МЕТАБОЛИЗМ | Энциклопедия Кругосвет
Содержание статьи
МЕТАБОЛИЗМ, или обмен веществ, химические превращения, протекающие от момента поступления питательных веществ в живой организм до момента, когда конечные продукты этих превращений выделяются во внешнюю среду. К метаболизму относятся все реакции, в результате которых строятся структурные элементы клеток и тканей, и процессы, в которых из содержащихся в клетках веществ извлекается энергия. Иногда для удобства рассматривают по отдельности две стороны метаболизма – анаболизм и катаболизм, т.е. процессы созидания органических веществ и процессы их разрушения. Анаболические процессы обычно связаны с затратой энергии и приводят к образованию сложных молекул из более простых, катаболические же сопровождаются высвобождением энергии и заканчиваются образованием таких конечных продуктов (отходов) метаболизма, как мочевина, диоксид углерода, аммиак и вода.
Термин «обмен веществ» вошел в повседневную жизнь с тех пор, как врачи стали связывать избыточный или недостаточный вес, чрезмерную нервозность или, наоборот, вялость больного с повышенным или пониженным обменом.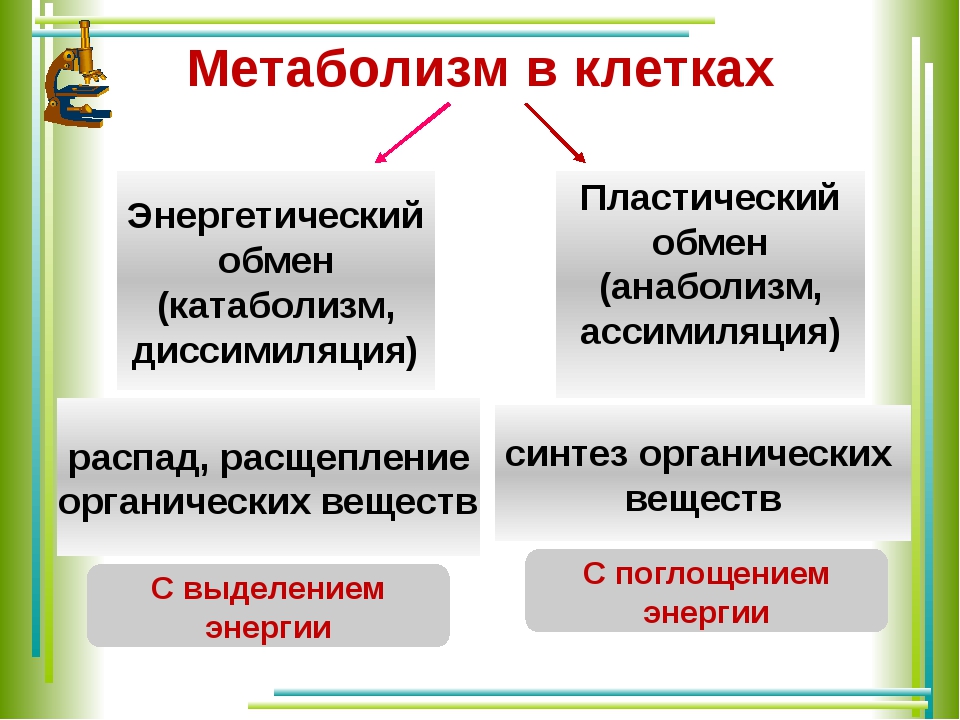 Для суждения об интенсивности метаболизма ставят тест на «основной обмен». Основной обмен – это показатель способности организма вырабатывать энергию. Тест проводят натощак в состоянии покоя; измеряют поглощение кислорода (О2) и выделение диоксида углерода (СО2). Сопоставляя эти величины, определяют, насколько полно организм использует («сжигает») питательные вещества. На интенсивность метаболизма влияют гормоны щитовидной железы, поэтому врачи при диагностике заболеваний, связанных с нарушениями обмена, в последнее время все чаще измеряют уровень этих гормонов в крови. См. также ЩИТОВИДНАЯ ЖЕЛЕЗА.
Для суждения об интенсивности метаболизма ставят тест на «основной обмен». Основной обмен – это показатель способности организма вырабатывать энергию. Тест проводят натощак в состоянии покоя; измеряют поглощение кислорода (О2) и выделение диоксида углерода (СО2). Сопоставляя эти величины, определяют, насколько полно организм использует («сжигает») питательные вещества. На интенсивность метаболизма влияют гормоны щитовидной железы, поэтому врачи при диагностике заболеваний, связанных с нарушениями обмена, в последнее время все чаще измеряют уровень этих гормонов в крови. См. также ЩИТОВИДНАЯ ЖЕЛЕЗА.
Методы исследования.
При изучении метаболизма какого-нибудь одного из питательных веществ прослеживают все его превращения от той формы, в какой оно поступает в организм, до конечных продуктов, выводимых из организма. В таких исследованиях применяется крайне разнообразный набор биохимических методов.
Использование интактных животных или органов.

Животному вводят изучаемое соединение, а затем в его моче и экскрементах определяют возможные продукты превращений (метаболиты) этого вещества. Более определенную информацию можно получить, исследуя метаболизм определенного органа, например печени или мозга. В этих случаях вещество вводят в соответствующий кровеносный сосуд, а метаболиты определяют в крови, оттекающей от данного органа.
Поскольку такого рода процедуры сопряжены с большими трудностями, часто для исследования используют тонкие срезы органов. Их инкубируют при комнатной температуре или при температуре тела в растворах с добавкой того вещества, метаболизм которого изучают. Клетки в таких препаратах не повреждены, и так как срезы очень тонкие, вещество легко проникает в клетки и легко выходит из них. Иногда затруднения возникают из-за слишком медленного прохождения вещества сквозь клеточные мембраны. В этих случаях ткани измельчают, чтобы разрушить мембраны, и с изучаемым веществом инкубируют клеточную кашицу. Именно в таких опытах было показано, что все живые клетки окисляют глюкозу до СО2 и воды и что только ткань печени способна синтезировать мочевину.
Именно в таких опытах было показано, что все живые клетки окисляют глюкозу до СО2 и воды и что только ткань печени способна синтезировать мочевину.
Использование клеток.
Даже клетки представляют собой очень сложно организованные системы. В них имеется ядро, а в окружающей его цитоплазме находятся более мелкие тельца, т.н. органеллы, различных размеров и консистенции. С помощью соответствующей методики ткань можно «гомогенизировать», а затем подвергнуть дифференциальному центрифугированию (разделению) и получить препараты, содержащие только митохондрии, только микросомы или прозрачную жидкость – цитоплазму. Эти препараты можно по отдельности инкубировать с тем соединением, метаболизм которого изучается, и таким путем установить, какие именно субклеточные структуры участвуют в его последовательных превращениях. Известны случаи, когда начальная реакция протекает в цитоплазме, ее продукт подвергается превращению в микросомах, а продукт этого превращения вступает в новую реакцию уже в митохондриях. Инкубация изучаемого вещества с живыми клетками или с гомогенатом ткани обычно не выявляет отдельные этапы его метаболизма, и только последовательные эксперименты, в которых для инкубации используются те или иные субклеточные структуры, позволяют понять всю цепочку событий.
Инкубация изучаемого вещества с живыми клетками или с гомогенатом ткани обычно не выявляет отдельные этапы его метаболизма, и только последовательные эксперименты, в которых для инкубации используются те или иные субклеточные структуры, позволяют понять всю цепочку событий.
Использование радиоактивных изотопов.
Для изучения метаболизма какого-либо вещества необходимы: 1) соответствующие аналитические методы для определения этого вещества и его метаболитов; и 2) методы, позволяющие отличать добавленное вещество от того же вещества, уже присутствующего в данном биологическом препарате. Эти требования служили главным препятствием при изучении метаболизма до тех пор, пока не были открыты радиоактивные изотопы элементов и в первую очередь радиоактивный углерод 14C. С появлением соединений, «меченных» 14C, а также приборов для измерения слабой радиоактивности эти трудности были преодолены. Если к биологическому препарату, например к суспензии митохондрий, добавляют меченную 14C жирную кислоту, то никаких специальных анализов для определения продуктов ее превращений не требуется; чтобы оценить скорость ее использования, достаточно просто измерять радиоактивность последовательно получаемых митохондриальных фракций. Эта же методика позволяет легко отличать молекулы радиоактивной жирной кислоты, введенной экспериментатором, от молекул жирной кислоты, уже присутствовавших в митохондриях к началу эксперимента.
Эта же методика позволяет легко отличать молекулы радиоактивной жирной кислоты, введенной экспериментатором, от молекул жирной кислоты, уже присутствовавших в митохондриях к началу эксперимента.
Хроматография и электрофорез.
В дополнение к вышеупомянутым требованиям биохимику необходимы и методы, позволяющие разделять смеси, состоящие из малых количеств органических веществ. Важнейший из них – хроматография, в основе которой лежит феномен адсорбции. Разделение компонентов смеси проводят при этом либо на бумаге, либо путем адсорбции на сорбенте, которым заполняют колонки (длинные стеклянные трубки), с последующей постепенной элюцией (вымыванием) каждого из компонентов.
Разделение методом электрофореза зависит от знака и числа зарядов ионизированных молекул. Электрофорез проводят на бумаге или на каком-нибудь инертном (неактивном) носителе, таком, как крахмал, целлюлоза или каучук.
Высокочувствительный и эффективный метод разделения – газовая хроматография. Им пользуются в тех случаях, когда подлежащие разделению вещества находятся в газообразном состоянии или могут быть в него переведены.
Выделение ферментов.
Последнее место в описываемом ряду – животное, орган, тканевой срез, гомогенат и фракция клеточных органелл – занимает фермент, способный катализировать определенную химическую реакцию. Выделение ферментов в очищенном виде – важный раздел в изучении метаболизма.
Сочетание перечисленных методов позволило проследить главные метаболические пути у большей части организмов (в том числе у человека), установить, где именно эти различные процессы протекают, и выяснить последовательные этапы главных метаболических путей. К настоящему времени известны тысячи отдельных биохимических реакций, изучены участвующие в них ферменты.
Клеточный метаболизм.
Живая клетка – это высокоорганизованная система. В ней имеются различные структуры, а также ферменты, способные их разрушить. Содержатся в ней и крупные макромолекулы, которые могут распадаться на более мелкие компоненты в результате гидролиза (расщепления под действием воды). В клетке обычно много калия и очень мало натрия, хотя клетка существует в среде, где натрия много, а калия относительно мало, и клеточная мембрана легко проницаема для обоих ионов. Следовательно, клетка – это химическая система, весьма далекая от равновесия. Равновесие наступает только в процессе посмертного автолиза (самопереваривания под действием собственных ферментов).
Следовательно, клетка – это химическая система, весьма далекая от равновесия. Равновесие наступает только в процессе посмертного автолиза (самопереваривания под действием собственных ферментов).
Потребность в энергии.
Чтобы удержать систему в состоянии, далеком от химического равновесия, требуется производить работу, а для этого необходима энергия. Получение этой энергии и выполнение этой работы – непременное условие для того, чтобы клетка оставалась в своем стационарном (нормальном) состоянии, далеком от равновесия. Одновременно в ней выполняется и иная работа, связанная со взаимодействием со средой, например: в мышечных клетках – сокращение; в нервных клетках – проведение нервного импульса; в клетках почек – образование мочи, значительно отличающейся по своему составу от плазмы крови; в специализированных клетках желудочно-кишечного тракта – синтез и выделение пищеварительных ферментов; в клетках эндокринных желез – секреция гормонов; в клетках светляков – свечение; в клетках некоторых рыб – генерирование электрических разрядов и т. д.
д.
Источники энергии.
В любом из перечисленных выше примеров непосредственным источником энергии, которую клетка использует для производства работы, служит энергия, заключенная в структуре аденозинтрифосфата (АТФ). В силу особенностей своей структуры это соединение богато энергией, и разрыв связей между его фосфатными группами может происходить таким образом, что высвобождающаяся энергия используется для производства работы. Однако энергия не может стать доступной для клетки при простом гидролитическом разрыве фосфатных связей АТФ: в этом случае она расходуется впустую, выделяясь в виде тепла. Процесс должен состоять из двух последовательных этапов, в каждом из которых участвует промежуточный продукт, обозначенный здесь X–Ф (в приведенных уравнениях X и Y означают два разных органических вещества; Ф – фосфат; АДФ – аденозиндифосфат):
Поскольку практически для любого проявления жизнедеятельности клеток необходим АТФ, неудивительно, что метаболическая активность живых клеток направлена в первую очередь на синтез АТФ. Этой цели служат различные сложные последовательности реакций, в которых используется потенциальная химическая энергия, заключенная в молекулах углеводов и жиров (липидов).
Этой цели служат различные сложные последовательности реакций, в которых используется потенциальная химическая энергия, заключенная в молекулах углеводов и жиров (липидов).
МЕТАБОЛИЗМ УГЛЕВОДОВ И ЛИПИДОВ
Синтез АТФ.
Анаэробный (без участия кислорода).
Главная роль углеводов и липидов в клеточном метаболизме состоит в том, что их расщепление на более простые соединения обеспечивает синтез АТФ. Несомненно, что те же процессы протекали и в первых, самых примитивных клетках. Однако в атмосфере, лишенной кислорода, полное окисление углеводов и жиров до CO2 было невозможно. У этих примитивных клеток имелись все же механизмы, с помощью которых перестройка структуры молекулы глюкозы обеспечивала синтез небольших количеств АТФ. Речь идет о процессах, которые у микроорганизмов называют брожением. Лучше всего изучено сбраживание глюкозы до этилового спирта и CO2 у дрожжей.
В ходе 11 последовательных реакций, необходимых для того, чтобы завершилось это превращение, образуется ряд промежуточных продуктов, представляющих собой эфиры фосфорной кислоты (фосфаты). Их фосфатная группа переносится на аденозиндифосфат (АДФ) с образованием АТФ. Чистый выход АТФ составляет 2 молекулы АТФ на каждую молекулу глюкозы, расщепленную в процессе брожения. Аналогичные процессы происходят во всех живых клетках; поскольку они поставляют необходимую для жизнедеятельности энергию, их иногда (не вполне корректно) называют анаэробным дыханием клеток.
Их фосфатная группа переносится на аденозиндифосфат (АДФ) с образованием АТФ. Чистый выход АТФ составляет 2 молекулы АТФ на каждую молекулу глюкозы, расщепленную в процессе брожения. Аналогичные процессы происходят во всех живых клетках; поскольку они поставляют необходимую для жизнедеятельности энергию, их иногда (не вполне корректно) называют анаэробным дыханием клеток.
У млекопитающих, в том числе у человека, такой процесс называется гликолизом и его конечным продуктом является молочная кислота, а не спирт и CO2. Вся последовательность реакций гликолиза, за исключением двух последних этапов, полностью идентична процессу, протекающему в дрожжевых клетках.
Аэробный (с использованием кислорода).
С появлением в атмосфере кислорода, источником которого послужил, очевидно, фотосинтез растений, в ходе эволюции развился механизм, обеспечивающий полное окисление глюкозы до CO2 и воды, – аэробный процесс, в котором чистый выход АТФ составляет 38 молекул АТФ на каждую окисленную молекулу глюкозы. Этот процесс потребления клетками кислорода для образования богатых энергией соединений известен как клеточное дыхание (аэробное). В отличие от анаэробного процесса, осуществляемого ферментами цитоплазмы, окислительные процессы протекают в митохондриях. В митохондриях пировиноградная кислота – промежуточный продукт, образовавшийся в анаэробной фазе – окисляется до СО2 в шести последовательных реакциях, в каждой из которых пара электронов переносится на общий акцептор – кофермент никотинамидадениндинуклеотид (НАД). Эту последовательность реакций называют циклом трикарбоновых кислот, циклом лимонной кислоты или циклом Кребса. Из каждой молекулы глюкозы образуется 2 молекулы пировиноградной кислоты; 12 пар электронов отщепляется от молекулы глюкозы в ходе ее окисления, описываемого уравнением:
Этот процесс потребления клетками кислорода для образования богатых энергией соединений известен как клеточное дыхание (аэробное). В отличие от анаэробного процесса, осуществляемого ферментами цитоплазмы, окислительные процессы протекают в митохондриях. В митохондриях пировиноградная кислота – промежуточный продукт, образовавшийся в анаэробной фазе – окисляется до СО2 в шести последовательных реакциях, в каждой из которых пара электронов переносится на общий акцептор – кофермент никотинамидадениндинуклеотид (НАД). Эту последовательность реакций называют циклом трикарбоновых кислот, циклом лимонной кислоты или циклом Кребса. Из каждой молекулы глюкозы образуется 2 молекулы пировиноградной кислоты; 12 пар электронов отщепляется от молекулы глюкозы в ходе ее окисления, описываемого уравнением:
Перенос электронов.
В каждой митохондрии имеется механизм, посредством которого восстановленный НАД (НАДЧН, где Н – водород), образовавшийся в цикле трикарбоновых кислот, передает свою пару электронов кислороду. Перенос, однако, не происходит напрямую. Электроны как бы передаются «из рук в руки» и, лишь пройдя цепь переносчиков, присоединяются к кислороду. Эта «цепь переноса электронов» состоит из следующих компонентов:
Перенос, однако, не происходит напрямую. Электроны как бы передаются «из рук в руки» и, лишь пройдя цепь переносчиков, присоединяются к кислороду. Эта «цепь переноса электронов» состоит из следующих компонентов:
НАДНЧН ® Флавинадениндинклеотид ® Кофермент Q ®
® Цитохром b ® Цитохром c ® Цитохром a ® O2
Все компоненты этой системы, находящиеся в митохондриях, фиксированы в пространстве и сцеплены друг с другом. Такое их состояние облегчает перенос электронов.
В состав НАД входит никотиновая кислота (витамин ниацин), а в состав флавинадениндинуклеотида – рибофлавин (витамин B2). Кофермент Q представляет собой высокомолекулярный хинон, синтезируемый в печени, а цитохромы – это три разных белка, каждый из которых, подобно гемоглобину, содержит гемогруппу.
В цепи переноса электронов на каждую пару электронов, перенесенную от НАДЧН на O2, синтезируется 3 молекулы АТФ. Поскольку от каждой молекулы глюкозы отщепляются и передаются молекулам НАД 12 пар электронов, в общей сложности на каждую молекулу глюкозы образуется 3ґ12 = 36 молекул АТФ. Этот процесс образования АТФ в ходе окисления называется окислительным фосфорилированием.
Этот процесс образования АТФ в ходе окисления называется окислительным фосфорилированием.
Липиды как источник энергии.
Жирные кислоты могут использоваться в качестве источника энергии приблизительно так же, как и углеводы. Окисление жирных кислот протекает путем последовательного отщепления от молекулы жирной кислоты двууглеродного фрагмента с образованием ацетилкофермента A (ацетил-КоА) и одновременной передачей двух пар электронов в цепь переноса электронов. Образовавшийся ацетил-КоА – нормальный компонент цикла трикарбоновых кислот, и в дальнейшем его судьба не отличается от судьбы ацетил-КоА, поставляемого углеводным обменом. Таким образом, механизмы синтеза АТФ при окислении как жирных кислот, так и метаболитов глюкозы практически одинаковы.
Если организм животного получает энергию почти целиком за счет одного только окисления жирных кислот, а это бывает, например, при голодании или при сахарном диабете, то скорость образования ацетил-КоА превышает скорость его окисления в цикле трикарбоновых кислот. В этом случае лишние молекулы ацетил-КоА реагируют друг с другом, в результате чего образуются в конечном счете ацетоуксусная и b-гидроксимасляная кислоты. Их накопление является причиной патологического состояния, т.н. кетоза (одного из видов ацидоза), который при тяжелом диабете может вызвать кому и смерть.
В этом случае лишние молекулы ацетил-КоА реагируют друг с другом, в результате чего образуются в конечном счете ацетоуксусная и b-гидроксимасляная кислоты. Их накопление является причиной патологического состояния, т.н. кетоза (одного из видов ацидоза), который при тяжелом диабете может вызвать кому и смерть.
Запасание энергии.
Животные питаются нерегулярно, и их организму нужно как-то запасать заключенную в пище энергию, источником которой являются поглощенные животным углеводы и жиры. Жирные кислоты могут запасаться в виде нейтральных жиров либо в печени, либо в жировой ткани. Углеводы, поступая в большом количестве, в желудочно-кишечном тракте гидролизуются до глюкозы или иных сахаров, которые затем в печени превращаются в ту же глюкозу. Здесь из глюкозы синтезируется гигантский полимер гликоген путем присоединения друг к другу остатков глюкозы с отщеплением молекул воды (число остатков глюкозы в молекулах гликогена доходит до 30 000). Когда возникает потребность в энергии, гликоген вновь распадается до глюкозы в реакции, продуктом которой является глюкозофосфат.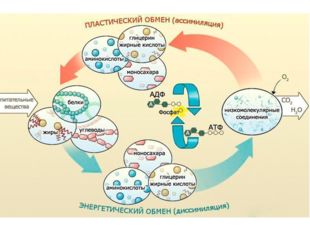 Этот глюкозофосфат направляется на путь гликолиза – процесса, составляющего часть пути окисления глюкозы. В печени глюкозофосфат может также подвергнуться гидролизу, и образующаяся глюкоза поступает в кровоток и доставляется кровью к клеткам в разных частях тела.
Этот глюкозофосфат направляется на путь гликолиза – процесса, составляющего часть пути окисления глюкозы. В печени глюкозофосфат может также подвергнуться гидролизу, и образующаяся глюкоза поступает в кровоток и доставляется кровью к клеткам в разных частях тела.
Синтез липидов из углеводов.
Если количество углеводов, поглощенных с пищей за один прием, больше того, какое может быть запасено в виде гликогена, то избыток углеводов превращается в жиры. Начальная последовательность реакций совпадает при этом с обычным окислительным путем, т.е. сначала из глюкозы образуется ацетил-КоА, но далее этот ацетил-КоА используется в цитоплазме клетки для синтеза длинноцепочечных жирных кислот. Процесс синтеза можно описать как обращение обычного процесса окисления жирных клеток. Затем жирные кислоты запасаются в виде нейтральных жиров (триглицеридов), отлагающихся в разных частях тела. Когда требуется энергия, нейтральные жиры подвергаются гидролизу и жирные кислоты поступают в кровь. Здесь они адсорбируются молекулами плазменных белков (альбуминов и глобулинов) и затем поглощаются клетками самых разных типов. Механизмов, способных осуществлять синтез глюкозы из жирных кислот, у животных нет, но у растений такие механизмы имеются.
Здесь они адсорбируются молекулами плазменных белков (альбуминов и глобулинов) и затем поглощаются клетками самых разных типов. Механизмов, способных осуществлять синтез глюкозы из жирных кислот, у животных нет, но у растений такие механизмы имеются.
Метаболизм липидов.
Липиды попадают в организм главным образом в форме триглицеридов жирных кислот. В кишечнике под действием ферментов поджелудочной железы они подвергаются гидролизу, продукты которого всасываются клетками стенки кишечника. Здесь из них вновь синтезируются нейтральные жиры, которые через лимфатическую систему поступают в кровь и либо транспортируются в печень, либо отлагаются в жировой ткани. Выше уже указывалось, что жирные кислоты могут также синтезироваться заново из углеводных предшественников. Следует отметить, что, хотя в клетках млекопитающих может происходить включение одной двойной связи в молекулы длинноцепочечных жирных кислот (между С–9 и С–10), включать вторую и третью двойную связь эти клетки неспособны.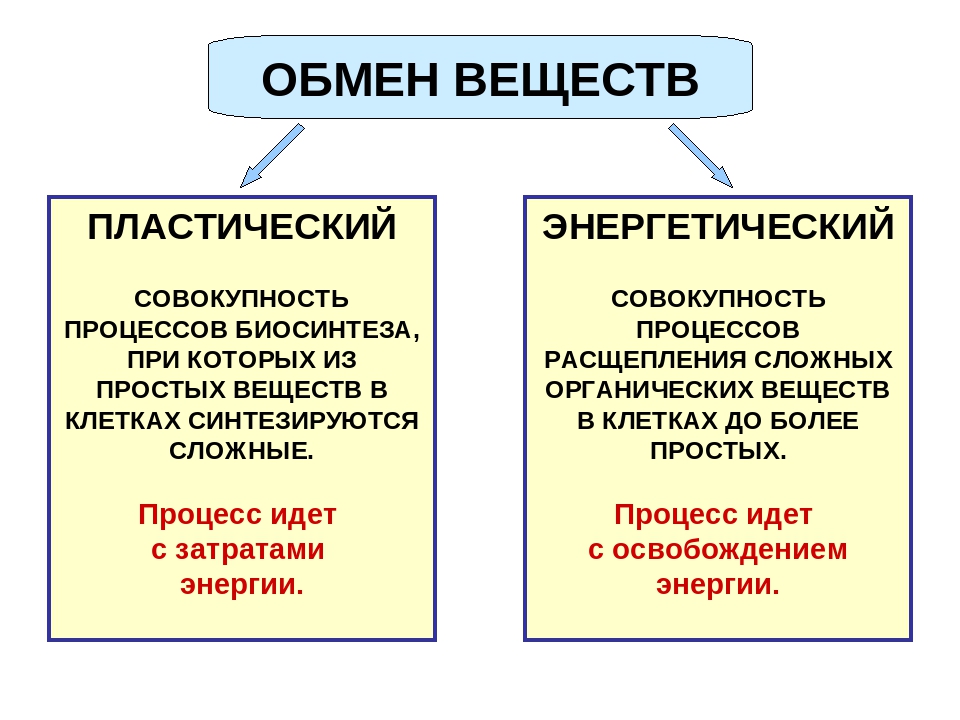 Поскольку жирные кислоты с двумя и тремя двойными связями играют важную роль в метаболизме млекопитающих, они в сущности являются витаминами. Поэтому линолевую (C18:2) и линоленовую (C18:3) кислоты называют незаменимыми жирными кислотами. В то же время в клетках млекопитающих в линоленовую кислоту может включаться четвертая двойная связь и путем удлинения углеродной цепи может образоваться арахидоновая кислота (C20:4), также необходимый участник метаболических процессов.
Поскольку жирные кислоты с двумя и тремя двойными связями играют важную роль в метаболизме млекопитающих, они в сущности являются витаминами. Поэтому линолевую (C18:2) и линоленовую (C18:3) кислоты называют незаменимыми жирными кислотами. В то же время в клетках млекопитающих в линоленовую кислоту может включаться четвертая двойная связь и путем удлинения углеродной цепи может образоваться арахидоновая кислота (C20:4), также необходимый участник метаболических процессов.
В процессе синтеза липидов остатки жирных кислот, связанные с коферментом А (ацил-КоА), переносятся на глицерофосфат – эфир фосфорной кислоты и глицерина. В результате образуется фосфатидная кислота – соединение, в котором одна гидроксильная группа глицерина этерифицирована фосфорной кислотой, а две группы – жирными кислотами. При образовании нейтральных жиров фосфорная кислота удаляется путем гидролиза, и ее место занимает третья жирная кислота в результате реакции с ацил-КоА. Кофермент А образуется из пантотеновой кислоты (одного из витаминов). В его молекуле имеется сульфгидрильная (– SH) группа, способная реагировать с кислотами с образованием тиоэфиров. При образовании фосфолипидов фосфатидная кислота реагирует непосредственно с активированным производным одного из азотистых оснований, таких, как холин, этаноламин или серин.
В его молекуле имеется сульфгидрильная (– SH) группа, способная реагировать с кислотами с образованием тиоэфиров. При образовании фосфолипидов фосфатидная кислота реагирует непосредственно с активированным производным одного из азотистых оснований, таких, как холин, этаноламин или серин.
За исключением витамина D, все встречающиеся в организме животных стероиды (производные сложных спиртов) легко синтезируются самим организмом. Сюда относятся холестерин (холестерол), желчные кислоты, мужские и женские половые гормоны и гормоны надпочечников. В каждом случае исходным материалом для синтеза служит ацетил-КоА: из ацетильных групп путем многократно повторяющейся конденсации строится углеродный скелет синтезируемого соединения.
МЕТАБОЛИЗМ БЕЛКОВ
Синтез аминокислот.
Растения и большинство микроорганизмов могут жить и расти в среде, в которой для их питания имеются только минеральные вещества, диоксид углерода и вода. Это значит, что все обнаруживаемые в них органические вещества эти организмы синтезируют сами. Встречающиеся во всех живых клетках белки построены из 21 вида аминокислот, соединенных в различной последовательности. Аминокислоты синтезируются живыми организмами. В каждом случае ряд химических реакций приводит к образованию a-кетокислоты. Одна такая a-кетокислота, а именно a-кетоглутаровая (обычный компонент цикла трикарбоновых кислот), участвует в связывании азота по следующему уравнению:
Встречающиеся во всех живых клетках белки построены из 21 вида аминокислот, соединенных в различной последовательности. Аминокислоты синтезируются живыми организмами. В каждом случае ряд химических реакций приводит к образованию a-кетокислоты. Одна такая a-кетокислота, а именно a-кетоглутаровая (обычный компонент цикла трикарбоновых кислот), участвует в связывании азота по следующему уравнению:
a-Кетоглутаровая кислота + NH3 + НАДЧН ®
® Глутаминовая кислота + НАД.
Азот глутаминовой кислоты может быть затем передан любой из других a-кетокислот с образованием соответствующей аминокислоты.
Организм человека и большинства других животных сохранил способность синтезировать все аминокислоты за исключением девяти т.н. незаменимых аминокислот. Поскольку кетокислоты, соответствующие этим девяти, не синтезируются, незаменимые аминокислоты должны поступать с пищей.
Синтез белков.
Аминокислоты нужны для биосинтеза белка. Процесс биосинтеза протекает обычно следующим образом. В цитоплазме клетки каждая аминокислота «активируется» в реакции с АТФ, а затем присоединяется к концевой группе молекулы рибонуклеиновой кислоты, специфичной именно для данной аминокислоты. Эта сложная молекула связывается с небольшим тельцем, т.н. рибосомой, в положении, определяемом более длинной молекулой рибонуклеиновой кислоты, прикрепленной к рибосоме. После того как все эти сложные молекулы соответствующим образом выстроились, связи между исходной аминокислотой и рибонуклеиновой кислотой разрываются и возникают связи между соседними аминокислотами – синтезируется специфичный белок. Процесс биосинтеза поставляет белки не только для роста организма или для секреции в среду. Все белки живых клеток со временем претерпевают распад до составляющих их аминокислот, и для поддержания жизни клетки должны синтезироваться вновь.
Процесс биосинтеза протекает обычно следующим образом. В цитоплазме клетки каждая аминокислота «активируется» в реакции с АТФ, а затем присоединяется к концевой группе молекулы рибонуклеиновой кислоты, специфичной именно для данной аминокислоты. Эта сложная молекула связывается с небольшим тельцем, т.н. рибосомой, в положении, определяемом более длинной молекулой рибонуклеиновой кислоты, прикрепленной к рибосоме. После того как все эти сложные молекулы соответствующим образом выстроились, связи между исходной аминокислотой и рибонуклеиновой кислотой разрываются и возникают связи между соседними аминокислотами – синтезируется специфичный белок. Процесс биосинтеза поставляет белки не только для роста организма или для секреции в среду. Все белки живых клеток со временем претерпевают распад до составляющих их аминокислот, и для поддержания жизни клетки должны синтезироваться вновь.
Синтез других азотсодержащих соединений.
В организме млекопитающих аминокислоты используются не только для биосинтеза белков, но и как исходный материал для синтеза многих азотсодержащих соединений. Аминокислота тирозин является предшественником гормонов адреналина и норадреналина. Простейшая аминокислота глицин служит исходным материалом для биосинтеза пуринов, входящих в состав нуклеиновых кислот, и порфиринов, входящих в состав цитохромов и гемоглобина. Аспарагиновая кислота – предшественник пиримидинов нуклеиновых кислот. Метильная группа метионина передается ряду других соединений в ходе биосинтеза креатина, холина и саркозина. При биосинтезе креатина от одного соединения к другому передается также и гуанидиновая группировка аргинина. Триптофан служит предшественником никотиновой кислоты, а из валина в растениях синтезируется такой витамин, как пантотеновая кислота. Все это лишь отдельные примеры использования аминокислот в процессах биосинтеза.
Аминокислота тирозин является предшественником гормонов адреналина и норадреналина. Простейшая аминокислота глицин служит исходным материалом для биосинтеза пуринов, входящих в состав нуклеиновых кислот, и порфиринов, входящих в состав цитохромов и гемоглобина. Аспарагиновая кислота – предшественник пиримидинов нуклеиновых кислот. Метильная группа метионина передается ряду других соединений в ходе биосинтеза креатина, холина и саркозина. При биосинтезе креатина от одного соединения к другому передается также и гуанидиновая группировка аргинина. Триптофан служит предшественником никотиновой кислоты, а из валина в растениях синтезируется такой витамин, как пантотеновая кислота. Все это лишь отдельные примеры использования аминокислот в процессах биосинтеза.
Азот, поглощаемый микроорганизмами и высшими растениями в виде иона аммония, расходуется почти целиком на образование аминокислот, из которых затем синтезируются многие азотсодержащие соединения живых клеток. Избыточных количеств азота ни растения, ни микроорганизмы не поглощают.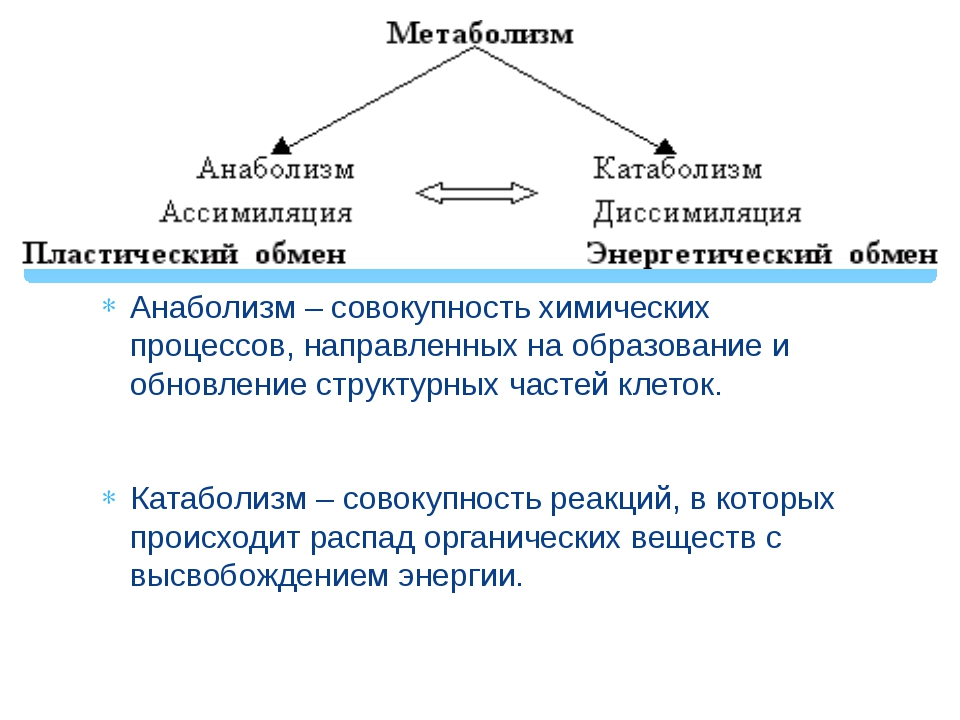 В отличие от них, у животных количество поглощенного азота зависит от содержащихся в пище белков. Весь азот, поступивший в организм в виде аминокислот и не израсходованный в процессах биосинтеза, довольно быстро выводится из организма с мочой. Происходит это следующим образом. В печени неиспользованные аминокислоты передают свой азот a-кетоглутаровой кислоте с образованием глутаминовой кислоты, которая дезаминируется, высвобождая аммиак. Далее азот аммиака может либо на время запасаться путем синтеза глутамина, либо сразу же использоваться для синтеза мочевины, протекающего в печени.
В отличие от них, у животных количество поглощенного азота зависит от содержащихся в пище белков. Весь азот, поступивший в организм в виде аминокислот и не израсходованный в процессах биосинтеза, довольно быстро выводится из организма с мочой. Происходит это следующим образом. В печени неиспользованные аминокислоты передают свой азот a-кетоглутаровой кислоте с образованием глутаминовой кислоты, которая дезаминируется, высвобождая аммиак. Далее азот аммиака может либо на время запасаться путем синтеза глутамина, либо сразу же использоваться для синтеза мочевины, протекающего в печени.
У глутамина есть и другая роль. Он может подвергаться гидролизу в почках с высвобождением аммиака, который поступает в мочу в обмен на ионы натрия. Этот процесс крайне важен как средство поддержания кислотно-щелочного равновесия в организме животного. Почти весь аммиак, происходящий из аминокислот и, возможно, из других источников, превращается в печени в мочевину, так что свободного аммиака в крови обычно почти нет. Однако при некоторых условиях довольно значительные количества аммиака содержит моча. Этот аммиак образуется в почках из глутамина и переходит в мочу в обмен на ионы натрия, которые таким образом реадсорбируются и задерживаются в организме. Этот процесс усиливается при развитии ацидоза – состояния, при котором организм нуждается в дополнительных количествах катионов натрия для связывания избытка ионов бикарбоната в крови.
Однако при некоторых условиях довольно значительные количества аммиака содержит моча. Этот аммиак образуется в почках из глутамина и переходит в мочу в обмен на ионы натрия, которые таким образом реадсорбируются и задерживаются в организме. Этот процесс усиливается при развитии ацидоза – состояния, при котором организм нуждается в дополнительных количествах катионов натрия для связывания избытка ионов бикарбоната в крови.
Избыточные количества пиримидинов тоже распадаются в печени через ряд реакций, в которых высвобождается аммиак. Что касается пуринов, то их избыток подвергается окислению с образованием мочевой кислоты, выделяющейся с мочой у человека и других приматов, но не у остальных млекопитающих. У птиц отсутствует механизм синтеза мочевины, и именно мочевая кислота, а не мочевина, является у них конечным продуктом обмена всех азотсодержащих соединений.
Нуклеиновые кислоты.
Структура и синтез этих азотсодержащих соединений подробно описаны в статье НУКЛЕИНОВЫЕ КИСЛОТЫ.
ОБЩИЕ ПРЕДСТАВЛЕНИЯ О МЕТАБОЛИЗМЕОРГАНИЧЕСКИХ ВЕЩЕСТВ
Можно сформулировать некоторые общие понятия, или «правила», касающиеся метаболизма. Приведенные ниже несколько главных «правил» позволяют лучше понять, как протекает и регулируется метаболизм.
1. Метаболические пути необратимы. Распад никогда не идет по пути, который являлся бы простым обращением реакций синтеза. В нем участвуют другие ферменты и другие промежуточные продукты. Нередко противоположно направленные процессы протекают в разных отсеках клетки. Так, жирные кислоты синтезируются в цитоплазме при участии одного набора ферментов, а окисляются в митохондриях при участии совсем другого набора.
2. Ферментов в живых клетках достаточно для того, чтобы все известные метаболические реакции могли протекать гораздо быстрее, чем это обычно наблюдается в организме. Следовательно, в клетках существуют какие-то регуляторные механизмы. Открыты разные типы таких механизмов.
а) Фактором, ограничивающим скорость метаболических превращений данного вещества, может быть поступление этого вещества в клетку; именно на этот процесс в таком случае и направлена регуляция.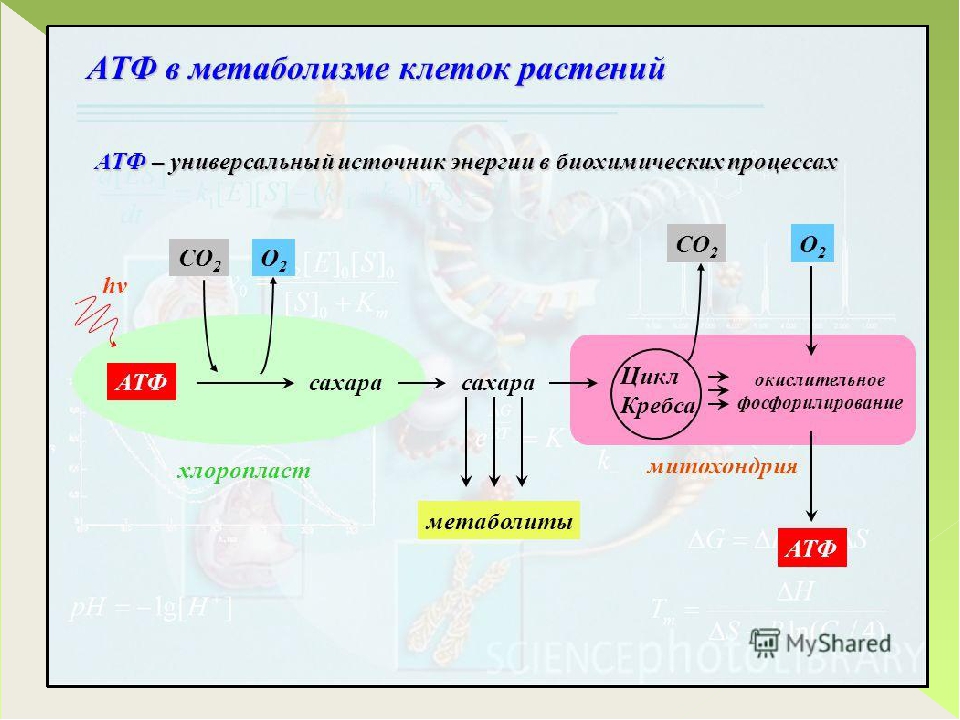 Роль инсулина, например, связана с тем, что он, по-видимому, облегчает проникновение глюкозы во все клетки, глюкоза же подвергается превращениям с той скоростью, с какой она поступает. Сходным образом проникновение железа и кальция из кишечника в кровь зависит от процессов, скорость которых регулируется.
Роль инсулина, например, связана с тем, что он, по-видимому, облегчает проникновение глюкозы во все клетки, глюкоза же подвергается превращениям с той скоростью, с какой она поступает. Сходным образом проникновение железа и кальция из кишечника в кровь зависит от процессов, скорость которых регулируется.
б) Вещества далеко не всегда могут свободно переходить из одного клеточного отсека в другой; есть данные, что внутриклеточный перенос регулируется некоторыми стероидными гормонами.
в) Выявлено два типа сервомеханизмов «отрицательной обратной связи».
У бактерий были обнаружены примеры того, что присутствие продукта какой-нибудь последовательности реакций, например аминокислоты, подавляет биосинтез одного из ферментов, необходимых для образования этой аминокислоты.
В каждом случае фермент, биосинтез которого оказывается затронутым, был ответствен за первый «определяющий» этап (на схеме реакция 4) метаболического пути, ведущего к синтезу данной аминокислоты.
Второй механизм хорошо изучен у млекопитающих. Это простое ингибирование конечным продуктом (в нашем случае – аминокислотой) фермента, ответственного за первый «определяющий» этап метаболического пути.
Еще один тип регулирования посредством обратной связи действует в тех случаях, когда окисление промежуточных продуктов цикла трикарбоновых кислот сопряжено с образованием АТФ из АДФ и фосфата в процессе окислительного фосфорилирования. Если весь имеющийся в клетке запас фосфата и (или) АДФ уже исчерпан, то окисление приостанавливается и может возобновиться лишь после того, как этот запас вновь станет достаточным. Таким образом, окисление, смысл которого в том, чтобы поставлять полезную энергию в форме АТФ, происходит только тогда, когда возможен синтез АТФ.
3. В биосинтетических процессах участвует сравнительно небольшое число строительных блоков, каждый из которых используется для синтеза многих соединений. Среди них можно назвать ацетилкофермент А, глицерофосфат, глицин, карбамилфосфат, поставляющий карбамильную (H2N–CO–) группу, производные фолиевой кислоты, служащие источником гидроксиметильной и формильной групп, S-аденозилметионин – источник метильных групп, глутаминовую и аспарагиновую кислоты, поставляющие аминогруппы, и наконец, глутамин – источник амидных групп..PNG) Из этого относительно небольшого числа компонентов строятся все те разнообразные соединения, которые мы находим в живых организмах.
Из этого относительно небольшого числа компонентов строятся все те разнообразные соединения, которые мы находим в живых организмах.
4. Простые органические соединения редко участвуют в метаболических реакциях непосредственно. Обычно они должны быть сначала «активированы» путем присоединения к одному из ряда соединений, универсально используемых в метаболизме. Глюкоза, например, может подвергнуться окислению лишь после того, как она будет этерифицирована фосфорной кислотой, для прочих же своих превращений она должна быть этерифицирована уридиндифосфатом. Жирные кислоты не могут быть вовлечены в метаболические превращения прежде, чем они образуют эфиры с коферментом А. Каждый из этих активаторов либо родствен одному из нуклеотидов, входящих в состав рибонуклеиновой кислоты, либо образуется из какого-нибудь витамина. Легко понять в связи с этим, почему витамины требуются в таких небольших количествах. Они расходуются на образование «коферментов», а каждая молекула кофермента на протяжении жизни организма используется многократно, в отличие от основных питательных веществ (например, глюкозы), каждая молекула которых используется только один раз.
В заключение следует сказать, что термин «метаболизм», означавший ранее нечто не более сложное, чем просто использование углеводов и жиров в организме, теперь применяется для обозначения тысяч ферментативных реакций, вся совокупность которых может быть представлена как огромная сеть метаболических путей, многократно пересекающихся (из-за наличия общих промежуточных продуктов) и управляемых очень тонкими регуляторными механизмами.
МЕТАБОЛИЗМ МИНЕРАЛЬНЫХ ВЕЩЕСТВ
Относительное содержание.
Различные элементы, встречающиеся в живых организмах, перечислены ниже в убывающем порядке в зависимости от их относительного содержания: 1) кислород, углерод, водород и азот; 2) кальций, фосфор, калий и сера; 3) натрий, хлор, магний и железо; 4) марганец, медь, молибден, селен, йод и цинк; 5) алюминий, фтор, кремний и литий; 6) бром, мышьяк, свинец и, возможно, некоторые другие.
Кислород, углерод, водород и азот – это те элементы, из которых построены мягкие ткани тела. Они входят в состав таких соединений, как углеводы, липиды, белки, вода, диоксид углерода и аммиак. Элементы, перечисленные в пп. 2 и 3, находятся в организме обычно в виде одного или нескольких неорганических соединений, а элементы пп. 4, 5 и 6 присутствуют только в следовых количествах и потому их называют микроэлементами.
Они входят в состав таких соединений, как углеводы, липиды, белки, вода, диоксид углерода и аммиак. Элементы, перечисленные в пп. 2 и 3, находятся в организме обычно в виде одного или нескольких неорганических соединений, а элементы пп. 4, 5 и 6 присутствуют только в следовых количествах и потому их называют микроэлементами.
Распределение в организме.
Кальций.
Кальций присутствует главным образом в костной ткани и в зубах, преимущественно в виде фосфата и в небольших количествах в виде карбоната и фторида. Поступающий с пищей кальций всасывается в основном в верхних отделах кишечника, имеющих слабокислую реакцию. Этому всасыванию (у человека здесь всасывается всего 20–30% кальция пищи) способствует витамин D. Под действием витамина D клетки кишечника вырабатывают особый белок, который связывает кальций и облегчает его перенос через стенку кишечника в кровь. На всасывание влияет также присутствие некоторых других веществ, в особенности фосфата и оксалата, которые в малых количествах способствуют всасыванию, а в больших, наоборот, подавляют его.
В крови около половины кальция связано с белком, остальное составляют ионы кальция. Соотношение ионизированной и неионизированной форм зависит от общей концентрации кальция в крови, а также от содержания белка и фосфата и концентрации водородных ионов (рН крови). Доля неионизированного кальция, на которую влияет уровень белка, позволяет косвенным образом судить о качестве питания и об эффективности работы печени, в которой идет синтез плазменных белков.
На количество ионизированного кальция влияют, с одной стороны, витамин D и факторы, воздействующие на всасывание, а с другой – паратиреоидный гормон и, возможно, также витамин D, поскольку оба эти вещества регулируют как скорость отложения кальция в костной ткани, так и его мобилизацию, т.е. вымывание из костей. Избыток паратиреоидного гормона стимулирует выход кальция из костной ткани, что приводит к повышению его концентрации в плазме. Изменяя скорости всасывания и экскреции кальция и фосфата, а также скорости образования костной ткани и ее разрушения, эти механизмы строго контролируют концентрацию кальция и фосфата в сыворотке крови. Ионы кальция играют регулирующую роль во многих физиологических процессах, в том числе в нервных реакциях, мышечном сокращении, свертывании крови. Выведение кальция из организма происходит в норме в основном (на 2/3) через желчь и кишечник и в меньшей степени (1/3) – через почки.
Ионы кальция играют регулирующую роль во многих физиологических процессах, в том числе в нервных реакциях, мышечном сокращении, свертывании крови. Выведение кальция из организма происходит в норме в основном (на 2/3) через желчь и кишечник и в меньшей степени (1/3) – через почки.
Фосфор.
Метаболизм фосфора – одного из главных компонентов костной ткани и зубов – во многом зависит от тех же факторов, что и метаболизм кальция. Фосфор в виде фосфата присутствует в организме также в сотнях различных физиологически важных органических эфиров. Паратиреоидный гормон стимулирует выведение фосфора с мочой и выход его из костной ткани; тем самым он регулирует концентрацию фосфора в плазме крови.
Натрий.
Натрий – главный катион внеклеточной жидкости – вместе с белком, хлоридом и бикарбонатом играет важнейшую роль в регулировании осмотического давления и pH (концентрации водородных ионов) крови. В клетках, напротив, содержится очень мало натрия, так как они обладают механизмом для выведения ионов натрия и удержания ионов калия. Весь натрий, превышающий потребности организма, очень быстро выводится через почки.
Весь натрий, превышающий потребности организма, очень быстро выводится через почки.
Поскольку во всех процессах выделения натрий теряется, он должен постоянно поступать в организм с пищей. При ацидозе, когда необходимо, чтобы из организма выводились большие количества анионов (например, хлорида или ацетоацетата), почки предотвращают чрезмерную потерю натрия благодаря образованию аммиака из глутамина. Выведение натрия через почки регулируется гормоном коры надпочечников альдостероном. Под действием этого гормона в кровь возвращается достаточно натрия для поддержания нормального осмотического давления и нормального объема внеклеточной жидкости.
Суточная потребность в хлористом натрии составляет 5–10 г. Эта величина возрастает при поглощении больших количеств жидкости, когда усиливается потоотделение и выделяется больше мочи.
Калий.
В отличие от натрия, калий содержится в клетках в больших количествах, но во внеклеточной жидкости его мало. Главная функция калия – регулирование внутриклеточного осмотического давления и поддержание кислотно-щелочного равновесия.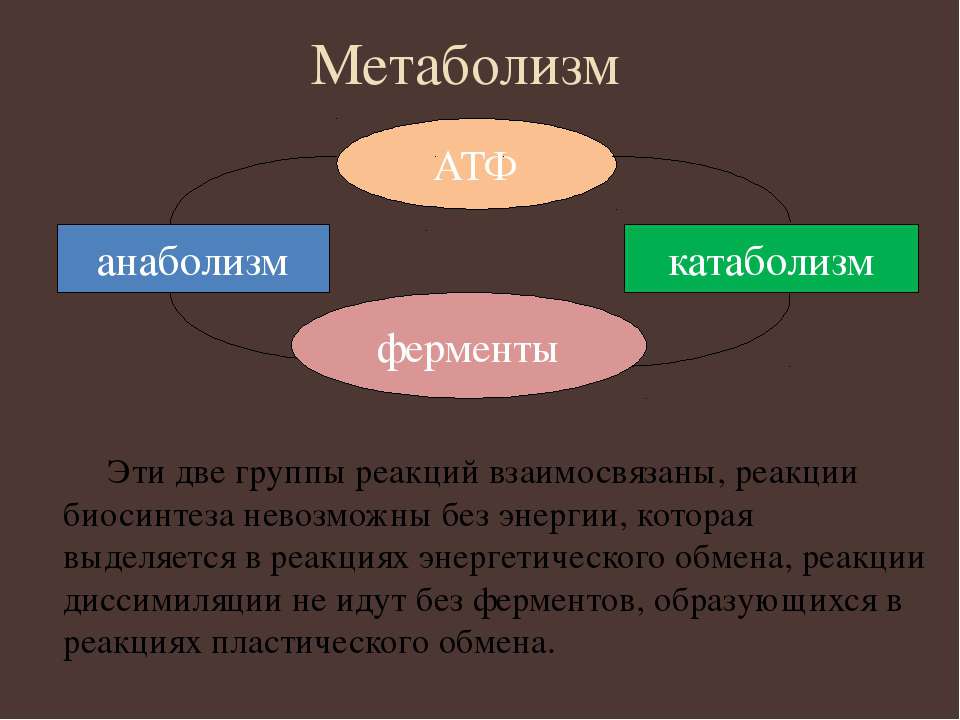 Он также играет важную роль в проведении нервного импульса и во многих ферментных системах, в том числе и в тех, которые участвуют в мышечном сокращении. Калий широко распространен в природе, и его много в любой пище, так что спонтанно калиевая недостаточность возникнуть не может. В плазме концентрация калия регулируется альдостероном, стимулирующим его экскрецию с мочой.
Он также играет важную роль в проведении нервного импульса и во многих ферментных системах, в том числе и в тех, которые участвуют в мышечном сокращении. Калий широко распространен в природе, и его много в любой пище, так что спонтанно калиевая недостаточность возникнуть не может. В плазме концентрация калия регулируется альдостероном, стимулирующим его экскрецию с мочой.
Сера.
С пищей сера поступает в организм главным образом в составе двух аминокислот – цистина и метионина. На конечных этапах метаболизма этих аминокислот сера высвобождается и в результате окисления переводится в неорганическую форму. В составе цистина и метионина сера присутствует в структурных белках. Важную роль играет также сульфгидрильная (–SH) группа цистеина, от которой зависит активность многих ферментов.
Большая часть серы выводится с мочой в виде сульфата. Небольшое количество экскретируемого сульфата обычно связано с органическими соединениями типа фенолов.
Магний.
Метаболизм магния сходен с метаболизмом кальция, и в виде комплекса с фосфатом этот элемент тоже входит в состав костной ткани. Магний присутствует во всех живых клетках, где он функционирует как необходимый компонент многих ферментных систем; эта его роль была убедительно продемонстрирована на примере углеводного обмена в мышцах. Магний, как и калий, широко распространен, и вероятность возникновения его недостаточности очень мала.
Магний присутствует во всех живых клетках, где он функционирует как необходимый компонент многих ферментных систем; эта его роль была убедительно продемонстрирована на примере углеводного обмена в мышцах. Магний, как и калий, широко распространен, и вероятность возникновения его недостаточности очень мала.
Железо.
Железо входит в состав гемоглобина и других гемопротеинов, а именно миоглобина (мышечного гемоглобина), цитохромов (дыхательных ферментов) и каталазы, а также в состав некоторых ферментов, не содержащих гемогруппы. Всасывается железо в верхних отделах кишечника, причем это единственный элемент, всасывающийся только тогда, когда его запас в организме полностью исчерпан. В плазме железо транспортируется в соединении с белком (трансферрином). Через почки железо не выводится; избыток его накапливается в печени в соединении с особым белком (ферритином).
Микроэлементы.
У каждого микроэлемента, присутствующего в организме, своя особая функция, связанная с тем, что он стимулирует действие того или иного фермента или как-либо иначе на него влияет. Цинк необходим для кристаллизации инсулина; кроме того, он является компонентом карбоангидразы (фермента, участвующего в транспорте диоксида углерода) и некоторых других ферментов. Молибден и медь – тоже необходимые компоненты различных ферментов. Иод требуется для синтеза трииодтиронина, гормона щитовидной железы. Фтор (входящий в состав зубной эмали) способствует предотвращению кариеса.
Цинк необходим для кристаллизации инсулина; кроме того, он является компонентом карбоангидразы (фермента, участвующего в транспорте диоксида углерода) и некоторых других ферментов. Молибден и медь – тоже необходимые компоненты различных ферментов. Иод требуется для синтеза трииодтиронина, гормона щитовидной железы. Фтор (входящий в состав зубной эмали) способствует предотвращению кариеса.
ИСПОЛЬЗОВАНИЕ МЕТАБОЛИТОВ
Углеводы.
Всасывание.
Моносахариды, или простые сахара, высвобождающиеся при переваривании углеводов пищи, переходят из кишечника в кровоток в результате процесса, называемого всасыванием. Механизм всасывания представляет собой сочетание простой диффузии и химической реакции (активного всасывания). Одна из гипотез, касающихся природы химической фазы процесса, предполагает, что в этой фазе моносахариды соединяются с фосфорной кислотой в реакции, катализируемой ферментом из группы киназ, после чего проникают в кровеносные сосуды и здесь высвобождаются в результате ферментативного дефосфорилирования (разрыва фосфатной связи), катализируемого одной из фосфатаз. Именно активным всасыванием объясняется то, что разные моносахариды всасываются с разной скоростью и что углеводы всасываются даже тогда, когда уровень сахара в крови выше, чем в кишечнике, т.е. в условиях, когда естественно было бы ожидать их перемещения в обратном направлении – из крови в кишечник.
Именно активным всасыванием объясняется то, что разные моносахариды всасываются с разной скоростью и что углеводы всасываются даже тогда, когда уровень сахара в крови выше, чем в кишечнике, т.е. в условиях, когда естественно было бы ожидать их перемещения в обратном направлении – из крови в кишечник.
Механизмы гомеостаза.
Поступающие в кровоток моносахариды повышают уровень сахара в крови. При голодании концентрация глюкозы в крови колеблется обычно от 70 до 100 мг на 100 мл крови. Этот уровень поддерживается с помощью механизмов, называемых механизмами гомеостаза (самостабилизации). Как только уровень сахара в крови в результате всасывания из кишечника повышается, в действие вступают процессы, выводящие сахар из крови, так что уровень его колеблется не слишком сильно.
Подобно глюкозе, все прочие моносахариды поступают из кровотока в печень, где превращаются в глюкозу. Теперь они неотличимы как от глюкозы, которая всосалась, так и от той, что уже была в организме, и подвергаются тем же метаболическим превращениям. Один из механизмов гомеостаза углеводов, функционирующий в печени, – это гликогенез, посредством которого глюкоза переходит из крови в клетки, где превращается в гликоген. Гликоген хранится в печени до тех пор, пока не произойдет снижение уровня сахара в крови: в этой ситуации гомеостатический механизм вызовет распад накопленного гликогена до глюкозы, которая вновь поступит в кровь.
Один из механизмов гомеостаза углеводов, функционирующий в печени, – это гликогенез, посредством которого глюкоза переходит из крови в клетки, где превращается в гликоген. Гликоген хранится в печени до тех пор, пока не произойдет снижение уровня сахара в крови: в этой ситуации гомеостатический механизм вызовет распад накопленного гликогена до глюкозы, которая вновь поступит в кровь.
Превращения и использование.
Поскольку кровь поставляет глюкозу во все ткани тела и все ткани используют ее для получения энергии, уровень глюкозы в крови снижается главным образом за счет ее использования.
В мышцах глюкоза крови превращается в гликоген. Однако мышечный гликоген не может быть использован для получения глюкозы, которая перешла бы в кровь. В нем заключен запас энергии, и скорость его использования зависит от мышечной активности. В мышечной ткани содержатся два соединения с большим запасом легко доступной энергии в форме богатых энергией фосфатных связей – креатинфосфат и аденозинтрифосфат (АТФ). При отщеплении от этих соединений их фосфатных групп высвобождается энергия для мышечного сокращения. Чтобы мышца вновь могла сокращаться, эти соединения должны быть восстановлены в своей исходной форме. Для этого требуется энергия, которую поставляет окисление продуктов распада гликогена. При мышечном сокращении гликоген превращается в глюкозофосфат, а затем – через ряд реакций – во фруктозодифосфат. Фруктозодифосфат распадается на два трехуглеродных соединения, из которых после ряда этапов образуется сначала пировиноградная кислота, а в конечном итоге – молочная кислота, как об этом уже говорилось при описании метаболизма углеводов. Это превращение гликогена в молочную кислоту, сопровождающееся высвобождением энергии, может происходить в отсутствие кислорода.
При отщеплении от этих соединений их фосфатных групп высвобождается энергия для мышечного сокращения. Чтобы мышца вновь могла сокращаться, эти соединения должны быть восстановлены в своей исходной форме. Для этого требуется энергия, которую поставляет окисление продуктов распада гликогена. При мышечном сокращении гликоген превращается в глюкозофосфат, а затем – через ряд реакций – во фруктозодифосфат. Фруктозодифосфат распадается на два трехуглеродных соединения, из которых после ряда этапов образуется сначала пировиноградная кислота, а в конечном итоге – молочная кислота, как об этом уже говорилось при описании метаболизма углеводов. Это превращение гликогена в молочную кислоту, сопровождающееся высвобождением энергии, может происходить в отсутствие кислорода.
При недостатке кислорода молочная кислота накапливается в мышцах, диффундирует в кровоток и поступает в печень, где из нее вновь образуется гликоген. Если кислорода достаточно, то молочная кислота в мышцах не накапливается. Вместо этого она, как это описано выше, полностью окисляется через цикл трикарбоновых кислот до диоксида углерода и воды с образованием АТФ, который может быть использован для сокращения.
Вместо этого она, как это описано выше, полностью окисляется через цикл трикарбоновых кислот до диоксида углерода и воды с образованием АТФ, который может быть использован для сокращения.
Метаболизм углеводов в нервной ткани и эритроцитах отличается от метаболизма в мышцах тем, что гликоген здесь не участвует. Однако и здесь промежуточными продуктами являются пировиноградная и молочная кислоты, образующиеся при расщеплении глюкозофосфата.
Глюкоза используется не только в клеточном дыхании, но и во многих других процессах: синтезе лактозы (молочного сахара), образовании жиров, а также особых сахаров, входящих в состав полисахаридов соединительной ткани и ряда других тканей.
Гликоген печени, синтезируемый при всасывании углеводов в кишечнике, служит самым доступным источником глюкозы, когда всасывание отсутствует. Если этот источник оказывается исчерпанным, в печени начинается процесс глюконеогенеза. Глюкоза образуется при этом из некоторых аминокислот (из 100 г белка образуется 58 г глюкозы) и нескольких других неуглеводных соединений, в том числе из глицериновых остатков нейтральных жиров.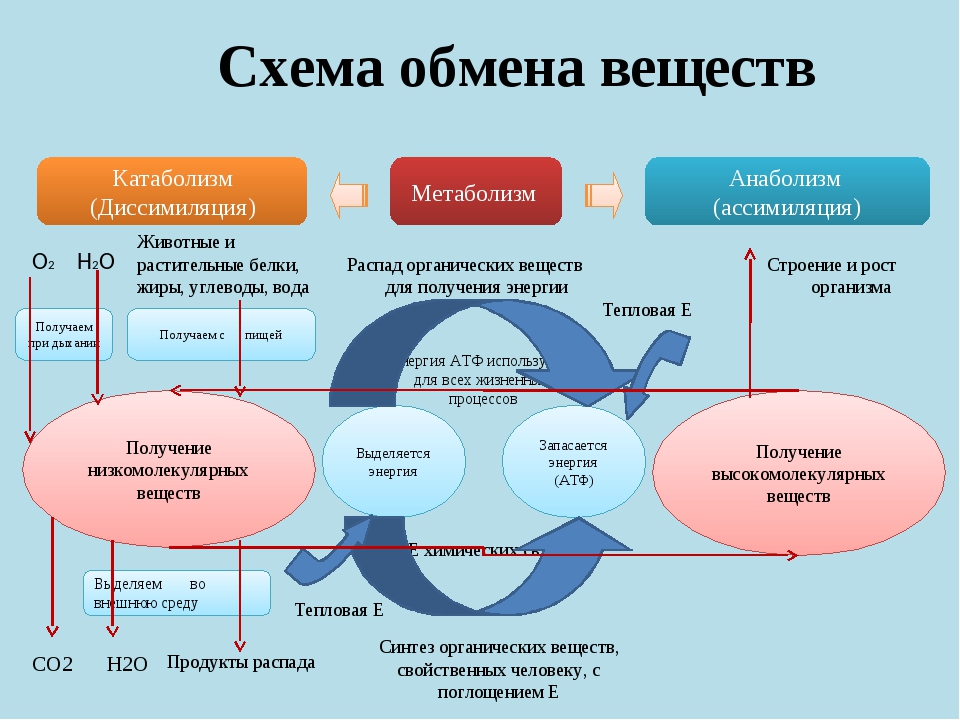
Некоторую, хотя и не столь важную, роль в метаболизме углеводов играют почки. Они выводят из организма избыток глюкозы, когда ее концентрация в крови слишком высока; при меньших концентрациях глюкоза практически не выводится.
В регулировании метаболизма углеводов участвует несколько гормонов, в том числе гормоны поджелудочной железы, передней доли гипофиза и коры надпочечников.
Гормон поджелудочной железы инсулин снижает концентрацию глюкозы в крови и повышает ее концентрацию в клетках. По-видимому, он стимулирует также и запасание гликогена в печени. Кортикостерон, гормон коры надпочечников, и адреналин, вырабатываемый мозговым веществом надпочечников, воздействуют на метаболизм углеводов, стимулируя распад гликогена (главным образом в мышцах и печени) и синтез глюкозы (в печени).
Липиды.
Всасывание.
В кишечнике после переваривания жиров остаются главным образом свободные жирные кислоты с небольшой примесью холестерина и лецитина и следами жирорастворимых витаминов. Все эти вещества очень тонко диспергированы благодаря эмульгирующему и солюбилизирующему действию солей желчных кислот. Солюбилизирующее действие обычно связывают с образованием нестойких химических соединений между жирными кислотами и солями желчных кислот. Эти комплексы проникают в клетки эпителия тонкого кишечника и здесь распадаются на жирные кислоты и соли желчных кислот. Последние переносятся в печень и вновь секретируются с желчью, а жирные кислоты вступают в соединение с глицерином или холестерином. Образовавшиеся реконструированные жиры поступают в лимфатические сосуды брыжейки в форме млечного сока, т.н. «хилуса». Из сосудов брыжейки хилус по лимфатической системе через грудной проток поступает в кровеносную систему.
Все эти вещества очень тонко диспергированы благодаря эмульгирующему и солюбилизирующему действию солей желчных кислот. Солюбилизирующее действие обычно связывают с образованием нестойких химических соединений между жирными кислотами и солями желчных кислот. Эти комплексы проникают в клетки эпителия тонкого кишечника и здесь распадаются на жирные кислоты и соли желчных кислот. Последние переносятся в печень и вновь секретируются с желчью, а жирные кислоты вступают в соединение с глицерином или холестерином. Образовавшиеся реконструированные жиры поступают в лимфатические сосуды брыжейки в форме млечного сока, т.н. «хилуса». Из сосудов брыжейки хилус по лимфатической системе через грудной проток поступает в кровеносную систему.
После переваривания пищи содержание липидов в крови возрастает приблизительно от 500 мг (уровень при голодании) до 1000 мг на 100 мл плазмы. Присутствующие в крови липиды представляют собой смесь жирных кислот, нейтральных жиров, фосфолипидов (лецитина и кефалина), холестерина и эфиров холестерина.
Распределение.
Кровь доставляет липиды в разные ткани тела и прежде всего в печень. Печень обладает способностью модифицировать поступающие в нее жирные кислоты. Это особенно выражено у видов, запасающих жиры с высоким содержанием насыщенных или, наоборот, ненасыщенных жирных кислот: в печени этих животных соотношение насыщенных и ненасыщенных кислот изменяется таким образом, что отлагающийся жир по своему составу соответствует жиру, свойственному данному организму.
Жиры в печени либо используются для получения энергии, либо переходят в кровь и доставляются ею в разные ткани. Здесь они могут включаться в структурные элементы тканей, но большая их часть отлагается в жировых депо, где они хранятся до тех пор, пока не возникнет потребность в энергии; тогда они снова переносятся в печень и подвергаются здесь окислению.
Метаболизм липидов, как и углеводов, регулируется гомеостатически. Механизмы гомеостаза, воздействующие на липидный и углеводный обмен, видимо, тесно связаны, поскольку при замедлении метаболизма углеводов усиливается метаболизм липидов, и наоборот.
Превращения и использование.
Четырехуглеродные кислоты – ацетоуксусная (продукт конденсации двух ацетатных единиц) и b-гидроксимасляная – и трехуглеродное соединение ацетон, образующийся при отщеплении одного атома углерода от ацетоуксусной кислоты, известны под общим названием кетоновых (ацетоновых) тел. В норме кетоновые тела присутствуют в крови в небольших количествах. Избыточное их образование при тяжелом диабете ведет к повышению их содержания в крови (кетонемия) и в моче (кетонурия) – это состояние обозначают термином «кетоз».
Белки.
Всасывание.
При переваривании белков пищеварительными ферментами образуется смесь из аминокислот и небольших пептидов, содержащих от двух до десяти остатков аминокислот. Эти продукты всасываются слизистой кишечника, и здесь гидролиз завершается – пептиды также распадаются до аминокислот. Поступившие в кровь аминокислоты смешиваются с находящимися здесь такими же аминокислотами. В крови содержится смесь из аминокислот, поступивших из кишечника, образовавшихся при распаде тканевых белков и синтезированных организмом заново.
Синтез.
В тканях непрерывно идет распад белков и их новообразование. Содержащиеся в крови аминокислоты избирательно поглощаются тканями как исходный материал для построения белков, а из тканей в кровь поступают другие аминокислоты. Синтезу и распаду подвергаются не только структурные белки, но и белки плазмы крови, а также белковые гормоны и ферменты.
Во взрослом организме аминокислоты или белки практически не запасаются, поэтому удаление аминокислот из крови происходит с такой же скоростью, как и их поступление из тканей в кровь. В растущем организме формируются новые ткани, и на этот процесс расходуется больше аминокислот, чем поступает в кровь за счет распада тканевых белков.
Печень участвует в метаболизме белков самым активным образом. Здесь синтезируются белки плазмы крови – альбумины и глобулины – а также собственные ферменты печени. Так, при потере плазменных белков содержание альбумина в плазме восстанавливается – за счет интенсивного синтеза – довольно быстро. Аминокислоты в печени используются не только для образования белков, но подвергаются также расщеплению, в ходе которого извлекается заключенная в них энергия.
Аминокислоты в печени используются не только для образования белков, но подвергаются также расщеплению, в ходе которого извлекается заключенная в них энергия.
Превращения и использование.
Если аминокислоты используются в качестве источника энергии, то отщепляемая от них аминогруппа (–NH2) направляется на образование мочевины, а не содержащий азота остаток молекулы окисляется приблизительно так же, как глюкоза или жирные кислоты.
Так называемый «орнитиновый цикл» описывает, как происходит превращение аммиака в мочевину. В этом цикле аминогруппа, отщепившаяся от аминокислоты в форме аммиака, присоединяется вместе с диоксидом углерода к молекуле орнитина с образованием цитруллина. Цитруллин присоединяет второй атом азота, на этот раз от аспарагиновой кислоты, и превращается в аргинин. Далее аргинин подвергается гидролизу с образованием мочевины и орнитина. Орнитин может теперь вновь вступить в цикл, а мочевина выводится из организма через почки как один из конечных продуктов метаболизма.
См. также ГОРМОНЫ; ФЕРМЕНТЫ; ЖИРЫ И МАСЛА; НУКЛЕИНОВЫЕ КИСЛОТЫ; БЕЛКИ; ВИТАМИНЫ.
Сайт преподавателя биологии и химии Дмитрия Андреевича Соловкова
Обновлены задания 3, 6, 11, 27 и 28 по биологии — добавлены новые номера из ЕГЭ-2020.
Рубрика: Абитуриенту
Обновил задания 1, 2, 20 и 21 в первой части — добавил около 30 новых номеров.
Рубрика: Абитуриенту
За последний месяц обновлены все задания второй части экзамена (№№30-35) — добавлены задания образца 2020 года. Согласно проекту ЕГЭ-2021, изменений во второй части по химии не планируется.
Рубрика: Абитуриенту
Обновлено задание 33 — добавлено около 15 новых цепочек.
Рубрика: Без рубрики
|
В задания 34 добавлен новый раздел — задачи на атомистику и молярные соотношения. Также обновлены задачи по электролизу с учетом новых задач ЕГЭ-2020.
Также обновлены задачи по электролизу с учетом новых задач ЕГЭ-2020.
Рубрика: Абитуриенту
|
Обновлены задания 32 и 35 — добавлены задачи 2020 года
Рубрика: Абитуриенту
|
Обновлены задания 30 и 31 — добавлены номера образца 2020 года. Посмотреть их можно здесь.
Рубрика: Абитуриенту
Задания 19 и 20 первой части обновлены в соответствии со спецификатором ЕГЭ-2021 года. В задание 24 добавлено около 10 номеров.
Рубрика: Абитуриенту
Выложены последние 2 презентации по цитологии — репликация ДНК и биосинтез белка. Таким образом, полностью закончен целый раздел общей биологии. Всем рекомендую для подготовки к ЕГЭ.
Рубрика: Абитуриенту
|
Добавлены очередные презентации по цитологии: по диссимиляции и автотрофному питанию
Рубрика: Абитуриенту
Метаболизм как основа жизнедеятельности клетки
Метаболизм как основа жизнедеятельности клетки
Под метаболизмом понимают постоянно происходящий в клетках живых организмов обмен веществ и энергии. Одни соединения, выполнив свою функцию, становятся ненужными, в других возникает насущная потребность. В различных процессах метаболизма из простых веществ при участии ферментов синтезируются высокомолекулярные соединения, в свою очередь сложные молекулы расщепляются на более простые.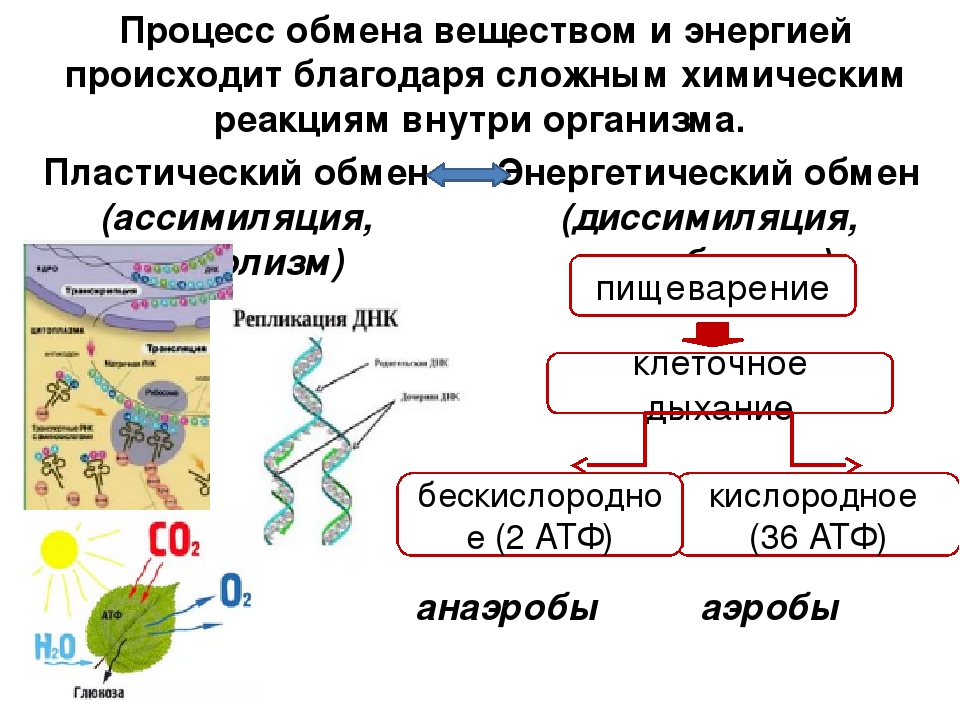
Реакции биологического синтеза называются анаболическими (греч. anabole подъем), а их совокупность в клетке — анаболизмом, или пластическим обменом (греч. plastos вылепленный, созданный).
В клетке протекает огромное количество процессов синтеза: липидов в эндоплазматической сети, белков на рибосомах, полисахаридов в комплексе Гольджи эукариот и в цитоплазме прокариот, углеводов в пластидах растений. Структура синтезируемых макромолекул обладает видовой и индивидуальной специфичностью. Набор характерных для клетки веществ соответствует последовательности нуклеотидов ДНК, составляющих генотип. Для обеспечения реакций синтеза клетке требуются существенные затраты энергии, получаемой при расщеплении веществ.
Совокупность реакций расщепления сложных молекул на более простые носит название катаболизма (греч. katabole разрушение), или энергетического обмена. Примерами таких реакций является расщепление липидов, полисахаридов, белков и нуклеиновых кислот в лизосомах, а также простых углеводов и жирных кислот в митохондриях.
В результате процессов катаболизма высвобождается энергия. Существенная ее часть запасается в виде высокоэнергетичных химических связей АТФ. Запасы АТФ позволяют организму быстро и эффективно обеспечивать различные процессы жизнедеятельности.
Молекулы белков функционируют в организме от нескольких часов до нескольких дней. За этот период в них накапливаются нарушения, и белки становятся непригодными для выполнения своих функций. Они расщепляются и заменяются на вновь синтезируемые. Требуют постоянного обновления и сами клеточные структуры.
Пластический и энергетический обмены неразрывно взаимосвязаны. Процессы расщепления осуществляют энергетическое обеспечение процессов синтеза, а также поставляют необходимые для синтеза строительные вещества. Правильный обмен веществ поддерживает постоянство химического состава биологических систем, их внутренней среды. Способность организмов сохранять внутренние параметры неизменными носит название гомеостаза. Процессы метаболизма происходят в соответствии с генетической программой клетки, реализуя ее наследственную информацию.
Энергетический обмен в клетке. Синтез АТФ
Человек и животные получают энергию за счет окисления органических соединений, поступающих с пищей. Биологическое окисление веществ — это, по сути, медленное горение. Конечные продукты сгорания дров (целлюлозы) — углекислый газ и вода. Полное окисление органических веществ (углеводов и липидов) в клетках также происходит до воды и углекислого газа. В отличие от горения, процесс биологического окисления происходит постепенно. Высвобождающаяся энергия также постепенно запасается в виде химических связей синтезируемых соединений. Некоторая ее часть рассеивается в клетках, поддерживая необходимую для жизнедеятельности температуру.
Синтез АТФ происходит главным образом в митохондриях (у растений еще и в хлоропластах) и обеспечивается в основном энергией, выделяющейся при расщеплении глюкозы, но могут использоваться и другие простые органические соединения — сахара, жирные кислоты и пр.
Гликолиз. Процесс расщепления глюкозы в живых организмах носит название гликолиза (греч. glykys сладкий + lysis расщепление). Рассмотрим основные его этапы.
glykys сладкий + lysis расщепление). Рассмотрим основные его этапы.
На первой, предварительной стадии в лизосомах происходит образование простых органических молекул путем расщепления ди- и полисахаридов. Выделяющееся при этом небольшое количество энергии рассеивается в виде тепла.
Второй этап гликолиза происходит в цитоплазме без участия кислорода и называется анаэробным (бескислородным — греч. ana без + aer воздух) гликолизом — неполным окислением глюкозы без участия кислорода.
Бескислородный гликолиз представляет собой сложный многоступенчатый процесс из десяти последовательных реакций. Каждая реакция катализируется специальным ферментом. В итоге глюкоза расщепляется до пировиноградной кислоты (ПВК):
С6Н12О6(глюкоза) + 2Н3РО4 + 2АДФ = 2С3Н4О3(ПВК) + 2АТФ + 2Н2О
Глюкоза в этом процессе не только расщепляется, но и окисляется (теряет атомы водорода). В мышцах человека и животных две молекулы ПВК, приобретая атомы водорода, восстанавливаются в молочную кислоту С3Н6О3. Этим же продуктом заканчивается гликолиз у молочнокислых бактерий и грибков, применяемый для приготовления кислого молока, простокваши, кефира, а также при силосовании кормов в животноводстве. Процесс превращения ПВК в клетках микроорганизмов и растений в устойчивые конечные продукты называют брожением.
Этим же продуктом заканчивается гликолиз у молочнокислых бактерий и грибков, применяемый для приготовления кислого молока, простокваши, кефира, а также при силосовании кормов в животноводстве. Процесс превращения ПВК в клетках микроорганизмов и растений в устойчивые конечные продукты называют брожением.
Так, дрожжевые грибки расщепляют ПВК на этиловый спирт и углекислый газ. Этот процесс, называемый спиртовым брожением, используют для приготовления кваса, пива и вина. Брожение других микроорганизмов завершается образованием ацетона, уксусной кислоты и т.д.
Главным результатом анаэробного гликолиза во всех организмах является образование двух молекул АТФ. Высвобождающаяся при расщеплении глюкозы энергия относительно невелика — 200 кДж/моль. Высокоэнергетичные связи АТФ запасают 40% этой величины. Остальные 60% рассеиваются в виде тепла. Основной выход энергии и молекул АТФ происходит на третьем, кислородном этапе гликолиза, называемом еще аэробным дыханием.
Кислородный гликолиз. При наличии достаточного количества кислорода дальнейший процесс расщепления ПВК происходит уже не в цитоплазме, а в митохондриях, и включает несколько десятков последовательных реакций, каждая из которых обслуживается своим комплексом ферментов.
При наличии достаточного количества кислорода дальнейший процесс расщепления ПВК происходит уже не в цитоплазме, а в митохондриях, и включает несколько десятков последовательных реакций, каждая из которых обслуживается своим комплексом ферментов.
Молекулы ПВК под действием ферментов (и кофермента НАД — никотинамидадениндинуклеотида) поэтапно окисляются сначала до уксусной кислоты, а затем, в так называемом цикле Кребса (или трикарбоновых кислот), до углекислого газа и воды (медленное горение). В процессе окисления образуются сложные молекулярные соединения с присоединенными к ним атомами водорода. Молекулы-переносчики подхватывают и перемещают электроны этих атомов по длинной цепи ферментов от одного к другому. На каждом шаге электроны вступают в окислительно-восстановительные реакции и отдают свою энергию, которая идет на перемещение протонов на внешнюю сторону внутренней мембраны митохондрии.
В результате оставшиеся протоны и перемещенные электроны оказываются на разных сторонах внутренней мембраны. На мембране создается разность потенциалов.
На мембране создается разность потенциалов.
Фермент, синтезирующий АТФ (АТФ-синтетаза), встроен во внутреннюю мембрану по всей ее толщине. Этот фермент имеет характерную особенность: небольшой каналец в молекулярной структуре. При накоплении на мембране разности потенциалов примерно в 200 мВ ионы Н+ начинают протискиваться через каналец в молекуле АТФ-синтетазы. В процессе энергичного продвижения ионов через фермент происходит синтез АТФ из АДФ с участием фосфорной кислоты.
В химических реакциях кислородного гликолиза освобождается большое количество энергии — 2600 кДж/моль. Существенная ее часть (55%) запасается в высокоэнергетичных связях образующихся молекул АТФ. Остальные 45% рассеиваются в виде тепла (поэтому при выполнении физической работы нам жарко). Итоговое уравнение кислородной стадии выглядит следующим образом:
2С3Н6О3(молочн.кислота) + 6О2 + 36Н3РО4 + 36АДФ = 6СО2 + 42Н2О + 36АТФ
Таким образом, кислородное расщепление резко увеличивает эффективность энергетического обмена и играет основную роль в аккумулировании энергии. Если гликолиз без участия кислорода дает только 2 молекулы АТФ, то кислородный гликолиз обеспечивает синтез 36 молекул АТФ. В итоге в полном цикле гликолиза на каждую молекулу глюкозы образуется 38 молекул АТФ.
Если гликолиз без участия кислорода дает только 2 молекулы АТФ, то кислородный гликолиз обеспечивает синтез 36 молекул АТФ. В итоге в полном цикле гликолиза на каждую молекулу глюкозы образуется 38 молекул АТФ.
При среднесуточных энергетических затратах в 10 тыс. кДж в организме человека ежедневно синтезируется около 170 кг АТФ, а содержится всего около 50 г АТФ, следовательно, возобновление запаса происходит с частотой 3400 раз в сутки!
При интенсивной физической работе клетки организма не успевают насытиться кислородом, и расщепление глюкозы ограничивается бескислородным гликолизом. В результате быстро накапливается молочная кислота — токсичное для нервных и мышечных клеток соединение (вспомним мышечные боли после тяжелой работы). Появление молочной кислоты возбуждает дыхательный центр и заставляет нас усиленно дышать. Насыщение клеток кислородом позволяет организму возобновить процесс кислородного расщепления, обеспечивающий необходимое количество энергии в виде молекул АТФ.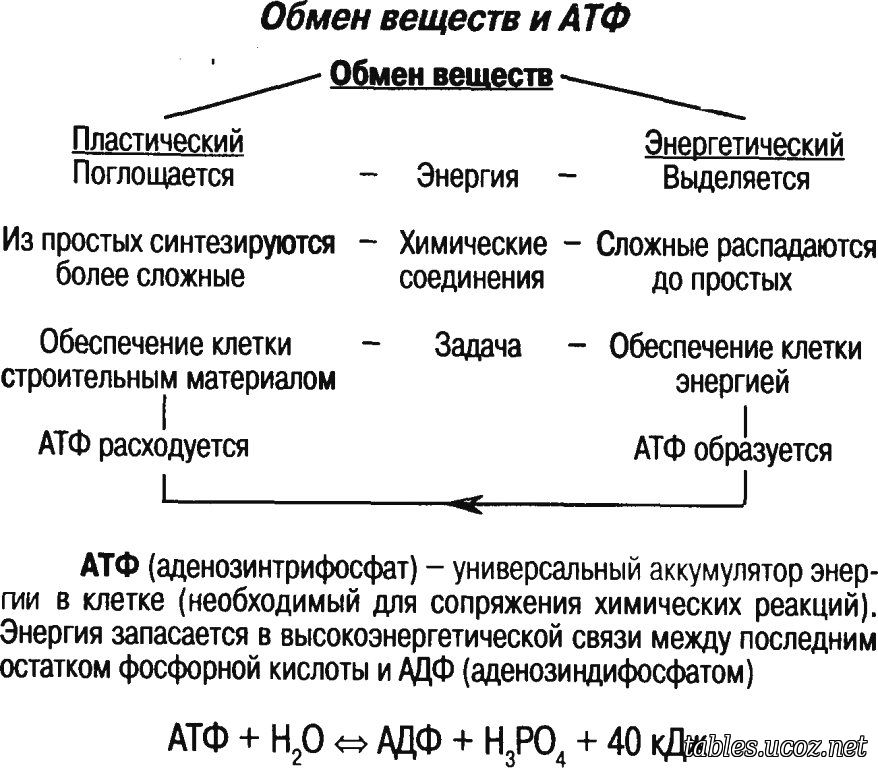 Наступает «второе дыхание». Гепардам после интенсивного бега требуется продолжительный отдых, порой они оказываются не в состоянии защитить свою добычу от менее сильных хищников. В большой скорости восстановления кислородного запаса, а значит, в лучшей приспособленности к длительной мышечной активности — преимущество многих мелких животных.
Наступает «второе дыхание». Гепардам после интенсивного бега требуется продолжительный отдых, порой они оказываются не в состоянии защитить свою добычу от менее сильных хищников. В большой скорости восстановления кислородного запаса, а значит, в лучшей приспособленности к длительной мышечной активности — преимущество многих мелких животных.
Митохондрии способны использовать для синтеза АТФ не только расщепление глюкозы. В их матриксе содержатся также ферменты, расщепляющие жирные кислоты. Особенностью этого цикла является большой энергетический выход — 51 молекула АТФ на каждую молекулу жирной кислоты. Не случайно медведи и другие животные, впадая в спячку, запасают именно жиры. Любопытно, что часть запасаемого жира имеет у них бурый цвет. Такие жировые клетки содержат множество митохондрий необычного строения: их внутренние мембраны пронизаны порами. Ионы водорода свободно проходят через эти поры, и синтез АТФ в клетках бурого жира не происходит. Вся энергия, освобождающаяся в процессе кислородного расщепления жирных кислот, выделяется в виде большого количества тепла, согревающего животных во время долгой зимней спячки.
Бурый жир составляет не более 1-2% массы тела, но повышает производство тепла до 400 Вт на каждый килограмм веса (теплопроизводство человека в состоянии покоя составляет 1 Вт/кг). Запасают жир и верблюды. При постоянном дефиците влаги это вдвойне выгодно, поскольку расщепление жиров дает еще и большое количество воды.
Кроме глюкозы и жирных кислот, митохондрии способны расщеплять аминокислоты, но они — дорогое топливо. Аминокислоты являются важным строительным материалом, из них организм синтезирует свои белки. К тому же использование аминокислот для синтеза АТФ требует предварительного удаления аминогруппы NН2 с образованием токсичного аммиака. Белки и составляющие их аминокислоты используются клеткой для получения энергии только в крайнем случае.
Этиловый спирт тоже может использоваться митохондриями для синтеза АТФ. Но спирт как «топливо» имеет для организма человека свои недостатки, постоянное употребление алкоголя приводит к тяжелым расстройствам, например, к жировому перерождению печени — циррозу.
Вопросы
1. Как связаны катаболизм, анаболизм и гомеостаз?
2. Что называют брожением? Приведите примеры.
3. Опишите ход кислородного гликолиза. В чем его основной результат?
4. Почему при выполнении физической работы нам жарко?
5. Каковы функции бурого жира?
Фотосинтез — превращение энергии света в энергию химических связей
Автотрофные организмы. В отличие от человека и животных, все зеленые растения и часть бактерий способны синтезировать органические вещества из неорганических соединений. Такой тип обмена веществ называется автотрофным (греч. autos сам + trophe пища). В зависимости от вида энергии, используемой автотрофами для синтеза органических молекул, их делят на фототрофов и хемотрофов. Фототрофы используют энергию солнечного света, а хемотрофы — химическую энергию, высвобождающуюся при окислении ими различных неорганических соединений.
Зеленые растения являются фототрофами. Их хлоропласты содержат хлорофилл, позволяющий растениям осуществлять фотосинтез — преобразование энергии солнечного света в энергию химических связей синтезируемых органических соединений. Из всего спектра солнечного излучения молекулы хлорофилла поглощают красную и синюю часть, а зеленая составляющая достигает сетчатки наших глаз. Поэтому большинство растений мы видим зелеными.
Из всего спектра солнечного излучения молекулы хлорофилла поглощают красную и синюю часть, а зеленая составляющая достигает сетчатки наших глаз. Поэтому большинство растений мы видим зелеными.
Для осуществления фотосинтеза растения поглощают из атмосферы углекислый газ, а из водоемов и почвы — воду, неорганические соли азота и фосфора. Итоговое уравнение фотосинтеза выглядит довольно просто:
6СО2 + 6Н2О = С6Н12О6(глюкоза) + 6О2,
но всем хорошо известно, что при смешивании углекислого газа и воды глюкоза не образуется. Фотосинтез — сложный многоступенчатый процесс, для прохождения которого необходим не только солнечный свет и хлорофилл, но и ряд ферментов, энергия АТФ и молекулы-переносчики. Выделяют две фазы фотосинтеза — световую и темновую.
С в е т о в а я ф а з а фотосинтеза начинается с освещения растений светом. Солнечные фотоны, передавая свою энергию молекуле хлорофилла, переводят молекулу в возбужденное состояние: ее электроны, получая дополнительную энергию, переходят на более высокие орбиты. Отрыв таких возбужденных электронов может происходить значительно легче, чем невозбужденных. Молекулы-переносчики захватывают их и перемещают на другую сторону мембраны тилакоида.
Отрыв таких возбужденных электронов может происходить значительно легче, чем невозбужденных. Молекулы-переносчики захватывают их и перемещают на другую сторону мембраны тилакоида.
Молекулы хлорофилла восполняют потерю электронов, отрывая их от молекул воды. В результате вода расщепляется на протоны и молекулярный кислород:
2Н2О – 4е = 4Н+ + О2
Процесс расщепления молекул воды до молекулярного кислорода, протонов и электронов под действием света называют фотолизом. Молекулярный кислород легко диффундирует сквозь мембраны тилакоидов и выделяется в атмосферу. Протоны неспособны к проникновению через мембрану и остаются внутри.
Таким образом, снаружи мембраны накапливаются электроны, доставленные молекулами-переносчиками с возбужденных молекул хлорофилла, а внутри — протоны, образовавшиеся в результате фотолиза воды. Возникает разность потенциалов. В мембраны тилакоидов хлоропласта, так же как и во внутренние мембраны митохондрий, встроены ферменты-синтетазы, осуществляющие синтез АТФ. В молекулярной структуре синтетаз растений также имеется каналец, через который могут проходить протоны. При достижении на мембране критической разности потенциалов протоны, влекомые силой электрического поля, протискиваются по канальцу АТФ-синтетазы, затрачивая энергию на синтез АТФ. Соединяясь на другой стороне мембраны с электронами, протоны образуют атомарный водород.
В молекулярной структуре синтетаз растений также имеется каналец, через который могут проходить протоны. При достижении на мембране критической разности потенциалов протоны, влекомые силой электрического поля, протискиваются по канальцу АТФ-синтетазы, затрачивая энергию на синтез АТФ. Соединяясь на другой стороне мембраны с электронами, протоны образуют атомарный водород.
Фотосинтез в хлоропластах весьма эффективен: он дает в 30 раз больше АТФ, чем кислородный гликолиз в митохондриях тех же растений.
Таким образом, во время световой фазы фотосинтеза происходят следующие главные процессы: выделение в атмосферу свободного кислорода, синтез АТФ и образование атомарного водорода.
Протекание дальнейших реакций может происходить и в темноте, потому носит название темновой фазы.
Т е м н о в а я ф а з а. Реакции этой фазы происходят в строме хлоропласта при участии атомарного водорода и АТФ, образовавшихся в световой фазе, а также ферментов, восстанавливающих СО2 до простого сахара — триозы (глицеральдегида) — и синтезирующих из нее глюкозу:
6СО2 + 24Н = С6Н12О6(глюкоза) + 6Н2О
Для образования одной молекулы глюкозы требуется 18 молекул АТФ. Комплекс реакций темновой фазы, осуществляемых ферментами (и коферментом НАД), носит название цикла Кальвина.
Комплекс реакций темновой фазы, осуществляемых ферментами (и коферментом НАД), носит название цикла Кальвина.
Кроме глюкозы, из триозы могут синтезироваться жирные кислоты, аминокислоты и пр. Углеводы и жирные кислоты далее транспортируются в лейкопласты, где из них формируются запасные питательные вещества — крахмал и жиры.
С наступлением темноты растения продолжают процесс фотосинтеза, используя запасенные на свету соединения. Когда этот запас исчерпывается, прекращается и фотосинтез. В ночной темноте растения напоминают по типу обмена веществ животных: они поглощают кислород из атмосферы (дышат) и окисляют при помощи его запасенные днем питательные вещества. На дыхание растения используют в 20-30 раз меньше кислорода, чем выделяют в атмосферу в процессе фотосинтеза.
Количество энергии, производимой растениями, значительно превышает количество тепла, выделяющегося при сжигании всем населением планеты горючих полезных ископаемых. Ежегодно растительность планеты дает 200 млрд. т кислорода и 150 млрд. т органических соединений, необходимых человеку и животным.
т кислорода и 150 млрд. т органических соединений, необходимых человеку и животным.
Хемосинтез. Большинство бактерий лишены хлорофилла. Некоторые из них являются хемотрофами: для синтеза органических веществ они используют не энергию света, а энергию, высвобождающуюся при окислении неорганических соединений. Такой способ получения энергии и синтеза органических веществ назвали хемосинтезом (греч. chemia химия). Явление хемосинтеза открыто в 1887 г. русским микробиологом С. Н. Виноградским.
Н и т р и ф и ц и р у ю щ и е б а к т е р и и. В корневищах растений, главным образом, бобовых, живут особые клубеньковые бактерии. Они способны усваивать недоступный растениям атмосферный азот и обогащать почву аммиаком. Нитрифицирующие бактерии окисляют аммиак клубеньковых бактерий до азотистой кислоты и далее — азотистую до азотной. В результате растения получают соли азотной кислоты, необходимые для синтеза аминокислот и азотистых оснований.
В о д о р о д н ы е б а к т е р и и также широко распространены в почвах. Они окисляют молекулы водорода, образующиеся в результате бескислородного окисления органических останков различными микроорганизмами:
Они окисляют молекулы водорода, образующиеся в результате бескислородного окисления органических останков различными микроорганизмами:
2Н2 + О2 = 2Н2О
Ж е л е з о б а к т е р и и используют энергию, высвобождающуюся при окислении двухвалентного железа до трехвалентного (закисные соли до окисных).
С е р о б а к т е р и и обитают в болотах и «питаются» сероводородом. В результате окисления сероводорода выделяется необходимая для жизнедеятельности бактерий энергия и накапливается сера. При окислении серы до серной кислоты высвобождается еще часть энергии. Суммарный выход энергии составляет существенную величину — 666 кДж/моль. Огромное количество серобактерий обитает в Черном море. Его воды, начиная со стометровой глубины, насыщены сероводородом.
Гетеротрофный тип обмена веществ. Человек и животные не способны синтезировать необходимые для жизнедеятельности органические вещества из неорганических и вынуждены поглощать их с пищей. Такие организмы называют гетеротрофами (греч. heteros другой). К гетеротрофам относятся также большинство бактерий и грибы. Вещества, поступившие с пищей, разлагаются в организмах животных на простые углеводы, аминокислоты, нуклеотиды, из которых далее синтезируются высокомолекулярные соединения, необходимые для конкретного вида существ в конкретной фазе жизненного цикла. Часть поступивших с пищей молекул расщепляется до конечных продуктов, а высвобождающаяся энергия используется в процессах жизнедеятельности. Некоторое количество энергии рассеивается в виде теплоты, служащей для поддержания температуры тела.
heteros другой). К гетеротрофам относятся также большинство бактерий и грибы. Вещества, поступившие с пищей, разлагаются в организмах животных на простые углеводы, аминокислоты, нуклеотиды, из которых далее синтезируются высокомолекулярные соединения, необходимые для конкретного вида существ в конкретной фазе жизненного цикла. Часть поступивших с пищей молекул расщепляется до конечных продуктов, а высвобождающаяся энергия используется в процессах жизнедеятельности. Некоторое количество энергии рассеивается в виде теплоты, служащей для поддержания температуры тела.
Многие одноклеточные водоросли имеют миксотрофное (смешанное) питание. На свету они фотосинтезируют, а в темноте переходят к фагоцитозу, т.е. становятся гетеротрофами.
Вопросы
1. Какова функция фотосинтеза в организмах растений?
2. В чем состоит основное предназначение световой и темновой фаз?
3. Опишите обмен веществ растений в ночное время.
3. Чем отличаются хемотрофы от фототрофов, в чем их сходство? Приведите примеры хемотрофов.
4. Отличается ли человек от растений по типу обмена веществ, кто такие гетеротрофы?
Пластический обмен.Биосинтез белков. Синтез иРНК
В процессах метаболизма реализуется наследственная информация. Клетка синтезирует только те вещества, которые записаны в ее генетической программе. Каждой группе клеток присущ свой комплекс химических соединений. Среди них особенно важными для организма являются белки.
Многие функции и признаки организма определяются его набором белков. Белки-ферменты расщепляют пищу, отвечают за поглощение и выделение солей, синтезируют жиры и углеводы, производят множество других биохимических превращений. Белки определяют цвет глаз, рост — словом, внешнюю специфичность организмов. Большинство белков, выполняющих одни и те же функции, несколько различны даже у особей одного и того же вида (к примеру, белки групп крови). Но некоторые однофункциональные белки могут иметь сходное строение у далеких групп организмов (к примеру, инсулин собаки и человека).
В процессе жизнедеятельности белковые молекулы постепенно разрушаются, теряют свою структуру — денатурируют. Их активность падает, и клетки заменяют их новыми. В организмах постоянно происходит синтез необходимых белков.
иосинтез белковых молекул — сложный ферментативный процесс, начинающийся в ядре и заканчивающийся на рибосомах. Центральную функцию в нем выполняют носители генетической информации — нуклеиновые кислоты ДНК и РНК.
Генетический код. Последовательность нуклеотидов ДНК задает последовательность аминокислот в белках — их первичную структуру. Молекулы ДНК являются матрицами для синтеза всех белков.
Отрезок ДНК, несущий информацию о первичной структуре конкретного белка, называют геном. Соответствующую последовательность нуклеотидов — генетическим кодом белка.
Идею о том, что наследственная информация записана на молекулярном уровне, а синтез белков идет по матричному принципу, впервые высказал еще в 1920-х годах русский биолог Н. К. Кольцов. В настоящее время код ДНК полностью расшифрован. В этом заслуга известных ученых: Г. Гамова (1954), а также Ф. Крика, С. Очоа, М. Ниренберга, Р. Холи и К. Хорана (1961-65). Значительную часть свойств генетического кода установил английский физик Ф. Крик, исследуя бактериофагов.
В настоящее время код ДНК полностью расшифрован. В этом заслуга известных ученых: Г. Гамова (1954), а также Ф. Крика, С. Очоа, М. Ниренберга, Р. Холи и К. Хорана (1961-65). Значительную часть свойств генетического кода установил английский физик Ф. Крик, исследуя бактериофагов.
К о д т р и п л е т е н. Каждая аминокислота в генетическом коде задается последовательностью трех нуклеотидов — триплетом, или кодоном. Различных нуклеотидов в ДНК четыре, следовательно, теоретически возможных кодонов — 64 (43). Большинству аминокислот соответствует от 2 до 6 кодонов — код, как говорят, вырожден. Чем чаще аминокислота встречается в белках, тем, как правило, большим числом кодонов она кодируется. Оставшиеся три кодона вместе с кодоном метионина (АУГ) служат знаками препинания при считывании информации — указывают начало и конец матриц конкретных белков. Если белок имеет несколько полимерных цепей (образующих отдельные глобулы), то знаки препинания выделяют полипептидные звенья. Считывание каждого звена происходит непрерывно, без знаков препинания и пропусков — триплет за триплетом.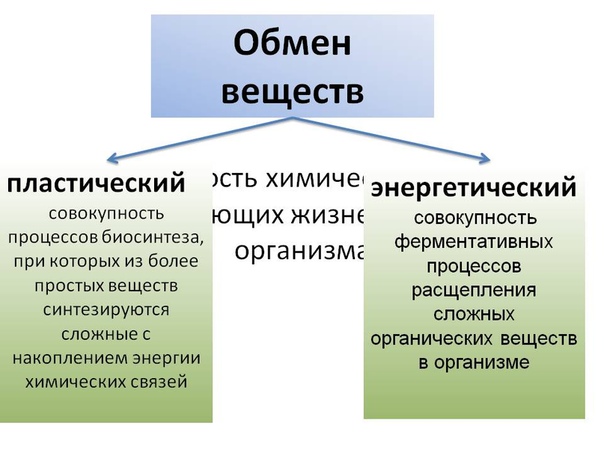
К о д о д н о з н а ч е н. Кроме триплетности, генетический код наделен рядом других характерных свойств. Его кодоны не перекрываются, каждый кодон начинается с нового нуклеотида, и ни один нуклеотид не может прочитываться дважды. Любой кодон соответствует только одной аминокислоте.
К о д у н и в е р с а л е н. Генетическому коду свойственна универсальность для всех организмов на Земле. Одинаковые аминокислоты кодируются одними и теми же триплетами нуклеотидов у бактерий и слонов, водорослей и лягушек, черепах и лошадей, птиц и даже человека. Несколько отличаются (на 1-5 кодонов) только коды митохондрий некоторых организмов, ряда дрожжей и бактерий.
Ошибка хотя бы в одном триплете приводит к серьезным нарушениям в организме. У больных серповидной анемией (их эритроциты имеют не дисковую, а серповидную форму) из 574 аминокислот белка гемоглобина одна аминокислота заменена другой в двух местах. В результате белок имеет измененную третичную и четвертичную структуру. Нарушенная геометрия активного центра, присоединяющего кислород, не позволяет гемоглобину эффективно справляться со своей задачей — связывать кислород в легких и снабжать им клетки организма.
Нарушенная геометрия активного центра, присоединяющего кислород, не позволяет гемоглобину эффективно справляться со своей задачей — связывать кислород в легких и снабжать им клетки организма.
Транскрипция. Синтез белка происходит в цитоплазме на рибосомах. Генетическую информацию от хромосом ядра к месту синтеза переносят иРНК:
ДНК — иРНК — белок
Информационная РНК синтезируется на отрезке одной из нитей ДНК как на матрице, хранящей информацию о первичной структуре конкретного белка или группы белков, выполняющих одну функцию. В основе синтеза лежит принцип комплементарности: напротив Цднк встает Грнк, напротив Гднк — Црнк, напротив Аднк — Урнк, напротив Тднк — Арнк. Затем мономерные звенья связываются в полимерную цепь. Таким образом, иРНК становится точной копией второй нити ДНК (с учетом замены Т- У). Молекула иРНК имеет одноцепочечную структуру, она в сотни раз короче ДНК.
Процесс перенесения генетической информации на синтезируемую иРНК носит название транскрипции. Перед началом каждого гена или группы однофункциональных генов расположена последовательность нуклеотидов, называемая инициатором (содержит кодон АУГ). В этой последовательности есть участок (промотор) для присоединения фермента РНК-полимеразы, осуществляющего транскрипцию. Полимераза распознает промотор благодаря химическому сродству. В конце матрицы синтеза находится стоп-кодон (один из трех в таблице), или терминатор.
Перед началом каждого гена или группы однофункциональных генов расположена последовательность нуклеотидов, называемая инициатором (содержит кодон АУГ). В этой последовательности есть участок (промотор) для присоединения фермента РНК-полимеразы, осуществляющего транскрипцию. Полимераза распознает промотор благодаря химическому сродству. В конце матрицы синтеза находится стоп-кодон (один из трех в таблице), или терминатор.
В ходе транскрипции РНК-полимераза в комплексе с другими ферментами разрывает водородные связи между азотистыми основаниями двух нитей ДНК, частично раскручивает ДНК и производит синтез иРНК по принципу комплементарности. На одной ДНК «работают» сразу несколько полимераз.
Готовая молекула иРНК после небольшой перестройки связывается в комплекс со специальными белками и транспортируется ими через ядерную оболочку на рибосомы. Эти белки выполняют и другую функцию — они защищают иРНК от действия различных ферментов цитоплазмы. В прокариотической клетке ДНК не отделена от цитоплазмы, и синтез белков рибосомы начинают еще во время транскрипции.
Транспортные РНК. Необходимые для синтеза белков аминокислоты всегда имеются в составе цитоплазмы. Они образуются в процессе расщепления лизосомами белков. Транспортные РНК связывают аминокислоты, доставляют их на рибосомы и производят точную пространственную ориентацию аминокислот на рибосоме.
Рассмотрим устройство тРНК, позволяющее ей успешно выполнять свои сложные функции. В цепочке, состоящей из 70-90 звеньев, имеется 4 пары комплементарных отрезков из 4-7 нуклеотидов — А, Б, В и Г. Комплементарные участки связываются водородными связями попарно (как в молекуле ДНК). В результате нить тРНК «слипается» в четырех местах с образованием петлистой структуры, напоминающей лист клевера. В верхушке «листа» располагается триплет, код которого комплементарен кодону иРНК, соответствующему транспортируемой аминокислоте. Так, если в иРНК код аминокислоты валина ГУГ, то на вершине валиновой тРНК ему будет соответствовать триплет ЦАЦ. Комплементарный триплет в тРНК называют антикодоном.
Специальный фермент распознает антикодон тРНК, присоединяет к «черенку листа» определенную аминокислоту (в нашем примере — валин), и затем тРНК перемещает ее к рибосоме. Каждая тРНК транспортирует только свою аминокислоту.
Вопросы
1. Какая группа органических соединений определяет основные свойства организмов? Докажите.
2. Что такое генетический код? Перечислите его основные свойства.
3. Как происходит транскрипция? Какой принцип лежит в основе этого процесса? Каковы особенности протекания транскрипции у прокариот?
4. В чем состоит функция иРНК?
5. Опишите строение и функции тРНК.
Страница
1 — 1 из 2
Начало | Пред. |
1
2
|
След. |
Конец
| Все
© Все права защищены http://www.portal-slovo. ru
ru
Обмен веществ в клетке | Дистанционные уроки
25-Июл-2013 | комментариев 8 | Лолита Окольнова
Во всех клетках живых организмов непрерывно идут процессы обмена веществ и энергии.
Это называется метаболизм.
Если рассмотреть этот процесс более детально, то это постоянные процессы образования и распада веществ и поглощения и выделения энергии.
Процесс синтеза веществ = пластический обмен = ассимиляция = анаболизм
Пластический обмен (анаболизм, или ассимиляция) – это совокупность физиолого-биохимических процессов, в ходе которых из простых органических и неорганических веществ образуются более сложные вещества. Пластический обмен протекает с затратой высокоорганизованной энергии (например, в виде АТФ)
Чтобы что-то построить, надо затратить энергию — этот процесс идет с поглощением энергии.
∗
Глюконеогенез — это процесс синтеза глюкозы из неуглеводных соединений, например, из пирувата. Реакции глюконеогенеза у человека происходят в клетках печени, почек и эпителия тонкого кишечника.
Гликогеногенез — это процесс синтеза гликогена из глюкозы. Реакции гликогеногенеза осуществляются в клетках мышечной ткани и в клетках печени
Синтез жирных кислот осуществляется в цитоплазме жировой ткани
Синтез нуклеотидов осуществляется в цитоплазме всех активных клеток организма
Процесс расщепления = энергетический обмен = диссимиляция = катаболизм
Энергетический обмен (катаболизм, или диссимиляция) – это совокупность физиолого-биохимических процессов, в ходе которых происходит окисление сложных органических веществ. В результате энергетического обмена образуются более простые органические или неорганические вещества, и выделяется высокоорганизованная энергия (например, в виде АТФ) .

В основном, это реакции окисления, происходят они в митохондриях, самый простой пример — дыхание. При дыхании сложные органические вещества расщепляются до простых, выделяется углекислый газ и энергия.
Вообще, эти два процесса взаимосвязаны и переходят один в другой. Суммарно уравнение метаболизма — обмена веществ в клетке — можно записать так:
катаболизм + анаболизм = обмен веществ в клетке = метаболизм
Энергетический обмен = Диссимиляция = Катаболизм
Этот процесс идет в несколько этапов и нам нужно рассмотреть как он проходит а различных организмах.
Организмов будет всего 2 — многоклеточный (человек, например) и одноклеточный (растительный и животный).
И запомните, сочетание букв АТФ (аденинтрифосфорная кислота) — означает “энергию”.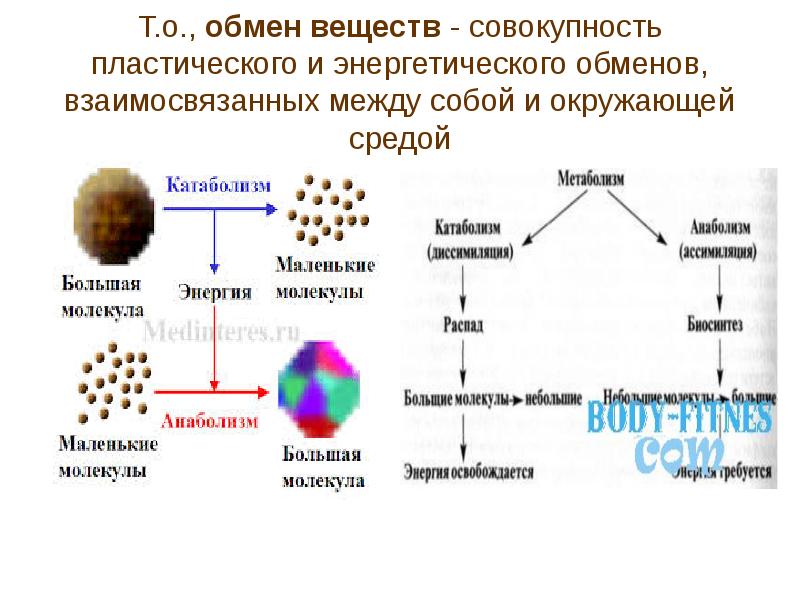 Просто эта энергия заключена в молекуле.
Просто эта энергия заключена в молекуле.
Обмен веществ в клетке
Этапы диссимиляции:
1 этап — подготовительный
Давайте проследим путь пищи от начала и до конца… Итак, пища поступила в организм. А что у нас за пища? Точнее, из чего она состоит? Из белков, жиров и углеводов.
Пища начинает перевариваться.
В чем суть пищеварения? Очень просто: полимеры: белки, жиры и углеводы расщепляются до мономеров:
- жиры → до глицерина и жирных кислот
- углеводы (полисахариды) → до моносахаридов
Такое расщепление возможно с помощью ферментов (био-катализаторов)
- у одноклеточных — в их “мини-желудочках” — лизосомах (пищеварительных вакуолях)
2 этап — бескислородный — гликолиз
Глюкоза, полученная в предыдущем этапе, превращается в пировиноградную кислоту (ПВК) и выделяется энергия (“+” — это выделение энергии, “-” — поглощение).
С6H12O6 → C3H4O3 + 2 АТФ
Происходит этот процесс уже в цитоплазме клеток (как много-, так и одноклеточных организмов).
3 этап — кислородный = Цикл Кребса + окислительное фосфорилирование
Здесь мы не будем детально разбирать цикл Кребса и фосфорилирование — это будет отдельная подробная тема в формате ЕГЭ…
Сама суть этого процесса в том, что в митохондриях (на кристах) ( а если митохондроий нет, то и процесс этот отсутствет, т.е. у анаэробов кислородного этапа нет) кислота превращается уже до конца: до CO2 (то, что мы выдыхаем) и H2O:
в цикле Кребса:
C3H4O3→CO2 + h3O + 36 АТФ
Общее уравнение диссимиляции:
С6H12O6 + 6O2 = 6CO2 + 6H2O + 38 АТФ
Взаимосвязь пластического и энергетического обмена:
- Пластический обмен обеспечивает клетку сложными органическими веществами (белками, жирами, углеводами, нуклеиновыми кислотами), в том числе белками-ферментами для энергетического обмена.

- Энергетический обмен обеспечивает клетку энергией. При выполнении работы (умственной, мышечной и т.п.) энергетический обмен усиливается.
Пластический и энергетический обмен – это сопряженные (взаимосвязанные) процессы.
Реакции метаболизма рано или поздно завершаются превращением всей исходной энергии в тепло.
Еще на эту тему:
Обсуждение: «Обмен веществ в клетке»
(Правила комментирования)
Урок 24. энергетика живой клетки — Естествознание — 10 класс
Конспект на интерактивный видео-урок
по предмету «Естествознание» для «10» класса
Урок № 24.Энергетика живой клетки
Перечень вопросов, рассматриваемых в теме:
- Как энергия запасается в клетке;
- Что такое метаболизм;
- В чем суть процессов гликолиза, брожения и клеточного дыхания;
- Какие процессы проходят на световой и темновой фазах фотосинтеза;
- Как связаны процессы энергетического и пластического обмена;
- Что представляет собой хемосинтез.

Глоссарий по теме:
Метаболизм (обмен веществ) — сложная цепь превращений веществ в организме начиная с момента их поступления из внешней среды и кончая удалением продуктов распада. Представляет собой совокупность процессов энергетического обмена (катаболизма диссимиляции) и пластического обмена (анаболизма, ассимиляции).
Энергетический обмен – это совокупность химических реакций постепенного распада органических соединений, сопровождающихся высвобождением энергии, часть которой расходуется на синтез АТФ. Синтезированная АТФ становится универсальным источником энергии для жизнедеятельности организмов. Значение энергетического обмена – снабжение клетки энергией, которая необходима для жизнедеятельности.
Пластический обмен – это совокупность химических реакций образования (синтеза) из простых веществ с затратой энергии более сложные. Непосредственным поставщиком энергии в клетках выступает АТФ.
Фотосинтез – процесс образования органических веществ из неорганических (углекислого газа и воды) с использованием солнечной энергии. Проходит в два этапа: световая фаза (происходит улавливание и фиксация энергии света в АТФ) и темновая (связывание углекислого газа в молекулы глюкозы с затратой энергии АТФ).
Проходит в два этапа: световая фаза (происходит улавливание и фиксация энергии света в АТФ) и темновая (связывание углекислого газа в молекулы глюкозы с затратой энергии АТФ).
Хемосинтез — процесс образования органических веществ из неорганических (углекислого газа и воды) с использованием энергии окисления неорганических веществ. Например, такой тип питания используют азотфиксирующие бактерии.
Клеточное или тканевое дыхание — совокупность биохимических реакций, протекающих в клетках живых организмов, в ходе которых происходит окисление углеводов, липидов и аминокислот до углекислого газа и воды.
Основная и дополнительная литература по теме урока :
Естествознание. 10 класс [Текст]: учебник для общеобразоват. организаций: базовый уровень / И.Ю. Алексашина, К.В. Галактионов, И.С. Дмитриев, А.В. Ляпцев и др. / под ред. И.Ю. Алексашиной. – 3-е изд., испр. – М.: Просвещение, 2017.: с 115 — 118.
Электронные ресурсы:
Обмен веществ. Портал открытая биология // Электронный доступ: https://biology.ru/textbook/content.html
Портал открытая биология // Электронный доступ: https://biology.ru/textbook/content.html
АТФ и её роль в клетке .Проект «вся биология» // Электронный доступ: http://www.sbio.info/materials/obbiology/obbkletka/stroenorg/12
Энергетика живой клетки. Научно-познавательный журнал «Познавайка» // Электронный доступ: http://www.poznavayka.org/biologiya/energiya-zhivoy-kletki/
Энергетика живой клетки. Журнал «В МИРЕ НАУКИ» №3, 2006 . БИОЛОГИЯ // электронный доступ: https://elementy.ru/nauchno-populyarnaya_biblioteka/430308/430310?SSL=1
Теоретический материал для самостоятельного изучения
Обязательным условием существования биологических систем являются потоки энергии. В этом заключается ключевое различие между живой и неживой природой. Энергия не хранится в клетке, а поступает извне. Ключевую роль в трансформации энергии обеспечивает клетка, как элементарная структура живого. Специальные биохимические механизмы трансформируют одни виды энергии в другие, для обеспечения необходимых функций клетки.
Основным источником энергии для всех живых существ планеты Земля, является энергия Солнца. Однако эта энергия может быть использована живым только после того, как она будет усвоена фотоавтотрофами (от греч. «фото» — свет, «авто» — сам, «трофос» — питание).
В процессе эволюции появились и другие организмы, которые научились потреблять готовые органические соединения для получения запасённой в них энергии – гетеротрофы (от греч. «гетерос» — другой, «трофос» — питание).
Некоторые виды микроорганизмов (хемоавтотрофы) приобрели способность к использованию энергии, выделяемой при окислении неорганических веществ.
Таким образом, из всего многообразия существующих форм энергии живые существа на нашей планете используют только две – световую и энергию химических связей.
Главный переносчик энергии в клетке
Световая энергия Солнца и энергия, заключённая в потребляемой пище, запасаются в особых бимолекулярных аккумуляторах – молекулах АТФ (аденозинтрифосфат). В молекулах АТФ энергия запасается в виде высокоэнергетических химических связях между остатками фосфорной кислоты, которая освобождается при отщеплении фосфата: АТФ → АДФ + Ф + E.
В молекулах АТФ энергия запасается в виде высокоэнергетических химических связях между остатками фосфорной кислоты, которая освобождается при отщеплении фосфата: АТФ → АДФ + Ф + E.
Выделяемая энергия используется клетками для процессов выработки тепла, мышечных сокращений (мышечная клетка), для проведения нервного импульса (нервные клетки) и т.п.
Обратный процесс образования АТФ с затратой энергии, получил название энергетический обмен.
Синтез макромолекул важнейших органических соединений, необходимых для построения структур клетки, обеспечения всех процессов жизнедеятельности клеток – пластический обмен — обеспечивается также энергией АТФ.
Независимо от типа питания, универсальным аккумулятором энергии живых организмов выступают молекулы АТФ, где добытая энергия извне запасается в виде химических связей. Такая схожесть иллюстрирует единство происхождения всего живого.
Метаболизм
Поступившие вместе с пищей (или в результате фотосинтеза) органические вещества расщепляются на более простые (катаболизм или диссимиляция), которые служат для постройки макромолекул органических соединений (анаболизм или ассимиляция). Эти процессы происходят в организме одновременно. Совокупность этих процессов получила название – метаболизм. В результате его организм осуществляет обмен веществом и энергией с окружающей средой. Наибольшее значение для энергетического обмена являются многостадийные реакции расщепления глюкозы.
Эти процессы происходят в организме одновременно. Совокупность этих процессов получила название – метаболизм. В результате его организм осуществляет обмен веществом и энергией с окружающей средой. Наибольшее значение для энергетического обмена являются многостадийные реакции расщепления глюкозы.
На стадии гликолиза в цитоплазме клетки происходит ферментативное расщепление молекулы глюкозы с образованием более простой пировиноградной кислоты и молекул АТФ: С6Н12О6 + 2 АДФ + 2 Ф → 2С3Н4О3 + 4Н+ + 2АТФ
Молекулы пировиноградной кислоты обладают значительной энергией, высвобождение которой происходит в митохондриях. В ходе так называемого клеточного дыхания (аэробного расщепления), вещество распадается на углекислый газ, который впоследствии выделяется из клетки и воду. По последним исследованиям, при этом образуется 30 молекул АТФ.
Суммарную реакцию окисления глюкозы можно представить следующим образом:
С6Н12О6 + 6О2 + 6Н2О + 32 АДФ + 32 Ф → 6 СО2 + 12 Н2О + 32АТФ
Некоторые микроорганизмы при недостатке кислорода расщепляют глюкозу в процессе анаэробного дыхания или брожения. В зависимости от конечных продуктов такого расцепления различают спиртовое брожение (с образование этанола), молочнокислое (молочная кислота). Последнее происходит и в мышцах, при недостатке кислорода, например во время длительной тренировки. Энергетический выход такого типа расщепления менее энергоэффективен.
В зависимости от конечных продуктов такого расцепления различают спиртовое брожение (с образование этанола), молочнокислое (молочная кислота). Последнее происходит и в мышцах, при недостатке кислорода, например во время длительной тренировки. Энергетический выход такого типа расщепления менее энергоэффективен.
Основным источником энергии для организмов является окисление глюкозы в митохондриях. При этом также может происходить окисление других органических соединений (белков, жиров), потребляемых, например, вместе с пищей.
Фотосинтез
Фотоавтотрофы имеют уникальные ферментативные системы, способные трансформировать энергию солнечного света в энергию химической связи. Процесс образования органических веществ из неорганических (углекислого газа и воды) с использованием солнечной энергии получил название фотосинтез. В растениях фотосинтезирующие комплексы сосредоточены в специальных органеллах – хлоропластах. Основной пигмент – хлорофилл – выполняет функцию световых «антенн», улавливая световые волны практически всех диапазонов, кроме зелёного. Стоит отметить, что это обуславливает окраску листьев растений.
Стоит отметить, что это обуславливает окраску листьев растений.
В так называемой, световой фазе, кванты света выбивают электроны из молекулы хлорофилла, и он начинает передаваться по специальным белковым переносчикам, расположенных на мембране хлоропластов. Под действием света одновременно происходит разложение воды (фотолиз). В реакции высвобождается, в том числе катион водорода (Н+), необходимый для последующего биосинтеза, который захватывает молекула НАДФ (никотинамидадениндинуклеотидфосфат): НАДФ+ + Н+→НАДФ∙Н
Энергия возбуждённого электрона заряжает известный нам биологический катализатор АТФ и молекулу НАДФ – в этом заключается биологический смысл световой фазы фотосинтеза.
Заметим, что побочными продуктами фотолиза воды становятся свободный кислород и свободные электроны, восстанавливающие хлорофилл: 2Н2О→ Н+ + 4е— + О2
Дальнейший процесс может уже проходить без света. Сущность реакций темновой фазы можно выразить следующим уравнением: СО2 + НАДФ∙Н + АТФ = С6Н12О6 +АДФ + НАДФ+
Не сложно заметить, что выделяются вещества необходимые на начальном этапе фотосинтеза, что замыкает цикл. Энергия молекулярных аккумуляторов была использована для фиксации углекислого газа в энергию химических связей углевода.
Энергия молекулярных аккумуляторов была использована для фиксации углекислого газа в энергию химических связей углевода.
Фотосинтез, таким образом, является процессом превращения одной (световой) формы энергии в другую(химическую). Вся энергия биосферы запускается благодаря этому процессу. Другими словами, фотосинтез является отражением космических потоков энергии. Помимо этого, фотосинтезирующие организмы не только обеспечивают первичный синтез органических соединений, но и создают условия необходимые для существования других живых организмов.
Взаимосвязь энергетического и пластического обмена
Не сложно заметить, что процессы аккумулирования энергии в молекулах АТФ (энергетический обмен) и использование запасённой энергии для синтеза необходимых веществ (белков, жиров, углеводов, нуклеиновых кислот) неразрывно связаны. Так синтез АТФ не возможен без разложения органических веществ, а синтез веществ клетки не возможен без энергии АТФ. Причём, заметим, что и фотосинтез представляет собой единство этих процессов: темновая фаза – пластический обмен, световая фаза – энергетический.
Оба процесса протекают одновременно и неотделимы друг от друга, обеспечивая жизнедеятельность организма. Таким образом, в клетках происходит трансформация вещества и энергии, которые лежат в основе существования жизни и непрерывного самообновления. Сходство процессов энергетического обмена в клетках всех живых организмов является доказательством единства их происхождения.
Вывод
В клетках происходят одновременно процессы энергетического и пластического обмена, это лежит в основе сохранения жизни. Взаимообмен энергией и веществом между живой и неживой природой является иллюстрацией принципа единства и взаимосвязи материального мира.
Примеры и разбор решения заданий тренировочного модуля:
Задание 1. Выберите один ответ:
- Универсальным аккумулятором энергии в клетке является:
- Жиры;
- Белки;
- АТФ;
- НАДФ∙Н.
Ответ: АТФ
Пояснение: универсальной «разменной валютой» в энергетике живой клетки выступает АТФ.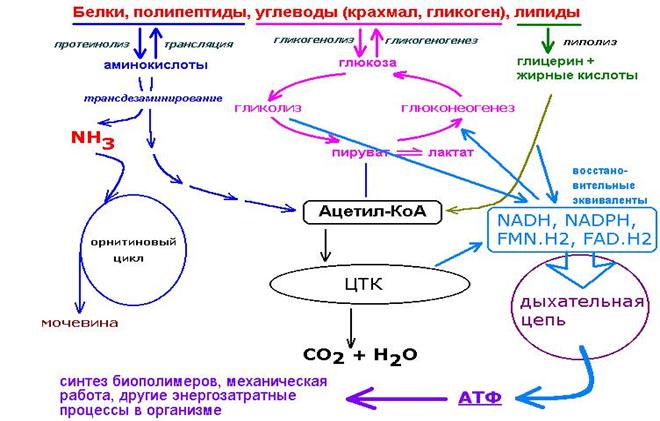 При его распаде выделяется энергия, которая расходуется на все жизненно важные процессы.
При его распаде выделяется энергия, которая расходуется на все жизненно важные процессы.
Задание 2. Исправьте ошибки, анализируя текст с позиции энергетического обмена:
В рационе питания человека помимо белков растительных и животных не обязательно должны присутствовать углеводы и жиры. Отсутствие жиров в пище не приводит к истощению. Человек толстеет, если употребляет в пищу избыточное количество углеводов. На сое и рисе можно прожить.
Ответ: В рационе питания человека помимо белков растительных и животных не обязательно должны присутствовать углеводы и жиры. Отсутствие жиров в пище не приводит к истощению. Человек толстеет, если употребляет в пищу много жиров. Исключительно на сое и рисе можно благополучно прожить.
Пояснение: с точки зрения энергетического обмена, наиболее энергоэффективными являются жиры. При этом, жиры, поступающие с пищей, используются в том числе, для построения многих важных соединений, например гормонов. «Быстрая» энергия углеводов в избыточном количестве может приводить к полноте. Употребление только растительного белка в пищу, по сравнению с животным, является менее энергоэффективным и при отсутствии других источников энергии может приводить к истощению организма.
«Быстрая» энергия углеводов в избыточном количестве может приводить к полноте. Употребление только растительного белка в пищу, по сравнению с животным, является менее энергоэффективным и при отсутствии других источников энергии может приводить к истощению организма.
Происхождение метаболизма установили, не воссоздавая живые клетки
Исследователи из Кембриджского университета (University of Cambridge) разобрались в том, как именно зародились метаболические процессы — каскады реакций, протекающие во всех клетках и обеспечивающие их необходимыми для выживания веществами.
Обмен веществ — это процесс, необходимый для любой жизнедеятельности, и потому знания о его происхождении должны помочь по-новому взглянуть на то, как зародилась первая жизнь на нашей планете. Как выяснилось, для первичного запуска метаболизма потребовалось не так много «ингредиентов».
«Многие считают, что это был очень сложный процесс, — говорит руководитель исследования Маркус Релсер (Markus Ralser). — Но наши результаты показывают, что многие из этих реакций произошли спонтанно в первичном бульоне Земли, благодаря лишь ионам металлов, а не ферментам, которые управляют обменными процессами сегодня. Если вы обратите внимание на метаболизм самых разных организмов со всего мира, то увидите, что цепь реакций выглядит почти одинаково. Должно быть, обмен веществ начался в очень ранней эволюции, но никто точно не знает, когда и как это произошло».
— Но наши результаты показывают, что многие из этих реакций произошли спонтанно в первичном бульоне Земли, благодаря лишь ионам металлов, а не ферментам, которые управляют обменными процессами сегодня. Если вы обратите внимание на метаболизм самых разных организмов со всего мира, то увидите, что цепь реакций выглядит почти одинаково. Должно быть, обмен веществ начался в очень ранней эволюции, но никто точно не знает, когда и как это произошло».
Одна из теорий гласит, что первым строительным блоком жизни была РНК, так как она помогает производить ферменты, способные катализировать сложные последовательности реакций. Также существует версия, что сначала появился обмен веществ, и именно он сгенерировал молекулы, необходимые для создания РНК. Последняя гипотеза не получила веских доказательств. Эксперимент учёных из Кембриджа впервые показал, что обменные процессы можно запустить даже в отсутствие РНК.
Открытие произошло случайно во время обычного тестирования контроля качества среды, используемой для культивирования клеток в лаборатории Релсера. Один из его студентов рассмотрел неиспользованные материалы через масс-спектрометр и обнаружил подписи пирувата — конечного продукта метаболического пути под названием гликолиз.
Один из его студентов рассмотрел неиспользованные материалы через масс-спектрометр и обнаружил подписи пирувата — конечного продукта метаболического пути под названием гликолиз.
Чтобы проверить, могли ли те же самые процессы породить жизнь на Земле, исследователи пригласили своих коллег, работавших над реконструкцией химии архейского океана, который покрывал планету почти четыре миллиарда лет назад. Это был бескислородный мир из горячей воды богатой железом и другими металлами и фосфатами.
Именно атомы и ионы железа сделали возможными многие ключевые реакции, протекающие сегодня в живых клетках с участием десятков ферментов.
Команда Релсера добавила к полученной субстанции вещества, которые являются отправными точками современных метаболических путей, а затем в течение 5 часов нагревала образцы до 50-70 градусов по Цельсию (подобная температура характерна для гидротермальных жерл). После был проанализирован молекулярный состав образцов.
«Сначала мы надеялись обнаружить одну-две реакции, но результаты оказались просто поразительными, — говорит Релсер. — Мы смогли реконструировать пару метаболических путей почти полностью».
— Мы смогли реконструировать пару метаболических путей почти полностью».
Исследователи реконструировали гликолиз и пентозофосфатный путь — реакции, которые составляют «ядро» метаболизма каждой живой клетки. Вместе эти пути производят одни из самых важных материалов в современных клетках, в том числе аденозинтрифосфат — нуклеотид, который играет исключительно важную роль в обмене энергии и веществ в организмах и является универсальным источником энергии для всех биохимических процессов, протекающих в живых системах. Клетки используют АТФ для формирования ДНК и РНК, а также молекул, необходимых для создания жиров и белков.
Как оказалось, в целом, без использования каких-либо ферментов, с одними лишь микроэлементами, можно выполнить 29 разных биохимических реакций, включая и ту, при которой образуется рибозо-5-фосфат, образующий цепь РНК. Метаболические пути не идентичны современным, но образуют множество таких же молекул: видимо, с появлением в клетках ферментов пути совершенствовались и улучшались.
Эти исследования показывают, что ключ к образованию жизни на планете может быть гораздо проще, чем казалось ранее, и жизнь на самом деле более распространена в нашей Солнечной системе и за её пределами. Подробности исследования были опубликованы в статье издания Molecular Systems Biology.
Также по теме: Объяснено происхождение механизма клеточного деления В девонских рифах обнаружены молекулы древнейшего жира Ископаемые существа донесли до исследователей старейшие биомолекулы Столкновение с метеоритом увековечило жизнь в стекле «Зловонный флюорит» оказался единственным убежищем для природного элементарного фтора
Метаболизм
Здравствуйте, уважаемые читатели блога репетитора биологии по Скайпу biorepet-ufa.ru.
Тема метаболизма или обмена веществ в клетке — это, можно сказать, «краеугольный камень» в понимании самих основ жизненных процессов, в описании того, что лежит вообще в основе жизни.
Мне, как репетитору биологии по Скайпу, приходится констатировать, что понимание проблемы клеточного метаболизма вызывает определенные трудности у многих.
Буквально на одной страничке этого поста с помощью наглядной схемы как «живет жизнь» попытаемся разобраться
В чем возможные причины плохого усвоения данной темы
На мой взгляд, вопросы обмена веществ в клетке или клеточный метаболизм вызывают затруднения в понимании сути процессов не потому, что это что-то сверхсложное, а скорее всего из-за:
1) обилия синонимической терминологии, которая буквально затмевает сознание;
2) неправильного двойственного понимания описываемого явления и
3) отсутствия наглядного схематичного представления метаболизма у организмов с разным типом питания (автотрофы, гетеротрофы).
Итак, по первому вопросу
Как репетитору по биологии, мне давно стало понятно, что если все термины объединить в какую-то систему в одном месте, то все быстро становится на свои места. Через пару минут и вам тема метаболизма уже не будет казаться свалкой терминов.
Через пару минут и вам тема метаболизма уже не будет казаться свалкой терминов.
Если у термина «обмен веществ» всего один синоним — «метаболизм», то названия ВСЕГО ДВУХ разнонаправленных процессов, из которых складывается обмен веществ в клетке, включают по три наименования!
Синонимами термину пластический обмен (или СИНТЕЗ органических веществ) являются еще термины ассимиляция и анаболизм.
Синонимами термину энергетический обмен (заключающемуся в РАСПАДЕ органических веществ с высвобождением энергии), являются еще и такие термины, как диссимиляция и катаболизм. Эти термины для удобства запоминания в табличке ниже расположены напротив друг друга. По второму вопросу
В чем проявляется двойственность в определении описываемых явлений? Да в том, что в учебных руководствах описанию процессов синтеза органических веществ часто сопутствует такая фраза: «при пластическом обмене идет запасание энергии«.
Как же так, выше я привожу табличку для лучшего запоминания, что энергия высвобождается лишь в процессах распада органических веществ. На создание новых органических веществ она тратится. А тут говорится, что и при пластическом обмене она запасается. И как это понимать?
А так: надо иметь четкое разграничение между понятиями универсального, реального на любой момент времени жизни клетки источника энергии, каковыми являются молекулы АТФ и потенциально возможными источниками извлечения энергии из органических веществ клетки.
Итак, важно помнить, что универсальный перманентный источник энергии в любых клетках, любых живых систем — аденозинтрифосфорная кислота — АТФ, в макроэнергетических связях которой временно сосредотачивается большое количество энергии. Откуда берется эта энергия? В результате энергетического обмена при распаде потенциально богатых энергией органических веществ.
Эта энергия макроэнергетических связей АТФ тратится на осуществление процессов синтеза, поскольку создаются богатые энергией органические вещества в пластическом обмене.
Любой многоклеточный организм существует пока в его клетках слаженно идут процессы расщепления поступивших веществ для выработки энергии и синтеза необходимых организму собственных веществ.
Третий момент — обещанная наглядность: схема как «живет жизнь»
Что здесь главное
* В основании жизни «лежат» совершенно особенные, присущие только живым системам органические вещества: крупные молекулы жиров (липидов), моно- и полисахариды (углеводы), и крупные гетерополимерные молекулы нуклеиновых кислот и белков, определяющие все разнообразие жизни на Земле.
* Существует всего два пути создания органических веществ в мире живого:
у автотрофов — путем автотрофной ассимиляции из неорганических веществ СО2, Н2О, NH3 за счет энергии солнечного света (у фототрофов) или энергии окисления неорганических веществ (у хемотрофов); Почему на схеме стрелка «автотрофная ассимиляция» имеет два цвета? Потому что у автотрофных организмов внешние источники энергии (красный цвет) и источники для создания органических веществ (желтый цвет) — разные.
у гетеротрофов — путем гетеротрофной ассимиляции чужеродных органических веществ за счет энергии окисления этих же веществ. Поэтому на схеме блок перевернутая трапеция «чужеродные органические вещества» — двухцветная. Это отражает то, что для гетеротрофных организмов источник энергии для жизни и источник создания собственных органических веществ один и тот же (чужеродные органические вещества).
* Само по себе существование такого сложного состояния материи как «жизнь» требует затраты энергии двоякого рода: а) для поддержания организма — энергии поддержания и б) для расширенного воспроизведения живой материи — функциональной энергии.
* Всю эту энергию в клетках поставляют высокоэнергетические соединения, прежде всего АТФ, образующиеся в процессах диссимиляции (расщепления) собственных органических веществ (у любых организмов) или поступивших извне (как у гетеротрофов).
____________________________________________
Ну как? Совсем ведь не сложно, видя эту схемку перед глазами, запомнить, что материальной основой жизни, её фундаментом, являются органические вещества.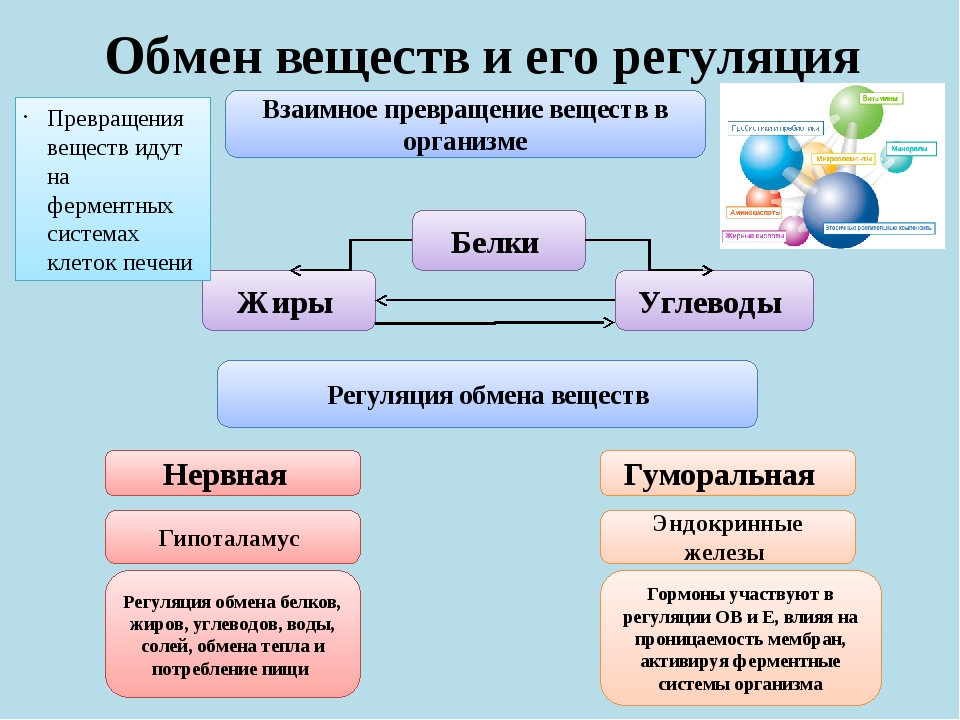
А энергию: одни организмов получают только за счет окисления органических веществ (гетеротрофы), а другие — способны еще использовать световую энергию (фототрофы) или энергию окисления неорганических веществ (хемотрофы).
По теме пластического обмена необходимо хорошо разбираться в процессах синтеза углеводов растениями при фотосинтезе и в биосинтезе белка.
Энергетическому обмену бескислородному у анаэробных организмов и кислородному (в митохондриях) у аэробных организмов посвящена следующая статья.
***************************************
У кого есть вопросы по статье к репетитору биологии по Скайпу, замечания, пожелания — прошу в комментарии.
Cell Metabolism — обзор
5.1 Внутренняя регуляция функции стволовых клеток
Клеточный метаболизм и воздействие окружающей среды могут привести к повреждению ДНК, особенно в коже, которая напрямую получает УФ-излучение от солнца и мутагенов окружающей среды, которые могут вызывать геномную нестабильность.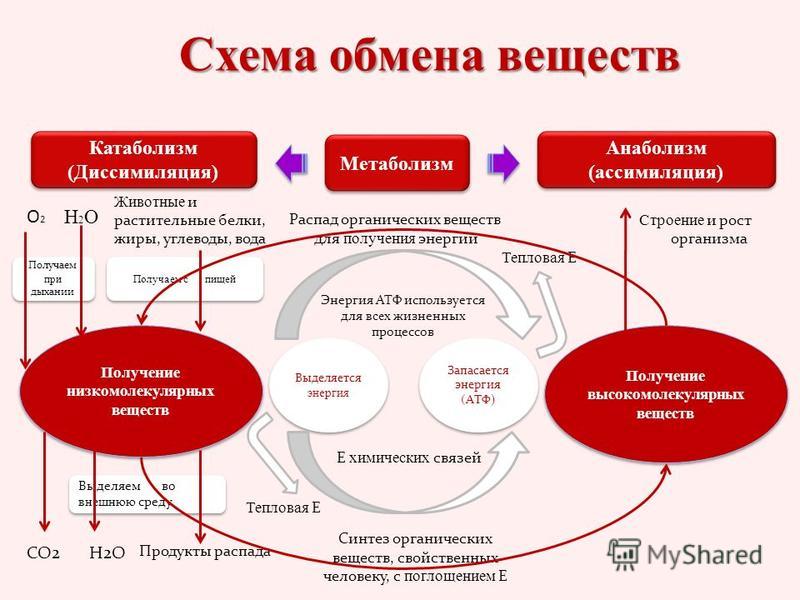 Интересно, что HFSC более устойчивы к радиационному повреждению по сравнению с другими эпителиальными клетками кожи (Sotiropoulou et al., 2010). Для достижения этой устойчивости HFSC экспрессируют высокие уровни антиапоптотического белка B-клеточной лимфомы 2 (Bcl2) и временно экспрессируют p53, что способствует выживанию.Кроме того, рак груди 1 (Brca1) необходим для восстановления повреждений ДНК (Gudmundsdottir & Ashworth, 2006; Moynahan & Jasin, 2010), а эпидермальная делеция Brca1 приводит к дефектам образования HF, а также к индукции каспазозависимого апоптоза, который приводит к к гиперпролиферации и последующему истощению взрослых SCs (Sotiropoulou et al., 2013). Дифференциальная регуляция повреждений ДНК в SCs также существует в других тканях (Mandal, Blanpain, & Rossi, 2011) и может быть сходной с механизмами, которые действуют в SCs в IFE и др. Эпидермальных придатках.
Интересно, что HFSC более устойчивы к радиационному повреждению по сравнению с другими эпителиальными клетками кожи (Sotiropoulou et al., 2010). Для достижения этой устойчивости HFSC экспрессируют высокие уровни антиапоптотического белка B-клеточной лимфомы 2 (Bcl2) и временно экспрессируют p53, что способствует выживанию.Кроме того, рак груди 1 (Brca1) необходим для восстановления повреждений ДНК (Gudmundsdottir & Ashworth, 2006; Moynahan & Jasin, 2010), а эпидермальная делеция Brca1 приводит к дефектам образования HF, а также к индукции каспазозависимого апоптоза, который приводит к к гиперпролиферации и последующему истощению взрослых SCs (Sotiropoulou et al., 2013). Дифференциальная регуляция повреждений ДНК в SCs также существует в других тканях (Mandal, Blanpain, & Rossi, 2011) и может быть сходной с механизмами, которые действуют в SCs в IFE и др. Эпидермальных придатках.
Было идентифицировано несколько транскрипционных регуляторов функции SC в HF, которые являются общими для SC других тканей, включая фактор транскрипции 3 и 4 (TCF3 / 4), ядерный фактор активированных Т-клеток 1 (NFATc1) и область Y, определяющую пол. -box 9 (Sox9) (Blanpain & Fuchs, 2006; Nguyen et al., 2009; Nguyen, Rendl, & Fuchs, 2006; Nowak, Polak, Pasolli, & Fuchs, 2008). Кроме того, Lgr5 (Barker et al., 2007) и атипичный гомеобоксный белок HOP Hopx экспрессируются с помощью кишечного SC эпителиального пула в основании крипты (Takeda, Jain, LeBoeuf, Wang, & Lu, 2011).В HF Hopx экспрессируется внутри клеток bulge и может вносить вклад во все клоны HF при росте HF, а также в клетки IFE при ранении (Takeda, Jain, LeBoeuf, & Padmanabhan, 2013). Клетки нижнего балджа, экспрессирующие SC маркер Lgr5, также экспрессируют Hopx, способны избегать апоптоза во время фазы смерти HF и вносить вклад в долгосрочное поддержание клеток балджа (Takeda et al., 2013).
-box 9 (Sox9) (Blanpain & Fuchs, 2006; Nguyen et al., 2009; Nguyen, Rendl, & Fuchs, 2006; Nowak, Polak, Pasolli, & Fuchs, 2008). Кроме того, Lgr5 (Barker et al., 2007) и атипичный гомеобоксный белок HOP Hopx экспрессируются с помощью кишечного SC эпителиального пула в основании крипты (Takeda, Jain, LeBoeuf, Wang, & Lu, 2011).В HF Hopx экспрессируется внутри клеток bulge и может вносить вклад во все клоны HF при росте HF, а также в клетки IFE при ранении (Takeda, Jain, LeBoeuf, & Padmanabhan, 2013). Клетки нижнего балджа, экспрессирующие SC маркер Lgr5, также экспрессируют Hopx, способны избегать апоптоза во время фазы смерти HF и вносить вклад в долгосрочное поддержание клеток балджа (Takeda et al., 2013).
Фактор транскрипции LIM гомеобокс-белок 2 (Lhx2) — еще один гомеобоксный белок, который участвует в регуляции морфогенеза и формирования паттерна эктодермальных производных, а также в поддержании и покое SC в нише HF SC (Mardaryev et al., 2011; Ри, Полак и Фукс, 2006 г .; Торнквист, Сандберг, Хэгглунд и Карлссон, 2010 г. ). Lhx2 экспрессируется в выпуклости и вторичном зачатке волоса, где он локализуется совместно с SC маркерами Sox9, Tcf4 и Lgr5. В ответ на повреждение кожи клетки Lhx2 + внутри выпуклости и вторичного волосяного зародыша пролиферируют и способствуют реэпителизации кожи за счет положительной регуляции Sox9 и Tcf4 , в то же время подавляя цикл HF за счет отрицательной регуляции Lgr5 ( Mardaryev et al. al., 2011). Эти и многие другие исследования предоставили новое понимание того, как пути передачи сигналов Wnt и BMP и сети регуляции транскрипции модулируют активность эпителиальных СК во время нормального гомеостаза и в ответ на повреждение (Blanpain & Fuchs, 2006; Lee & Tumbar, 2012; Sennett & Rendl, 2012).
). Lhx2 экспрессируется в выпуклости и вторичном зачатке волоса, где он локализуется совместно с SC маркерами Sox9, Tcf4 и Lgr5. В ответ на повреждение кожи клетки Lhx2 + внутри выпуклости и вторичного волосяного зародыша пролиферируют и способствуют реэпителизации кожи за счет положительной регуляции Sox9 и Tcf4 , в то же время подавляя цикл HF за счет отрицательной регуляции Lgr5 ( Mardaryev et al. al., 2011). Эти и многие другие исследования предоставили новое понимание того, как пути передачи сигналов Wnt и BMP и сети регуляции транскрипции модулируют активность эпителиальных СК во время нормального гомеостаза и в ответ на повреждение (Blanpain & Fuchs, 2006; Lee & Tumbar, 2012; Sennett & Rendl, 2012).
Эпителиальные SCs также регулируются посттранскрипционно и частично трансляционно с помощью микроРНК (miRNAs), которые представляют собой небольшие некодирующие РНК, которые изменяют трансляцию или стабильность РНК для контроля экспрессии генов.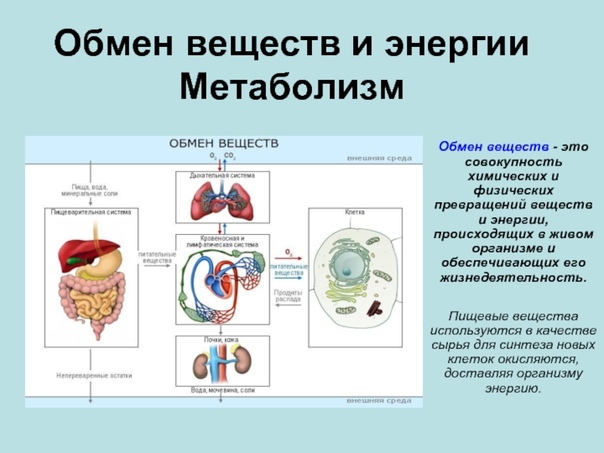 Полное устранение продукции miRNA путем делеции вышестоящего процессирующего фермента Dicer у мышей приводит к перинатальной летальности и тяжелым дефектам HF (Andl et al., 2006; Yi et al., 2006). К этим дефектам относятся неразвитые и смещенные HFs, повышенный апоптоз и отсутствие клеток K15 + и CD34 + в компартменте выпуклости, указывая тем самым, что miRNAs, в целом, важны для поддержания HF SC (Andl et al., 2006).
Полное устранение продукции miRNA путем делеции вышестоящего процессирующего фермента Dicer у мышей приводит к перинатальной летальности и тяжелым дефектам HF (Andl et al., 2006; Yi et al., 2006). К этим дефектам относятся неразвитые и смещенные HFs, повышенный апоптоз и отсутствие клеток K15 + и CD34 + в компартменте выпуклости, указывая тем самым, что miRNAs, в целом, важны для поддержания HF SC (Andl et al., 2006).
Несколько miRNAs пространственно-временно регулируются внутри IFE и HFSCs.Было показано, что MiR203 преимущественно обогащен IFE по сравнению с HF (Andl et al., 2006; Yi, Poy, Stoffel, & Fuchs, 2008) и достаточен для стимулирования дифференциации IFE и подавления самообновления в IFE путем контроля экспрессия p63 (Andl et al., 2006; Yi et al., 2008). Кроме того, miR203 транскрипционно активируется во время асимметричного клеточного деления в развивающемся эпидермисе, локализуясь в дифференцированной дочерней клетке, где он способствует выходу из клеточного цикла и устраняет собственное -обновление в процессе, включающем совместную супрессию p63, белка 2, связанного с S-фазой киназы (Skp2), и белка, связывающего РНК musashi 2 (Msi2) (Jackson et al. , 2013).
, 2013).
Дополнительная miRNA, miR125b, достаточна для изменения гомеостаза IFE и отмены спецификации волос (Zhang, Stokes, Polak, & Fuchs, 2011). MiR31 также может изменять активность HFSC, воздействуя на фактор роста фибробластов 10 (Fgf10), без дистального гомеобокса. 3 (Dlx3), несколько генов кератина, а также компоненты сигнальных путей Wnt и BMP (Mardaryev et al., 2010). Дифференциальная регуляция нескольких miRNAs в эпителии кожи предполагает, что роли дополнительных miRNAs будут определяться, поскольку это растущее поле продолжает расширяться.
Другой уровень регуляции SCs кожи происходит посредством модификации гистонов и ДНК для эпигенетической регуляции транскрипции (Calo & Wysocka, 2013). Несколько эпигенетических факторов играют роль в дифференцировке эпидермиса (Mulder et al., 2012). Ацетилирование и метилирование гистонов посредством активности гистондеацетилазы и метилтрансферазы, соответственно, регулируют развитие IFE (Driskell et al., 2012; LeBoeuf et al. , 2010) и гомеостаз (Driskell et al., 2012). Поддержание репрессивных модификаций гистонов через репрессорный комплекс polycomb, энхансер гомолога 1 zeste (Ezh2) и Ezh3 необходимы для дифференцировки IFE, а также для морфогенеза и поддержания HF (Bardot et al., 2013; Ежкова и др., 2011). Клеткам Меркеля также необходимы белки Ezh3 для своего поддержания посредством регуляции фактора транскрипции Sox2 (Bardot et al., 2013). Метилирование гистонов, контролируемое доменом деметилазы Jumonji, содержащим 3 (JmjD3), необходимо для дифференцировки IFE (Sen, Webster, Barragan, Chang, & Khavari, 2008), в то время как белок 2, содержащий домен деметилазы Jumonji / jmjc (Jarid2), необходим для поддерживать базальных предшественников IFE (Mejetta et al., 2011). Кроме того, ДНК-метилтрансфераза 1 (DNMT1) и убиквитин, содержащий PHD и домен-1 пальца RING (UHRF1), экспрессируются в базальных клетках и подавляются, как только клетки входят в программу дифференцировки, что позволяет предположить, что они также участвуют в регуляции стволовости.
, 2010) и гомеостаз (Driskell et al., 2012). Поддержание репрессивных модификаций гистонов через репрессорный комплекс polycomb, энхансер гомолога 1 zeste (Ezh2) и Ezh3 необходимы для дифференцировки IFE, а также для морфогенеза и поддержания HF (Bardot et al., 2013; Ежкова и др., 2011). Клеткам Меркеля также необходимы белки Ezh3 для своего поддержания посредством регуляции фактора транскрипции Sox2 (Bardot et al., 2013). Метилирование гистонов, контролируемое доменом деметилазы Jumonji, содержащим 3 (JmjD3), необходимо для дифференцировки IFE (Sen, Webster, Barragan, Chang, & Khavari, 2008), в то время как белок 2, содержащий домен деметилазы Jumonji / jmjc (Jarid2), необходим для поддерживать базальных предшественников IFE (Mejetta et al., 2011). Кроме того, ДНК-метилтрансфераза 1 (DNMT1) и убиквитин, содержащий PHD и домен-1 пальца RING (UHRF1), экспрессируются в базальных клетках и подавляются, как только клетки входят в программу дифференцировки, что позволяет предположить, что они также участвуют в регуляции стволовости. Удаление DNMT1 в анализах регенерации кожи человека вызывало преждевременную дифференцировку предшественников и прогрессирующую потерю ткани, что дополнительно демонстрирует его важность для самообновления (Sen, Reuter, Webster, Zhu, & Khavari, 2010).
Удаление DNMT1 в анализах регенерации кожи человека вызывало преждевременную дифференцировку предшественников и прогрессирующую потерю ткани, что дополнительно демонстрирует его важность для самообновления (Sen, Reuter, Webster, Zhu, & Khavari, 2010).
Дополнительный контроль функции SC происходит посредством регуляции экспрессии генов путем изменения положения нуклеосом за счет действия комплексов ремоделирования хроматина, таких как комплекс SWI / SNF (Kidder, Palmer, & Knott, 2009). Перестраивая положения нуклеосом в хроматине, эти комплексы регулируют занятость РНК-полимеразы II и, таким образом, инициацию транскрипции АТФ-зависимым образом (Liu, Balliano, & Hayes, 2011).В основе этих комплексов ген 1, связанный с brahma (Brg1), действует как каталитическая субъединица и регулирует пролиферацию и дифференцировку SC. В HF недавно было показано, что Brg1 динамически активируется после активации SC в коже. Делеция Brg1 с помощью bulge-специфического NFATc1-Cre индуцирует преждевременную регрессию HF, потерю HFSCs и прогрессирующую потерю волос (Xiong et al. , 2013). Молекулярно Brg1 и Shh действуют в молекулярной петле, где Brg1 регулирует экспрессию Shh, а Shh активирует экспрессию Brg1 в фолликуле (Xiong et al., 2013). Регулирует ли Brg1 дополнительные гены, чтобы контролировать функцию HFSC, будет интересной областью будущих исследований.
, 2013). Молекулярно Brg1 и Shh действуют в молекулярной петле, где Brg1 регулирует экспрессию Shh, а Shh активирует экспрессию Brg1 в фолликуле (Xiong et al., 2013). Регулирует ли Brg1 дополнительные гены, чтобы контролировать функцию HFSC, будет интересной областью будущих исследований.
Клеточный метаболизм: определение, процесс и роль ATP
Клеткам требуется энергия для движения, деления, размножения и других процессов. Они проводят большую часть своей жизни, сосредоточившись на получении и использовании этой энергии посредством метаболизма.
Клеточный метаболизм
Клеточный метаболизм — это серия процессов, которые происходят в живых организмах, чтобы поддерживать эти организмы.
В клеточной биологии и молекулярной биологии метаболизм относится к биохимическим реакциям, которые происходят внутри организмов с целью производства энергии. Разговорное или пищевое использование метаболизма относится к химическим процессам, , которые происходят в вашем теле, когда вы превращаете пищу в энергию.
Хотя термины похожи, есть и различия. Метаболизм важен для клеток, потому что эти процессы поддерживают жизнь организмов и позволяют им расти, воспроизводиться или делиться.
Что такое процесс клеточного метаболизма?
Основными этапами клеточного дыхания у эукариот являются:
- Гликолиз
- Окисление пирувата
- Лимонная кислота или цикл Кребса
- Окислительное фосфорилирование
Основными реагентами являются глюкоза и кислород, а основными продуктами — углекислый газ. вода и АТФ. Фотосинтез в клетках — это еще один тип метаболического пути, который организмы используют для производства сахара.
Растения, водоросли и цианобактерии используют фотосинтез.Основными этапами являются светозависимые реакции и цикл Кальвина или светонезависимые реакции. Основными реагентами являются световая энергия, углекислый газ и вода, а основными продуктами являются глюкоза и кислород.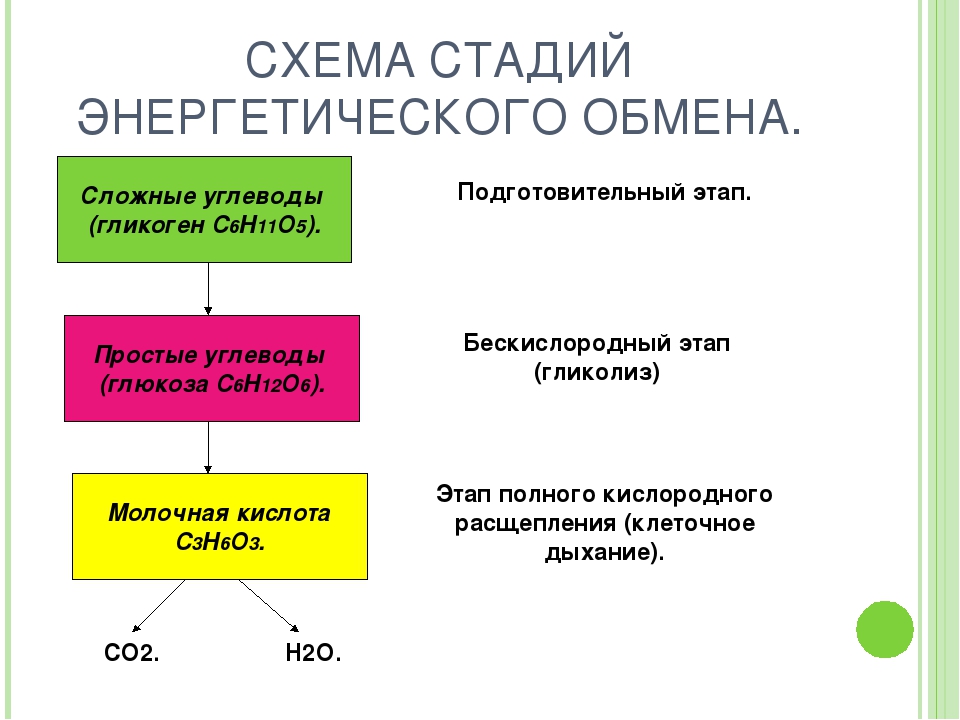
Метаболизм у прокариот может быть разным. Основные типы метаболических путей включают гетеротрофные, автотрофные, фототрофные и хемотрофные реакции. Тип метаболизма прокариот может влиять на то, где он живет и как взаимодействует с окружающей средой.
Их метаболические пути также играют роль в экологии, здоровье человека и болезнях. Например, есть прокариоты, которые не переносят кислород, такие как C. botulinum. Эти бактерии могут вызывать ботулизм, потому что они хорошо растут в местах без кислорода.
Ферменты: основы
Ферменты — это вещества, которые действуют как катализаторы , ускоряя или вызывая химические реакции. Работа большинства биохимических реакций в живых организмах зависит от ферментов.Они важны для клеточного метаболизма, поскольку могут влиять на многие процессы и способствовать их запуску.
Глюкоза и световая энергия являются наиболее распространенными источниками топлива для метаболизма клеток. Однако метаболические пути не работают без ферментов. Большинство ферментов в клетках являются белками и снижают энергию активации химических процессов.
Однако метаболические пути не работают без ферментов. Большинство ферментов в клетках являются белками и снижают энергию активации химических процессов.
Поскольку большинство реакций в клетке протекает при комнатной температуре, без ферментов они протекают слишком медленно. Например, во время гликолиза в клеточном дыхании фермент пируваткиназа играет важную роль, помогая переносить фосфатную группу.
Клеточное дыхание у эукариот
Клеточное дыхание у эукариот происходит главным образом в митохондриях. Выживание эукариотических клеток зависит от клеточного дыхания.
Во время гликолиза клетка расщепляет глюкозу в цитоплазме в присутствии кислорода или без него. Он расщепляет шестиуглеродную молекулу сахара на две трехуглеродные молекулы пирувата. Кроме того, гликолиз производит АТФ и превращает NAD + в NADH. Во время окисления пирувата пируваты попадают в митохондриальный матрикс и становятся коферментом A или ацетил-CoA . Это высвобождает углекислый газ и производит больше НАДН.
Это высвобождает углекислый газ и производит больше НАДН.
Во время цикла лимонной кислоты или цикла Кребса ацетил-КоА объединяется с оксалоацетатом с образованием цитрата . Затем цитрат вступает в реакцию с образованием диоксида углерода и НАДН. Цикл также производит FADh3 и ATP.
Во время окислительного фосфорилирования , цепь переноса электронов играет решающую роль. NADH и FADh3 отдают электроны цепи переноса электронов и становятся NAD + и FAD.Электроны движутся по этой цепочке и производят АТФ. В этом процессе также образуется вода. Большая часть производства АТФ во время клеточного дыхания находится на этой последней стадии.
Метаболизм в растениях: фотосинтез
Фотосинтез происходит в клетках растений, некоторых водорослях и некоторых бактериях, называемых цианобактериями. Этот метаболический процесс происходит в хлоропластах благодаря хлорофиллу, и вместе с кислородом он производит сахар. Светозависимые реакции плюс цикл Кальвина или светонезависимые реакции являются основными частями фотосинтеза.Это важно для общего здоровья планеты, потому что живые существа полагаются на кислород, производимый растениями.
Светозависимые реакции плюс цикл Кальвина или светонезависимые реакции являются основными частями фотосинтеза.Это важно для общего здоровья планеты, потому что живые существа полагаются на кислород, производимый растениями.
Во время светозависимых реакций в тилакоидной мембране хлоропласта пигменты хлорофилла поглощают световую энергию. Они производят АТФ, НАДФН и воду. Во время цикла Кальвина или светонезависимые реакции в строме , АТФ и НАДФН помогают производить глицеральдегид-3-фосфат, или G3P, который в конечном итоге становится глюкозой.
Как и клеточное дыхание, фотосинтез зависит от окислительно-восстановительных реакций , которые включают перенос электронов и цепь переноса электронов.
Существуют разные типы хлорофилла, наиболее распространенными из которых являются хлорофилл a, хлорофилл b и хлорофилл c. У большинства растений есть хлорофилл А, который поглощает волны синего и красного света. Некоторые растения и зеленые водоросли используют хлорофилл b. Вы можете найти хлорофилл c в динофлагеллятах.
Некоторые растения и зеленые водоросли используют хлорофилл b. Вы можете найти хлорофилл c в динофлагеллятах.
Метаболизм у прокариот
В отличие от людей и животных, прокариоты различаются по потребностям в кислороде.Некоторые прокариоты могут существовать без него, а другие зависят от него. Это означает, что у них может быть аэробный (требуется кислород) или анаэробный (не требующий кислорода) метаболизм.
Кроме того, некоторые прокариоты могут переключаться между двумя типами метаболизма в зависимости от обстоятельств или окружающей среды.
Прокариоты, метаболизм которых зависит от кислорода, — это облигатные аэробы . С другой стороны, прокариоты, которые не могут существовать в кислороде и не нуждаются в нем, — это облигатные анаэробы .Прокариоты, которые могут переключаться между аэробным и анаэробным метаболизмом в зависимости от наличия кислорода, — это факультативных анаэробов .
Молочная ферментация
Молочная ферментация — это тип анаэробной реакции, производящей энергию для бактерий. В ваших мышечных клетках также происходит ферментация молочной кислоты. Во время этого процесса клетки производят АТФ без кислорода посредством гликолиза. Процесс превращает пируват в молочную кислоту и производит НАД + и АТФ.
Этот процесс находит множество применений в промышленности, например, в производстве йогурта и этанола. Например, бактерии Lactobacillus bulgaricus помогают производить йогурт. Бактерии сбраживают лактозу, сахар в молоке, с образованием молочной кислоты. Это делает молочный сгусток и превращает его в йогурт.
Каков метаболизм клеток у различных типов прокариот?
Прокариоты можно разделить на разные группы в зависимости от их метаболизма. Основные типы — гетеротрофный, автотрофный, фототрофный и хемотрофный.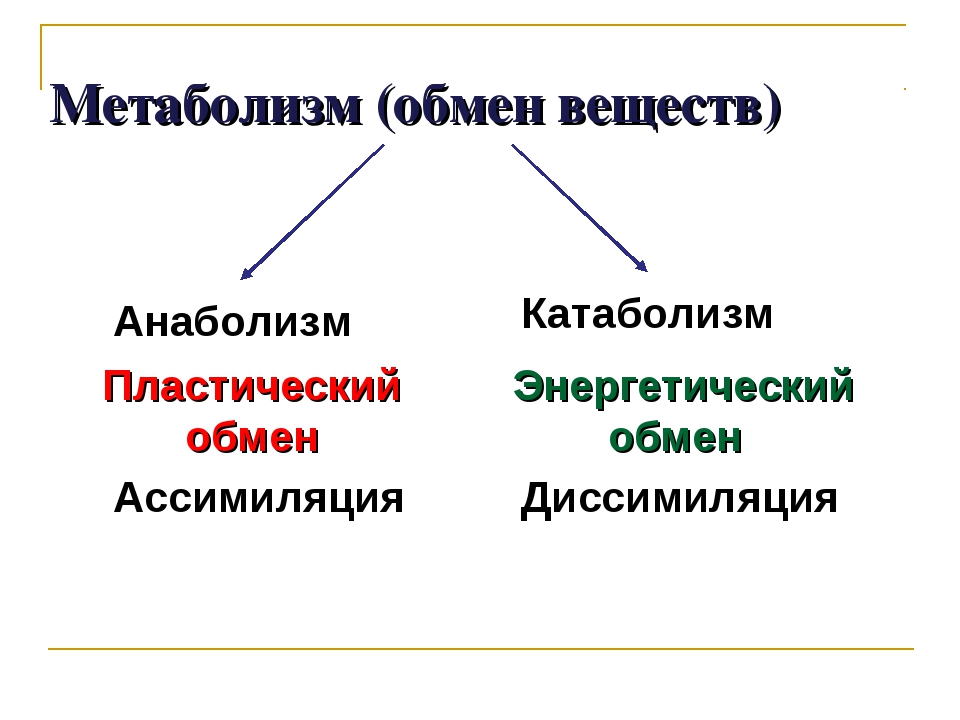 Тем не менее, всем прокариотам для жизни по-прежнему нужен какой-то тип энергии или топлива .
Тем не менее, всем прокариотам для жизни по-прежнему нужен какой-то тип энергии или топлива .
Гетеротрофные прокариоты получают органические соединения от других организмов для получения углерода. Автотрофные прокариоты используют углекислый газ в качестве источника углерода. Многие могут использовать фотосинтез для этого. Фототрофные прокариоты получают энергию от света.
Хемотрофные прокариоты получают энергию от химических соединений, которые они расщепляют.
Анаболический vs.Катаболический
Вы можете разделить метаболические пути на анаболические и катаболические категории. Анаболические означают, что им требуется энергия и они используют ее для создания больших молекул из маленьких. Катаболический означает, что они выделяют энергию и разбивают большие молекулы на более мелкие. Фотосинтез — это анаболический процесс, а клеточное дыхание — это катаболический процесс.
Эукариоты и прокариоты зависят от клеточного метаболизма, чтобы жить и развиваться. Хотя их процессы различны, они оба либо используют, либо создают энергию.Клеточное дыхание и фотосинтез — наиболее распространенные пути, наблюдаемые в клетках. Однако у некоторых прокариот разные метаболические пути, которые уникальны.
Хотя их процессы различны, они оба либо используют, либо создают энергию.Клеточное дыхание и фотосинтез — наиболее распространенные пути, наблюдаемые в клетках. Однако у некоторых прокариот разные метаболические пути, которые уникальны.
Метаболизм, клеточный — Биологическая энциклопедия
Автор фотографии: Luk Cox
Клеточный метаболизм — это сумма всех биохимических реакций.
происходит внутри клетки. Он включает в себя все реакции, связанные с
разлагая молекулы пищи, синтезируя
макромолекулы
необходимы клетке, а также в производстве небольших молекул-предшественников, таких как
немного
аминокислоты
, для сотовых нужд.Он также включает все реакции с участием электронов.
перенос (окислительно-восстановительные или окислительно-восстановительные реакции). Обмен веществ занимает
место в последовательности биохимических реакций, называемых путями.
Метаболические пути
Метаболические пути могут быть простыми линейными последовательностями нескольких реакций или
они могут быть сильно разветвленными, при этом реакции сходятся или расходятся.
от центральной главной дороги. Они могут быть циклическими, с предшественником
ранняя реакция регенерируется в конце пути (например,
Цикл Кребса
из
аэробный
дыхание, или цикл фотосинтеза Кальвина).Некоторые пути служат
несколько целей. Например, цикл Кребса наиболее известен своей ролью
в окисляющих сахарах и других
органический
молекулы для обеспечения аденозинтрифосфата (
АТФ
) для клетки, но он также используется в качестве источника молекул-предшественников для
клеточный биосинтез. Ясно, что эволюция неоднократно использовала существующие
метаболические пути для обеспечения новых функций.
Все биохимические реакции проходят
катализированный
к
белки
называется
ферменты
; для большинства реакций есть один фермент, который катализирует только
реакция.Ферменты могут точно регулироваться клеткой, обеспечивая
высокая степень контроля клеточного метаболизма. Активность ферментов
часто чувствительны к количеству определенных молекул в клетке. За
например, ферменты, участвующие в производстве АТФ, часто ингибируются АТФ;
когда в клетке достаточно этого
метаболит
, следовательно, пути, которые его производят, отключены, тем самым
предотвращение бесполезных реакций.В качестве альтернативы те же самые ферменты могут быть
сильно активируется предшественником АТФ, аденозиндифосфатом (
ADP
), уровень которого повышается, когда клетка выполняет работу и нуждается в
быстрое образование АТФ. Этот образец регуляции молекулами, которые
предшественники или продукты пути распространены в клеточном
метаболизм.
Анаболизм и катаболизм
Метаболизм делится на две большие категории.Катаболизм, или
деградация молекул, обычно включает удаление электронов из
молекул (окисление) и обычно сопровождается высвобождением
энергия. Анаболизм или синтез сложных молекул обычно включает:
обогащает молекулы электронами (восстановление) и обычно требует
клетка расходует энергию в виде АТФ. Реакции, дающие энергию,
такие как большинство катаболических реакций, называются экзергоническими, тогда как те, которые
требуют ввода энергии, например, большинство
анаболический
реакции, называются эндергоническими.
Основная функция анаболических путей — синтез четырех
классы макромолекул, необходимые клетке:
полисахариды
,
липиды
, нуклеиновые кислоты и белки. Хотя эти четыре категории
химически различны, все они синтезируются одним и тем же общим типом
реакция, конденсационный синтез отдельных малых субъединиц (мономеров)
в макромолекулы (полимеры).В реакции конденсации водород
атом удален из одного
мономер
, а
гидроксил
группа от другой, образуя воду. Между двумя
мономеры, из которых была удалена вода:
—А — ОН + Н — А— → Н
2
О + —А — А—
Например, нуклеиновые кислоты, такие как ДНК и РНК, синтезируются из их
мономеры,
нуклеотиды
, путем конденсационного синтеза.Полисахариды и белки производятся в
аналогичным образом из их мономеров, сахаров и аминокислот,
соответственно. Липиды, четвертый класс макромолекул, в некоторой степени
разные. В отличие от других макромолекул, которые
Сканирующая электронная микрофотография эпителиальной клетки кишечника.
показаны две лизомы, содержащие пищеварительные ферменты, разрушающие
поврежденные молекулы. Одна категория метаболизма — катаболизм,
деградация молекул.
состоит из длинных цепей мономеров, большинство липидов имеют всего три или четыре
различные молекулярные субъединицы, наиболее важными из которых являются жирные кислоты.
Жирные кислоты не соединены напрямую друг с другом, но соединены
другая молекула, такая как глицерин (для жиров и масел). Однако жирный
кислоты присоединяются к глицерину в результате той же реакции дегидратации, что и
в других группах макромолекул.
Обратный конденсационный синтез
гидролиз
, в котором молекула воды присоединена к связи между двумя мономерами,
его разрушение и разделение мономеров.Один из водородов из воды
присоединяется к одному из мономеров, а оставшийся гидроксил
к другому прикреплен:
ЧАС
2
О + -А-А- → -А-ОН +
H — A—
Например, нуклеиновые кислоты разлагаются до мономеров, когда вода
вставлен между отдельными нуклеотидными мономерами, разрывая связь
что присоединяется к ним. Реакции гидролиза — это разновидность катаболической реакции,
хотя они обычно не производят напрямую АТФ; они, однако,
производят мономеры, которые часто далее катаболизируются с образованием АТФ.
Оборот
Обмен веществ — это динамический процесс. Клетка постоянно деградирует и
синтезирующие молекулы. В целом катаболические пути обеспечивают
энергия в виде АТФ, которая используется для управления анаболическими процессами.
Это необходимо, поскольку для протекания эндергонических реакций требуется
вход энергии, который они получают от АТФ. Это достигается
сочетание эндергонической реакции с гидролизом АТФ до АДФ и
неорганический
фосфат, экзэргоническая реакция.Пока количество энергии
меньше, чем количество, высвобождаемое при гидролизе АТФ, связанная
реакции будут продолжаться.
Динамический характер метаболизма приводит к постоянной деградации и
восстановление большинства ячеистых материалов. Например, белки существуют в
ячейку в течение относительно короткого времени, от минут до недель, причем большинство
белки со средней продолжительностью жизни несколько дней. Структурные белки
обычно служат дольше, чем ферменты, но они тоже в конечном итоге разлагаются
и синтезировал заново.Аналогичным образом переворачиваются и другие ячеистые материалы.
аналогичным образом. Этот постоянный оборот ячеистых материалов сохраняет
клетка в хорошем состоянии. Молекулы, которые могли быть повреждены в результате
например, будучи частично окисленным, рано или поздно разрушится и
заменены.
Клеточный метаболизм — это самый фундаментальный уровень, на котором динамический
свойства жизни начинают проявляться. Сложные взаимодействия разнообразных
пути, их регулирование и их организация демонстрируют
изысканная утонченность биохимии жизни.Все происходящие процессы
внутри отдельных организмов можно проследить пути клеточного
метаболизм.
Библиография
Hinckle, P.C. и R.E. McCarthy. «Как клетки производят АТФ».
Scientific American
238 (март 1978 г.): 104–123.
Страйер, Люберт.
Биохимия.
Нью-Йорк: В. Х. Фриман и компания, 1995.
Другие статьи, которые могут вам понравиться:
Введение в клеточный метаболизм
Клеточный метаболизм относится к жизненно важным метаболическим реакциям, происходящим внутри клетки.Когда большинство людей думают о «метаболизме», они ассоциируют его с «сжиганием» или расщеплением питательных веществ. Однако в клеточной биологии метаболизм включает в себя «катаболизм», который представляет собой расщепление молекул, и «анаболизм», который представляет собой синтез новых биологических соединений. Эти процессы обеспечивают клетки энергией и соответственно помогают строить их компоненты.
В этом видео мы углубимся в основные открытия, которые внесли свой вклад в наше понимание клеточного метаболизма. Мы продолжим изучение ключевых вопросов в данной области и некоторых методов, используемых в настоящее время для изучения метаболических путей.
Давайте погрузимся в богатую историю клеточного метаболизма.
Между 1770 и 1805 годами четыре химика провели ключевые эксперименты, которые помогли объяснить, как растения производят «массу» для роста. Их работа привела к основной реакции фотосинтеза, которая установила, что при солнечном свете растения поглощают углекислый газ и воду и производят кислород и органические вещества. Позже, в 1860-х годах, Юлиус фон Сакс определил, что этот органический материал представляет собой крахмал, состоящий из сахарной глюкозы.
Итак, сахар производят заводы. Но мы потребляем это. Так что же происходит с сахаром в нашем организме? Возможный ответ пришел в 1930-х годах, когда Густав Эмбден, Отто Мейерхоф и Якоб Парнас описали гликолиз — путь, по которому глюкоза расщепляется на пируват. Теперь мы знаем, что гликолиз также производит аденозинтрифосфат или АТФ.
Структура
АТФ была определена в 1935 году в лаборатории Мейерхофа Карлом Ломанном. Мейерхоф и Ломанн предположили, что АТФ может «хранить» энергию, что было подтверждено Фрицем Липманном в 1941 году, который идентифицировал богатые энергией связи в АТФ и представил теорию, с помощью которой эти связи могут использоваться во время биосинтеза.
Параллельно Ганс Кребс обнаружил, что окисление глюкозы или пирувата может стимулироваться рядом кислот, каждая из которых является частью циклических реакций, образующих цикл трикарбоновых кислот, сокращенно называемый циклом TCA. Его главный вклад заключался в том, что оксалоацетат и пируват можно было преобразовать в цитрат, что дало этой серии окисления циклическую форму.
В 1946 году Липманн и Натан Каплан дополнительно объяснили реакцию превращения пирувата в цитрат, открыв кофермент А.Теперь мы знаем, что пируват взаимодействует с этим ферментом с образованием ацетил-кофермента A, который запускает цикл TCA.
Позже, между 1950-ми и 1970-ми годами, исследователи определили, что электроны, высвобождаемые во время цикла TCA, могут «переноситься» в белковые комплексы, расположенные в митохондриях, по пути, называемому цепью переноса электронов. Важно отметить, что в 1961 году Питер Митчелл предположил, что перенос электронов между этими комплексами создает протонный «градиент», который может управлять производством большей части АТФ в клетке.
В совокупности открытия фотосинтеза, гликолиза, цикла TCA и цепи переноса электронов сформировали основу, на которой сегодня зиждутся исследования клеточного метаболизма.
Хотя эти исторические открытия предоставили огромное понимание метаболических путей, они также породили несколько вопросов. Давайте рассмотрим некоторые из тех, на которые нет ответа.
Сегодня исследователи изучают, как на метаболические пути влияют стрессовые факторы окружающей среды, такие как токсины или радиация.В частности, есть интерес к тому, как такие факторы приводят к аномальному производству активных форм кислорода, таких как свободные радикалы, которые обладают неспаренными электронами на атомах кислорода, что делает их очень реактивными. Эти молекулы могут повредить другие клеточные компоненты и привести к окислительному стрессу.
Окислительный стресс вовлечен в клеточное старение и смерть, а также в инициирование и прогрессирование рака. Следовательно, клеточные биологи заинтересованы в определении того, как эти активные виды влияют на нормальные физиологические процессы клетки, такие как деление клетки.Обладая этой информацией, они могут сделать дальнейшие выводы о роли этих видов в патологических событиях.
Наконец, несколько исследователей интересуются метаболическими нарушениями — состояниями, при которых нарушаются определенные метаболические реакции. К ним относятся такие заболевания, как диабет, когда организм не может усваивать сахар. В настоящее время исследователи пытаются определить факторы, такие как гены или сигналы окружающей среды, которые способствуют возникновению таких заболеваний. В конечном итоге это поможет им разработать более эффективные методы лечения пациентов.
Теперь, когда вы услышали несколько неотложных вопросов в области клеточного метаболизма, давайте рассмотрим экспериментальные методы, которые ученые используют для их решения.
Конечная цель многих катаболических процессов в живых клетках — выработка АТФ, который является основной молекулой хранения энергии, используемой клетками. Следовательно, такие методы, как анализ биолюминесценции АТФ, который определяет количественно АТФ в образце с помощью реакции люминесценции, могут дать представление о метаболической активности клеток.
Другие методы сосредоточены на конкретных метаболических путях. Например, исследователи могут оценить метаболизм гликогена в его мономер глюкозу. Один из способов сделать это — переработать глюкозу, полученную из гликогена, в продукты, которые будут реагировать с детектирующими зондами и вызывать изменение цвета или флуоресценцию. Таким образом, исследователи могут рассчитать, сколько гликогена изначально присутствовало в их образцах.
Напротив, аномальный метаболизм может быть обнаружен путем измерения активных форм кислорода.Обычно исследователи используют зонд, который флуоресцирует после «нападения» представителя этих видов. Эти анализы напрямую определяют количество активных метаболитов кислорода и, следовательно, помогают в обнаружении окислительного стресса.
Наконец, исследователи анализируют метаболизм на уровне организма с помощью «Метаболического профилирования». С помощью передовых методов, таких как высокоэффективная жидкостная хроматография или ВЭЖХ, масс-спектрометрия или МС, ученые могут количественно определять метаболиты, присутствующие в биологических образцах, и определять, застопорились ли определенные метаболические пути или они чрезмерно активны.
Имея в своем распоряжении все эти инструменты, давайте посмотрим, как ученые применяют их в экспериментах.
Некоторые ученые применяют эти методы для разработки новых способов диагностики нарушений обмена веществ. Здесь был разработан протокол для выделения мононуклеарных клеток периферической крови или PBMC из образцов крови пациентов с целью оценки содержания в них гликогена. Используя анализ окрашивания, специфичного для метаболизма гликогена, исследователи получили представление о количестве гликогена, присутствующего в этих образцах.В будущем этот метод может помочь диагностировать пациентов с заболеваниями, связанными с метаболизмом гликогена.
Другие исследователи используют эти инструменты для изучения влияния стресса окружающей среды на метаболизм. В этом эксперименте ученые измерили активные формы кислорода у эмбрионов рыбок данио, обработанных химическим веществом, называемым ротеноном, или после повреждения их хвостов. Это было сделано с помощью зонда, который флуоресцирует красным при наведении на активные формы кислорода. Последующая оценка целых эмбрионов показала повышенную продукцию этих молекул в ответ на повреждение и химическое воздействие, что свидетельствует о защитной роли этих метаболитов.
Наконец, клеточные биологи также изучают метаболические характеристики раковых клеток. Здесь исследователи собрали содержимое клеток рака толстой кишки человека и подвергли этот экстракт метаболическому профилированию с помощью ВЭЖХ и МС. Это позволило исследователям идентифицировать метаболиты, присутствующие в этой больной ткани.
Вы только что посмотрели вводное видео JoVE о клеточном метаболизме. Многие сложные пути описывают метаболическую активность клеток, и теперь вы знаете, как эти пути были обнаружены и как исследователи все еще пытаются расшифровать неизвестные компоненты.Помните, обмен веществ — это хорошо, но избыток чего-либо может быть вредным. Как всегда, спасибо за просмотр!
Что такое метаболизм?
4 сентября 2015 г.
2 мин чтения
ДОБАВИТЬ ТЕМУ В СООБЩЕНИЯ ПО ЭЛЕКТРОННОЙ ПОЧТЕ
Получать электронное письмо, когда новые статьи публикуются на
Укажите свой адрес электронной почты, чтобы получать сообщения о публикации новых статей.Подписаться
Нам не удалось обработать ваш запрос. Пожалуйста, повторите попытку позже. Если проблема не исчезнет, обратитесь по адресу [email protected].
Вернуться в Healio
Метаболизм — это термин, обозначающий набор химических реакций, которые происходят в клетках живых организмов для поддержания жизни. Метаболические процессы приводят к росту и размножению и позволяют живым организмам поддерживать свои структуры и реагировать на окружающую среду.Все химические реакции, происходящие в живых организмах, от пищеварения до транспорта веществ от клетки к клетке, могут быть частью метаболизма.
Промежуточный или промежуточный метаболизм — это термин для переноса веществ в разные клетки и между ними.
Как это работает
Есть две категории метаболизма: катаболизм и анаболизм. Катаболизм — это распад органического вещества, а анаболизм использует энергию для создания компонентов клетки, таких как белки и нуклеиновые кислоты.
Химические реакции в метаболическом процессе организованы в метаболические пути, при которых одно химическое вещество за несколько этапов превращается в другое. Ферменты помогают в этом процессе, облегчая реакции и выступая в качестве катализаторов протекания реакций. Реакции не могли бы происходить без ферментов, которые отвечают на сигналы между клетками и регулируют метаболические пути. Скорость метаболизма называется скоростью метаболизма.
Метаболизм живого организма позволяет ему определять, какие вещества питательны и полезны, а какие ядовиты.
Некоторыми другими химическими веществами и частями организма, участвующими в метаболическом процессе, являются аминокислоты, белки, липиды, углеводы, нуклеотиды, коферменты, минералы и кофакторы.
Метаболический синдром
Метаболический синдром описывает группу черт и привычек, повышающих риск ишемической болезни сердца, диабета и инсульта. Факторы риска включают избыток жира в желудке, высокий уровень триглицеридов, низкий уровень холестерина ЛПВП, также известный как «хороший холестерин», высокое кровяное давление и высокий уровень сахара в крови натощак.
Эти факторы обычно встречаются вместе. Однако у пациентов должно быть не менее трех из них, чтобы диагностировать метаболический синдром.
У человека с метаболическим синдромом вдвое риск развития сердечных заболеваний и в пять раз выше вероятность диагностирования диабета, чем у человека без метаболического синдрома. Это становится все более распространенным в результате роста показателей ожирения среди взрослых. Можно предотвратить или отсрочить метаболический синдром с помощью здорового питания и физических упражнений.
Дополнительную информацию можно найти на следующих сайтах:
http://bloodjournal.hemologylibrary.org/cgi/collection/gene_expression
http://www.nlm.nih.gov/medlineplus/ency/article/003706.htm
http://www.mayoclinic.com/health/metabolism/WT00006/
http://www.nature.com/jcbfm/index.html
http://www.nutritionandmetabolism.com/
http://www.hormone.org/Public/endocrinologist.cfm
http: // www.nlm.nih.gov/medlineplus/ency/article/002257.htm
http://www.ncbi.nlm.nih.gov/books/NBK22/?depth=10
http://endo.endojournals.org/
http://www.mayoclinic.org/medicalprofs/glucocorticoid-induced-diabetes.html
http://www.nlm.nih.gov/medlineplus/steroids.html
http://www.cancer.gov/cancertopics/understandingcancer/estrogenreceptors
http://www.ncbi.nlm.nih.gov/gene/2099
http://ghr.nlm.nih.gov/glossary=enzyme
http: // www.nlm.nih.gov/medlineplus/ency/article/002353.htm
http://www.nhlbi.nih.gov/health/health-topics/topics/ms
ДОБАВИТЬ ТЕМУ В СООБЩЕНИЯ ПО ЭЛЕКТРОННОЙ ПОЧТЕ
Получать электронное письмо, когда новые статьи публикуются на
Укажите свой адрес электронной почты, чтобы получать сообщения о публикации новых статей.
Подписаться
Нам не удалось обработать ваш запрос.Пожалуйста, повторите попытку позже. Если проблема не исчезнет, обратитесь по адресу [email protected].
Вернуться в Healio
Энергия и обмен веществ | Безграничная биология
Роль энергии и метаболизма
Всем организмам требуется энергия для выполнения задач; метаболизм — это набор химических реакций, высвобождающих энергию для клеточных процессов.
Цели обучения
Объясните важность обмена веществ
Ключевые выводы
Ключевые моменты
- Всем живым организмам нужна энергия для роста и размножения, поддержания своей структуры и реакции на окружающую среду; метаболизм — это набор процессов, делающих энергию доступной для клеточных процессов.
- Метаболизм — это комбинация химических реакций, которые являются спонтанными и высвобождают энергию, и химических реакций, которые не являются спонтанными и требуют энергии для протекания.
- Живые организмы должны получать энергию через пищу, питательные вещества или солнечный свет, чтобы выполнять клеточные процессы.
- Транспортировка, синтез и распад питательных веществ и молекул в клетке требуют использования энергии.
Ключевые термины
- метаболизм : полный набор химических реакций, происходящих в живых клетках
- биоэнергетика : исследование превращений энергии, происходящих в живых организмах
- энергия : работоспособность
Энергия и обмен веществ
Всем живым организмам нужна энергия для роста и воспроизводства, поддержания своей структуры и реакции на окружающую среду.Метаболизм — это набор поддерживающих жизнь химических процессов, которые позволяют организмам преобразовывать химическую энергию, хранящуюся в молекулах, в энергию, которая может использоваться для клеточных процессов. Животные потребляют пищу для восполнения энергии; их метаболизм расщепляет углеводы, липиды, белки и нуклеиновые кислоты, чтобы обеспечить химическую энергию для этих процессов. В процессе фотосинтеза растения преобразуют световую энергию солнца в химическую энергию, хранящуюся в молекулах.
Биоэнергетика и химические реакции
Ученые используют термин биоэнергетика, чтобы обсудить концепцию потока энергии через живые системы, такие как клетки.Клеточные процессы, такие как построение и разрушение сложных молекул, происходят в результате пошаговых химических реакций. Некоторые из этих химических реакций являются спонтанными и высвобождают энергию, тогда как другие требуют энергии для протекания. Все химические реакции, происходящие внутри клеток, включая те, которые используют энергию, и те, которые высвобождают энергию, являются метаболизмом клетки.
Большая часть энергии прямо или косвенно исходит от Солнца. : Большинство форм жизни на Земле получают энергию от Солнца.Растения используют фотосинтез для улавливания солнечного света, а травоядные животные поедают эти растения для получения энергии. Плотоядные животные поедают травоядных, а разлагатели переваривают растительную и животную материю.
Клеточный метаболизм
Каждое задание, выполняемое живыми организмами, требует энергии. Энергия необходима для выполнения тяжелой работы и упражнений, но люди также расходуют много энергии во время размышлений и даже во время сна. При каждом действии, требующем энергии, происходит множество химических реакций, обеспечивающих химическую энергию системам тела, включая мышцы, нервы, сердце, легкие и мозг.
Живые клетки каждого организма постоянно используют энергию для выживания и роста. Клетки расщепляют сложные углеводы на простые сахара, которые клетка может использовать для получения энергии. Мышечные клетки могут потреблять энергию для построения длинных мышечных белков из небольших молекул аминокислот. Молекулы можно модифицировать и переносить по клетке или можно распространять по всему организму. Так же, как энергия требуется как для строительства, так и для сноса здания, энергия требуется как для синтеза, так и для разрушения молекул.
Многие клеточные процессы требуют постоянного снабжения энергией, обеспечиваемой метаболизмом клетки. Сигнальные молекулы, такие как гормоны и нейротрансмиттеры, должны быть синтезированы и затем транспортированы между клетками. Патогенные бактерии и вирусы попадают в организм и разрушаются клетками. Клетки также должны экспортировать отходы и токсины, чтобы оставаться здоровыми, и многие клетки должны плавать или перемещать окружающие материалы посредством биения клеточных придатков, таких как реснички и жгутики.
Еда дает энергию для таких действий, как полет. : Колибри нужна энергия, чтобы поддерживать длительные периоды полета.Колибри получает энергию, принимая пищу и превращая питательные вещества в энергию с помощью ряда биохимических реакций. Летательные мышцы птиц чрезвычайно эффективны в производстве энергии.
Типы энергии
Различные типы энергии включают кинетическую, потенциальную и химическую энергию.
Цели обучения
Различия между видами энергии
Ключевые выводы
Ключевые моменты
- Все организмы используют разные формы энергии для поддержания биологических процессов, которые позволяют им расти и выживать.
- Кинетическая энергия — это энергия, связанная с движущимися объектами.
- Потенциальная энергия — это тип энергии, связанный со способностью объекта выполнять работу.
- Химическая энергия — это энергия, выделяющаяся при разрыве химических связей, которая может быть использована для метаболических процессов.
Ключевые термины
- химическая энергия : Чистая потенциальная энергия, высвобождаемая или поглощаемая в ходе химической реакции.
- потенциальная энергия : энергия, которой обладает объект из-за его положения (в гравитационном или электрическом поле) или его состояния (в виде растянутой или сжатой пружины, в качестве химического реагента или из-за наличия массы покоя).
- кинетическая энергия : энергия, которой обладает объект из-за его движения, равная половине массы тела, умноженной на квадрат его скорости.
Энергия — это свойство объектов, которое может быть передано другим объектам или преобразовано в другие формы, но не может быть создано или уничтожено. Организмы используют энергию для выживания, роста, реакции на раздражители, воспроизводства и для всех типов биологических процессов. Потенциальная энергия, хранящаяся в молекулах, может быть преобразована в химическую энергию, которая в конечном итоге может быть преобразована в кинетическую энергию, позволяющую организму двигаться.В конце концов, большая часть энергии, используемой организмами, преобразуется в тепло и рассеивается.
Кинетическая энергия
Энергия, связанная с движущимися объектами, называется кинетической энергией. Например, когда самолет находится в полете, он очень быстро движется по воздуху, выполняя работу по изменению своего окружения. Реактивные двигатели преобразуют потенциальную энергию топлива в кинетическую энергию движения. Крушащий шар может нанести большой урон даже при медленном движении.Однако все еще разрушающийся шар не может выполнять никакой работы и, следовательно, не имеет кинетической энергии. Ускоряющаяся пуля, идущий человек, быстрое движение молекул в воздухе, выделяющих тепло, и электромагнитное излучение, такое как солнечный свет, — все это имеет кинетическую энергию.
Потенциальная энергия
Что, если тот же самый неподвижный шар для разрушения поднять на два этажа над автомобилем с краном? Если подвешенный шар для разрушения не движется, связана ли с ним энергия? Да, разрушающий шар обладает энергией, потому что разрушающий шар может выполнять свою работу.Эта форма энергии называется потенциальной энергией, потому что объект может выполнять работу в данном состоянии.
Объекты переносят свою энергию между потенциальным и кинетическим состояниями. Поскольку разрушающий шар неподвижно висит, он имеет [latex] \ text {0%} [/ latex] кинетическую и [latex] \ text {100%} [/ latex] потенциальную энергию. Когда мяч выпущен, его кинетическая энергия увеличивается по мере того, как мяч набирает скорость. В то же время мяч теряет потенциальную энергию при приближении к земле. Другие примеры потенциальной энергии включают энергию воды, удерживаемой за плотиной, или человека, который собирается прыгнуть с парашютом из самолета.
Зависимость потенциальной энергии от кинетической : Вода за плотиной имеет потенциальную энергию. Движущаяся вода, например, в водопаде или быстро текущей реке, обладает кинетической энергией.
Химическая энергия
Потенциальная энергия связана не только с местонахождением материи, но и со структурой материи. Пружина на земле обладает потенциальной энергией, если она сжата, как и натянутая резинка. Тот же принцип применим к молекулам. На химическом уровне связи, которые удерживают атомы молекул вместе, обладают потенциальной энергией.Этот тип потенциальной энергии называется химической энергией, и, как и вся потенциальная энергия, ее можно использовать для выполнения работы.
Например, химическая энергия содержится в молекулах бензина, которые используются в автомобилях. Когда газ воспламеняется в двигателе, связи в его молекулах разрываются, и выделяемая энергия используется для приведения в движение поршней. Потенциальная энергия, хранящаяся в химических связях, может быть использована для выполнения работы для биологических процессов. Различные метаболические процессы разрушают органические молекулы, высвобождая энергию для роста и выживания организма.
Химическая энергия : Молекулы в бензине (октановое число, указанная химическая формула) содержат химическую энергию. Эта энергия преобразуется в кинетическую энергию, которая позволяет автомобилю мчаться по гоночной трассе.
Метаболические пути
Анаболический путь требует энергии и строит молекулы, в то время как катаболический путь производит энергию и разрушает молекулы.
Цели обучения
Опишите два основных типа метаболических путей
Ключевые выводы
Ключевые моменты
- Метаболический путь — это серия химических реакций в клетке, которые создают и разрушают молекулы для клеточных процессов.
- Анаболические пути синтезируют молекулы и требуют энергии.
- Катаболические пути расщепляют молекулы и производят энергию.
- Поскольку почти все метаболические реакции происходят не спонтанно, белки, называемые ферментами, помогают облегчить эти химические реакции.
Ключевые термины
- катаболизм : деструктивный метаболизм, обычно включающий выделение энергии и расщепление материалов
- фермент : глобулярный белок, катализирующий биологическую химическую реакцию
- анаболизм : конструктивный метаболизм тела в отличие от катаболизма
Метаболические пути
Процессы производства и расщепления углеводных молекул иллюстрируют два типа метаболических путей.Метаболический путь — это последовательный ряд взаимосвязанных биохимических реакций, которые преобразуют молекулу или молекулы субстрата через ряд промежуточных продуктов метаболизма, в конечном итоге приводя к конечному продукту или продуктам. Например, один из метаболических путей углеводов расщепляет большие молекулы на глюкозу. Другой метаболический путь может превращать глюкозу в большие молекулы углеводов для хранения. Первый из этих процессов требует энергии и называется анаболическим. Второй процесс производит энергию и называется катаболическим.Следовательно, метаболизм состоит из этих двух противоположных путей:
- Анаболизм (построение молекул)
- Катаболизм (разрушение молекул)
Анаболические и катаболические пути : Анаболические пути — это те пути, которые требуют энергии для синтеза более крупных молекул. Катаболические пути — это те, которые генерируют энергию за счет разрушения более крупных молекул. Оба типа путей необходимы для поддержания энергетического баланса клетки.
Анаболические пути
Анаболические пути требуют ввода энергии для синтеза сложных молекул из более простых.Одним из примеров анаболического пути является синтез сахара из CO 2 . Другие примеры включают синтез больших белков из строительных блоков аминокислот и синтез новых цепей ДНК из строительных блоков нуклеиновых кислот. Эти процессы имеют решающее значение для жизни клетки, происходят постоянно и требуют энергии, обеспечиваемой АТФ и другими высокоэнергетическими молекулами, такими как НАДН (никотинамидадениндинуклеотид) и НАДФН.
Катаболические пути
Катаболические пути включают разложение сложных молекул на более простые, высвобождая химическую энергию, хранящуюся в связях этих молекул.Некоторые катаболические пути могут захватывать эту энергию для производства АТФ, молекулы, используемой для питания всех клеточных процессов. Другие запасающие энергию молекулы, такие как липиды, также расщепляются посредством аналогичных катаболических реакций, высвобождая энергию и производя АТФ.
Важность ферментов
Химические реакции в метаболических путях редко происходят спонтанно. Каждая стадия реакции облегчается или катализируется белком, называемым ферментом. Ферменты важны для катализирования всех типов биологических реакций: тех, которые требуют энергии, а также тех, которые выделяют энергию.
Метаболизм углеводов
Организмы расщепляют углеводы для производства энергии для клеточных процессов, а фотосинтезирующие растения производят углеводы.
Цели обучения
Анализируйте важность углеводного обмена для производства энергии
Ключевые выводы
Ключевые моменты
- Распад глюкозы, которую живые организмы используют для производства энергии, описывается уравнением: [латекс] {\ text {C}} _ {6} {\ text {H}} _ {12} {\ text {O}} _ {6} +6 {\ text {O}} _ {2} \ rightarrow 6 {\ text {CO}} _ {2} +6 {\ text {H}} _ {2} \ text {O} + \ text {энергия} [/ латекс].
- Процесс фотосинтеза, который растения используют для синтеза глюкозы, описывается уравнением: [латекс] 6 \ text {CO} _ {2} +6 {\ text {H}} _ {2} \ text {O} + \ text { энергия} \ rightarrow {\ text {C}} _ {6} {\ text {H}} _ {12} {\ text {O}} _ {6} +6 \ text {O} _ {2} [/ латекс].
- Потребляемая глюкоза используется для производства энергии в виде АТФ, который используется для выполнения работы и химических реакций в клетке.
- Во время фотосинтеза растения превращают световую энергию в химическую энергию, которая используется для создания молекул глюкозы.
Ключевые термины
- аденозинтрифосфат : многофункциональный нуклеозидтрифосфат, используемый в клетках в качестве кофермента, часто называемый «молекулярной единицей энергетической валюты» при внутриклеточной передаче энергии
- глюкоза : простой моносахарид (сахар) с молекулярной формулой C6h22O6; это основной источник энергии для клеточного метаболизма
Метаболизм углеводов
Углеводы — одна из основных форм энергии для животных и растений.Растения вырабатывают углеводы, используя световую энергию солнца (в процессе фотосинтеза), в то время как животные едят растения или других животных для получения углеводов. Растения хранят углеводы в длинных полисахаридных цепях, называемых крахмалом, в то время как животные хранят углеводы в виде молекулы гликогена. Эти большие полисахариды содержат много химических связей и, следовательно, хранят много химической энергии. Когда эти молекулы расщепляются во время метаболизма, энергия химических связей высвобождается и может использоваться для клеточных процессов.
Все живые существа используют углеводы как форму энергии. : Растения, такие как дуб и желудь, используют энергию солнечного света для производства сахара и других органических молекул. И растения, и животные (например, эта белка) используют клеточное дыхание для получения энергии из органических молекул, изначально производимых растениями
Производство энергии из углеводов (клеточное дыхание)
Метаболизм любого моносахарида (простого сахара) может производить энергию для использования клеткой.Избыточные углеводы хранятся в виде крахмала в растениях и в виде гликогена у животных, готовые к метаболизму, если потребность организма в энергии внезапно возрастет. Когда эта потребность в энергии увеличивается, углеводы расщепляются на составляющие моносахариды, которые затем распределяются по всем живым клеткам организма. Глюкоза (C 6 H 12 O 6 ) является типичным примером моносахаридов, используемых для производства энергии.
Внутри клетки каждая молекула сахара расщепляется в ходе сложной серии химических реакций.Поскольку химическая энергия высвобождается из связей в моносахариде, она используется для синтеза высокоэнергетических молекул аденозинтрифосфата (АТФ). АТФ — это основная энергетическая валюта всех клеток. Точно так же, как доллар используется в качестве валюты для покупки товаров, клетки используют молекулы АТФ для немедленной работы и проведения химических реакций.
Распад глюкозы во время метаболизма — клеточное дыхание — можно описать уравнением:
[латекс] {C} _ {6} {H} _ {12} {O} _ {6} +6 {O} _ {2} \ rightarrow 6 {CO} _ {2} +6 {H} _ {2} О + энергия [/ латекс]
Производство углеводов (фотосинтез)
Растения и некоторые другие виды организмов производят углеводы в процессе фотосинтеза.Во время фотосинтеза растения преобразуют энергию света в химическую энергию, превращая молекулы углекислого газа (CO 2 ) в молекулы сахара, такие как глюкоза. Поскольку этот процесс включает в себя создание связей для синтеза большой молекулы, для его продолжения требуется ввод энергии (света). Синтез глюкозы путем фотосинтеза описывается этим уравнением (обратите внимание, что оно является обратным предыдущему уравнению):
[латекс] 6CO_ {2} +6 {H} _ {2} O + энергия \ rightarrow {C} _ {6} {H} _ {12} {O} _ {6} + 6O_ {2} [/ латекс]
В рамках химических процессов растений молекулы глюкозы могут объединяться и превращаться в другие типы сахаров.В растениях глюкоза хранится в форме крахмала, который может быть снова расщеплен на глюкозу посредством клеточного дыхания для обеспечения АТФ.
Что такое клеточный метаболизм? 3 стадии клеточного метаболизма
3 стадии клеточного метаболизма
Клеточный метаболизм относится к химическим реакциям, происходящим внутри клеток. В эукариотических клетках эти реакции производят энергию, необходимую для поддержания гомеостаза среди других важных функций (например,грамм. метаболический оборот, деление клеток, сокращение и т. д.). Таким образом, клеточный метаболизм напрямую способствует процессам, связанным с ростом, воспроизводством, структурным поддержанием и т.д. клеточные компоненты.
Некоторые из ферментов, участвующих в этих реакциях, включают:
- Гексокиназа
- Пируваткиназа
- Фосфофруктокиназа
- Фосфоглицерат мутаза
- Фосфотриозо-изомераза
6
- Фосфотриозо-изомераза 14
- Фосфотриозо-изомераза 14
- Фосфотриозо-изомераза
-
Стадии клеточного метаболизма
Хотя расщепление глюкозы с образованием АТФ (химической энергии) в клеточном метаболизме является непрерывным процессом, его обычно делят на три основных этапа, которые включают:
1 / Гликолиз
Обычно гликолиз можно описать как окисление молекулы глюкозы (гексозы с шестью атомами углерода) с образованием двух молекул пирувата (каждая молекула пирувата состоит из трех атомов углерода).
Это включает в себя ряд важных шагов, которые будут подробно описаны ниже:
Учитывая, что молекулы глюкозы не могут просто диффундировать в клетку, они должны транспортироваться мембранными белками, известными как переносчики глюкозы (GLUT), через процесс известный как облегченная диффузия.
В то время как большинство этих транспортеров не инсулинозависимы, те, которые обнаруживаются в мышечных и жировых клетках, являются инсулинозависимыми.
Этап 1: Фосфорилирование 1 — Первый этап гликолиза, известный как фосфорилирование, происходит в цитоплазме клетки.Как следует из названия, фосфорилирование включает добавление фосфата к молекуле глюкозы, что позволяет перейти к следующему этапу.
Здесь фосфат, добавленный к молекуле сахара, образуется в результате распада АТФ в клетке, который не только преобразует АТФ в АДФ и фосфат, но также производит энергию, необходимую для добавления фосфата к молекуле сахара.
Этому процессу способствует фермент глюкокиназа в печени и поджелудочной железе, в то время как гексокиназа участвует в процессе в мышцах и превращает глюкозу в глюкозо-6-фосфат.
* Фосфорилирование глюкозы важно в том смысле, что оно создает отрицательный заряд на молекуле, который захватывает ее в клетке, таким образом предотвращая ее транспортировку из клетки.
Этап 2: Изомеризация глюкозо-6-фосфата в фруктозо-6-фосфат — Следующий этап гликолиза включает преобразование глюкозо-6-фосфата во фруктозо-6-фосфат. Хотя оба они состоят из шести (6) углеродных сахаров, фосфат фруктозы отличается от глюкозы, учитывая, что это кетон.
Углерод в карбонильной группе связан с двумя дополнительными атомами углерода, в то время как глюкозо-6-фосфат находится в форме альдегида — атом углерода в карбонильной группе связан с атомом водорода и атомом углерода.
Процесс изомеризации катализируется ферментом фосфогексозоизомеразой (также известной как фосфоглюкозоизомераза или глюкозо-6-фосфатизомераза) и требует наличия иона магния.
* Хотя фруктозо-6-фосфат можно превратить обратно в глюкозо-6-фосфат, следующий шаг необратим.По этой причине говорят, что молекула совершила гликолиз, когда перейдет на следующий этап.
Этап 3: Фосфорилирование 2 — На третьем этапе АТФ снова требуется для фосфорилирования фруктозо-6-фосфата с образованием 1,6-бисфосфата (дифосфата фруктозы). Как и в случае с первым фосфорилированием, АТФ снова расщепляется с образованием АДФ и фосфата.
Используя энергию, вырабатываемую АТФ, фермент фосфофруктокиназа типа 1 добавляет фосфат к фруктозо-6-фосфату, что приводит к образованию 1,6-бисфосфата.
* В отличие от фруктозо-6-фосфата, 1,6-бисфосфат содержит два фосфата, учитывая, что другой фосфат добавляется во время третьей стадии (фосфорилирование 2).
* Термин «бисфосфат» относится к факт, что между фосфатами есть атомы углерода.
Этап 4: Расщепление фруктозо-1,6-бисфосфата — Четвертый этап включает расщепление фруктозо-1,6-бисфосфата с образованием дигидроксиацетонфосфата (DHAP) и глицеральдегид-3-фосфата (GA3P). ).
Здесь фермент альдолаза участвует в расщеплении фруктозо-1,6-бисфосфата на две трехуглеродные молекулы. Хотя обе молекулы являются трехуглеродными, только одна из двух (глицеральдегид-3-фосфат) может перейти к следующему этапу гликолиза.
Стоит отметить, что одна молекула может трансформироваться / превращаться в другую (изомеризация). Фермент, ответственный за это, известен как триозофосфат-изомераза. В том случае, если организм имеет высокую потребность в энергии, большее количество молекул дигидроксиацетонфосфата преобразуется ферментом в глицеральдегид-3-фосфат, чтобы они могли перейти на следующий этап.
Шаг 5: Дегидрогенизация и фосфорилирование — На этой стадии гликолиза молекула глицеральдегид-3-фосфата превращается в 1,3-бис-фосфоглицерат под действием фермента глицеральдегид-3-фосфатдегидрогеназы.
В присутствии фермента НАД + (никотинамидадениндинуклеотид) реагирует с глицеральдегид-3-фосфатом (2 молекулы), в результате чего молекула (глицеральдегид-3-фосфат) теряет гидриды. В результате NAD + превращается в NADH.
* Учитывая, что дигидроксиацетонфосфат превращается в глицеральдегид-3-фосфат на четвертой стадии гликолиза, тогда две молекулы NAD + превращаются в 2 молекулы NADH после реакции 2NAD + с двумя молекулами глицеральдегида-3 -фосфат.
Помимо образования двух молекул НАДН, фермент глицеральдегид-3-фосфатдегидрогеназа также участвует в фосфорилировании двух молекул глицеральдегид-3-фосфата, что приводит к образованию 1,3-бис-фосфоглицерата. (в каждой из молекул по два фосфата).
Этап 6: Дефосфорилирование — На этом этапе 1,3-бис-фосфоглицерат подвергается дефосфорилированию с образованием 3-фосфоглицерата в присутствии фермента фосфоглицераткиназой.
На этом этапе две молекулы АДФ реагируют с двумя молекулами 1,3-бис-фосфоглицерата в присутствии фермента с образованием двух молекул АТФ и двух молекул глицеральдегид-3-фосфата.
Следовательно, фермент фосфоглицераткиназа играет важную роль в высвобождении фосфатов из двух молекул 1,3-бис-фосфоглицерата, что приводит к продукции / образованию двух молекул АТФ.
* На этапе 6 фермент фосфоглицераткиназа фосфорилирует АДФ с образованием АТФ. Это очень важный этап гликолиза, поскольку впервые на этой стадии клеточного метаболизма вырабатывается энергия в виде двух (2) молекул АТФ.
Этап 7: На следующем этапе фермент фосфоглицератмутаза превращает 3-фосфоглицерат в 2-фосфоглицерат, который, в свою очередь, превращается в фосфоенолпируват (PEP) ферментом энолазой.
Этап 8: Дефосфорилирование — На этом этапе фосфоенолпируват, образующийся на седьмой стадии, подвергается дефосфорилированию, что приводит к образованию двух (2) дополнительных молекул АТФ.
Здесь, в присутствии фермента пируваткиназы, две молекулы АДФ реагируют с двумя молекулами фосфат-пируват-фосфата, что приводит к дефосфорилированию двух молекул фосфат-пируват-фосфата. При фосфорилировании молекул АДФ образуются две молекулы АТФ и две молекулы пирувата.
В конце стадии 8 пируват может вступить в одну из двух реакций в зависимости от наличия или отсутствия кислорода. В анаэробных условиях (отсутствие кислорода) молекулы пирувата восстанавливаются в присутствии фермента лактатдегидрогеназы, что приводит к образованию молочной кислоты.
В этой реакции две молекулы НАДН окисляются с образованием двух (2) молекул НАД +. В печени молочная кислота может превращаться в глюкозу или использоваться для производства молекул АТФ.
Из-за кислотности молочной кислоты она может вызвать повышенную кислотность крови, вызывая падение pH. В случае, если клинический тест показывает повышение кислотности крови, это может указывать на такое состояние здоровья, как инфаркт миокарда.
В присутствии кислорода (аэробные условия) пируват переходит в следующую стадию клеточного метаболизма.
2 / Цикл Кребса (цикл лимонной кислоты или цикл трикарбоновой кислоты)В присутствии кислорода пируват входит в цикл Кребса, который является второй стадией клеточного метаболизма.Однако, прежде чем он действительно войдет в этот этап, он должен пройти этап перехода, также известный как подготовительный этап.
В аэробных условиях молекулы пирувата не превращаются в молочную кислоту и поэтому могут проникать в митохондрии, где они могут пройти важный этап перехода.
Декарбоксилирование — Эта переходная стадия известна как декарбоксилирование и включает превращение молекул пирувата в ацетил-КоА ферментом пируватдегидрогеназой.
Как следует из названия, этот этап включает удаление углерода (формы CO2) из пирувата ферментом пируватдегидрогеназой.
Фермент добавляет коэнзим А к 2 молекулам пирувата в присутствии НАД +, что не только приводит к образованию 2 ацетил-КоА, но также превращает молекулы НАД + в 2 молекулы НАДН.
Следующие основные этапы цикла Креба:
Этап 1: Синтез цитрата — На первом этапе цикла Креба ацетил-КоА, полученный на переходной стадии, соединяется с оксалоацетатом. (OAA) в присутствии фермента цитрат-синтазы с образованием цитрата.
Как следует из названия, фермент участвует в синтезе цитрата путем объединения ацетил-КоА, который представляет собой молекулу из двух (2) атомов углерода, и оксалоацетата, молекулу из четырех атомов углерода.
* Шаг 1 цикла Креба строго регулируется. Некоторые молекулы, регулирующие функцию фермента цитрат-синтазы, включают АТФ, НАДН и цитрат. Когда имеется большое количество цитрата (молекулы, синтезируемой ферментом), он посылает обратную связь, ограничивая его активность.
Этап 2: Изомеризация — Второй этап представляет собой реакцию изомеризации и приводит к получению изоцитрата. Здесь фермент аконитаза превращает цитрат в изоцитрат путем перегруппировки молекул углерода.
Здесь стоит отметить, что процесс обратим, что означает, что изоцитрат может быть преобразован обратно в цитрат, если это необходимо.
* Изоцитрат, полученный изомеризацией цитрата, менее стабилен по сравнению с цитратом.
Этап 3: Декарбоксилирование — На этапе 3 изоцитрат превращается в альфа-кетоглутарат посредством процесса, известного как декарбоксилирование. Как следует из названия, это включало удаление углерода из изоцитрата в виде диоксида углерода.
В процессе NAD + восстанавливается до NADH и иона водорода. Этот процесс катализируется ферментом изоцитратдегидрогеназой. На этом этапе НАД + реагирует с изоцитратом в присутствии фермента, который восстанавливает НАД +, превращая изоцитрат в альфа-кетоглутарат.
В присутствии слишком большого количества АТФ фермент, участвующий в этой реакции, ограничен, что снижает производство альфа-кетоглутарата. Однако большое количество АДФ способствует действию фермента, усиливая его активность.
Этап 4: Декарбоксилирование 2 — На этапе 4 альфа-кетоглутарат превращается в сукцинил-КоА ферментом альфа-кетоглутаратдегидрогеназой. В этой реакции НАД + реагирует с альфа-кетоглутаратом в присутствии фермента альфа-кетоглутаратдегидрогеназы, что снова приводит к его восстановлению.
Углерод также теряется в виде диоксида углерода, что приводит к образованию сукцинил-КоА. В случае, когда в клетке вырабатывается слишком много энергии, молекула сукцинил-КоА связывается с ферментом, ограничивая его активность. В результате производство сукцинил-КоА снижается. Некоторые из других веществ, которые ингибируют фермент, включают НАДН и кальций.
Этап 5: Гидролиз — На этапе 4 добавляется КоА, что приводит к получению сукцинил-КоА.С другой стороны, на этапе 5 КоА удаляют, что приводит к образованию сукцината.
Фермент, участвующий в этом этапе, известен как сукцинил-КоА-синтетаза, и действует, стимулируя превращение сукцинил-КоА в сукцинат. В этом процессе КоА высвобождается вместе с фосфатом.
Здесь молекула GDP (гуанозиндифосфата) поглощает фосфат с образованием GTP (гуанозинтрифосфата). Однако ГТФ теряет фосфат в пользу АДФ, что приводит к производству АТФ.Это известно как фосфорилирование на уровне субстрата.
* На стадии 5 гидролиз ГТФ дает АТФ.
Этап 6: Фумарат — На шестом (6) этапе цикла сукцинат превращается в фумарат ферментом сукцинатдегидрогеназой. Здесь FAD (флавинадениндинуклеотид) реагирует с сукцинатом в присутствии фермента, что приводит к его восстановлению до FADH.
Этап 7: Гидролиз — На этапе 7 фермент фумараза участвует в гидролизе фумарата с образованием малата.
Этап 8: Производство оксалоацетата — На этом этапе фермент малатдегидрогеназа участвует в превращении малата в оксалоацетат. Здесь НАД + реагирует с малатом в присутствии фермента, что приводит к его восстановлению до НАДН и иона водорода. Как только он произведен, оксалоацетат может войти в цикл, приняв другую молекулу ацетил-КоА по мере продолжения цикла.
* С двумя молекулами ацетил-КоА цикл Креба производит 4 молекулы углекислого газа, 6 молекул НАДН, 2 молекулы FADh3, а также 2 молекулы АТФ.Молекулы NADH и FADh3 важны для третьей и последней стадии клеточного метаболизма.
3 / Электронная транспортная система (Электронная транспортная цепь)
Электронная транспортная система / цепь является третьей и последней стадией клеточного метаболизма и происходит в складчатой внутренней мембране митохондрий (кристах). Это особенно важный этап, учитывая, что здесь вырабатывается большая часть молекул АТФ.
Этот этап также включает несколько важных этапов переноса электрона, которые включают:
Этап 1: На первом этапе системы переноса электронов молекулы НАДН контактируют с ферментным комплексом 1, который забирает электроны у этих молекул. таким образом превращая их из НАДН в НАД + (6 молекул).
В отличие от NADH, FADH имеет более высокое сродство к ферментному комплексу 2 и, таким образом, реагирует с ферментом, высвобождая два электрона (2 молекулы FADH высвобождают электроны в комплекс 2). Это превращает 2 молекулы FADH в 2 молекулы FAD и протонов.
Этап 2: На этапе 2 клеточного метаболизма комплекс 1 высвобождает электроны, полученные от НАДН, так что он может перейти с высокого уровня энергии на низкий уровень энергии. Это также приводит к открытию пор в комплексе, позволяя выкачивать протоны.
В то время как комплекс 2 также высвобождает электроны, чтобы переключиться на более низкое энергетическое состояние, он не имеет пор, и поэтому протоны не могут быть откачаны. На этапе 2 все электроны, высвобождаемые комплексом 1 и 2, захватываются коэнзимом Q (также известным как убихинон).
Шаг 3: Одной из определяющих характеристик коэнзима Q является то, что он подвижен и, следовательно, может перемещаться внутри крист. Это важно на этом этапе, так как позволяет молекуле перемещаться для передачи электронов.На этом этапе молекула перемещается и передает свои электроны ферментному комплексу 3.
Этап 4: Как и другие ферментные комплексы в этой системе, ферментный комплекс 3 также переходит на более высокий энергетический уровень, получив электроны. В этом состоянии он должен освободить эти электроны, чтобы вернуться на более низкий уровень энергии.
Опять же, электроны переносятся на другую мобильную молекулу, известную как цитохром C. Подобно ферментному комплексу 1, этот комплекс также имеет пору, которая открывается после высвобождения электронов, что позволяет протонам перекачиваться в межмембранное пространство.
Этап 5: На этапе 5 процесс снова повторяется с цитохромом C, высвобождающим электроны, чтобы переключиться на более низкий уровень энергии. Эти электроны затем принимаются ферментным комплексом 4.
Этап 6: На этапе 6 ферментный комплекс 4 высвобождает полученные электроны, таким образом переходя с высокого уровня энергии на более низкий уровень энергии. Однако здесь высвободившиеся электроны соединяются с кислородом и протонами, образуя воду. Высвобождение электронов здесь также приводит к открытию поры, что позволяет выкачивать протоны.
* До этого этапа очевидно, что комплексы (а также другие вовлеченные молекулы) получают и высвобождают электроны, создавая цепочку передачи электронов.
Здесь комплексы переключаются на более высокое энергетическое состояние каждый раз, когда они получают электроны, и должны высвободить эти электроны, чтобы переключиться обратно на более стабильный, более низкий энергетический уровень.
В результате высвобождения протонов в межмембранное пространство происходит высокая аккумуляция / концентрация этих электронов в этом пространстве по сравнению с комплексом митохондрий.
Из-за этой разницы создается градиент, который означает, что они перемещаются из области высокой концентрации протонов (в межмембранном пространстве) в область с более низкой концентрацией протонов (матрица митохондрий). Для этого протоны проходят через специальную структуру, известную как АТФ-синтаза.
Здесь протоны проходят через статорную часть структуры, что позволяет им двигаться к матрице митохондрий. Когда они проходят, другая часть конструкции, известная как ротор, начинает вращаться.Когда он вращается с движением протонов в матрицу, ротор начинает поглощать потенциальную энергию.
Затем эта энергия используется другой частью структуры АТФ-синтазы (каталитической ручкой) для создания молекул АТФ из фосфатов и АДФ. АДФ и фосфаты содержатся в каталитической ручке и используются для производства АТФ, когда потенциальная энергия поглощается в роторе.
Здесь процесс, используемый для производства АТФ из потенциальной энергии, создаваемой движением протонов, известен как окислительное фосфорилирование.
* В аэробных условиях три стадии клеточного метаболизма производят в общей сложности 36 молекул АТФ. Анаэробные условия приводят, в частности, к образованию 2 молекул АТФ в результате гликолиза.
Здесь можно узнать больше о делении клеток, дифференцировке клеток, пролиферации клеток и пентозофосфатном пути
См. Статьи о культуре клеток, окрашивании клеток и окрашивании по Граму.
В чем разница между растительной клеткой и животной клеткой?
Ознакомьтесь с информацией по теории клеток.
Почему так важна адгезия клеток?
Возвращение из того, что такое клеточный метаболизм? на главную страницу MicroscopeMaster
Ссылки
Кристиан М. Металло и Мэтью Г. Вандер Хайден. (2014). Понимание регуляции метаболизма и ее влияния на физиологию клетки.
Дональд С. Риццо. (2000). Основы анатомии и физиологии.
Jones and Bartlett Publishers. Глава 7: Метаболизм.
Майкл Палмер.(2019). Метаболизм человека. Конспект лекций.
Ссылки
https://www.