циркадной системы на физиологию желудочно-кишечного тракта.Периодичность кишечника в утренние часы более эффективна, чем в вечерние. Два исследования на здоровых взрослых показали, что скорость опорожнения желудка утром (8.00) была выше, чем вечером (20.00–11.00) [70]. Кроме того, нарушения циркадного ритма, такие как посменная работа и нарушение биоритмов, вызывают расстройства желудочно-кишечной системы, такие как боли в животе, вздутие живота, диарея или запор. Эти наблюдения показывают функциональную корреляцию между суточными ритмами и физиологией желудочно-кишечного тракта.Hoogerwerf et al. [71] показали, что экспрессия белков PER2 и BMAL1 была связана с циркадным ритмом в области мышечно-кишечного сплетения, который играет важную роль в координации эпителиальных клеток толстой кишки и моторики толстой кишки.
Влияние приема пищи и физической активности на циркадные ритмы
Здесь подчеркивается влияние циркадного ритма на компоненты расхода энергии и его роль в энергетическом балансе. Однако некоторые исследования показали, что некоторые факторы окружающей среды (т.е., пища, состав пищи, время кормления и упражнения) могут быть эффективны для периферических часов, которые существуют в нескольких частях тела, таких как печень, поджелудочная железа или сердце [72–75]. Периферические часы играют неотъемлемую и уникальную роль в каждой из своих тканей, управляя циркадной экспрессией определенных генов, участвующих в различных физиологических функциях. Существование всех этих часов, работающих вместе и синхронизируемых центральными часами, со многими гормонами и физиологическими переменными или переменными окружающей среды, изменяющимися в течение дня, делает это двунаправленное взаимодействие в циркадной системе довольно сложным [4].
Питание — один из внешних синхронизаторов наших периферийных часов. Основная роль циркадных часов — вовлекать организм в сигналы окружающей среды; это позволяет организмам прогнозировать наличие пищи. Ограничение доступа к пище определенным временем дня оказывает глубокое влияние на поведение и физиологию организмов [72]. Damiola et al. [76] показали, что временное ограничение питания в условиях свет-темнота или темнота-темнота может изменить фазу экспрессии циркадных генов в типах периферических клеток на срок до 12 часов, не затрагивая при этом фазу экспрессии циклических генов в SCN.
Время кормления оказывает значительное влияние на репертуар, фазу и амплитуду экспрессии ритмических генов. В исследовании было показано, что и временной режим приема пищи, и циркадные часы влияют на транскрипцию печеночных генов у мышей дикого типа [73].
Состав рациона — еще один важный фактор, влияющий на циркадные часы. Kohsaka et al. [74] показали, что диета с высоким содержанием жиров приводила к изменению периода ритма локомоторной активности и изменениям в экспрессии и цикличности генов канонических циркадных часов, ядерных рецепторов, которые регулируют факторы транскрипции часов, и задействованных генов, контролируемых часами. в использовании топлива в гипоталамусе, печени и жировой ткани.
Одним из примечательных нефотических сигналов для регулирования периферийных часов являются упражнения. Предполагается, что физическая активность или упражнения вызывают несколько физиологических изменений, таких как изменение температуры тела и гормонального статуса, которые, как известно, влияют на периферические часы через активацию симпатической нервной системы и высвобождение глюкокортикоидов [75]. Повышение температуры тела может действовать как входной сигнал для циркадного водителя ритма у млекопитающих [77]. Кроме того, упражнения способствуют выработке и высвобождению мелатонина.Благоприятный эффект 4-недельного лечения мелатонином для модуляции циркадных компонентов цикла сна и бодрствования обычно приводит к улучшению качества сна [78].
Напротив, молекулярные циркадные часы в периферических тканях могут реагировать на время выполнения упражнений, предполагая, что физическая активность обеспечивает важную временную информацию для синхронизации циркадных часов по всему телу. Хотя точная продолжительность и интенсивность упражнений, необходимых для изменения циркадной ритмики, не были определены, одно исследование на мышах показало, что упражнения на выносливость низкой интенсивности, поддерживаемые курсом по 2 часа в день в течение 4 недель, были достаточными, чтобы задействовать циркадные часы и изменить циркадную ритмичность [79].
Обычно, когда периферийные часы десинхронизируются с центральными часами, это приводит к сбою хронометража [80]. Это физиологическое изменение связано с различными заболеваниями, такими как рак, сердечно-сосудистые заболевания, депрессия, ожирение и метаболический синдром [72]. Например, при лечении ожирения основным подходом к диетическому лечению является ограничение потребления энергии [81]. Как правило, факторы, которые напрямую влияют на биологические ритмы, такие как время приема пищи и время сна, обычно не исследуются при планировании диеты.Циркадные часы играют важную роль в энергетическом гомеостазе и метаболических процессах. Следовательно, оценка факторов (сменная работа, нерегулярный сон, бессонница и т. Д.), Которые могут привести к нарушениям циркадного ритма у лиц с метаболическими заболеваниями, такими как ожирение, и планирование тренировок и времени приема пищи в соответствии с нормальными биологическими ритмами ( например, утренний выбор белковой пищи для увеличения термогенеза) может повысить эффективность лечения.
В последние годы для поддержания метаболического здоровья были разработаны методы лечения в зависимости от циркадного ритма, диетические вмешательства и упражнения, которые получили название «хронофармакология», «хроническое питание» и «хроноупражнения» соответственно.Хронофармакология изучает правильное время приема доз лекарственного средства для повышения эффективности, абсорбции и / или эффективности лекарственного средства [65]. Например, фермент HMG-COA, фермент, ограничивающий уровень холестерина, демонстрирует циркадный ритм у людей. Этот фермент достигает пика ночью, поэтому рекомендуется принимать на ночь препараты, снижающие уровень холестерина, такие как статины, чтобы максимизировать их эффективность [19]. Хроническое питание — это подход к определению оптимального усвоения питательных веществ для поддержания здоровья и регулирования циркадного ритма [82].Например, кофеин, нобилетин (флавоноид, содержащийся в цитрусовых) и ресвератрол в пищевых продуктах могут вызывать изменения циркадного ритма на молекулярном или поведенческом уровнях [83]. Chronoexercise в первую очередь исследует влияние продолжительности упражнений на поддержание здоровья и спортивные результаты, быстрые изменения в системе внутренних часов или повторное регулирование циркадных часов [84].
В результате циркадный ритм имеет двунаправленное взаимодействие почти со всеми метаболическими процессами и является основным фактором, влияющим на цикл сна-бодрствования.Следовательно, изучение и использование режима сна, информации о качестве и составление руководств по лечению с использованием циркадного ритма может повысить эффективность лечения заболевания. По этой причине могут быть разработаны новые подходы, перспективы и стратегии лечения метаболического баланса.
Благодарности
Нет.
Заявление об этике
Эта статья не содержит исследований с участием людей или животных, проведенных кем-либо из авторов.
Заявление о раскрытии информации
Авторы заявляют, что у них нет конфликта интересов.
Источник финансирования
Нет.
Вклад авторов
Ю.С. провели обзор литературы и N.A.T. организовал и подготовил рукопись. Все авторы читали и одобрили окончательный вариант рукописи.
Список литературы
- Акынджи Э., Орхан ФО.Sirkadiyen ritim uyku bozuklukları. Псикиятр Гунджел Якласимлар. 2016; 8 (2): 178–89.
- Özbayer C, Değirmenci İ. Sirkadiyen saat, hücre döngüsü ve kanser. Дикле Тип Дергиси; 2011. с. 38.
- Гумз М.Л., редактор.Циркадные часы: роль в здоровье и болезнях. Springer; 2016 г.
- Ричардс Дж., Гумз МЛ. Успехи в понимании периферических циркадных часов. FASEB J. 2012 сентябрь; 26 (9): 3602–13.
- Güldür T, Otlu HG.Циркадный ритм у млекопитающих: время есть и время спать. Biol Rhythm Res. 2017; 48 (2): 243–61.
- Аллен Р.П. В рецензируемой статье: увлечение мелатонином свободных циркадных ритмов у слепых людей. Sleep Med. Март 2001 г., 2 (2): 167–8.
- Санти Н., Лазар А.С., Маккейб П.Дж., Ло Дж.К., Грёгер Д.А., Дейк Д.Дж.Половые различия в суточной регуляции сна и познания в бодрствовании у людей. Труды Национальной академии наук. 2016: 201521637.
- Ферри Дж. Э., Кумари М., Сало П., Сингх-Ману А., Кивимяки М. Эпидемиология сна — быстро развивающаяся область. Издательство Оксфордского университета; 2011 г.
- Кронхольм Э., Партонен Т., Лаатикайнен Т., Пелтонен М., Хярма М., Хублин С. и др. Тенденции изменения продолжительности сна и симптомов, связанных с бессонницей, в Финляндии с 1972 по 2005 год: сравнительный обзор и повторный анализ выборок финского населения. J Sleep Res. Март 2008 г., 17 (1): 54–62.
- Чжу Л., Зи ПК. Расстройства циркадного ритма сна. Neurol Clin. 2012 ноябрь; 30 (4): 1167–91.
- Авраам Ф.Обзор функциональных причин бесплодия у коров. JFIV Reprod Med Genet. 2017; 5 (2): 203.
- Schernhammer ES, Laden F, Speizer FE, Willett WC, Hunter DJ, Kawachi I, et al. Работа в ночную смену и риск колоректального рака в исследовании состояния здоровья медсестер. J Natl Cancer Inst.2003 июн; 95 (11): 825–8.
- Гейл Дж. Э., Кокс Х. И., Цянь Дж., Блок Г. Д., Колвелл К. С., Матвеенко А. В.. Нарушение циркадных ритмов ускоряет развитие диабета из-за потери и дисфункции бета-клеток поджелудочной железы. J Biol Rhythms. 2011 Октябрь; 26 (5): 423–33.
- Мартино Т.А., Аудит Г.Ю., Герценберг А.М., Тата Н., Колетар М.М., Кабир Г.М. и др.Дезорганизация циркадного ритма вызывает у хомяков серьезные сердечно-сосудистые и почечные заболевания. Am J Physiol Regul Integr Comp Physiol. 2008 Май; 294 (5): R1675–83.
- Бакли П. Сон и нарушение циркадного ритма при шизофрении. Ежегодник психиатрии и прикладного психического здоровья; 2013.С. 411–2.
- Чхве СС, Ху Джи, Хван Ай Джей, Ким Джи, Ким Джи Би. Ремоделирование жировой ткани: его роль в энергетическом обмене и метаболических нарушениях. Фронт-эндокринол (Лозанна). 2016 Апрель; 7:30.
- On M, Dalğin D, Cenesiz M, Cenesiz S.Лептин ве адипонектинин enerji ve egzersiz ilişkisi. 2017 г.
- Цуджино Н., Сакураи Т. [Циркадный ритм лептина, орексина и грелина]. Нихон Риншо. 2012 июл; 70 (7): 1121–5.
- Сато Т, Ида Т, Кодзима М.Роль биологических ритмов в выполнении физических нагрузок. J Phys Fit Sports Med. 2017; 6 (3): 125–34.
- МакГиннис Г.Р., Молодой ME. Циркадная регуляция метаболического гомеостаза: причины и последствия. Nat Sci Sleep. 2016 Май; 8: 163–80.
- İbrahim Erdemir ET.Kortizol Sirkadiyen Ritmini Etkileyen Bazı Fiziksel ве Fizyolojik Parametrelerin Karşılaştırılması. Balıkesir Üniversitesi Sosyal Bilimler Enstitüsü Dergisi. 2008; 11: 1–10.
- Christiansen JJ, Djurhuus CB, Gravholt CH, Iversen P, Christiansen JS, Schmitz O, et al. Влияние кортизола на метаболизм углеводов, липидов и белков: исследования острого синдрома отмены кортизола при надпочечниковой недостаточности.J Clin Endocrinol Metab. 2007 сентябрь; 92 (9): 3553–9.
- Bolli GB, De Feo P, De Cosmo S, Perriello G, Ventura MM, Calcinaro F и др. Демонстрация феномена рассвета на нормальных людях-добровольцах. Диабет. 1984 декабрь; 33 (12): 1150–3.
- Рыбицка М., Крысяк Р., Окопень Б.Феномен рассвета и эффект Сомоги — два явления утренней гипергликемии. Эндокринол Pol. 2011. 62 (3): 276–84.
- Özçelik F, Erdem M, Bolu A, Gülsün M. Melatonin: genel özellikleri ve psikiyatrik bozukluklardaki rolü. Псикиятр Гунджел Якласимлар. 2013; 5 (2).
- Pan X, Zhang Y, Wang L, Hussain MM.Суточная регуляция MTP и триглицеридов плазмы с помощью CLOCK опосредуется SHP. Cell Metab. 2010 август; 12 (2): 174–86.
- Пан X, Хуссейн MM. Суточная регуляция микросомальных белков-переносчиков триглицеридов и уровней липидов в плазме. J Biol Chem. 2007 август; 282 (34): 24707–19.
- Пан X, Хуссейн MM.Часы важны для питания и суточной регуляции абсорбции макроэлементов у мышей. J Lipid Res. 2009 Сен; 50 (9): 1800–13.
- Бейли С.М., Удох США, Янг МЭ. Циркадная регуляция обмена веществ. J Endocrinol. 2014 Август; 222 (2): R75–96.
- Стивенсон Р.Циркадные ритмы и нарушения дыхания во сне. Sleep Med. 2007 сентябрь; 8 (6): 681–7.
- Muller JE, Stone PH, Turi ZG, Rutherford JD, Cheisler CA, Parker C, et al. Циркадные вариации частоты возникновения острого инфаркта миокарда. N Engl J Med. 1985 ноя; 313 (21): 1315–22.
- Фрой О. Метаболизм и циркадные ритмы — значение для ожирения. Endocr Rev.2010, февраль; 31 (1): 1–24.
- Крамер А., Мерроу М., редакторы.Циркадные часы. Springer; 2013.
- Браун С.А., Аззи А. Периферийные циркадные осцилляторы у млекопитающих; Циркадные часы. Springer; 2013. С. 45–66.
- Дэвидсон А.Дж., Лондон Б., Блок Г.Д., Менакер М.Сердечно-сосудистые ткани содержат независимые циркадные часы. Clin Exp Hypertens. 2005, февраль-апрель; 27 (2-3): 307–11.
- la Fleur SE, Kalsbeek A, Wortel J, Fekkes ML, Buijs RM. Суточный ритм толерантности к глюкозе: роль супрахиазматического ядра. Диабет. 2001 июн; 50 (6): 1237–43.
- Харада Н., Инагаки Н. Роль часовых генов в секреции инсулина. J. Исследование диабета. 2016 ноя; 7 (6): 822–3.
- Mıcılı S, Özoul C.Диябетте Кёк Хюкрелер. Dokuz Eylül Üniversitesi Tıp Fakültesi Dergisi. 2007; 21: 109–17.
- Sözlü S, anlier N. Sirkadiyen Ritim, Sağlık ve Beslenme İlişkisi. Turkiye Klinikleri Journal of Health Sciences. 2017; 2: 100–9.
- Кесслер К, Пивоварова О, Пфайффер АФ.[Циркадные часы и энергетический обмен: значение для здоровья]. Dtsch Med Wochenschr. 2014 Апрель; 139 (14): 684–6.
- Санкар Г., Бруннер М. Циркадные часы и энергетический метаболизм. Cell Mol Life Sci. Июль 2014 г .; 71 (14): 2667–80.
- Масри С.Сиртуин-зависимый контроль часов: новые достижения в метаболизме, старении и раке. Curr Opin Clin Nutr Metab Care. 2015 ноя; 18 (6): 521–7.
- Ян Х, Даунс М., Ю Р.Т., Буккаут А.Л., Хе В., Страуме М. и др. Экспрессия ядерных рецепторов связывает циркадные часы с метаболизмом. Клетка. 2006 август; 126 (4): 801–10.
- Фрой О., Мискин Р. Взаимосвязь кормления, циркадных ритмов и старения. Prog Neurobiol. 2007 июнь; 82 (3): 142–50.
- Байрам А., Мехри И.Sirtuin Genleri ve İşlevleri. Firat Tip Derg. 2013; 18: 136–40.
- Раттер Дж., Рейк М., Ву Л.С., Макнайт С.Л. Регулирование часов и связывания ДНК NPAS2 окислительно-восстановительным состоянием кофакторов NAD. Наука. Июль 2001 г., 293 (5529): 510–4.
- Ум Дж. Х., Ян С., Ямадзаки С., Кан Х., Виоллет Б., Форец М. и др.Активация 5′-AMP-активированной киназы лекарственным средством от диабета метформином индуцирует зависимую от казеинкиназы иепсилон (CKIepsilon) деградацию часового белка mPer2. J Biol Chem. Июль 2007 г., 282 (29): 20794–8.
- Кумар Джа П., Шалле Э., Калсбек А. Циркадные ритмы метаболизма глюкозы и липидов у ночных и дневных млекопитающих.Mol Cell Endocrinol. 2015 декабрь; 418 (Pt 1): 74–88.
- Бенедикт Ф.Г. Факторы, влияющие на основной обмен. Proc Natl Acad Sci USA. 1915 Февраль; 1 (2): 105–9.
- Haugen HA, Melanson EL, Tran ZV, Kearney JT, Hill JO.Вариабельность измеренной скорости метаболизма в состоянии покоя. Am J Clin Nutr. 2003 декабрь; 78 (6): 1141–5.
- Лапоски А.Д., Басс Дж., Кохсака А., Турек Ф.В. Сон и циркадные ритмы: ключевые компоненты регуляции энергетического обмена. FEBS Lett. Январь 2008 г., 582 (1): 142–51.
- Алгин Д.И., Акдаг Г., Эрдинч О.О.Калители уйку ве уйку бозуклуклари. Османгази медицинский журнал. 2016; 38.
- Шахин Л., Аджиоглу М., Ташкин Э. Уйку ве уйкунун дюзенленмеси. Sağlık Bilimleri Dergisi. 2013; 22: 93–8.
- Boscolo RA, Esteves AM, de Santana MG, Viana VAR, Grassmann V, Tufik S, de Mello MT.Есть ли связь между составом тела, основной скоростью метаболизма и сном у пожилых пациентов с синдромом обструктивного апноэ во сне и без него? Наука о сне. 2013. 6 (4): 129–134.
- Кнутсон К.Л., Шпигель К., Пенев П., Ван Каутер Э. Метаболические последствия лишения сна.Sleep Med Rev.2007 июн; 11 (3): 163–78.
- Chtourou H, Souissi N. Эффект тренировки в определенное время суток: обзор. J Strength Cond Res. 2012 Июль; 26 (7): 1984–2005.
- Суиси Н., Готье А., Сесбое Б., Лару Дж., Давенн Д.Циркадные ритмы в двух типах упражнений для ног анаэробного цикла: сила-скорость и 30-секундный тест Вингейта. Int J Sports Med. 2004, январь; 25 (1): 14–9.
- Kinişler A. Anaerobik Performansta sirkadiyen değişimlerin incelenmesi. Spor Bilimleri Dergisi. 2005. 16: 174–84.
- Кройчи К.Как регулируется циркадный ритм основной температуры тела? Clin Auton Res. 2002 июн; 12 (3): 147–149.
- Райт К.П. младший, Халл Дж. Т., Чейслер, Калифорния. Взаимосвязь между бдительностью, работоспособностью и температурой тела у людей. Am J Physiol Regul Integr Comp Physiol. 2002 декабрь; 283 (6): R1370–7.
- Беннард П., Дусет Э. Острые эффекты времени выполнения упражнений и гликемического индекса завтрака на окисление жиров, вызванное физической нагрузкой. Appl Physiol Nutr Metab. 2006 Октябрь; 31 (5): 502–11.
- Катсанос К.С., Моффатт Р.Дж.Острые эффекты упражнений перед едой по сравнению с упражнениями после еды на постпрандиальную гипертриглицеридемию. Clin J Sport Med. 2004, январь; 14 (1): 33–9.
- Фарах Н.М., Гилл Дж. М.. Влияние упражнений до или после приема пищи на баланс жиров и постпрандиальный метаболизм у мужчин с избыточным весом. Br J Nutr.2013 июн; 109 (12): 2297–307.
- Канг Дж., Рейнс Э., Розенберг Дж., Ратамесс Н., Наклерио Ф., Файгенбаум А. Метаболические реакции во время постпрандиальных упражнений. Res Sports Med. 2013. 21 (3): 240–52.
- Шибата С., Тахара Ю.Циркадный ритм и упражнения. J Phys Fit Sports Med. 2014; 3 (1): 65–72.
- Паоли А., Марколин Дж., Зонин Ф., Нери М., Сивьери А., Пачелли QF. Делать упражнения натощак или кормить для ускорения похудания? Влияние приема пищи на соотношение дыхательных путей и избыточное потребление кислорода после тренировки после тренировки на выносливость.Int J Sport Nutr Exerc Exerc Metab. 2011 Февраль; 21 (1): 48–54.
- Таппи Л. Термический эффект пищи и деятельность симпатической нервной системы человека. Reprod Nutr Dev. 1996. 36 (4): 391–7.
- Romon M, Edme JL, Boulenguez C, Lescroart JL, Frimat P.Циркадные вариации термогенеза, вызванного диетой. Am J Clin Nutr. 1993, апрель; 57 (4): 476–80.
- Моррис С.Дж., Гарсия Д.И., Майерс С., Ян Дж., Триенекенс Н., Шеер Ф.А. Циркадная система человека играет доминирующую роль в возникновении утренних и вечерних различий в термогенезе, вызванном диетой.Ожирение (Серебряная весна). 2015 Октябрь; 23 (10): 2053–8.
- Grammaticos PC, Doumas A, Koliakos G. Половина утреннего и ночного опорожнения желудка различалась более чем на 220% у двух молодых здоровых взрослых людей. Ад J Nucl Med. 2015, январь-апрель; 18 (1): 60–2.
- Hoogerwerf WA.Роль часовых генов в моторике желудочно-кишечного тракта. Am J Physiol Gastrointest Liver Physiol. 2010 сентябрь; 299 (3): G549–55.
- Гараулет М., Гомес-Абеллан П. Время приема пищи и ожирение: новая ассоциация. Physiol Behav. 2014 июль; 134: 44–50.
- Фоллмерс К., Гилл С., Ди Таккио Л., Пуливарти С.Р., Ле HD, Панда С.Время кормления и внутренние ритмы привода циркадных часов в экспрессии генов печени. Proc Natl Acad Sci U S. A. 2009, 15 декабря; 106 (50): 21453–8.
- Kohsaka A, Laposky AD, Ramsey KM, Estrada C, Joshu C., Kobayashi Y, et al. Диета с высоким содержанием жиров нарушает поведенческие и молекулярные циркадные ритмы у мышей.Cell Metab. 2007 ноябрь; 6 (5): 414–21.
- Тахара Ю., Аояма С., Шибата С. Циркадные часы млекопитающих и их увлечение стрессом и физическими упражнениями. J Physiol Sci. 2017 Янв; 67 (1): 1–10.
- Дамиола Ф., Ле Минь Н., Прейтнер Н., Корнманн Б., Флери-Олела Ф., Шиблер У.Ограниченное питание отделяет циркадные осцилляторы в периферических тканях от центрального водителя ритма в супрахиазматическом ядре. Genes Dev. 2000 декабрь; 14 (23): 2950–61.
- Brown SA, Zumbrunn G, Fleury-Olela F, Preitner N, Schibler U. Ритмы температуры тела млекопитающих могут поддерживать периферические циркадные часы.Curr Biol. 2002 Сен; 12 (18): 1574–83.
- Леонардо-Мендонса Р.К., Мартинес-Николас А., де Тереза Гальван С., Оканья-Вильхельми Дж., Русанова И., Герра-Эрнандес Е. и др. Преимущества четырехнедельного лечения мелатонином на циркадные ритмы у спортсменов, тренирующихся с отягощениями. Chronobiol Int.2015; 32 (8): 1125–34.
- Wolff G, Esser KA. Запланированная фаза упражнений сдвигает циркадные часы в скелетных мышцах. Медико-спортивные упражнения. 2012 сентябрь; 44 (9): 1663–70.
- Гараулет М., Ордовас Я. М., Мадрид, Я.Хронобиология, этиология и патофизиология ожирения. Int J Obes. 2010 декабрь; 34 (12): 1667–83.
- Фрой О. Циркадные ритмы и ожирение у млекопитающих. ISRN Obes. 2012 18 ноября; 2012: 437198.
- Тахара Ю., Шибата С.Хронобиология и питание. Неврология. 2013 декабрь; 253: 78–88.
- Ойке Х. Модуляция суточных часов питательными веществами и пищевыми факторами. Biosci Biotechnol Biochem. 2017 Май; 81 (5): 863–70.
- Накао Р.Циркадные часы скелетных мышц: текущие исследования. Хронофизиология и терапия. 2017; 7: 47–57.
Автор Контакты
Nilüfer Acar Tek
Факультет медицинских наук, Департамент питания и диетологии
Университет Гази, район Эмниет, улица Муаммера Яшара Бостанджи
Номер 16, TR – 06560 Анкара (Турция)
Электронная почта acarnil @ hotmail.com
Подробности статьи / публикации
Предварительный просмотр первой страницы
Поступила: 26 июня 2018 г.
Дата принятия: 30 марта 2019 г.
Опубликована онлайн: 23 апреля 2019 г.
Дата выпуска: июнь 2019 г.
Количество страниц для печати: 9
Количество рисунков: 1
Количество столов: 1
ISSN: 0250-6807 (печатный)
eISSN: 1421-9697 (онлайн)
Для дополнительной информации: https: // www.karger.com/ANM
Авторские права / Дозировка препарата / Заявление об ограничении ответственности
Авторские права: Все права защищены. Никакая часть данной публикации не может быть переведена на другие языки, воспроизведена или использована в любой форме или любыми средствами, электронными или механическими, включая фотокопирование, запись, микрокопирование, или с помощью какой-либо системы хранения и поиска информации, без письменного разрешения издателя. .
Дозировка лекарства: авторы и издатель приложили все усилия, чтобы гарантировать, что выбор и дозировка лекарства, указанные в этом тексте, соответствуют текущим рекомендациям и практике на момент публикации.Тем не менее, ввиду продолжающихся исследований, изменений в правительственных постановлениях и постоянного потока информации, касающейся лекарственной терапии и реакций на них, читателю рекомендуется проверять листок-вкладыш для каждого препарата на предмет любых изменений показаний и дозировки, а также дополнительных предупреждений. и меры предосторожности. Это особенно важно, когда рекомендованным средством является новое и / или редко применяемое лекарство.
Отказ от ответственности: утверждения, мнения и данные, содержащиеся в этой публикации, принадлежат исключительно отдельным авторам и соавторам, а не издателям и редакторам.Появление в публикации рекламы и / или ссылок на продукты не является гарантией, одобрением или одобрением рекламируемых продуктов или услуг или их эффективности, качества или безопасности. Издатель и редактор (-ы) не несут ответственности за любой ущерб, нанесенный людям или имуществу в результате любых идей, методов, инструкций или продуктов, упомянутых в контенте или рекламе.
Глава 16. Цинк
Глава 16. Цинк
Роль цинка в метаболизме человека
процессы
Метаболизм цинка и
гомеостаз
Диетические источники и доступность
цинк
Группы риска по цинку
дефицит
Потребность в цинке
Индивидуальные различия в цинке
потребности и рекомендуемое потребление питательных веществ
Верхний предел содержания цинка
потребление
Адекватность потребления цинка по отношению к
к оценкам требований
Заключение
Дальнейшее исследование
Ссылки
Роль цинка в метаболизме человека
процессы
Цинк присутствует во всех тканях и жидкостях организма.Все тело
содержание цинка оценивается в 30 ммоль (2
грамм). Скелетные мышцы составляют примерно 60 процентов всего тела.
содержание и костная масса, с концентрацией цинка 1,5-3 ммоль / г (100-200 мг / г), примерно на 30 процентов.
Концентрация цинка в безжировой массе тела составляет примерно 0,46 ммоль / г (30 мг / г). Плазменный цинк имеет быструю смену
и составляет лишь около 0,1 процента от общего содержания цинка в организме. Этот уровень
кажется, находится под тщательным гомеостатическим контролем.Высокие концентрации цинка
в сосудистой оболочке глаза 4,2 ммоль / г (274
мкг / г) и в жидкостях предстательной железы 4,6-7,7 ммоль / л (300-500 мг / л)
( 1 ).
Цинк является важным компонентом большого количества (> 300)
ферменты, участвующие в синтезе и расщеплении углеводов, липидов,
белков и нуклеиновых кислот, а также в метаболизме других
микроэлементы. Цинк стабилизирует молекулярную структуру клеточных компонентов.
и мембраны и таким образом способствует поддержанию клеток и органов
честность.Кроме того, цинк играет важную роль в полинуклеотиде.
транскрипции и, следовательно, в процессе генетической экспрессии. Его участие в
такая фундаментальная деятельность, вероятно, объясняет важность цинка для
все формы жизни.
Цинк играет центральную роль в иммунной системе, влияя на
ряд аспектов клеточного и гуморального иммунитета ( 2 ). Роль цинка
в области иммунитета был подробно рассмотрен Shanglar et al .
( 2 ).
Клинические признаки тяжелого дефицита цинка у людей:
задержка роста, задержка полового созревания и созревания костей, поражения кожи, диарея,
алопеция, нарушение аппетита, повышенная восприимчивость к инфекциям, опосредованным через
дефекты иммунной системы и появление изменений в поведении
( 1 ). Последствия маргинального или легкого дефицита цинка менее очевидны. А
снижение скорости роста и нарушение иммунной защиты — пока единственные
четко продемонстрированы признаки умеренного дефицита цинка у людей.Прочие эффекты,
такие как нарушение вкуса и заживление ран, которые, как утверждается, являются результатом
низкое потребление цинка наблюдаются реже.
Метаболизм цинка и
гомеостаз
Всасывание цинка зависит от концентрации и происходит
по всему тонкому кишечнику. В нормальных физиологических условиях транспорт
процессы поглощения не насыщены. Цинк вводят в водных растворах
у людей, голодных, всасывается эффективно (60-70 процентов), тогда как всасывание
от твердых диет менее эффективен и варьируется в зависимости от содержания цинка и диеты
состав ( 3 ).
Цинк выводится из организма через почки, кожу и
кишечник. Эндогенные кишечные потери могут варьироваться от 7 ммоль / день (0,5 мг / день) до более 45 ммоль / день (3 мг / день), в зависимости от потребления цинка.
( 4 ). Потери с мочой и кожей составляют порядка 7-10 ммоль / день (0,5-0,7 мг / день) каждая и в меньшей степени зависят от нормы.
вариации в потреблении цинка ( 4 ). Повышается голодание и мышечный катаболизм
потери цинка с мочой. Тяжелые упражнения и повышенная температура окружающей среды могут
приводят к потерям из-за потоотделения.
В теле нет запасов цинка в общепринятом понимании. В
условиях резорбции костей и катаболизма тканей, цинк высвобождается и может
в какой-то степени используется повторно. Экспериментальные исследования на людях с диетами с низким содержанием цинка
2,6-3,6 мг / день (40-55 ммоль / день) показали, что
уровни циркулирующего цинка и активность цинксодержащих ферментов могут быть
поддерживается в пределах нормы в течение нескольких месяцев ( 5 , 6 ), что
подчеркивает эффективность механизма гомеостаза цинка.Контролируемый
Исследования истощения-восполнения запасов на людях показали, что изменения в эндогенных
выведение цинка через почки, кишечник и кожу и изменения в
абсорбционная эффективность — это то, как поддерживается содержание цинка в организме
( 7 — 10 ). Основные механизмы плохо изучены.
Чувствительные индексы для оценки статуса цинка неизвестны в
настоящее время. Статические показатели, такие как концентрация цинка в плазме, клетках крови и
волосы и экскреция цинка с мочой уменьшаются при тяжелом дефиците цинка.А
количество состояний, не связанных со статусом цинка, может повлиять на все эти
индексы, особенно уровни цинка в плазме. Инфекция, стрессовые ситуации, такие как
лихорадка, прием пищи и беременность снижают концентрацию цинка в плазме, тогда как при
Например, длительное голодание увеличивает его ( 11 ). Однако по населению
По всей видимости, снижение концентрации цинка в плазме является маркером
снижение роста в зависимости от цинка ( 12 , 13 ). Экспериментальный цинк
Исследования истощения предполагают, что изменения иммунного ответа происходят раньше, чем
очевидно снижение концентрации цинка в плазме ( 14 ).Пока это
не удалось идентифицировать цинк-зависимые ферменты, которые могли бы служить
ранние маркеры статуса цинка.
Был проведен ряд функциональных показателей статуса цинка.
предложил, например, заживление ран, остроту вкуса и адаптацию к темноте
( 11 ). Однако изменения в этих функциях не характерны для цинка и
эти индексы до сих пор не доказали свою полезность для определения маргинального цинка.
дефицит у человека.
Внедрение методов стабильных изотопов в исследованиях цинка
( 15 ) создал возможности для оценки взаимосвязи между
диета и статус цинка и, вероятно, приведет к лучшему пониманию
механизмы, лежащие в основе гомеостатической регуляции цинка.Оценки
скорость оборота введенных изотопов в плазме или моче выявила
наличие относительно небольшого быстро заменяемого пула цинка в организме примерно
1,5-3 ммоль (100-200 мг) ( 16-19 ). Размер
бассейна, похоже, коррелирует с обычным диетическим потреблением, и оно сокращается
в исследованиях контролируемого истощения ( 18 ). Сменный цинковый пул также был
установлено, что это коррелирует с эндогенным фекальным экскрецией цинка ( 19 ) и
общее суточное всасывание цинка.Эти данные позволяют предположить, что размер
сменный пул зависит от недавно усвоенного цинка и от большего
обменный пул приводит к увеличению эндогенной экскреции. Изменения эндогенного
кишечная экскреция цинка кажется более важной, чем изменения в
абсорбционная эффективность для поддержания гомеостаза цинка ( 19 ).
Диетические источники и наличие
цинк
Постное красное мясо, цельнозерновые злаки, бобовые и бобовые
обеспечивают максимальные концентрации цинка 25-50 мг / кг (380-760 ммоль / кг) сырой массы.Обработанные крупы с низким
степень извлечения, шлифованный рис и нежирное мясо или мясо с высоким содержанием жира
имеют умеренное содержание цинка 10-25 мг / кг (150-380 ммоль / кг). Рыба, корнеплоды и клубни, зеленые листовые
овощи и фрукты являются лишь скромными источниками цинка <10 мг / кг (<150
ммоль / кг) ( 20 ). Сепарированные жиры и масла,
сахар и алкоголь имеют очень низкое содержание цинка.
Утилизация цинка зависит от общего состава
диета. Экспериментальные исследования выявили ряд диетических факторов, как
потенциальные промоторы или антагонисты абсорбции цинка ( 21) .Растворимый
низкомолекулярные органические вещества, такие как амино- и гидроксикислоты,
облегчают всасывание цинка. Напротив, органические соединения, образующие стабильные и
плохо растворимые комплексы с цинком могут ухудшить абсорбцию. Кроме того,
конкурентные взаимодействия между цинком и другими ионами с аналогичными
физико-химические свойства могут влиять на поглощение и всасывание в кишечнике
цинк. Риск конкурентного взаимодействия, по-видимому, в основном связан с высокой
дозы в виде добавок или в водных растворах.Однако на уровнях
присутствует в пище и при реалистичных уровнях обогащения, появляется абсорбция цинка
не подвергаться влиянию, например, железа и меди ( 21 ).
Изотопные исследования с участием людей выявили два
факторы, которые вместе с общим содержанием цинка в рационе являются основными
детерминанты всасывания и использования пищевого цинка. Первый — это
содержание гексафосфата инозита (фитата), а второе — уровень и
источник диетического белка.Фитаты присутствуют в цельнозерновых злаках и
бобовые и в меньших количествах другие овощи. У них большой потенциал
для связывания двухвалентных катионов и их угнетающего действия на всасывание цинка имеет
продемонстрировано на людях ( 21) . Молярное соотношение между фитатами и
цинк в еде или диетах является полезным индикатором влияния фитатов на
подавляет всасывание цинка. При молярных соотношениях выше 6-10 цинк
абсорбция начинает снижаться; при соотношении выше 15 абсорбция обычно меньше
более 15 процентов ( 20 ).Однако действие фитата изменяется из-за
источник и количество потребляемых белков. Белки животного происхождения улучшают содержание цинка
абсорбция из диеты, содержащей фитаты. Поглощение цинка некоторыми
диеты на основе бобовых сравнимы с диетами на основе животных белков.
несмотря на более высокое содержание фитатов в первом. Повышенный уровень кальция в рационе
антагонистические эффекты фитатов на абсорбцию цинка в экспериментальных исследованиях.
Результаты исследований на людях менее последовательны, и любые эффекты кажутся
зависят от источника кальция и состава рациона
( 22 ).
Некоторые примеры недавно опубликованных исследований абсорбции
проиллюстрировать влияние содержания цинка и состава рациона на фракционное содержание цинка
абсорбция ( Таблица 53 ) ( 19, 23-25 ). Результаты
исследования общей диеты, когда все основные приемы пищи в течение дня были
с внешней маркировкой, демонстрируют замечательную стабильность фракционного поглощения
несмотря на относительно большие различия в составе еды и содержании цинка. Таким образом,
примерно в два раза больше цинка было усвоено из невегетарианцев или мяса с высоким содержанием мяса.
диета ( 24 , 25 ), чем из диеты в сельских районах Китая на основе риса и
мука пшеничная ( 20 ).Отсутствуют данные о всасывании цинка из типичных диет.
развивающихся стран, которые обычно имеют высокое содержание фитатов.
Доступность цинка из рациона можно повысить, если
снижение содержания фитатов и включение источников животного белка. Ниже
степень извлечения зерна злаков приведет к более низкому содержанию фитата, но при
в то же время содержание цинка снижается, так что чистое влияние на поставку цинка
ограничено. Содержание фитата можно снизить, активируя присутствующую фитазу.
в большинстве пищевых продуктов, содержащих фитат, или за счет добавления микробных или грибковых
фитазы.Фитазы гидролизуют фитат с понижением содержания фосфатов инозита, в результате чего
в улучшенном всасывании цинка ( 26 , 27 ). Активность фитаз
в тропических злаках, таких как кукуруза и сорго, ниже, чем в пшенице и
рожь ( 28 ). Прорастание зерновых и бобовых культур увеличивает активность фитазы.
и добавление некоторого количества пророщенной муки к непроработанной кукурузе или сорго с последующим добавлением
замачивание при температуре окружающей среды в течение 12-24 часов может снизить содержание фитатов
по существу ( 28 ).Дополнительного снижения можно добиться за счет
ферментация каши для прикорма или теста для выпечки хлеба.
Также можно использовать коммерчески доступные препараты фитазы, но их нельзя
экономически доступны для многих групп населения.
Группы риска по цинку
дефицит
Центральная роль цинка в делении клеток, синтезе белков,
и рост делает младенцев, детей, подростков и беременных женщин особенно
существует риск неадекватного потребления цинка.Задержка в росте, вызванная воздействием цинка, была
выявлено в нескольких исследованиях ( 29 ), и более быстрое увеличение массы тела у
Сообщалось о детях с недостаточным питанием, получавших добавки цинка. В других исследованиях есть
не смогли продемонстрировать стимулирующий рост эффект от приема цинка ( 13 ). А
недавний метаанализ 25 интервенционных исследований с участием 1834 детей в возрасте до 13 лет.
лет, со средней продолжительностью около 7 месяцев и средней дозой
цинка 14 мг / день (214 ммоль / день), показал небольшой
но значительный положительный эффект от приема цинка на рост и вес
увеличивается ( 13 ).Первоначальное присутствие задержки роста было значительно
связан с эффектом приема цинка на рост, тогда как исходный низкий
концентрации цинка в плазме были связаны с более выраженным влиянием на
увеличение веса.
Результаты исследований добавок цинка показывают, что низкий
статус цинка у детей не только влияет на рост, но также связан с
повышенный риск тяжелых инфекционных заболеваний ( 30 ). Эпизоды острого
диарея с меньшей продолжительностью и меньшей тяжестью, а также снижение заболеваемости
сообщалось о диарее в группах, принимавших цинк.Другие исследования указывают на
что заболеваемость острыми инфекциями нижних дыхательных путей и малярией может
также может быть уменьшено добавлением цинка. Профилактика неоптимального цинкового статуса
и дефицит цинка у детей из-за повышенного потребления и доступности цинка
может, следовательно, оказать значительное влияние на здоровье ребенка в процессе развития
страны.
Роль цинкового статуса матери в исходе беременности все еще остается
не понятно. Положительные и отрицательные ассоциации между содержанием цинка в плазме
концентрация и рост плода или роды и осложнения при родах были
сообщил ( 31 ).Результаты исследований добавок цинка также остаются
безрезультатно ( 31 ). Интерпретация концентрации цинка в плазме крови
беременность осложняется действием гемодилюции и низким содержанием цинка в плазме
уровни могут отражать другие метаболические нарушения ( 11 ). Цинк
Исследования добавок беременных женщин проводились в основном в
относительно хорошо обеспеченное население, что может быть одной из причин
смешанные результаты ( 31 ). Недавнее исследование среди американских женщин с низким доходом с
концентрации цинка в плазме ниже среднего при включении в дородовую помощь показали
что потребление цинка 25 мг / день привело к увеличению веса новорожденного при рождении и
окружность головы и снижение очень низкой массы тела при рождении среди людей, не страдающих ожирением
женщин по сравнению с группой плацебо ( 12 ).
Таблица 53
Примеры фракционного поглощения цинка из общего рациона
измерено изотопными методами
|
Характеристики объекта (исх.) |
Диета / особенности приема пищи |
Изотопная техника |
Содержание цинка м моль |
Мольное соотношение фитат-цинк |
Поглощение цинка,% (x ± стандартное отклонение) а |
|
Молодые люди (n = 8) (22) |
Диета с высоким содержанием клетчатки |
Радиоизотопы |
163 (10.7) |
7 |
27 ± 6 |
|
Привычный рацион |
Стабильные изотопы |
124 (8,1) |
10 |
34 ± ± 9 |
|
|
Девушки (n = 10) (19) |
Лакто-ово-вегетарианские |
Радиоизотопы |
139 (9.1) |
14 |
26 б |
|
Женщины (20-42 года) (n = 21) (24) |
Невегетарианец |
Радиоизотопы |
169 (11,1) |
5 |
33 б |
|
Женщины (20-42 года) (n = 21) (24) |
Мясное мясо |
Радиоизотопы |
102 (6.7) |
– |
30 в |
|
Женщины в постменопаузе (n-14) (25) |
Высокое мясо |
Радиоизотопы |
198 (13,0) |
– |
28 в |
a SD, стандартное отклонение. b Объединенный
SD = 5. c Объединенное SD = 4,6.
Источник: адаптировано из материалов ФАО / ВОЗ по следовым минералам.
Отчет ( 32 )
Требования к цинку
Отсутствие специфических и чувствительных индексов статуса цинка
ограничивает возможности оценки потребности в цинке на основании эпидемиологических
наблюдения. В отчете ФАО / МАГАТЭ / ВОЗ за 1996 год ( 32 ) потребности в цинке
были оценены с использованием факторного метода (т. е. путем добавления
требования для роста тканей, поддержания, метаболизма и эндогенных потерь).Экспериментальные исследования восполнения запасов цинка при низком потреблении цинка ясно показали, что
что организм имеет ярко выраженную способность адаптироваться к разным уровням цинка
поступления за счет изменения эндогенных потерь цинка через почки, кишечник,
и кожа ( 5-9,33 ). Нормативные требования к абсорбированному цинку составляли
определяется как обязательная потеря на ранней стадии истощения цинка перед
имеет место адаптивное снижение экскреции, которое было установлено на уровне 1,4 мг / день для мужчин
и 1.0 мг / день для женщин. Оценить нормативные требования к содержанию
другие возрастные группы, соответствующие показатели основного метаболизма использовались для
экстраполяция. У растущих особей скорость прироста и содержание цинка
вновь сформированные ткани были использованы для получения данных, необходимых для роста тканей.
Аналогичным образом, удержание цинка во время беременности и концентрация цинка в
молоко на разных стадиях лактации использовалось для оценки физиологического
потребности при беременности и кормлении грудью ( 32 ).
Перевод этих оценок поглощенного цинка в
Потребность в диетическом цинке включает несколько соображений. Во-первых, природа
диеты (то есть содержание в ней промоторов и ингибиторов абсорбции цинка)
определяет потенциально усваиваемую долю цинка в пище.
Во-вторых, эффективность поглощения потенциально доступного цинка обратно пропорциональна
связано с содержанием цинка в рационе. Обзор имеющихся данных из
экспериментальные исследования абсорбции цинка при однократном или полном рационе питания привели к
разделение диет на три категории — с высоким, умеренным и низким содержанием цинка
биодоступность — как указано в Таблица 54 ( 32 ).Это было
затем обнаружил, что взаимосвязь между эффективностью абсорбции и цинком
содержание различалось для этих диет ( 32 ). Разработаны алгоритмы
( 32 ) и применен к оценке потребности в поглощенном цинке в
достичь набора цифр для средних индивидуальных диетических потребностей в цинке
( Таблица 55 ). Показатели дробного поглощения, применяемые для
три категории диеты составляли 50 процентов, 30 процентов и 15 процентов соответственно.На основании этих оценок и оценки данных о потреблении с пищей
исследований, были определены средние поступления в популяции, которые были сочтены достаточными для
обеспечить низкую распространенность людей с риском недостаточного потребления цинка
( 32 ).
Младенцы, дети и подростки
Эндогенные потери цинка у младенцев, вскармливаемых грудным молоком, составили
предполагается, что составляет 20 мкг / кг / день (0,31 ммоль / кг / день)
тогда как 40 мкг / кг / день (0,6 ммоль / кг / день) было
принято для младенцев, получающих смесь или продукты для отлучения от груди ( 32 ).Для другого возраста
группирует среднюю потерю 0,002 ммоль / базальный кДж
(0,57 мкг / базальная ккал) было получено из оценок для взрослых. По оценкам
повышение содержания цинка для роста ребенка было установлено на 120 и 140 мкг / кг / день.
(1,83-2,14 ммоль / кг / день) для женщин и мужчин
младенцы соответственно в первые 3 мес ( 32 ). Эти значения уменьшаются
до 33 мкг / кг / день (0,50 ммоль / кг / день) в возрасте
6-12 месяцев. Для возраста 1-10 лет требования к росту были основаны на
предположение, что новая ткань содержит 30 мкг / г (0.46 ммоль цинка / г) ( 32 ). Для подросткового роста цинк
содержание 23 мкг / г (0,35 ммоль / г) увеличение
масса тела была принята. Вспышки полового созревания повышают физиологический уровень цинка
требования существенно. Рост самцов-подростков соответствует
увеличение потребности организма в цинке примерно на 0,5 мкг / день (7,6 ммоль / день) ( 32 ).
Беременность
Общее количество цинка, оставшегося во время беременности, было
оценивается в 1.5 ммоль (100 мг) ( 34).
В течение третьего триместра физиологическая потребность в цинке составляет примерно
вдвое выше, чем у небеременных женщин ( 32 ).
Лактация
Концентрация цинка в грудном молоке высока в раннем периоде лактации,
2-3 мг / л (31-46 ммоль / л) в первый месяц, и
снижается до 0,9 мг / л (14 ммоль / л) через 3 месяца
( 35 ). Исходя из данных об объеме материнского молока и содержании цинка, было
подсчитано, что суточный выход цинка с молоком в течение первых 3 месяцев
лактация могла составлять 1.4 мг / день (21,4 ммоль / л), что теоретически
в три раза больше физиологической потребности в цинке у кормящих женщин по сравнению с
Не кормящим, небеременным женщинам. При установлении ориентировочных требований к
в раннем периоде лактации предполагалось, что часть этого требования была покрыта
послеродовая инволюция матки и резорбция скелета
( 32 ).
Пожилые люди
Требования к пожилым людям примерно такие же, как
те для других взрослых.Сообщается о более низкой абсорбционной эффективности
пожилые люди, что может оправдать более высокие диетические требования. С другой стороны,
эндогенные потери, по-видимому, ниже у пожилых людей. Из-за предложенной роли
цинка при инфекционных заболеваниях, оптимальный статус цинка у пожилых людей может иметь
оказывает значительное влияние на здоровье населения и является областью метаболизма цинка, требующей
дальнейшие исследования.
Различия цинка между индивидуумами
потребности и рекомендуемое потребление питательных веществ
Исследования ( 6 — 10 ), использованные для оценки среднего
физиологические потребности с факторной техникой считались
относительно небольшое количество предметов и не позволяют оценить
межиндивидуальные различия в обязательных потерях цинка при разных потребностях.Поскольку потребности в цинке связаны со скоростью обновления и роста тканей, это
разумно предположить, что вариации физиологической потребности в цинке
такая же величина, как и вариации потребности в белке ( 36 ), и что
такой же показатель (12,5%) для межиндивидуального коэффициента вариации
(CV) может быть принято. Однако оценки потребности в цинке с пищей
предполагают оценку поглощения. Следовательно, вариации поглощающей способности
эффективность, не имеющая отношения к оценке потребности в белке, может
должны быть приняты во внимание при оценке общей межличностной
изменение требований к цинку.Систематические исследования межличностного
вариации в абсорбции цинка в различных условиях немногочисленны. В малых
группы здоровых, хорошо питающихся субъектов, сообщаемые различия в содержании цинка
абсорбция из определенного приема пищи или диеты составляет порядка 20-40 процентов и
кажутся независимыми от возраста, пола или особенностей диеты. Сколько этих
вариации, помимо того, что они связаны с методологической неточностью, отражают
вариации физиологической потребности, последствия предшествующего приема цинка,
и т. Д. .не известно. Из имеющихся данных исследований абсорбции цинка
( 19, 20, 23-27 ) предварительно предполагается, что различия в диете
требования к цинку, которые включают изменение требований к абсорбированному цинку.
(т.е. вариации метаболизма и скорости оборота цинка) и вариации в
абсорбционная эффективность соответствует CV 25 процентов. Рекомендуемый
потребление питательных веществ, полученное на основе оценок среднего индивидуального диетического питания.
потребности ( Таблица 55 ) с добавлением 50 процентов (2
стандартные отклонения) приведены в Таблица 56 .
Таблица 54
Критерии для категоризации диет в соответствии с потенциалом
наличие цинка
|
Номинальная категория |
Основные диетические характеристики |
|
Высокая доступность |
Рафинированные диеты с низким содержанием зерновой клетчатки и фитиновой кислоты, |
|
Средняя доступность |
Смешанные диеты, содержащие белок животного или рыбного происхождения. Лакто-ово, |
|
Низкая доступность |
Рационы с высоким содержанием нерафинированных, неферментированных и непроросших злаков |
a Прорастание таких зерен или ферментация (например,
закваска) многих видов муки может снизить антагонистическое действие; диета должна тогда
быть классифицированным как умеренная доступность. b Овощные диеты с
соотношения фитат-цинк, превышающие 30, известны; для таких диет предположение
наличие 10 процентов цинка или менее может быть оправдано, особенно если
потребление белка низкое, солей кальция чрезмерно или и то, и другое (например,г., кальций
соли, обеспечивающие> 1,5 г Ca 2+ / день).
Таблица 55
Средние индивидуальные нормативные требования по цинку
(мкг / кг массы тела / день) из рационов, различающихся содержанием цинка
биодоступность a
|
Возраст, лет |
Высокая биодоступность b |
Умеренная биодоступность c |
Низкая биодоступность d |
|
|
мкг / кг массы тела / сутки |
||||
|
Младенцы и дети |
||||
|
суки, 0-0.25 |
175 e |
457 ф |
1067 г |
|
|
Мужчины, 0-0,25 |
200 e |
514 ф |
1200 г |
|
|
0.25-0,5 |
79 e |
204 из |
477 г |
|
|
0,5–1 |
66 e |
– |
– |
|
|
0,5–1 |
186 |
311 |
621 |
|
|
1-3 |
138 |
230 |
459 |
|
|
3-6 |
114 |
190 |
380 |
|
|
6-10 |
90 |
149 |
299 |
|
|
Подростки |
||||
|
Женщины, 10-12 |
68 |
113 |
227 |
|
|
Мужчины, 10-12 |
80 |
133 |
267 |
|
|
суки, 12-15 |
64 |
107 |
215 |
|
|
Мужчины, 12-15 |
76 |
126 |
253 |
|
|
Женщины, 15-18 лет |
56 |
93 |
187 |
|
|
Мужчины, 15-18 лет |
61 |
102 |
205 |
|
|
Взрослые |
||||
|
Женщины, 18-60 лет + |
36 |
59 |
119 |
|
|
Мужчины, 18-60 лет + |
43 |
72 |
144 |
|
Источник: по материалам ФАО / МАГАТЭ / ВОЗ ( 32 ).
a Для получения информации о диетах см. Таблица 54
b Предполагаемая биодоступность пищевого цинка 50
процентов.
c Предполагаемая биодоступность пищевого цинка 30
процентов.
d Предполагаемая биодоступность пищевого цинка 15
процентов.
e Применяется исключительно для младенцев, вскармливаемых матерью.
только молоко, для которого биодоступность цинка не учитывается.
место хранения.
f Применимо к младенцам, частично вскармливаемым грудным молоком или
кормление кормом из коровьего молока с добавлением коровьего молока или молока с низким содержанием фитатов. Без надбавки
для хранения.
г Для младенцев, получающих богатые фитатами
Детская смесь на основе растительного белка с цельнозерновыми злаками или без них. Нет
припуск на хранение.
Верхний предел цинка
потребление
Было зарегистрировано всего несколько случаев острого отравления цинком.
сообщил. Признаками токсичности являются тошнота, рвота, диарея, лихорадка и
вялость и наблюдались после приема внутрь 4-8 г (60-120 ммоль) цинка.Долгосрочное потребление цинка выше, чем
требования могут, однако, взаимодействовать с метаболизмом других следов
элементы. Медь особенно чувствительна к высоким дозам цинка. Цинк
прием 50 мг / день (760 ммоль) влияет на медь
индексы статуса, такие как CuZn-супероксиддисмутаза в эритроцитах ( 37,
38 ). Наблюдались низкие уровни меди и церулоплазмина и анемия.
после повышенного потребления цинка 450-660 мг / день (6,9-10 ммоль / день) ( 39, 40 ).Изменения липидного состава сыворотки и иммунного ответа также наблюдались у
исследования добавок цинка ( 41, 42 ). Поскольку медь также имеет центральную
роль в иммунной защите, эти наблюдения требуют осторожности перед крупномасштабными
реализуются программы приема добавок цинка. Любые положительные эффекты цинка
добавки на рост или инфекционные заболевания могут быть замаскированы или
уравновешивается негативным воздействием на функции, связанные с медью.
Верхний уровень потребления цинка для взрослого мужчины установлен на уровне 45
мг / день (690 ммоль / день) и экстраполировано на другие
группы по отношению к основной скорости метаболизма.Для детей эта экстраполяция
означает верхний предел потребления 23-28 мг / день (350-430 ммоль / день), что близко к тому, что использовалось в некоторых
исследований добавок цинка. За исключением чрезмерного потребления некоторых видов
Из морепродуктов такое потребление вряд ли будет достигнуто при большинстве диет.
Посторонний цинк в воде из загрязненных колодцев и при гальванике.
посуда также может привести к высокому потреблению цинка.
Таблица 56
Рекомендуемое потребление питательных веществ (RNI) для пищевого цинка
(мг / день) для соблюдения нормативных требований к хранению из рационов, различающихся содержанием цинка.
биодоступность a
|
Возрастная группа |
Масса тела, кг |
Высокая биодоступность |
Умеренная биодоступность |
Низкая биодоступность |
|
Младенцы и дети |
||||
|
0-6 месяцев |
6 |
1.1 б |
2,8 в |
6,6 д |
|
7-12 месяцев |
9 |
0,8 б |
– |
– |
|
7-12 месяцев |
9 |
2.5 e |
4,1 |
8,4 |
|
1-3 года |
12 |
2,4 |
4,1 |
8,3 |
|
4-6 лет |
17 |
2.9 |
4,8 |
9,6 |
|
7-9 лет |
25 |
3,3 |
5,6 |
11,2 |
|
Подростки |
||||
|
Женщины, 10-18 лет, |
47 |
4.3 |
7,2 |
14,4 |
|
Мужчины, 10-18 лет |
49 |
5,1 |
8,6 |
17,1 |
|
Взрослые |
||||
|
Женщины, 19-65 лет |
55 |
3.0 |
4,9 |
9,8 |
|
Мужчины, 19-65 лет |
65 |
4,2 |
7,0 |
14,0 |
|
Женщины, 65+ лет |
55 |
3.0 |
4,9 |
9,8 |
|
Мужчины, 65+ лет |
65 |
4,2 |
7,0 |
14,0 |
|
Беременные |
||||
|
Первый триместр |
– |
3.4 |
5,5 |
11,0 |
|
Второй триместр |
– |
4,2 |
7,0 |
14,0 |
|
Третий триместр |
– |
6.0 |
10,0 |
20,0 |
|
Кормящие женщины |
||||
|
0-3 месяца |
– |
5,8 |
9.5 |
19,0 |
|
3-6 месяцев |
– |
5,3 |
8,8 |
17,5 |
|
6-12 месяцев |
– |
4,3 |
7.2 |
14,4 |
a Информацию о диетах см. В таблице .
54 . Если не указано иное, внутрииндивидуальные вариации цинка
Предполагается, что требования составляют 25 процентов. Данные о весе интерполированы из ФАО / ВОЗ,
1988 г. ( 36 ).
b Младенцы на грудном вскармливании, получающие материнское молоко
Только; предполагаемый коэффициент вариации (CV) 12,5 процента; предполагаемая доступность
80 процентов.
c Младенцы на искусственном вскармливании: умеренная биодоступность для
молочные смеси с добавлением сыворотки и для младенцев, частично вскармливаемых грудным молоком или даваемых
корма с низким содержанием фитатов с добавлением другого жидкого молока; предполагается CV 12,5
процентов.
d Младенцы, находящиеся на искусственном вскармливании; низкая биодоступность
применимо к смеси на основе растительного белка, богатой фитатами, с или без
цельнозерновые крупы; CV 12,5 процента.
e Не применимо к младенцам, потребляющим грудное молоко.
Только.
Адекватность потребления цинка по отношению к
в соответствии с оценками требований
Риск неадекватного потребления цинка детьми был снижен.
оценено с использованием предложенных оценок потребности в цинке ( 32 ) и
используя имеющиеся данные о составе пищевых продуктов и рационе питания в различных
части света ( 43 ). Для этой оценки предполагалось, что
Распределение потребностей в цинке является гауссовым с CV 15 процентов, и что
корреляция между потреблением и потребностью очень мала.Поглощение цинка из
диеты в Малави, Кении, Мексике и Гватемале оценивались в 15 процентов
на основе высокого мольного соотношения фитат-цинк ( 37-42 ) в этих диетах,
в то время как потребление 30 процентов предполагалось для диет в Гане, Гватемале,
Египет и Папуа-Новая Гвинея. Продукты из ферментированной кукурузы и маниоки
( kenkey , banku и gari ) в Гане, дрожжевой
хлеб на основе пшеницы в Египте и использование саго с низким содержанием фитатов в качестве
основной продукт в рационе Новой Гвинеи, как предполагалось, приводит к снижению
мольное соотношение фитат-цинк и лучшая доступность.При таком подходе 68-94
процент детей, по оценкам, находятся в группе риска дефицита цинка в
эти группы населения, за исключением Египта, где оценка составляла 36 процентов
( 43 ). Среднесуточное потребление цинка этими детьми составляло 3,7-6,9 мг.
(56-105 ммоль). Большая часть добавок цинка
исследования не предоставили данных о рационе питания, которые можно было бы использовать для определения
потребление цинка имеет решающее значение для эффектов роста. В недавнем исследовании, проведенном в Чили,
положительный эффект на увеличение роста у мальчиков после 14 месяцев приема добавок цинка
был отмечен ( 44 ).Потребление в группе плацебо в начале составляло 6,3.
± 1,3 мг / сут (96 ± 20 ммоль / сут) (n = 49).
Потому что только 15 процентов цинка, потребляемого чилийскими детьми, было получено
предполагалось, что наличие мясных продуктов относительно невелико.
Krebs et al ( 45 ) не наблюдали влияния цинка.
добавка по содержанию цинка в грудном молоке или по материнскому статусу группы
кормящим женщинам и сочли, что их потребление достаточно для поддержания адекватного уровня цинка
статус через 7 месяцев и более лактации.Среднее потребление цинка
женщины, не принимавшие добавки, составляли 13,0 ± 3,4 мг / день (199 ± 52 ммоль / день).
Эффективные гомеостатические механизмы поддержания цинка в организме
содержание при низком потреблении, которое легло в основу оценок физиологического
требований в отчете ФАО / МАГАТЭ / ВОЗ за 1996 год ( 32 ), а также
предполагаемое негативное влияние диеты с высоким содержанием фитатов на статус цинка, были подтверждены
в недавних экспериментальных исследованиях ( 10, 44, 46, 47, ). Уменьшение мочевыделения
и фекальные потери поддерживали нормальную концентрацию цинка в плазме в течение 5 недель в
11 человек при приеме 2.45 мг цинка / день (37 ммоль / день) или выше при диете с предположительно высоким
наличие ( 10 ). В аналогичном исследовании восполнения-истощения с участием 15 мужчин,
прием 4 мг / сутки (61 ммоль / сутки) из диеты с
молярное соотношение фитат-цинк 58 в течение 7 недель привело к снижению мочевыделения.
экскреция цинка от 0,52 ± 0,18 до 0,28 ± 0,15 мг / сут (7,9 ± 2,8
ммоль / сут до 4,3 ± 2,3 ммоль / сут) ( 46 ). Значительное снижение плазмы
Наблюдались концентрации цинка и изменения клеточного иммунного ответа.Влияние на иммунитет также наблюдалось при диете с ограничением цинка с высоким содержанием цинка.
с содержанием фитата (молярное соотношение примерно 20) потребляли пять молодых мужчин
волонтеры на 20-24 недели ( 14 ). Неоптимальный статус цинка также был
документально подтвержден у беременных женщин, соблюдающих диеты с высоким содержанием фитата и цинка.
(> 17) ( 47 ). Частые репродуктивные циклы и высокая распространенность малярии
казалось, способствовал ухудшению статуса цинка.
Заключение
В заключение, подход, использованный для получения среднего
индивидуальные потребности в цинке, использованные в отчете ФАО / МАГАТЭ / ВОЗ за 1996 г. (32) и
полученные оценки все еще кажутся действительными и полезными для оценки
адекватность потребления цинка в группах населения и для планирования диеты для определенных
группы населения.
Будущие исследования
Как уже указано в отчете ФАО / МАГАТЭ / ВОЗ за 1996 год
( 32 ), существует острая необходимость охарактеризовать ранний функциональный
последствия дефицита цинка и определить их связь с патологическими изменениями.
Эти знания особенно необходимы для понимания роли цинка.
недостаточность этиологии задержки роста и нарушения иммунокомпетентности.
Для лучшего понимания взаимосвязи диеты
и поставки цинка, необходимы дальнейшие исследования для тщательной оценки
наличие цинка из рациона, типичного для развивающихся стран.Исследование
должен включать оценку эффекта доступности принятия реалистичных
и принятые в культуре методы приготовления пищи, такие как ферментация,
проращивание, замачивание и включение недорогого и доступного животного белка
источники в диетах, основанных на растительной пище.
Список литературы
1. Hambidge, K.M. 1987. Цинк. В: Микроэлементы в
питание человека и животных. Mertz, W., ed. 5-е, т. 1. С. 1-137. Орландо,
Флорида, Academic Press, Inc.
2. Shankar, A.H. & Prasad A.S. 1998. Цинк и
иммунная функция: биологическая основа измененной устойчивости к инфекции. Am.
J. Clin. Nutr., 68 (доп.): 447S-463S
3. Sandström, B. 1997. Биодоступность цинка.
Eur. J. Clin. Nutr., 51 (дополнение 1): S17-S19
4. King, J.C. & Turnlund, J.R. 1989. Человеческий цинк
требования. В: Цинк в биологии человека . Миллс К.Кормили. стр.335-350. Девон
, Великобритания, Springer-Verlag.
5. Лукаски Х.С., Болончук У., Клевай Л.М., Милн,
Д. & Sandstead, H.H. 1984. Изменения содержания цинка в плазме после
упражнения у мужчин, соблюдающих диету с низким содержанием цинка. Am. J. Physiol. , 247:
Е88-93.
6. Милн, Д. Б., Кэнфилд, В. К., Галлахер, С. К., Хант, Дж. Р.
И Клевай, Л.М. 1987. Метаболизм этанола у женщин в постменопаузе, получавших
диета маргинальная по цинку. Am.J. Clin. Nutr., 46: 688-93.
7. Baer, M.J. & King, J.C. 1984. Уровни цинка в тканях.
и выведение цинка во время экспериментального истощения цинка у молодых мужчин. Am. Дж.
Clin. Nutr., 39: 556-70.
8. Hess, F.M., King, J.C. & Margen, S. 1977. Цинк.
экскреция у молодых женщин, принимающих низкие дозы цинка и пероральные контрацептивы.
J. Nutr., 107: 1610-20.
9. Милн, Д. Б., Кэнфилд, В. К., Махалко, Дж.Р. &
Sandstead, H.H. 1983. Влияние пищевого цинка на потерю поверхностной массы тела
цинк: влияние на оценку удержания цинка балансовым методом. Am. J. Clin.
Nutr., 38: 181-6.
10. Джонсон, П.Э., Хант, С.Д., Милн, Д.Б. И Маллен,
Л.К. 1993. Гомеостатический контроль метаболизма цинка у мужчин: экскреция цинка и
баланс у мужчин, питающихся диетами с низким содержанием цинка. Am. J. Clin. Нутр., 57:
557-565.
11. Эджет П.J. & Favier A. 1993. Цинк. Внутр. Дж.
Vit. Nutr. Res., 63: 247-316.
12. Гольденберг, Р.Л., Тамура, Т., Неггерс, Ю., Медь,
Р.Л., Джонстон, К.Е., ДюБард, М. & Hauth, J.C. 1995. Эффект
добавки цинка на исход беременности. JAMA , 274:
463-468
13. Браун К., Пеерсон Дж. М. и Аллен Л. Х. 1998.
Влияние добавок цинка на рост детей. В: Роль трассировки
элементы для укрепления здоровья и профилактики заболеваний. Сандстрём, Б.,
Уолтер П., ред. Bibliotheca Nutritio et Dieta , Базель: Каргер; 54:
76-83.
14. Beck, F.W.J., Prasad, A.S. И Каплан Дж. 1997.
Изменения продукции цитокинов и субпопуляций Т-клеток в экспериментальных
индуцированный дефицит цинка у людей. Am. J. Physiol. 272: E1002-7.
15. Сандстрём, Б. Фэйрвезер-Тейт, С., Харрелл, Р.
И Ван Доккум, W. 1993. Методы изучения минералов и микроэлементов.
абсорбция у людей с использованием стабильных изотопов. Nutr. Res. Revs. , 6:
71-95.
16. Wastney, M.E., Gökmen, I.G., Aarmodt, R.L.,
Рамбл, У.Ф., Гордон, Г. & Хенкин Р.И. 1991. Кинетический анализ.
метаболизма цинка у человека и одновременное введение 65 Zn
и 70 Zn. Am. J. Physiol. , 260: R134-41.
17. Фэйрвезер-Тейт, С.Дж., Джексон, М.Дж., Фокс, Т.Э.,
Wharf, S.G., Eagles, J. & Groghan, P.C. 1993.Измерение
обменные пулы цинка с использованием стабильного изотопа 70 Zn. руб. Дж.
Nutr., 70: 221-34.
18. Миллер, Л.В., Хамбидж, К.М., Нааке, В.Л., Хонг, З.,
Весткотт, Дж. Л., Феннесси, П. В. 1994. Размер цинковых бассейнов,
быстро обмениваются с плазменным цинком у людей: альтернативные методы
измерение и отношение к потреблению цинка с пищей. J. Nutr., 124:
268-76.
19. Сиан, Л., Мингян, X., Миллер, Л.В., Тонг, Л., Кребс,
Н.Ф. И Хамбидж, К. 1996. Всасывание цинка и кишечные потери
эндогенный цинк у молодых китаянок с минимальным потреблением цинка. Am. Дж.
Clin. Nutr., 63: 348-53.
20. Сандстрём Б. 1989. Диета и цинк.
поставка. В: Цинк в биологии человека . Миллс К.Ф. изд. п. 350-363. Девон,
Великобритания, Springer-Verlag.
21. Sandström, B. & Lönnerdal, B. 1989.
Промоторы и антагонисты абсорбции цинка. В: Цинк в биологии человека .
Миллс К.Ф. изд. с.57-78. Девон, Великобритания, Springer-Verlag.
22. Петтерсон, Д., Сандстрём, Б. и Седерблад,
Å. 1994. Поглощение цинка люпином (Lupinus angustifolius)
— продукты на основе. руб. J. Nutr., 72: 865-71.
23. Кнудсен, Э., Йенсен, М., Солгаард, П., Соренсен,
S.S. & Sandström, B. 1995. Поглощение цинка оценивается фекалиями.
мониторинг стабильных изотопов цинка подтвержден сравнением с данными для всего тела
удержание радиоизотопов цинка в организме человека. J. Nutr., 125:
1274-1282.
24. Хант, Дж. Р., Маттис, Л. А. и Джонсон, Л. К. 1998.
Абсорбция цинка, минеральный баланс и липиды крови у женщин с контролируемым потреблением.
лактоовегетарианская и всеядная диеты на 8 недель. Am. J. Clin. Нутр., 67:
421-30.
25. Хант, Дж. Р., Галлахер, С. К., Джонсон, Л. К. &
Ликкен, Г. 1995. Диеты с высоким и низким содержанием мяса: влияние на усвоение цинка,
статус железа, а также кальций, медь, железо, магний, марганец, азот,
баланс фосфора и цинка у женщин в постменопаузе. Am. J. Clin. Нутр.,
62: 621-32.
26. Нэверт, Б., Сандстрём, Б. & Седерблад,
Å. 1985. Снижение содержания фитатов в отрубях путем закваски в
хлеб и его влияние на усвоение цинка человеком. руб. J. Nutr., 53:
47-53.
27. Sandström, B. & Sandberg, A.S. 1992.
Ингибирующее действие изолированных фосфатов инозитола на абсорбцию цинка у людей.
J Trace Elem Electrolyte Health Dis., 6: 99-103.
28. Гибсон, Р.С., Йеудалл, Ф., Дрост, Н., Мтитимуни, Б.
& Cullinan, T. 1998. Диетические вмешательства для предотвращения дефицита цинка.
Am. J. Clin. Нутрь., 68 (доп.): 484С-487С.
29. Симмер, К., Ханум, С., Карлссон, Л. и Томпсон,
R.P.H. 1988. Нутритивная реабилитация в Бангладеш — важность
цинк. Am. J. Clin. Nutr., 47: 1036-40.
30. Черный, М.М. 1998. Дефицит цинка и ребенок.
разработка. Am. J. Clin. Nutr., 68 (доп.): 464S-469S
31. Колфилд, Л.Е., Завалета, Н., Шанкар, А.Х. и
Merialdi, M. 1998. Возможный вклад материнского цинка.
во время беременности для выживания матери и ребенка. Am. J. Clin. Nutr.,
68 (доп.): 499S-508S
32. АО / МАГАТЭ / ВОЗ. 1996. Микроэлементы в организме человека.
питание и здоровье. Женева, Всемирная организация здравоохранения.
33. Тейлор, К.М., Бэкон, Дж. Р., Аггетт, П. Дж. И Бремнер,
I. 1991. Гомеостатическая регуляция всасывания цинка и эндогенных потерь в организме.
мужчины, лишенные цинка. Am. J. Clin. Nutr., 53: 755-63.
34. Swanson, C.A. & King, J.C. 1987. Цинк и
исход беременности. Am. J. Clin. Nutr., 46: 763-771.
35. Всемирная организация здравоохранения. 1998. Дополнительный
кормление маленьких детей в развивающихся странах: обзор современных
научные знания.Женева, Всемирная организация здравоохранения.
36. ФАО / ВОЗ / УООН. 1985. Потребности в энергии и белке.
Отчет совместной консультации экспертов ФАО / ВОЗ / УООН. Серия технических отчетов 724.
Женева, Всемирная организация здравоохранения.
37. Fischer, P.W.F., Giroux, A. & L’Abbé,
M.R. 1984. Влияние добавок цинка на статус меди у взрослого мужчины.
Am. J. Clin. Nutr., 40: 743-6.
38. Ядрик М.К., Кенни М.А. и Винтерфельдт Э.А.
1989. Состояние железа, меди и цинка: реакция на добавки с цинком или
цинк и железо у взрослых самок. Am. J. Clin. Nutr., , 49: 145-50.
39. Паттерсон, У.П., Винкельманн, М. и Перри, М.С.
1985. Цинк-индуцированный дефицит меди: мегаминеральная сидеробластная анемия. Ann.
Internal Med. . 103: 385-6.
40. Портер, К.Г., Макмастер, Д., Элмс, М.Э. и Лав,
A.H.G. 1977. Анемия и низкое содержание меди в сыворотке крови во время цинковой терапии. Ланцет ,
II: 774.
41. Хупер П.Л., Висконти Л., Гарри П.Дж. и Джонсон,
G.E. 1980. Цинк снижает уровень холестерина липопротеинов высокой плотности.
JAMA , 244: 1960-2.
42. Chandra, R.K. 1984. Чрезмерное потребление цинка.
ухудшает иммунные реакции. JAMA , 252: 1443-6.
43. Гибсон, Р.С. И Фергюсон, Э. 1998. Оценка
диетического цинка в популяции. Am.J. Clin. Nutr., 68 (доп.):
430S-434S
44. Руз, М., Кастильо-Дюран, К., Лара, X., Кодосео, Дж.,
Rebolledo, A. & Atalah, E. 1997. 14-месячное испытание приема цинка в
внешне здоровые чилийские дошкольники. Am. J. Clin. Нутр., 66:
1406-13.
45. Кребс, Н.Ф., Рейдингер, К.Дж., Хартли, С., Робертсон,
A.D. & Hambidge, K.M. 1995. Добавки цинка в период лактации:
влияние на материнский статус и концентрацию цинка в молоке. Am. J. Clin.
Nutr., 61: 1030-6.
46. Руз, М., Каван, К.Р., Беттгер, В.Дж. и Гибсон,
Р.С. 1992.