Леденцы ‘Витаминные’, повышение иммунитета, нормализация обмена веществ, 50 г
Леденцы «Витаминные», повышение иммунитета, нормализация обмена веществ, 50 г
Бета-каротин (провитамин А) рекомендован для постоянного применения всем возрастным группам. Основные функции в организме:
- эффективный антиоксидант;
- повышает иммунитет;
- способствует нормализации обмена веществ;
- способствует поддержанию нормального состояния органов зрения, слизистых оболочек и
- кожных покровов организма.
Способ применения: по 3-4 леденца 3 раза в день.
Состав: сахар, патока, бета-каротин (Бетавитон), аскорбиновая кислота E300, лимонная кислота E330, ароматизатор натуральный или идентичный натуральному.
- Вес: 56 г
- Тип индивидуальной упаковки: Без упаковки
- Длина упаковки: 15
- Высота упаковки: 2.5
- Ширина упаковки: 13
- Объем упаковки: 0.488
- Объем продукта: 0.
 1989
1989 - Объем бокса: 9.945
- Материал: См. на упаковке
- Вес нетто, г: 50
- РосЗакуп: Да
- Температурное ограничение: Нет
- Назначение: Для зрения, Для иммунной системы, Антиоксиданты
- Форма выпуска: Леденцы
- ШК WB: 6900071417094
- Производитель: Вулкан
- Страна производитель: Россия
Отправка заказа будет осуществляться из города Костанай.
Стоимость доставки оплачивается при получении заказа в пункте выдачи вашего города.
Для выставления счета юр. лицам и ИП просим связаться с нашим менеджером
Ваш заказ будет готов к отправке в течение шести рабочих дней после оплаты.
Доставка Вашего заказа возможна следующими способами:
- Казпочта
- Транспортной компанией «КИТ»
- Пункты выдачи компании «СДЭК»
- Экспресс-доставкой «СДЭК»
- Транспортной компанией «Авис»
Хрупкие товары (стеклянные товары, зеркала и прочее) отправляются в жесткой упаковке, что влечет за собой удорожание доставки
Страница не найдена |
Страница не найдена |
404. Страница не найдена
Страница не найдена
Архив за месяц
ПнВтСрЧтПтСбВс
13141516171819
20212223242526
2728293031
12
12
1
3031
12
15161718192021
25262728293031
123
45678910
12
17181920212223
31
2728293031
1
1234
567891011
12
891011121314
11121314151617
28293031
1234
12
12345
6789101112
567891011
12131415161718
19202122232425
3456789
17181920212223
24252627282930
12345
13141516171819
20212223242526
2728293031
15161718192021
22232425262728
2930
Архивы
Метки
Настройки
для слабовидящих
Кафедра наследственных болезней обмена веществ
Ученая степень: доктор медицинских наук.
Преподаваемые дисциплины:
- клинико-биохимическое обоснование применения диетотерапии при наследственных болезнях обмена веществ, выявляемых путем неонатального и селективного скрининга
- современные подходы к организации диетотерапии НБО
- перспективы применения новых методов терапии НБО у детей и взрослых
Образование
Уровень образования: высшее
Окончила педиатрический факультет II Московского ордена В.И.Ленина государственного медицинского института им.Н.И.Пирогова в 1984 году по специальности «Педиатрия». С 1984 по 1987 гг. проходила клиническую ординатуру при Научно –исследовательском институте педиатрии Академии медицинских наук СССР по специальности «Педиатрия», по окончании которой работала в отделении реанимации и интенсивной терапии новорожденных детей ВНИИЦ охраны матери и ребенка. В 1995 г окончила аспирантуру в Институте питания АМН СССР по специальности «Педиатрия». Защитила диссертацию на соискание ученой степени кандидата медицинских наук на тему: «Обмен кальция и его регуляция у детей, больных фенилкетонурией, на фоне диетотерапии».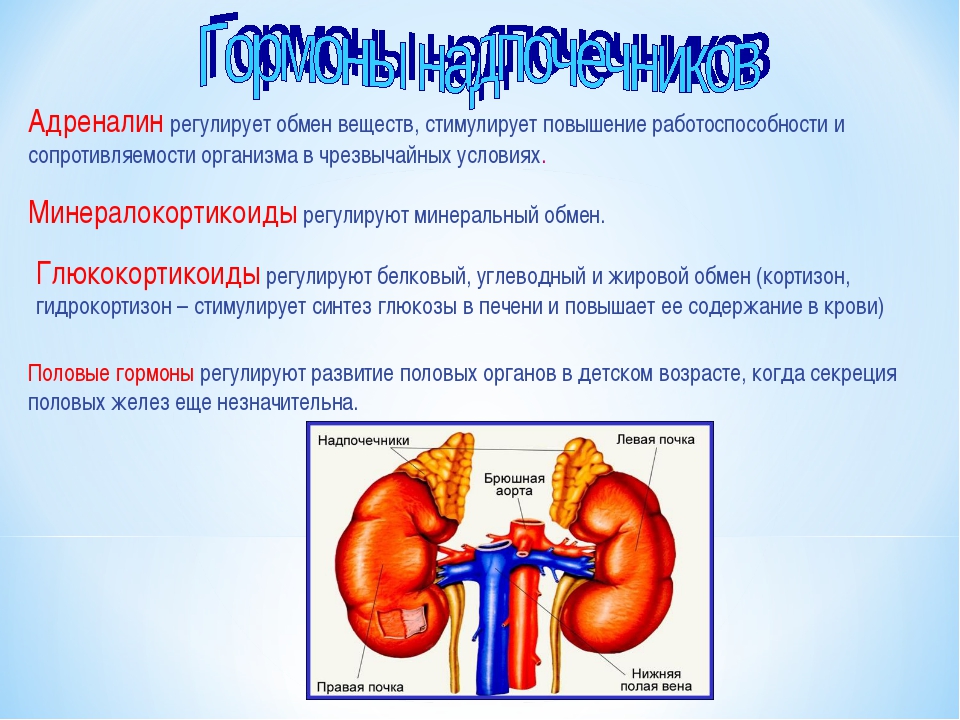 С 2001 года работает в лаборатории питания здорового и больного ребенка ФГАУ НМИЦ здоровья детей МЗ РФ. В 2016 году защитила диссертацию на соискание ученой степени доктора медицинских наук на тему «Диетотерапия болезней обмена веществ, выявляемых по неонатальному скринингу». Совмещает научную деятельность с лечебно-консультативной и педагогической работой.
С 2001 года работает в лаборатории питания здорового и больного ребенка ФГАУ НМИЦ здоровья детей МЗ РФ. В 2016 году защитила диссертацию на соискание ученой степени доктора медицинских наук на тему «Диетотерапия болезней обмена веществ, выявляемых по неонатальному скринингу». Совмещает научную деятельность с лечебно-консультативной и педагогической работой.
Повышение квалификации и (или) профессиональная переподготовка
Имеет действующие сертификаты по специальностям:
- «Педиатрия» регистрационный номер 02-С/66.6 от 06.02.2018г.
- «Диетология» регистрационный номер 29186 от 09.06 2017
- «Гастроэнтерология» диплом о профессиональной переподготовке 772407738602 от 31.05.2019 г.
Повышение квалификации за последние 5 лет:
- «Педиатрия»; удостоверение 180001339621 от 06.02.2018г.
- «Диетология», удостоверение 772405498996 о 09.06.2017г.
- «Гастроэнтерология», регистрационный номер 6367 о 31.05.
 2019 г.
2019 г. - «Инновационные направления в информационных технологиях», удостоверение 180001917152 от 18.12.2019 г.
- «Педагогика высшей школы: современные аспекты медицинского образования» 180001690077 от 02.03.2019 г.
Стаж: общий стаж работы 35 лет; стаж работы по специальности «Педиатрия» 35 лет.
Научные интересы
Сфера научных интересов: клиническое питание при хирургической и соматической патологии, диагностика и лечение наследственных болезней обмена веществ, синдрома короткой кишки, синдрома мальабсорбции, значение диетотерапии в лечении синдромальной патологии, профилактика и диетотерапия алиментарно-зависимых состояний раннего возраста, современные методы оценки нутритивного статуса у пациентов с различной патологией, способы коррекции нутритивной недостаточности, ожирения и дефицитных состояний.
В соответствии с базой РИНЦ является автором 178 публикаций в рецензируемых научных изданиях, индекс Хирша 13.
Дополнительная информация: член Российской Ассоциации медицинских генетиков, член Европейского общества педиатров, гастроэнтерологов, гепатологов, нутрициологов (ESPGHAN).
Как ускорить обмен веществ
Обменом веществ (метаболизмом) называют процесс переработки калорий пищи в энергию, благодаря которой организм поддерживает жизнь. Метаболизм дает силы, чтобы расти, размножаться, отвечать на влияние окружающей среды и сохранять структуры тела.
Но понятие «скорость метаболизма» – условное, поскольку в научной среде этого термина нет. Не существует единиц, чтобы измерить скорость обмена веществ, а значит, нельзя точно вывести норму и определить, медленный ли у человека метаболизм или быстрый.
Понятие «скорость метаболизма» используют для обозначения интенсивности обменных процессов, основываясь на внешних признаках, которые связаны с либо полнотой, либо с невозможностью набрать вес.
Условное понятие «скорости метаболизма» делят на 3 условные же категории – быстрый, нормальный или медленный. А подразумевают под этим следующее:
А подразумевают под этим следующее:
- Быстрый (ускоренный) метаболизм – это такая интенсивность обменных процессов, когда питательные вещества мгновенно перерабатываются в энергию. Человек при этом не может набрать вес, сколько бы ни старался, и даже если он очень много ест, ни жира, ни мышц не вырастет;
- Нормальный метаболизм же означает естественную интенсивность обменных процессов, когда никаких нарушений нет. А человек при этом не набирает вес, если не переедает и допускает умеренные физические нагрузки;
- Медленным метаболизмом называют нарушенный обмен веществ, при котором организм усиленно запасает подкожный жир, готовясь к дефициту калорий. Обычно такое случается после того, как человек пережил резкое повышение уровня физических нагрузок при сокращении рациона. Но может повлиять и гормональный дисбаланс.
Дополнительными признаками нарушения метаболизма в сторону снижения его интенсивности считаются: вялость, быстрая утомляемость, замедление реакций и спад интеллектуальных способностей.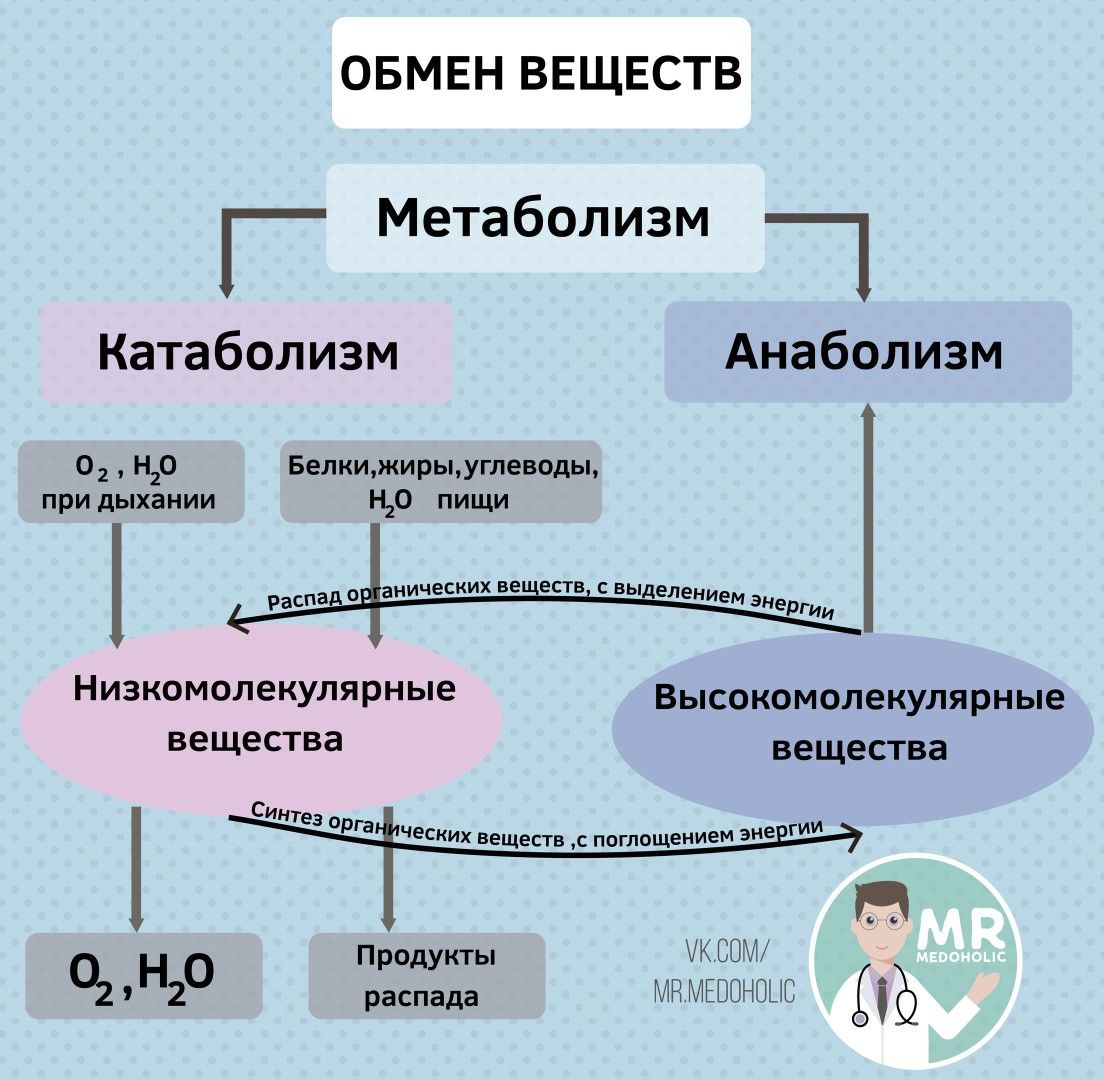 Серьезное влияние на обменные процессы оказывают гормоны щитовидной железы: когда их много, повышается интенсивность обмена белков, жиров и углеводов, увеличивается поглощение кислорода в тканях.
Серьезное влияние на обменные процессы оказывают гормоны щитовидной железы: когда их много, повышается интенсивность обмена белков, жиров и углеводов, увеличивается поглощение кислорода в тканях.
Ускорить или нормализовать метаболизм можно двумя способами: первый – это регулярные занятия спортом с питанием, усиленным на 15, а то и 20%, второй – диета. Каждый из них применим в домашних условиях и способствует похудению.
Первый способ основан на том, чтобы перестроить обменные процессы организма так, чтобы он готовил тело к предстоящим нагрузкам, запасая энергию в мышцах, а не в жировой прослойке. Дело тут в том, что чем больше у человека мышечная масса, тем больше расходуется калорий на ее поддержание, а в жир отложить уже нечего.
Как лечить нарушение обмена веществ?
« Назад
04.09.2019 00:00
Обмен веществ во многом определяет состояние нашего здоровья. Замедление обмена веществ ведет к постепенному увеличению веса, ухудшению состояния кожи, волос и общего самочувствия.
Замедление обмена веществ ведет к постепенному увеличению веса, ухудшению состояния кожи, волос и общего самочувствия.
Ухудшение обмена веществ нередко связано с неправильным питанием, поэтому в первую очередь следует пересмотреть свой рацион. Все прекрасно знают о пользе клетчатки, которая очищает наш кишечник и выводит токсины. Особенно полезны в этом плане отруби, морковь и свекла.
А вот большое количество пищи животного происхождения приводит к нарушениям обменных процессов и ожирению, потому что организм тяжелее и дольше справляется с такой пищей. Чтобы дать организму восстановиться, иногда надо проводить разгрузочные дни.
Как лечить нарушение обмена веществ народными методами?
1. Облепиховое масло.
Регуляцию обмена веществ хорошо восстанавливает облепиховое масло. Кроме этого оно имеет противоопухолевые свойства. Чтобы приготовить масло из ягод облепихи, их надо промыть и просушить. После этого ягоды надо подавить, насыпать в емкость и залить растительным маслом, чтобы оно немного покрыло ягоды. Оставьте настояться в темном месте одну неделю при периодическом перемешивании. Готовый настой отжать, процедить и оставить еще настояться до прозрачности. Затем масло слить в емкость с непрозрачным стеклом, держать в холодильнике. Перед употреблением облепиховое масло разводят растительным маслом в пропорции 1:5.
Оставьте настояться в темном месте одну неделю при периодическом перемешивании. Готовый настой отжать, процедить и оставить еще настояться до прозрачности. Затем масло слить в емкость с непрозрачным стеклом, держать в холодильнике. Перед употреблением облепиховое масло разводят растительным маслом в пропорции 1:5.
2. Йод.
Йод является обязательным условием для нормального протекания обменных процессов. Так как во многих районах нашей страны йода не хватает в продуктах, то следует добавлять в питание морепродукты и морскую капусту, хурму и редьку. А пищу присаливать йодированной солью.
3. Лечебное голодание.
Обмен веществ хорошо восстанавливается после проведения процедур очищения. Сюда относится в первую очередь лечебное голодание, во время которого организм вынужден начать переваривать собственные ткани. Однако в первую очередь он начнет использовать самое ненужное, куда входят отложения солей, лишний жир, кисты, жировики, чужеродные патологические новообразования. Такой процесс начинается от того, что усиливается ферментативная и энзимная активность лейкоцитов.
Такой процесс начинается от того, что усиливается ферментативная и энзимная активность лейкоцитов.
К примеру, голодание длительностью в 36 ч увеличивает их активность в 3 раза, чего уже достаточно для очищения организма. Более длительные голодовки приводят к полному обновлению и омоложению организма, а также к усилению обмена веществ. После лечебного голодания первый месяц наблюдается улучшение обмена веществ на 5-6 % и лучшее усвоение пищи. Это объясняется отдыхом во время голодания всех органов, восстановлением поврежденных структур и избавлением от шлаков.
4. Рецепт тибетских лекарей.
Этот сбор улучшает обмен веществ, приводит к очищению организма и омоложению. Для него необходимо по 100 г травы зверобоя продырявленного, березовых почек, бессмертника песчаного, цветков ромашки аптечной.
Чтобы его приготовить надо все составляющие измельчить с помощью мясорубки. Затем отделить 1 ст. л. сбора и залить вечером пол-литром кипятка, настоять 20 минут и процедить. Пить 1 стакан настоя перед сном с добавлением 1 ч. л. меда. После выпитого настоя нельзя ничего пить и есть. А утром следует немного подогреть стакан оставшегося настоя и выпить натощак за полчаса до еды с 1 ч. л. меда. Пить отвар до окончания первоначальных компонентов. Курс можно повторять раз в 5 лет.
Пить 1 стакан настоя перед сном с добавлением 1 ч. л. меда. После выпитого настоя нельзя ничего пить и есть. А утром следует немного подогреть стакан оставшегося настоя и выпить натощак за полчаса до еды с 1 ч. л. меда. Пить отвар до окончания первоначальных компонентов. Курс можно повторять раз в 5 лет.
Как видите, рекомендации по улучшению обмена веществ, несложные. Главное, как считает Здоровая жизнь, желать помочь своему организму быть здоровым.
Источник
Диетология в Краснодаре официальный сайт центра
Лечебное воздействие осуществляется с помощью диетотерапии (индивидуально разработанного режима питания) с использованием методов:
- физиотерапии,
- гомеопатии,
- иглорефлексотерапии,
- биоимпедансометрия.
Комплексный метод лечения, позволяющего внести регуляцию и равновесие в работу всех систем организма.
С помощью акупунктуры (иглотерапии) достигается:
- улучшение психо-эмоционального состояния и нормализация пищевого поведения,
- снижение аппетита,
- повышение насыщаемости,
- активизация основного обмена веществ,
- улучшение кровообращения.

Применяются различные варианты иглотерапии – поверхностная рефлексотерапия и фармакопунктура (в косметологической практике называтся — «мезотерапия»).
При выполнении поверхностной рефлексотерапии с помощью специального многоигольчатого валика стимулируется рецепторная зона в области локальной зоны жировых отложений, что способствует улучшению микроциркуляции и обменных процессов в данной зоне. Процедура дополняется аппликацией термоактивных гелей.
Метод фармакопунктуры (медицинская «мезотерапия») заключается во введении с помощью тонкоигольного шприца в точки акупунктуры определенной области тела специальных лекарственных смесей аллопатического или гомеопатического происхождения.
Также применяется озонотерапия и физиотерапевтическое воздействие.
Озонотерапия – внутривенное или внутритканевое введение озоно-кислородной смеси позволяет активизировать процессы окисления жиров в организме.
Сочетание различных физиотерапевтических методов (вакуумный массаж, компьютерная миостимуляция, обертывания) обеспечивают лифтинговый и лимфодренажный эффекты, что имеет важное значение в коррекции фигуры. Достигается стойкое уменьшение объемов за счет жировой ткани.
Достигается стойкое уменьшение объемов за счет жировой ткани.
Результат снижения веса находится в физиологических рамках – 8-12 кг за 2 месяца. Это позволяет похудеть за счет жировой ткани, а не в результате временного обезвоживания организма.
Диетологические рекомендации учитывают пожелания пациента. Нет ограничения в потреблении воды и времени последнего приема пищи.
Комплексная лечебная программа позволяет нормализовать обмен веществ, снизить избыточное количество жировой ткани в организме и обеспечивает результат, который можно назвать эффектом «бесхирургической липосакции».
Формула метаболизма — расчет базового уровня метаболизма
Постоянно пользуясь термином «метаболизм» — мы мало знаем о нем. Не о термине, конечно — об обмене веществ.
Оказывается, даже ученые мужи не знают о метаболизме всего. Но зато создали формулу, по которой можно вычислить собственный базовый уровень метаболизма. И это поможет нам худеть осознанно.
Мечтая – или приняв решение – похудеть, что мы начинаем делать? Слушать знающих приятельниц и читать соответствующие тексты. В которых твердят: одно из главных условий сброса веса — ускорить (повысить) обмен веществ (метаболизм).
В которых твердят: одно из главных условий сброса веса — ускорить (повысить) обмен веществ (метаболизм).
Ничего не понимая ни в метаболизме, ни в способах его ускорения, мы, тем не менее, часто «покупаемся» на наукообразные рассуждения рекламы и покупаем то или иное средство (таблетки, капсулы) или способ (чаще всего диеты) повысить этот самый обмен веществ.
Давайте попробуем, насколько это возможно для неспециалистов, разобраться, что же такое метаболизм, можно ли его ускорить и как это сделать.
Для этого придется забыть о рекламе и почитать кое-что более серьезное. А почитав – огорчиться. Потому что становится ясно, что наука сегодня не имеет полного знания о том, что происходит в нашем организме. В том числе – нет ясности и с метаболизмом.
В нашем организме непрерывно происходят тысячи биохимических реакций, поддерживающих его жизнеспособность и жизнедеятельность. Отдельные реакции, их взаимосвязи и взаимозависимости особенно пристально изучаются последние полтора столетия.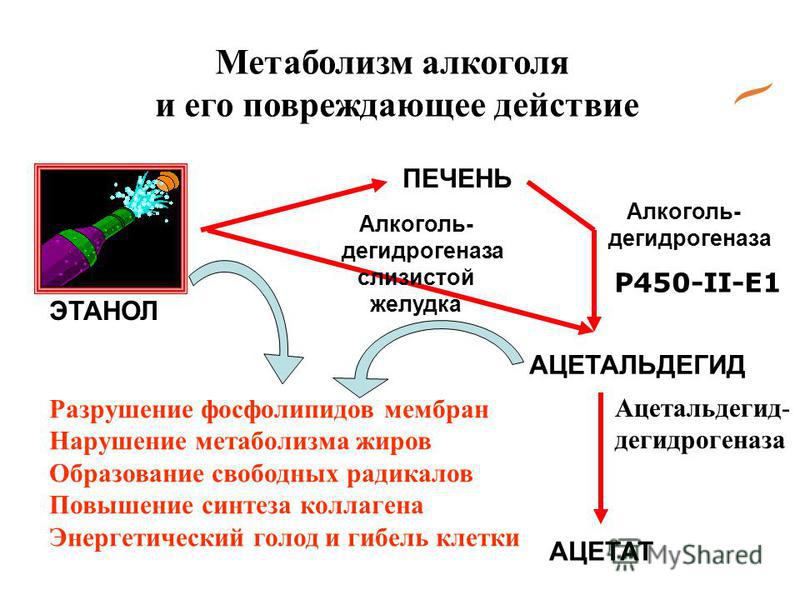 Появилась новая наука – биохимия, проводились и проводятся тысячи исследований, накапливается информация.
Появилась новая наука – биохимия, проводились и проводятся тысячи исследований, накапливается информация.
Обработка всей накопленной информации позволила ученым из Калифорнийского университета несколько лет назад вплотную подойти к решению задачи: с помощью новой отрасли знания – системной биологии – создать комплексную модель метаболизма человека. Она пока несовершенна, в общей картине обмена веществ есть еще не проясненные детали.
Но нам некогда ждать, пока ученые разберутся досконально. Мы хотим понять, что лично нам – здесь и сейчас – надо делать, чтобы ускорить обмен, чтобы избавиться от лишнего веса.
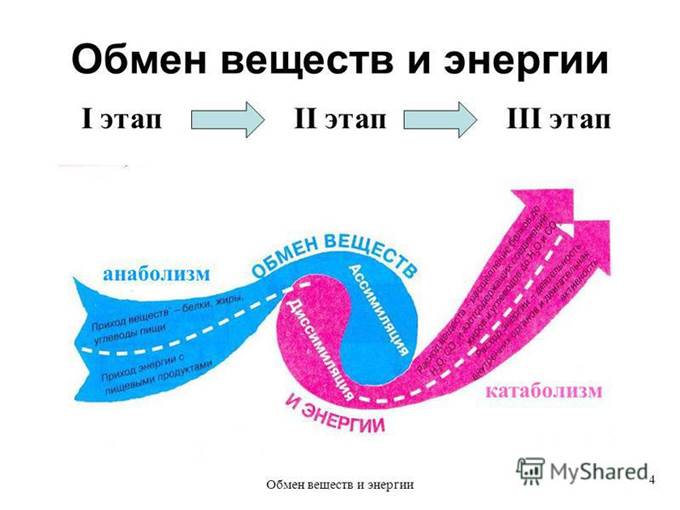
Начнем с определений. Греческое слово μεταβολ обозначает «превращение», «изменение». «Метаболизм» (обмен веществ) в современном понимании — это целый комплекс множества взаимозависимых биохимических и энергетических процессов, обеспечивающих использование пищи и прочих поступлений в наш организм, их переработку для нужд организма и вывод продуктов распада.
Обмен веществ состоит из двух противоположных, одновременно протекающих процессов — анаболизма и катаболизма.
Анаболизм – объединяет все те реакции, которые обеспечивают синтез необходимых организму сложных органических веществ из более простых, и проходит он с поглощением энергии.
Катаболизм – включает реакции, которые обеспечивают расщепление органических веществ и выведение из организма продуктов распада. Проходит с выделением энергии.
Те, кто понимает в этих процессах, утверждают: «повысить» метаболизм – что действительно ведет к избавлению от лишнего веса – это значит ускорить катаболизм. То есть тот процесс, в ходе которого органические вещества в организме расщепляются и выводятся.
Самый простой и верный путь сделать это – вы не поверите – знаком нам давно. Он заключается в том, чтобы отклеить «пятую точку» от любимого дивана-кресла-стула и заставить себя двигаться. Хотя бы понемногу. Но каждый день. Стать физически активными, чтобы больше тратить калорий.
Хотя бы понемногу. Но каждый день. Стать физически активными, чтобы больше тратить калорий.
Но ведь это надо себя заставить! Нам же почему-то кажется более легким и надежным получать меньше калорий – и мы садимся на очередную низкокалорийную диету. Не зная или забыв о том, что это и вредно для здоровья, и — в абсолютном большинстве случаев — бесполезно, ведь мало кто способен выдерживать длительные многомесячные диеты. В результате начинается обратный стремительный набор веса, потому что организм, напуганный голодным рационом, ускоренно запасается жиром. И отдавать свои запасы больше не собирается.
Как только мы снижаем уровень потребления пищи ниже той планки, которая необходима нашему организму для нормальной жизнедеятельности – обмен веществ дает обратный ход: он замедляется.
Такая вот печальная зависимость. Есть о чем подумать, не так ли?
Но как понять, каков у нас уровень метаболизма?
Вряд ли все мы побежим в немногочисленные специальные лаборатории или клиники, где проведут соответствующий достаточно сложный тест на «скорость основного обмена». За деньги, естественно.
За деньги, естественно.
Давайте попробуем обойтись «домашними» расчетами. Потому что такая формула существует. Формула расчета базового уровня метаболизма – RMR. Она не может претендовать на особую точность, но способна стать поворотным пунктом в понимании того, куда и как надо двигаться. Выглядит эта формула так.
RMR = 655 + (9,6 х вес) + (1,8 х рост) — (4,7 х возраст в годах).
Предположим, что ваш вес 70 кг, рост – 160 см и возраст – 30 лет. Подставляем в формулу значения.
RMR= 655 + (9,6х70) + (1,8х160) – (4,7х30) = 1474 (ккал)
Теперь находим свой коэффициент активности и умножаем на него полученную цифру.
Коэффициенты следующие.
- 1,2 при сидячем образе жизни
- 1,375 при легкой активности (легкие тренировки 1—3 раза в неделю)
- 1,55 при умеренной активности (интенсивные тренировки 3-5 раз в неделю)
- 1,725 при повышенной активности (интенсивные тренировки 6-7 раз в неделю)
- 1,9 при экстра-активности (сверхинтенсивные тренировки или тяжелая физическая работа)
Предположим, что вы весь день сидите у компьютера и в спортзале не бываете – ваш коэффициент – 1,2 . Умножаем на него RMR. 1474х1,2 = 1768.8. Немногим более 1700 ккал – примерно столько требуется вашему организму, чтобы он нормально работал. Ему этого достаточно.
Умножаем на него RMR. 1474х1,2 = 1768.8. Немногим более 1700 ккал – примерно столько требуется вашему организму, чтобы он нормально работал. Ему этого достаточно.
А теперь, не лукавя, посчитаем, каким количеством ккал в день вы свой организм насыщаете. Таблицы калорийности есть в интернете. Подсчитали? Теперь отнимите ту цифру, что мы вывели раньше. Feel the difference, как любит выражаться реклама. Чувствуете разницу? Ту самую, которая оседает в «проблемных местах»?
Информация к размышлению: 7000 ккал этой самой неизрасходованной разницы «дарят» нам 1000 г жира.
Нет, конечно, каждый из нас волен по-прежнему с надеждой читать рекламу, уверенно обещающую средство или диету, которая поможет похудеть на …надцать кг в месяц, не сходя с дивана. Это особенно впечатляет, если знать, что человек физиологически способен «отдать» не более 3-4 кг жира в месяц, все остальное — жидкость, белки, мышцы. Конечно, если они у вас лишние…
Или… включить мозги и определить для себя комплексную программу безопасного надежного сброса веса, в которой есть место и правильному полноценному питанию, и физическим упражнением, и питию потребного количества воды, и целенаправленной настройке подсознания. И тогда наш собственный организм поможет нам стать стройнее и здоровее.
И тогда наш собственный организм поможет нам стать стройнее и здоровее.
Повысьте метаболизм для улучшения здоровья
Основной метаболизм, или скорость основного обмена (BMR), представляет собой все непроизвольные действия, выполняемые человеческим телом для поддержания жизни в состоянии покоя в состоянии голодания. Эти действия включают дыхание, кровообращение, регулирование температуры тела, нервную активность, секрецию гормонов и синтез новой ткани. Дополнительные калории расходуются на физическую активность и пищеварение. BMR использует от 60 до 70% затрат энергии / калорий человеческого тела.
Есть несколько факторов, которые влияют на ваш метаболизм. BMR частично определяется нашими генами. Факторы, снижающие наш BMR, включают: старение, голодание или соблюдение низкокалорийной диеты, а также увеличение жировых отложений. Факторы, которые увеличивают наш BMR, включают: более высокое соотношение мышечной массы к телу, принадлежность к мужскому полу, беременность, растущие дети, стресс и лихорадка. Температура окружающей среды и гормоны щитовидной железы также влияют на обмен веществ.
Температура окружающей среды и гормоны щитовидной железы также влияют на обмен веществ.
Ставьте цели
Вот несколько способов улучшить метаболизм:
- Получить физический урон. Выполняйте упражнения для сердечно-сосудистой системы большую часть дней недели, включая силовые и силовые тренировки, которые ускоряют метаболизм за счет наращивания мышечной массы. Сердечно-сосудистые упражнения, такие как ходьба, бег трусцой, танцы и езда на велосипеде, увеличивают частоту сердечных сокращений и помогают организму сжигать больше калорий и жира. Силовые тренировки наращивают мышечную массу, что увеличивает ваш метаболизм, потому что мышцы сжигают больше калорий, чем жировая ткань. Каждый день каждый фунт мышц сжигает 6 калорий, а каждый фунт жира сжигает 2 калории.
- Подпитите свой метаболизм правильным питанием. Низкокалорийная диета может помочь вам сбросить несколько килограммов, но также приводит к потере мышечной массы, что затем снижает скорость метаболизма.
 Когда вы едите слишком мало калорий и / или пропускаете приемы пищи, чтобы снизить потребление калорий, ваше тело думает, что есть голод, из-за которого ваш метаболизм замедляется, чтобы сохранить запасы энергии. В конце концов, ваше тело будет сжигать меньше калорий, и потеря веса остановится. Поддерживайте свой метаболизм с помощью здоровой пищи, включая цельнозерновые, растительные и / или нежирные животные белки, фрукты, овощи, обезжиренные молочные продукты, бобовые, бобы и полезные жиры, орехи и семена.
Когда вы едите слишком мало калорий и / или пропускаете приемы пищи, чтобы снизить потребление калорий, ваше тело думает, что есть голод, из-за которого ваш метаболизм замедляется, чтобы сохранить запасы энергии. В конце концов, ваше тело будет сжигать меньше калорий, и потеря веса остановится. Поддерживайте свой метаболизм с помощью здоровой пищи, включая цельнозерновые, растительные и / или нежирные животные белки, фрукты, овощи, обезжиренные молочные продукты, бобовые, бобы и полезные жиры, орехи и семена. - Ешьте часто. Чтобы стимулировать метаболизм, ешьте каждые 3-5 часов бодрствования. Включите нежирный белок и продукты со сложными углеводами / волокнами, чтобы перекусить. Примеры включают дольки яблока с арахисовым маслом; чашка зелени и сырых овощей, ½ стакана черной фасоли с легкой заправкой для салата или сырые овощи, обмакнутые в нежирный греческий йогурт или хумус. Увеличение потребления клетчатки приведет к сжиганию большего количества калорий во время пищеварения.
- Напитки для напитков. Организму требуется вода для переработки клетчатки и калорий.Обезвоживание замедляет метаболизм. Выпивайте стакан ледяной воды перед каждым приемом пищи и перекусом. Каждые 8 унций ледяной воды сжигают дополнительные 25 калорий, а 8 унций воды комнатной температуры сжигают 16 калорий. Умеренный кофе приводит к кратковременному увеличению скорости метаболизма и повышает выносливость во время тренировок. Зеленый чай ускоряет метаболизм на пару часов за счет активного ингредиента катехина и помогает организму сжигать больше калорий во время умеренно интенсивных упражнений.Энергетические напитки ускоряют метаболизм за счет кофеина и, при добавлении, аминокислоты таурина. Однако было замечено, что энергетические напитки вызывают проблемы с высоким кровяным давлением, нарушениями сна и беспокойством. Американская академия педиатрии не рекомендует их детям и подросткам.
- Ешьте жирную рыбу. Омега-3 жирные кислоты ускоряют метаболизм и увеличивают количество ферментов сжигания жира, уменьшая количество ферментов, запасающих жир.
 Дважды в неделю рекомендуются дикий лосось, тунец, скумбрия, сардины, сельдь, форель, консервированный лосось и тунец альбакор.
Дважды в неделю рекомендуются дикий лосось, тунец, скумбрия, сардины, сельдь, форель, консервированный лосось и тунец альбакор. - Приправить. Капсаицин, химическое вещество, содержащееся в остром красном и зеленом перце и карри, немного ускоряет метаболизм. Если вы едите острую пищу и приправы, польза от этого часто увеличивается.
- Получите ZZZ’s. Сон менее 7-8 часов в сутки может изменить ваш метаболизм и снизить количество сжигаемых калорий в состоянии покоя. Недостаток сна связан с тягой к сладкой или жирной пище и увеличивает выработку гормона стресса кортизола. Это вызывает повышенный аппетит и стресс.
- Контрольная температура в помещении. Жаркая и очень прохладная комнатная температура заставляет нас есть больше. Будьте зеленым и опускайте термостат зимой и поднимайте летом, чтобы вы меньше ели и ускоряли метаболизм, чтобы оставаться в тепле или прохладнее.
Преимущества ускоренного метаболизма обеспечат вас большей энергией, помогут сбросить вес и удержать его, а также помогут вам получить качественный сон, сжигая больше калорий перед сном.
Всегда консультируйтесь со своим врачом или поставщиком медицинских услуг, прежде чем вносить какие-либо изменения в диету / режим питания или начинать или изменять свою физическую активность.
Как это влияет на контроль веса / питание
Может возникнуть соблазн обвинить в неудачных попытках похудания или набора веса медленный метаболизм. Однако в редких случаях чрезмерная прибавка в весе из-за медицинских проблем замедляет метаболизм, как в случае синдрома Кушинга или недостаточной активности щитовидной железы (гипотиреоза).
Упражнения для ускорения метаболизма
Хотя некоторые факторы, влияющие на скорость метаболизма, изменить нельзя, существуют способы его максимизировать.Среди лучших способов — упражнения. Это включает в себя аэробные тренировки, чтобы сжечь больше калорий в краткосрочной перспективе, и силовые тренировки для наращивания мышц, которые улучшат ваш метаболизм в долгосрочной перспективе. Поскольку мышцы сжигают больше калорий, чем жира, даже в состоянии покоя, чем больше у вас мышц, тем выше уровень метаболизма в состоянии покоя. Это означает, что ваше тело будет сжигать больше калорий, чтобы поддерживать вас.
Поскольку мышцы сжигают больше калорий, чем жира, даже в состоянии покоя, чем больше у вас мышц, тем выше уровень метаболизма в состоянии покоя. Это означает, что ваше тело будет сжигать больше калорий, чтобы поддерживать вас.
Для получения дополнительной информации:
Понимание метаболизма: как повысить свой с помощью упражнений
Понимание метаболизма: что определяет ваш BMR?
Как питание влияет на метаболизм
Помимо упражнений, вы можете соблюдать некоторые диетические рекомендации, чтобы повысить скорость метаболизма.Например, частые приемы пищи небольшими порциями помогают поддерживать метаболизм на высоком уровне, а это означает, что в целом вы сжигаете больше калорий. Когда вы слишком долго ждете между приемами пищи, ваш метаболизм замедляется, чтобы компенсировать это. Лучший совет — есть 3 раза в день и пару небольших перекусов каждый день. Частое питание также помогает вам есть меньше при регулярном приеме пищи.
Ешьте, чтобы улучшить метаболизм
Всегда завтракайте. Здоровый завтрак запускает вашу метаболическую печь. Исследования показали, что люди, которые едят здоровый, сбалансированный завтрак каждый день, имеют значительно меньшее количество жира в организме.
Поддерживайте достаточный уровень гидратации. Обезвоживание может способствовать неэффективному метаболизму. Температура вашего тела немного падает, если вы обезвожены, заставляя ваше тело накапливать жир, чтобы поддерживать температуру. Вода помогает мобилизовать жировые отложения, а обезвоживание стимулирует жировые отложения.
Продукты, способствующие увеличению метаболизма
А как насчет утверждений о том, что определенные продукты увеличивают метаболизм? Белок требует больше энергии для переваривания, чем углеводы.Итак, теоретически еда с высоким содержанием белка может ускорить метаболизм немного больше, чем еда с высоким содержанием углеводов и тем же количеством калорий. Белок также заставляет вас чувствовать себя более удовлетворенным после еды, чтобы избежать переедания. Но имейте в виду, что большая часть потери веса объясняется дефицитом калорий и увеличением физических нагрузок, а не изменениями процентного содержания белков или углеводов в рационе.
Омега-3 жиры могут ускорить обмен веществ, а потребление насыщенных жиров замедляет метаболизм.Необработанные продукты, такие как фрукты, овощи и цельнозерновые, перевариваются дольше, потому что организму приходится усерднее работать, чтобы переваривать клетчатку. По сравнению с рафинированными и обработанными продуктами, простое переваривание цельных продуктов сжигает больше калорий.
Острая пища, такая как красный перец, может усиливать метаболизм примерно на 30 минут после еды, но исследования не показали, что эффект длится дольше. Кроме того, наибольший эффект наблюдался при употреблении острой пищи с продуктами с высоким содержанием жиров, которых лучше избегать. Зеленый чай или EGCG и кофеин также могут улучшить сжигание калорий, но неясно, достаточно ли этого для ускорения похудания.Прирост по-прежнему незначителен по сравнению с тем, что вам нужно для похудения.
Могут ли добавки или таблетки помочь снизить метаболизм?
Некоторые продавцы добавок для похудения заставляют вас поверить, что их продукт поможет ускорить обмен веществ и избавиться от жира. Не обманывайтесь: волшебной пилюли не существует. Как правило, ингредиенты лишь незначительно ускоряют обмен веществ и несут в себе риски. Некоторые таблетки имеют опасные побочные эффекты, а такие ингредиенты, как «ускоряющие метаболизм», такие как белая ива, кофеин и ма хуанг, вызывают чрезмерную нагрузку на щитовидную железу.Стимуляторы, такие как гуарана, мате, кофеин и горький апельсин, увеличивают частоту сердечных сокращений и могут вызвать повышение артериального давления. Управление по санитарному надзору за качеством пищевых продуктов и медикаментов не требует от производителей пищевых добавок доказывать, что их продукты безопасны или эффективны, поэтому разумно относиться к этим продуктам скептически.
Однако прием витаминно-минеральных добавок имеет смысл. Эффективный метаболизм также требует плавного протекания многих сложных процессов в организме, от которых зависит достаточно.Недостаточное количество B6, B12, фолиевой кислоты, тиамина и ниацина может замедлить метаболизм в организме. Магний также играет роль в метаболизме, поскольку он используется в более чем 300 биохимических реакциях в организме.
Калорийность и обмен веществ
Хотя ваш метаболизм влияет на основные потребности вашего тела в энергии, увеличение веса чаще всего является результатом потребления большего количества калорий, чем вы сжигаете. Таким образом, чтобы похудеть, вам нужно создать дефицит энергии, потребляя меньше калорий, увеличивая количество калорий, которые вы сжигаете за счет физической активности, или и то, и другое.Формула победы для поддержания веса заключается в том, что количество потребляемых вами калорий должно равняться затрачиваемым калориям. Так что сосредоточьтесь на факторах, которые вы контролируете: своей диете и образе жизни. Они могут помочь вам контролировать свой вес и, возможно, улучшить метаболизм.
Если вас беспокоит свой вес или вы думаете, что ваш метаболизм слишком медленный, проконсультируйтесь с врачом. Ваш врач может проверить наличие медицинских причин и помочь вам внести изменения в здоровый образ жизни, чтобы помочь вам сбросить вес.
Мария Фейрес, RD — зарегистрированный диетолог, личный тренер, специалист по здоровью и фитнесу и писатель-фрилансер из Саммамиша, штат Вашингтон.Мария считается одним из ведущих экспертов Западного Вашингтона по фитнесу и питанию. Как владелец компании Active Nutrition Fitness & Consulting, Мария предоставляет персонализированные услуги по питанию, индивидуальные тренировки и профилактические и постреабилитационные фитнес-программы в своей частной тренировочной студии. Она также предоставляет консультации по питанию и тренинги по Skype, телефону и онлайн для удаленных клиентов. Мария возглавляет отрасль в разработке передовых методов фитнеса и питания, а также инновационных и уникальных фитнес-программ.Мария профессионально разрабатывает каждую тренировку, план питания и обеспечивает личное внимание, дополнительную мотивацию, поддержку и ответственность, которые помогают ее клиентам достичь оптимальной производительности и здоровья. Свяжитесь с Марией или узнайте больше о ней на сайте www.myactivenutrition.com.
Скорость метаболизма — обзор
Авторадиография: методы и ограничения
LCMRglcs измеряются с помощью метода 2DG, а показатели LCBF оцениваются с помощью метода IAP.Оба метода адаптированы для взрослых мышей и неполовозрелых крыс. Пространственное разрешение этих методов превосходное и основано на обнаружении зерен серебра (на авторадиографических пленках), которые возбуждаются электронами, испускаемыми меченной 14 C молекулой, которая накопилась в различных областях мозга. Оптическую плотность интересующих областей можно измерить (например, на серийных корональных срезах головного мозга) и сравнить с калиброванными стандартами [ 14 C] -метилметакрилатного пластика.Последним этапом является вычисление с помощью специальных операционных уравнений отношения концентрации 14 C к скорости использования церебральной глюкозы или кровотока в каждой конкретной области мозга, представляющей интерес, с использованием системы обработки изображений. Для этих процедур сейчас доступно множество пакетов программного обеспечения (включая бесплатное программное обеспечение, предлагаемое на веб-сайте NIH).
Основная техническая трудность обоих методов связана с тем, что эксперименты требуют введения полиэтиленовых катетеров, обычно в бедренную артерию и вену, для количественной оценки результатов.Венозный катетер позволяет вводить в / в. инъекция (2DG) или инфузия индикатора (IAP), в то время как артериальный катетер необходим для забора пробы крови, рассчитанных по времени, для количественной оценки временной эволюции специфической активности индикатора в крови. У взрослой крысы введение катетеров не представляет каких-либо технических трудностей, а объем крови достаточен для повторного отбора проб без значительных последствий для состояния здоровья животных. Однако незрелым крысам (или мышам) было труднее ввести катетеры; эта трудность усугубляется у неполовозрелых животных, у которых малые размеры сосудов сочетаются с незрелостью стенки сосуда (особенно вены).Кроме того, из-за небольшого объема крови в организме необходимо уменьшить объем и количество проб.
Выбор интересующей техники также ограничен временным разрешением. В то время как LCBF можно измерить за очень короткие промежутки времени от 20 до 60 с, количественная оценка LCMRglcs требует 45-минутного периода стабильного метаболизма, чего невозможно достичь с такими событиями, как короткие судороги. Это ограничение привело к тому, что некоторые авторы использовали метод 2DG для неколичественного картирования метаболизма мозга во время припадков.В этом последнем случае данные выражаются в виде соотношений оптической плотности интересующей структуры и структуры, которая должна оставаться неизменной (чаще всего мозолистого тела или мозжечка). Однако этот полуколичественный подход не следует использовать в слишком короткие сроки; более длительные периоды необходимы, чтобы свести к минимуму возможные ошибки из-за большой доли свободных 2DG, присутствующих в крови и ткани мозга во время жертвоприношения. При изучении коротких событий предпочтительно выбирать альтернативные методы, такие как авторадиографическое измерение LCBF, которое можно количественно оценить за очень короткие промежутки времени.
Флавоноид процианидин C1 обладает сенотерапевтической активностью и увеличивает продолжительность жизни у мышей
Низкие концентрации GSE сдерживают экспрессию SASP
В попытке идентифицировать новые соединения, которые могут эффективно модулировать стареющие клетки, был проведен беспристрастный скрининг агентов с помощью фитохимической библиотеки, состоящей из 46 лекарственных средств растительного происхождения (библиотека PDMA). Мы использовали первичную нормальную линию стромальных клеток простаты человека, PSC27, в качестве клеточной модели для этой цели.Состоящий в основном из фибробластов, но с небольшим процентом нефибробластных клеточных линий, включая эндотелиальные клетки и гладкомышечные клетки, PSC27 сам по себе является первичной линией клеток и развивает типичный SASP после воздействия стрессоров, таких как генотоксическая химиотерапия или ионизирующее излучение 14, 15,16,17 . Мы обработали эти клетки предварительно оптимизированной сублетальной дозой блеомицина (50 мкг / мл -1 ) и наблюдали усиление окрашивания связанной со старением β-галактозидазы (SA-β-Gal), снижение включения 5-бромдезоксиуридина и повышенное повреждение ДНК. очаги репарации (DDR) через 7–10 дней после (дополнительный рис.1а – в). Мы разработали стратегию скрининга, чтобы сравнить влияние отдельных лекарственных средств на выживаемость и профиль экспрессии стареющих клеток (расширенные данные, рис. 1а).
Одним из многообещающих преимуществ сенолитических агентов является избирательная индукция запрограммированной гибели стареющих клеток, таких как ABT-263, ABT-737 и комбинированное использование дазатиниба и кверцетина 11,18,19 . Сначала мы протестировали эффективность этих геропротекторных препаратов против стареющих клеток PSC27, чтобы продемонстрировать их потенциал в качестве экспериментальной клеточной модели для скрининга лекарств.Наши предварительные данные предполагают, что каждое из этих соединений значительно истощает стареющие клетки, но не пролиферирует клетки, тем самым подтверждая возможность использования этой стромальной линии для дальнейших исследований (расширенные данные, рис. 1b). После крупномасштабного скрининга библиотеки PDMA мы идентифицировали несколько соединений, способных избирательно убивать стареющие клетки в культуре (расширенные данные, рис. 1c – e).
Среди агентов, демонстрирующих предварительные эффекты против старения, были GSE, кверцетин, физетин, куркумин и пиперлонгумин (расширенные данные, рис.1г, д). Кверцетин и физетин имеют схожие химические структуры, обладают схожим лечебным действием и оба являются известными сенолитиками 11,20,21 . Куркумин и пиперлонгумин также являются природными соединениями с недавно обнаруженным сенолитическим потенциалом 22,23 . Мы решили сосредоточиться на GSE, который оставался в значительной степени малоизученным источником. В условиях in vitro GSE подавлял SASP с максимальной эффективностью при 0,1875 мкг / мл -1 (расширенные данные, рис. 2a), что соответствует свойству сеноморфиков 24 .Более низкие или более высокие концентрации GSE были менее эффективными, возможно, из-за индукции клеточных стрессовых реакций в результате повышенной цитотоксичности (расширенные данные, рис. 2а). Используя RNA-seq, мы обнаружили, что обработка GSE значительно изменяла профиль экспрессии клеток старения: 2644 гена подавлялись, а 1472 гена усиливались при изменении в 2,0 раза на ген ( P <0,01) (расширенные данные, рис. 2b). . Хотя экспрессия нескольких генов, не связанных с SASP, показала сходную тенденцию с экспрессией типичных факторов SASP (расширенные данные, рис.2c), данные нашего анализа обогащения набора генов (GSEA) подтвердили снижение экспрессии молекулярных сигнатур SASP или активацию комплекса ядерного фактора (NF) -κB, который является ключевым медиатором провоспалительного фенотипа (расширенные данные рис. 2г, д).
Ядерная транслокация p65, одной из основных субъединиц комплекса NF-κB, наблюдалась в стареющих клетках, что согласуется с его функциональным участием в экспрессии SASP 14 (Extended Data Fig. 2f). Следует отметить, что этой тенденции существенно противодействовал GSE в низких концентрациях (например, 0.1875 мкг мл -1 ). Напротив, активация передачи сигналов NF-κB не подавлялась, а, скорее, казалась усиленной, когда GSE использовался в более высоких концентрациях (например, 3,7500 мкг / мл -1 ), что свидетельствует о дифференциальных ответах стареющих клеток в этих условиях лечения. Активация передачи сигналов DDR, о чем свидетельствует фосфорилирование киназы ATM в ядерных фракциях, и экспрессия хемокинового лиганда мотива C – X – C (CXCL) 8, одного из определяющих факторов SASP, наблюдаемого в цитоплазматических фракциях, соответствовали NF -κB активация в этих настройках (Расширенные данные рис.2е).
Профили белок-белковых взаимодействий выявили высокоактивную сеть, включающую множество факторов, которые значительно усиливаются при клеточном старении, но подавляются, как только клетки подвергаются воздействию GSE (Extended Data Fig. 3a). Профилирование генной онтологии показало, что эти молекулы функционально вовлечены в биологические процессы и связаны с клеточными компонентами, в целом согласующимися с секретируемой природой SASP (расширенные данные рис. 3b, c). Таким образом, GSE — это натуральный продукт, который может контролировать провоспалительный профиль стареющих клеток, SASP, при использовании в определенном диапазоне концентраций.Хотя GSE был не единственным натуральным продуктом с сенолитической эффективностью в наших клеточных анализах (расширенные данные, рис. 1d, e), наше последующее исследование в основном было сосредоточено на GSE, поскольку его геропротекторная способность оказалась особенно поразительной.
GSE обладает сенолитической активностью при высоких концентрациях
Учитывая эффективность GSE в снижении SASP как сеноморфного агента, мы затем исследовали потенциал этого природного продукта в уничтожении стареющих клеток в более высоких концентрациях, действуя как сенолитик.Окрашивание SA-β-Gal показало, что стареющие клетки элиминировались при концентрации GSE 0,75 мкг / мл -1 (рис. 1a, b). При 3,75 мкг / мл -1 GSE было достигнуто плато 20% выживаемости стареющих клеток (рис. 1a, b).
Рис. 1: Характеристика способности GSE устранять стареющие клетки.
a , Количественная оценка выживаемости стареющих клеток PSC27 по положительности SA-β-Gal. GSE вносили в среду в возрастающих концентрациях. CTRL — контрольные (пролиферирующие) клетки; SEN — стареющие клетки; NS, не имеет значения.Значения P рассчитывали с помощью однофакторного дисперсионного анализа с тестом множественного сравнения Тьюки. b , типичные изображения, демонстрирующие окрашивание SA-β-Gal после обработки клеток PSC27 различными концентрациями GSE. Масштабная линейка, 20 мкм. Данные представляют из трех независимых экспериментов. ДМСО, диметилсульфоксид. c , Анализ выживаемости контрольных и стареющих клеток PSC27 после обработки GSE (при концентрациях 0,3750, 0,7500, 1,8750, 3,7500, 7,5000 и 15,0000 мкг / мл -1 , соответственно).Данные представлены как среднее ± стандартное отклонение. и были получены из трех биологических повторов ( n = 3 независимых анализа). Значения P были рассчитаны с помощью двусторонних тестов t . d , Измерение динамики жизнеспособности in vitro при обработке контрольных и стареющих клеток PSC27 GSE (3,75 мкг мл -1 ). Данные представлены как среднее ± стандартное отклонение. и были получены из трех биологических повторов ( n = 3 независимых эксперимента). Значения P были рассчитаны с помощью двусторонних тестов t . e , Измерение проточной цитометрии контрольных и стареющих клеток PSC27 после обработки с помощью набора аннексина V – FITC и пропидия йодида (PI) и окрашивания 4,6-диамидино-2-фенилиндолом (DAPI) для определения степени апоптоза. Q1 – Q4, 1–4 квартили. f , Сравнительная количественная оценка процента жизнеспособных (Q4, PI — аннексина V — ) и апоптического (Q2 и Q3, PI + аннексина V + и PI — аннексина V + соответственно) клетки в контрольных или стареющих популяциях, обработанных носителем или GSE в течение 3 дней ( n = 3 биологически независимых анализа).Значения P были рассчитаны с помощью двусторонних тестов t . g , Измерение сигнала флуоресценции MitoSOX Red, индикатора митохондриального супероксида, в клетках PSC27 в различных условиях. Значения P были рассчитаны с помощью двусторонних тестов t . h , Масс-спектры высокого разрешения, показывающие хроматограмму общих ионов (TIC) и хроматограмму основного пика GSE после выполнения HPLC – ESI-QTOF-MS. Если не указано иное, клетки подвергали соответствующему анализу через 3 дня после обработки GSE в условиях культивирования.cps, отсчетов в секунду. Данные на гистограммах и кривых регрессии показаны как среднее ± стандартное отклонение. и представляют три биологических повтора. NS, P > 0,05; * P <0,05; ** P <0,01; *** P <0,001; **** П <0,0001.
Исходные данные
Анализы жизнеспособности клеток показали, что GSE индуцировал гибель стареющих клеток, но не гибель пролиферирующих клеток, начиная с концентрации 0,75 мкг / мл -1 (рис.1в). При концентрации 7,50 мкг / мл -1 процент выживших стареющих клеток снизился примерно до 10%, тогда как жизнеспособность пролиферирующих клеток не пострадала даже при 15,00 мкг / мл -1 GSE (рис. 1c). самая высокая концентрация, используемая в наших клеточных анализах, что свидетельствует о выдающейся селективности и специфичности GSE против стареющих клеток, которые являются основными чертами сенолитиков.
Затем мы измерили способность GSE по-разному воздействовать на стареющие клетки с течением времени.После обработки GSE в концентрации 3,75 мкг / мл -1 жизнеспособность стареющих клеток существенно не снижалась до 20 часов. Разница в жизнеспособности стареющих клеток и контроля (пролиферирующих клеток) достигла максимума через 32 часа, что предполагает гетерогенность внутренней устойчивости к сенолитикам в популяциях стареющих клеток (рис. 1d).
Поскольку GSE оказывает явное действие против стареющих клеток, мы проанализировали эффективность GSE в индукции апоптоза клеток.Проточная цитометрия показала значительно сниженную жизнеспособность, в то время как апоптоз стареющих клеток, но не апоптоз пролиферирующих клеток, был повышенным (рис. 1e, f и дополнительный рис. 2a). Дисфункция митохондрий и метаболические изменения являются одними из признаков стареющих клеток и старения организма, событий, вызывающих окислительный стресс и производство активных форм кислорода (АФК), таких как супероксид 3,25 . Мы использовали MitoSOX Red, индикатор митохондриального супероксида 26 , для исследования межклеточных изменений и обнаружили, что GSE способствует генерации митохондриальных АФК в стареющих клетках, но не в пролиферирующих клетках (рис.1г). Таким образом, наши данные согласуются с моделью, в которой GSE убивает стареющие клетки посредством индукции апоптоза и обострения митохондриального стресса in vitro.
Виноградные косточки составляют 38–52% по сухому веществу винограда и являются обильным источником антиоксидантов 27 . Мы применили жидкостную хроматографию высокого давления (ВЭЖХ) в сочетании с квадрупольной времяпролетной масс-спектрометрией (QTOF-MS), оснащенной интерфейсом ионизации электрораспылением (ESI), для идентификации основных компонентов GSE.Мы обнаружили три основные категории фитохимических веществ, включая фенольные кислоты, флавоноиды (такие как флаван-3-ол, процианидины) и другие соединения (рис. 1h и дополнительная таблица 1). Среди них несколько компонентов были идентифицированы как процианидины и их производные, которые, как сообщалось, нацелены на митохондриальные белки и облегчают множественные хронические заболевания 28 . Однако основные компоненты, опосредующие сенолитическую функцию GSE, остаются в значительной степени неясными.
Компонент PCC1 GSE обладает сенолитической активностью.
Сообщенные биологические активности процианидинов виноградных косточек включают уменьшение окислительного повреждения, подавление воспаления и индукцию апоптоза раковых клеток 29,30,31,32 .Среди отдельных соединений, обнаруженных в GSE, особого внимания заслуживает PCC1, поскольку было показано, что он вызывает повреждение ДНК, вызывает остановку клеточного цикла и увеличивает экспрессию киназ контрольных точек 33 . Данные предварительного анализа (общая ионная хроматограмма) GSE, смеси фитохимических агентов как таковой, с помощью HPLC – QTOF-MS предполагают присутствие PCC1, поскольку профиль GSE на определенных пиках MS совпадает с профилем хроматограммы химически чистого PCC1 приобретено из коммерческого источника (рис. 1h и дополнительный рис.2б).
PCC1, как было показано, снижает уровень BCL-2, но увеличивает экспрессию регулятора BAX и активность каспаз 3 и 9 в культивируемых раковых клетках, потенциально вызывая противораковые эффекты за счет индукции апоптоза 33 . Следовательно, мы затем оценили способность и селективность PCC1 устранять стареющие клетки в культуре. Данные свидетельствуют о том, что PCC1 является сенолитическим для стареющих стромальных клеток, начиная с концентрации 50 мкМ, при которой пролиферирующие клетки остаются в значительной степени незатронутыми (рис.2а, б и дополнительная таблица 2). Хотя более высокие концентрации вызывали более низкую выживаемость стареющих клеток, с порогом примерно 200 мкМ, PCC1 проявлял токсичность по отношению к контрольным клеткам только при использовании в 600 мкМ или выше (фиг. 2b). Динамика активности каспазы 3/7 показала, что PCC1 проявлял апоптотические эффекты в течение 12 часов, достигая плато через 24 часа (рис. 2c). Этот результат в значительной степени согласуется с измерениями жизнеспособности (рис. 2d). Сенолитическая природа PCC1 была подтверждена клетками, которые вступили в стадию старения из-за репликативного истощения или старения (RS) или сверхэкспрессии онкогена (HRAS G12V ) (OIS), что вызывает стрессовые воздействия, подобные таковым при старении, вызванном терапией (рис.2e, расширенные данные рис. 4b – e и дополнительная таблица 2). В совокупности результаты предполагают, что PCC1 избирательно очищает стареющие стромальные клетки человека, индуцированные различными стимулами, дозозависимым образом, но без значительного воздействия на незрелые клетки при использовании в соответствующих концентрациях.
Рис. 2: Характеристика сенолитического потенциала PCC1.
a , Измерение выживаемости стареющих клеток PSC27 путем окрашивания SA-β-Gal. PCC1 применяли в возрастающих концентрациях.Значения P рассчитывали с помощью однофакторного дисперсионного анализа с тестом множественного сравнения Тьюки. b , Выживание стареющих клеток PSC27, индуцированных блеомицином при увеличении концентрации PCC1. c , Апоптотический анализ активности каспазы 3/7. d , Кривые зависимости выживаемости от времени для оценки жизнеспособности клеток PSC27 после обработки PCC1. и , Изображения окрашивания SA-β-Gal. TIS, старение, вызванное терапией (блеомицином). Масштабная линейка, 20 мкм. Данные представляют из трех независимых экспериментов. f , Проточная цитометрия после обработки с помощью набора аннексина V – FITC и PI и окрашивания DAPI для определения уровней апоптоза. г , Количественное определение процента жизнеспособных (Q4, PI — , аннексин V — ) и апоптотического (Q2 и Q3, PI + , аннексин V + и PI — , аннексин V + , соответственно) клетки после обработки носителем или PCC1 в течение 3 дней ( n = 3 биологически независимых анализа). ч , Иммунофлуоресцентное окрашивание клеток PSC27.RS индуцировали последовательным пассированием перед обработкой PCC1. Красный, p16 INK4a . Клетки на раннем пассаже (P10) использовали в качестве отрицательного контроля. ABT-263 (1,25 мкМ) тестировали в качестве положительного контроля. Масштабная линейка, 20 мкм. и , Статистика иммунофлуоресцентного окрашивания. j , сенолитическая активность, индуцированная PCC1 после ингибирования панкаспазы (20 мкМ QVD-OPh). k , PD-анализ МСК человека. PCC1 применяли на 8-й день после начала экспериментов, как указано. БЛЕО, блеомицин.Для c , d , k данные показаны как среднее ± стандартное отклонение. и были получены из трех биологических повторов ( n = 3 независимых анализа). Для данных в b — d , g , i , j , P значения были рассчитаны с помощью двусторонних t -тестов. В экспериментах для c — k использовали PCC1 в концентрации 100 мкМ. Если не указано иное, образцы собирали для анализов через 3 дня после обработки PCC1.Данные на гистограммах показаны как среднее ± стандартное отклонение. и представляют три биологических повтора. NS, P > 0,05; * P <0,05; ** P <0,01; *** P <0,001; **** П <0,0001.
Исходные данные
Чтобы экспериментально расширить и установить эффективность PCC1 по всем клеточным линиям, мы обработали фибробласты легких плода человека (WI38), первичные эндотелиальные клетки пупочной вены человека (HUVEC) и мезенхимальные стволовые клетки человека (MSC) с помощью PCC1 и обнаружили, что клетки всех этих клонов проявляли сходную восприимчивость к селективной абляции PCC1, тогда как их нестареющие аналоги оставались жизнеспособными (расширенные данные, рис.4f – h и дополнительная таблица 3). Мы также подтвердили индукцию апоптоза в стареющих клетках в ответ на PCC1 с помощью проточной цитометрии, в то время как пролиферирующие клетки оставались в значительной степени незатронутыми PCC1 (рис. 2f, g). В целом, наши данные показывают, что PCC1 избирательно устраняет стареющие клетки из разных типов клеток и возникающие в результате различных триггеров старения.
Чтобы визуализировать истощение стареющих клеток под действием PCC1, мы исследовали экспрессию p16 INK4a , широко используемого маркера старения, в стромальных клетках, которые испытывали RS.PCC1 эффективно удалял p16-позитивные стареющие клетки, которые появлялись только в популяциях позднего пассажа PSC27, с эффективностью, в значительной степени напоминающей эффективность ABT-263 (1,25 мкМ), хорошо зарекомендовавшего себя синтетического сенолитического агента 18,21 (рис. 2h ,я).
Чтобы подтвердить, что PCC1-опосредованная элиминация стареющих клеток происходит в основном за счет индукции апоптоза, а не через другие формы запрограммированной гибели клеток, мы обрабатывали клетки ингибитором апоптоза панкаспазы хинолил-валил- O -метиласпартил- ( -2,6-дифторфенокси) метилкетон (QVD-OPh).QVD-OPh обращает способность PCC1 убивать стареющие клетки. PCC1, таким образом, разделяет свою зависимую от каспазы индукцию апоптоза как сенолитический признак с ABT-263 (Fig. 2j). Дальнейший анализ с химическими ингибиторами исключил гибель клеток, вызванную PCC1, в результате ферроптоза, пироптоза или некроптоза (расширенные данные, рис. 4i).
Чтобы оценить потенциал удвоения клеточной популяции (PD) после лечения генотоксичными препаратами, мы использовали МСК, которые могут самообновляться и возобновлять пролиферацию на основе колоний даже после воздействия стрессов окружающей среды 34 , вероятно, из-за гетерогенности повреждение, при этом клетки, испытывающие меньшее повреждение, предположительно способны сохранять потенциал для самовосстановления и повторного входа в клеточный цикл 24,35 .В отличие от поврежденных блеомицином клеток, рост которых после обработки быстро прекращался, обработка PCC1 после старения значительно увеличивала способность МСК к PD, особенно после удаления стареющих клеток, развивающих SASP и сохраняющих потенциал для индукции паракринного старения в популяциях клеток (рис. 2k). Однако обработка PCC1 не влияла на PD пролиферирующих клеток, что дополнительно свидетельствует о селективности PCC1 для стареющих клеток по сравнению с их нормальными аналогами.
Поскольку GSE представляет собой сложную фитохимическую смесь, многие из ее компонентов обладают антиоксидантной и противовоспалительной активностью 27,36 , мы исследовали, был ли PCC1 основным компонентом GSE, участвующим в истощении стареющих клеток, или альтернативные фитохимические вещества в GSE может способствовать его общему сенолитическому эффекту. С этой целью мы исследовали влияние отдельных фитохимических молекул на выживаемость стареющих клеток PSC27. Большинство компонентов GSE не проявляют сенолитической активности в диапазоне доз PCC1 и не вызывают значительной гибели пролиферирующих клеток (дополнительные фиг.3 и 4). Хотя флавоноид кверцетин проявлял сенолитическую активность, как и в наших предыдущих исследованиях, свойство, разделяемое с естественными флавонами 11,21 , «восстановленным GSE», состоящим из основных компонентов, смешанных в соответствии с их массовым процентным содержанием, как было выявлено с помощью нашей ВЭЖХ – QTOF-MS. данные (дополнительная таблица 1, обратите внимание, что кверцетин составляет только 0,9%), но целенаправленно исключая PCC1, не показали результатов, аналогичных результатам, наблюдаемым для PCC1 в обоих анализах (дополнительные рисунки 3 и 4). Хотя мы не можем сделать вывод о том, вносят ли вклад другие компоненты, наши данные ясно указывают на то, что PCC1 является первичным медиатором сенолитического эффекта GSE.
PCC1 вызывает митохондриальную дисфункцию в стареющих клетках
Учитывая выдающуюся эффективность PCC1 в селективном индуцировании гибели стареющих клеток, мы исследовали лежащие в основе механизмы. PCC1 принадлежит к суперсемейству флавоноидов, которые могут улавливать свободные радикалы, хелатировать металлы и снижать образование гидропероксидов, антиоксидантные свойства, приписываемые функциональной группе ‘-OH’ в структуре и ее положению в кольце молекулы флавоноида 27 . Антиоксидантная способность процианидинов частично определяется степенью их полимеризации, в то время как PCC1 по своей природе является тримером процианидин-эпикатехина (рис.3а).
Рис. 3: PCC1 индуцирует апоптоз стареющих клеток, задействуя проапоптотические пути.
a , Химическая структура тримерного эпикатехина PCC1. b , тепловая карта, изображающая верхние гены (50), значительно повышенная в стареющих клетках PSC27, но подавляющаяся при обработке PCC1 (50 мкМ). Красные звезды, факторы SASP. c , график GSEA значимого набора генов в спектре SASP. FDR — коэффициент ложного обнаружения; NES, нормализованная оценка обогащения. d , график GSEA значительного набора генов, связанных с передачей сигналов, опосредованной NF-κB. и , NetworkAnalyst карта белок-белковых взаимодействий типичных SASP-ассоциированных факторов, которые значительно усиливаются в стареющих клетках, но подавляются обработкой PCC1. f , Тепловая карта, показывающая дифференциальную экспрессию генов семейства BCL-2 в контрольных, стареющих и стареющих клетках, обработанных PCC1. г , Иммуноблоттинг клеток PSC27, подвергнутых действию различных агентов. Была исследована экспрессия проапоптотических и антиапоптотических факторов и молекул, связанных с передачей сигналов DDR.Каспаза 3 (t), общая каспаза 3; каспаза 3 (с), расщепленная каспаза 3; p, фосфорилированный. β-актин и GAPDH, контроль загрузки. Данные представляют из трех независимых экспериментов. ч г. Клетки инфицировали тремя разными видами коротких шпилечных РНК, нацеленных на NOXA или PUMA , перед тем, как подвергнуть их воздействию блеомицина для индукции старения. Через семь дней клетки обрабатывали PCC1 (100 мкМ) в течение 3-х дней для индукции апоптоза. SCR, схватка. i , Экспрессия NOXA определялась с помощью количественной ПЦР с обратной транскрипцией (RT – qPCR).Клетки обрабатывали блеомицином для индукции старения перед воздействием 100 мкМ PCC1 в течение 3 дней в отсутствие или в присутствии 10 мкМ MK2206, 10 мкМ руксолитиниба или 20 нМ BMS-582949 для ингибирования активности AKT, JAK1 и / или JAK2 или p38. МАПК соответственно. j , Аналогичный набор анализов экспрессии RT – qPCR для PUMA с использованием условий, описанных в i . k — m , Измерение жизнеспособности клеток после обработки PCC1 в отсутствие или в присутствии MK2206 ( k ), руксолитиниба ( l ) или BMS-582949 ( m ), включенных для подавления ферментативной активности AKT, JAK1 и / или JAK2 или p38 MAPK, соответственно.Для данных в c , d , P значения были рассчитаны с помощью однофакторного дисперсионного анализа с апостериорным сравнением Тьюки. Статистическая значимость в диапазоне ч, — м. была рассчитана с использованием двусторонних тестов t или одностороннего дисперсионного анализа (критерий Даннета). Данные на всех гистограммах показаны как среднее ± стандартное отклонение. и представляют собой три биологические копии. NS, P > 0,05; * P <0,05; ** P <0,01; *** P <0,001; **** П <0.0001.
Исходные данные
Мы сначала проанализировали влияние PCC1 на транскриптомную экспрессию стареющих клеток. Биоинформатика показала, что 4406 генов были значительно усилены, а 2766 генов были подавлены в стромальных клетках после обработки PCC1 (дополнительный рис. 5a). Мы наблюдали большой набор факторов SASP, экспрессия которых заметно повышалась во время клеточного старения, но существенно снижалась, когда стареющие клетки подвергались воздействию PCC1 (рис. 3b).Профилирование GSEA показало, что сигнатуры SASP и NF-κB заметно подавлялись обработкой PCC1 (Fig. 3c, d). Мы также заметили множественные взаимные взаимодействия или функциональные связи между этими факторами, которые усиливаются во время старения и снижаются после лечения PCC1, появляющиеся в списке самых дифференциально экспрессируемых генов, большинство из которых обычно были секретируемыми факторами (рис. 3e).
Чтобы понять избирательность PCC1 для стареющих клеток, мы дополнительно оценили профиль транскриптомной экспрессии и заметили, что PCC1 индуцировал изменения экспрессии у нескольких членов надсемейства BCL-2 (рис.3е). Хотя передача сигналов DDR оставалась в значительной степени незатронутой, наблюдалась PCC1-зависимая активация или активация митоген-активируемой протеинкиназы p38 (MAPK), при этом в этих клетках происходило расщепление каспазой 3 (рис. 3g). Хотя экспрессия BCL-xL была повышена в стареющих клетках по сравнению с их пролиферирующими контролями, обработка PCC1 не приводила к дальнейшему повышению уровня его белка. Уровни двух других факторов BCL-2, а именно BCL-2 и BAX, практически не изменились. В то время как NOXA и PUMA (два члена подсемейства 3 (Bh4) гомологии BCL-2 — только проапоптотическое подсемейство) проявляли разные паттерны экспрессии во время клеточного старения, обработка PCC1 приводила к усилению активности обоих факторов (рис.3г).
Нокдаун проапоптотических факторов BCL-2 предполагает, что NOXA и PUMA частично опосредуют сенолитические действия PCC1 (рис. 3h и расширенные данные рис. 5a – c). Обработка химическими ингибиторами киназы AKT, киназы Janus (JAK) 1, JAK2 и p38 MAPK также предполагала участие этих сигнальных путей в экспрессии PMAIP1 ( NOXA ) и BBC3 ( PUMA ) и стареющих клеток. апоптоз после обработки PCC1 (рис. 3i – m).
Поскольку сбивающие с ног NOXA и PUMA только частично подавляли сенолитический эффект PCC1 (рис.3h, k – m), мы исследовали другие возможные механизмы, приводящие к гибели стареющих клеток. Поскольку процианидины обычно повышают жизнеспособность клеток, снижают продукцию ROS и сдерживают окислительный стресс в клетках млекопитающих 37,38 , мы затем спросили, могут ли аналогичные или антиоксидантные эффекты наблюдаться в стареющих клетках, подвергшихся воздействию PCC1. Неожиданно мы обнаружили обратное, поскольку стареющие клетки PSC27 демонстрировали повышенные уровни АФК при обработке PCC1, в отличие от их пролиферирующих аналогов (рис.4a и расширенные данные на рис. 5d, обратите внимание на сигналы от зонда 2′-7′-дихлородигидрофлуоресцеина диацетата (DCFH-DA)). Обработка HS-1793, стабильным аналогом ресвератрола, который обладает активностью улавливания свободных радикалов 39 , эффективно блокировала продукцию ROS в стареющих клетках, обработанных PCC1 (расширенные данные, рис. 5e, f), тогда как уровни ROS были дополнительно увеличены после воздействия Обработанные PCC1 стареющие клетки по отношению к CCCP, протонофорному митохондриальному разобщителю 40 или руксотемитиду (LTX-315), амфипатическому катионному пептиду, который индуцирует проницаемость внешней митохондриальной мембраны 41 , каждый применяется в концентрациях, не являющихся контрольными. ячеек (Расширенные данные рис.5д, е). Хотя лечение CCCP или руксотемитидом само по себе также вызывало усиленную продукцию ROS, эффекты, как правило, были меньше, чем эффекты, вызванные PCC1, что позволяет предположить, что PCC1 запускает митохондриальную дисфункцию в стареющих клетках. Измеряя индекс апоптоза стареющих клеток (активность каспазы 3/7), мы обнаружили, что эффект, индуцированный PCC1, может быть дополнительно усилен при комбинации PCC1 с каждым разрушителем митохондрий, но подавлен при совместной обработке с HS-1793 (расширенные данные рис. .5г).
Рис. 4: Апоптоз, индуцированный PCC1, частично опосредуется митохондриальной дисфункцией.
a , Измерение уровней ROS с помощью DCFH-DA, проницаемого для клеток флуоресцентного зонда, чувствительного к изменениям в окислительно-восстановительном состоянии клеток. Эксперименты проводили через 1 день после обработки PCC1. Слева, репрезентативные изображения. Масштабная линейка 10 мкм. Верно, статистика. DCF, дихлородигидрофлуоресцеин. b , Иммуноблоттинг после воздействия на клетки различных обработок. Распределение цитохрома с между митохондриями и цитоплазмой определяли путем выделения митохондрий из надосадочных жидкостей цитозоля через 3 дня после обработки PCC1.ЦОГ IV — это конечный фермент дыхательной цепи митохондрий и митохондриальный маркер. c , Кривые зависимости выживаемости от времени для оценки жизнеспособности клеток PSC27 после обработки PCB2, другим членом семейства природных процианидинов. Данные представлены как среднее ± стандартное отклонение. и были получены из трех биологических повторов ( n = 3 независимых анализа). d , анализ продукции ROS проводили аналогично тому, как описано в a , за исключением того, что клетки подвергались воздействию PCB2.Масштабная линейка 10 мкм. e , Иммуноблоттинг экспрессии и распределения ATM, p53 и каспазы 3 между цитоплазмой и ядром. GAPDH и ламин A / C, контролируют загрузку цитоплазмы и ядер, соответственно. C1, PCC1; B2, PCB2. f , Конфокальная микроскопия иммунофлуоресцентного окрашивания после обработки клеток носителем (ДМСО) или РСС1. Применяли первичные антитела, специфичные к p53 или COX IV. Масштабная линейка 10 мкм. г , Иммуноблот-анализ клеток PSC27, подвергнутых действию различных агентов.Цито, цитоплазматический; мито, митохондриальный. ч , Анализ окрашивания JC-1, флуоресцентный зонд, показывающий Δψm. Сигналы измерялись в течение 3 дней. Зеленая флуоресценция указывает на мономеры JC-1 (они появляются в цитозоле после деполяризации митохондриальной мембраны и указывают на раннюю стадию апоптоза). Красная флуоресценция указывает на агрегацию JC-1 (находится в интактных митохондриях). Слева, репрезентативные изображения. Верно, статистика. И PCC1, и PCB2 использовали при 100 мкМ в соответствующих анализах. Данные в b , e — g являются репрезентативными для трех независимых экспериментов.Статистическая значимость в a (справа), d (справа) и h (справа) была рассчитана с использованием двухсторонних тестов t , а в c была рассчитана с помощью одностороннего дисперсионного анализа (дисперсионный анализ Даннета). контрольная работа). Данные на всех гистограммах показаны как среднее ± стандартное отклонение. и представляют три биологических повтора. NS, P > 0,05; * P <0,05; ** P <0,01.
Исходные данные
Высвобождение цитохрома c и нарушение митохондриальной мембраны — это внутриклеточные события, связанные с апоптозом, и часто действуют как прямые драйверы апоптоза 42 .Наши данные предполагают, что обработка PCC1 увеличивала высвобождение цитохрома c из митохондрий в окружающее цитоплазматическое пространство (рис. 4b и расширенные данные рис. 5h). Высвобождение цитохрома c из митохондрий в значительной степени согласуется с биохимическими реакциями, такими как активация каспазы в стареющих клетках, обработанных PCC1 (рис. 3g).
Члены семейства процианидинов обладают широким спектром фармакологических свойств, включая антиоксидантные и противовоспалительные, которые противоположны тем, которые мы наблюдали при лечении стареющих клеток с помощью PCC1.Текущие данные побудили нас задуматься, воспроизводятся ли эффекты PCC1 другими процианидинами. Процианидин B2 (PCB2) представляет собой типичный флавоноид, который существует в виде димера и снижает уровни ROS во время окислительного стресса в культивируемых клетках 43 . PCB2 не смог устранить стареющие клетки (фиг. 4c и дополнительные фиг. 3b и 4b) и не усилил продукцию ROS и не индуцировал митохондриальное высвобождение цитохрома c в стареющих клетках (фиг. 4d и расширенные данные фиг. 5i). Значительное количество p65 (RelA), одной из основных субъединиц комплекса NF-κB, перемещается в ядро стареющих клеток (рис.4д). Хотя обработка PCB2 противодействовала ядерной транслокации p65, что согласуется с его противовоспалительной способностью, этот эффект не воспроизводился PCC1 (рис. 4e). Стареющие клетки, подвергшиеся воздействию PCC1, демонстрировали заметное расщепление каспазой 3, тогда как клетки, обработанные PCB2, этого не делали, что дополнительно дифференцирует биологическую активность этих двух молекул процианидина (рис. 4e).
Как фактор, который функционально управляет судьбой клеток, p53 может индуцировать апоптоз либо путем трансактивации проапоптотических генов, либо независимым от транскрипции способом путем транслокации в митохондрии 44 .Мы наблюдали повышенную ядерную транслокацию p53 при клеточном старении, паттерн заметно снижается с помощью PCC1, но намного меньше, чем с помощью PCB2 (рис. 4e, f). Поскольку ядерное исключение p53 является критическим шагом в индукции апоптоза стареющих клеток 45 , мы дополнительно оценили распределение p53. Иммунофлуоресцентное окрашивание показало существенно увеличенное перекрытие p53 с субъединицей IV цитохром с-оксидазы (COX IV) (трансмембранный белковый комплекс в дыхательной электронной цепи митохондрий, часто используемый в качестве маркера резидентного белка митохондрий) в стареющих клетках, обработанных PCC1, что свидетельствует об усилении транслокации p53 в митохондриальный матрикс.Хотя мы наблюдали некоторое количество p53 в митохондриях пролиферирующих клеток, PCC1 не вызывал заметного или полного притока белка p53 в митохондриальный матрикс пролиферирующих клеток (рис. 4f). Однако в стареющих клетках уровни p53 снижались в ядрах, но повышались в митохондриях после воздействия PCC1 (рис. 4g).
Снижение митохондриального мембранного потенциала (Δψm) — это событие, которое может запускать апоптоз через митохондриально-опосредованный внутренний путь 46 .Мы обнаружили, что Δψm значительно снижается в стареющих клетках, в то время как пролиферирующие клетки остаются в основном незатронутыми в присутствии PCC1, на что указывает профиль сигналов зонда JC-1 (фиг. 4h). Таким образом, PCC1 способствует генерации АФК, запускает высвобождение цитохрома с и вызывает нарушение Δψm в стареющих клетках, события, по своей сути связанные с митохондриальной недостаточностью и функционально управляющим апоптозом клеток.
В совокупности наши экспериментальные данные предполагают, что стареющие клетки подвержены апоптозу, индуцированному PCC1, процессу, частично опосредованному активацией NOXA и PUMA и связанным с усилением продукции ROS и митохондриальной дисфункцией.
PCC1 способствует регрессии опухоли и снижает химиорезистентность.
Учитывая способность и избирательность PCC1 устранять стареющие клетки in vitro, мы затем исследовали, можно ли использовать этот агент для вмешательства против возрастных патологий in vivo. В клинической онкологии лекарственная устойчивость ограничивает эффективность большинства противоопухолевых методов лечения, в то время как стареющие клетки часто вносят вклад в терапевтическую резистентность за счет развития in vivo SASP в микросреде опухоли, поврежденной лекарственными средствами (TME) 15,16,47 .Фармакологическое устранение стареющих клеток, вызванных терапией, сводит к минимуму побочные эффекты химиотерапии и предотвращает рецидив рака у животных 48 . Однако возможность PCC1-опосредованного истощения стареющих клеток первичных опухолей для повышения эффективности противоопухолевого лечения остается в значительной степени неизвестной.
Во-первых, мы решили создать тканевые рекомбинанты путем смешивания клеток PSC27 с клетками PC3, которые являются типичной линией клеток рака простаты с высокой степенью злокачественности, в предварительно оптимизированном соотношении (1: 4) 14 .Затем клетки подкожно имплантировали в задний бок мышей с диабетом без ожирения и тяжелым комбинированным иммунодефицитом (NOD – SCID). Опухоли животных измеряли в конце 8-недельного периода, а ткани собирали для патологической оценки. По сравнению с опухолями, содержащими раковые клетки PC3 и наивные стромальные клетки PSC27, ксенотрансплантаты, состоящие из клеток PC3 и стареющих клеток PSC27, демонстрировали значительно увеличенный объем, подтверждая стимулирующий рост опухоли эффект стареющих клеток (расширенные данные, рис.6а).
Чтобы имитировать клинические условия, мы экспериментально разработали доклиническую схему, включающую генотоксические терапевтические препараты и / или сенолитики (рис. 5а). Через две недели после подкожной имплантации, когда наблюдалось стабильное поглощение опухолями in vivo, животным вводили однократную дозу митоксантрона (MIT, химиотерапевтический препарат) или плацебо в 1-й день 3-й, 5-й и 7-й недель до конца 8-недельный режим (расширенные данные рис. 6b). В отличие от группы, получавшей плацебо, введение MIT заметно задерживало рост опухоли, подтверждая эффективность MIT в качестве химиотерапевтического агента (44.0% уменьшение размера опухоли) (рис. 5b). Примечательно, что хотя введение PCC1 само по себе не приводило к уменьшению размера опухоли, лечение MIT с последующей доставкой PCC1 (в дозе 20 мг на кг посредством внутрибрюшинной (ip) инъекции через 2 недели после первой дозы MIT и затем вводимой раз в две недели) значительно усилило регрессию опухоли (55,2 % уменьшение размера опухоли по сравнению с одним MIT; уменьшение объема опухоли на 74,9% по сравнению с лечением плацебо) (рис. 5b).
Рис. 5: Сенолизис PCC1 в поврежденном TME снижает устойчивость к раку, обусловленную SASP.
a , Иллюстративная схема доклинической схемы. Через две недели после подкожной имплантации и поглощения тканевых рекомбинантов in vivo самцы мышей NOD-SCID получали либо одиночные (моно), либо комбинированные (двойные) агенты по метрономному графику, состоящему из нескольких циклов. BLI, биолюминесцентная визуализация. b , Статистическое профилирование конечных объемов опухоли. Клетки PC3 были ксенотрансплантаты отдельно или вместе с клетками PSC27 в задний бок животных. c , Сравнительная оценка старения in vivo путем окрашивания SA-β-Gal.Опухоли после умерщвления животных препарировали и обрабатывали как замороженные срезы для гистологического окрашивания. Шкала 200 мкм. d , Графики скрипки, отображающие сравнительную статистику окрашивания SA-β-Gal в опухолевых тканях. и , Анализ транскриптов для экспрессии in vivo нескольких канонических факторов SASP в стромальных клетках, выделенных из опухолей. Ткани животных, ксенотрансплантированных как стромальными, так и раковыми клетками, подвергали выделению с использованием микродиссекции с помощью лазерного захвата и последующим процессам.Данные являются репрезентативными для трех биологических повторов ( n = 10 животных на группу). Наборы данных отображаются в виде графиков в виде прямоугольников, на которых прямоугольник простирается от 25-го до 75-го процентиля, при этом медиана отображается в виде линии посередине, а усы указывают наименьшие и наибольшие значения. f , Профилирование транскриптов SASP в стромальных клетках. Сигналы, соответствующие каждому фактору, были нормализованы к сигналам из группы, обработанной носителем. Примечание p16 INK4a также известен как CDKN2A и p21 CIP1 также известен как CDKN1A . g , Статистическое измерение клеток с повреждением ДНК и апоптозных клеток в биопробах, собранных, как описано в a , b . Значения представлены как процент клеток, положительно окрашенных с помощью иммуногистохимии (ИГХ) антителами, специфичными к гистону γh3AX или каспазе 3 (расщепленные). Для b , d — g , P значения были рассчитаны с помощью двусторонних тестов t . h , Репрезентативные изображения IHC каспазы 3 (расщепленная, CC3) в конце терапевтических режимов.Шкала 100 мкм. i , Сравнительная выживаемость мышей, убитых после развития тяжелой тяжелой болезни. Продолжительность выживания рассчитывалась от времени инъекции рекомбинантной ткани до гибели животного. МС, медиана выживаемости. Значения P были рассчитаны с помощью двусторонних логранговых тестов (Мантела – Кокса). Данные в c , h являются репрезентативными для трех независимых экспериментов. Данные на всех гистограммах показаны как среднее ± стандартное отклонение. и представляют три биологических повтора.
Исходные данные
Затем мы проверили, происходит ли клеточное старение в опухолевых очагах этих животных. Неудивительно, что введение MIT вызывало появление большого количества стареющих клеток в опухолевой ткани. Однако доставка PCC1 этим обработанным химиотерапией животным истощила большинство стареющих клеток (рис. 5c, d). Микродиссекция с помощью лазерного захвата с последующими анализами транскриптов показала значительно повышенную экспрессию факторов SASP, включая IL6 , CXCL8 , SPINK1 , WNT16B (также известный как WNT16 ), CSF (также известный как ), MMP3 и IL1A , тенденция, сопровождающаяся активацией гена, кодирующего маркер старения p16 INK4a у животных, получавших химиотерапию (рис.5e и расширенные данные рис. 6c). Эти изменения в основном наблюдались в стромальных клетках, а не в соседних раковых клетках, что подразумевает возможность репопуляции остаточных раковых клеток, которые часто развивают приобретенную устойчивость в поврежденных лечением TME. Однако при введении PCC1 связанные с SASP изменения в значительной степени обращались вспять, как предполагают анализы транскриптов и RNA-seq (фиг. 5f и расширенные данные фиг. 6d).
Для исследования механизмов, лежащих в основе экспрессии SASP у мышей, получавших MIT, мы вскрыли опухоли у животных, получавших эти два агента, через 7 дней после доставки первой дозы GSE, момент времени до развития устойчивых колоний.В отличие от лечения плацебо, введение MIT увеличивало повреждение ДНК и апоптоз, тогда как лечение только PCC1 не усиливало (фиг. 5g). Однако, когда животным, получавшим MIT, одновременно вводили PCC1, повреждение ДНК и апоптоз значительно усиливались, что подразумевает усиление цитотоксичности у животных, получавших как химиотерапию, так и сенолитики. В качестве подтверждающего доказательства мы наблюдали повышенное расщепление каспазой 3, типичный признак клеточного апоптоза, когда PCC1 вводили вместе с MIT (рис. 5h).
Затем мы оценили последствия прогрессирования опухоли, сравнив выживаемость различных групп животных с течением времени. В этой доклинической когорте животных наблюдали на предмет роста опухоли, при этом считалось, что объемное заболевание возникло, как только опухолевая нагрузка стала заметной (размер ≥ 2000 мм 3 ), подход, использованный в предыдущих исследованиях 14,49 . Мыши, получавшие комбинаторную терапию MIT – PCC1, показали наиболее длительную медианную выживаемость, прожив по крайней мере на 48,1% дольше, чем в группе, получавшей только MIT (рис.5i, зеленый против синего). Однако лечение только PCC1 лишь незначительно увеличивало выживаемость. Наши данные предполагают, что одно только введение PCC1 не изменяет рост опухоли и не способствует выживанию животных, тогда как совместное введение PCC1 с MIT имеет значительный синергетический эффект.
Следует отметить, что лечение, проведенное в этих исследованиях, оказалось хорошо переносимым животными, поскольку не наблюдалось значительных изменений уровней мочевины, креатинина или печеночных ферментов или массы тела (расширенные данные, рис.6д, е). Что еще более важно, химиотерапевтические и геропротективные агенты, вводимые в дозах, оптимизированных в этом исследовании, не оказывали значительного воздействия на целостность иммунной системы или тканевый гомеостаз критических органов, даже у иммунокомпетентных мышей (дополнительный рис. 6a – c). Эти результаты подтверждают обоснование того, что антивозрастные агенты в сочетании с традиционной химиотерапией могут усиливать ответ опухоли, не вызывая серьезной системной токсичности.
Удаление стареющих клеток в результате лечения PCC1 уменьшает физическую дисфункцию
Даже небольшое количество стареющих клеток может вызвать физическую дисфункцию у молодых животных 50 .Мы спросили, может ли PCC1 избирательно убивать стареющие клетки in vivo и тем самым предотвращать физическую дисфункцию. Чтобы ответить на этот вопрос, мы выполнили параллельную имплантацию контрольных и стареющих эмбриональных фибробластов мыши (MEF, 0,5 × 10 6 клеток на сторону), конститутивно экспрессирующих люциферазу (LUC + ), подкожно сингенным мышам дикого типа (WT). Сразу после имплантации животных обрабатывали PCC1 (в дозе 20 мг на кг через внутрибрюшинную инъекцию) или носителем (этанол-полиэтиленгликоль 400-Phosal 50 пропиленгликоль (PG) в 10:30:60) в течение 7 дней (рис.6а). Мы обнаружили, что интенсивность люминесцентного сигнала была значительно ниже у мышей, которым имплантировали стареющие клетки и лечили PCC1, чем у мышей, получавших носитель из однопометников, хотя не наблюдали разницы после обработки мышей, которым трансплантировали контрольные клетки LUC + (рис. 6b, c). ), что подтверждает сенолитическую эффективность PCC1 in vivo.
Рис. 6: PCC1-опосредованный сенолиз предотвращает физическую дисфункцию и облегчает патологические симптомы.
a , Схема экспериментальных процедур трансплантации клеток и тестов физических функций на 5-месячных самцах мышей C57BL / 6J.b , типичные изображения, показывающие активность люциферазы in vivo через 2 дня после последней обработки мышей. Шкала линейки 20 мм. c , Люминесценция трансплантированных клеток в процентах относительно средних сигналов у животных, которым вводили носитель. d — f , Измерение максимальной скорости ходьбы (относительно базовой линии) ( d ), выносливости в висе ( e ) и силы захвата ( f ) у 5-месячных мышей-самцов C57BL / 6J , с тестами, проведенными через 1 месяц после последнего лечения. г , Схема экспериментального дизайна для трансплантации и измерения физических функций. h — j , Измерение максимальной скорости ходьбы (относительно базовой линии) ( h ), выносливости в висе ( i ) и силы захвата ( j ) у самцов мышей C57BL / 6J в возрасте 28 недель (Через 2 недели после последнего лечения). k , Годовые кривые выживаемости 17-месячных животных, которым имплантировали 0,5 × 10 6 контрольных клеток MEF и обрабатывали носителем (CTRL-носитель), и мышей, которым имплантировали 0.5 × 10 6 стареющих клеток MEF, обработанных либо носителем (SEN-носитель), либо PCC1 (SEN-PCC1). Красные стрелки, имплантация клеток (на 528-й день возраста) или конец измерения выживаемости (890-й день возраста). Значения P были рассчитаны с помощью двусторонних логранговых тестов (Мантела – Кокса). l , Сравнительная количественная оценка бремени болезни (слева) и бремени опухоли (справа) (показано как медиана с межквартильным размахом) после имплантации стареющих клеток и лечения носителем или PCC1. m , Причина смерти животных, которым вводили имплантированные клетки и лечили носителем или PCC1. Для d — f , h — j данные показаны в виде диаграмм с прямоугольными ячейками, на которых прямоугольники простираются от 25-го до 75-го процентиля, а медиана показана в виде линии посередине, а усы указывают наименьшие и наибольшие значения. Для c — f , h — j , P значения рассчитывались с помощью двусторонних тестов t .Количество животных, n = 5 на группу для c , n = 10 на группу для d — j , n = 27 для k и n = 13 для л , м . NS, P > 0,05; * P <0,05; ** P <0,01; *** P <0,001; **** П <0,0001.
Исходные данные
Затем мы исследовали, может ли уничтожение имплантированных стареющих клеток с помощью PCC1 ослабить патологические явления, в частности физическую дисфункцию.Обработка молодых животных PCC1 после имплантации стареющих клеток в течение 1 недели предотвратила снижение максимальной скорости ходьбы (RotaRod), выносливости при подвешивании (тест на подвешивание) и силы захвата (измеритель захвата), изменений, наблюдаемых в течение 1 месяца после обработки носителем другой группы мышей, несущих стареющие клетки, что согласуется с потенциалом PCC1 уменьшать физическую дисфункцию (Fig. 6d-f). Введение PCC1 также предотвращало физическую дисфункцию, которая возникала у животных через 5 недель после имплантации стареющих клеток (рис.6г). У мышей, несущих стареющие клетки, однократный 5-дневный курс лечения PCC1 улучшил физическую функцию по сравнению с лечением носителем (рис. 6h – j). Следует отметить, что улучшение было обнаружено через 2 недели после лечения PCC1 и даже длилось несколько месяцев (расширенные данные, рис. 7a, b). В эти два момента времени введения PCC1 (сразу по сравнению с 5 неделями после имплантации стареющих клеток) положительные эффекты PCC1, по-видимому, были сопоставимы. Данные предполагают, что график введения PCC1 может быть гибким, что указывает на его потенциальную клиническую осуществимость.Поскольку процианидины, полученные из семян растений, обычно имеют период полувыведения <12 часов 51,52 , такое устойчивое улучшение физических функций после однократного курса лечения PCC1 позволяет обойтись без непрерывного лечения сенолитическим агентом, что также означает, что активности PCC1 достаточно, чтобы предотвратить физическую дисфункцию, вызванную стареющими клетками.
Затем мы попытались оценить влияние стареющих клеток или пользу их устранения у животных среднего возраста.Для этой цели мы использовали 17-месячных мышей C57BL / 6J, которым имплантировали контрольные или стареющие MEF. Примечательно, что выживаемость животных, несущих стареющие клетки и получавших лечение носителем, в следующем году была значительно ниже, чем у животных, получавших лечение PCC1, с повышенным в 2,4 раза риском смерти (отношение рисков, P = 0,0172) (рис. 6k ). Однако бремя болезни, бремя опухоли на момент смерти и причины смерти существенно не различались между мышами, получавшими носитель, и мышами, получавшими PCC1 (рис.6л, м). Эти данные предполагают, что небольшое количество стареющих клеток может влиять на выживаемость посредством общего процесса, такого как ускорение прогрессирования старения, а не вызывая какую-либо конкретную патологию или несколько индивидуальных состояний. Увеличение количества стареющих клеток приводит к физической дисфункции, тенденции, которая связана со смертностью среднего возраста, но может быть отложена введением сенолитиков, таких как PCC1.
PCC1 поддерживает физические функции и продлевает выживание старых мышей
Сенолитики истощают стареющие клетки в различных тканях и органах в различных патофизиологических ситуациях, большинство из которых коррелирует со старением 53 .Для дальнейшего изучения влияния PCC1 на стареющие клетки организмов и старение организма мы выбрали две независимые животные модели старения in vivo, в том числе мышей, подвергшихся воздействию терапии, и мышей, стареющих естественным путем. Во-первых, мы индуцировали клеточное старение, подвергая мышей WT облучению всего тела (WBI) в сублетальной дозе (5 Гр), после чего следовала геропротекторная обработка PCC1 (20 мг на кг через внутрибрюшинную инъекцию) или носителем (этанол-полиэтилен). гликоль 400 – Phosal 50 PG в 10:30:60) (один раз в неделю) (Рис.7а). Следует отметить, что животные, которые подверглись WBI, демонстрировали ненормальный внешний вид тела, включая заметно поседевшие волосы, которые, однако, в значительной степени были устранены введением PCC1 (рис. 7b, c). SA-β-Gal-положительные стареющие клетки индуцировались in vivo у этих животных, о чем свидетельствует повышенная положительность окрашивания в сердечной и легочной тканях (фиг. 7d, e). Однако, когда мы лечили PCC1 i.p. инъекции, процент SA-β-Gal-позитивных клеток в рассеченных тканях был значительно снижен, в отличие от мышей, получавших носитель, на стадии после WBI (рис.7е, г). Обработка PCC1 также снижает экспрессию маркеров старения и подмножества ключевых факторов SASP по сравнению с обработкой носителем (фиг. 7h). В целом, данные свидетельствуют о том, что PCC1 может эффективно истощать SA-β-Gal-положительные клетки, контролировать экспрессию SASP и минимизировать нагрузку стареющими клетками в условиях in vivo у мышей.
Рис. 7: Обработка PCC1 облегчает физическую дисфункцию у животных, подвергшихся воздействию WBI.
a , Схема экспериментальной процедуры для мышей, испытывающих WBI и физические функциональные тесты.b , Сравнение снимков всего тела самцов мышей C57BL / 6J, которые были наивными, подвергались воздействию WBI с последующей обработкой носителем или подвергались воздействию WBI и лечились PCC1, соответственно. c , Изображение в клетке животных, описанных в a , в доклинических условиях. d , Типичные изображения окрашивания SA-β-Gal сердечной ткани необработанных (наивных) мышей и мышей, получавших WBI, подвергнутых обработке носителем или PCC1. Масштабная линейка 200 мкм. e , Типичные изображения окрашивания SA-β-Gal легочной ткани мышей, как описано в d .Масштабная линейка 200 мкм. f , Сравнительная статистика окрашивания SA-β-Gal сердечной ткани животных, исследованных в d . g , Сравнительная статистика окрашивания SA-β-Gal легочной ткани животных, исследованных в e . h . Количественное измерение экспрессии SASP на уровне транскрипции в тканях, взятых у животных, получавших лечение в условиях, описанных в a . i , j , Измерение дистанции бега на беговой дорожке ( i ) и силы захвата ( j ) экспериментальных мышей.Для f — j , P значения рассчитывались с помощью двусторонних тестов t . k , анализ выживаемости Каплана-Мейера мышей C57BL / 6J, подвергшихся воздействию WBI и получавших еженедельно носитель или PCC1, с наивными мышами в качестве необработанного контроля. ДИ — доверительный интервал; ЧСС, степень опасности; DFI, безрецидивный интервал. Значения P были рассчитаны с помощью двусторонних логранговых тестов (Мантела – Кокса). Данные на гистограммах показаны как среднее ± стандартное отклонение. и представляют три независимых эксперимента.NS, P > 0,05; * P <0,05; ** P <0,01; *** P <0,001; **** П <0,0001.
Исходные данные
Затем мы оценили влияние доклинического лечения на физические параметры мышей. Как и ожидалось, WBI значительно снизил нагрузочную способность и мышечную силу, измеренную с помощью тестов на беговой дорожке и силы захвата в группе с транспортным средством (рис. 7i, j). Напротив, введение PCC1 дало существенную пользу, восстанавливая эти возможности.Что еще более важно, лечение PCC1 увеличивало выживаемость (рис. 7k). Наши результаты показывают, что вызванное PCC1 устранение SA-β-Gal-позитивных стареющих клеток может быть эффективной стратегией для облегчения физического регресса, связанного со старением, и снижения смертности в условиях преждевременного старения, вызванного факторами окружающей среды, такими как цитотоксическая терапия.
Затем мы попытались определить влияние стареющих клеток на физические функции у естественно стареющих животных. С этой целью мы обрабатывали нормальных 20-месячных мышей WT носителем (этанол – полиэтиленгликоль 400 – Phosal 50 PG в 10:30:60) или PCC1 (20 мг на кг через i.п. инъекция) (1 раз в 2 недели) в течение 4 месяцев (рис. 8а). Гистологическая оценка выявила значительно повышенный процент SA-β-Gal-положительных стареющих клеток в почках, печени, легких и предстательной железе у пожилых животных, что было устранено обработкой PCC1 (рис. 8b, c и расширенные данные, рис. 8a-f). ). Результаты физических испытаний показали, что PCC1 облегчает физическую дисфункцию за счет увеличения максимальной скорости ходьбы, выносливости при висении, силы захвата, выносливости на беговой дорожке, дневной активности и показателей баланса пучка у животных, которым вводили PCC1, по сравнению с животными, получавшими носитель (рис.8d – i), масса тела и потребление пищи оставались в значительной степени неизменными у мышей, получавших PCC1 (расширенные данные, рис. 8g, h). Примечательно, что экспрессия SASP была значительно снижена в тканях, таких как легкие старых мышей, получавших PCC1, по сравнению с таковой в группе, получавшей носитель (рис. 8j), что соответствует меньшей секреции факторов SASP обработанными стромальными тканями человека. с PCC1 (рис. 5е).
Рис. 8: Прерывистое введение PCC1 продлевает как продолжительность здоровья, так и продолжительность жизни старых мышей.
a , Схематический дизайн для физического осмотра 20-месячных мышей-самцов C57BL / 6J, получавших PCC1 один раз каждые 2 недели (раз в две недели) в течение 4 месяцев. b , Типичные изображения окрашивания почек SA-β-Gal у молодых и старых мышей, получавших носитель или PCC1. Масштабная линейка 200 мкм. c , Количественная оценка окрашивания SA-β-Gal, как описано в b . Данные представляют собой среднее значение ± стандартное отклонение. d — h , Количественная оценка максимальной скорости ходьбы (относительно базовой) ( d ), выносливости в висе ( e ), силы захвата ( f ), выносливости на беговой дорожке ( g ) и ежедневной активности ( ч ) 20-месячных мышей-самцов C57BL / 6J после 4-месячного лечения. i , Количественное определение времени, необходимого для пересечения балансира. Точки данных до и после лечения каждого животного соединяются для прямого сравнения эффектов лечения. j , Количественное профилирование транскриптов экспрессии SASP в тканях легких, собранных у 6-месячных необработанных (6M), 24-месячных мышей, получавших носитель (24M-носитель), и 24-месячных мышей, получавших PCC1 (24M -PCC1). Данные представлены как среднее ± стандартное отклонение. и были получены из трех биологических повторов ( n = 3 независимых анализа). k , Схематический дизайн для анализа продолжительности жизни мышей (обоих полов) в возрасте 24–27 месяцев. l , m , кривые выживаемости после лечения ( l ) и выживаемости в течение всей жизни ( m ) животных C57BL / 6J, которым каждые две недели вводили PCC1 ( n = 91; 48 самцов, 43 самки) или транспортное средство ( n = 80; 42 мужчины, 38 женщин), начиная с возраста 24–27 месяцев. n , максимальная скорость ходьбы и выносливость в зависании, усредненные за последние 2 месяца жизни ( n = 10 мышей на группу) и продолжительность жизни самых долгоживущих мышей (первые 20 мышей) в обеих группах. o , Бремя болезней и опухолей на момент смерти. Для обоих полов n = 60 мышей на руку. Для мужчин n = 31 для PCC1 и n = 33 для автомобиля. Для женщин n = 29 для PCC1 и n = 27 для транспортного средства. Для c — h , j , n = 3 биологически независимых анализа. Данные отображаются в виде диаграмм в виде прямоугольников, на которых прямоугольник простирается от 25-го до 75-го процентиля, при этом медиана отображается в виде линии посередине, а усы указывают на наименьшие и наибольшие значения ( d — h , n ) или как среднее ± s.d. ( o ). Для определения статистической значимости использовались непарные двусторонние тесты t ( c — j , n , o ) и модели регрессии пропорциональных рисков Кокса ( l , m ).
Исходные данные
Чтобы установить потенциал элиминации стареющих клеток для увеличения оставшейся продолжительности жизни мышей WT, мы провели лечение PCC1, начиная с очень пожилого возраста (рис. 8k). У мышей, получавших введение PCC1 (один раз в 2 недели или раз в две недели), начиная с возраста 24–27 месяцев (что примерно соответствует возрасту 75–90 лет у людей), было 64.На 2% больше медиана продолжительности жизни после лечения (или на 9,4% больше общей продолжительности жизни) и более низкий риск смертности (65,0%, P <0,0001), чем в группе, получавшей носитель (рис. 8l, m). Эти данные показывают, что PCC1 может значительно снизить риск возрастной смертности у старых мышей.
Затем мы спросили, не произошло ли снижение смертности старых животных за счет увеличения заболеваемости в позднем возрасте. Мы измеряли физическую функцию у экспериментальных мышей, получавших PCC1 или носитель ежемесячно до смерти.Несмотря на более длительную оставшуюся продолжительность жизни у мышей, получавших PCC1, физическая функция в последние 2 месяца жизни не была значительно ниже, чем у мышей, получавших носитель (фиг. 8n). После вскрытия частота нескольких возрастных патологий, опухолевой нагрузки и причин смерти существенно не различалась у мышей, получавших PCC1, и мышей, получавших носитель (фиг. 8o и расширенные данные, фиг. 9a, b). Однако экспрессия SASP была снижена в твердых органах, что было в значительной степени совместимо со снижением циркулирующих уровней интерлейкина (IL) -6, колониестимулирующего фактора (CSF) 2 и моноцитарного хемоаттрактантного белка (MCP) 1, репрезентативных маркеров SASP. в периферической крови (расширенные данные рис.9c – f). Мы также наблюдали снижение экспрессии SASP в CD3 + Т-клетках в периферической крови (расширенные данные, рис. 9g), клеточной линии, которая демонстрирует устойчивое увеличение экспрессии p16 INK4a во время старения человека 54 . Кроме того, обработка PCC1 снижает окислительный стресс в тканях печени, о чем свидетельствует уменьшение аддуктов продукта перекисного окисления липидов 4-гидроксиноненаля (HNE) и увеличение соотношения восстановленного и окисленного глутатиона (расширенные данные, рис.9h, i), что согласуется с общими свойствами флавоноидов, которые проявляют антиоксидантную активность, противодействуя свободным радикалам и задействуя систему антиоксидантной защиты 55,56 .
Таким образом, сенолитический агент PCC1, фитохимический компонент, полученный из GSE (или, альтернативно, в меньшем количестве, из натуральных продуктов, таких как экстракты корицы, какао, яблочной кожуры и сосновой коры), может уменьшить бремя старения и, возможно, другие клетки развивают провоспалительный фенотип и по своей природе зависят от антиапоптотических путей, связанных со старением, способствующими выживанию, и увеличивают продолжительность жизни после лечения, не вызывая повышенной заболеваемости у мышей.Настоящим мы представляем принципиальные доказательства того, что даже при применении в пожилом возрасте такой терапевтический метод обладает значительным потенциалом для значительного замедления возрастной дисфункции, уменьшения возрастных заболеваний и улучшения состояния здоровья, тем самым открывая новые возможности для улучшения продолжительность здоровья и продолжительность жизни в гериатрической медицине будущего.
Термогенез, индуцированный водой | Журнал клинической эндокринологии и метаболизма
Аннотация
Питье большого количества воды обычно используется в схемах похудания и считается полезным для здоровья; тем не менее, несколько систематических исследований посвящены этому понятию.У 14 здоровых субъектов с нормальным весом (семь мужчин и семь женщин) мы оценили влияние употребления 500 мл воды на расход энергии и скорость окисления субстрата с помощью непрямой калориметрии в помещении. Влияние питья воды на метаболизм жировой ткани оценивали методом микродиализа. Употребление 500 мл воды увеличивает скорость обмена веществ на 30%. Увеличение происходило в течение 10 мин и достигало максимума через 30–40 мин. Общий термогенный ответ составил около 100 кДж. Около 40% термогенного эффекта было вызвано нагреванием воды с 22 до 37 C.У мужчин липиды в основном способствовали увеличению скорости метаболизма. Напротив, у женщин углеводы в основном использовались в качестве источника энергии. Увеличение расхода энергии с водой уменьшалось при системной блокаде β-адренорецепторов. Таким образом, употребление 2 литров воды в день увеличит расход энергии примерно на 400 кДж. Следовательно, термогенный эффект воды следует учитывать при оценке расхода энергии, особенно во время программ похудания.
«Пейте много воды и придерживайтесь расписания» — старая поговорка о здоровье.Тем не менее, немного данных подтверждают эту проповедь. Недавние исследования показали, что питьевая вода действительно вызывает значительную физиологическую реакцию. Например, употребление воды повышает систолическое артериальное давление более чем на 30 мм рт. Ст. У пациентов с тяжелой вегетативной недостаточностью. Прессорный ответ проявлялся в течение 5 минут, достигал максимума примерно через 35 минут и сохранялся более 60 минут (1, 2). Питье воды умеренно повышает кровяное давление у пожилых, но не у более молодых контрольных субъектов (2).Прессорный ответ может быть опосредован симпатической нервной системой (2–4). У здоровых людей употребление воды увеличивает трафик симпатических нервов в мышцах (3) и концентрацию норэпинефрина в венозной плазме (2, 3, 5). Кроме того, прессорный ответ можно устранить с помощью системной ганглионарной блокады (2). Симпатическая нервная система играет важную роль в регулировании энергетического обмена и использования топлива. Симпатическая активация увеличивает клеточное поглощение глюкозы и метаболизм, а также стимулирует липолиз (6).Мы проверили гипотезу о том, что симпатический стимул, вызванный употреблением воды, может увеличить скорость метаболизма. Кроме того, мы определили влияние питья воды на мобилизацию и использование топлива на системном уровне и на уровне жировой ткани у нормальных мужчин и женщин. Наконец, мы пришли к выводу, что метаболический эффект воды можно ослабить с помощью системной или местной блокады β-адренорецепторов.
Объекты и методы
Субъектов
Четырнадцать здоровых субъектов [семеро мужчин: возраст 29 ± 3 года; индекс массы тела (ИМТ), 24.20 ± 0,94 кг / м 2 ; семь женщин: возраст 27 ± 2 года; ИМТ 20,80 ± 0,88 кг / м 2 ; P <0,05 для ИМТ] участвовали в метаболических исследованиях. У восьми здоровых испытуемых (двое мужчин и шесть женщин; возраст 25 ± 1 год) мы определили влияние питья воды на осмолярность плазмы. Все испытуемые не курили и не употребляли наркотики. Наблюдательный совет учреждения одобрил все исследования, и до включения в исследование было получено письменное информированное согласие.
Протокол
Субъекты не ели 12.За 5 ч до исследования и за 1,5 ч до тестирования. Были проведены три отдельных исследования. В первом исследовании мы оценили влияние употребления 500 мл воды на расход энергии и скорость окисления субстрата с помощью косвенной калориметрии. Во втором исследовании мы использовали метод микродиализа, чтобы охарактеризовать влияние употребления 500 мл воды на кровоток и метаболизм жировой ткани. В подгруппе исследования проводили дважды: один раз после приема плацебо и один раз после приема 100 мг блокатора β-адренорецепторов метопролола (Stada Arzneimittel AG, Бад-Вильбель, Германия).Лекарства принимали вслепую за 1 час до питья воды. В третьем исследовании мы определяли осмолярность венозной плазмы на исходном уровне и через 30 и 60 минут после приема воды.
Калориметрия
Поглощение кислорода и производство углекислого газа измеряли с помощью дыхательной камеры для оценки изменений в расходе энергии, дыхательном коэффициенте (RQ; произведено CO 2 / потреблено O 2 ), а также скорости окисления углеводов и липидов, соответственно.В предыдущих исследованиях с участием 16 здоровых субъектов максимальное спонтанное изменение скорости метаболизма за 3-часовой период составляло 0,2 ± 0,09 кДж / мин (3%). На протяжении всего исследования испытуемые оставались сидя. После 15-минутной приработки определяли расход энергии в покое в течение 30 минут. Затем испытуемые выпили 500 мл воды (22 ° C). В подгруппе мы также проверили эффект 500 мл теплой воды 37 ° C. После завершения питья измерения продолжали еще 90 мин.
Микродиализ
Исследования микродиализа проводили в положении лежа на спине, как описано ранее (7, 8).Вкратце, один (системная блокада β-адренорецепторов) или два (местная блокада β-адренорецепторов) микродиализных зондов вводили в подкожно жировую ткань на уровне пупка. Перед введением зондов соответствующая область подвергалась поверхностной анестезии кремом EMLA (AstraZeneca GmbH, Ведель, Германия). В день системной блокады β-адренорецепторов субъекты принимали метопролол (Stada Arzneimittel AG) перед тестированием, а зонд перфузировали раствором Рингера (Serumwerke Bernburg AG, Бернбург, Германия) с добавлением 50 мм этанола (B.Braun Melsungen AG, Melsungen, Германия) для мониторинга изменений кровотока и аскорбата 10 мкм (Jenapharm GmbH & Co. KG, Йена, Германия). В день местной блокады β-адренорецепторов субъекты принимали плацебо перед тестированием, а в перфузат для одного микродиализного зонда добавляли 100 нМ неселективного блокатора β-адренорецепторов пропранолола (Obsidan; ALPHARMA-ISIS, Langenfeld, Германия). Использовали датчики для микродиализа CMA / 60 и насосы для микродиализа CMA / 102 (оба от CMA Microdialysis AB, Солна, Швеция).Скорость потока составляла 2 мкл / мин.
Аналитические методы
Концентрация этанола определялась в перфузате (приток) и диализате (отток) с помощью стандартного ферментативного анализа (6). Концентрации глицерина, глюкозы и лактата в диализате определяли стандартизированными ферментативными колориметрическими методами с использованием автоматического анализатора CMA / 600 (CMA Microdialysis AB). Осмолярность плазмы определяли методом снижения точки замерзания (Осмометр Model A, Precision Scientific, Винчестер, Вирджиния).
Расчеты и статистика
Энергозатраты и скорости окисления субстрата рассчитывались по Ферраннини (9). Изменения кровотока определяли с использованием метода разбавления этанолом, основанного на принципе Фика (9–11). Соответственно, уменьшение соотношения этанола в диализате и перфузате [(EtOH) d / (EtOH) p ] соответствует увеличению кровотока и наоборот. Концентрацию глицерина в диализате измеряли для оценки изменений липолиза и / или мобилизации липидов (12).Концентрации глюкозы и лактата в диализате определяли для характеристики поступления глюкозы и гликолиза, соответственно. В предыдущих исследованиях было обнаружено, что in situ и извлечение глицерина, глюкозы и лактата в диализате составили примерно 30% при использовании почти равновесного диализа при 0,3 мкл / мин (13). Все данные представлены как средние ± средн. Статистический анализ проводился с помощью ANOVA с повторными измерениями с использованием блокады β-адренорецепторов или без блокады β-адренорецепторов и времени в качестве факторов для определения значимости различий в энергетическом метаболизме, гемодинамике и метаболическом ответе жировой ткани на воду у мужчин с нормальным весом и женщины соответственно.Для тестирования использовалась статистическая программа (InStat, версия 3.0; Graphpad Software Inc., Сан-Диего, Калифорния). Значимые коэффициенты F из дисперсионного анализа сопровождались апостериорными сравнениями между средними значениями с использованием множественного критерия Бонферрони t .
Результаты
Энергетический обмен
Расход энергии в покое составил 5,06 ± 0,30 кДж / мин у мужчин и 4,02 ± 0,17 кДж / мин у женщин ( P <0,001, мужчины против .женщины). Через 10 мин после питья воды расход энергии начал увеличиваться. Через шестьдесят минут после питья воды расход энергии увеличился на 30% у мужчин и на 30% у женщин (рис. 1A, P <0,001). RQ в состоянии покоя составлял 0,841 ± 0,013 и 0,794 ± 0,009 у мужчин и женщин, соответственно (рис. 1B, P <0,01, мужчины против женщин). У женщин RQ существенно не изменился в течение 30 минут после питья воды (рис. 1B). Через 40 минут RQ значительно снизился до минимума 0.75. Резкое снижение RQ сопровождалось увеличением до 0,88 между 50 и 90 мин (рис. 1B). Напротив, у мужчин RQ снизился до 0,79 через 30 минут и оставался на этом уровне в течение следующих 30 минут (рис. 1B). RQ приблизился к исходному значению через 90 мин (рис. 1B). Скорость окисления углеводов существенно не изменилась у мужчин в течение 90 мин после питья воды (рис. 1С). Напротив, у женщин окисление углеводов увеличивалось примерно в 2 раза (незначительно) в течение первых 50 минут после питья воды и примерно в 3 раза ( P <0.05) в течение следующих 40 минут после питья воды. (Рис. 1C). В течение первых 40 мин скорость окисления липидов увеличивалась как у мужчин (+ 100%), так и у женщин (+ 50%). В течение следующих 30 минут скорость окисления липидов оставалась повышенной у мужчин, тогда как у женщин она снизилась до исходных значений (рис. 1D). Через 90 минут скорость окисления липидов все еще была повышена у мужчин, тогда как у женщин она снизилась ниже исходных значений (рис. 1D).
Рис. 1.
Изменения в расходе энергии (EE), RQ, скорости окисления углеводов (COX) и скорости окисления липидов (LOX) после употребления 500 мл воды (22 ° C).Значения при t = 0 мин относятся к исходному уровню (до питья воды). Данные представлены как средние значения ± стандартная ошибка (n = 7 как для мужчин, так и для женщин). ЭЭ увеличивалось примерно на 30% как у мужчин, так и у женщин ( P <0,001) после употребления воды. RQ в состоянии покоя был значительно выше у мужчин против . женщины ( P <0,05). Через шестьдесят минут после питья воды RQ был значительно ниже у мужчин, но значительно выше у женщин по сравнению с исходным уровнем ( P <0,05). ЦОГ был значительно повышен у женщин ( P <0.001), но не изменяется у мужчин после питья воды. Напротив, LOX был значительно увеличен у мужчин ( P <0,001), но не изменился у женщин после употребления воды.
Рис. 1.
Изменения в расходе энергии (EE), RQ, скорости окисления углеводов (COX) и скорости окисления липидов (LOX) после употребления 500 мл воды (22 ° C). Значения при t = 0 мин относятся к исходному уровню (до питья воды). Данные представлены как средние значения ± стандартная ошибка (n = 7 как для мужчин, так и для женщин). ЭЭ увеличилась примерно на 30% как у мужчин, так и у женщин ( P <0.001) после питья воды. RQ в состоянии покоя был значительно выше у мужчин против . женщины ( P <0,05). Через шестьдесят минут после питья воды RQ был значительно ниже у мужчин, но значительно выше у женщин по сравнению с исходным уровнем ( P <0,05). ЦОГ был значительно повышен у женщин ( P <0,001), но не изменился у мужчин после употребления воды. Напротив, LOX был значительно увеличен у мужчин ( P <0,001), но не изменился у женщин после употребления воды.
В подгруппе добровольцев (n = 7) было протестировано влияние системной блокады β-адренорецепторов на вызванное водой увеличение расхода энергии. Опять же, все семь испытуемых показали увеличение расхода энергии с 292 ± 21 до 359 ± 23 кДж / ч (рис. 2А). У шести пациентов блокада β-адренорецепторов почти полностью предотвратила увеличение расхода энергии после употребления воды (рис. 2B). У одной женщины энергия была лишь немного ослаблена блокадой β-адренорецепторов.
Рис.2.
Изменения в расходе энергии (EE) после употребления 500 мл воды (22 C) отдельно (A) или при системной β-адренергической блокаде метопрололом (B). Приведены кумулятивные значения за 1 час. Данные представлены как средние значения ± se. *, P <0,05 по сравнению с исходным уровнем (остальные).
Рис. 2.
Изменения в расходе энергии (ЭЭ) после употребления только 500 мл воды (22 C) (A) или при системной β-адренергической блокаде метопрололом (B). Приведены кумулятивные значения за 1 час. Данные представлены как средние значения ± se.*, P <0,05 по сравнению с исходным уровнем (остальные).
На другой подгруппе добровольцев (n = 4) было протестировано влияние температуры воды на вызванное водой увеличение расхода энергии. Изменение расхода энергии, вызванное водой, составило около 70 кДж при 22 ° C и около 40 кДж при 37 ° C, т.е. разница между двумя температурами составляет около 30 кДж. (Рис. 3). Употребление воды вызывало стойкое снижение осмолярности вен. Осмолярность плазмы составила 296 ± 1 мосмоль / л до питья воды и 289 ± 1 мосмоль / л после питья воды ( P <0.01).
Рис. 3.
Влияние температуры воды (22 C или 37 C) на изменение расхода энергии после употребления 500 мл воды. Приведены кумулятивные значения за 1 час. Данные представлены как средние значения ± se. *, P <0,05, 22 C против . 37 C.
Рис. 3.
Влияние температуры воды (22 C или 37 C) на изменение расхода энергии после употребления 500 мл воды. Приведены кумулятивные значения за 1 час. Данные представлены как средние значения ± se.*, P <0,05, 22 C против . 37 C.
Метаболизм жировой ткани
Исходное соотношение этанола составляло 0,39 ± 0,03 и 0,29 ± 0,05 ( P <0,05, мужчины против . женщин) в жировой ткани мужчин и женщин, соответственно. Питьевая вода не влияла на соотношение этанола. Кроме того, соотношение этанола оставалось неизменным во время как системной, так и местной блокады β-адренорецепторов (данные не показаны). Исходный уровень глюкозы в диализате был равен 0.76 ± 0,14 и 1,02 ± 0,21 ммоль / л ( P <0,05, мужчины против . женщин) у мужчин и женщин соответственно. Эти значения существенно не изменились после употребления воды в обеих группах ни в отсутствие, ни в присутствии местной или системной блокады β-адренорецепторов (данные не показаны). Исходный уровень лактата диализата составлял 0,42 ± 0,12 и 0,47 ± 0,08 ммоль / л у мужчин и женщин, соответственно. Питье воды вызвало значительное увеличение лактата диализата у мужчин на 45% (рис. 4).Интересно, что это увеличение было почти полностью предотвращено системной, а не местной блокадой β-адренорецепторов (рис. 4). Напротив, лактат диализата не изменился у женщин после питья воды. Однако содержание лактата в диализате повышалось на 50% у женщин при наличии системной, но не местной блокады β-адренорецепторов (рис. 4). Исходный уровень глицерина в диализате составлял 58 ± 15 и 96 ± 16 мкмоль / л у мужчин и женщин, соответственно ( P <0,05, у мужчин против у женщин). После питья воды у мужчин отмечалось небольшое, но незначительное повышение содержания глицерина в диализате (+ 20%), чему препятствовала системная, а не локальная блокада β-адренорецепторов (рис.4). Однако у женщин изменений содержания глицерина в диализате не наблюдалось (рис. 4).
Рис. 4.
Относительные изменения лактата диализата и глицерина в жировой ткани у мужчин (n = 7) и у женщин (n = 7) после употребления 500 мл воды (22 C). CTRL, Control, только решение Рингера; BBL, местная β-адренергическая блокада 100 нм пропранололом, добавленным в перфузионную среду; BBS, системная β-адренергическая блокада при приеме внутрь 100 мг метопролола. Данные представлены как средние значения ± se.У мужчин содержание лактата в диализате значительно увеличивалось ( P <0,01) после употребления воды, и этому эффекту препятствовала системная, но не местная β-адренергическая блокада. Напротив, у женщин уровень лактата диализата существенно не изменился после употребления только воды, но значительно увеличился ( P <0,05) при наличии системной β-адренергической блокады. Уровень глицерина в диализате увеличился незначительно, но незначительно у мужчин. Однако этого увеличения не наблюдалось во время β-адренергической блокады.У женщин вообще не наблюдалось никаких изменений в диализатном глицерине при любом используемом протоколе.
Рис. 4.
Относительные изменения лактата диализата и глицерина в жировой ткани у мужчин (n = 7) и у женщин (n = 7) после употребления 500 мл воды (22 C). CTRL, Control, только решение Рингера; BBL, местная β-адренергическая блокада 100 нм пропранололом, добавленным в перфузионную среду; BBS, системная β-адренергическая блокада при приеме внутрь 100 мг метопролола. Данные представлены как средние значения ± se. У мужчин уровень лактата диализата значительно увеличился ( P <0.01) после питья воды, и этот эффект был предотвращен системной, но не местной β-адренергической блокадой. Напротив, у женщин уровень лактата диализата существенно не изменился после употребления только воды, но значительно увеличился ( P <0,05) при наличии системной β-адренергической блокады. Уровень глицерина в диализате увеличился незначительно, но незначительно у мужчин. Однако этого увеличения не наблюдалось во время β-адренергической блокады. У женщин вообще не наблюдалось никаких изменений в диализатном глицерине при любом используемом протоколе.
Обсуждение
Новым открытием этого исследования является то, что употребление 500 мл воды увеличивает скорость метаболизма на 30% как у мужчин, так и у женщин. Увеличение скорости метаболизма наблюдалось через 10 мин после завершения и достигало максимума через 30–40 мин после употребления воды. Эффект держался более часа. Сердечно-сосудистые изменения после употребления воды, которые мы описали ранее, имели аналогичную динамику (1–3, 10–12). Основываясь на наших измерениях, мы оцениваем, что потребление воды увеличивается на 1.5 литров увеличивают ежедневный расход энергии примерно на 200 кДж. За год расход энергии увеличится на 73 000 кДж (17 400 ккал), что составляет 2,4 кг жировой ткани. Для сравнения, прием 50 мг эфедрина трижды в день увеличивает расход энергии примерно на 320 кДж / день (13). Субстраты, которые способствовали увеличению скорости метаболизма, различались у мужчин и женщин. У мужчин употребление воды привело к заметному усилению окисления липидов. Окисление углеводов не изменилось после питья воды.Напротив, у женщин углеводы в основном способствовали увеличению скорости метаболизма после употребления воды.
Наши данные убедительно свидетельствуют о том, что увеличение скорости метаболизма с водой связано с активацией симпатической нервной системы и повышенной стимуляцией β-адренорецепторов. Действительно, максимальное увеличение скорости метаболизма после употребления воды соответствует максимальной активации симпатической нервной системы в предыдущих исследованиях (2, 3). Системная блокада β-адренорецепторов существенно ослабляла вызванное водой повышение скорости метаболизма.Основываясь на этом наблюдении, можно предположить, что употребление воды во время блокады β-адренорецепторов может снизить температуру тела. К сожалению, мы не определяли температуру тела. Мы предполагаем, что сужение сосудов конечностей после питья воды (3) может быть достаточным для поддержания теплового гомеостаза даже при отсутствии увеличения скорости метаболизма.
Мы использовали метод микродиализа для мониторинга метаболических изменений как на системном, так и на тканевом уровне. Питье воды не повлияло на кровоток в жировой ткани, что было определено методом разбавления этанолом (7, 8, 14, 15).Следовательно, изменение концентрации метаболитов после употребления воды нельзя объяснить локальными изменениями кровотока. Нам было особенно интересно узнать, получены ли окисленные липиды из п / к. У мужчин содержание интерстициального глицерина значительно увеличивается после употребления воды. Ответ был устранен системной, но не местной блокадой β-адренорецепторов. Таким образом, у мужчин употребление воды увеличивает мобилизацию липидов за счет стимуляции β-адренорецепторов. Однако липиды не происходят из подкожной жировой ткани брюшной полости.У женщин содержание глицерина в диализате не изменялось после питья воды независимо от наличия или отсутствия блокады β-адренорецепторов.
Концентрация глюкозы в диализате не изменилась после питья воды. Это наблюдение предполагает, что баланс между поступлением глюкозы и использованием глюкозы в жировой ткани не изменился с водой. У мужчин уровень лактата диализата увеличился, хотя системное окисление углеводов не увеличилось. Увеличение лактата согласуется с усилением гликолиза.Эффект подавлялся системной, но не местной блокадой β-адренорецепторов. Таким образом, лактат не образуется в подкожно жировой ткани. Мы предполагаем, что увеличение выработки лактата может быть результатом увеличения высвобождения глюкозы из печени, которое подавляется блокадой β-адренорецепторов. У женщин уровень лактата диализата не увеличивался. Предположительно, глюкоза более полно окислялась у женщин, о чем свидетельствует усиление окисления углеводов. Как ни парадоксально, уровень лактата диализата повышается после питья воды во время системной, но не местной блокады β-адренорецепторов.Возможно, снижение мобилизации и окисления липидов сопровождалось усилением гликолиза. Гендерный эффект воды может быть связан с различиями в составе тела или гормональными факторами (16).
Механизм, который вызывает симпатическую активацию при питье воды, остается неясным (17). На реакцию прессора, похоже, не влияет температура воды (2). Частично увеличение расхода энергии могло быть связано с энергией, необходимой для нагрева воды от комнатной температуры до температуры тела (500 мл × 15 C = 7500 кал = 30 кДж).Расчетные затраты энергии, связанные с нагревом воды, точно соответствовали разнице между термогенным эффектом воды 22 ° C и воды 37 ° C в наших исследованиях метаболической камеры. Таким образом, примерно 60–70% термогенеза, вызванного водой, нельзя отнести к нагреванию проглоченной воды. Растяжение желудка увеличивает симпатическую активность у людей (18). Однако во время максимальной реакции в желудке остается менее 125 мл воды (19). Мы наблюдали умеренное, но, тем не менее, стойкое снижение осмолярности плазмы после употребления воды, что отражало динамику метаболической реакции.У людей инфузия гипоосмолярных растворов через желудочный зонд вызывает большее усиление потоотделения, симпатический ответ, чем инфузия изосмолярных растворов (20). Возможно, симпатическая активация при питье воды включает осморецептивные или чувствительные к натрию афферентные нервные волокна (21, 22). Одним из важных выводов нашего исследования является то, что влияние воды на расход энергии и использование топлива следует признать мощным смешивающим фактором в метаболических исследованиях. Действительно, термогенез, вызванный питьем воды, является важным и непризнанным компонентом ежедневных затрат энергии.Если это подтвердится в других исследованиях, это бесплатное вмешательство может быть полезным дополнительным лечением у людей с избыточным весом и ожирением для увеличения расхода энергии.
Благодарности
Эта работа была частично поддержана Deutsche Forschungsgemeinschaft. J.J. является стипендиатом Гельмгольца Центра молекулярной медицины Макса-Дельбрюка.
Сокращения:
-
BMI,
-
RQ,
1
Jordan
J
,
Shannon
JR
,
Grogan
E
,
Biaggioni
000
000
000 Biaggioni
1999
Сильный прессорный ответ, вызванный питьевой водой.
Ланцет
353
:
723
2
Иордания
J
,
Шеннон
JR
,
Черный
BK
,
Али
Y
Farley3
F
,
Diedrich
A
,
Robertson
RM
,
Biaggioni
I
,
Robertson
D
at 2000
?
Обращение
101
:
504
—
509
3
Скотт
EM
,
Гринвуд
JP
,
Гилби
SG
9000 9000
DA Stoker
,
DA
2001
Употребление воды увеличивает симпатические сосудосуживающие выделения у нормальных людей.
Clin Sci Colch
100
:
335
—
342
4
Резервуар
J
,
Schroeder
C
,
Stoffels
M
,
D
AM
,
Luft
FC
,
Jordan
J
2003
Давление от питья воды у пациентов с тетраплегией может быть спинальным рефлексом.
Гипертония
41
:
1234
—
1239
5
Geelen
G
,
Greenleaf
JE
,
Keil
LC
, вызванные изменениями плазмы, вызываемыми парами у обезвоженных людей.
J Clin Endocrinol Metab
81
:
2131
—
2135
6
Nonogaki
K
2000
Новое понимание симпатической регуляции метаболизма глюкозы и жиров.
Diabetologia
43
:
533
—
549
7
Иордания
J
,
Резервуар
J
,
Stoffels
M
,
000
000 Franke G2
,
000 Franke G2
,
Luft
FC
,
Boschmann
M
2001
Взаимодействие между стимуляцией β-адренорецепторов и высвобождением оксида азота на перфузию тканей и метаболизм.
J Clin Endocrinol Metab
86
:
2803
–
2810
8
Boschmann
M
,
Krupp
G
,
Luft
FC
us
J
2002
Ответ in vivo на стимуляцию α (1) -адренорецептора в белой жировой ткани человека.
Obes Res
10
:
555
—
558
9
Ferrannini
E
1988
Теоретические основы косвенной калориметрии: обзор.
Метаболизм
37
:
287
—
301
10
Карига
P
,
Матиас
CJ
2001
Оральная
Гемодинамика за счет вегетативной динамики давления воды отказ.
Clin Sci (Лондон)
101
:
313
—
319
11
Schroeder
C
,
Bush
VE
,
Norcliffe
000
LJ2000
LJ2000 Tank
J
,
Jordan
J
,
Hainsworth
R
2002
Употребление воды резко улучшает ортостатическую переносимость у здоровых людей.
Обращение
106
:
2806
—
2811
12
Рутледж
HC
,
Чоудхари
S
,
Coote
JH
000
000
000 JH
nd
вагусная реакция на проглатывание воды у нормальных людей.
Clin Sci (Лондон)
103
:
157
—
162
13
Шеннон
JR
,
Gottesdiener
K
,
K0003
J
Лесть
S
,
Larson
PJ
,
Candelore
MR
,
Gertz
B
,
Robertson
D
,
Sun
Эффект
24-часовой энергетический баланс.
Clin Sci Colch
96
:
483
—
491
14
Hickner
RC
,
Ekelund
U
,
Mellander
000
S
t U
S
,
J
1995
Мышечный кровоток у кошек: сравнение методики микродиализа этанолом с прямым измерением.
J Appl Physiol
79
:
638
–
647
15
Hickner
RC
,
Rosdahl
H
,
Borg
Uorfd2
, Uном
L
,
Henriksson
J
1992
Метод этанола для мониторинга локальных изменений кровотока в скелетных мышцах крысы: значение для микродиализа.
Acta Physiol Scand
146
:
87
—
97
16
Simoneau
JA
,
Lortie
G
,
Boulay
MR
MR
, MC
,
G
,
Bouchard
C
1985
Гистохимические и биохимические характеристики скелетных мышц у мужчин и женщин, ведущих сидячий образ жизни.
Can J Physiol Pharmacol
63
:
30
—
35
17
Jordan
J
2002
Острое влияние воды на артериальное давление.Что мы знаем?
Clin Auton Res
12
:
250
—
255
18
Росси
P
,
Andriesse
GI
,
Oey
PL
,
000
000 GIeneke2
JM
,
Akkermans
LM
1998
Растяжение желудка увеличивает активность симпатических нервов эфферентных мышц и кровяное давление у здоровых людей.
J Neurol Sci
161
:
148
—
155
19
Ploutz-Snyder
L
,
Foley
J
,
Ploutz-Snyder 9000 R
,
Sagendorf
K
,
Meyer
R
1999
Опорожнение желудочного газа и жидкости по данным магнитно-резонансной томографии.
Eur J Appl Physiol Occup Physiol
79
:
212
—
220
20
Haberich
FJ
1968
Osmoreception в обращении портала.
Fed Proc
27
:
1137
—
1141
21
Adachi
A
1984
Термочувствительные и осморецептивные афферентные волокна в печеночной ветви блуждающего нерва.
J Auton Nerv Syst
10
:
269
—
273
22
Andrews
WH
,
Orbach
J
1974
Нервы, активирующие некоторые нервы кролика.
Am J Physiol
227
:
1273
—
1275
Авторские права © 2003 Эндокринологическое общество
Узнайте, почему метаболизм замедляется с возрастом
Чувствуете ли вы, что набираете пять фунтов после того, как съели кусок торта, когда вы могли бы съесть весь торт в свои 20 лет и не набрать ни грамма? Это не твое воображение. С возрастом наш метаболизм замедляется, и скорость расщепления пищи снижается на 10 процентов каждые десять лет после 20 лет.
Метаболизм — это количество энергии (калорий), которое ваше тело использует для поддержания себя. Количество калорий, сжигаемых вашим телом в состоянии покоя, зависит от вашей основной скорости метаболизма (BMR), а также от того, сколько вы тренируетесь, и от отношения мышечной массы к жировой ткани.
«К тому времени, когда нам исполнится 50, мы увидим 30-процентное снижение», — говорит Карен Андри, лицензированный и зарегистрированный диетолог и координатор по бариатрии в больнице Пьемонт, Атланта.
Почему так легко набрать вес
Замедление метаболизма может приводить к увеличению веса каждый год, даже если вы не едите намного больше, чем обычно.
Например, если вы каждую ночь съедаете по тарелке мороженого — около 500 калорий, — за эту неделю вы потребляете 3500 дополнительных калорий, что составляет фунт жира. Умножьте это количество на 52 недели в году, и вы увидите значительное увеличение веса, если будете употреблять всего одно дополнительное лакомство за ночь.
«Набрать вес очень просто, но очень сложно его сбросить», — говорит Андри.
Поддержание мышечной массы — ключ к предотвращению увеличения веса, но это может быть сложной задачей, потому что после 45 лет средний человек теряет примерно 1 процент мышечной массы каждый год.Щелкните здесь, чтобы найти укрепляющие упражнения для всего тела.
Поддержание здорового веса требует усилий, поэтому внимательно относитесь к тому, что вы едите каждый день, и увеличивайте физическую активность.
Идеальная масса тела
Как узнать, что у вас идеальный вес? Медицинский персонал обычно либо определяет ваш индекс массы тела, либо использует простую формулу, подобную этой:
- Женский — 5’0 дюймов = 100 фунтов
- Женский — 5 футов 1 дюйм = 105 фунтов
- Женский — 5 футов 2 дюйма = 110 фунтов
- Самка — 5 футов 3 дюйма = 115 фунтов.
- Самка — 5 футов 4 дюйма = 120 фунтов.
- Самка — 5 футов 5 дюймов = 125 фунтов.
«Мы используем таблицы Metropolitan Life Insurance для определения идеальной массы тела много лет назад», — говорит Андри. «Они на самом деле не очень реалистичны; однако они используются повсеместно, поэтому вы можете сравнить яблоки с яблоками ».
Поскольку вес настолько уникален для образа жизни и факторов риска каждого человека, лучше всего поговорить с лечащим врачом, чтобы определить ваш личный целевой вес.Щелкните здесь, чтобы найти ближайшего к вам врача.
Чтобы получить дополнительные советы по фитнесу и питанию, посетите страницу «Улучшение здоровья и благополучия».
Вам нужно записаться на прием к врачу из Пьемонта? Экономьте время, бронируйте онлайн.
14 лучших способов улучшить метаболизм
На буквальном уровне ваш уровень метаболизма описывает, сколько энергии вы тратите на выполнение повседневных физиологических функций. Однако это имеет много практических последствий, потому что ваш уровень метаболизма также влияет на то, как вы себя чувствуете, сколько калорий вы сжигаете, сколько калорий вы можете съесть, не набирая вес, ваше либидо, фертильность, переносимость холода, сколько у вас субъективной энергии. , как вы восстанавливаетесь после травм и стресса, как определенные продукты влияют на вас и как вы тренируетесь в тренажерном зале.Короче говоря, обычно лучше иметь более высокий уровень метаболизма.
Вот несколько способов улучшить метаболизм здоровым и продуктивным образом.
1. Оптимизируйте здоровье щитовидной железы.
Щитовидная железа является основным регулятором скорости метаболизма. Чтобы увеличить второе, мы должны поддержать первое.
- Определенные питательные вещества играют огромную роль в функции щитовидной железы, например, йод (для создания гормона щитовидной железы) и селен (для преобразования его в активную форму). Обязательно ешьте водоросли и бразильские орехи.
- Многие заболевания щитовидной железы являются аутоиммунными по своей природе, поэтому сосредоточьтесь на том, чтобы избегать распространенных аутоиммунных триггеров, таких как пшеница и другие злаки, укреплять кишечный барьер и поддерживать здоровье кишечника с помощью пребиотиков и пробиотиков.
- Чрезмерное потребление овощей семейства крестоцветных может иметь зобогенный (подавляющий щитовидную железу) эффект. Не бойтесь брокколи и капусты, но не ешьте несколько фунтов в день.
- Если вы низкоуглеводны, обратите внимание, что слишком низкое потребление калорий может подавить функцию щитовидной железы.Есть некоторые свидетельства того, что это может быть адаптивным и в какой-то мере благотворно влиять на долголетие, но вы не хотите подавлять его так сильно, что вы все время мерзнете, постоянно истощены и не можете похудеть. Если вы так себя чувствуете, употребляя мало углеводов, увеличьте количество калорий (будь то углеводы, белок или жир).
Если у вас действительно гипотиреоз, не бойтесь лечить его. Иногда добавление щитовидной железы — правильный ответ, и он менее «современный», чем вы думаете; в традиционных культурах используется для дополнения щитовидной железы животных.
2. Ешьте достаточно калорий.
Недостаточное потребление калорий посылает вашему организму сигнал о голоде. Конечно, есть разные степени — 400 калорий в день посылают совершенно другой сигнал, чем 1200 калорий в день, — но любой дефицит будет восприниматься как фактор стресса, каким бы незначительным он ни был. Это нормально. Стрессоры важны, и часть пользы мы получаем от них. Но в условиях постоянного и постоянного потребления низких калорий организм имеет тенденцию снижать скорость метаболизма, чтобы соответствовать количеству поступающей энергии.Это замедляет потерю веса, если не останавливает ее полностью, и объясняет, почему многие люди чувствуют себя усталыми, холодными и несчастными при длительной диете.
3. Следите за своим лептином.
Лептин является основным фактором, определяющим скорость метаболизма и расход энергии. Поскольку он секретируется жировыми отложениями и в меньшей степени реагирует на потребление углеводов, использование низкоуглеводной диеты для похудания может в конечном итоге, как это ни парадоксально, работать против вашего метаболизма. Вы можете исправить это, включив рефид углеводов один или два раза в неделю, при этом вы сохраните низкий уровень жиров и высокий уровень углеводов, чтобы повысить уровень лептина.Другие возможные решения включают отказ от зерен, лектины которых могут мешать рецепторам лептина.
4. Спринт.
Я всегда говорю, что спринт — единственное лучшее упражнение для высыхания. По сравнению с тренировками с отягощениями и традиционными кардиотренировками, интервалы между спринтами больше всего увеличивают расход энергии в состоянии покоя. Похоже, что большая часть увеличенных расходов происходит из-за окисления жиров. Лучше всего то, что повышенные расходы после спринта не приводят к увеличению количества потребляемой пищи.
Кроме того, спринт превращает ваши мышцы в гликогеновые губки, поэтому углеводы, которые вы едите для увеличения лептина и метаболизма, будут перемещаться в сторону мышц.
5. Поднимать тяжелые предметы.
Силовые тренировки необходимы для увеличения скорости метаболизма по трем основным причинам:
- Поднятие тяжестей наращивает мышцы. Мышцы стоят дорого. Он жаждет энергии. Ему нужна энергия. Он сжигает энергию. Он активирует метаболизм просто в силу своего существования.
- Непосредственный подъем тяжестей метаболически интенсивен. Подъем тяжелых предметов, будь то штанга, камень или собственный вес, требует энергии в данный момент.
- Силовые тренировки увеличивают скорость метаболизма в покое в краткосрочной и долгосрочной перспективе. Хорошая силовая тренировка даже после этого на несколько часов активизирует метаболизм.
6. Ешьте острую пищу.
Капсаицин, пряный компонент чили, обладает термогенными свойствами. Он активирует метаболически активный бурый жир, который может уменьшить жировые отложения. В сочетании с упражнениями увеличивает расход энергии. В сочетании с пищей капсаицин увеличивает термический эффект еды (количество энергии, сжигаемой во время пищеварения).Чуда здесь не жди. Но небольшие изменения складываются. Кроме того, острая еда — если она у вас есть — восхитительна.
7. Ешьте в обычное время.
Некоторым людям нравится беспорядочный график приема пищи. У некоторых людей непостоянный режим питания снижает метаболизм, потому что их организм «ожидает» еды в определенное время. В одном исследовании у здоровых и стройных взрослых людей наблюдался более низкий термический эффект от пищи — дополнительный прилив энергии, необходимый для обработки и переваривания пищи, которую мы едим, — когда они ели в соответствии с беспорядочным, неустойчивым графиком.Тот же эффект наблюдался у полных женщин, пытающихся похудеть. Снижение термического эффекта пищи означает снижение расхода энергии и метаболизма.
8. Погрузитесь в холод.
Чтобы согреться в холодную погоду, люди делают несколько вещей, связанных с повышением метаболизма. Мы дрожим, что сжигает калории для поддержания температуры тела. Мы активируем бурый жир, тип жировых отложений, который сжигает энергию и увеличивает метаболизм, а не накапливает и притупляет его. Чем более вы адаптированы к холоду, тем больше вы полагаетесь на бурый жир, чтобы согреться.
Необязательно принимать ледяные ванны. Просто оставив тепло в доме и выйдя на улицу с короткими рукавами в холодную погоду, вы увеличите свои энергозатраты.
9. Ешьте углеводы, которые вы зарабатываете.
Если вы «заработали углеводы» тяжелыми тренировками, решили не есть их и продолжаете выполнять тяжелую работу в тренажерном зале, ваш метаболизм пострадает. Кроссфиттер не делает себе никаких одолжений, не пополняя запасы гликогена после тяжелых тренировок; это усугубит дефицит энергии и вызовет реакцию голодания, угнетающую метаболизм.
10. Ешьте больше белка.
Протеин обладает наивысшим термическим эффектом из всех макроэлементов, что означает, что он требует больше всего калорий для переваривания и приводит к более высокому расходу энергии. То, что он помогает наращивать термогенную ткань — мышцы, — тоже не повредит.
11. Управляйте стрессом.
Острый стресс, по-видимому, увеличивает скорость метаболизма, вероятно, за счет увеличения адреналина и кортизола. Но в контексте хронического стресса, когда кортизол постоянно повышен и менее стимулирует, скорость метаболизма может снизиться.Это могло бы объяснить, почему женщины, которые сообщают о большем количестве «стрессовых событий», имеют более низкую термическую реакцию на еду, которую они едят. Это хронический стресс, и он наносит гораздо больший ущерб, чем острый стресс, от которого мы можем и действительно восстанавливаемся, и даже получаем пользу.
12. Стойте больше, чем сидите.
Прямые сравнения показывают, что люди, пользующиеся партами стоя, расходуют больше энергии, чем люди, которые сидят.
13. Часто двигайтесь в течение дня (беспокойство считается).
Термогенез физической активности (NEAT) — недооцененный путь к усилению метаболизма.Тот парень в вашем офисе, который раскачивается взад и вперед на своем стуле, постоянно выгибает шею, чтобы посмотреть на всех, постоянно жует жевательную резинку, часто ходит в ванную и, кажется, никогда не сидит на месте, возможно, страдает от привычки к кристаллическому метамфетамину. или он мог быть просто энергичным парнем с повышенным уровнем метаболизма.
14. Выпейте кофе или чай.
Неважно, страдаете ли вы ожирением или имеете нормальный вес, употребление кофе увеличивает скорость метаболизма. Повышение, опосредованное в первую очередь ускорением метаболизма жиров, носит временный характер, но прежде чем вы поймете, что снова наступило утро, вы готовы выпить еще одну чашку (или три).
Чай тоже работает. В нем есть кофеин (хотя и не так много, как в кофе), и, если вы пьете зеленый чай, особые соединения, которые способствуют расходу энергии независимо от кофеина.
Это 14 проверенных и верных способов улучшить метаболизм как в краткосрочной, так и в долгосрочной перспективе. Я уверен, что есть и другие. Что у вас, ребята?
Спасибо за внимание всем!
Предпочитаете слушать чтение? Получите аудиозапись этого сообщения в блоге и подпишитесь на подкаст Primal Blueprint в iTunes, чтобы получить мгновенный доступ ко всем прошлым, настоящим и будущим выпускам здесь.
об авторе
Марк Сиссон является основателем Mark’s Daily Apple, крестным отцом движения Primal food and lifestyle, а также автором бестселлера New York Times « The Keto Reset Diet ». Его последняя книга — Keto for Life , в которой он обсуждает, как он сочетает кето-диету с основным образом жизни для оптимального здоровья и долголетия. Марк также является автором множества других книг, в том числе The Primal Blueprint , которой приписывают ускоренный рост движения Primal / Paleo еще в 2009 году.Проведя три десятилетия в изучении и обучении людей тому, почему еда является ключевым компонентом достижения и поддержания оптимального самочувствия, Марк основал Primal Kitchen, компанию по производству настоящих продуктов питания, которая создает основные продукты для кухни, подходящие для Primal / палео, кето и Whole30.
Сообщение навигации
Если вы хотите добавить аватар ко всем своим комментариям, нажмите здесь!
.