Катаболизм углеводов
Катаболизм
глюкозы. Аэробный, анаэробный гликолиз.
Гликолиз
– процесс окисления глюкозы в результате
которого происходит расщепление глюкозы
с образование 2 молекул пирувата (аэробный
гликолиз) или 2 молекул лактата (анаэробный
гликолиз). Аэробный и анаэробный
гликолитический путь начинается с
фосфорилирования глюкозы. Во многих
тканях эту реакцию катализирует
гексокиназа (Км<0,1мМоль/л),
а в печени эту реакцию может катализировать
и глюкокиназа (Км<12мМоль/л).
Образование глюкозо-6-фосфата — своеобразная
«ловушка» для глюкозы.
В
аэробном и анаэробном гликолизе можно
выделить 2 стадии.
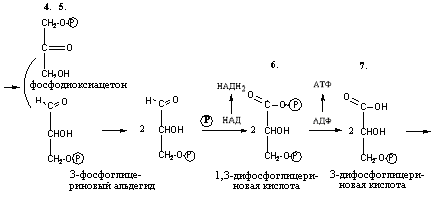
На стадии а) глюкоза
распадается с образованием 2 фосфотриоз
(фосфоглицериновый альдегид и
фосфодиоксиацетон). Эти реакции протекают
потреблением двух молекул АТФ.
Стадия б) аэробного
гликолиза — обеспечивает окисление
фосфотриоз до пировиноградной кислоты
и образование АТФ по механизму
субстратного фосфорилирования. В ходе
реакций происходит дегидрирование
различных метаболитов и восстановление
НАД и НАДФ до НАДН2. Кислород
используется клетками (митохондриями)
значительно позже — на стадии окисления
НАДН2в цепи биологического
окисления. Это приводит к образованию
АТФ по механизму окислительного
фосфорилирования. Все этапы гликолитического
пути окисления глюкозы происходят в
цитозоле. Мембрана митохондрий не
проницаема для водорода. Водород
транспортируется в митохондрии с помощью
челночного механизма. Существует два
вида челночных механизмов:
1) глицеро-фосфатный
челночный механизм (функционирует в
клетках скелетных мышц и гепатоцитах
печени).
2) малат-аспартатный
челночный механизм (функционирует в
клетках сердечных мышц)
Обе челночные
системы существенно отличаются по
количеству синтезируемого АТФ. В первой
системе соотношение Р/О=2, т.к водород
вводится в ЦПЭ на уровне КоQ.
Вторая система энергетически более
эффективна, т.к. передает водород в ЦПЭ
через митохондриальный НАД и соотношение
Р/О=3.
Стадия
в анаэробного гликолиза отличается
от аэробного тем, что НАДН2
(восстановленныйкофактор
фермента глицерофосфатдегидрогеназы)
окисляется не кислородом, а за счет
переноса водорода ферментом
лактатдегидрогеназой на пировиноградную
кислоту с образованием лактата. Движущей
силой анаэробного гликолиза является
гликолитическая оксидоредукция.
Гликолитическая оксидоредукция
обеспечивает возможность продолжения
гликолиза и образования энергии при
недостаточном поступлении кислорода
к органам и тканям по механизму только
субстратного фосфорилирования. Значение
реакции восстановления пирувата
заключается не в образовании лактата,
а в том, что данная реакция обеспечивает
регенерацию НАД. Анаэробный распад
глюкозы происходит в мышцах в первые
минуты работы, в эритроцитах, в которых
нет митохондрий, а также в различных
органах при недостаточном снабжении
их кислородом (гипоксия). При достаточном
поступлении О2происходит
подавление анаэробного гликолиза,
концентрация лактата снижается, а
окисление глюкозы переключается на
более энергетически выгодный аэробный
путь окисления.
Пируват, образовавшийся
в аэробном гликолизе окисляется в общем
пути катаболизма – в процессе
окислительного декарбоксилирования,
а полученный ацетил-КоА окисляется в
цикле трикарбоновых кислот.
Баланс энергии
окисления одной молекулы глюкозы.
|
Этапы |
Количество |
Количество |
|
I. Глюкоза→2пируват II. 2 III. 2 Суммарный |
-2 — |
+10 +6 +24 +38 |
|
Этапы |
Количество |
Количество |
|
Гликолиз: Глюкоза→2фосфотриозы Гликолиз: 2фосфотриозы Суммарный |
-2 |
+4 +2 |
Регуляция
катаболизма глюкозы.
Значение гликолиза
состоит в синтезе АТФ. Показателем
потребления АТФ является накопление
АДФ и АМФ. Отношение уровня АТФ к АМФ и
АДФ характеризует энергетический статус
клетки. Три реакции гликолиза катализируемые
гексокиназой (или глюкокиназой),
фосфофруктокиназой и пируваткиназой
необратимы. Существенное значение для
регуляции гликолиза имеет изменение
активности фермента фосфофруктокиназы,
т.к. этот фермент катализирует наиболее
медленную реакцию гликолиза.
Фосфофруктокиназа аллостерически
активируется АМФ, но ингибируется АТФ.
При высоком уровне АТФ снижается скорость
ЦТК и ЦПЭ, в этих условиях процесс
гликолиза замедляется. На скорость
катаболизма глюкозы влияет скорость
регенерации НАДН2в дыхательной
цепи.
Глюконеогенез,
цикл Кори. Пентозофосфатный путь
Пентозофосфатный
путь превращения глюкозы (ПФП).
ПФП называют также апотомическим
(прямым) путем окисления глюкозы или
гексомонофосфатным шунтом. ПФП состоит
из двух стадий: окислительной и
неокислительной.
studfile.net
Катаболизм углеводов
Основными источниками углерода и энергии, а значит, и пищевыми субстратами для большинства организмов (исключая растения) служат углеводы. Среди них на планете в наибольшей мере распространены целлюлоза, ее производные и крахмал. Кроме этих полисахаридов, большое значение имеют гликоген, инулин, хитин, ксиланы, пектиновые вещества и др. Большинство перечисленных полисахаридов расщепляется при участии специфических ферментов на моносахариды, среди которых преобладают гексозы и пентозы. Зачастую расщепление поли- и олигосахаридов осуществляется при участии ферментов фосфорилаз, и тогда образованные продукты оказываются фосфорилированными.
Особенностью сахаров является наличие при каждом атоме углерода атома кислорода, что делает возможным химическую атаку этих субстратов практически в любой точке молекулы. Кроме того, моносахариды и в первую очередь их фосфорилированные формы способны к изомеризации: карбонильные группы, атомы водорода могут легко перемещаться в соседнее положение или изменять свое пространственное положение в молекуле с помощью изомераз. Таким образом, появляется возможность перехода от любой гексозы или пентозы к любой другой, изомерной ей. По этой причине, несмотря на многообразие и сложность процессов обмена углеводов, можно выделить несколько типичных путей их превращения, в частности катаболизма, имеющих выраженные отличительные особенности. Такими путями служат: гликолиз, пентозофосфатные пути и путь Энтнера—Дудорова.
К закономерностям катаболизма моносахаридов относится обязательная начальная стадия активации свободных моноз, которая осуществляется в ходе фосфорилирования. В результате образуются фосфорные эфиры моносахаридов, способные вступать в дальнейшие превращения.
Гликолиз. Этот способ катаболизма сахаров называют иначе фруктозо-1,6-дифосфатным путем (по названию ключевого соединения) или путем Эмбдена—Мейергофа—Парнаса (по именам его исследователей).
Гликолиз считается наиболее универсальным и самым выгодным с энергетической точки зрения путем катаболизма гексоз. Процесс открыт в 1897 г. братьями Бухнерами, и его название происходит от двух греческих корней: glicos — сахар и lysis — растворять. В ходе гликолиза происходит не требующее участия молекулярного кислорода многоэтапное превращение гексоз в пируват, что сопровождается образованием АТР и восстановительных эквивалентов.
Реакции гликолитического пути осуществляются в цитозоле. Все промежуточные соединения имеют фосфорилированную форму. Для запасания энергии используется механизм субстратного фосфорилирования.
Превращение глюкозы в пируват (рис. 9.3) требует участия десяти ферментов и осуществляется в ходе следующих стадий: подготовка к разрыву гексозной цепи, разрыв цепи и образование глицеральдегид-3-фосфата, первое и второе субстратное фосфорилирование.
Процесс начинается с фосфорилирования глюкозы (активация молекулы) с участием АТР (донор фосфатной группы) и фермента гексокиназы. Образующийся глюкозо-6-фосфат в следующей реакции изомеризуется глюкозофосфатизомеразой во фруктозо-6-фосфат, который претерпевает второе фосфорилирование за счет АТР с образованием фруктозо-1,6-дифосфата.
Фруктозо-1,6-дифосфат является ключевым промежуточным продуктом гликолитического пути: именно данное соединение отличает гликолиз от других путей катаболизма сахаров. Кроме того, на уровне этого вещества осуществляется регуляция скорости всего процесса гликолиза. Активность катализирующего данную реакцию фермента (фосфо-фруктокиназы) ингибируется высокими концентрациями АТР, при этом снижается сродство фосфофруктокиназы к субстрату — фруктозо-6-фосфату. Кроме того, фосфофруктокиназа ингибируется цитратом —ранним промежуточным продуктом цикла трикарбоновых кислот. Известна и активация фосфофруктокиназы: ее осуществляют ADP и неорганический фосфат. Таким образом, фосфофруктокиназа наиболее активна в условиях, когда в клетке мало АТР (много ADP) и недостает строительных блоков.
На следующем этапе гликолиза фруктозодифосфат расщепляется с участием фруктозодифосфатальдолазы на два триозофосфата: дигидроксиацетонфосфат и глицеральдегидтрифосфат. Эти продукты представляют собой изомеры и легко переходят один в другой под действием триозофосфатизомеразы. Однако направление этой реакции сдвинуто в сторону образования глицеральдегид-3-фосфата, поскольку он все время выводится из реакционной смеси, претерпевая дальнейшие превращения. Глицеральдегид-3-фосфат служит субстратом первого субстратного фосфорилирования.
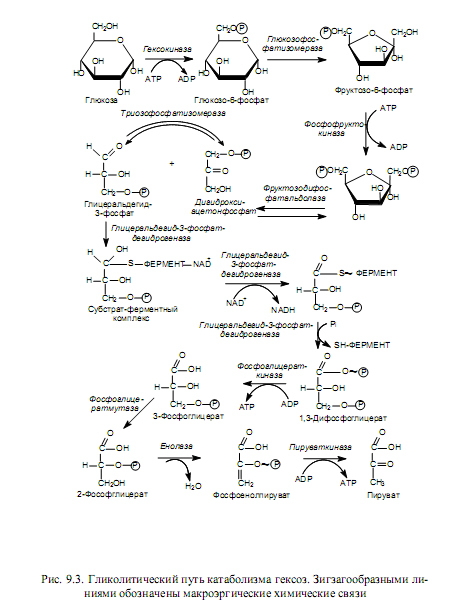
В ходе оставшихся этапов гликолиза, в отличие от предыдущих, энергия выделяется и запасается в форме АТР. Одной из подобных реакций является окисление глицеральдегид-3-фосфата. Фермент, катализирующий данную реакцию (глицеральдегид-3-фосфатдегидрогеназа), использует в качестве кофермента NAD и характеризуется содержанием большого количества сульфгидрильных групп (SH-групп). Окисление начинается со связывания глицеральдегид-3-фосфата с SH-группой фермента — формируется субстратферментный комплекс. Далее фермент катализирует перенос водорода с субстрата на NAD+, и восстановленный кофермент отделяется. Реакция дегидрирования и служит окислительной реакцией, поставляющей энергию: возникает комплекс фермента с ацильным остатком (тиоэфир), богатый энергией. Затем осуществляется фосфоролиз — перенос остатка глицеральдегид-3-фосфата совместно с макроэргической связью на фосфорную кислоту, что приводит к образованию 1,3-дифосфоглицериновой кислоты и исходной формы фермента. 1,3-Дифосфоглицерат представляет собой смешанный ангидрид фосфорной и карбоновой кислот и имеет высокий потенциал переноса фосфатной группы. Богатая энергией фосфатная группа переносится с 1,3-дифосфоглицерата на ADP с участием фосфоглицераткиназы и образованием продуктов: 3-фосфоглицерата и АТР. Так осуществляется первое субстратное фосфорилирование в гликолизе.
Заключительный этап гликолиза — второе субстратное фосфорилирование — начинается со внутримолекулярной перестройки, в ходе которой 3-фосфоглицерат изомеризуется в 2-фосфоглицерат (фермент фосфоглицератмутаза). 2-Фосфоглицерат дегидратируется в фосфоенолпируват при участии енолазы. В результате этой реакции образуется соединение, характеризующееся высоким потенциалом переноса фосфатной группы, таким образом, отщепление молекулы воды от 2-фосфоглицерата сопровождается перераспределением энергии внутри молекулы, и фосфатная связь у 2 атома углерода превращается из низко- в высокоэнергетическую. Последняя реакция гликолиза катализируется пируваткиназой, и в ходе нее фосфатная группа переносится на молекулу ADP и образуется конечный продукт пути — пируват.
Гликолитическое расщепление одной молекулы глюкозы приводит к образованию 4 молекул АТР (по две на каждую молекулу глицеральдегид-3-фосфата), из которых 2 расходуются на образование фруктозодифосфата. Таким образом, запасается всего 2 молекулы АТР. Кроме того, на одну молекулу глюкозы в этом процессе запасается 2 молекулы NADH (при окислении 2 молекул глицеральдегид-3-фосфата). Баланс гликолиза выглядит следующим образом:
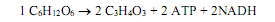
Гликолитический процесс служит клетке для запасания энергии и восстановительных эквивалентов, а также является поставщиком «строительных блоков» в виде трехуглеродного, частично окисленного соединения —пировиноградной кислоты и некоторых промежуточных продуктов, в частности глицеральдегид-3-фосфата.
Пентозофосфатные пути. Эти пути катаболизма сахаров довольно разнообразны. В зависимости от условий и видовой принадлежности организма результатом данных процессов могут быть различные вещества. Осуществление реакций пентозофосфатных путей в обратном направлении используется клетками при фиксации СО2. Основной отличительной особенностью путей является образование в качестве промежуточных соединений пятиуглеродных сахаров, а также последовательное отщепление от 6-углеродного фосфорилированного сахара по одному атому углерода, который высвобождается в среду в виде СО2.
Пентозофосфатные пути (иначе, схема Варбурга—Диккенса—Хореккера, гексозомонофосфатный шунт, фосфоглюконатный путь) реализуются организмами реже, чем гликолиз, и обнаруживаются чаще всего в клетках бактерий.
После традиционной реакции активации глюкозы в ходе фосфорилирования образованный глюкозо-6-фосфат подвергается дегидрированию. Данную реакцию катализирует глюкозо-6-фосфатдегидрогеназа, а восстановительные эквиваленты акцептирует ее кофермент — NADP+. Образуется 6-фосфоглюконолактон (рис. 9.4). Данное соединение подвергается гидролизу (раскрытие кольца) с участием глюконолактоназы, а образованная 6-фосфоглюконовая кислота претерпевает второе дегидрирование (фосфоглюконатдегидрогеназа), после которого сразу же следует декарбоксилирование и образуется рибулозо-5-фосфат. Рибулозо-5-фосфат изомеризуется в два пятиуглеродных сахара — ксилулозо-5-фосфат и рибозо-5-фосфат (рис. 9.4).
Следующий этап пентозофосфатных путей представляет собой многократные межмолекулярные перестройки и изомеризацию промежуточных соединений, формирующихся из рибулозо-5-фосфата. Эти реакции направлены на образование 6-углеродного соединения (глюкозо-6-фосфата), которое может снова подвергнуться окислению и декарбоксилированию. В результате из 6 молекул рибулозо-5-фосфата получается 5 молекул глюкозо-6-фосфата (рис.9.5).
Система структурной перестройки сахаров включает два фермента —транскетолазу и трансальдолазу, которые катализируют перенос двухуглеродных и трехуглеродных фрагментов, разрывая С—С-связи в двух положениях: по соседству с карбонильной (a) и у соседнего с карбонильной группой углеродного атома (b) (на рис. 9.5 места, в которых связи подвергаются расщеплению, обозначены пунктирной линией).
При полном окислении глюкозы в пентозофосфатных путях молекула глицеральдегид-3-фосфата, образующаяся на конечном этапе, подвергается изомеризации с участием триозофосфатизомеразы в дигидроксиацетонфосфат, который затем вступает в реакцию альдольной конденсации со второй молекулой глицеральдегид-3-фосфата, и образуется фруктозодифосфат. Данная реакция является обратимой процессу расщепления фруктозодифосфата, которое имеет место в гликолизе (рис. 9.3), а фермент фруктозодифосфатальдолаза катализирует и прямое, и обратное превращения. Молекула фруктозо-1,6-дифосфата дефосфорилируется с образованием фруктозо-6-фосфата (фермент фосфатаза), а он, в свою очередь, изомеризуется в глюкозо-6-фосфат и возвращается к начальному этапу дегидрирования.
Таким образом, на одну молекулу глюкозы при полном окислении в пентозофосфатных путях затрачивается 1 молекула АТР и образуется 6 молекул СО2 и 12 молекул NADPH. Таким образом, основной задачей пентозофосфатных путей является обеспечение клетки NADPH, который используется в процессах биосинтеза.
В ходе пентозофосфатных путей может и не происходить полного окисления глюкозы. В этом случае глицеральдегид-3-фосфат претерпевает иные превращения, например подвергается реакциям субстратного фосфорилирования под действием ферментов гликолитического пути.
Большое значение пентозофосфатные пути имеют для биосинтетических процессов: здесь образуются промежуточные соединения, играющие роль предшественников многих веществ. В частности, эритрозо-4-фосфат используется бактериями и растениями для синтеза ароматических ами-


нокислот. Рибозо-5-фосфат служит субстратом для синтеза азотистых оснований и некоторых аминокислот.
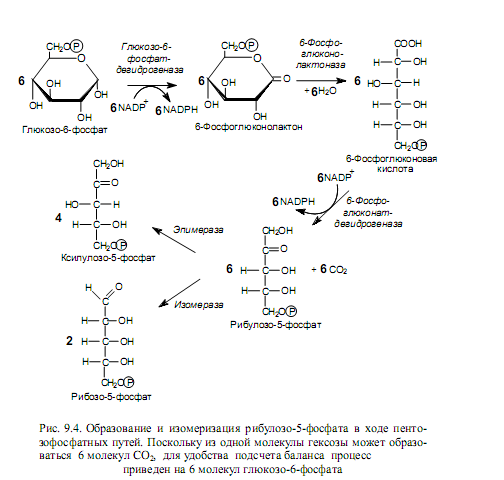
Путь Энтнера—Дудорова (2-кето-3-дезокси-6-фосфоглюконатный). Этот процесс является еще одним способом катаболизма гексоз и представлен в основном в клетках микроорганизмов. Особое значение данный путь имеет для расщепления глюконата. Начальные стадии превращения глюкозо-6-фосфата совпадают с таковыми для пентозофосфатных путей, вплоть до образования 6-фосфоглюконовой кислоты. Далее, однако, 6-фофоглюконат подвергается а не окислению, а реакции дегидратации, с участием фермента 6-фосфоглюконат-дегидратазы. Образуется ключевое соединение данного пути — 2-кето-3-дезокси-6-фосфоглюко-нат. Стадия дегидратации осуществляется через образование промежуточного соединения (енола), который в результате таутомерного превращения переходит в 2-кето-3-дезокси-6-фосфоглюконат (рис. 9.6).
Далее альдолаза катализирует расщепление ключевого соединения на пируват и глицеральдегид-3-фосфат. Последний может вступать в гликолитический путь и подвергаться дальнейшим превращениям.
Как следует из баланса пути Энтнера—Дудорова, этот процесс менее выгоден с энергетической точки зрения, чем гликолиз: на молекулу глюкозы здесь запасается только 1 молекула АТР (из двух синтезированных одна тратится на фосфорилирование глюкозы).
Обзор основных катаболических путей позволяет увидеть, что расщепление «топливных» молекул при их неполном окислении сопровождается образованием двух основных метаболитов — пировиноградной кислоты и ацетил СоА. Данные вещества могут использоваться как в биосинтетических путях в качестве «строительных блоков», так и подвергаться дальнейшим превращениям, обеспечивающим запасание клетками энергии. Поскольку в данном разделе обсуждаются процессы, приводящие к запасанию энергии, следует отметить, что судьба пирувата и ацетил-СоА зависит в первую очередь от наличия молекулярного кислорода в клетке, а также от ее ферментативного оснащения. Так, в клетках анаэробных микроорганизмов и тканей высших организмов пируват и ацетил-СоА могут подвергаться различного рода брожениям, завершающим процессы катаболического расщепления субстратов. В аэробных и факультативно-анаэробных клетках при наличии молекулярного кислорода пируват может подвергаться процессу окислительного декарбоксилирования, и образующийся ацетил-СоА вступает в цикл трикарбоновых кислот, где осуществляется его полное окисление до СО2 и Н2О, а формирующиеся восстановительные эквиваленты поступают в дыхательную цепь. Перечисленные процессы будут рассмотрены в следующих главах.
Глава 10. БРОЖЕНИЕ
Брожение со времен Пастера называют «Жизнью без воздуха». Этот тип метаболизма возник на нашей планете одним из самых первых — еще в то время, когда в атмосфере отсутствовал молекулярный кислород. Наиболее древние обитатели Земли использовали этот самый примитивный и наименее выгодный с энергетической точки зрения способ метаболизма для своей жизнедеятельности. В ходе эволюции возникли новые, более перспективные процессы запасания энергии, однако брожение по прежнему используется многими организмами, поскольку позволяет им осуществлять жизнедеятельность в условиях отсутствия молекулярного кислорода.
Брожение происходит в клетках бактерий, дрожжей (для этих микроорганизмов брожение является преобладающим типом метаболизма), простейших, моллюсков, в некоторых тканях рыб и птиц, а также высших животных, включая человека (мышцы, хрусталик и роговица глаза).
Известно большое разнообразие процессов брожения, которые отличаются друг от друга субстратами, продуктами, а также химическими превращениями. При этом можно различить несколько типов брожения, встречающихся наиболее часто: спиртовое, молочнокислое, пропионовокислое, муравьинокислое, маслянокислое, ацетоно-бутиловое. Свое название эти типы брожения получили от доминирующих или характерных продуктов, которые образуются в результате определенной последовательности реакций. Нетрудно заметить, что среди продуктов преобладают органические кислоты и спирты. Субстратами для разных типов брожения служат также органические вещества — в большинстве случаев пируват, а также лактат, ацетил-СоА, некоторые промежуточные метаболиты гликолитического пути, аминокислоты. Таким образом, брожение можно определить как катаболический процесс, происходящий в условиях, не требующих участия молекулярного кислорода, в котором и донорами, и акцепторами электронов являются органические вещества.
Реакциям брожения всегда предшествуют предварительные этапы катаболизма субстратов: чаще гликолиз, реже — пентозофосфатные и фосфоглюконатный пути. Поэтому часто реакции собственно брожения рассматривают в совокупности с предшествующими этапами, и в этом случае можно говорить о запасании энергии в ходе таких процессов. Однако следует помнить, что энергия в форме макроэргических связей АТР запасается в процессе субстратного фосфорилирования, как правило, еще до образования пирувата. В процессах собственно брожения энергия запасается редко, и можно назвать всего три реакции, в которых фосфорилируется ADP: образование масляной, уксусной, а также ацетоуксусной кислот. В брожениях других типов дополнительного количества энергии не образуется, и их основной задачей является регенерация окисленной формы переносчика восстановительных эквивалентов — NAD+. Это необходимо для бесперебойного осуществления реакций гликолиза и других катаболических путей, в ходе которых, как известно, происходит расщепление субстратов, сопровождающееся их окислением, что требует постоянного присутствия в клетке акцепторов восстановительных эквивалентов — NAD+.
Рассмотрев более детально несколько типов брожения, можно убедиться в том, что соблюдается эквивалентность между количеством молекул NADН, образованных в ходе гликолиза, и числом молекул NAD+, сформировавшихся в ходе собственно брожения (при условии, что в метаболизме не принимает участие молекулярный кислород). Таким образом, в анаэробных условиях использующие бродильный тип метаболизма клетки вынуждены тратить существенную часть двух-, трехуглеродных органических веществ для акцептирования водорода, выводя их затем из клетки. Этот крайне непроизводительный процесс приводит к тому, что в анаэробных условиях клетки перерабатывают огромные количества субстрата, чтобы извлечь нужное для жизнедеятельности количество энергии. Неслучайно у многих факультативноанаэробных микроорганизмов выработался механизм подавления процессов брожения в аэробных условиях с переключением на дыхательный метаболизм. При дыхании из одной молекулы глюкозы клетка может извлечь максимально 38 молекул АТР, а при брожении — только 4 (в большинстве случаев всего 2).
Спиртовое брожение
Преобладающим продуктом спиртового брожения является этиловый спирт. Этот тип брожения характерен для определенных видов дрожжей, бактерий, а также мицелиальных грибов. Известно 3 основных способа спиртового брожения, которые отличаются друг от друга тем, какой из трех катаболических путей расщепления глюкозы (и некоторых других сахаров) реализуется: гликолиз, пентозофосфатные пути либо путь Энтнера—Дудорова. Наиболее часто, как отмечалось выше, клетки используют гликолиз, и его конечный продукт — пируват чаще других служит субстратом спиртового брожения.
На рис. 10.1 представлены схемы двух способов спиртового брожения. Первый способ предполагает катаболизм глюкозы по гликолитическому пути (характерен для дрожжей и бактерий Sarcina ventriculi), а второй — по пентозофосфатному (гетероферментативные молочнокислые бактерии). Соответственно различаются участвующие в этих процессах ферменты и спектр продуктов. Третий способ спиртового брожения, который используют бактерии Zymomonas mobilis, отличается тем, что глюкоза катаболизируется по пути Энтнера—Дудорова. Образующийся пируват подвергается декарбоксилированию, как и в первом случае, и ступенчато восстанавливается в этанол. Часть пирувата восстанавливается в молочную кислоту.
На рис. 10.1 наглядно продемонстрировано, что количество затраченных в ходе гликолиза и пентозофосфатного пути молекул NAD+ (NADP+) строго соответствует количеству регенерированных в процессе спиртового брожения молекул NAD+ (NADP+). При этом реакции субстратного фосфорилирования, приводящие к синтезу АТР, имеют место только в катаболических путях до образования пирувата. В реакциях собственно спиртового брожения, связанных с преобразованием пирувата в этанол или лактат, запасание энергии не происходит.
Ход брожения очень сильно зависит от условий, в которых функционируют клетки. Кроме того, у некоторых микроорганизмов существуют отдельные отклонения от приведенных выше классических схем спиртового брожения. В результате спектр продуктов брожения может сильно различаться и включать кроме этанола и лактата примеси: глицерол, ацетальдегид, другие альдегиды, эфиры, кетоны, спирты, в том числе высшие органические кислоты и др.
Похожие статьи:
poznayka.org
15.16.Катаболизм основных пищевых веществ — углеводов, жиров, белков. Понятие о специфических путях катаболизма и общих путях катаболизма.
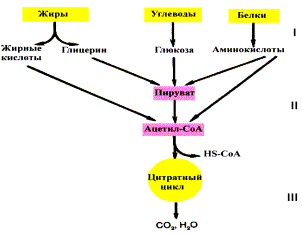
Если
процесс катаболизма рассматривать с
общей точки зрения, то можно выделить
три основные его части:
-
Расщепление в
пищеварительном тракте.
Это гидролитические реакции, превращающие
сложные пищевые вещества в относительно
небольшое число простых метаболитов:
глюкоза, аминокислоты, глицерин, жирные
кислоты. -
Специфические пути
катаболизма.
Простые метаболиты подвергаются
специфическим реакциям расщепления,
в результате которых образуется либо
пировиноградная кислота, либо ацетил
— КоА. Причем ацетил — КоА может
образоваться из пирувата в результате
окислительного декарбоксилирования.
Могут также образоваться другие
соединения, непосредственно включающиеся
в цитратный цикл. -
Цитратный цикл и
дыхательная цепь завершают
расщепление пищевых веществ до конечных
продуктов — СО2 и
Н2О.
Следовательно, начиная со
стадии образования пирувата происходит
унификация путей катаболизма. Из большого
числа исходных соединений образуется
всего два — пируват и ацетил — СоА.
Процесс, начинающийся от пирувата,
называется общим
путем катаболизма и
в свою очередь включает:
Именно в общем пути катаболизма
образуется основная масса субстратов
для реакций дегидрирования. Совместно
с дыхательной цепью и окислительным
фосфорилированием общий путь катаболизма
является основным источником энергии
в форме АТР.
17.Окислительное декарбоксилирование пировиноградной кислоты. Последовательность реакций. Строение пируватдекарбоксилазного комплекса.
Окислительное
декарбоксилирование пирувата происходит
в матриксе митохондрий. Транспорт
пирувата в митохондриальный матрикс
через внутреннюю мембрану митохондрий
осуществляется при участии специального
белка-переносчика по механизму симпорта
с Н+.
Превращение пирувата в ацетил-КоА
описывают следующим суммарным уравнением:
СН3-СО-СООН
+ NAD+ +
HSKoA
→ Ch4-CO
∼SKoA
+ NADH
+ H+ +
CO2
В
ходе этой реакции происходит окислительное
декарбоксилирование пирувата, в
результате которого карбоксильная
группа удаляется в виде СО2,
а ацетильная группа включается в состав
ацетил- КоА. Один атом водорода оказывается
в составе NADH, а другой в виде Н+ поступает
в среду. Реакция необратима, поскольку
ΔG0′ =
-33,5 кДж/моль.
Строение
пируватдегидрогеназного комплекса
Процесс
окислительного декарбоксилирования
пирувата катализирует сложнооргани-зованный
пируватдегидрогеназный комплекс. В
пируватдегидрогеназный комплекс (ПДК)
входят 3 фермента: пируватдекарбоксилаза
(Е1),
дигидролипоилтрансацетилаза (Е2)
и дигидролипоилдегидрогеназа (Е3),
а также 5 коферментов: тиаминдифосфат
(ТДФ), липоевая кислота,. FAD, NAD+ и
КоА. Кроме того, в состав комплекса
входят регуляторные субъединицы:
протеинкиназа и фосфопротеинфосфатаза.
Все эти ферменты и коферменты объединены
в мультиферментную систему, содержащую
разные количества каждого из ферментов
и имеющую молекулярную массу более
6×106.
В центре комплекса располагается
дигидролипоилтрансацетилаза (Е2),
образуя его ядро. К дигидролипоилтрансацетилазе
присоединены молекулы: пируватдекарбоксилазы
(Е1)
и дигидролипоилдегидрогеназы (Е3).
Пируватдекарбоксилаза содержит прочно
связанный с белковой частью ТДФ, а
дигидроли-поилдегидрогеназа —
FAD.Липоиллизиновые группы центрального
фермента (Е2)
функционируют как поворотные «кронштейны»,
переносящие атомы водорода и ацетильные
группы от одной ферментной молекулы
комплекса к другой.
Окислительное
декарбоксилирование пирувата
Превращение
пирувата в ацетил-КоА включает 5 стадий
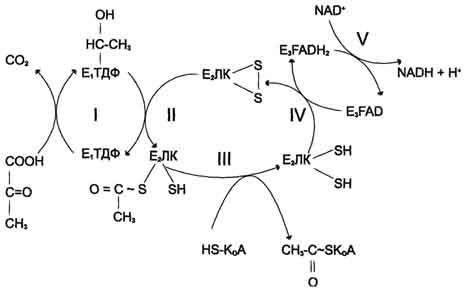
Пируват + Е1-ТДФ
→ Гидроксиэтил-ТДФ + CO2.
-
Стадия П. Дигидролипоилтрансацетилаза
(Е2)
катализирует перенос атома водорода
и ацетильной группы от ТДФ на окисленную
форму липоиллизиновых групп с образованием
ацетилтиоэфира липоевой кислоты. -
Стадия III. На
стадии III КоА взаимодействует с ацетильным
производным Е2,
в результате чего образуются ацетил-КоА
и полностью восстановленный липоильный
остаток, простетическая группа Е2. -
Стадия IV. На
стадии IV дигидролипоилде-гидрогеназа
(Е3)
катализирует перенос атомов водорода
от восстановленных липоильных групп
на FAD — простетическую группу фермента
Е3. -
Стадия V .
На стадии V восстановленный FADH2 передаёт
водород на NAD+ с
образованием NADH.
Пируватдегидрогеназный
комплекс характеризуется большим
отрицательным окислительно-восстановительным
потенциалом, который обеспечивает
наряду с восстановлением кофермента
(NADH) образование высокоэнергетической
тиоэфирной связи в ацетил-КоА
studfile.net
43.Катаболизм основных пищевых веществ — углеводов, жиров, белков. Понятие о специфических путях катаболизма и общих путях катаболизма.

Если
процесс катаболизма рассматривать с
общей точки зрения, то можно выделить
три основные его части:
-
Расщепление в
пищеварительном тракте.
Это гидролитические реакции, превращающие
сложные пищевые вещества в относительно
небольшое число простых метаболитов:
глюкоза, аминокислоты, глицерин, жирные
кислоты. -
Специфические пути
катаболизма.
Простые метаболиты подвергаются
специфическим реакциям расщепления,
в результате которых образуется либо
пировиноградная кислота, либо ацетил
— КоА. Причем ацетил — КоА может
образоваться из пирувата в результате
окислительного декарбоксилирования.
Могут также образоваться другие
соединения, непосредственно включающиеся
в цитратный цикл. -
Цитратный цикл и
дыхательная цепь завершают
расщепление пищевых веществ до конечных
продуктов — СО2 и
Н2О.
Следовательно, начиная со
стадии образования пирувата происходит
унификация путей катаболизма. Из большого
числа исходных соединений образуется
всего два — пируват и ацетил — СоА.
Процесс, начинающийся от пирувата,
называется общим
путем катаболизма и
в свою очередь включает:
Именно в общем пути катаболизма образуется
основная масса субстратов для реакций
дегидрирования. Совместно с дыхательной
цепью и окислительным фосфорилированием
общий путь катаболизма является основным
источником энергии в форме АТР.
44.Окислительное декарбоксилирование пировиноградной кислоты. Последовательность реакций. Строение пируватдекарбоксилазного комплекса.
Окислительное
декарбоксилирование пирувата происходит
в матриксе митохондрий. Транспорт
пирувата в митохондриальный матрикс
через внутреннюю мембрану митохондрий
осуществляется при участии специального
белка-переносчика по механизму симпорта
с Н+.
Превращение пирувата в ацетил-КоА
описывают следующим суммарным уравнением:
СН3-СО-СООН
+ NAD+ +
HSKoA → Ch4-CO ∼SKoA
+ NADH + H+ +
CO2
В
ходе этой реакции происходит окислительное
декарбоксилирование пирувата, в
результате которого карбоксильная
группа удаляется в виде СО2,
а ацетильная группа включается в состав
ацетил- КоА. Один атом водорода оказывается
в составе NADH, а другой в виде Н+ поступает
в среду. Реакция необратима, поскольку
ΔG0′ =
-33,5 кДж/моль.
Строение
пируватдегидрогеназного комплекса
Процесс
окислительного декарбоксилирования
пирувата катализирует сложнооргани-зованный
пируватдегидрогеназный комплекс. В
пируватдегидрогеназный комплекс (ПДК)
входят 3 фермента: пируватдекарбоксилаза
(Е1),
дигидролипоилтрансацетилаза (Е2)
и дигидролипоилдегидрогеназа (Е3),
а также 5 коферментов: тиаминдифосфат
(ТДФ), липоевая кислота,. FAD, NAD+ и
КоА. Кроме того, в состав комплекса
входят регуляторные субъединицы:
протеинкиназа и фосфопротеинфосфатаза.
Все эти ферменты и коферменты объединены
в мультиферментную систему, содержащую
разные количества каждого из ферментов
и имеющую молекулярную массу более
6×106.
В центре комплекса располагается
дигидролипоилтрансацетилаза (Е2),
образуя его ядро. К дигидролипоилтрансацетилазе
присоединены молекулы: пируватдекарбоксилазы
(Е1)
и дигидролипоилдегидрогеназы (Е3).
Пируватдекарбоксилаза содержит прочно
связанный с белковой частью ТДФ, а
дигидроли-поилдегидрогеназа —
FAD.Липоиллизиновые группы центрального
фермента (Е2)
функционируют как поворотные «кронштейны»,
переносящие атомы водорода и ацетильные
группы от одной ферментной молекулы
комплекса к другой.
Окислительное
декарбоксилирование пирувата
Превращение
пирувата в ацетил-КоА включает 5 стадий

Пируват + Е1-ТДФ
→ Гидроксиэтил-ТДФ + CO2.
-
Стадия П. Дигидролипоилтрансацетилаза
(Е2)
катализирует перенос атома водорода
и ацетильной группы от ТДФ на окисленную
форму липоиллизиновых групп с образованием
ацетилтиоэфира липоевой кислоты. -
Стадия III. На
стадии III КоА взаимодействует с ацетильным
производным Е2,
в результате чего образуются ацетил-КоА
и полностью восстановленный липоильный
остаток, простетическая группа Е2. -
Стадия IV. На
стадии IV дигидролипоилде-гидрогеназа
(Е3)
катализирует перенос атомов водорода
от восстановленных липоильных групп
на FAD — простетическую группу фермента
Е3. -
Стадия V .
На стадии V восстановленный FADH2 передаёт
водород на NAD+ с
образованием NADH.
Пируватдегидрогеназный
комплекс характеризуется большим
отрицательным окислительно-восстановительным
потенциалом, который обеспечивает
наряду с восстановлением кофермента
(NADH) образование высокоэнергетической
тиоэфирной связи в ацетил-КоА
studfile.net
43.Катаболизм основных пищевых веществ — углеводов, жиров, белков. Понятие о специфических путях катаболизма и общих путях катаболизма.

Если
процесс катаболизма рассматривать с
общей точки зрения, то можно выделить
три основные его части:
-
Расщепление в
пищеварительном тракте.
Это гидролитические реакции, превращающие
сложные пищевые вещества в относительно
небольшое число простых метаболитов:
глюкоза, аминокислоты, глицерин, жирные
кислоты. -
Специфические пути
катаболизма.
Простые метаболиты подвергаются
специфическим реакциям расщепления,
в результате которых образуется либо
пировиноградная кислота, либо ацетил
— КоА. Причем ацетил — КоА может
образоваться из пирувата в результате
окислительного декарбоксилирования.
Могут также образоваться другие
соединения, непосредственно включающиеся
в цитратный цикл. -
Цитратный цикл и
дыхательная цепь завершают
расщепление пищевых веществ до конечных
продуктов — СО2 и
Н2О.
Следовательно, начиная со
стадии образования пирувата происходит
унификация путей катаболизма. Из большого
числа исходных соединений образуется
всего два — пируват и ацетил — СоА.
Процесс, начинающийся от пирувата,
называется общим
путем катаболизма и
в свою очередь включает:
Именно в общем пути катаболизма образуется
основная масса субстратов для реакций
дегидрирования. Совместно с дыхательной
цепью и окислительным фосфорилированием
общий путь катаболизма является основным
источником энергии в форме АТР.
44.Окислительное декарбоксилирование пировиноградной кислоты. Последовательность реакций. Строение пируватдекарбоксилазного комплекса.
Окислительное
декарбоксилирование пирувата происходит
в матриксе митохондрий. Транспорт
пирувата в митохондриальный матрикс
через внутреннюю мембрану митохондрий
осуществляется при участии специального
белка-переносчика по механизму симпорта
с Н+.
Превращение пирувата в ацетил-КоА
описывают следующим суммарным уравнением:
СН3-СО-СООН
+ NAD+ +
HSKoA → Ch4-CO ∼SKoA
+ NADH + H+ +
CO2
В
ходе этой реакции происходит окислительное
декарбоксилирование пирувата, в
результате которого карбоксильная
группа удаляется в виде СО2,
а ацетильная группа включается в состав
ацетил- КоА. Один атом водорода оказывается
в составе NADH, а другой в виде Н+ поступает
в среду. Реакция необратима, поскольку
ΔG0′ =
-33,5 кДж/моль.
Строение
пируватдегидрогеназного комплекса
Процесс
окислительного декарбоксилирования
пирувата катализирует сложнооргани-зованный
пируватдегидрогеназный комплекс. В
пируватдегидрогеназный комплекс (ПДК)
входят 3 фермента: пируватдекарбоксилаза
(Е1),
дигидролипоилтрансацетилаза (Е2)
и дигидролипоилдегидрогеназа (Е3),
а также 5 коферментов: тиаминдифосфат
(ТДФ), липоевая кислота,. FAD, NAD+ и
КоА. Кроме того, в состав комплекса
входят регуляторные субъединицы:
протеинкиназа и фосфопротеинфосфатаза.
Все эти ферменты и коферменты объединены
в мультиферментную систему, содержащую
разные количества каждого из ферментов
и имеющую молекулярную массу более
6×106.
В центре комплекса располагается
дигидролипоилтрансацетилаза (Е2),
образуя его ядро. К дигидролипоилтрансацетилазе
присоединены молекулы: пируватдекарбоксилазы
(Е1)
и дигидролипоилдегидрогеназы (Е3).
Пируватдекарбоксилаза содержит прочно
связанный с белковой частью ТДФ, а
дигидроли-поилдегидрогеназа —
FAD.Липоиллизиновые группы центрального
фермента (Е2)
функционируют как поворотные «кронштейны»,
переносящие атомы водорода и ацетильные
группы от одной ферментной молекулы
комплекса к другой.
Окислительное
декарбоксилирование пирувата
Превращение
пирувата в ацетил-КоА включает 5 стадий
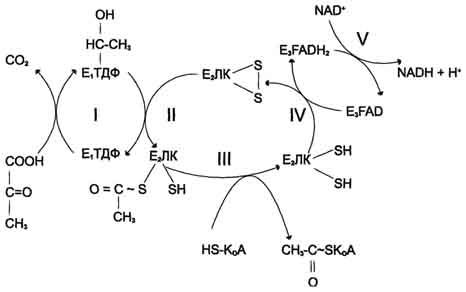
Пируват + Е1-ТДФ
→ Гидроксиэтил-ТДФ + CO2.
-
Стадия П. Дигидролипоилтрансацетилаза
(Е2)
катализирует перенос атома водорода
и ацетильной группы от ТДФ на окисленную
форму липоиллизиновых групп с образованием
ацетилтиоэфира липоевой кислоты. -
Стадия III. На
стадии III КоА взаимодействует с ацетильным
производным Е2,
в результате чего образуются ацетил-КоА
и полностью восстановленный липоильный
остаток, простетическая группа Е2. -
Стадия IV. На
стадии IV дигидролипоилде-гидрогеназа
(Е3)
катализирует перенос атомов водорода
от восстановленных липоильных групп
на FAD — простетическую группу фермента
Е3. -
Стадия V .
На стадии V восстановленный FADH2 передаёт
водород на NAD+ с
образованием NADH.
Пируватдегидрогеназный
комплекс характеризуется большим
отрицательным окислительно-восстановительным
потенциалом, который обеспечивает
наряду с восстановлением кофермента
(NADH) образование высокоэнергетической
тиоэфирной связи в ацетил-КоА
studfile.net
Катаболизм глюкозы
Расщепление
глюкозы в тканях происходит аэробно –
с участием кислорода – это основной
путь катаболизма. Окисление глюкозы
без доступа кислорода – анаэробное
превращение, которое начинается с
гликогена и заканчивается с образованием
молочной кислоты называется гликогенолизом.
Если этот процесс начинается
с
глюкозы,
то называется
гликолизом.
Гликогенолиз
Глюкоза,
депонированная в форме гликогена,
освобождается из него при участии
фермента гликогенфосфорилазы. Этот
фермент катализирует фосфорилирование
1,4-гликозидной связи невосстанавливающих
концов гликогена.
Глюкозо-1-фосфат,
образующийся из гликогена, при участии
фосфоглюкомутазы превращается в
глюкозо-6-фосфат, то есть остаток фосфорной
кислоты переносится на шестой углеродный
атом глюкозы. Дальнейшие превращения
глюкозо-6-фосфата такие же, как при
гликолизе.
Гликолиз
– распад глюкозы начинается с
фосфорилирования с участием АТФ
[остаток
фосфорной кислоты обозначается -PO-3H2
или
(P)].

Глюкоза
Глюкозо-6-фосфат
В
результате образуется глюкозо-6-фосфат.
Фермент, осуществляющий эту реакцию –
фосфогексокиназа (реже фосфоглюкокиназа).
Следует подчеркнуть, что глюкоза способна
проходить через клеточные мембраны, а
глюкозо-6-фосфат не может, в результате
фосфорилирования глюкозы она «запирается»
в клетке.
Глюкозо-6-фосфат,
образующийся при гликолизе и гликогенолизе
может подвергаться следующим обменным
превращениям:
а)
образуется фруктозо-6-фосфат и происходят
дальнейшие этапы гликолиза;
б)
происходит дефосфорилирование с
образованием глюкозы и неорганического
фосфата;
в) вовлекается в
пентозный путь распада.
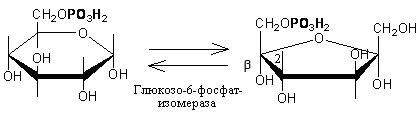
1.
Образование фруктозо-6-фосфата происходит
в результате изомерного превращения с
участием фермента фосфоглюкоизомеразы:
2.
В следующей реакции происходит
фосфорилирование фруктозо-6-фосфата с
участием АТФ и фермента фосфофруктокиназы,
в результате образуется фруктозо-1,6-ди-фосфат.

Фруктозо
— 6 — фосфат Фруктозо -.1,6-
дифосфат
3.
Наличие двух остатков фосфата на
противоположных концах молекулы гексозы
приводит к резкому ослаблению связей
между третьим и четвертым атомами
углерода. В результате этого углевод
легко расщепляется на 2 фосфотриозы под
влиянием фермента альдолазы:
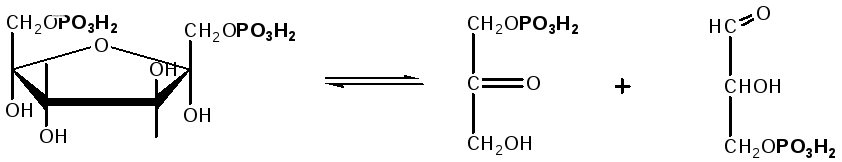
Фруктозо-1,6-дифосфат
Диоксиацетон-
Глицеральдегид
фосфат
— 3- фосфат
При
этом образуется 3-фосфоглицериновый
альдегид (3%) и диоксиацетонфосфат (97%),
который превращается в 3-фосфоглицериновый
альдегид:

Диоксиацетонфосфат
Глицеральдегид-3-фосфат
На
этом первый этап гликолиза завершается.
На втором этапе происходит окисление
3-фосфоглицеринового альдегида до
пировиноградной кислоты.
4.
3-фосфоглицериновый альдегид под влиянием
фермента глицеральдегиддигидрогеназы
и НАД вступает в реакцию образования
1,3-дифосфоглицериновой кислоты:

D-глицеральдегид-
1,3-дифосфоглицериновая
3-фосфат
кислота
При
этом в начале реакции образуется
фермент-субстратный комплекс, который
в присутствии фосфорной кислоты
распадается с образованием нативного
HS-фермента
(глицеральдегид-3-фосфатдегидрогеназы)
и 1,3-дифосфоглицериновой кислоты. При
этом энергия окисления 3-фосфоглицеринового
альдегида не рассеивается в виде тепла,
а используется для образования богатых
энергией фосфорных связей
1,3-дифосфоглицериновой кислоты. Богатые
энергией (макроэргические) связи в
формуле обозначаются в виде «~». Следует
подчеркнуть, что при окислении на
субстратном уровне происходит
восстановлении НАД с образованием
НАДН2,
который не может передавать водород
непосредственно на дыхательную цепь,
поскольку митохондриальная мембрана
непроницаема для НАДН2.
Перенос водорода цитозольного НАДН2
в
митохондрии происходит при участии
специальных механизмов.
-
Реакция
переноса остатка фосфорной кислоты с
богатой энергией связью 1,3-дифосфоглицериновой
кислоты на АДФ, в результате образуется
АТФ. Реакция происходит с участием
фермента фосфоглицераткиназы:
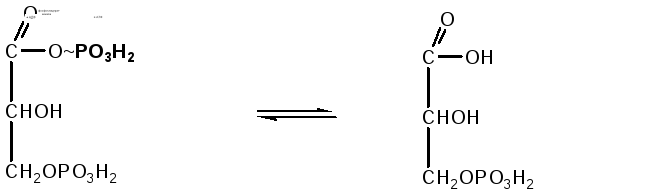
1,3-дифосфоглицериновая
3-фосфоглицериновая
кислота
кислота
Таким
образом, энергия, высвобождающаяся при
окислении альдегидной группы
глицеральдегид-3-фосфата до карбоксильной
группы, запасается в форме энергии АТФ.
В отличие от окислительного фосфорилирования,
образование АТФ из высокоэнергетических
соединений называется субстратным
фосфорилированием.
6.
3-фосфоглицериновая кислота под влиянием
фермента фосфоглицеромутазы превращается
в 2-фосфоглицериновую кислоту, то есть
остаток фосфорной кислоты с третьего
углеродного атома переносится ко второму
углеродному атому глицериновой кислоты:

3-фосфоглицериновая
2-фосфоглицериновая
кислота
кислота
7.
2-фосфоглицериновая кислота теряет
молекулу воды под действием фермента
енолгидратазы, в результате образуется
енольная форма 2-фосфопировиноградной
кислоты, имеющая макроэргическую
фосфатную связь:
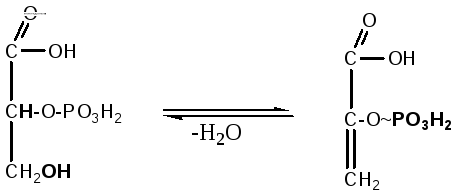
2-фосфоглицериновая
2-фосфоенолпировиноградная
кислота
кислота
8.
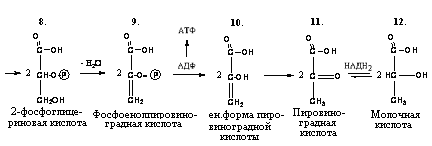
2-фосфоенолпировиноградная кислота под
действием фермента пируваткиназы отдает
макроэргическую связь на АДФ, в результате
синтезируется молекула АТФ (субстратное
фосфорилирование). Енольная форма
пировиноградной кислоты затем превращается
в кетоформу пирувата:
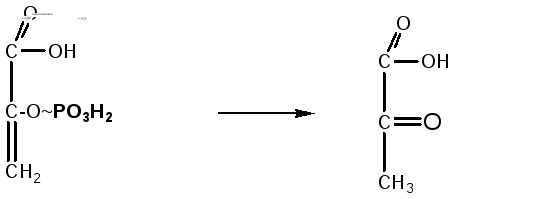
Фосфоенолпировиноградная
Пировиноградная
кислота
кислота
9.
При недостатке кислорода пировиноградная
кислота под действием лактатдегидрогеназы
с участием НАДН2
превращается в молочную кислоту, которая
является конечным продуктом анаэробного
расщепления глюкозы в животных тканях:
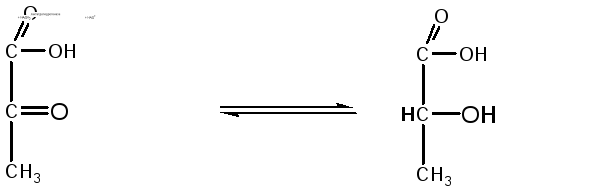
Пируват
Лактат (молочная кислота)
Лактатдегидрогеназа
в аэробных условиях может катализировать
образование пировиноградной кислоты
из молочной, то есть вести реакцию в
обратном направлении.
При
гликолизе из одной молекулы глюкозы
образуется 2 молекулы молочной кислоты
и синтезируется 4 молекулы АТФ, из них
2 молекулы расходуется на фосфорилирование
глюкозы (образование глюкозы 6-фосфата)
и фруктозы-6-фосфата (образование фруктозы
1,6-дифосфата). Суммарную реакцию гликолиза
можно записать в виде следующего
уравнения:
C6H12O6
+ 2АДФ + 2Фнеорг. → 2C3H6O3
+ 2 АТФ
Последовательность
реакций гликолиза можно представить в
виде:
 глюкоза
глюкоза
глюкозо- фруктозо-6-фосфат
фруктозо-
6-фосфат
1,6-фосфат
Молочнокислое
брожение
отличается от гликолиза тем, что при
этом в качестве конечного продукта
распада образуется две молекулы молочной
кислоты. Обычно брожением называют
микробиологическое, а гликолизом
тканевое окисление глюкозы без доступа
кислорода.
Этаноловое
(спиртовое) брожение
происходит под действием ферментов
дрожжевых клеток. При этом пировиноградная
кислота декарбоксилируется с участием
пируватдекарбоксилазы с образованием
уксусного альдегида:
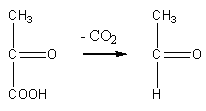
пируват
уксусный альдегид
Затем
уксусный альдегид восстанавливается
с участием НАДН2
в этанол:
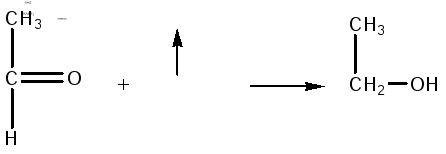
Таким
образом, из одной молекулы глюкозы
получается 2 молекулы этилового спирта.
Аэробный
путь распада глюкозы является основной
формой ее катаболизма. Десять ферментов,
катализирующих распад глюкозы до
пировиноградной кислоты, локализованы
в цитозоле, все остальные – в митохондриях.
Пировиноградная кислота, образованная
в цитозоле, проходит через мембраны
митохондрий с помощью малатного
челночного механизма и подвергается
окислительному декарбоксилированию с
образованием активированной уксусной
кислоты (ацетил-КоА).
Реакция
происходит под действием сложного
мультиферментного комплекса метаболона
с молекулярной массой свыше 6000000 Да,
состоящего из ферментов, осуществляющих
многоступенчатый процесс биохимических
превращений. В результате образуется
ацетил~SКоА,
1 молекула углекислоты, 1 молекула воды
и 3 молекулы АТФ:

Реакция
окислительного декарбоксилирования
подавляется молекулами АТФ, то есть
накопление молекул АТФ является сигналом
для прекращения реакции декарбоксилирования
пировиноградной кислоты.
Цикл
трикарбоновых кислот
или цикл
Кребса
является завершающим этапом распада
энергетических материалов в клетке. В
нем происходит окисление активированной
уксусной кислоты (ацетил~SКоА)
– промежуточного метаболита углеводов,
белков и липидов – до конечных продуктов
– углекислоты, воды и энергии. Реакции
цикла трикарбоновых кислот происходят
во внутренних отсеках митохондрий, то
есть на внутренней мембране.
Митохондрии
обычно имеют форму цилиндра с закругленными
концами, длиной 1-4 мкм и поперечником
0,3-0,7 мкм. Количество митохондрий в разных
клетках различно, например, в клетках
печени их может быть до 2000 (рис. 8.5.).
Система
ферментов тканевого окисления (дыхания)
расположены на внутренней мембране,
где происходит окисление СН3-СО~КоА
до конечных продуктов – CO2,
H2O,
с выделением энергии. В цикле трикарбоновых
кислот происходит Дегидрирование ди-
и трикарбоновых кислот, тканевое дыхание
и окислительное фосфорилирование, в
этих реакциях участвуют ферменты
дыхательной цепи и окислительного
фосфорилирования.

Рис.8.5.
Строение митохондрий.
1 —
наружная мембрана:
2 —
внутренняя мембрана;
3 — матрикс;
4 — кристы, образованные
внутренней
мембраной.
Реакции
цикла трикарбоновых кислот происходят
в несколько этапов:
-
Образование
лимонной кислоты происходит с участием
щавелевоуксусной кислоты и ацетил-SKoA.
В результате образуется лимонная
кислота под действием цитратсинтетазы,
a
HS-KoA
освобождается.
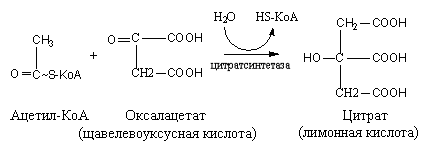
-
Превращение
лимонной кислоты в изолимонную происходит
под действием фермента аконитазы:
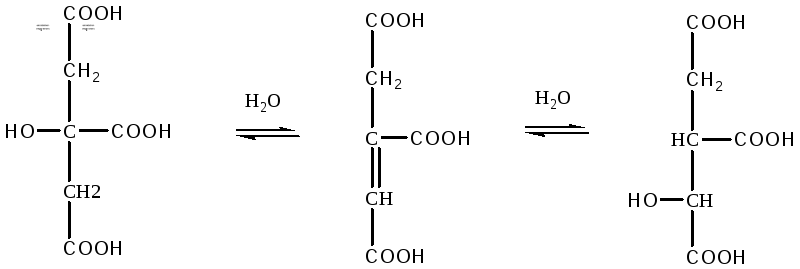
Цитрат
цис – Аконитат Изоцитрат
Фермент
аконитаза осуществляет отнятие молекулы
воды от лимонной кислоты, присоединение
молекулы воды к цис-аконитовой кислоте
и изомерное превращение цитрата в
изоцитрат.
3.
Отнятие двух атомов водорода от
изолимонной кислоты с образованием
щавелевоянтарной кислоты происходит
под действием фермента изоцитратдегидрогеназы.
В последующем с участием этого же
фермента происходит декарбоксилирование
щавелевоянтарной кислоты с образованием
а-кетоглутаровой кислоты:
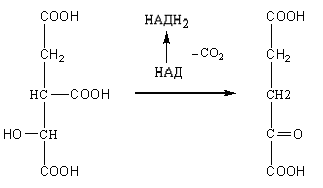
Изоцитрат
α-Кетоглутарат
При
отнятии двух атомов водорода от
изолимонной кислоты происходит
восстановление НАД в НАДН2
(или в НАДФН2),
который при окислении в дыхательной
цепи обеспечивает синтез трех молекул
АТФ.
4.
α-Кетоглутаровая кислота под влиянием
фермента α-кетоглутаратдегидрогеназы
декарбоксилируется, в результате
образуется активированная янтарная
кислота – сукцинил-КоА, содержащая
макроэргическую связь:

При
этом НАД восстанавливается в НАДН2,
который в дыхательной цепи обеспечивает
синтез трех молекул АТФ.
В
последующем активированная янтарная
кислота под влиянием фермента
сукцинил-КоА-синтетазы передает
макроэргическую связь ГДФ:
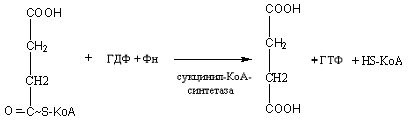
Активированная
Янтарная кислота
янтарная кислота
Образовавшийся
при этом ГТФ вступает в реакцию
перефосфорилирования с АДФ по следующей
схеме:
Г
 ТФ
ТФ
+ АДФ АТФ + ГДФ
5.
Янтарная кислота под воздействием
фермента сукцинатдегидрогеназы
окисляется до фумаровой кислоты.
Коферментом при этом является ФАД:
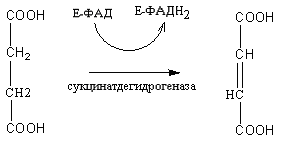
Янтарная
кислота Фумаровая кислота
ФАДН2
окисляется в дыхательной цепи
митохондрий и обеспечивает синтез двух
молекул АТФ.
6.
Фумаровая кислота под воздействием
фермента фумаратгидратазы превращается
в яблочную кислоту:
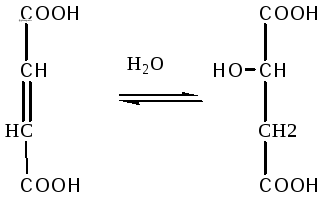
Фумаровая
Яблочная
кислота
кислота
7.
Яблочная кислота дегидрируется под
действием фермента малатдегидрогеназы,
в результате образуется щавелевоуксусная
кислота (ЩУК) и происходит восстановление
НАД в НАДН2,
последний в дыхательной цепи окисляется
и обеспечивает образование трех молекул
АТФ.
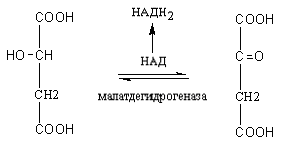
Яблочная
кислота Щавелевоуксусная
кислота
С
момента образования щавелевоуксусной
кислоты цикл трикарбоновых кислот
замыкается и может повторяться в том
же порядке: при наличии в системе
ацетил-КоА, ЩУК снова включается в цикл
трикарбоновых кислот.
Суммарная
реакция окисления ацетил-КоА в цикле
трикарбоновых кислот:
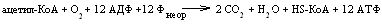
Таким
образом, в одном цикле трикарбоновых
кислот до конечных продуктов – CO2
и H2O
окисляется одна молекула активированной
уксусной кислоты (ацетил-КоА), а энергия,
высвобожденная при этом аккумулируется
в 12 молекулах АТФ.
Суммарное
выражение цикла трикарбоновых кислот
представлено на рис. 8.6
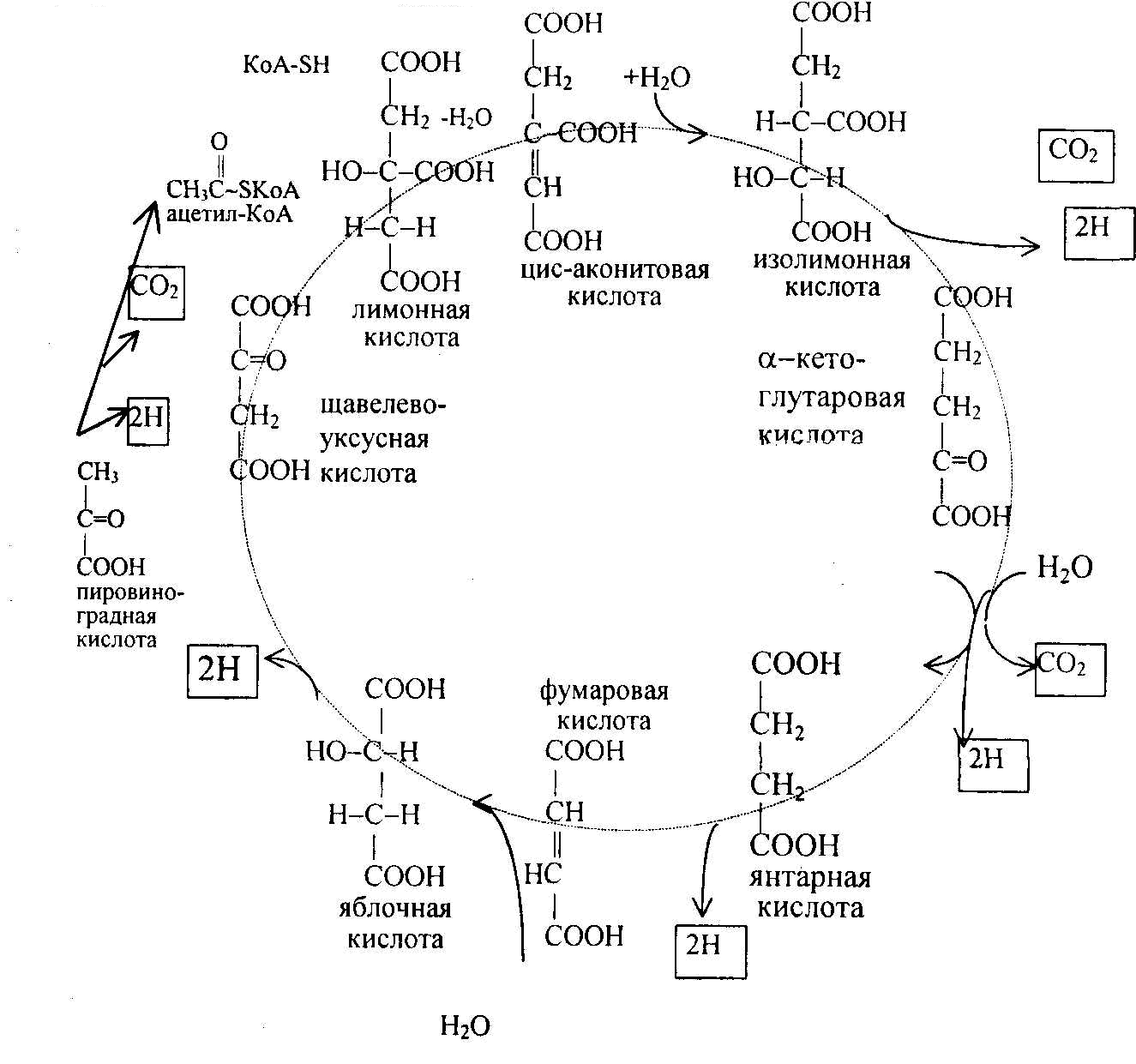
Рис. 8.6. Цикл
трикарбоновых кислот; конечные продукты
обведены рамкой.
Окисление
одной молекулы глюкозы до конечных
продуктов – CO2
и
H2O
обеспечивает синтез 38 молекул АТФ:
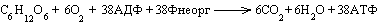
Из них
24 молекулы АТФ образуется в цикле
трикарбоновых кислот, 6 – при
декарбоксилировании пировиноградной
кислоты в ацетил-КоА, 2 – при анаэробном
гликолизе, 6 – при окислении НАДН2,
образующегося при субстратном окислении
фосфоглицеринового альдегида в цитозоле.
Пентозофосфатный
путь окисления глюкозо-6-фосфата
–
это цепь последовательных химических
превращений глюкозы, в результате
которых образуется энергия и пентозы,
необходимые для синтеза нуклеиновых
кислот, нуклеотидов и коферментов.
В
пентозофосфатном пути можно выделить
две части: окислительный и неокислительный
пути образования пентоз.
Окислительный
путь включает две реакции дегидрирования,
в которых акцептором водорода является
НАДФ. Во второй из этих реакций одновременно
происходит декарбоксилирование –
углеродная цепь укорачивается на один
атом углерода и получаются пентозы:
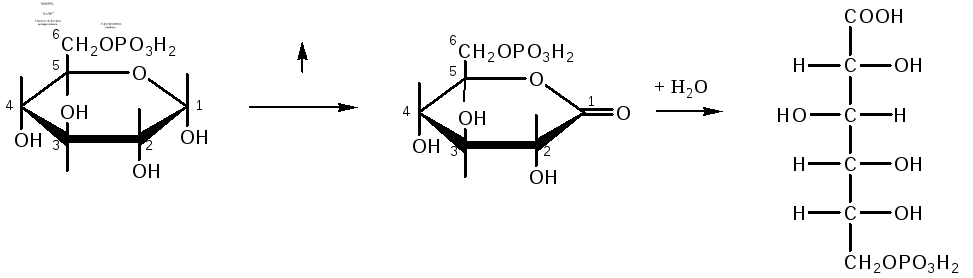
глюкозо-6-фосфат
6-фосфоглюконо-δ-лактон
6-фосфоглюконат
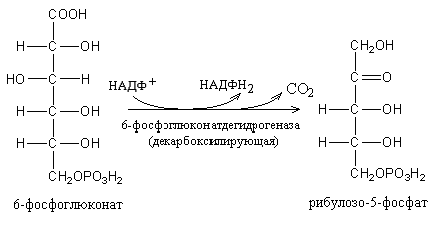
Распад
шести молекул глюкозо-6-фосфата начинается
с их дегидрирования в 6-фосфоглюконолактон,
который, присоединяя H2O,
превращается в 6-фосфоглюконовую кислоту.
Последняя окисляется в З-кето-6-фосфоглюконовую
кислоту, которая декарбоксилируясь,
превращается в D-рибулозо-5-фосфат.
На
втором этапе 6 молекул рибозо-5-фосфата
превращается в 5 молекул глюкозо-6-фосфата.
В
процессе пентозного цикла из 6 молекул
глюкоза-6-фосфата расщепляется 1 молекула
гексозы. Суммарная реакция:
6 Глюкозо-6-фосфат+12НАДФ
Глюкозо-6-фосфат+12НАДФ
 5
5
глюкозо-6-фосфат+6СО2+12НАДФН2+НзРО4+36АТФ
Значение
цикла не только в генерации энергии (36
АТФ), но и в образовании пентозы, НАДФН2.
Например, восстановленный НАДФН2
используется
в различных процессах синтеза, в
частности, при синтезе жирных кислот,
а пентозы – в биосинтезе нуклеиновых
кислот.
В
организме в целом различные пути распада
углеводов зависят от физических условий,
интенсивности работы. Гликолиз и аэробный
путь (дихотомический путь) имеет большее
значение, чем пентозный.
studfile.net
43.Катаболизм основных пищевых веществ — углеводов, жиров, белков. Понятие о специфических путях катаболизма и общих путях катаболизма.

Если
процесс катаболизма рассматривать с
общей точки зрения, то можно выделить
три основные его части:
-
Расщепление в
пищеварительном тракте.
Это гидролитические реакции, превращающие
сложные пищевые вещества в относительно
небольшое число простых метаболитов:
глюкоза, аминокислоты, глицерин, жирные
кислоты. -
Специфические пути
катаболизма.
Простые метаболиты подвергаются
специфическим реакциям расщепления,
в результате которых образуется либо
пировиноградная кислота, либо ацетил
— КоА. Причем ацетил — КоА может
образоваться из пирувата в результате
окислительного декарбоксилирования.
Могут также образоваться другие
соединения, непосредственно включающиеся
в цитратный цикл. -
Цитратный цикл и
дыхательная цепь завершают
расщепление пищевых веществ до конечных
продуктов — СО2 и
Н2О.
Следовательно, начиная со
стадии образования пирувата происходит
унификация путей катаболизма. Из большого
числа исходных соединений образуется
всего два — пируват и ацетил — СоА.
Процесс, начинающийся от пирувата,
называется общим
путем катаболизма и
в свою очередь включает:
Именно в общем пути катаболизма образуется
основная масса субстратов для реакций
дегидрирования. Совместно с дыхательной
цепью и окислительным фосфорилированием
общий путь катаболизма является основным
источником энергии в форме АТР.
44.Окислительное декарбоксилирование пировиноградной кислоты. Последовательность реакций. Строение пируватдекарбоксилазного комплекса.
Окислительное
декарбоксилирование пирувата происходит
в матриксе митохондрий. Транспорт
пирувата в митохондриальный матрикс
через внутреннюю мембрану митохондрий
осуществляется при участии специального
белка-переносчика по механизму симпорта
с Н+.
Превращение пирувата в ацетил-КоА
описывают следующим суммарным уравнением:
СН3-СО-СООН
+ NAD+ +
HSKoA → Ch4-CO ∼SKoA
+ NADH + H+ +
CO2
В
ходе этой реакции происходит окислительное
декарбоксилирование пирувата, в
результате которого карбоксильная
группа удаляется в виде СО2,
а ацетильная группа включается в состав
ацетил- КоА. Один атом водорода оказывается
в составе NADH, а другой в виде Н+ поступает
в среду. Реакция необратима, поскольку
ΔG0′ =
-33,5 кДж/моль.
Строение
пируватдегидрогеназного комплекса
Процесс
окислительного декарбоксилирования
пирувата катализирует сложнооргани-зованный
пируватдегидрогеназный комплекс. В
пируватдегидрогеназный комплекс (ПДК)
входят 3 фермента: пируватдекарбоксилаза
(Е1),
дигидролипоилтрансацетилаза (Е2)
и дигидролипоилдегидрогеназа (Е3),
а также 5 коферментов: тиаминдифосфат
(ТДФ), липоевая кислота,. FAD, NAD+ и
КоА. Кроме того, в состав комплекса
входят регуляторные субъединицы:
протеинкиназа и фосфопротеинфосфатаза.
Все эти ферменты и коферменты объединены
в мультиферментную систему, содержащую
разные количества каждого из ферментов
и имеющую молекулярную массу более
6×106.
В центре комплекса располагается
дигидролипоилтрансацетилаза (Е2),
образуя его ядро. К дигидролипоилтрансацетилазе
присоединены молекулы: пируватдекарбоксилазы
(Е1)
и дигидролипоилдегидрогеназы (Е3).
Пируватдекарбоксилаза содержит прочно
связанный с белковой частью ТДФ, а
дигидроли-поилдегидрогеназа —
FAD.Липоиллизиновые группы центрального
фермента (Е2)
функционируют как поворотные «кронштейны»,
переносящие атомы водорода и ацетильные
группы от одной ферментной молекулы
комплекса к другой.
Окислительное
декарбоксилирование пирувата
Превращение
пирувата в ацетил-КоА включает 5 стадий
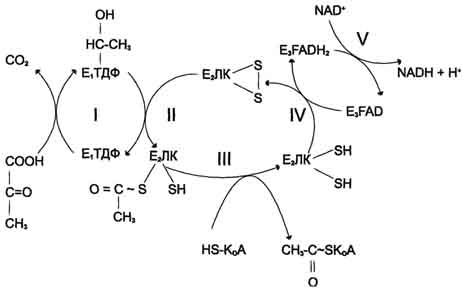
Пируват + Е1-ТДФ
→ Гидроксиэтил-ТДФ + CO2.
-
Стадия П. Дигидролипоилтрансацетилаза
(Е2)
катализирует перенос атома водорода
и ацетильной группы от ТДФ на окисленную
форму липоиллизиновых групп с образованием
ацетилтиоэфира липоевой кислоты. -
Стадия III. На
стадии III КоА взаимодействует с ацетильным
производным Е2,
в результате чего образуются ацетил-КоА
и полностью восстановленный липоильный
остаток, простетическая группа Е2. -
Стадия IV. На
стадии IV дигидролипоилде-гидрогеназа
(Е3)
катализирует перенос атомов водорода
от восстановленных липоильных групп
на FAD — простетическую группу фермента
Е3. -
Стадия V .
На стадии V восстановленный FADH2 передаёт
водород на NAD+ с
образованием NADH.
Пируватдегидрогеназный
комплекс характеризуется большим
отрицательным окислительно-восстановительным
потенциалом, который обеспечивает
наряду с восстановлением кофермента
(NADH) образование высокоэнергетической
тиоэфирной связи в ацетил-КоА
studfile.net