Белки
Белки — наиболее важные биологические вещества живых организмов. Они являются основным строительным материалом, из которого строятся клетки, ткани и органы человека. Энергетическая ценность 1 г белка равна 4,1 ккал (16,7 кДж).
Недостаток белка приводит к ряду тяжелых расстройств в организме взрослых, а у детей —к задержке роста.
В состав белков входят аминокислоты. Некоторые из них могут синтезироваться из других веществ, их называют заменимыми. Девять аминокислот, названных незаменимыми, должны поступать в организм в готовом виде. Белки, содержащие весь комплекс незаменимых аминокислот, называют Полноценными (белки рыбы, мяса, молока, яиц и др.). Белки, в состав которых не входит хотя бы одна из незаменимых аминокислот называют неполноценными.
Белки находятся в организме животных и растений в жидком состоянии (молоко, кровь), полужидком (яйца) и твердом (волосы, ногти, сухожилия).
По составу белки подразделяют на простые (протеины) и сложные (протеиды):
— простые белки — альбумины, глобулины, глютелины, проламины, протамины и гистоны;
— сложные белки — фосфоропротеиды, глюкопротеиды,_хромопротеиды, липопротеиды, нуклеопротеиды.
Под действием ферментов кислот, щелочей происходит расщепление белков — гидролиз (созревание сыров). При глубоком расщеплении белков появляется неприятный гнилостный запах (порча мяса, рыбы).
В воде большинство белков набухает и увеличивается в объеме. Процесс набухания белков играет важную роль при производстве макаронных и мучных кондитерских изделий. Яичный белок при взбивании образует пену. Это свойство используется при изготовлении кондитерских изделий.
При нагревании свыше 60°С белки свертываются — денатурируют, выпадают в осадок и вновь в воде уже не растворяются. Денатурация белков наблюдается при варке яиц, а также при производстве кисломолочных продуктов и сыров (образуется сгусток). Кроме того, денатурация белков происходит в процессе длительного хранения круп, бобовых культур, мороженого мяса, рыбы и др. В результате «старения» белки постепенно уплотняются, теряют способность к набуханию и растворимости, и продукты приобретают жесткую консистенцию, труднее развариваются и усваиваются.
Суточная потребность взрослого человека в белках составляет 80—100 г, половину должны составлять животные белки.
Калорийность Белка. Химический состав и пищевая ценность.
Химический состав и анализ пищевой ценности
Пищевая ценность и химический состав
«Белка».
В таблице приведено содержание пищевых веществ (калорийности, белков, жиров, углеводов, витаминов и минералов) на 100 грамм съедобной части.
| Нутриент | Количество | Норма** | % от нормы в 100 г |
% от нормы в 100 ккал |
100% нормы |
| Калорийность | 120 кКал | 1684 кКал | 7. 1% 1% |
5.9% | 1403 г |
| Белки | 21.23 г | 76 г | 27.9% | 23.3% | 358 г |
| Жиры | 3.21 г | 56 г | 5.7% | 4.8% | 1745 г |
| Вода | 73.83 г | 2273 г | 3.2% | 2.7% | 3079 г |
| Зола | 1. 16 г 16 г |
~ | |||
| Витамины | |||||
| Витамин В1, тиамин | 0.07 мг | 1.5 мг | 4.7% | 3.9% | 2143 г |
| Витамин В2, рибофлавин | 0.21 мг | 1.8 мг | 11.7% | 9.8% | 857 г |
| Витамин РР, НЭ | 4 мг | 20 мг | 20% | 16. 7% 7% |
500 г |
| Макроэлементы | |||||
| Калий, K | 304 мг | 2500 мг | 12.2% | 10.2% | 822 г |
| Кальций, Ca | 2 мг | 1000 мг | 0.2% | 0.2% | 50000 г |
| Магний, Mg | 24 мг | 400 мг | 6% | 5% | 1667 г |
| Натрий, Na | 103 мг | 1300 мг | 7. 9% 9% |
6.6% | 1262 г |
| Сера, S | 212.3 мг | 1000 мг | 21.2% | 17.7% | 471 г |
| Фосфор, P | 172 мг | 800 мг | 21.5% | 17.9% | 465 г |
| Микроэлементы | |||||
| Железо, Fe | 4. 7 мг 7 мг |
18 мг | 26.1% | 21.8% | 383 г |
| Селен, Se | 9.3 мкг | 55 мкг | 16.9% | 14.1% | 591 г |
| Незаменимые аминокислоты | |||||
| Аргинин* | 1.107 г | ~ | |||
| Валин | 0. 839 г 839 г |
~ | |||
| Гистидин* | 0.554 г | ~ | |||
| Изолейцин | 0.809 г | ~ | |||
| Лейцин | 1.525 г | ~ | |||
| Лизин | 1. 536 г 536 г |
~ | |||
| Метионин | 0.465 г | ~ | |||
| Треонин | 0.809 г | ~ | |||
| Фенилаланин | 0.822 г | ~ | |||
| Заменимые аминокислоты | |||||
| Аланин | 1. 006 г 006 г |
~ | |||
| Аспарагиновая кислота | 1.688 г | ~ | |||
| Глицин | 0.921 г | ~ | |||
| Глутаминовая кислота | 2.871 г | ~ | |||
| Пролин | 0. 792 г 792 г |
~ | |||
| Серин | 0.696 г | ~ | |||
| Тирозин | 0.635 г | ~ | |||
| Стеролы (стерины) | |||||
| Холестерин | 83 мг | max 300 мг | |||
| Насыщенные жирные кислоты | |||||
| Насыщеные жирные кислоты | 0. 38 г 38 г |
max 18.7 г | |||
| 14:0 Миристиновая | 0.02 г | ~ | |||
| 16:0 Пальмитиновая | 0.3 г | ~ | |||
| 18:0 Стеариновая | 0.05 г | ~ | |||
| Мононенасыщенные жирные кислоты | 1. 18 г 18 г |
min 16.8 г | 7% | 5.8% | |
| 16:1 Пальмитолеиновая | 0.1 г | ~ | |||
| 18:1 Олеиновая (омега-9) | 1.08 г | ~ | |||
| Полиненасыщенные жирные кислоты | 0.94 г | от 11.2 до 20.6 г | 8.4% | 7% | |
| 18:2 Линолевая | 0. 92 г 92 г |
~ | |||
| 18:3 Линоленовая | 0.02 г | ~ | |||
| Омега-3 жирные кислоты | 0.02 г | от 0.9 до 3.7 г | 2.2% | 1.8% | |
| Омега-6 жирные кислоты | 0.92 г | от 4.7 до 16.8 г | 19.6% | 16.3% |
Энергетическая ценность Белка составляет 120 кКал.
- oz = 28.35 гр (34 кКал)
- lb = 453.6 гр (544.3 кКал)
Основной источник: USDA National Nutrient Database for Standard Reference. Подробнее.
** В данной таблице указаны средние нормы витаминов и минералов для взрослого человека. Если вы хотите узнать нормы с учетом вашего пола, возраста и других факторов, тогда воспользуйтесь приложением
«Мой здоровый рацион».
Химический состав пшеничной и ржаной муки: Белки
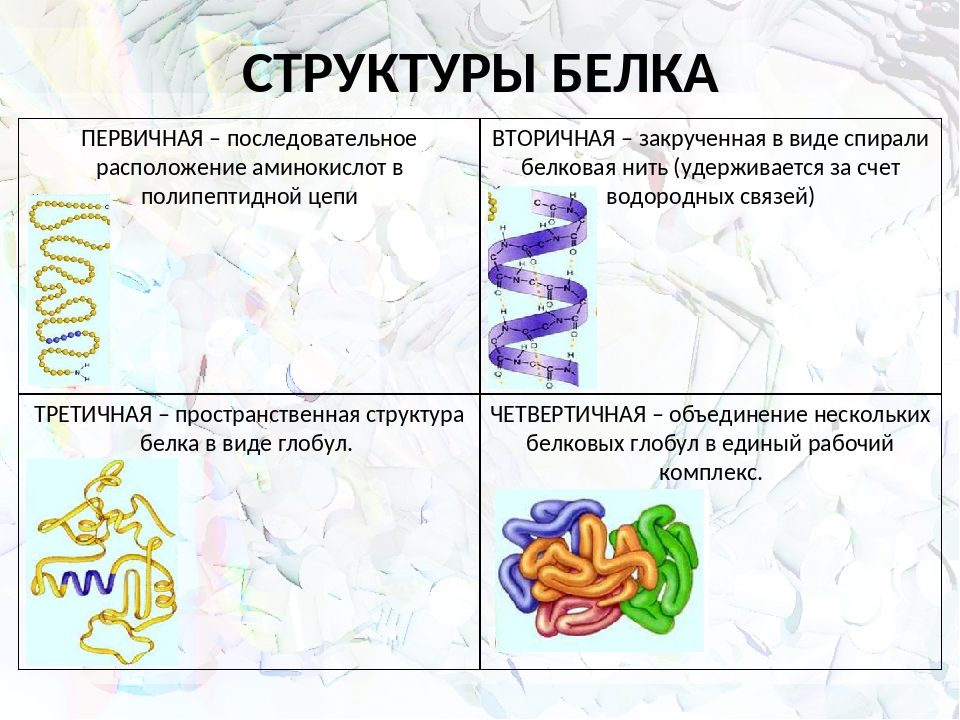
Белки — это органические высокомолекулярные соединения, состоящие из аминокислот. В молекуле белка аминокислоты соединены между собой пептидными связями. Разнообразие белков определяется последовательностью размещения остатков аминокислот в полипептидной цепи (первичная структура белка). Кроме того, существуют вторичная структура белка, характеризующая тип укладки полипептидных цепей (правая сс-спираль, а-структура и (J-изгиб), третичная структура белка, характеризующая расположение его полипептидной цепи в пространстве, и четвертичная структура, характеризующая белки, в состав которых входит несколько полипептидных цепей, связанных между собой нековалентными связями.
В состав белков пшеничной и ржаной муки входят белки простые (протеины), состоящие только из аминокислотных остатков, и сложные (протеиды). Сложные белки могут включать ионы металлов, пигменты, образовывать комплексы с липидами, нуклеиновыми кислотами, а также ковалентно связывать остаток фосфорной или нуклеиновой кислоты, углеводов. Их называют металло-протеиды, хромопротеиды, липопротеиды, нуклеопротеиды, фосфопротеиды, гликопротеиды.
Технологическая роль белков муки в приготовлении хлеба велика. Структура белковых молекул и физико-химические свойства белков определяют свойства теста, влияют на форму и качество хлеба. Белки обладают рядом свойств, которые особенно важны для приготовления хлеба.
Содержание белковых веществ в пшеничной и ржаной муке колеблется от 9 до 26% в зависимости от сорта зерна и условий его выращивания. Для белков характерны многие физико-химические свойства, из которых более всего важны растворимость, способность к набуханию, к денатурации и гидролизу.
По растворимости белки разделяют на альбумины — растворимые в воде, проламины — растворимые в спирте, глютелины — растворимые в слабых щелочах и глобулины — растворимые в солевых растворах. Белки пшеничной и ржаной муки представлены в основном про-ламинами (глиадин) и глютелинами (глютенин). Содержание этих белков составляет 2/3 или ¾ от всей массы белков муки.
Глиадин и глютенин в воде нерастворимы и поэтому при отмывании клейковины являются основными ее компонентами. В связи с этим их называют клейковинными белками. Эти белки находятся в эндосперме зерна и поэтому их больше содержится в муке высших сортов. Альбумин и глобулин содержатся в белке зародыша и алейронового слоя зерна, поэтому их больше содержится в муке низких сортов.
В сырой клейковине содержится 65—70% влаги и 35-30% сухих веществ, в сухой клейковине 90% белков и 10% крахмала, жира, сахара и других веществ муки, поглощенных белками при набухании. Количество сырой клейковины колеблется в широких пределах (15- 50% от массы муки). Чем больше белков содержится в муке и чем сильнее их способность к набуханию, тем больше получится сырой клейковины. Качество клейковины характеризуется цветом, эластичностью (способность клейковины восстанавливать свою форму после растягивания), растяжимостью (способность растягиваться на определенную длину) и упругостью (способность оказывать сопротивление при деформации).
Чем больше белков содержится в муке и чем сильнее их способность к набуханию, тем больше получится сырой клейковины. Качество клейковины характеризуется цветом, эластичностью (способность клейковины восстанавливать свою форму после растягивания), растяжимостью (способность растягиваться на определенную длину) и упругостью (способность оказывать сопротивление при деформации).
Количество клейковины и ее свойства определяют хлебопекарное достоинство муки и качество хлеба. Желательно, чтобы клейковина была эластичной, в меру упругой и имела среднюю растяжимость.
Значительная часть белков муки в воде не растворяется, но хорошо в ней набухает. Белки особенно хорошо набухают при температуре около 30° С, поглощая при этом воды в 2—3 раза больше их собственной массы.
Необратимая денатурация (изменение естественной структуры белка) происходит под действием некоторых реагентов или при нагревании свыше 60° С. Денатурированный белок теряет способность к растворимости и набуханию. Начальную стадию денатурации белков иногда специально вызывают при сушке и горячем кондиционировании зерна, чтобы несколько укрепить слабую клейковину. Значительная денатурация портит хлебопекарные свойства белковых веществ (клейковина становится неэластичной и короткорвущейся). Во время выпечки хлеба белки денатурируются полностью, свернувшийся белок образует при этом прочный каркас, закрепляющий форму изделия.
Начальную стадию денатурации белков иногда специально вызывают при сушке и горячем кондиционировании зерна, чтобы несколько укрепить слабую клейковину. Значительная денатурация портит хлебопекарные свойства белковых веществ (клейковина становится неэластичной и короткорвущейся). Во время выпечки хлеба белки денатурируются полностью, свернувшийся белок образует при этом прочный каркас, закрепляющий форму изделия.
Под действием протеолитических ферментов сложная структура белковой молекулы упрощается, уменьшается ее способность к набуханию, увеличивается растворимость белков.
Белки ржаной муки по составу и свойствам отличаются от белков пшеницы. Около половины ржаных белков растворимы в воде или в растворах солей. Белки ржаной муки имеют большую пищевую ценность, чем пшеничные (содержат много незаменимых аминокислот), однако технологические свойства их значительно ниже.
Белковые вещества ржи клейковину не образуют. В ржаном тесте большая часть белков находится в виде вязкого раствора, поэтому ржаное тесто лишено упругости и эластичности, свойственных пшеничному тесту.
ПОХОЖИЕ СТАТЬИ
На главную
Просмотрено: 29,244 раз
Коронавирус SARS-Cov-2: особенности структурных белков, контагиозность и возможные иммунные коллизии | Харченко
1. Koonin E, Dolja V, Krupovic M. Origins and evolution of viruses of eukaryotes: The ultimate modularity. Virology, 2015; 479–480: 2–25. doi: 10.1016/j.virol.2015.02.039
2. Stedman KM. Deep recombination: RNA and ssDNA virus genes in DNA virus and host genomes. Annu. Rev. Virol. 2015; 2: 203–217. doi: 10.1146/annurev-virology-100114-055127.
3. Харченко Е.П. Распространенность в геноме вирусов человека малых гомологичных и комплементарных фрагментов и возможная их роль. Инфекция и иммунитет. 2017. Т. 7, № 4. С. 393–404. doi: 10.15789/2220-7619-2017-4-393-404.
Инфекция и иммунитет. 2017. Т. 7, № 4. С. 393–404. doi: 10.15789/2220-7619-2017-4-393-404.
4. Selin LK, Wlodarczyk MF, Kraft AR et al. Heterologous immunity: immunopathology, autoimmunity and protection during viral infections. Autoimmunity. 2011; 44: 328–347. doi:10.3109/08916934.2011.523277.
5. Харченко Е.П. Вирус гонконгского гриппа: штрихи к портрету 50 лет спустя и будущая пандемия гриппа. Эпидемиология и Вакцинопрофилактика. 2020; 19 (1): 24–34. https://doi:10.31631/2073-3046-2020-19-1-24-34.
6. Song Z, Xu Y, Bao L, Zhang L et al. From SARS to MERS, Thrusting Coronaviruses into the Spotlight. Viruses. 2019;11(1). pii: E59. doi: 10.3390/v11010059.
7. Kwong PD, Mascola JR. HIV-1 Vaccines based on antibody identification, b cell ontogeny, and epitope structure. Immunity. 2018; 48: 5: 855-871. doi: 10.1016/j.immuni.2018.04.029.
HIV-1 Vaccines based on antibody identification, b cell ontogeny, and epitope structure. Immunity. 2018; 48: 5: 855-871. doi: 10.1016/j.immuni.2018.04.029.
8. Bajic G, van der Poel CE, Kuraoka M et al. Autoreactivity profiles of influenza hemagglutinin broadly neutralizing antibodies. Sci Rep. 2019; 9 (1): 3492. doi: 10.1038/s41598-019-40175-8.
9. Kanyavuz A, Marey-Jarossay A, Lacroix-Desmazes Set al. Breaking the law: unconventional strategies for antibody diversification. Nat Rev Immunol. 2019; 19 (6): 355– 368. doi: 10.1038/s41577-019-0126-7.
10. Харченко Е.П. Инвариантность генетического кода и возможный путь его эволюции. Ж.эвол.биохим. и физиол. 1986. Т.22 , N 5, С.351-354.
11. Харченко Е.П. Иммуноэпитопный континуум родства белков и полиреактивность и аутореактивность антител //Медицинская иммунология. 2015. Т. 17, № 4. C. 335–346.
Харченко Е.П. Иммуноэпитопный континуум родства белков и полиреактивность и аутореактивность антител //Медицинская иммунология. 2015. Т. 17, № 4. C. 335–346.
12. Sharma S., Thomas P.G. The two faces of heterologous immunity: protection or immunopathology. J. Leukoc. Biol., 2014, vol. 95, pp. 405–416. doi: 10.1189/jlb.0713386.
13. Shen Z.T., Nguyen T.T., Daniels K.A. et al. Disparate epitopes mediating protective heterologous immunity to unrelated viruses share peptide-MHC structural features recognized by cross-reactive T cells. J. Immunol., 2013, vol. 191, no. 10, pp. 5139–5152. doi: 10.4049/jimmunol.1300852 .
14. Welsh R.M., Che J.W., Brehm M.A et al.. Heterologous immunity between viruses. Immunol. Rev., 2010, vol. 235, no. 1, pp. 244–266. doi: 10. 1111/j.0105-2896.2010.00897.
1111/j.0105-2896.2010.00897.
15. Miller A., Reandelar M.J., Fasciglione K et al. Roumenova V., Li Y., Otazu G. H. Correlation between universal BCG vaccination policy and reduced morbidity and mortality for COVID-19: an epidemiological study. https://doi.org/10.1101/2020.03.24.20042937 .
16. Gil A., Kenney L.L., Mishra R. et al. Vaccination and heterologous immunity: educating the immune system. Trans. R. Soc. Trop. Med. Hyg., 2015, vol. 109, no. 1, pp. 62–69. doi: 10.1093/trstmh/tru198.
17. Харченко Е.П. Оптимизация прогнозирования вакцинных штаммов гриппа. Эпидемиология и Вакцинопрофилактика. 2019; Т. 18. № 1.С 4-17. doi: 10.31631/2073-3046-2019-18-1-4-17.
18. Czub, M., Weingartl, H. , Czub, S. et al. Evaluation of modified vaccinia virus Ankara based recombinant SARS vaccine in ferrets. Vaccine 2005; vol. 23, pp. 2273–2279. doi: 10.1016/j.vaccine.2005.01.033.
, Czub, S. et al. Evaluation of modified vaccinia virus Ankara based recombinant SARS vaccine in ferrets. Vaccine 2005; vol. 23, pp. 2273–2279. doi: 10.1016/j.vaccine.2005.01.033.
19. Weingartl, H., Czub, M., Czub, S. et al. Immunization with modified vaccinia virus Ankara-based recombinant vaccine against severe acute respiratory syndrome is associated with enhanced hepatitis in ferrets. J. Virol. 2004; vol. 78, pp. 12672–12676. doi: 10.1128/JVI.78.22.12672-12676.2004.
20. Pardi N, Hogan MJ, Porter FW et al. mRNA vaccines – a new era in vaccinology. Nat Rev Drug Discov. 2018; vol. 17, no.4, pp.261 –279. doi: 10.1038/nrd.2017.243.
21. Ge D, Du Q, Ran B et al. The neurotoxicity induced by engineered nanomaterials. Int J Nanomedicine. 2019; vol. 6. no.14, pp. 4167-4186. doi: 10.2147/IJN.S203352.
4167-4186. doi: 10.2147/IJN.S203352.
22. Харченко Е. П. Поиски универсальной противогриппозной вакцины: возможности и ограничения. Эпидемиология и Вакцинопрофилактика. 2019; 18 (5):70–84. doi: 10.31631/2073-3046-2019-18-5-70-84.
23. Ahmed S.S., Volkmuth W., Duca J. et al. Antibodies to influenza nucleoprotein cross-react with human hypocretin receptor 2. Sci. Transl. Med., 2015, vol. 7, no. 294: ra105. doi: 10.1126/scitranslmed.aab2354.
24. Jiang S, Bottazzi ME, Du L et al. Roadmap to developing a recombinant coronavirus S protein receptor-binding domain vaccine for severe acute respiratory syndrome.2012 Expert Review of Vaccines. 2012 Dec;11(12):1405-13. doi: 10.1586/erv.12.126.
25. Харченко, Е. П., Калихевич, В. Н., Соколова, Т. В., Шестак, К. И., Ардемасова, З. А. . Высокоактивные аналоги опиатоподобных пептидов гисторфина и каппорфина. 1989; Вопросы медицинской химии, Т. 35. № 2. С 106–109.
Харченко, Е. П., Калихевич, В. Н., Соколова, Т. В., Шестак, К. И., Ардемасова, З. А. . Высокоактивные аналоги опиатоподобных пептидов гисторфина и каппорфина. 1989; Вопросы медицинской химии, Т. 35. № 2. С 106–109.
Белки состав — Справочник химика 21
Человеческое тело может синтезировать 12 из 20 аминокислот. Остальные восемь должны поступать в организм в готовом виде вместе с белками пищи, поэтому они называются незаменимыми. Незаменимые аминокислоты включают изолейцин, лейцин, лизин, метионин, фенилаланин, треонин, триптофан, валин и (для детей) гистидин. При ограниченном поступлении такой аминокислоты в организм она становится лимитирующим веществом при построении любого белка, в состав которого она должна входить. Если такое случается, то единственное, что может предпринять организм, — это разрушить собственный белок, содержащий эту же аминокислоту. [c. 262]
262]
Важнейшее значение для питания растений имеют азот, фосфор и калий, от которых зависят обмен веществ в растении и его рост. Азот входит в состав белков и хлорофилла, принимает участие в фотосинтезе. Соединения фосфора играют важную роль в дыхании и размножении растений, участвуя в процессах превращения углеводов и азотсодержащих веществ. Калий регулирует жизненные процессы, происходящие в растении, улучшает водный режим, способствует обмену веществ и образованию углеводов в тканях растений. [c.240]
Азот является одним из четырех элементов, входящих в состав молекулы белка, и, подобно сере, содержание его в большинстве нефтей незначительно — до 0,1%. В отдельных фракциях содержание азота увеличивается с температурой кипения последних. [c.104]
Хотя ни один из растительных белков не может обеспечить нас всеми незаменимыми аминокислотами, смеси таких белков — могут. Такие комбинированные продукты питания, которые содержат взаимодополняющие (комплементарные) белки (табл. IV.7), входят в состав традиционной кухни всех народов мира. [c.262]
IV.7), входят в состав традиционной кухни всех народов мира. [c.262]
Минеральными удобрениями называют соли, содержащие элементы, необходимые для питания растений и вносимые в почву для получения высоких и устойчивых урожаев. В состав растений входят около 60 химических элементов. Для образования ткани растения, его роста и развития требуются в первую очередь углерод, кислород и водород, образующие основную часть растительной массы, далее азот, фосфор, калий, магний, сера, кальций и железо. Источниками веществ, необходимых для питания растений, служат воздух и почва. Из воздуха растения извлекают основную массу углерода в виде диоксида углерода, усваиваемого путем фотосинтеза, а из почвы — воду и минеральные вещества. Некоторое количество диоксида углерода воспринимается корневой системой растений из почвы. Среди минеральных веществ особенно важны для жизнедеятельности растений азот, фосфор и калий. Эти элементы способствуют обмену веществ в растительных клетках, росту растений и особенно плодов, повышают содержание ценных веществ (крахмала в картофеле, сахара в све-кле, фруктах и ягодах, белка в зерне), повышают морозостойкость и засухоустойчивость растений, а также их стойкость к заболеваниям. При интенсивном земледелии почва истощается, т. е. в ней резко снижается содержание усваиваемых растениями минеральных веществ, в первую очередь растворимых в воде и почвенных кислотах соединений азота, фосфора и калия. Истощение почвы снижает урожайность и качество сельскохозяйственных культур. Уменьшение содержания питательных веществ в почве необходимо постоянно компенсировать внесением удобрений. Ввиду огромных масштабов потребления минеральные удобрения— наиболее крупнотоннажный вид химической продукции, годовое количество которой составляет десятки миллионов тонн. [c.143]
При интенсивном земледелии почва истощается, т. е. в ней резко снижается содержание усваиваемых растениями минеральных веществ, в первую очередь растворимых в воде и почвенных кислотах соединений азота, фосфора и калия. Истощение почвы снижает урожайность и качество сельскохозяйственных культур. Уменьшение содержания питательных веществ в почве необходимо постоянно компенсировать внесением удобрений. Ввиду огромных масштабов потребления минеральные удобрения— наиболее крупнотоннажный вид химической продукции, годовое количество которой составляет десятки миллионов тонн. [c.143]
В состав белков входят углерод, водород, кислород, азот, v часто сера, фосфор, нелезо. Молекулярные массы белков очень велики — от 1500 до нескольких миллионов. [c.498]
Биологическая роль ДНК. Все признаки организма проявляются через свойства синтезирующихся в нем белков. Состав и структура каждого белка закодированы в отдельных участках молекулы ДНК, которые называются генами. Таким образом, основная функция ДНК — хранение и передача наследственной или генетической информации. [c.218]
Таким образом, основная функция ДНК — хранение и передача наследственной или генетической информации. [c.218]
Ферментами называются белки, входящие в состав клеток и тканей, катализирующие химические реакции, протекающие в организме. Ферменты часто называют биологическими катализаторами. [c.631]
Химический состав мембран прокариот. ЦПМ — белково-липидный комплекс, в котором белки составляют 50—75%, липиды — от 15 до 45%. Кроме того, в составе мембран обнаружено небольшое количество углеводов (табл. 4). Как правило, липиды и белки состав- [c.38]
Для большинства веществ частицы представляют собой молекулы. Молекула — наименьшая частица вещества, обладающая его химическими свойствами. Молекулы в свою очередь состоят из атомов. Атом — наименьшая частица элемента, обладающая его химическими свойствами. В состав молекулы может входить раз личное число атомов. Так, молекулы благородных газов одно-атомны, молекулы таких веществ, как водород, азот,— двухатомны, воды — трехатомны и т. д. Молекулы наиболее сложных веществ — высших белков и нуклеиновых кислот — построены из такого количества атомов, которое измеряется сотнями тысяч. При этом атомы могут соединяться друг с другом не только в различных соотношениях, но и различным образом. Поэтому при сравнительно небольшом числе химических элементов, число различных веществ очень велико. [c.20]
д. Молекулы наиболее сложных веществ — высших белков и нуклеиновых кислот — построены из такого количества атомов, которое измеряется сотнями тысяч. При этом атомы могут соединяться друг с другом не только в различных соотношениях, но и различным образом. Поэтому при сравнительно небольшом числе химических элементов, число различных веществ очень велико. [c.20]
Сера Все белки Входит в состав биологических молекул и ионов [c.277]
Все растительные и животные организмы содержат белковые вещества. Это сложные высокомолекулярные соединения, которые обладают коллоидными свойствами. Независимо от разнообразного строения и различных размеров молекул отдельные белковые вещества имеют очень близкий элементный состав. Некоторые белки содержат фосфор, железо, иод и т. д. [c.25]
Углеводороды представляют собой соединения, включающие только атомы С и Н. Простейшими углеводородами являются линейные полимеры с повторяющейся структурной единицей —СН2—, которые оканчиваются атомами водорода. Другие углеводороды состоят из разветвленных цепей или циклически связанных атомов. Бутан-газ, используемый для отопления и приготовления пищи,-представляет собой тетрамер (четыре структурные единицы). Полимеры, содержащие от 5 до 12 углеродных звеньев, входят в состав бензина одним из примеров является гептан (см. рис. 21-1). Керосин представляет собой смесь молекул, содержащих от 12 до 16 атомов углерода, а смазочные масла и парафиновый воск-смеси цепей с 17 и более атомами углерода. Полиэтилен содержит от 5000 до 50000 мономерных единиц —СН2— в каждой цепи. Существует много других органических цепей, содержащих кроме С и Н еще и другие атомы. Неопреновый каучук, тефлон и дакрон (см. рис. 21-1) являются синтетическими полимерами, а полипептидная цепь, показанная в самой нижней части рис. 21-1, представляет собой полимер, из которого построены все белки-шелк, шерсть, волосы, кол- [c.265]
Другие углеводороды состоят из разветвленных цепей или циклически связанных атомов. Бутан-газ, используемый для отопления и приготовления пищи,-представляет собой тетрамер (четыре структурные единицы). Полимеры, содержащие от 5 до 12 углеродных звеньев, входят в состав бензина одним из примеров является гептан (см. рис. 21-1). Керосин представляет собой смесь молекул, содержащих от 12 до 16 атомов углерода, а смазочные масла и парафиновый воск-смеси цепей с 17 и более атомами углерода. Полиэтилен содержит от 5000 до 50000 мономерных единиц —СН2— в каждой цепи. Существует много других органических цепей, содержащих кроме С и Н еще и другие атомы. Неопреновый каучук, тефлон и дакрон (см. рис. 21-1) являются синтетическими полимерами, а полипептидная цепь, показанная в самой нижней части рис. 21-1, представляет собой полимер, из которого построены все белки-шелк, шерсть, волосы, кол- [c.265]
Углеводы в форме крахмала являются важнейшими источниками энергии в пище. Для получения этой энергии мы либо употребляем в пищу зерна, в которых накапливается крахмал, либо скармливаем эти зерна животным, которые синтезируют мясные белки, а затем съедаем их. В любом случае потребляемая нами энергия в конце концов поставляется крахмалом, полимерным продуктом фотосинтеза. Целлюлоза входит в состав хлопка и льна, а также искусственных продуктов — ацетата целлюлозы и вискозного волокна. Дерево, из которого сделана наша мебель, также содержит целлюлозу. Бумага этой книги получена в процессе обработки целлюлозы. Даже деньги давно перестали делать из благородных металлов, заменив их целлюлозой. В этом разделе будет кратко рассмотрено, что представляют собой углеводы и как они используются. [c.308]
Для получения этой энергии мы либо употребляем в пищу зерна, в которых накапливается крахмал, либо скармливаем эти зерна животным, которые синтезируют мясные белки, а затем съедаем их. В любом случае потребляемая нами энергия в конце концов поставляется крахмалом, полимерным продуктом фотосинтеза. Целлюлоза входит в состав хлопка и льна, а также искусственных продуктов — ацетата целлюлозы и вискозного волокна. Дерево, из которого сделана наша мебель, также содержит целлюлозу. Бумага этой книги получена в процессе обработки целлюлозы. Даже деньги давно перестали делать из благородных металлов, заменив их целлюлозой. В этом разделе будет кратко рассмотрено, что представляют собой углеводы и как они используются. [c.308]
Белковые вещества входят в состав протоплазмы и часто составляют больше половины ее массы. Общее содержание белков в растениях зависит от их принадлежности к тому или иному виду (см. табл. 4). В деревьях оно меньше и колеблется от 1 до 10%. Значительно больше белковых веществ в простых водорослях (20—30%), а в некоторых бактериях их содержание достигает 80%. Молекулярная масса различных белков колеблется в широких пределах от (17500 до 6800000). Изучение белков затруднено тем, что они представляют собой сложные смеси, выделение которых из растений в неизмененном виде почти невозможно. Основной способ выяснения их строения состоит в изучении продуктов их гидролитического распада, осуществленного с помощью минеральных кислот или оснований. Белковые вещества легко гидролизуются не только в присутствии кислот и оснований, но и под действием различных ферментов (протеаз, пепсина, трипсина и др.). При их распаде образуется смесь до 30 различных аминокислот. Большинство из них относится к группе аминокарбоновых кислот, а некоторые имеют ароматический и гидроароматический характер [10, с. 90]. [c.25]
Значительно больше белковых веществ в простых водорослях (20—30%), а в некоторых бактериях их содержание достигает 80%. Молекулярная масса различных белков колеблется в широких пределах от (17500 до 6800000). Изучение белков затруднено тем, что они представляют собой сложные смеси, выделение которых из растений в неизмененном виде почти невозможно. Основной способ выяснения их строения состоит в изучении продуктов их гидролитического распада, осуществленного с помощью минеральных кислот или оснований. Белковые вещества легко гидролизуются не только в присутствии кислот и оснований, но и под действием различных ферментов (протеаз, пепсина, трипсина и др.). При их распаде образуется смесь до 30 различных аминокислот. Большинство из них относится к группе аминокарбоновых кислот, а некоторые имеют ароматический и гидроароматический характер [10, с. 90]. [c.25]
В белках всех живых организмов обычно встречается только 20 различных типов аминокислот, которые указаны в табл. 21-5. Некоторые из них имеют углеводородный состав, например валин (Вал), лейцин (Лей), изолейцин (Иле) и фенилаланин (Фен). Гидрофобные группы молекул всегда более устойчивы, если их можно удалить из водного окружения. Поэтому белковые цепи в водном растворе складываются в молекулы, у которьгх такие группы обращены вовнутрь. Некоторые остатки аминокислот оказываются заряженными например, аспарагиновая (Асп) и глутаминовая (Глу) кислоты входят в белки в ионизованной форме и несут на себе отрицательный заряд, а основания лизин (Лиз) и аргинин (Apr) при pH 7 положительно заряжены. Несмотря на то что некоторые другие группы, например аспарагин (Асн), глутамин (Глу) и серии (Сер), незаряжены, они имеют полярность и поэтому совместимы с водным окружением. Одним из наиболее важных факторов, определяющих свертывание белковой цепи в глобулярную молекулу, является устойчивость, достигаемая при ориентации гидрофобных групп вовнутрь молекулы, а заряженных групп-наружу. Хотя каждый из двух оптических изомеров, показанных на рис.
21-5. Некоторые из них имеют углеводородный состав, например валин (Вал), лейцин (Лей), изолейцин (Иле) и фенилаланин (Фен). Гидрофобные группы молекул всегда более устойчивы, если их можно удалить из водного окружения. Поэтому белковые цепи в водном растворе складываются в молекулы, у которьгх такие группы обращены вовнутрь. Некоторые остатки аминокислот оказываются заряженными например, аспарагиновая (Асп) и глутаминовая (Глу) кислоты входят в белки в ионизованной форме и несут на себе отрицательный заряд, а основания лизин (Лиз) и аргинин (Apr) при pH 7 положительно заряжены. Несмотря на то что некоторые другие группы, например аспарагин (Асн), глутамин (Глу) и серии (Сер), незаряжены, они имеют полярность и поэтому совместимы с водным окружением. Одним из наиболее важных факторов, определяющих свертывание белковой цепи в глобулярную молекулу, является устойчивость, достигаемая при ориентации гидрофобных групп вовнутрь молекулы, а заряженных групп-наружу. Хотя каждый из двух оптических изомеров, показанных на рис. 21-12, пред- [c.314]
21-12, пред- [c.314]
Белки имеют и другие свободные группы и радикалы (—ОН, —5Н, —5—5—), которые позволяют им реагировать со многими химическими соединениями, входящими в состав растений-угле-образователей, и с продуктами их распада. Многие авторы считают, что азот, сера и фосфор, содержащиеся во всех твердых топливах, являются результатом превращения белков, входящих в состав исходных растительных остатков. [c.26]
Круговорот серы в природе поддерживается микроорганизмами. При их участии сульфиды окисляются до сульфатов, сульфаты поглощаются живыми организмами, где сера восстанавливается и входит в состав белков. При гниении отмерших организмов сера возвращается в круговорот. [c.113]
Азот входит в состав разнообразных органических соединений — аминов, аминокислот, белков и многих других. [c.124]
Высокомолекулярные соединения подразделяют на природные и синтетические. К важнейшим природным полимерам относятся белки и полисахариды. Белки являются основой всего живого, они составляют существенную часть живой клетки и обеспечивают ее жизнедеятельность. Белки входят в состав кожи, мышц, сухожилий, нервов и крови, а также ферментов и гормонов, содержатся. во многих растительных и животных продуктах молоке, яйцах, зернах пшеницы, бобах и др. К белкам относятся широко применяемые в технике желатина, козеии, яичный альбумин. Из нерастворимых белков наиболее известны шерсть и шелк, отличающиеся волокнистым строением. [c.307]
Белки являются основой всего живого, они составляют существенную часть живой клетки и обеспечивают ее жизнедеятельность. Белки входят в состав кожи, мышц, сухожилий, нервов и крови, а также ферментов и гормонов, содержатся. во многих растительных и животных продуктах молоке, яйцах, зернах пшеницы, бобах и др. К белкам относятся широко применяемые в технике желатина, козеии, яичный альбумин. Из нерастворимых белков наиболее известны шерсть и шелк, отличающиеся волокнистым строением. [c.307]
Ксантопротеиновая реакция позволяет обнаруживать и белки, состав которых входят остатки ароматических аминокислот. Кстг ти, желтая окраска на коже при попадании на нее азотной кислоТ обусловлена именно этой реакцией. [c.412]
Степень точности анализов естественных неоднородных белков, состав которых может колебаться в зависи.мости от условий их выделения и от различий в составе аминокислот в тканях или организмах, не обязательно должна быть столь большой, как в случае тех не. многих белков, гомогенность которых была доказана. Хотя в аналитической работе надо бороться за абсолютные цифры, но можно получить много полезных сведений также и путем сравнительного а.минокислотного анализа, особенно в вопросах белкового питания. [c.8]
многих белков, гомогенность которых была доказана. Хотя в аналитической работе надо бороться за абсолютные цифры, но можно получить много полезных сведений также и путем сравнительного а.минокислотного анализа, особенно в вопросах белкового питания. [c.8]
Альбумины и глобулины сыворотки представляют очень сложную смесь белков, состав которой изменяется в зави9имости от метода приготовления. Серумальбумины являются превосходным источником для получения лизина. Отсюда логически следует, что в тех случаях, когда в организме происходит восстановление альбумина сыворотки, пищ,а должна быть богата лизином. [c.74]
Другой большой класс белков образуют фибриллярные белки. Они выполняют в организме главным образом роль структурных материалов. К их числу относится кератин, входящий в состав кожи, волос, шерсти, ногтей и других роговых тканей. К другому типу фибриллярных белков относится коллаген, находяищйся в сухожилиях, подкожном слое и роговице глаз к фибриллярным относятся белки шелка и тканей насекомых. Белки, углеводы и липиды (жиры с длинными цепями и жирные кислоты) играют роль строительных материалов в любых живых организмах. [c.313]
Белки, углеводы и липиды (жиры с длинными цепями и жирные кислоты) играют роль строительных материалов в любых живых организмах. [c.313]
КО на неферментные белки иногда может приходиться значительная часть общего белка. Так, например, на долю двух запасных глобулинов в клетках семядолей гороха приходится более 80% общего количества белка (гл. 29) другие примеры подобного рода — глиадин пшеницы и гордеин ячменя. Каково же происхождение таких запасных белков Состав глобулинов семядолей гороха не представляет ничего необычного для белков. Может быть, глобулины — ферменты, которые утратили свои активные центры Мутации, приводящие к нарушению активного центра фермента, могут и не препятствовать синтезу ставшего неактивным белка. А если синтез фермента контролировался путем репрессии продуктом катализируемой реакции, то тем в больших количествах могли бы образоваться молекулы фермента, уже не обладающие ферментативной активностью. Однако состав глиадина и гордеина в достаточной мере необычен (40% глутамина и 14% пролина). Поэтому трудно представить, что они также возникли в результате утраты активного центра, но что впоследствии они сильно изменились, превратившись в эффективную форму запаса углерода и азота в легко доступном для растения виде. Оболочки меристематических клеток, а также клеток, выросших в культуре ткани, содержат до 40% общего белка клетг п [c.16]
Поэтому трудно представить, что они также возникли в результате утраты активного центра, но что впоследствии они сильно изменились, превратившись в эффективную форму запаса углерода и азота в легко доступном для растения виде. Оболочки меристематических клеток, а также клеток, выросших в культуре ткани, содержат до 40% общего белка клетг п [c.16]
Вёлер, Кольбе и Бертло синтезировали относительно простые органические соединения, тогда как для живой природы характерны значительно более сложные соединения типа крахмала, жиров и белков. Изучать такие соединения гораздо труднее непросто даже установить их точный элементный состав. В целом изучение органических веществ обещало разгадку многих проблем, но подступиться к этим веществам химику прошлого века было совсем непросто. [c.71]
Групповой химический состав растений. Все живые организ — мы состоят в основном из следующих четырех классов органических веществ углеводов, липидов, белков и лигнина. [c.47]
[c.47]
Обычно в состав простетических групп в растительных и животных системах входят порфириновые ядра, представляющие собой хелатные структуры с включением ионов металлов (Ре , Со «, и т. д.). Так, гемоглобин животных содержит такую группу с Ре » , присоединенную к белковой половине (глобин). Эта группа аналогична по структуре простетической группе, содержащей в хлорофилле растений и одноклеточных животных. Молекулярный вес белков обычно лежит в пределах от 30 ООО до 80 ООО. Однако молекулярный вес может быть и меньше и значительно больше этих величин. Ферменты являются очень специфичными катализаторами. Зачастую их активность может проявляться только в какой-либо одной реакции. Так, например, фумараза катализирует только обратимую реакцию превращения малеиновой кислоты в фумаровую [98] [c.561]
Химический состав опорных тканей позвоночных отличается от состава скелетных тканей беспозвоночных — спонгина, хитина и др. В покровах позвоночных присутствует особый белок — кератин.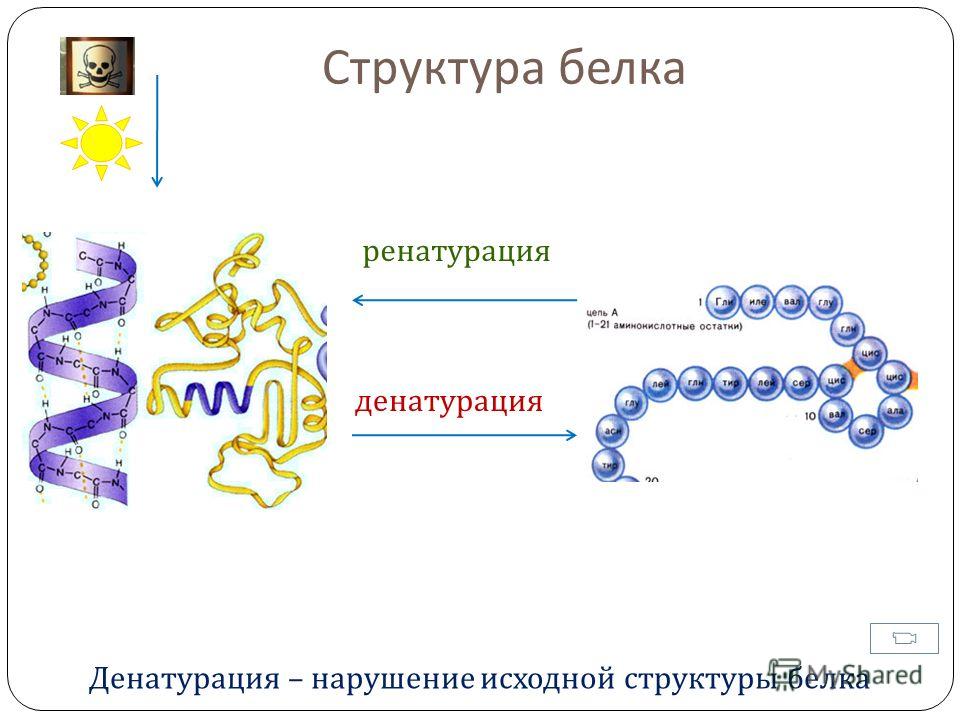 Позвоночные отличаются от беспозвоночных и действием пищерастительных ферментов, более высоким отношением (Ма + К)/ Са + Мд) в жидкой фазе внутренней среды. Среди беспозвоночных только у оболочников есть целлюлозная оболочка, имеется ванадий в крови в особых окрашенных клетках, а у круглоротых — соединительно-тканный скелет и хрящ, а также особый дыхательный пигмент — аритрокруорин с наименьшей для позвоночных молекулярной массой (17 600). Отличительная черта сипункулид — древних групп морских беспозвоночных — наличие специального переносчика кислорода — гемэритрина и наличие в эритроцитах значительного количества аллантоиновой кислоты. Для насекомых характерно высокое содержание в крови аминокислот, мочевой кислоты и редуцирующих и несбраживаемых веществ, в хитиновом покрове отсутствуют смолы, для членистоногих — наличие специфической (только для их групп) фенолазы в крови. Таким образом, можно констатировать, что систематические группы животных имеют свои биохимические особенности. Такие же особенности наблюдаются и у растений для различных систематических групп — наличие специфических белков, жиров, углеводов, алкалоидов, глюкозидов, ферментных систем.
Позвоночные отличаются от беспозвоночных и действием пищерастительных ферментов, более высоким отношением (Ма + К)/ Са + Мд) в жидкой фазе внутренней среды. Среди беспозвоночных только у оболочников есть целлюлозная оболочка, имеется ванадий в крови в особых окрашенных клетках, а у круглоротых — соединительно-тканный скелет и хрящ, а также особый дыхательный пигмент — аритрокруорин с наименьшей для позвоночных молекулярной массой (17 600). Отличительная черта сипункулид — древних групп морских беспозвоночных — наличие специального переносчика кислорода — гемэритрина и наличие в эритроцитах значительного количества аллантоиновой кислоты. Для насекомых характерно высокое содержание в крови аминокислот, мочевой кислоты и редуцирующих и несбраживаемых веществ, в хитиновом покрове отсутствуют смолы, для членистоногих — наличие специфической (только для их групп) фенолазы в крови. Таким образом, можно констатировать, что систематические группы животных имеют свои биохимические особенности. Такие же особенности наблюдаются и у растений для различных систематических групп — наличие специфических белков, жиров, углеводов, алкалоидов, глюкозидов, ферментных систем. [c.189]
[c.189]
В данном случае и кислотные, и основные функции определяются свойствами одной и той же группы ОН . Но существуют ам-фолиты и другого типа. Их кислотные и основные свойства определяются нал 1чием двух различных функциональных групп. Наиболее характерным примером соединений подобного типа М01 ут служить аминокислоты ЫНгНСООН. Аминокислоты входят и состав белков, поэтому исследование последних невозможно без учета явлений, обусловленных амфотерными свойствами аминокислот. [c.509]
Все многообразие белков образовано 20 различными аминокис-атами при этом для каждого белка строго специфичной является оследовательность, в которой остатки входящих в его состав ами-окислот соединяются друг с другом. Найдены методы выяснения гой последовательности в результате уже точно установлено троение некоторых белков. И самым замечательным достижением этой области явилось осуществление синтеза кз аминокислот ростейших белков как уже указывалось, в 50—60- гормон инсулин и фермент рибонуклеаза. аким образом, доказана принципиальная возможность синтеза ще более сложных белков. [c.499]
аким образом, доказана принципиальная возможность синтеза ще более сложных белков. [c.499]
Ионизирующая радиация разрушает химические связи и таким образом расщепляет молекулу. При низком уровне радиации молекула разрушается немного, и систем1Я организма могут ликвидировать опасность. Однако при большей дозе, попющинной организмом, повреждается слишком много молекул. Наибольшую опасность представляют повреждения белков и нуклеиновых кислот. Белки охл-авляют основу мягких тканей организма, и входят в состав ферментов, молекул, контролирующих все жизненные процессы. Когда большое число молекул в непосредственной близости друг от друга разрушаются, организму н( хв 1тает молекул для поддержания жизнедеятельности. [c.353]
В особую группу следует выделить синтезы на основе оксида углерода, водорода и азота метанола (3 процесса), муравьиной кислоты (2 процесса), метиламинов (2 процесса), метилформиата, аммиака (4 процесса), нитрата аммония (2 процесса), азотной кислоты (2 процесса), карбамида и одноклеточных белков. В каталог современных нефтехимических процессов последняя группа синтезов входит вследствие привязки к нефтяному углеводородному сырью через процессы конверсии метана и жидких нефтяных дистиллятов в оксид углерода н водород. Главным ядром данной группы процессов являются метанол и аммиак, которые потребляются в значительных количествах для производства эфиров различных алифатических и ароматических кислот, а также, аминонроизводных, поэтому входят в состав нефтехимической продукции и нефтехимического сырья. [c.358]
В каталог современных нефтехимических процессов последняя группа синтезов входит вследствие привязки к нефтяному углеводородному сырью через процессы конверсии метана и жидких нефтяных дистиллятов в оксид углерода н водород. Главным ядром данной группы процессов являются метанол и аммиак, которые потребляются в значительных количествах для производства эфиров различных алифатических и ароматических кислот, а также, аминонроизводных, поэтому входят в состав нефтехимической продукции и нефтехимического сырья. [c.358]
Полисахариды (полимерные углеводы) представляЕот собой соединения, состоящие из многих сотен нли даже тысяч моносаха-ридных звеньев. Их состав отвечает общей формуле (СеНюОз) . Наиболее важными среди полисахаридов являются целлюлоза и крахмал. Оба эти вещества образуются в растениях из диоксида углерода и воды в результате фотосинтеза. Целлюлоза — основной строительный материал растений, крахмал служит запасным пищевым фондом растений и находится в основном в семенах (кукуруза, картофель, рис, пшеница и др. ). Углеводы служат источником питания человека. В организме человека и животных они превращаются в жиры и белки. Целлюлоза в виде хлопка и вискозы применяется для изготовления одежды и бумаги. [c.307]
). Углеводы служат источником питания человека. В организме человека и животных они превращаются в жиры и белки. Целлюлоза в виде хлопка и вискозы применяется для изготовления одежды и бумаги. [c.307]
Сколько аминокислот входит в состав белков, как они называются?
Отказ от ответсвенности
Обращаем ваше внимание, что вся информация, размещённая на сайте
Prowellness предоставлена исключительно в ознакомительных целях и не является персональной программой, прямой рекомендацией к действию или врачебными советами. Не используйте данные материалы для диагностики, лечения или проведения любых медицинских манипуляций. Перед применением любой методики или употреблением любого продукта проконсультируйтесь с врачом. Данный сайт не является специализированным медицинским порталом и не заменяет профессиональной консультации специалиста. Владелец Сайта не несет никакой ответственности ни перед какой стороной, понесший косвенный или прямой ущерб в результате неправильного использования материалов, размещенных на данном ресурсе.
Сколько аминокислот входит в состав белков, как они называются?
Белок является главным строительным материалом для организма. Его структура сложная и многоступенчатая. Какие аминокислоты входят в его состав?
Белок очень важен для организма, так как является строительным материалом. Его основой являются аминокислоты. Эти вещества отвечают за разные функции и нужны для поддержания нормального состояния организма.
Основные свойства аминокислот
Аминокислоты обладают следующими свойствами:
- Быстро и хорошо растворяются в жидкой среде.
- По форме напоминают кристаллы.
- При воздействии высоких температур могут расплавиться.
- Имеют некоторые признаки кислот и оснований одновременно.
- Получаются в процессе белкового гидролиза.
Для чего аминокислоты нужны организму?
Биологическая роль аминокислот заключается в следующем:
- обеспечивают правильную работу иммунной системы;
- синтезируют глюкозу и участвуют в углеводном обмене;
- выступают в роли строительного материала для мышц и сухожилий;
- помогают спортсменам при наборе мышечной массы;
- поддерживают здоровье соединительных тканей;
- помогают поврежденным тканям быстрее восстановиться;
- участвуют в выработке энергии;
- выводят из организма вредные вещества и токсины;
- участвуют в образовании гормонов;
- поддерживают печень в порядке;
- помогают поддерживать нормальную работу головного мозга;
- обеспечивают бодрость духа и хорошее настроение;
- повышают работоспособность человека и его творческий потенциал;
- помогают нервным клеткам защититься от вредного воздействия алкогольных напитков;
- улучшают психическое здоровье человека;
- участвуют в жировом обмене;
- поддерживают работу органов желудочно-кишечного тракта;
- регулируют работу щитовидной железы;
- поддерживают в норме массу тела;
- замедляют естественные процессы старения.

Сколько аминокислот входит в состав белков?
В состав белков входят следующие незаменимые аминокислоты:
- Лейцин, нужный для набора мышечной массы и контроля массы тела.
- Изолейцин, стимулирующий выделение энергии.
- Лизин, отвечающий за укрепление иммунитета и повышение защитных сил организма.
- Фенилаланин, обеспечивающий правильную работу центральной нервной системы.
- Метионин, ответственный за эффективное и быстрое сжигание подкожного жира.
- Треонин, оказывающий положительное влияние на центральную нервную систему.
- Триптофан, формирующий полезные для жизнедеятельности гормоны.
- Валин, ускоряющий процессы обмена веществ.
Также белок образуют несколько заменимых аминокислот. К ним относятся:
- Аланин, необходимый для процессов углеводного обмена и выведения из организма токсических веществ.
- Аспарагиновая кислота, обеспечивающая человеку энергичность и прилив бодрости.
- Аспарагин, обеспечивающий работу центральной нервной системы и головного мозга.
- Гистидин, вырабатывающий кровяные тельца красного цвета.
- Серин, отвечающий за правильную и эффективную работу головного мозга и за протекание когнитивных процессов.
- Цистеин, подающий в организм кератин.
- Аргинин, оздоравливающий кожу, кости, мышечную ткань и сухожилия.
- Глютаминовая кислота, без которой невозможна нормальная работа головного и спинного мозга.
- Глютамин, предотвращающий атрофию мышечных волокон.
- Глицин, ускоряющий процессы свертываемости крови.
- Пролин, стимулирующий выработку коллагена.
- Тирозин, поддерживающий в норме аппетит и артериальное давление.
Внимание! В состав белков входят разные аминокислоты, заменимые и незаменимые.Они обеспечивают правильное формирование протеинов и эффективную работу всего организма.
Отказ от ответсвенности
Обращаем ваше внимание, что вся информация, размещённая на сайте
Prowellness предоставлена исключительно в ознакомительных целях и не является персональной программой, прямой рекомендацией к действию или врачебными советами. Не используйте данные материалы для диагностики, лечения или проведения любых медицинских манипуляций. Перед применением любой методики или употреблением любого продукта проконсультируйтесь с врачом. Данный сайт не является специализированным медицинским порталом и не заменяет профессиональной консультации специалиста. Владелец Сайта не несет никакой ответственности ни перед какой стороной, понесший косвенный или прямой ущерб в результате неправильного использования материалов, размещенных на данном ресурсе.
10.
 Химический состав клетки (белки, их структура и функции).
Химический состав клетки (белки, их структура и функции).
Белки,
или протеины, составляюи ри 50 до 85%
органических соединений, входящих в
состав живых организмов. Во всех тканях
любых существ важнейшей частью являются
белки. Они входят в состав всех клеток,
клеточных органоидов и межклеточных
жидкостей. Основными элементами белка
являются: кислород, водород, азот и сера.
Кроме того, в их состав могут входить
фосфор, железо, магний и другие.
Молекула
белка — типичный полимер, она состоит
из аминокислот. При соединении аминокислот
в молекуле белка образуется химическая
связь между карбоксильной группой
аминокислоты и аминной группой другой.
Связь которая образуется между молекулами
аминокислота, называется пептидной.
Белки имеют 4 структуры белка:
Первичной
структурой белковой молекулы является
полипептидная цепь.
Внутримолекулярные
силы заставляют цепь изгибаться –
возникает вторичная структура. Молекула
Молекула
белка бывает складчатой и спиральной.
Складчатая структура при этом характерна
для белков с низким метаболизмом.
Большинству
белковых молекул присуща третичная
структура, получавшая название третичной.
Полипептидные цепи скручиваются, образуя
глобулу.
Группы белковых молекул
образуют устойчивые комплексы, которые
называются четвертичными структурными.
Функции
белков. В клетке белки выполняют
структурную, сократительную. Ферментативную
функции. Структурная функция выражается
в том, что белки – основной строительный
материал цитоплазмы, наружной и внутренней
мембран – входят в состав хромосом и
других органоидов клетки. Сократительная
функция обеспечивает одно из основных
свойств жизни – явления раздражительности
и движения. С ферментативной функцией
белков связано то, что они катализируюи
все реакции, протекающие в организме.
11. Нуклеиновые кислоты, их строение, локализация, значение
Простейшие
нуклеиновые кислоты – мононуклеотиды.
Более сложные нуклеиновые кислоты
состоят из двух или более нуклеотидов
– полинуклеотиды.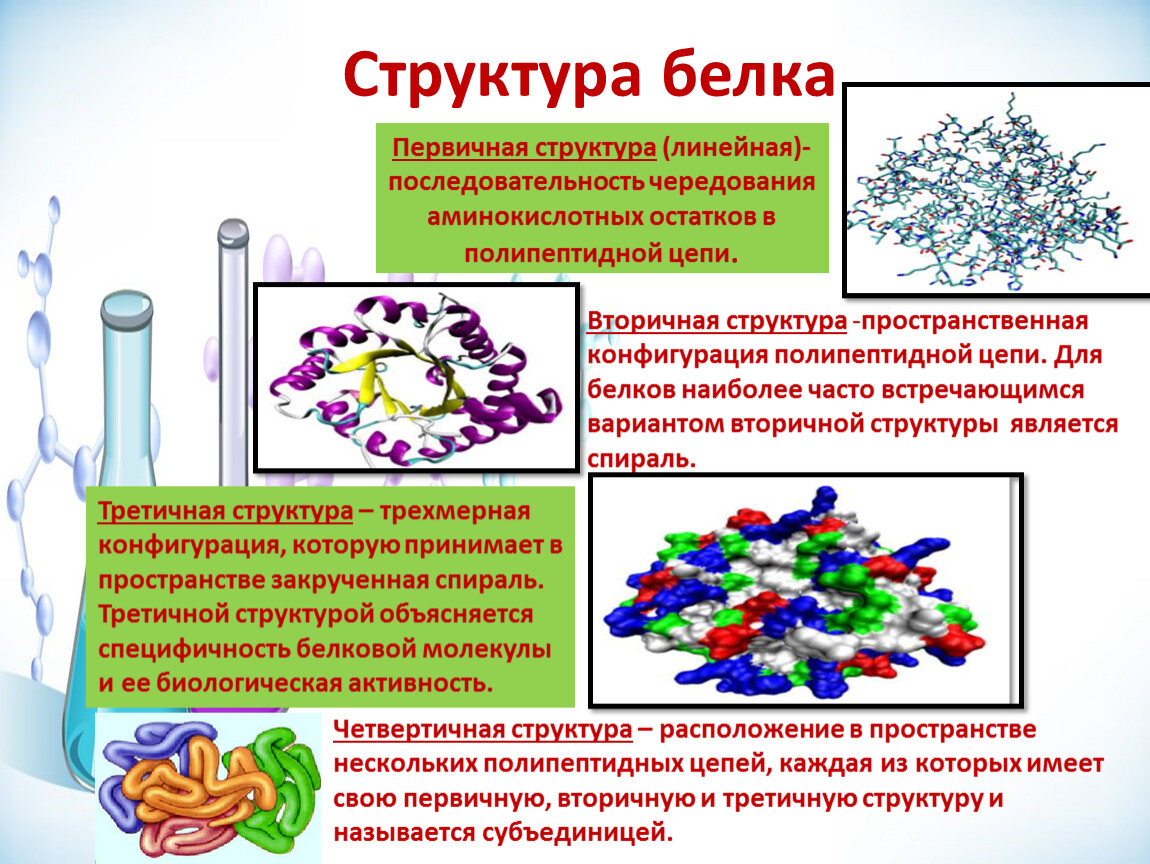 В состав нуклеиновых
В состав нуклеиновых
кислот входят углерод, кислород, водород,
азот и фосфор. Известны 2 типа нуклеиновых
кислот: ДНК и РНК. Они отличаются и
строением и биологическими свойствами.
ДНК и РНК в клетке имеют различную
локализацию. ДНК имеется в ядре, входит
в состав хроматина, сосредоточена в
хромосомах, имеется внутри митохондрий
и пластид. В ядре ДНК вступает соединение
с гистонами и протаминами, образуя
нуклепротеиды. Основные хранители РНК
– ядрышки, находящиеся в ядре, и рибосомы,
расположенные в цитоплазме. Кроме того,
РНК находится в гиалоплазме.
В состав
нуклеотида входит молекула фосфорной
кислоты, моносахарида и 4 азотистых
оснований: Аденин, Гуанин, Цитозин, Тимин
или Урацил.
РНК содержит моносахарид
рибозу, в то время как в состав ДНК входит
дезоксирибоза. Азотистые основания
аденин, гуанин, цитозин есть в составе
как ДНК, так и РНК, но тимин входит в
состав ДНК, а урацил – в состав РНК.
С
нуклеиновыми кислотами связаны процессы
синтеза белка, а этим в свою очередь
определяется характер обмена веществ,
закономерности роста и развития, явления
наследственности и изменчивости.
12. Роль
ДНК и РНК в передаче наследственной
информации. Основные этапы: транскрипция,
процессинг, трансляция.
Главную
роль в процессе передачи и реализации
наследственной информации играют
нуклеиновые кислоты. Основная биологическая
функция ДНК заключается в хранении,
постоянном самовозобновлении,
самовоспроизведении и передаче
генетической информации клетке.
Информация хранится в последовательности
нуклеотидов. Эта последовательность
нуклеотидов, или генетический код,
контролирует последовательность
аминокислот в молекуле белка. ДНК
является матрицей для построения иРНК.
ДНК принимает участие только в одном
этапе биосинтеза белка: транскрипции.
Транскрипция
– процесс переноса генетического кода,
записанного на молекуле ДНК на молекулу
иРНК. Транскрипция происходит при
синтезе молекул иРНК, нуклеотиды которой
присоединяются к нуклеотидам ДНК по
принципу комплементарности. Молекула
иРНК снимается с ДНК, как с матрицы,
после чего она отделяется и перемещается
в цитоплазму, где в специальных органоидах
– рибосомах происходит процесс
трансляции.
Непосредственное участие
в синтезе белка принимает иРНК.
Биологическая роль иРНК связана
преимущественно синтезом белка, т.е.
реализацией наследственной информации.
Именно РНК является посредником между
ДНК и строящейся в клетке белковой
молекулой. Выделяют иРНК, тРНК и рРНК.
иРНК обеспечивает перенос информации
о структуре белка от молекулы ДНК в
рибосомы, где синтезируется белок. рРНК
содержится в рибосомах и участвует в
синтезе белка. тРНК доставляет аминокислоты
к месту синтеза белка, т.е. к рибосомам.
Трансляция – процесс перевода
генетической информации, записанной
на иРНК в структуру белковой молекулы,
синтезируемой на рибосомах при участии
тРНК. На иРНК генетический код записан
«языком» триплетов нуклеотидов. Они
передают информацию только тем тРНК,
кодовый триплет которых комплементарен
триплету иРНК. При образовании связи
между кодовыми триплетами происходит
передача информации и аминокислота
присоединяется к цепочке белковой
молекулы.
Структура белка | Изучайте науку в Scitable
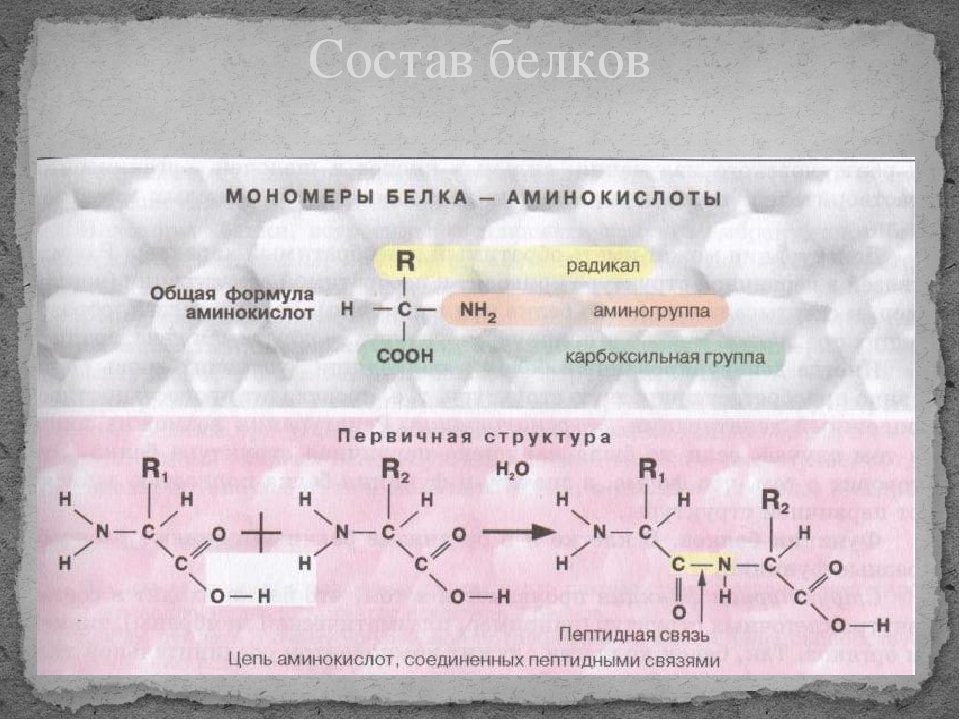
Строительными блоками белков являются аминокислоты, которые представляют собой небольшие органические молекулы, которые состоят из альфа (центрального) атома углерода, связанного с аминогруппой, карбоксильной группы, атома водорода и вариабельного компонента, называемого боковой цепью (см. Ниже ). Внутри белка несколько аминокислот связаны между собой пептидными связями , тем самым образуя длинную цепь. Пептидные связи образуются в результате биохимической реакции, которая извлекает молекулу воды, поскольку она соединяет аминогруппу одной аминокислоты с карбоксильной группой соседней аминокислоты.Линейная последовательность аминокислот в белке считается первичной структурой белка.
Ниже ). Внутри белка несколько аминокислот связаны между собой пептидными связями , тем самым образуя длинную цепь. Пептидные связи образуются в результате биохимической реакции, которая извлекает молекулу воды, поскольку она соединяет аминогруппу одной аминокислоты с карбоксильной группой соседней аминокислоты.Линейная последовательность аминокислот в белке считается первичной структурой белка.
Белки состоят из набора всего из двадцати аминокислот, каждая из которых имеет уникальную боковую цепь. Боковые цепи аминокислот имеют разный химический состав. Самая большая группа аминокислот имеет неполярные боковые цепи. Некоторые другие аминокислоты имеют боковые цепи с положительными или отрицательными зарядами, а другие имеют полярные, но незаряженные боковые цепи. Химический состав боковых цепей аминокислот имеет решающее значение для структуры белка, потому что эти боковые цепи могут связываться друг с другом, чтобы удерживать длину белка в определенной форме или конформации. Боковые цепи заряженных аминокислот могут образовывать ионные связи, а полярные аминокислоты способны образовывать водородные связи. Гидрофобные боковые цепи взаимодействуют друг с другом посредством слабых ван-дер-ваальсовых взаимодействий. Подавляющее большинство связей, образованных этими боковыми цепями, нековалентны. Фактически, цистеины — единственные аминокислоты, способные образовывать ковалентные связи, что они и делают со своими конкретными боковыми цепями. Из-за взаимодействий боковых цепей последовательность и расположение аминокислот в конкретном белке определяют, где в этом белке происходят изгибы и складки (рис. 1).
Боковые цепи заряженных аминокислот могут образовывать ионные связи, а полярные аминокислоты способны образовывать водородные связи. Гидрофобные боковые цепи взаимодействуют друг с другом посредством слабых ван-дер-ваальсовых взаимодействий. Подавляющее большинство связей, образованных этими боковыми цепями, нековалентны. Фактически, цистеины — единственные аминокислоты, способные образовывать ковалентные связи, что они и делают со своими конкретными боковыми цепями. Из-за взаимодействий боковых цепей последовательность и расположение аминокислот в конкретном белке определяют, где в этом белке происходят изгибы и складки (рис. 1).
Рис. 1. Взаимосвязь между боковыми цепями аминокислот и конформацией белка
Определяющим признаком аминокислоты является ее боковая цепь (вверху — синий кружок; внизу — все цветные кружки). Когда аминокислоты соединяются серией пептидных связей, они образуют полипептид, другое слово для обозначения белка. Затем полипептид сворачивается в определенную конформацию в зависимости от взаимодействий (пунктирные линии) между его боковыми аминокислотными цепями.
Рисунок 2: Структура белка бактериородопсина
Бактериородопсин — это мембранный белок бактерий, который действует как протонный насос. Его форма важна для его функции. Общая структура белка включает как альфа-спирали (зеленый), так и бета-листы (красный).
Первичная структура белка — его аминокислотная последовательность — управляет складыванием и внутримолекулярным связыванием линейной аминокислотной цепи, что в конечном итоге определяет уникальную трехмерную форму белка. Водородная связь между аминогруппами и карбоксильными группами в соседних областях белковой цепи иногда вызывает определенные паттерны сворачивания. Эти стабильные паттерны сворачивания, известные как альфа-спирали и бета-листов , составляют вторичную структуру белка.Большинство белков содержат несколько спиралей и листов в дополнение к другим, менее распространенным паттернам (рис. 2). Совокупность образований и складок в единой линейной цепи аминокислот — иногда называемой полипептидом — составляет третичную структуру белка.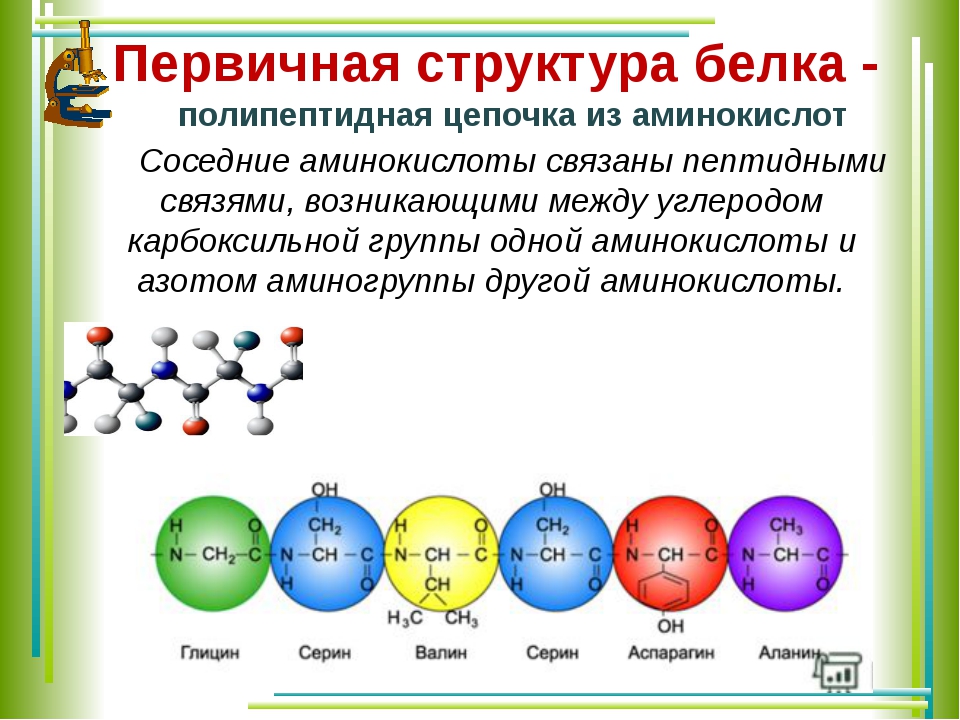 Наконец, четвертичная структура белка относится к тем макромолекулам с множеством полипептидных цепей или субъединиц.
Наконец, четвертичная структура белка относится к тем макромолекулам с множеством полипептидных цепей или субъединиц.
Окончательная форма, принятая вновь синтезированным белком, обычно является наиболее энергетически выгодной.По мере сворачивания белков они проверяют различные конформации, прежде чем достичь своей окончательной формы, которая является уникальной и компактной. Сложенные белки стабилизированы тысячами нековалентных связей между аминокислотами. Кроме того, химические силы между белком и его непосредственным окружением способствуют формированию и стабильности белка. Например, белки, которые растворены в цитоплазме клетки, имеют на своей поверхности гидрофильные (водолюбивые) химические группы, тогда как их гидрофобные (водоотталкивающие) элементы имеют тенденцию скрываться внутри.Напротив, белки, которые вставлены в клеточные мембраны, имеют на своей поверхности некоторые гидрофобные химические группы, особенно в тех областях, где поверхность белка подвергается воздействию липидов мембран.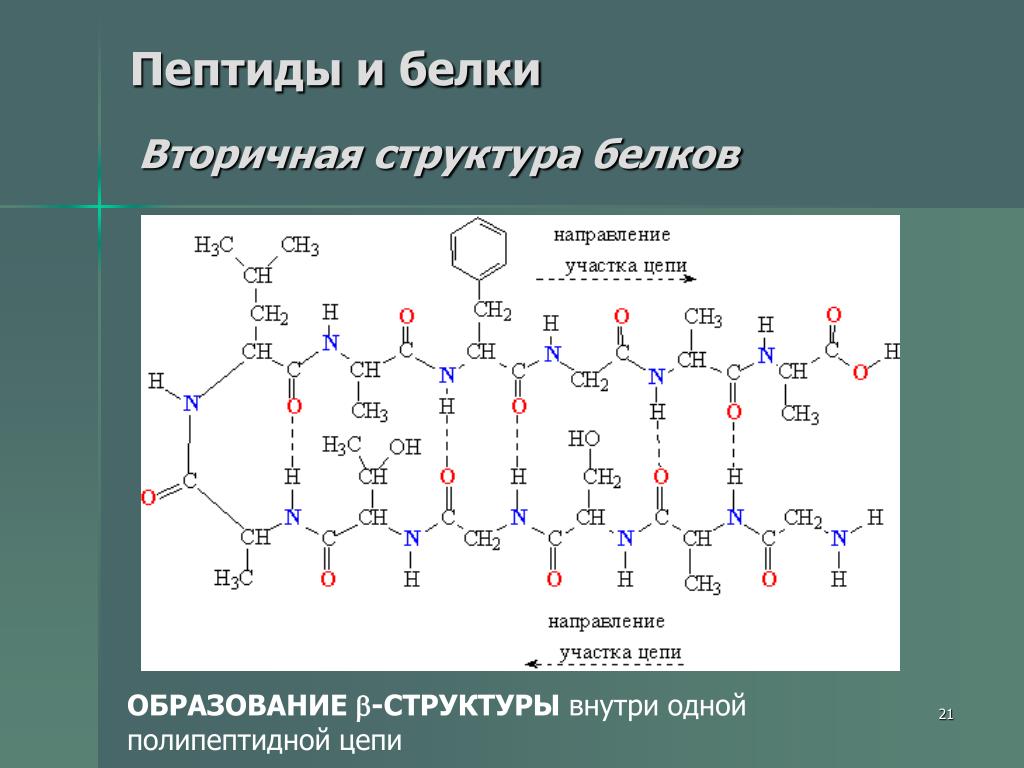 Однако важно отметить, что полностью свернутые белки не принимают форму. Скорее, атомы в этих белках остаются способными совершать небольшие движения.
Однако важно отметить, что полностью свернутые белки не принимают форму. Скорее, атомы в этих белках остаются способными совершать небольшие движения.
Несмотря на то, что белки считаются макромолекулами, они слишком малы, чтобы их можно было визуализировать даже в микроскоп.Итак, ученые должны использовать косвенные методы, чтобы выяснить, как они выглядят и как сложены. Наиболее распространенным методом исследования структуры белков является рентгеновская кристаллография . С помощью этого метода твердые кристаллы очищенного белка помещают в пучок рентгеновских лучей, а диаграмма отклоненных рентгеновских лучей используется для прогнозирования положений тысяч атомов в кристалле белка.
Состав и структура белка
— Biology LibreTexts
-
- Последнее обновление
- Сохранить как PDF
- Вторичная структура полипептидной цепи
- График Рамачандрана
- α-Спирали
- α-спираль
- β-ПЛИТИРОВАННЫЕ ЛИСТЫ
- α-ПЛИТИРОВАННЫЕ ЛИСТЫ
- Третичная структура
- Четвертичная структура
кодирует ДНК цепочка одномерных аминокислот, которые переводятся в трехмерные белки. Белки имеют первичную, вторичную и третичную структуры. Первичная структура состоит из аминокислот, которые связаны пептидными связями и образуют линейные цепи полипептидов. Вторичная структура состоит из полипептидных цепей, которые складываются в трехмерные структуры. Третичные структуры — это водорастворимые белки, которые конденсируются в уплотненные структуры.
Белки имеют первичную, вторичную и третичную структуры. Первичная структура состоит из аминокислот, которые связаны пептидными связями и образуют линейные цепи полипептидов. Вторичная структура состоит из полипептидных цепей, которые складываются в трехмерные структуры. Третичные структуры — это водорастворимые белки, которые конденсируются в уплотненные структуры.
Белковые структуры
• Первичная структура (линейный полимер аминокислот)
(удерживается вместе пептидными связями)
• Вторичная структура (стандартные трехмерные модели)
(a-спираль, ß-лист, удерживается вместе с Н-связями между атомами основной цепи)
• Третичная структура (детальная трехмерная конформация)
(связи между атомами боковых цепей)
• Четвертичная структура (комбинированные полимерные цепи)
Вторичная структура полипептидной цепи
Вторичная структура относится к форме сворачивающегося белка, обусловленной исключительно водородными связями между его амидной и карбонильной группами основной цепи. Вторичная структура не включает связывание между R-группами аминокислот, гидрофобные взаимодействия или другие взаимодействия, связанные с третичной структурой.
Вторичная структура не включает связывание между R-группами аминокислот, гидрофобные взаимодействия или другие взаимодействия, связанные с третичной структурой.
Две наиболее часто встречающиеся вторичные структуры полипептидной цепи — это альфа-спирали и бета-складчатые листы. Эти структуры являются первыми основными этапами сворачивания полипептидной цепи, и они устанавливают важные топологические мотивы, которые определяют последующую третичную структуру и конечную функцию белка.
Пептидные связи влияют на вторичную структуру
Напомним, что плоская амидная связь ограничивает плоскости изгиба цепи: вращения вокруг связей CO-N нет, но плоскости вращаются вокруг связей
α-CN (Φ) и α-CC = O связей (Ψ)
График Рамачандрана
Показывает группировку φψ и связывает их со структурами в реальных белках. Часто встречаются повторяющиеся структуры (α-спирали, β-листы)
http://commons.wikimedia.org/wiki/Fi …neral_100K.jpg
α-Спирали
Альфа-спираль представляет собой правостороннюю спираль аминокислотных остатков на полипептидной цепи, обычно в диапазоне от 4 до 40 остатков. Этот змеевик удерживается вместе водородными связями между кислородом C = O на верхнем витке и водородом N-H на нижнем витке. Такая водородная связь образуется ровно через каждые 4 аминокислотных остатка, а каждый полный виток спирали составляет всего 3,6 аминокислотных остатка. Этот регулярный узор придает альфа-спирали очень определенные характеристики в отношении толщины катушки и длины каждого полного витка вдоль оси спирали.
Этот змеевик удерживается вместе водородными связями между кислородом C = O на верхнем витке и водородом N-H на нижнем витке. Такая водородная связь образуется ровно через каждые 4 аминокислотных остатка, а каждый полный виток спирали составляет всего 3,6 аминокислотных остатка. Этот регулярный узор придает альфа-спирали очень определенные характеристики в отношении толщины катушки и длины каждого полного витка вдоль оси спирали.
Структурная целостность альфа-спирали частично зависит от правильной стерической конфигурации. Аминокислоты, R-группы которых слишком велики (триптофан, тирозин) или слишком малы (глицин), дестабилизируют альфа-спирали. Пролин также дестабилизирует альфа-спирали из-за своей неправильной геометрии; его R-группа соединяется обратно с азотом амидной группы, что вызывает стерические затруднения. Кроме того, отсутствие водорода в азоте пролина не позволяет ему участвовать в образовании водородных связей.
Еще одним фактором, влияющим на стабильность альфа-спирали, является общий дипольный момент всей спирали, обусловленный отдельными диполями групп C = O, участвующих в водородных связях. Стабильные альфа-спирали обычно заканчиваются заряженной аминокислотой, чтобы нейтрализовать дипольный момент.
Стабильные альфа-спирали обычно заканчиваются заряженной аминокислотой, чтобы нейтрализовать дипольный момент.
α-спираль
- 3,6 аминокислот на оборот
- 0,54 нм на оборот
- боковые цепи указаны
- Н-связи параллельно оси
- Н-связи n-4
- дипольный момент (отрицательный при C конец)
- no pro, less gly, ser
- ограниченные аналогичные заряды боковой цепи
α-спирали обладают дипольным моментом; некоторые боковые цепи являются предпочтительными
β
-ГЛИРОВАННЫЕ ЛИСТЫ
Эта структура возникает, когда два (или более, например.грамм. psi-loop) сегменты полипептидной цепи перекрывают друг друга и образуют ряд водородных связей друг с другом. Это может происходить в параллельном расположении:
Или в антипараллельном расположении:
Параллельное и антипараллельное расположение является прямым следствием направленности полипептидной цепи. В антипараллельной компоновке конец С-конца одного сегмента находится на той же стороне, что и конец N-конца другого сегмента. При параллельном расположении конец С-конца и конец N-конца находятся на одних и тех же сторонах для обоих сегментов.«Складка» возникает из-за чередования плоскостей пептидных связей между аминокислотами; выровненные амино- и карбонильная группы каждого противоположного сегмента меняют свою ориентацию от обращенных друг к другу на противоположные направления.
Параллельное расположение менее стабильно, поскольку геометрия отдельных молекул аминокислоты заставляет водородные связи располагаться под углом, делая их длиннее и, следовательно, слабее. Напротив, в антипараллельном расположении водородные связи выровнены прямо напротив друг друга, что делает связи более прочными и стабильными.
Обычно антипараллельный бета-складчатый лист образуется, когда полипептидная цепь резко меняет направление. Это может происходить в присутствии двух последовательных остатков пролина, которые создают угловой изгиб в полипептидной цепи и загибают ее обратно на себя. Это не обязательно для отдаленных сегментов полипептидной цепи для образования бета-складчатых листов, но для проксимальных сегментов это определенное требование. На коротких расстояниях два сегмента бета-складчатого листа разделены 4 + 2n аминокислотными остатками, причем 4 — это минимальное количество остатков.
Обратный поворот (ß-изгиб):
- R2 (сторона C = O) часто G, A
- R3 (сторона NH) часто D
- Proline часто R2 или R3
α
— ПЛИССИРОВАННЫЕ ЛИСТЫ
Альфа-гофрированные листы имеют структуру, аналогичную структуре бета-гофрированных листов. Эта структура энергетически менее выгодна, чем бета-складчатый лист, и довольно необычна для белков. Альфа-гофрированный лист характеризуется выравниванием карбонильных и аминогрупп; все карбонильные группы ориентированы в одном направлении, тогда как все группы N-H ориентированы в противоположном направлении.Поляризация амино- и карбонильных групп приводит к суммарному дипольному моменту на альфа-складчатом листе. Карбонильная сторона приобретает чистый отрицательный заряд, а амино-сторона приобретает чистый положительный заряд.
Третичная структура
http://commons.wikimedia.org/wiki/Fi…_Structure.png
Третичная структура включает связи между боковыми цепями и между ними:
• Водород (-OH… O -)
• Ионный (обычно отталкивание: -Ch3-Nh5 + ::::::: + h5N-Ch3-)
• Ван-дер-Вааль (притяжение на короткие расстояния)
• Дисульфид (ковалентный: -Ch3-SS- Ch3-)
• Гидрофобные
Третичные связи влияют на положение вторичных структур.
А положение вторичной структуры в белке будет влиять на типы боковых цепей (третичная структура).
α-Спираль на поверхности белка будет иметь гидрофильные боковые цепи с одной стороны оси спирали и гидрофобные боковые цепи с другой. Α-Спираль внутри белка будет иметь в основном гидрофобные боковые цепи. Α-Спираль, подверженная воздействию раствора со всех сторон (необычно), будет иметь гидрофильные боковые цепи со всех сторон от оси спирали (в основном).
Четвертичная структура
Четвертичная структура включает отдельные полипептиды, удерживаемые вместе слабыми связями в различной симметрии
Симметрии:
Гомомультимер :: гетеромультимер
гомомультимер: белок с несколькими полипептидными цепями, содержащий два или более идентичных компонента
: белок с множеством полипептидных цепей, содержащий два или более различных компонента
Изологичный :: гетерологичный
Закрытый :: открытый
Сворачивание белка снижает свободную энергию (ΔG) системы.
Сворачивание белка включает как белок, так и растворитель.
ΔG = GF- GU
= ΔH — TΔS
=
+ ΔH (белок)
+ ΔH (растворитель)
— TΔS (белок)
— TΔS (растворитель)
ΔG для сворачивание
невелико (от -20 до -60 кДж / моль) и в основном из-за гидрофобных взаимодействий
Почему так мало?
Изменения формы являются важной частью функции и контроля белка. Например: изменение формы позволяет ДНК-метилтрансферазе выбирать полуметилированный me CG / GC для биметилирования до me CG / Gme C
Белки: определение, состав, структура, примеры
Термин «протеин » происходит от греческого слова « proteios », что означает первичный или выдающийся, и впервые был предложен Йенсом Якобом Берцелиусом, одним из отцов современной химии, своему коллеге Герарду Йоханнесу. Малдер, изучавший химический состав альбуминов в 1839 году.Фактически, Берцелиус полагал, основываясь на формуле, данной Малдером альбумину, C 40 H 62 O 12 N 10 , неправильной формуле, что белки могут быть наиболее важными биологическими веществами.
Несмотря на ошибку Малдера, Берцелиус обладал «пророческой интуицией».
Они представляют собой класс молекул, присутствующих во всех живых организмах и во всех отделах клетки; в клетках животных они могут составлять более 50% их сухой массы.
Белки животных, растений, бактерий и вирусов представляют собой линейные полимеры, состоящие из субъединиц, называемых аминокислотами .Идентифицировано около 20 аминокислот, присутствующих почти исключительно в L-форме и связанных ковалентной связью, называемой пептидной связью, которая является жесткой и плоской. Аминокислотная последовательность, кодируемая конкретным геном, называется полипептидной цепью или белком. Каждая аминокислота повторяется более или менее большое количество раз.
Иногда D-аминокислоты обнаруживаются в некоторых бактериальных белках.
Белки имеют очень разные структуры, даже в одном и том же типе клеток, где мы можем найти сотни разных типов, которые выполняют разные функции.
Следует отметить, что пептидная связь очень устойчива при физиологическом pH: при отсутствии внешних вмешательств ее срок службы составляет около 1100 лет.
СОДЕРЖАНИЕ
Структура белков
Белки — это самые универсальные молекулы, присутствующие в живых организмах, где они выполняют функции, необходимые для жизни. Большое разнообразие функций, которые способны выполнять, проистекает из возможности сворачивания полипептидной цепи в конкретные трехмерные структуры , которые обеспечивают способность связывать различные молекулы и выполнять различные функции.
При описании того, как полипептидные цепи складываются в свои трехмерные структуры, полезно различать разные уровни организации, которые будут проанализированы ниже.
Примечание: в глобулярных белках присутствуют структуры, следующие за вторичной.
Первичная структура белка
Бычий инсулин был первым белком, первичная структура которого была определена благодаря работе Фредерика Сэнгера в 1953 году.
Первичная структура — это аминокислотная последовательность белков , их самый низкий уровень организации и, как было сказано ранее, он уникален и генетически детерминирован.
Он может состоять из 40-4000 аминокислотных остатков и определяет трехмерную структуру самого белка, которая, в свою очередь, определяет его функцию.
Полипептидная цепь имеет полярность, потому что ее два конца различны: один имеет свободную аминогруппу и называется NH 2 -конец или амино-конец, другой — свободная карбоксильная группа и называется COOH-концом или карбоксильным концом. . Два конца полипептидной цепи также известны как N-конец и C-конец, чтобы отличать их от карбоксильных и аминогрупп, присутствующих в цепи.По соглашению, N-концевой конец принимается за начало аминокислотной цепи и всегда помещается слева.
Первичная структура интересна еще и тем, что, сравнивая структуру одного и того же белка у разных видов, мы можем идентифицировать вариации, которым подвергся соответствующий ген, что является индикатором дивергенции видов в ходе эволюции.
Термины дипептид, трипептид, олигопептид и полипептид используются для обозначения цепей разной длины, соответственно, состоящих из 2, 3, менее 50 и более 50 аминокислот.
Вторичная структура белка
Открытие вторичной структуры белков связано с работой Линуса Полинга и Роберта Кори в 1951 году, которые предложили две структуры, названные α-спиралью и β-пластинчатой структурой или β-складчатым листом.
Вторичная структура возникает в результате образования водородных связей между смежными частями полипептидной цепи с определенными аминокислотными последовательностями. Следовательно, он описывает расположение в пространстве аминокислот не очень далеко друг от друга вдоль первичной структуры .
В дополнение к вышеупомянутым структурам, другие были идентифицированы как β-витки (β-витки), γ-витки (гамма-витки) и Ω-петли (петли омега), все они принадлежат к группе, называемой обратными витками. Эти структуры часто встречаются там, где полипептидная цепь меняет направление, и обычно расположены на поверхности молекулы.
Примечание: около 32–38% аминокислот в глобулярных белках находятся в структурах α-спирали.
Сверхвторичные структуры или мотивы
Они представляют собой комбинацию вторичных структур для формирования области молекулы с определенной трехмерной структурой и топологией.Супервторичные структуры связаны друг с другом петлевыми областями с неопределенной структурой.
Распространенные мотивы:
- «цинковый палец» (β-α-β), который часто встречается в белках, связывающих РНК или ДНК;
- греческий ключ, β-меандр и β-ствол.
Домены
Домены — это следующий уровень организации. Они представляют собой глобулярные области , которые являются результатом комбинации мотивов , которые сворачиваются независимо от остальной части полипептидной цепи, давая стабильную структуру.
Они состоят из 40-400 аминокислот, за исключением моторных и киназных доменов, которые образованы гораздо большим количеством аминокислот.
Домены были разделены на три основные группы на основе присутствующих вторичных структур и мотивов:
- α-домены;
- β-доменов;
- α / β-доменов.
Было обнаружено более 1000 доменных семейств (члены каждого семейства называются «гомологами»), и, похоже, они произошли от общего предка.
Очень часто каждый домен выполняет определенную функцию, то есть является функциональной единицей белка, в котором он содержится.
Белки могут состоять из одного домена, более мелких или из нескольких доменов. Например, химотрипсин состоит из одного домена, а папаин — из двух доменов.
Третичная структура белка
Третичная структура, также называемая «нативной структурой», представляет собой трехмерную структуру белков . Первый белок, третичная структура которого была определена, был миоглобин в 1958 году благодаря работе Джона Кендрю.
В этом типе структуры сворачивание белковой цепи отвечает за размещение аминокислотных остатков в тесном контакте далеко друг от друга вдоль цепи, то есть это относится к трехмерному расположению аминокислот вдали друг от друга вдоль первичная структура.
Рис. 3 — Окси-миоглобин
Третичная структура белков, в частности белков, состоящих из более чем 200 аминокислотных остатков, образована различными доменами, связанными короткими полипептидными сегментами. Он часто стабилизируется дисульфидными мостиками между остатками цистеина, мостиками, которые образуются после того, как молекула достигла своей нативной конформации.
Следует отметить, что не все глобулярные белки имеют третичную структуру.
Примером являются казеины молока, полипептидная цепь которых принимает неупорядоченную трехмерную конформацию, также известную как случайная спиральная структура .Неупорядоченная структура делает их очень восприимчивыми к действию кишечных протеаз и, следовательно, к высвобождению составляющих аминокислот. Это делает их очень подходящими для выполнения своей пищевой роли.
Другой пример белка со случайной спиралью — эластин.
Четвертичная структура белка
Этот дополнительный уровень структурной организации описывает, как более одной полипептидной цепи связывают с образованием единой белковой структуры. Следовательно, это относится к пространственному расположению отдельных цепей и природе сил, связывающих их вместе, например:
- гидрофобный эффект, который является основной движущей силой сворачивания белка;
- водородная связь;
- ван-дер-Ваальсовых взаимодействий;
- ионных взаимодействий;
- ковалентные сшивки.
Полученная структура называется олигомером (олигомерным белком) и составляющими полипептидами, которые могут быть одинаковыми или разными, мономерами или просто субъединицами.
В целом, большинство внутриклеточных белков являются олигомерами, в отличие от большинства внеклеточных. Классическим примером белка с четвертичной структурой является гемоглобин .
Этот уровень структуры явно отсутствует у глобулярных белков, состоящих из одной полипептидной цепи, то есть у мономерных белков.
Белки также способны взаимодействовать между собой, образуя структуры, в которых, действуя синергетическим образом, они выполняют функции, которые они не смогли бы выполнить в одиночку.
Примерами являются « макромолекулярные машины », участвующие в синтезе ДНК, РНК и самих белков, в сокращении мышц или в передаче сигналов между соседними клетками.
Список литературы
Лодиш Х., Берк А., Зипурский С.Л. и др. Молекулярная клеточная биология.4-е издание. Нью-Йорк: У. Х. Фриман; 2000. Раздел 3.1, Иерархическая структура белков. Доступно по ссылке: https://www.ncbi.nlm.nih.gov/books/NBK21475/
Кессель А., Бен-Тал Н. Введение в белки: структура, функции и движение. CRC Press, 2011 doi: 10.1002 / cbic.201100254
Нельсон Д.Л., Кокс М.М. Ленингер. Основы биохимии. 6-е издание. W.H. Фриман и компания, 2012
Стипанук М.Х., Кодилл М.А. Биохимические, физиологические и молекулярные аспекты питания человека.3-е издание. Elsevier Health Sciences, 2012 г.
Белковый состав определяет предпочтительное потребление аминокислот во время анаэробной ферментации смешанной культуры.
Основные моменты
- •
-
Белковая ферментация обычно приводит к неполному подкислению.
- •
-
Сбалансированные окислительно-восстановительные роли аминокислот не гарантируют более высоких степеней подкисления.
- •
-
Показано преимущественное потребление аминокислот.
- •
-
Белковый состав влияет на подкисление, селективность и предпочтительное потребление.
- •
-
Принятая стехиометрия недостаточна для описания превращения белка в ЛЖК.
Abstract
Валоризация богатых белком остаточных потоков с помощью анаэробной ферментации смешанных культур (MCF) практически не изучена в отличие от отходов, богатых углеводами. Таким образом, целью данной работы было изучить, как состав белков, т.е.е. профиль аминокислот (АК) влияет на индивидуальное потребление аминокислот и, следовательно, на результат процесса. Ферментации смешанных культур выполняли с двумя модельными белками (казеином и желатином) с использованием реакторов непрерывного и периодического действия при нейтральных значениях pH и 25 ° C. Подкисление обоих белков было неполным, казеин достиг более высокой ценности, чем желатин. Несмотря на то, что преобладала уксусная кислота, спектры продуктов также были разными: н-масляная кислота была вторым основным продуктом для казеина и пропионовой кислотой для желатина.Было продемонстрировано преимущественное потребление аминокислот, которое интересно зависит от белкового состава. Ранее принятая стехиометрия точно описывает образование изо- и н-масляной кислоты, но не дает образования пропионовой, изо и н-валериановой кислоты. В целом, это исследование предлагает лучшее понимание механизмов ферментации белка, что поможет улучшить модели деградации и разработать процессы ферментации на основе оптимального выбора субстрата.
Ключевые слова
Подкисление
Биоперерабатывающий завод
Производительность
Белковый состав
Селективность
Производство летучих жирных кислот
Рекомендуемые статьи Цитирующие статьи (0)
Полный текст
© 2020 Elsevier Ltd.Все права защищены.
Рекомендуемые статьи
Цитирующие статьи
Отношения на уровне протеома между локальным аминокислотным составом и судьбами и функциями белков
Цитирование: Cascarina SM, Ross ED (2018) Отношения на уровне протеома между локальным аминокислотным составом и судьбами белков и функции. PLoS Comput Biol 14 (9):
e1006256.
https://doi.org/10.1371/journal.pcbi.1006256
Редактор: Авнер Шлессингер, Медицинская школа Икана на горе Синай, США
Поступила: 30 мая 2018 г .; Принята к печати: 16 августа 2018 г .; Опубликовано: 24 сентября 2018 г.
Авторские права: © 2018 Cascarina, Ross.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все соответствующие данные находятся в документе и его файлах с вспомогательной информацией.
Финансирование: Эта работа была поддержана Национальным научным фондом (MCB-1517231; https://www.nsf.gov/div/index.jsp?div=MCB) EDR и Национальным институтом общей медицины. Наук (GM105991; https: // www.nigms.nih.gov/) в EDR. Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Домены низкой сложности (LCD) в белках — это области, обогащенные только подмножеством возможных аминокислот. ЖКД могут состоять из гомополимерных повторов одной аминокислоты, коротких тандемных повторов, состоящих только из нескольких различных аминокислот, или апериодических участков с небольшим аминокислотным разнообразием [1].Белки, содержащие ЖКД, относительно распространены среди организмов из всех сфер жизни и особенно распространены среди эукариот [2–4]. Например, примерно 70% генов в геноме Saccharomyces cerevisiae обладают по крайней мере одним классически определенным ЖКД [3]. Кроме того, общее количество ЖК-дисплеев намного превышает общее количество дрожжевых генов (примерно в 2 раза больше ЖК-дисплеев, чем генов), что указывает на то, что многие гены содержат несколько различных ЖК-дисплеев.
Разработаны различные методы оценки сложности биополимерной последовательности [1,5–9].Одним из наиболее часто используемых методов определения ЖК-дисплеев является алгоритм SEG [1], который сканирует последовательности белков (или нуклеиновых кислот) с использованием короткого скользящего окна и вычисляет локальную энтропию Шеннона для каждого окна (см. [10] для получения подробной информации. описание). Подпоследовательности со значением энтропии Шеннона ниже заранее определенного «триггерного» порога классифицируются как ЖК-дисплеи. Границы ЖК-дисплея позже расширяются и уточняются путем объединения перекрывающихся ЖК-дисплеев и вычисления вероятностей комбинаторной последовательности. Другой показатель, обычно используемый для оценки относительной сложности последовательностей, — это систематическая ошибка, которая включает определение статистической вероятности последовательности с учетом частот отдельных аминокислот для всего протеома [11,12].Эти подходы (или близкие к ним подходы) широко используются для изучения LCDs в масштабе протеома [1,3,12-17].
LCD-содержащих белков вовлечены во множество нормальных и патологических клеточных процессов. Например, дрожжевые белки, богатые Q / N, часто играют роль в регуляции транскрипции, эндоцитоза и регуляции клеточного цикла, среди других функций [11,18]. Многие белки, содержащие Q / N-богатые ЖК-дисплеи или ЖК-дисплеи родственных типов (Q / N / G / S / Y-богатые ЖК-дисплеи), были связаны с прионами или связанными с прионами процессами [11,18–21].Кроме того, многие прионоподобные LCDs, которые часто состоят из коротких тандемных повторов низкой сложности [22], были связаны со стрессовыми гранулами и процессинговыми телами (P-тельцами) у эукариот (см. [23] для недавнего обзора). Аминокислотный состав этих ЖК-дисплеев придает этим доменам необычные биофизические свойства [24], что, вероятно, связано с их уникальным поведением in vitro и in vivo [25–30]. Однако эти необычные характеристики оказываются неразрывно связаны и с патологическими процессами.Например, генетическая экспансия областей, кодирующих гомополимерные глутаминовые повторы (простейший тип ЖКД) в различных белках, может привести к множеству нейродегенеративных расстройств, включая болезнь Хантингтона и спиноцеребеллярную атаксию (см. Обзор [31]). Более того, мутации ЖКД белков стрессовых гранул могут изменять динамику стрессовых гранул и приводить к дегенеративным заболеваниям [26,28,30,32,33]. Важность ЖК-дисплеев выходит далеко за пределы ЖК-дисплеев с высоким соотношением Q / N, поскольку ЖК-дисплеи других составов также связаны с нормальными и патологическими клеточными процессами [12,14,17,34,35].
Хотя ЖК-дисплеи могут явно влиять на регуляцию и функцию белков, ряд проблем до сих пор ограничивают понимание этих взаимосвязей на уровне протеома. Одна из основных проблем заключается в определении ЖК-дисплеев. Современные подходы используют статистически определенные пороги сложности последовательности или систематической ошибки [1,11], или произвольно выбранные длины повторов для белков с гомополимерными повторами [34–41]. Хотя эти определения ЖКД, композиционно смещенных последовательностей (здесь называемых «статистически смещенные домены», чтобы избежать путаницы в дальнейшем) или гомополимерных повторов способствовали важным открытиям, биологическая значимость этих пороговых значений не была тщательно изучена.Более того, эти белки часто группируются в один класс, хотя их состав и, следовательно, физические свойства могут сильно различаться (ограничение, которое было оценено в недавнем обзоре [42]).
Чтобы устранить эти ограничения, мы разработали альтернативный подход к выводу о взаимосвязи между аминокислотным составом и метаболизмом и функцией белка. Сосредоточившись на аминокислотном составе, который является фундаментальной особенностью, лежащей в основе как сложности последовательности, так и статистической ошибки аминокислот, мы исследовали связи между локальным композиционным обогащением и различными аспектами регуляции и функции белка, не обращаясь к заранее определенной сложности последовательности или пороговым значениям статистической погрешности.Мы обнаружили, что локальное композиционное обогащение коррелирует с различиями почти во всех основных аспектах пребывания белка в клетке, включая эффективность трансляции, численность, период полужизни, беспорядочные связи белок-белок, субклеточную локализацию и функцию. Однако обогащение различными аминокислотами связано с разными эффектами, даже для остатков, часто группируемых на основе физико-химического сходства, что подчеркивает важность различения ЖКД разных типов. Эти взаимосвязи заметны при составах ниже тех, которые требуются для классификации как низко-сложные или статистически предвзятые, что позволяет предположить, что пороговые значения в традиционных методах не могут быть оптимизированы биологически.Наконец, анализ экспериментально определенных белковых компонентов стрессовых гранул и Р-тел выявляет как общие, так и отдельные особенности состава, связанные с этими органеллами.
Результаты
Систематическое исследование местного аминокислотного состава
По сути, сложность последовательности и статистическая ошибка аминокислот являются косвенными показателями локального аминокислотного состава. Поскольку состав является более прямым показателем общих свойств белкового домена, мы стремились изучить, можно ли использовать только состав для вывода специфичных для остатков взаимоотношений между локальным аминокислотным составом и регуляцией и функцией белка.Сначала мы разработали алгоритм разделения протеома дрожжей на основе максимального локального состава для каждой аминокислоты с использованием серии размеров окна сканирования (рис. 1; см. Методы). Для всех аминокислот большинство белков распределяется по составным ячейкам ≤ 25% (рис. 2 и таблица S1). Однако количество белков, достигающих более высокого локального состава, на что указывает правое плечо или хвост в распределении, сильно зависело от остатков. Например, белки, содержащие локальное обогащение высокогидрофобных остатков (I, L, M и V), ароматических остатков (F, W и Y) или цистеина, почти исключительно ограничены ячейками состава ≤ 45% для наименьшего окна. размер, тогда как распределение аланина и пролина простирается до несколько более высоких диапазонов состава (до 60–65%).Белки, содержащие локальное обогащение полярных (G, N, Q, S и T) или заряженных (D, E и K) остатков в ячейках композиции ≥ 40%, относительно распространены даже среди окон больших размеров (хотя и в разной степени) , тогда как регионы, богатые гистидином и аргинином, относительно редки. Эти данные показывают, что относительно высокое локальное обогащение допустимо для некоторых аминокислот, в то время как композиционное обогащение для других аминокислот, по-видимому, ограничено у дрожжей.
Рис. 1. Изображение сортировки протеома на основе максимальной локальной композиции.
(A) Для каждой комбинации аминокислоты и размера окна каждый дрожжевой белок сортируется в интервалы процентного состава на основе максимального локального состава аминокислоты в пределах заданного размера скользящего окна. Это эффективно сортирует протеом дрожжей 200 различными способами (20 аминокислот x 10 различных размеров скользящего окна). (B) Визуальное представление белков, отсортированных на основе максимального локального состава аспарагиновой кислоты с скользящим окном из 20 аминокислот.
https: // doi.org / 10.1371 / journal.pcbi.1006256.g001
Рис. 2. Распределение протеома дрожжей на основе максимального локального аминокислотного состава.
Количество белков, разделенных на каждый размер окна / процентный интервал состава для каждой из 20 канонических аминокислот, наносится на график как функция максимального локального состава для каждого размера окна. Точки разброса соединены отрезками линии только для наглядности.
https://doi.org/10.1371/journal.pcbi.1006256.g002
Зависимость от остатков между локальным обогащением состава и метаболизмом белка
Хотя происхождение и эволюция ЖКД широко изучены [3,4,14,38,43,44], регуляция и метаболизм ЖК-содержащих белков остаются малоизученными.Белки с внутренне неупорядоченными сегментами, которые часто квалифицируются как LCD [45,46], связаны с более низкими периодами полужизни белка [47]. Однако не все внутренне неупорядоченные области приводят к короткому периоду полураспада белков, и не все ЖК-дисплеи являются внутренне неупорядоченными [15]. Кроме того, белки с гомополимерными повторами, если их рассматривать как один класс, связаны с более низкой эффективностью трансляции, более низким содержанием белка и более низким периодом полужизни белка по сравнению с белками, лишенными гомополимерных повторов [37].Однако регуляция и структурные свойства белков с ЖК или гомополимерными повторами, вероятно, сильно зависят от преобладающих аминокислот в интересующем домене [42].
Чтобы изучить взаимосвязь между локальным обогащением состава и метаболизмом белка, мы сначала исследовали возможные связи между локальным обогащением состава и изобилием белка. Недавние успехи в протеомных методах способствовали значительному покрытию протеомом как для измерения содержания белка [48], так и для измерения периода полужизни белка [49] в дрожжах.В каждой ячейке состава размер окна / процентное соотношение распределение значений содержания белка для всех белков, распределенных в эту ячейку, сравнивали с распределением содержания белка для всех других дрожжевых белков (тест Mann-Whitney U ). Переходы от значительно более низкой медианной численности к значительно более высокой медианной численности или наоборот наблюдаются при обогащении по многим аминокислотам по отдельности (рис. 3). Однако направление тенденций прогрессивного обогащения состава зависит от типа аминокислоты.Для большинства аминокислот (C, D, F, H, I, L, M, N, P, Q, R, S, T, W или Y) композиционное обогащение связано с более низким медианным содержанием белка. Однако композиционное обогащение A, G или V связано с более высоким медианным содержанием белка. Два очень похожих перехода наблюдаются как для E-богатых, так и для K-богатых последовательностей: по мере увеличения композиционного обогащения относительное медианное содержание белка переходит от высокого к низкому, а затем обратно к высокому. В совокупности эти тенденции согласуются с ранее наблюдаемыми корреляциями между обилием белка и составом цельного белка, хотя и намного сильнее [50,51].Это говорит о том, что наблюдаемые ранее тенденции могут фактически отражать эффекты локального обогащения состава, которые увеличивают видимый состав цельного белка для обогащенной аминокислоты, но при этом ослабляются смешивающими эффектами остальной части белковой последовательности.
Рис. 3. Максимальный местный аминокислотный состав связан с остаточно-специфическими различиями в содержании белка.
Для каждой аминокислоты значения обилия белков, соответствующие белкам, разделенным на заданный размер окна и процентный интервал состава, сравнивали со значениями для всех белков длиной ≥ соответствующего размера окна, которые были исключены из интервала.Для этого рисунка и соответствующих последующих рисунков красные и синие точки обозначают ячейки, для которых распределение значений содержания белка значительно отличается (с поправкой Бонферрони p ≤ 0,05) от таковых для исключенных белков: красные точки указывают ячейки с более низким средним значением относительного для исключенных белков, тогда как синие точки указывают на ячейки с более высоким относительным медианным значением. Серые точки указывают на отсутствие статистической значимости сравнений. Индивидуальные точки масштабируются в пределах каждого участка, чтобы отразить размеры образцов белков, содержащихся в каждом бункере.
https://doi.org/10.1371/journal.pcbi.1006256.g003
Аналогичные тенденции наблюдаются при сравнении обогащения состава с периодом полураспада белка (рис. 4). Обогащение композиции по большинству аминокислот (C, H, K, M, N, P, S или T) связано с более низким периодом полужизни белка, тогда как обогащение по A, G, I или V связано с более высоким период полураспада белка. Обогащение F приводит к начальному переходу от более низкого периода полураспада к более высокому, в то время как дальнейшее обогащение приводит к переходу обратно к более низкому периоду полураспада.Стоит отметить, что аналогичные тенденции наблюдались в независимом наборе данных о периоде полужизни белка, когда белки анализировались на основе аминокислотного состава цельного белка [52], предполагая, что максимального локального состава достаточно для выявления ассоциаций между аминокислотным составом и половиной. -жизнь. Хотя для многих аминокислот тенденции очевидны, сила связи между композиционным обогащением и периодом полужизни белка, по-видимому, немного слабее, чем связь между композиционным обогащением и изобилием белка.Вероятно, это связано, по крайней мере частично, с ограниченным покрытием протеомом (по сравнению с набором данных о содержании белка). Однако недавнее исследование также показало, что период полураспада белка сильно зависит от факторов, отличных от характеристик последовательности [53], что, вероятно, еще больше ослабит взаимосвязь между обогащением состава и периодом полураспада белка. Наконец, период полужизни белка, как правило, менее консервативен, чем его изобилие [54], что, возможно, предполагает, что конкретные отношения между консервативными характеристиками последовательности и периодом полужизни белка могут быть не особенно сильными.Поэтому довольно удивительно, что мы наблюдаем указанные тенденции, несмотря на эти ограничения, и можно предположить, что период полураспада в большей степени зависит от местного состава, чем от конкретных мотивов первичной последовательности.
Рис. 4. Максимальный местный аминокислотный состав связан с остаточно-специфическими различиями в периоде полужизни белка.
Для каждой аминокислоты значения периода полужизни белка, соответствующие белкам, разделенным на заданный размер окна и процентный интервал состава, сравнивали со значениями периода полужизни для всех белков длиной ≥ соответствующего размера окна, которые были исключены из интервала.
https://doi.org/10.1371/journal.pcbi.1006256.g004
Прямое измерение скорости синтеза белка является более сложной экспериментальной задачей. Следовательно, охват протеома экспериментально полученной эффективностью трансляции остается существенно ниже, чем охват белком изобилия и периода полужизни. Нормализованная эффективность трансляции (nTE), показатель эффективности элонгации трансляции [55], основан на частотах использования кодонов и количестве копий гена тРНК, что позволяет рассчитать эффективность трансляции для всего протеома.Таким образом, мы сначала исследовали взаимосвязь между локальным композиционным обогащением и рассчитанной эффективностью трансляционного удлинения. nTE были рассчитаны для последовательностей цельного белка с использованием соответствующей кодирующей области на транскриптах мРНК (см. Методы). Эффективность трансляции сильно зависит от локально обогащенной аминокислоты (рис. 5). Для большинства аминокислот (C, D, E, F, H, I, K, L, M, N, P, Q, R или Y) локальное обогащение связано со значительно более низкими средними значениями nTE, что позволяет предположить, что, поскольку Один класс, белки с локальным обогащением состава, как правило, переводятся относительно неэффективно.Белки с доменами, обогащенными S, T или W, обычно связаны со значительно более низкими медианными nTE, хотя белки с очень высоким обогащением S, T или W связаны со значительно более высокими медианными nTE. Однако белки с доменами, обогащенными остатками A, G или V, последовательно связаны со значительно более высокими медианными nTE, что позволяет предположить, что эти белки могут транслироваться относительно эффективно. Примечательно, что наблюдаются почти идентичные тенденции между локальным композиционным обогащением и экспериментально полученными скоростями синтеза белка, о которых сообщалось для ограниченного протеома (S1 Рис.), Несмотря на значительное сокращение размера выборки ( n = 1115; [56]), что позволяет предположить, что nTE может служить хорошим суррогатом общей эффективности синтеза белка.В совокупности эти результаты показывают, что локальное обогащение аминокислот связано с различиями в скорости продукции белка в зависимости от состава.
Рис. 5. Максимальный местный аминокислотный состав связан с остаточно-специфическими различиями в nTE.
Для каждой аминокислоты значения nTE, соответствующие белкам, разделенным на заданный размер окна и процентный интервал состава, сравнивали со значениями nTE для всех белков длиной ≥ соответствующего размера окна, которые были исключены из интервала.
https://doi.org/10.1371/journal.pcbi.1006256.g005
Для большинства аминокислот мы заметили замечательное соответствие в тенденциях эффективности трансляции, изобилия белка и периода полужизни белка, несмотря на то, что эти значения получены на основе совершенно разных методов и экспериментов. Например, локальное обогащение многих типов аминокислот связано с низкими значениями nTE, низким содержанием белка и низким периодом полужизни белка (Таблица 1). В то время как эффективность трансляции и скорость деградации белка в значительной степени функционально независимы в клетках, обилие белка зависит, по крайней мере частично, как от эффективности трансляции, так и от периода полужизни белка [49].Это может указывать на то, что изобилие этих белков в клетках ограничено сочетанием низкой эффективности трансляции и быстрой скорости деградации. Напротив, локальное обогащение некоторыми аминокислотами связано с высоким содержанием белка, а также имеет тенденцию иметь более высокие значения nTE и более высокие периоды полураспада, что, возможно, предполагает, что высокое содержание белка для этих белков достигается за счет комбинации эффективной трансляции и плохой деградации.
Таблица 1. Жизненный цикл белков с высоким локальным составом отдельных аминокислот включает скоординированное регулирование эффективности трансляции, изобилия белка и периода полужизни белка.
Для каждой аминокислоты указаны тенденции средних значений nTE, содержания белка и периода полужизни при обогащении (т.е. приближении к более высокому процентному составу) данной аминокислоты. «Более высокий» указывает на то, что белки в ячейках с более высоким процентным содержанием имеют тенденцию иметь большее среднее значение по сравнению со всеми другими белками, в то время как «Ниже» указывает, что белки в ячейках с более высоким процентным содержанием, как правило, имеют большее среднее значение по сравнению со всеми другими белками. «Смешанные» обозначают аминокислоты, для которых наблюдаются множественные переходы при постепенном обогащении состава.Также указаны наборы данных без четких статистически значимых пороговых значений перехода («н.у.»). Цвета соответствуют цветам, используемым на рис. 3–5.
https://doi.org/10.1371/journal.pcbi.1006256.t001
Почти идентичные, специфичные для остатков отношения между локальным обогащением состава и обилием белка наблюдаются в
C . elegans
В качестве модели эукариотического организма S . cerevisiae обеспечивает ряд важных преимуществ в исследованиях протеомного масштаба, связывающих последовательность белка с метаболизмом и функцией белка.В дополнение к непревзойденному охвату протеомами в наборах данных об изобилии и периоде полужизни белка и доступности инструментов, специфичных для дрожжей, таких как nTE, анализ функции последовательности в дрожжах дополнительно упрощается за счет отсутствия тканеспецифических эффектов и ограниченного альтернативного сплайсинга (только ~ 4% дрожжевых генов содержат интроны, и лишь небольшая часть этих генов способна продуцировать альтернативные белковые продукты [57,58]).
Имея в виду эти предостережения, мы стремились изучить, могут ли подобные отношения между локальным аминокислотным составом и обилием белка быть обнаружены в модельном многоклеточном эукариотическом организме.Мы решили сосредоточиться на измерениях содержания белка в организме в C . elegans [59] по четырем основным причинам: 1) из-за технических экспериментальных проблем, измерения содержания белка в C . elegans существенно более надежны, чем измерения периода полужизни белка; 2) в масштабе протеома изобилие белка более консервативно у разных видов дрожжей, чем период полужизни белка [49], что позволяет предположить, что конечные уровни белка имеют тенденцию ограничиваться у разных организмов, в то время как регуляция метаболических путей, которые вносят вклад в изобилие белка. может различаться; 3) изобилие белка является, по крайней мере частично, функцией эффективности трансляции и периода полужизни белка; и 4) параметры, лежащие в основе метода эффективности трансляции (а именно « s -вектор», или эффективность спаривания оснований колебания между изоакцепторами тРНК) были оптимизированы для дрожжей [60].Следовательно, метод nTE может быть неприменим для других организмов.
Чтобы изучить взаимосвязь между максимальным локальным составом и обилием белка, мы сначала определили протеомное распределение C . elegans белков в зависимости от максимального локального состава каждой аминокислоты. Модель C . elegans -специфические распределения протеома (S2 фиг. И S2 таблица) в целом были очень похожи на распределения протеомов дрожжей (фиг. 2).Однако максимальный локальный состав для S и N, по-видимому, несколько более ограничен в C . elegans (обозначено сокращением плеча до более низких максимальных композиций), тогда как G, P и T достигают немного более высоких максимальных локальных композиций, что указывает на ослабление ограничений на локальное обогащение этих остатков. Эти результаты согласуются с предыдущими наблюдениями, в которых отмечены как общие, так и специфичные для организма сигнатуры гомополимерных повторов или объемные протеомные композиции в протеомах разных организмов [4,38,40,41,44–46,61].
Как наблюдали у дрожжей, прогрессивное обогащение состава приводит к переходу от более высокого к более низкому медианному содержанию большинства аминокислот с четкой тенденцией (C, F, I, M, N, P, S, W и Y; рис. 6). Кроме того, все три аминокислоты (A, G и V), которые демонстрируют переход от более низкого к более высокому среднему содержанию при прогрессивном обогащении дрожжей, демонстрируют ту же тенденцию в C . elegans тоже. Действительно, только одна аминокислота с четким переходом в содержании белка при локальном обогащении различается между C . elegans и S . cerevisiae : у дрожжей локальное обогащение калием связано со смешанными эффектами на изобилие белка (в зависимости от степени обогащения калием), тогда как у дрожжей C . elegans локальное обогащение калием слабо (но постоянно) связано с более высоким содержанием белка. Следовательно, между локальным обогащением аминокислот и обилием белка в более сложных эукариотах наблюдаются почти идентичные отношения, специфичные для остатков.
Рис 6.Связь между максимальным локальным составом и содержанием белка в C . elegans .
Для каждой аминокислоты значения обилия белков, соответствующие белкам, разделенным на заданный размер окна и процентный интервал состава, сравнивали со значениями для всех белков длиной ≥ соответствующего размера окна, которые были исключены из интервала. Тенденции в изобилии белка в зависимости от максимального локального состава для многих аминокислот очень похожи между дрожжами и C . elegans (сравните с рис. 3).
https://doi.org/10.1371/journal.pcbi.1006256.g006
Обогащение композиции связано с влиянием на метаболизм белка в отсутствие классических доменов низкой сложности, статистически предвзятых и гомополимерных доменов
Важным преимуществом подхода к ЖК-дисплеям с точки зрения композиции является возможность исследовать взаимосвязи между аминокислотным составом и исходами белка без обращения к заранее определенным пороговым значениям статистического смещения аминокислот [11] или сложности последовательности [1,10], которые могут не отражать биологически значимые пороговые значения.Действительно, наблюдаемые переходы в средней эффективности трансляции, содержании белка и периоде полураспада белка часто происходят при удивительно умеренных уровнях композиционного обогащения, что позволяет предположить, что эти тенденции могут наблюдаться даже в отсутствие классически определенных статистически предвзятых или низких значений. домены сложности.
Статистическая ошибка смещения аминокислот концептуально параллельна нашему исследованию обогащения состава и использовалась для исследования функций белков со статистически смещенными доменами [11,12].Чтобы проверить, может ли композиционное обогащение быть связано с биологически значимыми эффектами на метаболизм белков независимо от статистически смещенных доменов, был использован консервативный порог смещения для определения статистически смещенных доменов с использованием ранее разработанной методологии [12] (также см. Методы). Затем белки со статистически смещенными доменами отфильтровывали из протеома дрожжей ( n = 866 статистически смещенных белков для транслированного протеома дрожжей с последовательностями длиной ≥ 30 остатков).Однако, даже в отсутствие статистически смещенных доменов, обогащение состава привело к устойчивым тенденциям в эффективности трансляции, изобилии белка и полужизни белка, которые повторно капитулировали с первоначально наблюдаемыми (S3 – S5 Figs). Это говорит о том, что обогащение состава влияет на метаболизм белков на порогах, предшествующих пороговым значениям, которые требуются для классификации как статистически необъективные альтернативными методами.
Алгоритм SEG по умолчанию использует существенно более мягкие критерии при классификации белковых доменов как низко-сложные [1].Действительно, из 5 901 белка длиной ≥30 аминокислот в транслированном протеоме ORF 4 147 белков содержат по крайней мере один LCD, что согласуется с предыдущими оценками [3]. Тем не менее, несмотря на значительное уменьшение размера протеома, многие тенденции в метаболизме белков различимы, даже когда все белки с SEG-положительной последовательностью отфильтрованы из протеома (S6-S8 Figs). Это говорит о том, что обогащение состава оказывает биологически значимое воздействие даже среди белков, не содержащих ЖКД.
Белки, содержащие гомополимерные аминокислотные повторы (часто определяемые как пять или более идентичных аминокислот подряд), как недавно сообщалось, имеют более низкую эффективность трансляции, меньшее содержание белка и более низкий период полужизни белка по сравнению с белками без гомополимерных повторов [37] . Гомополимерные повторы фактически представляют собой короткие последовательности с максимально возможной плотностью одной аминокислоты. Следовательно, ожидается, что белки с гомополимерными повторами будут непропорционально распространены среди доменов, обогащенных составом, что повышает вероятность того, что тенденции, наблюдаемые в настоящем исследовании, были ошибочно приписаны только обогащению состава.Чтобы напрямую изучить эту возможность, взаимосвязь между обогащением состава и nTE, численностью и периодом полураспада была повторно оценена для отфильтрованного протеома, который исключает все белки, содержащие по крайней мере один гомополимерный повтор ( n = 755 белков исключены). В то время как исключение этих белков предпочтительно уменьшает размеры выборки при более высоких процентах обогащения по составу, отсутствие гомополимерных повторяющихся белков мало влияет на тенденции в nTE, численности и периоде полужизни в зависимости от обогащения состава (S9 – S11, рис.).Это не исключает окончательно возможность того, что гомополимерные повторы могут каким-то образом специфически влиять на эффективность трансляции, численность и период полураспада. Однако, поскольку гомополимерные повторы per se не являются абсолютно необходимыми, эффекты гомополимерных повторов вместо этого можно объяснить просто локальным композиционным обогащением.
В совокупности эти результаты предполагают, что обогащение состава влияет на эффективность трансляции, обилие белка и период полужизни белка на порогах, предшествующих пороговым значениям, которые требуются для классификации как низко-сложные или статистически смещенные традиционными методами.Стоит отметить, что в процессе удаления белков с классически определенными доменами низкой сложности, статистически смещенными или гомополимерными доменами белки с несколькими отдельными доменами, сильно обогащенными разными типами аминокислот, или с отдельными доменами, сильно обогащенными более чем одним аминокислоты, удаляются из протеома перед повторной оценкой. Следовательно, тенденции в метаболизме белков, наблюдаемые при обогащении данной аминокислотой, не связаны с смешивающими эффектами доменов, сильно обогащенных другими аминокислотами, встречающихся в тех же белковых последовательностях.
Локальное обогащение состава влияет на беспорядочные связи белок-белок специфическим для остатков образом
Локальное обогащение отдельной аминокислотой может сильно влиять на физико-химические свойства данного белкового домена [24]. В клеточном контексте эти физико-химические свойства, вероятно, влияют на взаимодействия между белками и окружающими молекулами, включая другие белки.
Чтобы проверить, влияет ли локальное композиционное обогащение на взаимодействия белок-белок, мы исследовали взаимосвязь между обогащением по каждой из аминокислот и беспорядочной половой связью белок-белок (определяемой как количество уникальных взаимодействующих партнеров на белок).Белки, обнаруженные в ряде бункеров с высоким процентным содержанием большинства аминокислот (A, D, E, G, K, N, P, Q, R и V), связаны со значительно большим количеством взаимодействующих партнеров по сравнению со всеми другими белками. (Рис. 7), предполагая, что эти домены относительно разнородны. Кроме того, белки с умеренным обогащением выбранных гидрофобных остатков (I, L и M) обычно связаны с большим количеством взаимодействующих партнеров, хотя меньшее количество сравнений достигает статистической значимости (синие или красные точки). Эти результаты согласуются с предыдущими сообщениями о том, что как один класс белки с ЖК-дисплеями или гомополимерными повторами имеют тенденцию иметь больше партнеров по взаимодействию белок-белок [16,37].Однако белки в диапазоне ячеек с высоким процентным содержанием для каждого из ароматических остатков (F, W и Y) связаны со значительно меньшим количеством взаимодействующих партнеров по сравнению с другими белками, что позволяет предположить, что ароматические остатки имеют тенденцию к отсутствию наблюдаемой беспорядочной связи. при более высоком процентном содержании других аминокислот. Кроме того, белки с локальным содержанием C от умеренного до высокого и белки с чрезвычайно высоким максимальным локальным содержанием S или T также связаны со значительно меньшим количеством взаимодействующих партнеров по сравнению с другими белками, что позволяет предположить, что эти домены также относительно не беспорядочные.Это особенно интересно, учитывая, что эти тенденции не наблюдались при обогащении другими полярными остатками. Опять же, это подчеркивает потенциальную ловушку объединения аминокислот со связанными физико-химическими свойствами в одну категорию. В совокупности эти результаты указывают на то, что беспорядочные связи белок-белок варьируются для белков с высоким обогащением состава в зависимости от остатка.
Рис. 7. Максимальный локальный аминокислотный состав соответствует беспорядочной связи белок-белок в зависимости от остатка.
Локальное обогащение отдельных аминокислот соответствует зависимым от состава изменениям числа уникальных партнеров белок-белкового взаимодействия.
https://doi.org/10.1371/journal.pcbi.1006256.g007
Белки с высоким обогащением по составу могут выполнять перекрывающиеся или специализированные молекулярные роли в клетке
Предыдущие исследования пытались связать белки, содержащие LCDs, статистически смещенные домены и гомополимерные повторы, с конкретными клеточными функциями [12,16–18,34,37].Однако одним важным соображением при выводе взаимосвязи между белками с ЖК-дисплеями и клеточными функциями, например, является преобладание белков с несколькими ЖК-дисплеями [3] и ЖК-дисплеев, сильно обогащенных более чем одним типом аминокислот [11,14,18, 36]. Следовательно, попытки связать клеточные функции с определенными типами ЖК-дисплеев без контроля других ЖК-дисплеев в рамках тех же белковых последовательностей, рискуют ошибочно приписать функции несвязанным свойствам белков [12,14,34,36]. Хотя несколько ЖК-дисплеев в одном и том же белке (или несколько типов аминокислот, обогащенных в одном и том же ЖК-дисплее) могут взаимодействовать для создания новых структур или функций, это усложняет интерпретацию роли каждого отдельного типа аминокислот в ЖК-дисплеях.Кроме того, поскольку некоторые типы ЖК-дисплеев более распространены, чем другие, общие попытки связать клеточные функции с ЖК-дисплеями, статистически смещенными доменами или гомополимерными повторами, вероятно, отражают функции, связанные только с наиболее распространенными типами, если их рассматривать как единый унифицированный класс [ 16,37]. Следовательно, окончательное присвоение клеточных функций каждому индивидуальному классу ЖК-дисплеев требует исключения белков с другими типами ЖК-дисплеев.
Чтобы свести к минимуму возможные мешающие эффекты, вносимые белками с множеством участков, обогащенных разными типами аминокислот, модифицированная версия начального расчета, выполненного алгоритмом SEG (а именно энтропия Шеннона; см. Методы), была использована для определения белков только с единственный тип композиционно-обогащенного домена (CED).Чтобы включить наши результаты (которые показывают, что обогащение состава может оказывать биологически значимое влияние на составы, предшествующие пороговому значению алгоритма SEG) в наше определение белков с одним CED, процент составных ячеек, для которых содержалось не менее 75% основных белков. SEG-положительные последовательности (как определено выше) объединяли для создания единого списка CED-содержащих белков для каждой аминокислоты. Затем из набора данных были удалены белки, содержащие несколько типов CED, в результате чего получился неизбыточный набор белков только с одним типом CED.Важно отметить, что этот метод позволяет исключить белки, содержащие более одного типа CED, а также белки с CED, сильно обогащенные более чем одним типом аминокислот.
Анализ терминов онтологии генов
(GO) был выполнен отдельно для каждого размера окна в каждой категории CED. Для каждого типа CED существует сильное перекрытие в терминах обогащенного GO во всем диапазоне размеров окна, предполагая, что связи между функциями и CED, специфичными для остатков, не сильно зависят от длины в этом масштабе.Поэтому для простоты интерпретации значительно расширенные термины GO для каждого размера окна были объединены в единый неизбыточный список расширенных терминов GO для каждого типа CED.
Удаление белков с несколькими типами CED выявляет значительную степень специализации для CED разных типов (рис. 8 и таблица S3), которая часто не наблюдается для CED, если рассматривать ее как одну категорию или когда белки с несколькими CED не являются Исключенный. Например, L-богатые белки преимущественно связаны с функциями на мембранах ER и вакуолей, тогда как I-богатые белки сильнее связаны с транспортом углеводов на плазматической мембране.A-богатые белки связаны с множеством процессов или клеточных компонентов, включая трансляцию, активность протеинкиназы, клеточную стенку и катаболизм углеводов / алкоголя. N-богатые белки сильно (и, возможно, исключительно) связаны с функциями, связанными с транскрипцией, тогда как Q-богатые белки, по-видимому, более слабо связаны с транскрипцией и, вместо этого, связаны с большим разнообразием функций, включая эндоцитоз, проекцию спаривания мембрана и ответ на глюкозу.Наконец, хотя белки клеточной стенки дрожжей часто радикально богаты S / T, после контроля совместного обогащения S и T теми же белками, S-богатые белки сильнее связаны с процессами, связанными с мембранами (клеточная стенка, клеточный зачаток кончик, шейка клеточного зачатка, проекция кончика спаривания и т. д.), активность протеинкиназы и транскрипция, тогда как Т-богатые белки, как правило, связаны со связыванием нуклеиновых кислот и активностью геликазы, с меньшим количеством ассоциаций с мембранными процессами. Следовательно, после контроля наличия нескольких CED в одних и тех же белках специализированные функции проявляются даже среди обычно сгруппированных аминокислот.
Рис. 8. Ячеечная модель, изображающая преобладающие функции CED.
Конкретные CED для остатков связаны как с перекрывающимися, так и с отдельными функциями. Основные клеточные / молекулярные процессы, связанные с каждым типом CED, происходят из значительно обогащенных (с поправкой Бонферрони, p ≤ 0,05) GO-терминов в таблице S3.
https://doi.org/10.1371/journal.pcbi.1006256.g008
Кроме того, CED, обогащенные некоторыми аминокислотами, имеют общие функции, несмотря на удаление белков multi-CED, что предполагает некоторую степень совместной специализации.Например, каждый из D-, E- и K-богатых CED был связан с функциями в ядре / ядрышке, включая процессинг рибосомной РНК, связывание нуклеиновых кислот, транскрипцию и связывание гистона / хроматина. Интересно, что внутренне неупорядоченные домены с противоположными суммарными зарядами (наряду с другими заряженными макромолекулами, такими как нуклеиновые кислоты и полиАДФ-рибоза) могут управлять фазовым разделением или комплексной коацервацией в ядре [62–64]. Возможно, что эти домены, наряду с нуклеиновыми кислотами и другими полиионными молекулами, могут участвовать в ядерных процессах посредством динамической электростатической ассоциации с этими или другими безмембранными ансамблями.Напротив, H-богатые CED связаны с процессами, связанными с транспортом и регулированием ионов цинка. Не было никаких терминов GO, существенно связанных с R-богатыми CED. Однако композиционное обогащение R, по-видимому, ограничено, о чем свидетельствует резкое снижение количества белков с R-богатыми доменами в сторону более высоких максимальных локальных процентных составов (см. Рис. 2), на которое может дополнительно повлиять удаление белков с другие типы CED.
Таким образом, если рассматривать как отдельные классы, различные типы CED могут иметь перекрывающиеся или специализированные роли в ячейке.
Обогащение состава соответствует преимущественной локализации в определенных субклеточных компартментах
Молекулярная специализация, наблюдаемая для CED, указывает на то, что белки с обогащением определенных остатков могут локализоваться в определенных субклеточных компартментах для выполнения своих специализированных функций. Более того, факторы контроля качества белка могут различаться между субклеточными компартментами (см. Обзор [65]), что может вносить вклад в зависимые от состава различия в метаболизме белков.Таким образом, мы применили восходящий подход, чтобы вывести профили состава, связанные с основными субклеточными компартментами (см. Методы).
В основном водные субклеточные компартменты почти исключительно связаны с белками, содержащими домены, обогащенные заряженными остатками, полярными остатками и пролином (Рис. 9; см. Также S12 Рис.). Однако различия в профилях обогащения состава очевидны даже среди родственных водных компартментов. Например, значимые ассоциации с заряженными остатками Q или N достигают более экстремальных процентных составов в ядре, тогда как значимые ассоциации с обогащением P достигают более высоких процентных составов в цитоплазме.Напротив, высоко мембранные внутренние органеллы (например, эндоплазматический ретикулум и аппарат Гольджи) преимущественно связаны с обогащением гидрофобных или ароматических остатков (рис.9 и S13). Вакуоль дрожжей также связана с профилями состава, напоминающими профили мембранных компартментов, с дополнительными более слабыми ассоциациями с обогащением S и C. Для митохондрий наблюдается немного слабых ассоциаций. Стенка дрожжевых клеток сильно связана с обогащением S (вероятно, связана с его способностью к гликозилированию), с дополнительными умеренными ассоциациями с обогащением T и A и слабой связью с умеренным обогащением V (рис. 9 и S14).Как и ожидалось, плазматическая мембрана связана с обогащением различных гидрофобных и ароматических остатков. Однако плазматическая мембрана также в значительной степени связана с обогащением избранного подмножества полярных остатков (а именно, C, G, S и T), дополнительно подтверждая особые роли, наблюдаемые для этих CED на внешней мембране. В самом деле, богатые G CED в значительной степени связаны с транспортом аминокислот (см. Таблицу S3), а богатые S или T CED плазматической мембраны могут иметь перекрывающиеся функции или взаимодействия с S- и T-богатыми CED клеточной стенки.Вместе эти наблюдения показывают, что субклеточные компартменты могут терпеть или предпочитать белки с определенными типами CED.
Рис. 9. Профили обогащения состава, связанные с основными субклеточными компартментами.
Все нанесенные на график точки указывают наборы белков, для которых ассоциация с указанным субклеточным компартментом является статистически значимой (точный тест Фишера с поправкой Бонферонни p <0,05). Теплые цвета (красный, оранжевый и желтый) соответствуют заряженным остаткам.Зеленые цвета обозначают полярные остатки. Холодные цвета (пурпурный и синий) соответствуют гидрофобным и ароматическим остаткам соответственно.
https://doi.org/10.1371/journal.pcbi.1006256.g009
Компоненты стресс-гранул и обрабатывающих тел обладают общими и уникальными композиционными особенностями
Недавние наблюдения показывают, что различные Q / N-богатые и G-богатые домены могут образовывать высокодинамичные богатые белком капли в водной среде [25–30], процесс, называемый разделением фаз жидкость-жидкость.Эти типы ЖК-дисплеев преобладают среди компонентов безмембранных органелл, таких как стресс-гранулы и Р-тела [23]. Более того, стрессовые гранулы и Р-тела имеют много общих свойств с богатыми белком жидкими каплями, образованными in vitro , предполагая, что фундаментальные биофизические свойства этих доменов связаны с образованием безмембранных органелл in vivo . Однако, хотя аминокислотный состав признан решающим фактором этого поведения, точные требования к составу, связанные с безмембранными органеллами, остаются в значительной степени неопределенными.
Таким образом, мы также применили наш восходящий подход, чтобы вывести профили обогащения состава, связанные с белковыми компонентами стрессовых гранул и Р-тел (как определено в [66]). Стресс-гранулы и Р-тельца имеют перекрывающиеся белковые компоненты и могут обмениваться белковыми компонентами [67,68], предполагая, что они являются тесно связанными, но разными органеллами. Соответственно, мы наблюдаем как общие, так и уникальные особенности в профилях состава, связанные со стрессовыми гранулами и белками Р-тельца (рис. 10).Как и ожидалось, как стрессовые гранулы, так и Р-тельца прочно связаны с белками, содержащими Q-богатые или N-богатые домены. Например, минимальные составы Q или N, в значительной степени связанные со стрессовыми гранулами, находятся в диапазоне от ~ 15–100% при малых размерах окна (≤30 аминокислот) до ~ 10–30% при больших размерах окна (≥80 аминокислот), хотя эти значения незначительно варьироваться в зависимости от размера окна и остатков. Точно так же минимальные составы Q или N, существенно связанные с P-телами, находятся в диапазоне от ~ 15–100% при малых размерах окна до ~ 10-40% при больших размерах окон.
Рис. 10. Профили состава, связанные с безмембранными органеллами.
Все цветные точки указывают минимальные процентные пороги состава, при которых компоненты стрессовых гранул (A) или P-тел (B) значительно обогащены ( p <0,05). Показаны только аминокислоты, для которых наблюдалось значительное обогащение стрессовых гранул или белков Р-тел по крайней мере в двух ячейках состава. Для большей чувствительности графики были построены с использованием нескорректированных значений p .Поэтому к любой отдельной точке следует относиться с некоторым скептицизмом: однако наличие нескольких последовательных значимых точек в пределах каждого размера окна предполагает, что наблюдаемая тенденция, вероятно, не является артефактом проверки нескольких гипотез.
https://doi.org/10.1371/journal.pcbi.1006256.g010
В дополнение к общепризнанной связи между стрессовыми гранулами / компонентами P-тела и доменами, богатыми Q / N, мы идентифицируем и определяем множество текущих недооцененные композиционные особенности, общие для стресс-гранул и компонентов P-body.Компоненты как стрессовых гранул, так и Р-тел сильно связаны с богатыми P доменами, слабо связаны с доменами, богатыми K, и очень слабо (но в значительной степени) связаны с доменами, богатыми Y. Более того, хотя как стрессовые гранулы, так и Р-тела связаны с белками, содержащими G-богатые домены, компоненты стрессовых гранул связаны с гораздо более широким диапазоном обогащения G, что позволяет предположить, что обогащение G может быть более характерной чертой стрессовых гранул, чем P- тела. Это особенно поразительно в свете недавних наблюдений, показывающих, что высокое содержание глицина помогает поддерживать жидкие характеристики капель с разделенными фазами и предотвращает затвердевание капель in vitro [69].
Кроме того, некоторые особенности состава уникальны как для гранул напряжения, так и для P-тел. Например, компоненты стрессовых гранул в значительной степени связаны с доменами, богатыми A, M, E и R, в то время как компоненты P-телец проявляют незначительное предпочтение или не проявляют никакого предпочтения к этим композиционным характеристикам (ключевую роль аргинина в Также недавно сообщалось о фазовом разделении белков, связанных со стрессовыми гранулами [69]). Напротив, компоненты P-телец слабо связаны с H-богатыми доменами, тогда как компоненты стрессовых гранул не обогащены среди белков, содержащих H-богатые домены.
Насколько нам известно, это первая попытка систематически определить диапазон аминокислотных составов, связанных с безмембранными органеллами, такими как стресс-гранулы и Р-тела. Эти наблюдения указывают на то, что компоненты родственных безмембранных органелл имеют частично совпадающие, но разные композиционные предпочтения. Возможно, что общие композиционные особенности облегчают физическое взаимодействие между стрессовыми гранулами и Р-телами и позволяют обмениваться компонентами, в то время как различия в композиционных характеристиках облегчают их способность функционировать как независимые органеллы.
Обсуждение
Белковые домены, классифицируемые как низко-сложные, статистически предвзятые или гомополимерные, охватывают широкие, гетерогенные классы последовательностей с различными физическими свойствами и клеточными функциями. Эти домены могут играть важную роль в нормальных и патологических процессах. Однако проблемы с категоризацией белков на основе сложности последовательности или статистической ошибки до сих пор препятствовали полному, общему для протеома взгляду на эффекты этих доменов на регуляцию и функцию белков.Здесь мы применяем альтернативный, беспристрастный подход для изучения протеомных взаимосвязей между локальным обогащением аминокислот и рождением, численностью, функциями, субклеточной локализацией и гибелью белков. Почти для всех аминокислот прогрессирующее локальное обогащение соответствует четким порогам перехода в отношении эффективности трансляции, изобилия и периода полужизни белка. Пороги перехода повсеместно встречались в композициях, предшествующих тем, которые требуются для классификации как низко-сложные или статистически смещенные традиционными методами, что указывает на то, что наблюдаемые нами пороги перехода более точно отражают биологически значимые критерии состава.
Последовательности белков могут варьироваться от совершенно разных (т. Е. Полностью гомогенной смеси аминокислот с максимальным расстоянием между идентичными аминокислотами) до отсутствия какого-либо разнообразия (т. Е. Гомополимерных последовательностей). В то время как гомополимерные области представляют собой крайность в этом спектре и могут влиять на метаболизм белков [37], классически определенные гомополимерные области не являются абсолютно необходимыми для этих эффектов (см. S9 – S11 Figs). Это предполагает, что обогащение состава может влиять на метаболизм белка даже при некоторой степени дисперсии первичной последовательности (т.е.е. большее линейное расстояние между идентичными аминокислотами). Определение пределов этой дисперсии может пролить дополнительный свет на взаимосвязь между аминокислотным составом и метаболизмом белка.
Преимущество оценки композиционного обогащения (в отличие от сложности последовательности) заключается в возможности различать эффекты композиционного обогащения для каждого типа аминокислот. Характер тенденций в эффективности трансляции, обилии белка и периоде полужизни белка зависит от аминокислоты, обогащенной в белковых последовательностях, что указывает на то, что локальное обогащение различными аминокислотами может иметь противоположные эффекты.Это подчеркивает ключевое ограничение при рассмотрении доменов низкой сложности, статистически предвзятых или гомополимерных доменов как одного класса — группирование доменов, состоящих из радикально разных аминокислот, эффективно искажает любые наблюдаемые тенденции в сторону наиболее распространенных типов и, в некоторых случаях, может полностью маскируют эффекты менее распространенных доменов низкой сложности, статистически предвзятых или гомополимерных доменов. Более того, даже группирование этих доменов на основе общих физико-химических свойств может вызвать такие же сложности.Примером являются неароматические гидрофобные аминокислоты; в то время как I-богатые, L-богатые и M-богатые домены связаны с низкой эффективностью трансляции, низкой численностью и быстрой скоростью деградации, A-богатые и V-богатые домены связаны с высокой эффективностью трансляции, высокой численностью и медленной деградацией. показатель. Кроме того, клеточные функции, связанные с доменами, обогащенными гидрофобными остатками, имеют тенденцию различаться; L-богатые домены преимущественно связаны с ER или вакуольной мембраной, тогда как I-богатые домены преимущественно связаны с транспортом углеводов на плазматической мембране.Сходным образом N-богатые домены сильно связаны с процессами, связанными с транскрипцией, тогда как Q-богатые домены более сильно связаны с эндоцитозом и другими процессами в цитоплазме. Хотя между этими двумя группами существует некоторое совпадение, это предполагает, что домены, обогащенные очень похожими аминокислотами, все же могут быть предпочтительными для выполнения специализированных ролей в клетке.
Наконец, восходящее применение нашего композиционно-ориентированного алгоритма к безмембранным органеллам обеспечивает первый шаг в определении отдельных композиционных профилей, связанных с каждым типом органелл.Мы обнаружили, что даже тесно связанные и физически взаимодействующие органеллы связаны с заметными различиями в обогащении состава, что может быть связано с различиями в их свойствах, регуляции и функции in vivo . Важно отметить, что, хотя наблюдаемые тенденции в обогащении состава в значительной степени связаны с белками стрессовых гранул или белками P-тельца как соответствующими группами, эти особенности могут не быть абсолютно необходимыми для включения отдельных белков в стрессовые гранулы и / или P -тела.Возможно, например, что два белка, обладающие неперекрывающимися подмножествами связанных композиционных характеристик, все еще могут быть задействованы в стрессовых гранулах, и что некоторые белки стрессовых гранул могут быть задействованы по причинам, полностью отличным от композиционного обогащения (например, через связывание РНК). домены). Можно даже представить, что различия в композиционных характеристиках, хотя все еще позволяют задействовать стрессовые гранулы и / или Р-тела, могут способствовать различиям в динамике отдельных белковых компонентов (например,грамм. кинетика входа / выхода, время выдержки, сила взаимодействий или глубина проникновения в гранулу напряжения / P-тело). Следовательно, хотя наблюдаемые здесь диапазоны ассоциированного состава в совокупности обогащены белками, связанными с этими безмембранными органеллами, каждый отдельный белок не обязательно должен обладать всеми характеристиками состава одновременно, чтобы функционировать как стрессовая гранула или белок Р-тельца.
В то время как большое внимание по праву уделяется пониманию влияния первичной аминокислотной последовательности на судьбы белков (включая фолдинг, регуляцию и функции), все больше полагают, что аминокислотный состав управляет множеством клеточных и молекулярных процессов.Здесь мы разработали подход для изучения взаимосвязи между локальным композиционным обогащением и судьбами белков для каждой из канонических аминокислот в отсутствие априорных предположений или заранее определенных пороговых значений. Наши результаты обеспечивают последовательный, протеомный взгляд на взаимосвязь между композиционным обогащением и фундаментальными аспектами жизненного цикла белка, субклеточной локализации и функции в модельных эукариотических организмах.
Методы
Алгоритм сканирования протеома на основе композиции
белковых последовательностей анализировали с использованием модуля анализа последовательностей FASTA из пакета Biopython [70].Для каждой аминокислоты в наборе из 20 канонических аминокислот каждый белок в транслированном протеоме ORF (последний выпуск с веб-сайта базы данных генома Saccharomyces, последнее изменение 13 января 2015 г.) или кодирующие последовательности ORF (идентификатор организма: UP000001940_6239, дата выпуска 23 мая 2018 г., скачанный с веб-сайта UniProt) сканировали с использованием скользящего окна определенного размера (от 10 до 100 аминокислот с шагом 10). Процентный состав представляющей интерес аминокислоты (AAoI) рассчитывается для каждого окна, и белок сортируется по ячейкам на основе максимального процентного состава, достигнутого для AAoI (в диапазоне от 0 до 100 процентов состава с шагом 5 процентов).Анализы были выполнены для всех возможных комбинаций AAoI, размера окна и процентного состава.
Нормализованная эффективность трансляции (nTE)
Эффективность трансляции для каждого гена оценивалась с использованием шкалы нормализованной эффективности трансляции (nTE) [55], которая основана на количестве копий гена тРНК, эффективности спаривания оснований колебания кодон-антикодон и использовании кодонов в масштабе всего транскриптома. Однако исходный алгоритм nTE отображает все значения nTE для каждого кодона, чтобы создать отдельный профиль эффективности трансляции для каждого гена.Чтобы свести информацию об эффективности трансляции к одному значению для каждого гена (способом, аналогичным индексу адаптации тРНК; [60]), среднее геометрическое значение nTE по транскрипту было рассчитано как
(1)
где nTE iks представляет собой значение эффективности трансляции кодона i -го, определенного k -м триплетом в нуклеотидной последовательности s , а l s представляет длину нуклеотидная последовательность, исключая стоп-кодоны.Следовательно, значения nTE, представленные в настоящем исследовании, представляют собой значения nTE для всего гена. Анализ nTE проводился с использованием собственного скрипта Python.
Определение энтропии Шеннона, статистического смещения аминокислот и гомополимерных повторов
Энтропия Шеннона каждой последовательности рассчитывалась как
(2)
где N представляет размер алфавита остатков ( N = 20, для канонических аминокислот), n i представляет количество вхождений i -го остатка в заданном окне последовательности длиной L .Для сравнения с установленными показателями сложности последовательности мы определили области низкой сложности, используя размер окна по умолчанию (12 аминокислот) и порог энтропии Шеннона (SE ≤ 2,2 бита), используемые на первом проходе алгоритма SEG для первоначальной идентификации ЖК-дисплеев [ 1,10].
В алгоритме SEG вектор состояния сложности, используемый для вычисления энтропии Шеннона, не учитывает аминокислотный состав (то есть значения n i в уравнении 2 не относятся к их соответствующим аминокислотам).Следовательно, если указано, чтобы различать ЖКД на основе преобладающей аминокислоты, последовательности, для которых SE ≤ 2,2 бита и n AAoI ≥ n max в пределах состояния сложности ( что указывает на то, что AAoI вносит основной вклад в классификацию последовательности как ЖКД) были отнесены к соответствующей аминокислотной категории (например, ЖК-дисплеи с высоким содержанием А, ЖК-дисплеи с высоким содержанием углерода и т. д.). Белки Single-LCD / CED — это белки, классифицируемые как LCD или CED, которые не появляются в списках LCD / CED для нескольких аминокислот.
Статистическая ошибка аминокислот рассчитывалась, как описано в [12]. Вкратце, подпоследовательность с наименьшей вероятностью для каждого белка определялась путем тщательного сканирования белков с размерами окна в диапазоне от 25 до 2500 аминокислот. Для каждого окна вероятность смещения подпоследовательности (P смещение ) была определена как
(3)
где w обозначает размер окна, n обозначает количество появлений интересующей аминокислоты в подпоследовательности, а f x обозначает долю представляющей интерес аминокислоты в транслируемом протеоме дрожжей. .Подпоследовательность с наименьшей вероятностью для каждого белка — это подпоследовательность с наименьшим смещением P .
Подходящий порог для определения статистически смещенных белков в дрожжевом белке был определен, как описано ранее [12], за исключением того, что использовались более мягкие критерии для включения дополнительных белков с меньшими крайними смещениями. Вкратце, смещение P , соответствующее субпоследовательности с наименьшей вероятностью (P мин ) для каждого белка, наносили на логарифмический график в зависимости от длины последовательности целого белка.Линия была подогнана, затем Y-пересечение было уменьшено до тех пор, пока только 15% протеома не имели значений P мин ниже линии (в предыдущих анализах использовалось более строгое ограничение в 10% для определения статистически смещенных белков [12]. ). Кроме того, независимый от длины порог был обозначен как значение P min , при котором 15% протеома имели меньшие абсолютные значения P min . Этот порог использовался, когда он был меньше порога P мин , заданного методом, зависящим от длины, чтобы избежать необоснованно ослабленных критериев смещения для небольших последовательностей белка.Аминокислотное смещение было рассчитано с использованием значений только из транслированного протеома orf и реализовано с помощью собственного скрипта Python с предварительно рассчитанными справочными таблицами для повышения вычислительной эффективности.
Белки, содержащие гомополимерные последовательности, были определены просто как любой белок с подпоследовательностью из пяти или более смежных остатков одной и той же аминокислоты, как описано ранее [37].
Данные о содержании и периоде полужизни белка
Значения содержания дрожжевого белка (в среднем количестве молекул на клетку на белок) были получены из [48] ( n = 5,391).Значения содержания белка для C . elegans были получены из [59]. Данные о периоде полужизни дрожжевого белка были получены из [49]. Для простоты интерпретации в наборы данных были включены только белки с однозначными ненулевыми значениями периода полураспада или содержания. Белки, перечисленные в отдельных строках с идентичными значениями периода полураспада или содержания, были сохранены, тогда как измерения периода полужизни или содержания белка, относящиеся к более чем одному белку одной и той же линии, были исключены (часто это были высокогомологичные гены, что позволяет предположить, что измерение не могло однозначно относиться к одному из белков).Кроме того, все белки, соответствующие измерениям с «низкой достоверностью» в наборе данных о периоде полужизни, были исключены (критерии см. В [49]). n = 3,525 для отфильтрованного набора данных о периоде полураспада дрожжей и n = 5 952 для отфильтрованного C . elegans набор данных о содержании белка.
Статистика и построение
Для всех бинов AAoI / размер окна / процент состава, распределение значений nTE, численности или периода полужизни для белков, включенных в данную ячейку, сравнивали с распределением соответствующих значений всех белков, исключенных из данной ячейки.Статистическая значимость оценивалась с использованием двустороннего критерия Манна-Уитни и (также называемого критерием суммы рангов Вилкоксона; подробное описание и обоснование см. В дополнительных экспериментальных процедурах из [47]). Где указано, значения p были скорректированы в каждом окне с использованием метода коррекции Бонферрони для проверки множественных гипотез. Все статистические тесты были выполнены с использованием модулей, доступных в пакете SciPy, с настройками по умолчанию, если не указано иное.Все графики были созданы с использованием модулей Matplotlib или Seaborn.
Анализ обогащения терминов генной онтологии (GO)
Тесты обогащения по
GO-члену были выполнены с использованием пакета GOATOOLS (версия 0.7.9) [71] для каждого набора белков, содержащихся в заданной аминокислоте / размере окна / процентном соотношении состава. Для каждого теста набор фоновых белков определяли как все белки из транслированного протеома ORF с длиной последовательности, большей или равной заданному размеру окна. Все зарегистрированные значения p были скорректированы с использованием поправки Бонферрони во время временной ассоциации GO.Чтобы оценить составные профили обогащения, связанные с терминами GO, относящимися к субклеточным компартментам, мы применили подход минимального порогового сканирования ко всем разделенным протеомам. Для каждого AAoI, размера окна и корзины процентного состава все белки с максимальным локальным составом, превышающим или равным текущему рассматриваемому процентному составу, объединяются и оцениваются на предмет возможных условий обогащения GO. Это эффективно оценивает возможное обогащение терминов GO итеративно с увеличением критериев максимального локального состава.Результаты GO-термина были впоследствии оценены на предмет значительного обогащения одного GO-термина, описывающего каждый субклеточный компартмент (или двух родственных GO-терминов, «внешняя мембрана» и «плазматическая мембрана» в случае плазматической мембраны). Значения p были дополнительно скорректированы в пределах каждого размера окна с использованием метода коррекции Бонферрони.
Подобные анализы были выполнены для наборов экспериментально определенных стрессовых гранул ( n = 83) и белков P-body ( n = 52) белков [66].В частности, подход минимального порогового сканирования был применен ко всем разделенным протеомам. Для каждого AAoI, размера окна и бункера процентного состава объединяются все белки с максимальным локальным составом, большим или равным текущему рассматриваемому процентному составу. Значительное обогащение экспериментально определенных стрессовых гранул или белков Р-тел в каждом пуле белков оценивали с использованием точного критерия Фишера ( p <0,05).
Вспомогательная информация
S1 Таблица.Максимальные значения локального состава для каждой аминокислоты и комбинации размера окна для всех транслированных дрожжевых белков.
Для каждого белка в транслированном протеоме ORF указаны значения nTE, содержание белка и период полужизни белка. nTE рассчитывали по методике, описанной в [55]. Значения содержания и периода полужизни белка были представлены в [48] и [54] соответственно. Остальные столбцы содержат максимальное значение локального состава (на 100) для каждой аминокислоты и комбинацию размера окна для каждого белка.
https://doi.org/10.1371/journal.pcbi.1006256.s001
(CSV)
S2 Таблица. Максимальные значения локального состава для каждой аминокислоты и комбинации размера окна для всех белков C. elegans
.
Для каждого белка в C . Протеом elegans , указаны значения обилия белка (средняя измеренная интенсивность в 3 биологических повторностях из [59]; критерии включения см. В разделе «Методы»). Остальные столбцы содержат максимальное значение локального состава (на 100) для каждой аминокислоты и комбинацию размера окна для каждого белка. https://doi.org/10.1371/journal.pcbi.1006256.s002 (CSV) Все перечисленные термины GO представляют термины, значимо связанные с набором специфичных для остатков CED по крайней мере для одного размера окна (с поправкой Бонферрони p ≤ 0,05). https://doi.org/10.1371/journal.pcbi.1006256.s003 (XLSX) Локальное обогащение отдельных аминокислот соответствует зависимым от состава изменениям в экспериментально полученной эффективности синтеза белка [56]. https://doi.org/10.1371/journal.pcbi.1006256.s004 (TIF) C. elegans на основе максимального локального аминокислотного состава.
Количество белков, разделенных на каждый размер окна / процентный интервал состава для каждой из 20 канонических аминокислот, нанесено на график как функция максимального локального состава для каждого размера окна.Точки разброса соединены отрезками линии только для наглядности. https://doi.org/10.1371/journal.pcbi.1006256.s005 (TIF) Для каждой аминокислоты значения nTE, соответствующие белкам, разделенным на заданный размер окна и процентный интервал состава, сравнивали со значениями для всех белков длиной ≥ соответствующего размера окна, которые были исключены из интервала.Красные и синие точки обозначают ячейки, для которых распределение значений периода полужизни белка значительно отличается (с поправкой Бонферрони, p ≤ 0,05) от таковых для исключенных белков: красные точки указывают ячейки с более низким медианным значением по сравнению с таковым для исключенных белков, тогда как синие точки обозначают интервалы с более высоким относительным медианным значением. Серые точки указывают на отсутствие статистической значимости сравнений. Индивидуальные точки масштабируются в пределах каждого участка, чтобы отразить размеры образцов белков, содержащихся в каждом бункере. https://doi.org/10.1371/journal.pcbi.1006256.s006 (TIF) Для каждой аминокислоты значения обилия белков, соответствующие белкам, разделенным на заданный размер окна и процентный интервал состава, сравнивали со значениями для всех белков длиной ≥ соответствующего размера окна, которые были исключены из интервала.Красные и синие точки обозначают ячейки, для которых распределение значений периода полужизни белка значительно отличается (с поправкой Бонферрони, p ≤ 0,05) от таковых для исключенных белков: красные точки указывают ячейки с более низким медианным значением по сравнению с таковым для исключенных белков, тогда как синие точки обозначают интервалы с более высоким относительным медианным значением. Серые точки указывают на отсутствие статистической значимости сравнений. Индивидуальные точки масштабируются в пределах каждого участка, чтобы отразить размеры образцов белков, содержащихся в каждом бункере. https://doi.org/10.1371/journal.pcbi.1006256.s007 (TIF) Для каждой аминокислоты значения периода полужизни белка, соответствующие белкам, разделенным на заданный размер окна и процентный интервал состава, сравнивали со значениями для всех белков длиной ≥ соответствующего размера окна, которые были исключены из интервала.Красные и синие точки обозначают ячейки, для которых распределение значений периода полужизни белка значительно отличается (с поправкой Бонферрони, p ≤ 0,05) от таковых для исключенных белков: красные точки указывают ячейки с более низким медианным значением по сравнению с таковым для исключенных белков, тогда как синие точки обозначают интервалы с более высоким относительным медианным значением. Серые точки указывают на отсутствие статистической значимости сравнений. Индивидуальные точки масштабируются в пределах каждого участка, чтобы отразить размеры образцов белков, содержащихся в каждом бункере. https://doi.org/10.1371/journal.pcbi.1006256.s008 (TIF) Для каждой аминокислоты значения nTE, соответствующие белкам, разделенным на заданный размер окна и процентный интервал состава, сравнивали со значениями для всех белков длиной ≥ соответствующего размера окна, которые были исключены из интервала. Красные и синие точки обозначают интервалы, для которых распределение значений периода полужизни белка значительно различается (с поправкой Бонферрони p ≤ 0.05) от значений исключенных белков: красные точки указывают ячейки с более низким медианным значением по сравнению со средним значением исключенных белков, тогда как синие точки указывают ячейки с более высоким относительным медианным значением. Серые точки указывают на отсутствие статистической значимости сравнений. Индивидуальные точки масштабируются в пределах каждого участка, чтобы отразить размеры образцов белков, содержащихся в каждом бункере. https://doi.org/10.1371/journal.pcbi.1006256.s009 (TIF) Для каждой аминокислоты значения обилия белков, соответствующие белкам, разделенным на заданный размер окна и процентный интервал состава, сравнивали со значениями для всех белков длиной ≥ соответствующего размера окна, которые были исключены из интервала. Красные и синие точки обозначают ячейки, для которых распределение значений периода полужизни белка значительно отличается (с поправкой Бонферрони, p ≤ 0,05) от таковых для исключенных белков: красные точки указывают ячейки с более низким медианным значением по сравнению с таковым для исключенных белков, тогда как синие точки обозначают интервалы с более высоким относительным медианным значением.Серые точки указывают на отсутствие статистической значимости сравнений. Индивидуальные точки масштабируются в пределах каждого участка, чтобы отразить размеры образцов белков, содержащихся в каждом бункере. https://doi.org/10.1371/journal.pcbi.1006256.s010 (TIF) Для каждой аминокислоты значения периода полужизни белка, соответствующие белкам, разделенным на заданный размер окна и процентный интервал состава, сравнивали со значениями для всех белков длиной ≥ соответствующего размера окна, которые были исключены из интервала.Красные и синие точки обозначают ячейки, для которых распределение значений периода полужизни белка значительно отличается (с поправкой Бонферрони, p ≤ 0,05) от таковых для исключенных белков: красные точки указывают ячейки с более низким медианным значением по сравнению с таковым для исключенных белков, тогда как синие точки обозначают интервалы с более высоким относительным медианным значением. Серые точки указывают на отсутствие статистической значимости сравнений. Индивидуальные точки масштабируются в пределах каждого участка, чтобы отразить размеры образцов белков, содержащихся в каждом бункере. https://doi.org/10.1371/journal.pcbi.1006256.s011 (TIF) Для каждой аминокислоты значения nTE, соответствующие белкам, разделенным на заданный размер окна и процентный интервал состава, сравнивали со значениями для всех белков длиной ≥ соответствующего размера окна, которые были исключены из интервала. Красные и синие точки обозначают интервалы, для которых распределение значений периода полужизни белка значительно различается (с поправкой Бонферрони p ≤ 0.05) от значений исключенных белков: красные точки указывают ячейки с более низким медианным значением по сравнению со средним значением исключенных белков, тогда как синие точки указывают ячейки с более высоким относительным медианным значением. Серые точки указывают на отсутствие статистической значимости сравнений. Индивидуальные точки масштабируются в пределах каждого участка, чтобы отразить размеры образцов белков, содержащихся в каждом бункере. https://doi.org/10.1371/journal.pcbi.1006256.s012 (TIF) Для каждой аминокислоты значения обилия белков, соответствующие белкам, разделенным на заданный размер окна и процентный интервал состава, сравнивали со значениями для всех белков длиной ≥ соответствующего размера окна, которые были исключены из интервала. Красные и синие точки обозначают ячейки, для которых распределение значений периода полужизни белка значительно отличается (с поправкой Бонферрони, p ≤ 0,05) от таковых для исключенных белков: красные точки указывают ячейки с более низким медианным значением по сравнению с таковым для исключенных белков, тогда как синие точки обозначают интервалы с более высоким относительным медианным значением.Серые точки указывают на отсутствие статистической значимости сравнений. Индивидуальные точки масштабируются в пределах каждого участка, чтобы отразить размеры образцов белков, содержащихся в каждом бункере. https://doi.org/10.1371/journal.pcbi.1006256.s013 (TIF) Для каждой аминокислоты значения периода полужизни белка, соответствующие белкам, разделенным на заданный размер окна и процентный интервал состава, сравнивали со значениями для всех белков длиной ≥ соответствующего размера окна, которые были исключены из интервала.Красные и синие точки обозначают ячейки, для которых распределение значений периода полужизни белка значительно отличается (с поправкой Бонферрони, p ≤ 0,05) от таковых для исключенных белков: красные точки указывают ячейки с более низким медианным значением по сравнению с таковым для исключенных белков, тогда как синие точки обозначают интервалы с более высоким относительным медианным значением. Серые точки указывают на отсутствие статистической значимости сравнений. Индивидуальные точки масштабируются в пределах каждого участка, чтобы отразить размеры образцов белков, содержащихся в каждом бункере. https://doi.org/10.1371/journal.pcbi.1006256.s014 (TIF) Указаны диапазоны состава для каждой аминокислоты, в значительной степени связанной с цитоплазмой (A) и ядром (B). Все нанесенные на график точки указывают на наборы белков, для которых ассоциация с указанным субклеточным компартментом является статистически значимой (с поправкой Бонферонни p <0.05). Графики показаны только для аминокислот с по меньшей мере двумя ячейками состава, значимо связанными с указанным субклеточным компартментом. https://doi.org/10.1371/journal.pcbi.1006256.s015 (TIF) Указаны диапазоны состава для каждой аминокислоты, в значительной степени связанной с эндоплазматическим ретикулумом (A), аппаратом Гольджи (B), вакуолью (C) и митохондриями (D).Все нанесенные на график точки указывают наборы белков, для которых ассоциация с указанным субклеточным компартментом является статистически значимой (с поправкой Бонферонни p <0,05). Графики показаны только для аминокислот с по меньшей мере двумя ячейками состава, значимо связанными с указанным субклеточным компартментом. https://doi.org/10.1371/journal.pcbi.1006256.s016 (TIF) Указаны диапазоны состава для каждой аминокислоты, в значительной степени связанной с плазматической мембраной (A) и клеточной стенкой (B). Все нанесенные на график точки указывают наборы белков, для которых ассоциация с указанным субклеточным компартментом является статистически значимой (с поправкой Бонферонни p <0,05). Графики показаны только для аминокислот с по меньшей мере двумя ячейками состава, значимо связанными с указанным субклеточным компартментом. https://doi.org/10.1371/journal.pcbi.1006256.s017 (TIF) Наше исследование состояло из рандомизированного двойного слепого протокола, в котором участвовали люди, которым давали 48 граммов риса или 48 граммов изолята сывороточного протеина после тренировки с отягощениями (фаза 1) и после каждой тренировки во время тренировки. 8-недельный периодизированный тренировочный протокол (Фаза 2).На этапе 1 исследования изучалось влияние источников белка на восстановление через 48 часов после интенсивной тренировки с отягощениями, ориентированной на гипертрофию. Вторая фаза наступила для оставшегося восьминедельного протокола RT, который состоял из тренировки каждой группы мышц два раза в неделю с использованием нелинейной периодизированной модели RT. Прямое ультразвуковое определение мышечной массы, двойная рентгеновская абсорбциометрия (DXA), определяющая состав тела, максимальная сила и мощность, оценивались коллективно в конце 0, 4 и 8 недель. В исследовании приняли участие 24 здоровых мужчины (21,3 ± 1,9 года, 76,08 ± 5,6 кг, 177,8 ± 12,3 см). В качестве критериев включения требовалось, чтобы все субъекты прекратили прием пищевых добавок за три месяца до исследования, участвовали в ЛТ не менее 3 раз в неделю в течение последних шести месяцев и имели как минимум 1-летний опыт ЛТ. Субъекты были тщательно подобраны по возрасту, массе тела, силе и опыту тренировок с отягощениями, а затем случайным образом помещены в группу риса (n = 12) или сыворотки (n = 12).Все процедуры были одобрены Экспертным советом Университета Тампы. Все испытуемые участвовали в тренировке с отягощениями большого объема, состоящей из 3 подходов: жима ногами, жима лежа и военного жима, подтягиваний, тяги в наклоне, сгибаний со штангой и разгибаний. Сразу после тренировки испытуемые потребляли 48 граммов RPI или WPI соответственно. Непосредственно перед тренировкой и через 48 часов после тренировки были проведены измерения болезненности, предполагаемой готовности к тренировке и шкалы предполагаемого восстановления (PRS).Болезненность измерялась по визуальной аналоговой шкале от 0 до 10. При этом ноль означает отсутствие болезненных ощущений в мышцах, а 10 означает самую сильную мышечную болезненность из когда-либо испытанных. PRS состоит из значений от 0 до 10, где 0–2 — очень плохо с ожидаемым снижением производительности, 4–6 — от низкого до умеренного с ожидаемой аналогичной производительностью, а 8–10 — с высоким восприятием восстановления с ожидаемым увеличением производительности. Воспринимаемая готовность показывает, насколько испытуемый чувствовал себя готовым к тренировке.По этой шкале 10 — это максимальная готовность человека к тренировкам, а 0 означает, что испытуемый чувствует, что он совсем не готов к тренировкам. Наш протокол тренировки с отягощениями представлял собой модифицированную комбинацию Kraemer et al. [22] и Монтейро и др. [3]. Эти исследователи обнаружили, что у спортсменов нелинейная программа тренировок с отягощениями дала лучшие результаты, чем традиционная или непериодизированная программа. Программа была разработана для тренировки всех основных групп мышц с использованием в основном сложных движений для верхней и нижней части тела.Запрограммированный нелинейный тренировочный сплит был разделен на дни гипертрофии, состоящие из 8–12 нагрузок ПМ на 3 подхода, с отдыхом 60–120 секунд и силовые дни, состоящие из 2–5 нагрузок ПМ на 3 подхода для всех упражнений, кроме жима ногами. и жим лежа, в котором было выполнено 5 подходов. По мере выполнения предписанных повторений веса постепенно увеличивались на 2–5%. Исследователи внимательно следили за всеми тренировками, чтобы гарантировать максимальные усилия и интенсивность на каждой тренировке. Сила оценивалась с помощью теста 1-RM жима ногами и жима лежа. Каждое упражнение считалось успешным в соответствии с правилами Международной федерации пауэрлифтинга [23]. Состав тела (безжировая масса, жировая масса и общая масса) определяли на приборе Lunar Prodigy DXA (версия программного обеспечения, enCORE 2008, Мэдисон, Висконсин, США). Гипертрофию скелетных мышц определяли с помощью изменений в ультразвуковом исследовании, определяя комбинированную толщину двуглавой мышцы плеча и широкой мышцы бедра (VL) и широкой мышцы бедра (VI) (General Electric Medical Systems, Милуоки, Висконсин, США). Мощность оценивалась во время теста максимальной велосипедной эргометрии. Во время теста на велосипеде добровольцев проинструктировали ехать на велосипеде с заданным сопротивлением (7,5% веса тела) как можно быстрее в течение 10 секунд [24]. Высота седла регулировалась в соответствии с ростом человека, чтобы обеспечить сгибание колена на 5–10 °, в то время как ступня находилась в нижнем положении центральной полости. Испытуемым был предоставлен стандартный вербальный стимул. Выходная мощность регистрировалась в реальном времени компьютером, подключенным к стандартному велоэргометру Monark (модель Monark 894e, Vansbro, Швеция) во время 10-секундного спринтерского теста.Пиковую мощность (PP) регистрировали с использованием программного обеспечения для анаэробных испытаний Monark (Monark Anaerobic Wingate Software, версия 1.0, Monark, Vansbro, Швеция). После завершения испытаний крыла, проведенных в течение нескольких дней, коэффициент межклассовой корреляции для пиковой мощности составил 0,96. За две недели до и на протяжении всего исследования испытуемые были переведены на диету, состоящую из 25% белка, 50% углеводов и 25% жира, зарегистрированным диетологом, специализирующимся на спортивном питании.Испытуемые встречались в группе с диетологом, и в начале исследования им давали индивидуальные планы питания. Ежедневное количество калорий определялось по уравнению Харриса-Бенедикта и отслеживалось еженедельными журналами для обеспечения соответствия. Белковая добавка вводилась под наблюдением лаборанта после тренировки с отягощениями, и она состояла из 48 г изолята сывороточного протеина (Nutra Bio Whey Protein Isolate (Dutch Chocolate), Мидлсекс, штат Нью-Джерси) или 48 г изолята рисового протеина (Growing Изолят рисового протеина Naturals (шоколадная сила), изготовленный из рисового протеина Oryzatein ® , Axiom Foods, Oro Valley, AZ), растворенного в 500 мл воды.Аминокислотный профиль исследуемого материала был проанализирован независимой аналитической лабораторией (Eurofins Analytical Laboratories, Metairie, LA) и показан в таблице 2. И добавка сывороточного белка, и добавка рисового белка были изоназотными, изокалорийными и соотношениями макроэлементов совпадающими. Таблица 2 Все добавки были протестированы HFL Sports Science перед использованием, чтобы гарантировать отсутствие загрязнения стероидами или стимуляторами в соответствии с аккредитованными тестами ISO 17025. Модель ANOVA использовалась для измерения группового, временного и группового по времени взаимодействий как для фазы 1, так и для фазы 2. Если наблюдались какие-либо основные эффекты, применялся апостериорный метод Тьюки, чтобы определить, где произошли различия. Вся статистика была проведена с использованием программного обеспечения Statistica (Statsoft, 2011). Детальное знание состава и структуры сплайсосомы и ее промежуточных соединений является предпосылкой для понимания сложного процесса сплайсинга пре-мРНК.С этой целью мы разработали метод аффинного отбора тобрамицина, который обычно применим для очистки природных комплексов РНП. Используя этот метод, мы выделили пресплицеосомы человека, которые идеально подходят как для биохимических, так и для структурных исследований. MS идентифицировала> 70 белков, связанных с пресплицеосомами, включая почти все известные белки snRNP U1 и U2, а также ожидаемые факторы сплайсинга, не относящиеся к snRNP. Кроме того, были обнаружены DEAD-бокс-белок p68, РНК-геликаза A и ряд белков, которые, по-видимому, выполняют в клетке множество функций, таких как YB-1 и TLS.Также были идентифицированы несколько ранее не охарактеризованных белков с неизвестной функцией, предполагая, что они играют роль в сплайсинге и потенциально действуют во время сборки пресплицеосом. Эти данные дают представление о сложности сварочного оборудования на ранней стадии его сборки. Сплайсосомы, сложные ферменты, ответственные за сплайсинг пре-мРНК, состоят из малых ядерных рибонуклеопротеидов (snRNP) U1, U2, U5 и U4 / U6, а также множества белков, не относящихся к snRNP. Во время пошаговой сборки сплайсосом несколько различных комплексов, которые образуются в определенном порядке (т.е., комплекс E, за которым следуют A, B и C), можно биохимически выделить in vitro (см. ссылки 1 и 2). Перед ассоциацией snRNPs и факторов сплайсинга пре-мРНК связывается с гетерогенными белками ядерного рибонуклеопротеина (hnRNP), образуя H-комплекс. Сборка сплайсосомы инициируется АТФ-независимым взаимодействием мяРНК U1 с 5′-сайтом сплайсинга, которое частично включает спаривание оснований между мяРНК U1 и пре-мРНК. Этот начальный комплекс, названный E (ранний), также содержит U2 snRNP, который на этой стадии слабо связан с пре-мРНК (3).На последующем АТФ-зависимом этапе пары оснований мяРНК U2 с сайтом разветвления приводят к стабильной ассоциации мяРНП U2 и образованию комплекса А (также называемого пресплицеосомой). Контакты между мяРНК / белками и пре-мРНК, которые устанавливаются во время этих ранних стадий сборки сплайсосом, играют решающую роль в распознавании и выборе сайтов сплайсинга. Таким образом, регуляция альтернативно сплайсированных пре-мРНК часто достигается путем модуляции ассоциации сплайсосомных компонентов во время образования E и A.На следующем этапе три-мяРНП U4 / U6.U5 связывается, что приводит к образованию сплайсосомного комплекса B. После значительного конформационного изменения и первой реакции переэтерификации сплайсинга образуется комплекс C. После второй стадии переэтерификации мРНК и вырезанный интрон высвобождаются, и сплайсосома диссоциирует. На сегодняшний день более 200 сплайсосомных белков идентифицировано у млекопитающих с помощью МС (4–6). Однако в этих исследованиях была проанализирована смесь очищенных комплексов сплайсинга, предотвращающая отнесение конкретного белка к конкретному комплексу сплайсинга (4).Совсем недавно аффинный отбор с амилозными гранулами сплайсинговых комплексов, образованных на пре-мРНК, предварительно связанной с мальтозосвязывающим белком, был использован для выделения высокоочищенных нативных комплексов Е и С (3, 7). Однако к настоящему времени белковый состав только сплайсосомного комплекса С детально определен методом МС (7). Таким образом, исчерпывающая информация о составе сплайсинговых комплексов, образованных перед каталитическими стадиями сплайсинга, в настоящее время отсутствует. Точно так же подходы к выделению нативных сплайсинговых комплексов, которые подходят как для биохимических, так и для структурных исследований, весьма ограничены. Здесь мы разработали метод аффинной селекции тобрамицина и использовали его для очистки препаративных количеств пресплицеосом человека в естественных условиях. Анализ МС показал, что пресплицеосомы очень сложные, с> 70 ассоциированными белками. Эти исследования предоставляют подробную информацию о составе пресплицеосом и предполагают роль недавно идентифицированных пресплицеосомных белков на ранних стадиях сборки сплайсосом. Олигонуклеотидов, использованных в этом исследовании, были M13f (GTAAAACGACGGCCAGT), HPV23 (GATCCGGCTTAGTATAGCGAGGTTTTAGCTACACTCGTGCTGAGCCGGATCCGCATG) и HPV24 (CTATTAGATCCGGCTCGTCAGCAGCCCTAG). Некодирующая и кодирующая последовательности аптамер РНК J6f1 (8) подчеркнуты соответственно. Вставку Eco RI- Bam h2 pMINX (9) сначала клонировали в pGEM-3Zf (+), получая pGEM-MINX. Олигонуклеотиды HPV23 и HPV24 были отожжены и клонированы в линеаризованную pGEM-MINX Bam h2 / Sph I с получением pGEM MINX-T5.ПЦР с праймерами HPV24 и M13f генерировала матрицу транскрипции для пре-мРНК, содержащей аптамер. Для получения контрольной пре-мРНК, лишенной аптамера, продукт ПЦР расщепляли Bam HI. Транскрипты с кэпом m7G были синтезированы с помощью РНК-полимеразы Т7 (MegaScript, Ambion, Austin, TX). Для количественного определения [α- 32 P] UTP (3000 Ки / ммоль; 1 Ки = 37 ГБк) добавляли до 0,23 мкМ. N -гидроксисукцинимид-активированная сефароза 4 Fast Flow была дериватизирована 5 мМ тобрамицином, как описано (10). Все дальнейшие процедуры выполнялись при 4 ° C, если не указано иное. Для селекции по аффинности 4 × связывающий буфер (4 × BP) / 80 мМ Tris mHCl, pH 9,1 при 4 ° C / 4 мМ CaCl 2 /4 мМ MgCl 2 /2 мМ DTT был свежеприготовлен. Матрицу тобрамицина блокировали 400 мкл блокирующего буфера (1 × BP / 300 мМ KCl / 0,1 мг / мл тРНК / 0,5 мг / мл BSA / 0,01% Nonidet P-40) на аликвоту 15 мкл головой к хвосту (HOT ) вращение за ночь.Матрикс собирали, 400 мкл смеси, содержащей ≈60 пмоль меченой тобрамицином пре-мРНК и 40 мкг тРНК в связывающем буфере (1 × BP / 145 мМ KCl), добавляли к 15 мкл матрицы и инкубировали в течение 1,5 ч. Затем матрицу трижды промывали (по 750 мкл) связывающим буфером, содержащим 0,1% Nonidet P-40. Было обнаружено, что концентрация пре-мРНК 22-25 нМ является оптимальной для образования комплекса А (не показано). Для стандартного анализа сплайсинга были приготовлены четыре аликвоты по 15 мкл матрицы, связанной с пре-мРНК.Смесь для сплайсинга (1,5 мл) добавляли на 15 мкл матрицы и инкубировали в указанные время и температуру при постоянном горячем перемешивании. Смесь для сплайсинга содержала 35% ядерного экстракта клеток HeLa (11) в буфере D [20 мМ Hepes-KOH, pH 7,9 / 100 мМ KCl / 1,5 мМ MgCl 2 / 0,2 мМ EDTA, pH 8,0 / 0,5 мМ DTT / 0,5 мМ PMSF / 10% (об. / Об.) Глицерин] и добавляли 25 мМ KCl / 3 мМ MgCl 2 /2 мМ АТФ / 20 мМ креатинфосфат. После сращивания реакционные смеси охлаждали на льду и собирали матрицу центрифугированием.Аликвоты матрицы объединяли и трижды промывали 750 мкл промывочного буфера (1 × BP / 75 мМ KCl / 0,1% Nonidet P-40). Комплексы элюировали путем инкубации с 250 мкл буфера для элюции (1 × BP / 5 мМ тобрамицин / 145 мМ KCl / 2 мМ MgCl 2 ) при HOT вращении в течение 10 мин. Обычно можно было элюировать ≈75% связанных комплексов. РНК и белок выделяли и анализировали на гелях 8,3 М мочевина / 10% полиакриламида (РНК) или SDS / 10–13% PAGE (белки), или для сплайсинга на гелях 8,3 М мочевина-15% полиакриламида. Элюат тобрамицина (220 мкл) загружали в 3,8 мл линейный градиент 10–30% глицерина в 20 мМ Hepes-KOH, pH 7,9 / 75 мМ KCl / 1,5 мМ MgCl 2 / 0,5 мМ DTT. Градиенты центрифугировали в течение 107 мин при 60 000 об / мин в роторе Sorvall TH660 и собирали вручную фракциями по 175 мкл сверху. РНК и белок экстрагировали из 100 мкл каждой нечетной фракции и анализировали, как указано выше. Сшивание инициировали добавлением псоралена (4′-аминометил-4,5 ‘, 8-триметилпсорален гидрохлорид) в ДМСО (2 мг / мл) к 75 мкл фракций четного градиента до конечной концентрации 40 мкг / мл и инкубации в течение 10 мин на льду. Образцы облучали светом 365 нм в течение 30 мин при 4 ° C. РНК выделяли (12) и разделяли в геле 5% полиакриламида / 8,3 М мочевины и анализировали методом Нозерн-блоттинга, как описано (13). ДНК-зонды были получены путем случайного праймирования (Prime-it II, Promega) с использованием ДНК pMINX (пре-мРНК) или кодирующих последовательностей мяРНК U1 или U2, амплифицированных с помощью ПЦР, и [α- 32 P] dATP (6000 Ки / ммоль). Белки, разделенные с помощью SDS / PAGE, анализировали с помощью масс-спектрометрии с лазерной десорбцией / ионизацией с использованием матрицы (MALDI-MS) или тандемной масс-спектрометрии с жидкостной хроматографией [LC-MSMS, как описано в исх. 14; для получения дополнительной информации см. Supporting Methods , который опубликован в качестве вспомогательной информации на веб-сайте PNAS, www.pnas.org) и идентифицирован в неизбыточной базе данных Национального центра биотехнологической информации с использованием MASCOT (Matrix Science, Лондон) в качестве поиска. двигатель. Для аффинной очистки сплайсосом в нативных условиях мы ввели аптамер РНК 40 нт на 3′-конец пре-мРНК MINX, полученной из аденовируса. Этот аптамер с высоким сродством (5 нМ) связывает аминогликозидный антибиотик тобрамицин в физиологических условиях (8). Действительно, пре-мРНК MINX, меченная аптамером, эффективно связывалась с гранулами дериватизированной тобрамицином сефарозы, когда реакцию связывания проводили при pH 9.1 в буфере, содержащем 145 мМ KCl и 1 мМ Mg 2+ . В этих условиях связывается 60–70% пре-мРНК, меченной аптамером, но только ≈2% пре-мРНК без метки. Что еще более важно, 80% связанной пре-мРНК можно элюировать путем инкубации с 5 мМ тобрамицином. Меченная аптамером пре-мРНК MINX образовывала сплайсосомные комплексы и подвергалась сплайсингу in vitro с эффективностью и кинетикой, аналогичными таковым для немаркированной пре-мРНК MINX (не показано). Однако в присутствии ядерного экстракта <1% меченой пре-мРНК связывается с матрицей тобрамицина, что позволяет предположить, что аптамер РНК больше не доступен или условия, используемые для сплайсинга, неоптимальны для связывания.Таким образом, твердофазный сплайсинг выполняли, сначала иммобилизуя меченую пре-мРНК на матриксе тобрамицина, а затем инкубируя с ядерным экстрактом в условиях сплайсинга (фиг. 1 A ). Затем матрицу тщательно промывали и связанный материал элюировали тобрамицином. РНК выделяли из элюата или надосадочной жидкости реакции и анализировали денатурирующим ПААГ (фиг. 1 B ). Промежуточные продукты / продукты сплайсинга сначала наблюдались в тех комплексах, которые элюировались из матрицы через 90 мин, и заметно увеличивались через 180 и 270 мин, когда продукт мРНК становился легко обнаруживаемым (рис.1 B , полосы 2–4). Таким образом, сплайсинг протекает со значительно сниженной кинетикой по сравнению с неиммобилизованной, меченой пре-мРНК MINX, которая давала продукты сплайсинга уже через 15 мин (не показано). Большая часть пре-мРНК оставалась связанной с матрицей даже после продолжительного времени инкубации, о чем свидетельствует присутствие только очень низкого уровня (максимум 7%) пре-мРНК и промежуточных продуктов сплайсинга в супернатанте твердофазной реакции сплайсинга. (Рис. 1 B , дорожки 5–7).Однако, в соответствии с его высвобождением из сплайсосом после второго этапа сплайсинга, было обнаружено повышенное количество вырезанного интрона. Эти результаты демонстрируют, что, несмотря на пониженную кинетику реакции, связанная с матриксом пре-мРНК, меченная аптамером, эффективно сплайсируется и, следовательно, функциональные сплайсосомные комплексы E, A, B и C также должны быть собраны. Рис 1. ( A ) Стратегия очистки. ( B ) Связанная с матрицей пре-мРНК, меченная аптамером, эффективно сплайсируется.Твердофазное сращивание выполняли в течение указанного времени при 30 ° C. РНК выделяли после элюирования (дорожки 2–4) или из реакционного супернатанта (дорожки 5–7; обратите внимание, что было проанализировано в три раза больше супернатанта, чем элюата), анализировали денатурирующим PAGE и визуализировали с помощью авторадиографии. Дорожка 1, ввод пре-мРНК. Положения пре-мРНК и промежуточных продуктов / продуктов сплайсинга указаны справа. Чтобы изолировать пресплицеосомы, мы выполнили твердофазный сплайсинг всего за 45 мин, чтобы минимизировать образование комплексов B / C.Затем содержание РНК и белка в элюате тобрамицина анализировали денатурирующим ПААГ или SDS / ПААГ, соответственно. В качестве контроля неспецифических взаимодействий аналогичным образом выполняли отбор по аффинности, но без пре-мРНК. Как показано на фиг. 2 A , в отсутствие пре-мРНК в элюате детектировалось относительно небольшое количество мяРНК (дорожка 2). Напротив, элюат пре-мРНК, меченной аптамером (дорожка 1), содержал аналогичные количества пре-мРНК, мяРНК U1 и U2 и только низкий уровень мяРНК U4, U5 и U6.Этот результат показал, что преимущественно сплайсинговый комплекс A (и / или потенциально E) образовался через 45 минут и впоследствии был элюирован тобрамицином. Соответственно, элюат от селекции по аффинности, выполненной с пре-мРНК, показал сложный, но отчетливый белковый паттерн, тогда как только несколько белков, мигрирующих в диапазоне от 55 до 65 кДа, наблюдались без пре-мРНК (рис. 2 A , сравните дорожки 3 и 4). Рис 2. ( A ) РНК и содержание белка в комплексах сплайсинга, элюированных тобрамицином.Твердофазный сплайсинг выполняли в течение 45 минут при 30 ° C в присутствии (дорожки 1 и 3) или в отсутствие (дорожки 2 и 4) пре-мРНК, а затем комплексы элюировали тобрамицином. РНК и белок были выделены, проанализированы с помощью PAGE и визуализированы окрашиванием серебром. ( B ) Анализ градиента глицерина аффинно-очищенных сплайсосомных комплексов. Голая пре-мРНК (•) или комплексы, образованные после сплайсинга в течение 15 минут при 4 ° C (○) или в течение 45 минут при 30 ° C (), подвергали центрифугированию в градиенте 10–30% глицерина, и процентное содержание 32 Р-меченую пре-мРНК в каждой фракции определяли методом Черенкова.( C ) Профиль РНК фракционированных в градиенте глицерина, 45-минутных сплайсинговых комплексов. РНК выделяли из фракций с нечетными номерами и анализировали, как в A . PRE, пре-мРНК; *, загрязняющие РНК. Для более подробной характеристики аффинно-очищенных сплайсинговых комплексов элюат подвергали центрифугированию в градиенте глицерина (рис. 2 B ), и седиментационное поведение комплексов, содержащих 32 P-меченую пре-мРНК, сравнивали с таковой. голой пре-мРНК или аффинно-отобранных H-комплексов (т.е.е. комплексы, образовавшиеся после 15-минутной инкубации с ядерным экстрактом при 4 ° C). Подавляющее большинство комплексов в 45-минутном элюате достигли пика во фракции 11 с величиной седиментации ≈22S; также наблюдались небольшие пики, осаждающиеся при ≈28–42S. Напротив, голые пре-мРНК и H комплексы осаждались при ≈12 и ≈13S соответственно. Таким образом, 45-минутный элюат, по-видимому, состоит из относительно однородной популяции сплайсинговых комплексов, которые показывают значение S значительно выше, чем комплекс H, что согласуется с идеей, что преимущественно были изолированы комплексы A или E.Действительно, анализ состава РНК фракций градиента из 45-минутного элюата выявил эквимолярные количества пре-мРНК и мяРНК U1 и U2 во фракциях 9–13 (рис. 2 C ). Эти фракции почти не содержали мяРНК U4, U5 и U6, которые, по большей части, были обнаружены либо в более медленных, либо в быстро мигрирующих комплексах; последние, которые содержат эквимолярные количества пре-мРНК и всех основных сплайсосомных мяРНК (дорожки 17–23), вероятно, соответствуют сплайсосомному комплексу B.Таким образом, центрифугирование в градиенте глицерина позволяет эффективно отделить H- и B-комплексы от выбранных по аффинности A / E-комплексов. Комплексы E и A содержат U1 и U2 snRNP, но взаимодействие спаривания оснований U2 / пре-мРНК сначала наблюдается в комплексе A (3). Таким образом, перекрестное сшивание РНК-РНК (12) было выполнено для точной идентификации комплексов, мигрирующих в области 22S градиента.Фракции градиента подвергали УФ-облучению в присутствии псоралена, и РНК анализировали методом Нозерн-блоттинга, последовательно исследуя пре-мРНК, U1 и U2 мяРНК (фиг. 3 A — C ). Множественные сшивки, содержащие мяРНК U1, которые достигли пика во фракции 10, наблюдались во фракциях 8–14 (рис. 3 C ). Каждую из этих полос также детектировали с помощью зонда, специфичного для пре-мРНК, подтверждая, что они представляют собой поперечные связи между субстратом сплайсинга и U1 (рис.3 А ). Напротив, сшивка U2 / пре-мРНК была впервые обнаружена во фракции 12, и ее интенсивность постепенно уменьшалась от фракции 14 до 22 (фиг. 3 B ). В независимом эксперименте сшивки U2 / пре-мРНК также наблюдались во фракции 11, но не в предыдущих фракциях (не показаны). Таким образом, фракции 11–14 содержат преимущественно сплайсосомальный комплекс A, тогда как фракции 8–10, вероятно, содержат в основном комплекс E. Основная сшивка U2 / U6 (подтвержденная зондированием на U6; не показана), которая указывает на комплекс B, достигла пика во фракциях 16–22.Однако лишь небольшие количества были обнаружены ниже фракции 14, что подтверждает, что фракции 11–13 (пик комплекса А) практически не содержат комплекса B. Рис. 3. Характеристика взаимодействий РНК-РНК в градиентно-фракционированных сплайсосомных комплексах путем сшивания псораленом. Отобранные по аффинности комплексы фракционировали в градиенте глицерина 10-30%, и сшивание выполняли с фракциями градиента с четными номерами (как указано над каждой полосой) или ≈2% нефракционированного элюата (дорожки E).В самой левой полосе E элюат подвергали УФ-облучению в отсутствие псоралена для контроля неспецифического сшивания. РНК переносили на нейлоновую мембрану, которую гибридизовали последовательно с 32 P-меченными зондами, специфичными для пре-мРНК ( A ) мяРНК U2 ( B ) и мяРНК U1 ( C ). Идентичность сшитых видов указана справа. АМТ, псорален; PRE, пре-мРНК; intPRE, внутренне сшитая пре-мРНК. Белковый состав различных фракций градиента анализировали с помощью SDS / PAGE (фиг. 4 A ). Наблюдали дифференциальное распределение белков по градиенту, при этом белки U2 и U5 с высокой молекулярной массой достигают пика во фракциях 9-13 и 17-23 соответственно. Кроме того, отчетливый белковый паттерн был обнаружен в самых верхних фракциях, в которых по существу отсутствует пре-мРНК, что позволяет предположить, что некоторые белки (или мяРНП; см. Также рис. 2 C для определения состава РНК) диссоциировали во время элюирования и / или градиентного центрифугирования. .Белки, присутствующие во фракциях 11-13 (пик комплекса А), объединяли, фракционировали с помощью SDS / PAGE и анализировали с помощью MALDI-MS или, дополнительно, с помощью LC-MSMS. Рис 4. Содержание белка в фракционированных в градиенте глицерина, 45-минутных комплексах сплайсинга ( A ) или в объединенных фракциях градиента 11–13 ( B ). Белок выделяли из нечетных или объединенных фракций и анализировали, как показано на рис. 2. Было идентифицировано более 70 белков, включая почти все белки, которые, как известно, присутствуют в мяРНП 12S U1 и 17S U2 (рис.4 B и таблица 1). Напротив, не было обнаружено ни U4 / U6, а только высокомолекулярные белки U5 и / или три-мяРНП, что подтверждает отсутствие больших количеств три-мяРНП 25S U4 / U6.U5. В соответствии с их известной ролью во время образования комплекса E / A было идентифицировано большинство членов семейства белков SR и основные факторы U2AF65 и U2AF35. Также было идентифицировано большое количество белков hnRNP и обоих белков, связывающих m7G-cap (CBP20 и CBP80). Помимо 17S U2 snRNP-ассоциированных белков hPrp5 и hPrp43, были обнаружены еще два члена DExH / D-box семейства РНК-уиндаз / РНПаз, а именно РНК-геликаза A (DDX9) и p68.Были идентифицированы несколько белков, участвующих в сплайсинге пре-мРНК, которые, по-видимому, выполняют множество ролей в клетке, таких как Y-бокс-связывающий белок 1 (YB-1), TLS / FUS и NFAR-2 (15-19), а также факторы экспорта мРНК Aly и HuR (20–22). Интересно, что аффинно очищенные пресплицеосомы также содержали ряд белков, которые, как ранее не было известно, участвуют в сплайсинге (Таблица 1). Таблица 1. Белки, ассоциированные с пресплицеосомами Используя процедуру отбора по аффинности тобрамицина с последующим центрифугированием в градиенте глицерина, можно было очистить пресплицеосомы человека (т.е.е., комплекс) в нативных условиях. Несколько линий доказательств подтверждают вывод о том, что комплексы, охарактеризованные с помощью MS, представляют собой пре-сплицеосомы в отличие от комплексов B / C: ( i ) фракции элюата тобрамицина и анализируемые методом MS градиентные фракции содержали почти эквимолярные количества пре-мРНК, U1 и U2. мяРНК и только минимальные количества мяРНК U5 или U4 / U6; ( ii ) большинство белков, идентифицированных с помощью MS, были белками snRNP U1 и U2 или известными факторами пресплицеосомного сплайсинга; и ( iii ) сшивки РНК-РНК, характерные для комплекса B / C, не наблюдались.Хотя несколько высокомолекулярных белков три-мяРНП присутствовали в небольших количествах (о чем свидетельствует окрашивание белкового геля серебром), никакие факторы сплайсинга, о которых известно, что они связываются первыми во время или после образования В-комплекса, не были обнаружены с помощью МС (таблица 1). Белки Tri-snRNP могут присутствовать из-за низкого уровня загрязнения комплекса B / C (или его продукта диссоциации) или они могут быть добросовестными пресплицеосомными компонентами. Действительно, недавние исследования показывают, что комплекс три-мяРНП U4 / U6.U5 взаимодействует с пре-мРНК уже на ранних стадиях образования комплекса сплайсинга (23). Комплексы E и A содержат U1 и U2 snRNP, и, таким образом, трудно различить их, основываясь только на составе белков snRNA и snRNP. Однако два наблюдения подтверждают вывод о том, что комплексы, проанализированные с помощью MS, являются преимущественно комплексами A: ( i ) комплексный белок E SF1 / mBBP не был обнаружен, и ( ii ) сшивание псоралена идентифицировало U1 / пре-мРНК и U2. Взаимодействия спаривания оснований / пре-мРНК во фракциях, проанализированных с помощью МС.Поскольку U2 snRNP первые пары оснований с пре-мРНК в комплексе A (3), эти данные являются лучшим доказательством того, что комплексы A, в отличие от комплексов E, были проанализированы. Однако из-за изначально низкой эффективности сшивания псораленом мы не можем исключить, что градиентные фракции 11-13 содержат комплексы E в дополнение к A. Градиентно-фракционированные, аффинно-очищенные сплайсинговые комплексы не выдерживают нативный PAGE, и, таким образом, этот метод не может быть использован для ответа на этот вопрос. Известно, что большинство белков, идентифицированных в наших аффинно очищенных пресплицеосомах, участвуют в сплайсинге пре-мРНК, что свидетельствует о высокой чистоте этих комплексов. Более того, были обнаружены почти все белки, которые, как ожидается, должны присутствовать в пресплицеосомах млекопитающих, что позволяет предположить, что перечень пресплицеосомных белков, представленный в таблице 1, относительно полный. Интересно, что большинство недавно идентифицированных белков snRNP 17S U2, таких как hPrp5, hPrp43, SPF45, SF3b14 и SF3b10, присутствовали в очищенных пре-сплицеосомах, что согласуется с идеей о том, что они потенциально способствуют ассоциации U2 с пре-мРНК (24 ).Судя по совокупной общей массе идентифицированных белков, маловероятно, что все они обнаруживаются в каждой пресплицеосоме. Действительно, некоторые белки явно присутствуют в субстехиометрических количествах, что указывает на их присутствие в субпопуляции аффинно-очищенных комплексов. Поскольку были обнаружены несколько членов семейств белков SR и hnRNP, также вероятно, что проанализированные нами комплексы могут состоять из смеси пресплицеосом, содержащих различные подмножества этих белков. Помимо белков SR, hnRNP и snRNP, было идентифицировано ≈20 дополнительных полипептидов. Каждый из недавно идентифицированных пресплицеосомных белков в силу их совместной очистки с этими комплексами потенциально играет роль в сплайсинге. Среди идентифицированных были несколько многофункциональных белков, включая YB-1 и TLS / FUS, что согласуется с идеей, что они являются общими факторами сплайсинга и могут действовать на ранней стадии процесса сплайсинга. Действительно, многочисленные исследования показали участие TLS / FUS в сплайсинге пре-мРНК (17, 25–27), а YB-1, как известно, играет роль в альтернативном сплайсинге (15).В дополнение к hPrp5 и hPrp43 были также идентифицированы белки DExH-бокса, p68 и РНК-геликаза A (DDX9), что позволяет предположить, что они могут вносить вклад в динамику образования сплайсосом. Соответственно, недавние данные подтверждают, что p68 дестабилизирует взаимодействие U1 / 5 ‘сайта сплайсинга во время сборки сплайсосом (28). MS выявила несколько белков, совместно фракционирующих со сплайсинговым комплексом A, точная функция которого неизвестна. Три из них (CC1.3, F23858_1 и RBM 5) содержат домены (т.е.e., SURP, SR), которые очень характерны для факторов сплайсинга пре-мРНК, что делает их хорошими кандидатами для ранее не охарактеризованных, истинных пре-сплицеосомных белков. Однако необходимы дополнительные эксперименты, чтобы выяснить, действительно ли какие-либо из них участвуют в сплайсинге пре-мРНК. Хотя большинство белков, идентифицированных с помощью MS, скорее всего, являются настоящими факторами сплайсинга, некоторые (например, белки hnRNP) могут связывать фланкирующие области пре-мРНК или внутренних интронных последовательностей, которые не участвуют в процессе сплайсинга.Будущие исследования усеченных пре-мРНК MINX должны выявить, связывают ли эти и потенциально другие белки субстрат, но не участвуют ли они напрямую в сплайсинге. В настоящее время также не ясно, какие из недавно идентифицированных пре-сплицеосомных белков являются основными компонентами (т.е. обычно присутствуют в пре-сплицеосомах) и которые могут специфически связываться с пре-мРНК MINX, используемой в наших исследованиях. Наши данные предоставляют первую подробную инвентаризацию компонентов сплайсосомы на ранних стадиях ее сборки.Характеристика комплексов, образующихся на более поздних стадиях, должна позволить четко определить порядок и динамику ассоциации белков со сплайсосомой во время ее сборки, каталитической активности и последующей разборки. Аналогичным образом, описанный здесь метод аффинной очистки потенциально может быть использован для идентификации субстрат-специфических факторов сплайсинга, таких как те, которые участвуют в альтернативных событиях сплайсинга. Наконец, способность изолировать пресплицеосомы, а также потенциально др. Сплайсосомные комплексы в естественных условиях, является ключевым шагом к выяснению их молекулярной архитектуры.Ультраструктурные исследования, например электронная микроскопия, проводимые с нативными сплайсосомными комплексами на определенных стадиях реакции сплайсинга, могут в конечном итоге выявить структурную динамику аппарата сплайсинга пре-мРНК. Мы благодарим Ирен Экснер и Монику Раабе за отличную техническую помощь. Работа выполнена при поддержке Deutsche Forschungsgemeinschaft Grant Lu 294 / 12-1 и Bundesministerium für Bildung und Forschung (Германия), грант 031U215B (R.Л.). ↵ † K.H., H.U. и H.-P.V. внес равный вклад в эту работу. ↵ ‡ Текущий адрес: Ribopharma AG, Fritz-Hornschuch-Strasse 9, D-95326 Kulmbach, Германия. ↵¶ Кому следует обращаться по адресу: Департамент клеточной биохимии, Институт биофизической химии Макса Планка, Am Fassberg 11, D-37077 Göttingen, Германия.Электронная почта: reinhard.luehrmann {at} mpi-bpc.mpg.de. Этот документ был отправлен напрямую (Трек II) в офис PNAS. snRNP, малый ядерный рибонуклеопротеин hnRNP, гетерогенный ядерный рибонуклеопротеин SR, с высоким содержанием серина / аргинина . S3 Таблица. CED, специфичные для остатков, связаны с уникальными клеточными структурами и процессами.
S1 Рис. Максимальный местный аминокислотный состав соответствует остаточно-зависимым различиям в эффективности синтеза белка.
S2 Рис. Распределение протеома
S3 Рис. Связи между локальным композиционным обогащением и nTE сохраняются в отсутствие белков со статистически смещенными доменами.
S4 Рис. Связь между локальным композиционным обогащением и обилием белка сохраняется в отсутствие белков со статистически смещенными доменами.
S5 Рис. Связь между локальным композиционным обогащением и периодом полужизни белка сохраняется в отсутствие белков со статистически смещенными доменами.
S6 Рис. Связь между локальным композиционным обогащением и nTE сохраняется в отсутствие ЖК-содержащих белков.
S7 Рис. Связь между локальным композиционным обогащением и обилием белка сохраняется в отсутствие ЖК-содержащих белков.
S8 Рис. Связь между локальным композиционным обогащением и периодом полужизни белка сохраняется в отсутствие ЖК-содержащих белков.
S9 Рис. Связи между локальным композиционным обогащением и nTE сохраняются в отсутствие белков с гомополимерными повторами.
S10 Рис. Связь между локальным обогащением состава и обилием белка сохраняется в отсутствие белков с гомополимерными повторами.
S11 Рис. Связь между локальным композиционным обогащением и периодом полужизни белка сохраняется в отсутствие белков с гомополимерными повторами.
S12 Рис. Индивидуальные профили аминокислотного состава для субклеточных компартментов, преимущественно связанные с обогащением полярными и заряженными остатками.
S13 Рис. Индивидуальные профили аминокислотного состава для субклеточных компартментов, преимущественно связанные с обогащением гидрофобных и ароматических остатков.
S14 Рис. Индивидуальные профили аминокислотного состава для субклеточных компартментов, преимущественно связанные с обогащением как полярных, так и гидрофобных / ароматических остатков.
Влияние 8-недельного приема добавок сывороточного или рисового протеина на композицию тела и работоспособность | Nutrition Journal
План эксперимента
Субъекты
Протокол фазы 1 тренировки с отягощениями
Протокол тренировки с отягощениями
Тестирование силы, мощности, состава тела и гипертрофии скелетных мышц
Добавки и контроль диеты
Аминокислотный профиль учебных материалов
Статистика
Белковый состав пресплицеосом человека, выделенных методом аффинной селекции по тобрамицину
Реферат
Методы
Плазмиды, олигонуклеотиды и получение РНК.
Твердофазная реакция сплайсинга и очистка сплайсосом.
Центрифугирование с градиентом глицерина.
Сшивание псораленом и Нозерн-блоттинг.
MS.
Результаты
Отбор сродства к тобрамицину нативных сплайсосомных комплексов.
Очистка сплайсосомального комплекса А.
Идентификация комплексов E и A посредством сшивания РНК-РНК.
Белковый состав пресплицеосом.
Обсуждение
Благодарности
Сноски
Сокращения