Ученые выяснили, как синтез белка влияет на старение
Биологи из Московского государственного университета совместно с коллегами из Гарвардской школы медицины изучили, как меняется с возрастом биосинтез белка в органах мышей. Это дает понимание одного из фундаментальных механизмов процесса старения. Результаты исследования опубликованы в журнале Proceedings of the National Academy of Sciences.
В основе старения лежат повреждения, которые со временем накапливаются в клетках, тканях и органах живых организмов. Биосинтез белка — важнейший метаболический процесс, на который клетка тратит большую часть вырабатываемой энергии. Его нарушения вносят большой вклад в старение организма в целом.
Известно, что некоторые воздействия, снижающие эффективность белкового синтеза, увеличивают продолжительность жизни животных. Например, при отсутствии аминокислот — когда животное голодает — или в условиях, способствующих денатурации белков — при перегреве или интоксикации — уровень синтеза белка падает, и эта адаптация позволяет организму выжить в неблагоприятных условиях.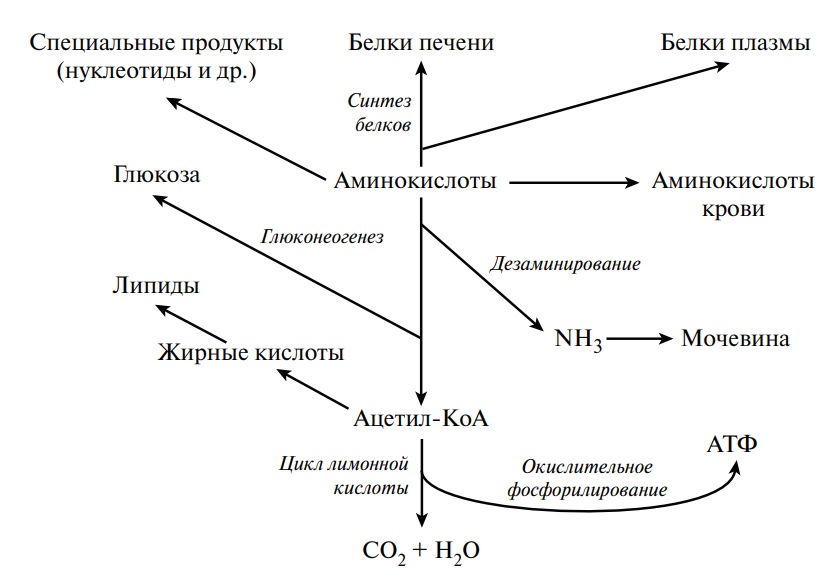
Эффект замедления синтеза белка, продлевающий жизнь, возникает в организмах некоторых животных при низкокалорийной диете или применении лекарственных средств, снижающих активность одного из главных регуляторов белкового синтеза — протеинкиназы mTOR.
Авторы выяснили, что с возрастом в печени и почке мышей происходит нечто похожее: стареющие клетки, как бы чувствуя накопление повреждений, снижают уровень белкового синтеза, чтобы отсрочить наступление неблагоприятных последствий. Задачей ученых было понять механизм этого процесса.
Исследованием изменений, происходящих с возрастом в различных организмах — от дрожжей до человека — ученые из НИИ Физико-химической биологии имени А.Н. Белозерского МГУ занимаются с 2017 года. Главным проектом лаборатории стало изучение картины биосинтеза белка в органах мышей разного возраста методом рибосомного профайлинга.
«Этот ультрасовременный метод появился относительно недавно и совершил настоящую революцию, привнеся все преимущества системной биологии в область изучения биосинтеза белка, — приводятся в пресс-релизе МГУ слова Сергея Дмитриева, одного из авторов исследования, старшего научного сотрудника НИИ ФХБ МГУ и ИМБ РАН.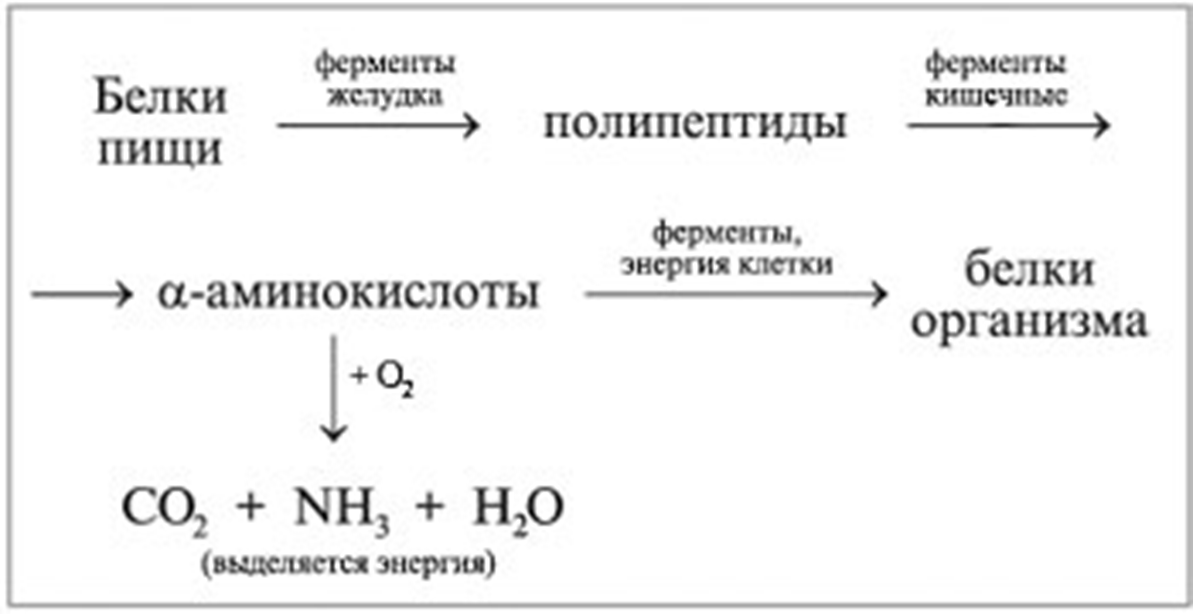 — В клетке белок синтезируют специальные молекулярные машины – рибосомы. Они «переводят» (транслируют) последовательности генов с языка азотистых оснований, на котором записана информация в ДНК и РНК, на язык аминокислот, из которых и состоят белки. Метод рибосомного профайлинга позволяет определить одновременно все РНК, которые в данный конкретный момент транслируются рибосомами в клетке. Для этого используется секвенирование нового поколения и сложная биоинформатическая обработка данных».
— В клетке белок синтезируют специальные молекулярные машины – рибосомы. Они «переводят» (транслируют) последовательности генов с языка азотистых оснований, на котором записана информация в ДНК и РНК, на язык аминокислот, из которых и состоят белки. Метод рибосомного профайлинга позволяет определить одновременно все РНК, которые в данный конкретный момент транслируются рибосомами в клетке. Для этого используется секвенирование нового поколения и сложная биоинформатическая обработка данных».
Как биохимическую, так и «компьютерную» части исследования выполняли в МГУ с учетом опыта американских коллег.
Выпускница факультета биоинженерии и биоинформатики МГУ Александра Анисимова, первый автор статьи, рассказывает: «Благодаря уникальному опыту, накопленному в лаборатории Вадима Гладышева в Бостоне, мы смогли применить метод рибосомного профайлинга к двум органам — печени и почке — мышей разного возраста. Это позволило детально охарактеризовать изменения в экспрессии генов на уровне биосинтеза белка при старении. Мы обнаружили изменения в синтезе компонентов многих важных процессов и регуляторных путей, в том числе связанных с иммунитетом, воспалением, внеклеточным матриксом и метаболизмом жиров. Но самое яркое наблюдение, которое мы сделали, касается снижения темпов наработки самих компонентов белок-синтезирующего аппарата — рибосомных белков и трансляционных факторов».
Мы обнаружили изменения в синтезе компонентов многих важных процессов и регуляторных путей, в том числе связанных с иммунитетом, воспалением, внеклеточным матриксом и метаболизмом жиров. Но самое яркое наблюдение, которое мы сделали, касается снижения темпов наработки самих компонентов белок-синтезирующего аппарата — рибосомных белков и трансляционных факторов».
«Полученные результаты помогают понять фундаментальные механизмы, лежащие в основе процессов старения. Одновременно мы узнаем много нового о молекулярных механизмах биосинтеза белка», — отмечает один из руководителей работы, профессор Гарвардской школы медицины Вадим Гладышев.
В настоящее время ученые продолжает исследования с целью выяснения, какие именно воздействия на организм мышей приводят к увеличению продолжительности жизни.
Белки синтез в организме — Справочник химика 21
Незаменимые аминокислоты [13 — 16]. Растения и некоторые микроорганизмы могут производить все аминокислоты, нужные им для синтеза клеточных белков.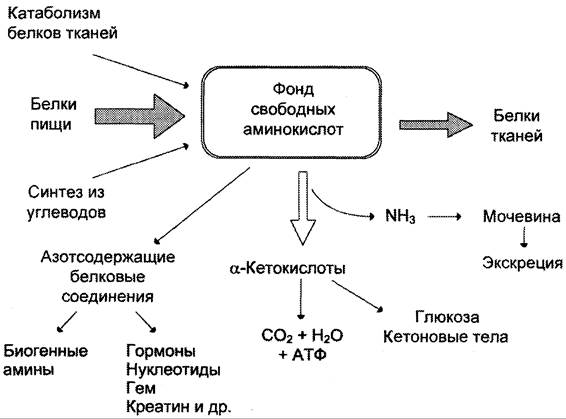 Животные организмы способны синтезировать только 10 протеиногенных аминокислот. Остальные 10 ие могут быть получены с помощью биосинтеза и должны постоянно поступать в организм в виде пищевых белков. Отсутствие их в организме ведет к угрожающим жизни явлениям (задержка роста, отрицательный азотный баланс, расстройство биосинтеза белков и т. д.). Розе и сотр. [17] предложили для этих аминокислот название незаменимые аминокислоты (НАК). В табл. 1-2 приведены незаменимые для организма человека аминокислоты и минимальная суточная потребность в них. [c.18]
Животные организмы способны синтезировать только 10 протеиногенных аминокислот. Остальные 10 ие могут быть получены с помощью биосинтеза и должны постоянно поступать в организм в виде пищевых белков. Отсутствие их в организме ведет к угрожающим жизни явлениям (задержка роста, отрицательный азотный баланс, расстройство биосинтеза белков и т. д.). Розе и сотр. [17] предложили для этих аминокислот название незаменимые аминокислоты (НАК). В табл. 1-2 приведены незаменимые для организма человека аминокислоты и минимальная суточная потребность в них. [c.18]
Синтез белка подчиняется закону все или ничего и осуществляется при условии наличия в клетке полного набора всех 20 аминокислот. Даже при поступлении всех аминокислот с пищей организм может испытывать состояние белковой недостаточности, если всасывание какой-либо одной аминокислоты в кишечнике замедлено или если она разрушается в большей степени, чем в норме, под действием кишечной микрофлоры.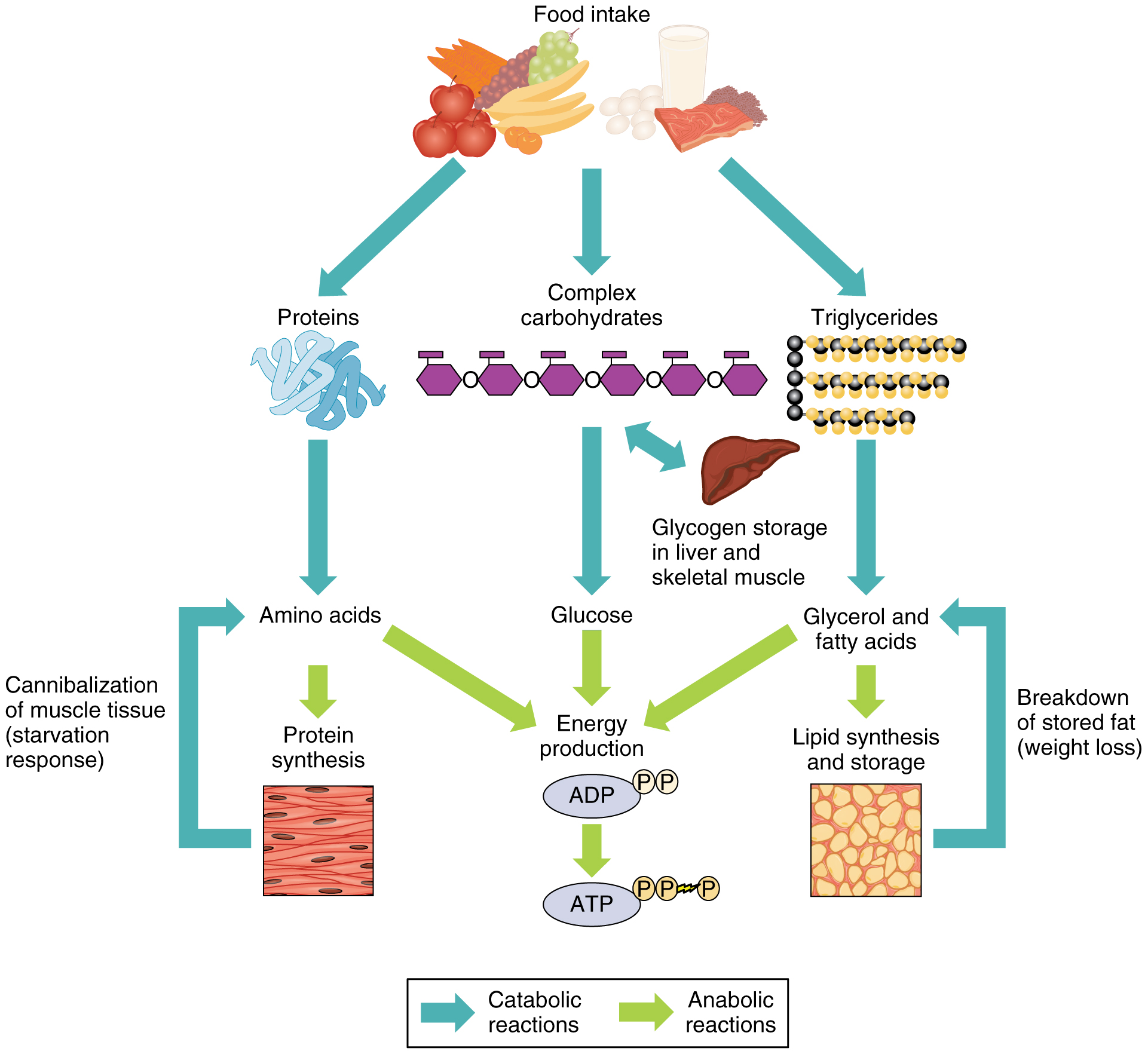 В этих случаях будет происходить ограниченный синтез белка или организм будет компенсировать недостаток аминокислоты для биосинтеза белка за счет распада собственных белков. Степень усвоения белков и аминокислот пищи зависит также от количественного и качественного состава углеводов и липидов, которые резко сокращают энергетические потребности организма за счет белков. Экспериментальный и клинический материал свидетельствует, что диета с недостаточным содержанием жиров и низкокалорийная пища способствуют повышению экскреции аминокислот и продуктов их распада с мочой. [c.412]
В этих случаях будет происходить ограниченный синтез белка или организм будет компенсировать недостаток аминокислоты для биосинтеза белка за счет распада собственных белков. Степень усвоения белков и аминокислот пищи зависит также от количественного и качественного состава углеводов и липидов, которые резко сокращают энергетические потребности организма за счет белков. Экспериментальный и клинический материал свидетельствует, что диета с недостаточным содержанием жиров и низкокалорийная пища способствуют повышению экскреции аминокислот и продуктов их распада с мочой. [c.412]
Синтез белка в организме [c.451]
Буквенные коды ДНК, которыми являются сочетания АТ и ГЦ, а также буквенные коды РНК — АУ и ГЦ — могут быть связаны в слова и предложения . В молекуле ДНК, управляющей синтезом лишь одного из белков в организме человека, содержится такое количество подобных слов , что из них составляется предложение , занимающее объем полномерной книги (150000 слов).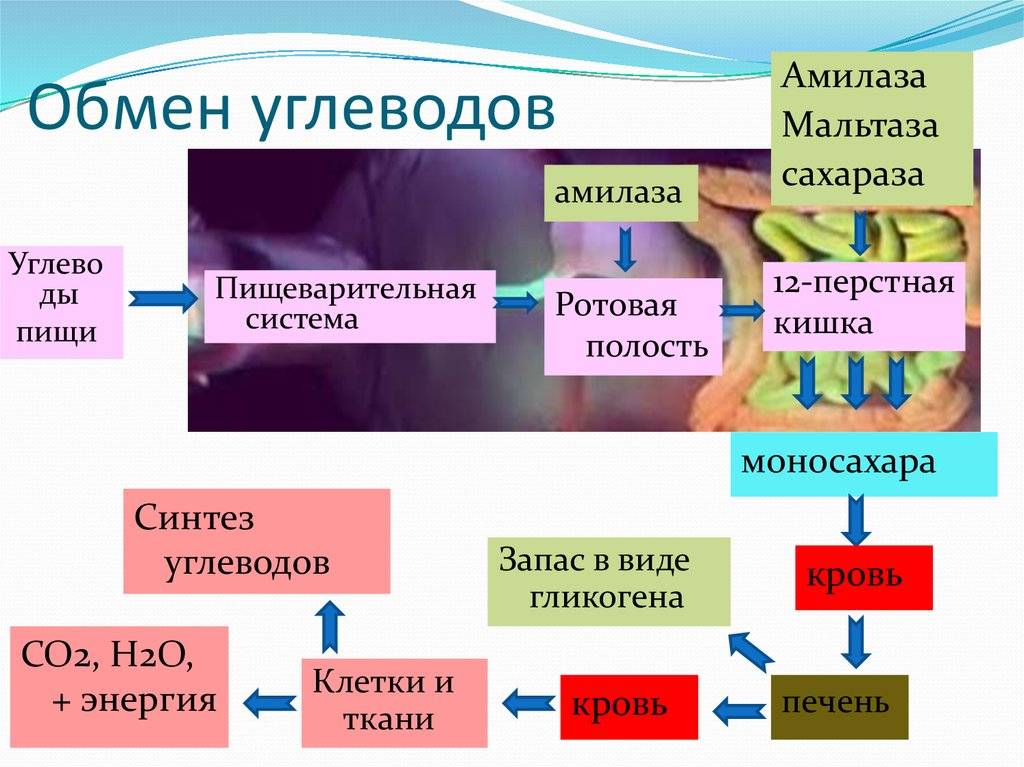 У низших организмов предложения , описывающие синтез белков, как правило, гораздо короче, поскольку их белки имеют меньщие размеры и проще по своему составу. Для построения одной клетки человеческого тела необходима информация, эквивалентная содержащейся в читальном зале библиотеки на 20000 книг. Такой гигантский объем информации требуется для синтеза каждого из многочисленных белков человеческого организма. Поскольку белки печени совершенно не похожи, скажем, на белки волос, для хранения всех книг, полностью описывающих [c.486]
У низших организмов предложения , описывающие синтез белков, как правило, гораздо короче, поскольку их белки имеют меньщие размеры и проще по своему составу. Для построения одной клетки человеческого тела необходима информация, эквивалентная содержащейся в читальном зале библиотеки на 20000 книг. Такой гигантский объем информации требуется для синтеза каждого из многочисленных белков человеческого организма. Поскольку белки печени совершенно не похожи, скажем, на белки волос, для хранения всех книг, полностью описывающих [c.486]
Синтез инсулина — замечательное достижение науки. Чтобы осуществить его, потребовалось последовательно провести 223 реакции. Удалось соединить в точно определенном порядке все остатки а-аминокислот, образующих молекулу инсулина (а их 51 ). Работа продолжалась три года. Таким образом, подтвердилась правильность материалистических представлений о принципиальной возможности синтеза белков вне организма. И несомненно, что с развитием науки будут осуществлены синтезы еще более сложных белковых веществ.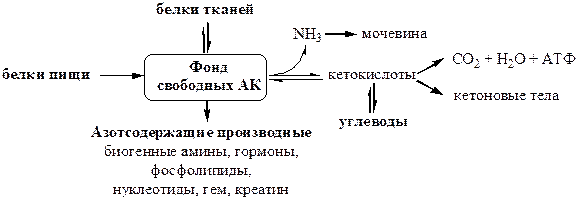 [c.294]
[c.294]
Мы ограничимся изложением известных в настоящее время данных о структуре и биологической функции наиболее важных соединений — белков, нуклеиновых кислот, жиров и углеводов, а также сообщим некоторые сведения о путях синтеза белка в организме. [c.435]
Аминокислоты пищевых белков потребляются организмом в первую очередь для построения белков, необходимых организму для роста, возобновления тканей и синтеза ферментов и гормонов. Избыток аминокислот, введенный с пищей, дезаминируется, причем образующийся аммиак удаляется в виде мочевины или мочевой кислоты, а органический остаток превращается в углеводы или жиры, т.е. в горючее , которое служит источником энергии. (Нормальный животный организм не откладывает запасов белков, подобно тому как он откладывает гликоген или жиры.) [c.387]
Белки поставляют организму вещества, необходимые для роста и восстановления тканей, а также для синтеза ферментов и некоторых гормонов (см.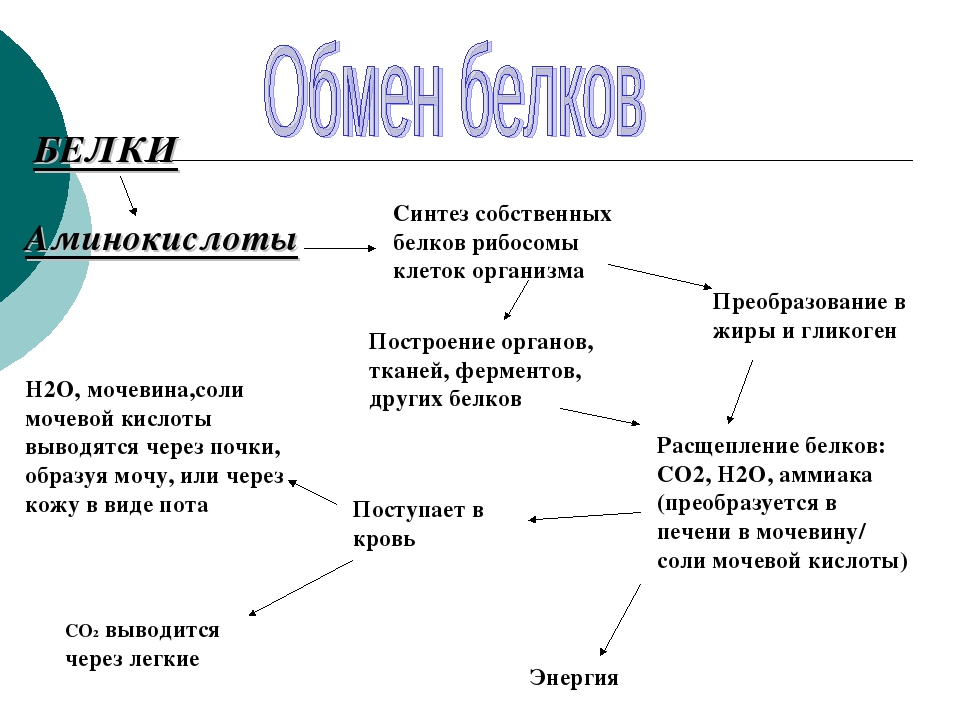 разд. 28.7). Питательная ценность бел- [c.486]
разд. 28.7). Питательная ценность бел- [c.486]
Алании и глутамин в крови. В плазме крови содержатся все аминокислоты, необходимые для синтеза белков в организме, но в разных количествах. При этом концентрации двух аминокислот, а именно аланина и глутамина намного выше, чем остальных. Объясните возможные причины высокого содержания этих двух аминокислот. [c.777]
Нуклеотиды и полинуклеотиды. Синтез белка в организме Ферменты [c.8]
Синтез и расщепление белков, в организмах растений, животных и микроорганизмов происходит с помощью ферментов. Каждой аминокислоте соответствует свой фермент, который привязывает к растущей молекуле пептида или белка только одну конкретную аминокислоту. [c.723]
Мы все время обсуждаем вопросы, относящиеся к структуре белков. Наряду со структурой необходимы точные ответы на три вопроса сколько, когда и где Сколько производится данного белка в организме, на какой стадии онтогенетического развития, в каких клетках и тканях Иными словами, определяющее значение имеет регуляция синтеза белков, о которой шла речь в 8. 8, Мутации регуляторных генов, мутации, нарушающие ди- [c.560]
8, Мутации регуляторных генов, мутации, нарушающие ди- [c.560]
Белки, попадающие в организм в качестве продуктов питания, подвергаются гидролизу. Как уже отмечалось, они легко гидролизуются в кислой среде с образованием отдельных аминокислот. Расщепление белков в организме начинается в желудке под действием фермента пепсина и соляной кислоты. При этом белки превращаются в смеси различных полипептидов. Гидролиз в желудке — лишь одна из стадий переработки белков. Смесь пептидов поступает из желудка в двенадцатиперстную кишку (верхний отдел кишечника), а затем — в тонкий кишечник, где под действием специальных ферментов — пеп-сидаз — завершается гидролиз полипептидов до свободных аминокислот. Образовавшиеся таким образом аминокислоты всасываются из тонкого кишечника в кровеносную систему, чтобы принять участие в синтезе именно тех белков, которые в данный период развития необходимы живому организму. [c.523]
Процесс усвоения белков животными организмами заключается первоначально в распаде гигантской молекулы белка на составляющие ее звенья — аминокислоты, а затем в синтезе из аминокислот таких белков, которые свойственны данному организму.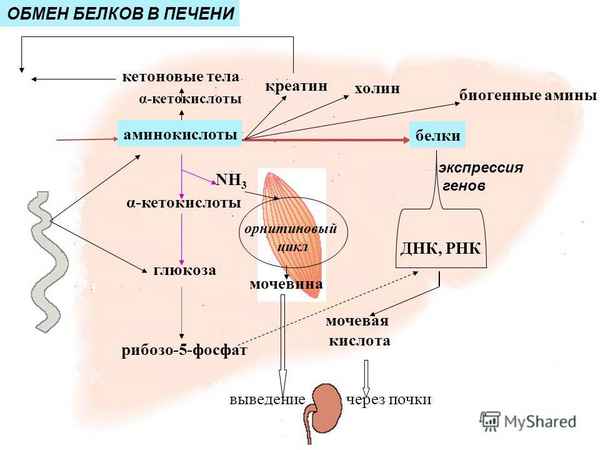 Одна из важнейших проблем естествознания, заключающаяся в искусственном получении белковых веществ, вероятно, близка к своему разрешению. [c.309]
Одна из важнейших проблем естествознания, заключающаяся в искусственном получении белковых веществ, вероятно, близка к своему разрешению. [c.309]
Ввиду того что антитела представляют собой типичные белки, их образование непосредственно связано с синтезом белков в организме наши знания относительно этого процесса пока еще крайне ограничены. [c.449]
Для синтеза аминокислот автотрофные организмы используют азот неорганических соединений (аммонийных солей и нитратов). Гетеротрофные организмы не способны к синтезу части аминокислот, необходимых для образования клеточных белков. Такие организмы для синтеза собственных белков пспользуют аминокислоты, входящие в состав белков пищи. [c.192]
Биосинтез белков — одна из самых важных и интересных проблем современной биохимии. В настоящее время расшифрованы многие процессы, приводящие к синтезу белков в организме. [c.289]
Превращение белков в организме. В организмах животных и человека под влиянием ферментов (пепсина, трипси—на, эрепсина и др.) происходит гидролиз белков. В результате этого образуются аминокислоты, которые всасываются ворсинками кишечника в кровь и используются для образования белков, специфических данному организму. Синтез белков идет с поглощением энергии. Эту энергию доставляют молекулы АТФ. (Повторите из учебника Общая биология 42.) В организме одновременно с синтезом белков непрерывно происходит и полное их разрушение, вначале до аминокислот, а затем до оксида углерода (IV), аммиака, мочевины и воды. При этих процессах выделяется энергия, но Б меньшем количестве, чем при распаде углеводов и жиров. [c.21]
В организмах животных и человека под влиянием ферментов (пепсина, трипси—на, эрепсина и др.) происходит гидролиз белков. В результате этого образуются аминокислоты, которые всасываются ворсинками кишечника в кровь и используются для образования белков, специфических данному организму. Синтез белков идет с поглощением энергии. Эту энергию доставляют молекулы АТФ. (Повторите из учебника Общая биология 42.) В организме одновременно с синтезом белков непрерывно происходит и полное их разрушение, вначале до аминокислот, а затем до оксида углерода (IV), аммиака, мочевины и воды. При этих процессах выделяется энергия, но Б меньшем количестве, чем при распаде углеводов и жиров. [c.21]
АНАБОЛИЧЕСКИЕ ВЕЩЕСТВА (анаболики) (от греч. апаЬо1ё-подъем), лек. синтетич. препараты, стимулирующие синтез белка в организме и кальцификацию костной ткани. Действие А. в. проявляется, в частности, в увеличении массы скелетной мускулатуры. При этом в связи с уси- [c.157]
Сходным образом осуществляется регуляция О.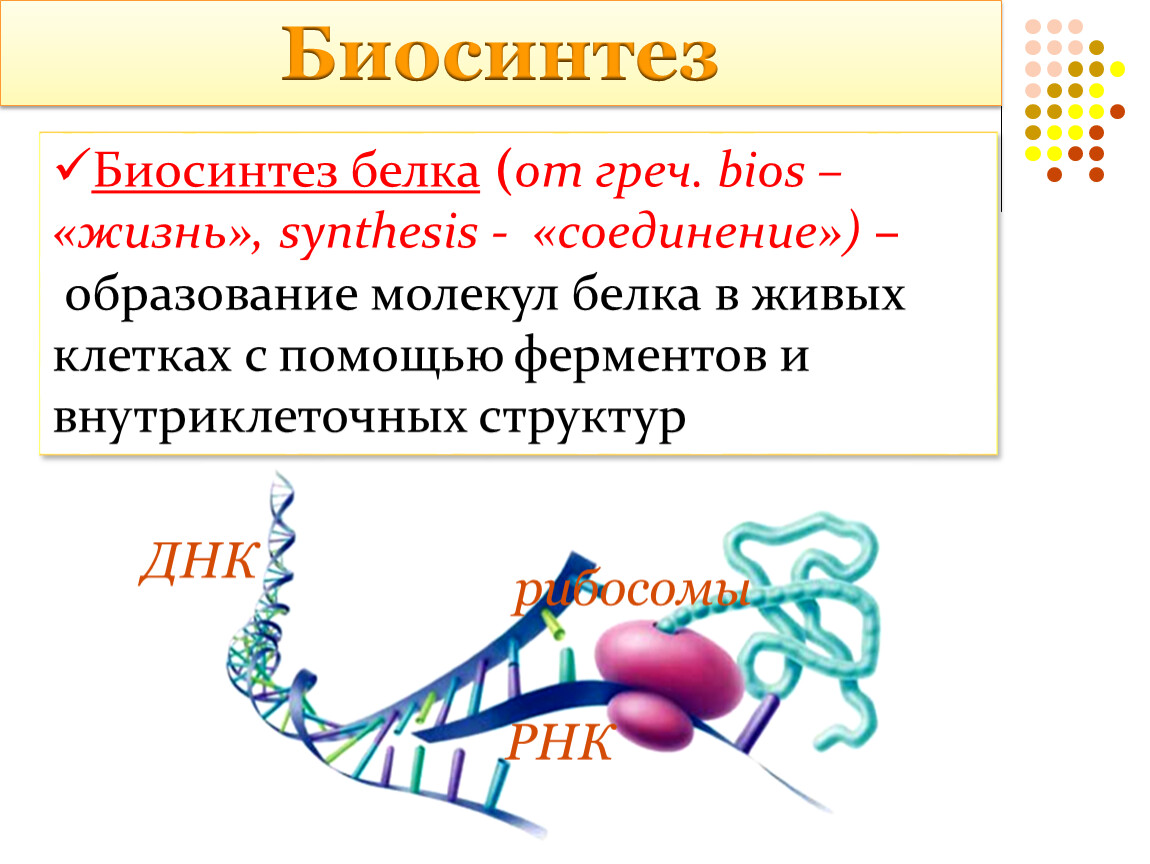 в. на уровне биосинтеза ферментов. При этом субстрат или продукт р-ции регулирует активность белкового репрессора, подавляющего транскрипцию (синтез матричной РНК на ДНК-матрице) соответствующего оперона (участок ДНК, кодирующий одну молекулу матричной РНК под контролем белка-репрессора). Примером регуляции при помощи положит. прямой связи может служить в данном случае управление расщеплением лактозы. Появление в среде лактозы инактивирует у бактерии Es heri hia oli соответствующий репрессор и тем самым разрешает транскрипцию оперона, кодирующего ферменты, катализирующие расщепление лактозы. Пример регуляции при помощи отрицат. обратной связи — управление биосинтезом гистидина. Избыток гистидина активирует репрессор, ингибирующий транскрипцию оперона, кодирующего ферменты биосинтеза гистидина. Если репрессор и белки, синтез к-рых он подавляет, кодируются одним опероном, то отрицат. обратная связь осуществляется без участия внеш. модуляторов активности репрессора. Аналогичным образом осуществляется регуляция биосинтеза белка на уровне трансляции (синтез белка ка РНК-матрице).
в. на уровне биосинтеза ферментов. При этом субстрат или продукт р-ции регулирует активность белкового репрессора, подавляющего транскрипцию (синтез матричной РНК на ДНК-матрице) соответствующего оперона (участок ДНК, кодирующий одну молекулу матричной РНК под контролем белка-репрессора). Примером регуляции при помощи положит. прямой связи может служить в данном случае управление расщеплением лактозы. Появление в среде лактозы инактивирует у бактерии Es heri hia oli соответствующий репрессор и тем самым разрешает транскрипцию оперона, кодирующего ферменты, катализирующие расщепление лактозы. Пример регуляции при помощи отрицат. обратной связи — управление биосинтезом гистидина. Избыток гистидина активирует репрессор, ингибирующий транскрипцию оперона, кодирующего ферменты биосинтеза гистидина. Если репрессор и белки, синтез к-рых он подавляет, кодируются одним опероном, то отрицат. обратная связь осуществляется без участия внеш. модуляторов активности репрессора. Аналогичным образом осуществляется регуляция биосинтеза белка на уровне трансляции (синтез белка ка РНК-матрице).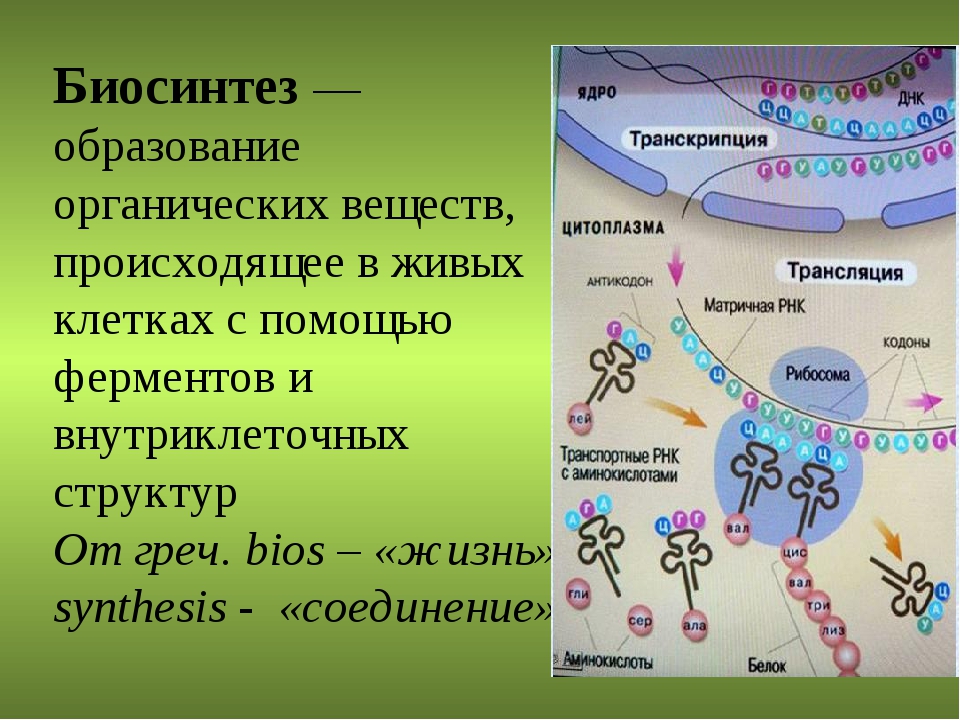 Такой механизм регуляции позволяет синтезировать белок в строгом соответствии с потребностью в нем на данном этапе существования организма. [c.317]
Такой механизм регуляции позволяет синтезировать белок в строгом соответствии с потребностью в нем на данном этапе существования организма. [c.317]
Однако в связи с прогрессирующим ростом населения земного шара и с ограниченностью площади земель, пригодных для земледелия, возникла необходимость получения синтетической и искусственной пищи. Уже давно для пополнения пищи животных соединениями фосфора применяется кормовой преципитат СаНР04, а также карбамид СО ЫН2)2 как один из источников синтеза белка в организме. [c.11]
В отличие от сложных белков, белки одноклеточных организмов (БОО) используются как пищевая добавка. Обогащением белковыми добавками на основе БОО улучшают качество растительного белка. Эти добавки повышают содержание витаминов, микроэлементов, а главное — аминокислот, несинтезируемых многими растениями. Производство пищевых белков измеряется миллионами тонн в год и постоянно растет. Микробиологический синтез белка, продукт которого представляет собой инактивированную массу клеток, — основной [c. 429]
429]
Ачдрогенные гормоны применяются при расстройствах мужской половой сферы, особенно если причиной являются переутомление, при раке молочной железы, при различных заболеваниях сосудистой системы (гипертонии, гипотонии) и т. д. Некоторые стероиды, близкие по строению к андрогенам, обладают так называемой анаболической активностью, т. е. свойством промотировать синтез белка в организме. При этом их андрогенные свойства являются излишними. Сейчас найден ряд соединений с высокой анаболической и малой андрогенной активностью, например, 17а-этилтестостерон и фенилпропионат 19-нортестостерона, кото- [c.322]
Аминокислоты как основные составные части белков участвуют во всех жизненных процессах наряду с нуклеиновыми кислотами, углеводами и липидами. Кроме аминокислот, входящих в состав белков, живые организмы обладают постоянным резервом свободных аминокислот, содержащихся в тканях и в клеточном соке. Они находятся в динамическом равновесии при многочисленных обменных реакциях. Аминокислоты используются в биосинтезе полипептидов и белков, а также в синтезе фосфатидов, порфи-ринов и нуклеотидов. [c.10]
Аминокислоты используются в биосинтезе полипептидов и белков, а также в синтезе фосфатидов, порфи-ринов и нуклеотидов. [c.10]
Модификации различных групп в полипептидной цепи. Если в синтезе белков участвуют 20 аминокислот генетического кода Ниренберга (Nirenberg), то остается еще не менее 140 аминокислот или их производных, идентифицированных в составе белков различных организмов [174]. [c.44]
Внерибосомный механизм синтеза нентидов. Накопленные данные, действительно, свидетельствуют о том, что матричный механизм синтеза лежит в основе биосинтеза почти всех белков живых организмов. Тем не [c.533]
Технология выделения и экспрессии чужеродных генов в Е. соН и в некоторых других микроорганизмах достаточно хорошо отработана, однако не стоит забывать, что синтез гетерологичного белка в организме-хозяине может оказывать на него негативное влияние. Например, сверхпродукция такого белка может привести к истощению метаболических ресурсов хозяйского организма и отрицательно повлиять на его рост. Присутствие гетерологичного белка может оказаться даже губительным для клетки-хозяина. Так, сайты рестрикции имеются во всех молекулах ДНК, и если продуктом клонированного гена является эндонуклеаза рестрикции, то в отсутствие специальных защитных механизмов хозяйская ДНК будет расщепляться ею. [c.247]
Присутствие гетерологичного белка может оказаться даже губительным для клетки-хозяина. Так, сайты рестрикции имеются во всех молекулах ДНК, и если продуктом клонированного гена является эндонуклеаза рестрикции, то в отсутствие специальных защитных механизмов хозяйская ДНК будет расщепляться ею. [c.247]
Как уже говорилось, в ДНК содержится информация, необходимая для синтеза всего набора белков, присущего данному организму. Аминокислотная последовательность в ДНК записана с помощью специального кода. Кодирующим элементом для каждой определенной аминокислоты является тридезоксирибону-клеотидный фрагмент. Общее число таких кодирующих элементов составляет величину, равную 4 = 64,. что превышает число аминокислот, участвующих в биосинтезе белков. Как уже говорилось, все белки живых организмов строятся из 20 аминокислот. Таким образом, некоторые аминокислоты имеют несколько кодирующих элементов — от одного до шести. Соответствие между аминокислотами и кодирующими их трину1 леотидами называют генетически. и кодом. [c.18]
и кодом. [c.18]
НИИ процесса деспирализации получаются уже две абсолютно тождественные исходной и друг другу молекулы ДНК. Аналогично на деснирализующейся молекуле ДНК происходит репликация молекул и-РНК, последовательность нуклеотидов в которой определяет всю информацию о синтезе белков в организме (рис. 93). [c.559]
Строение нуклеиновых кислот. Участие их в синтезе клеточных белков. Синтез белков лежит в основе построения новых клеточных структур. Организмы синтезируют свои собственные гбелки, отличающиеся от белков других видов характером чередования аминокислот. Первичная структура белков определяет многие их биохимические особенности. Изменение чередования аминокислот в молекулах ферментов в некоторых случаях приводит к потере свойств катализатора. Чем же определяется последовательность расположения аминокислот при синтезе белков Для ответа на этот вопрос была выдвинута теория матриц. Согласно этой теории, в клетках имеется нечто подобное типографским матрицам или штампам, каждый из которых штампует белок определенного вида или точнее белок со строго определенным порядком расположения аминокислот в его полипептидной цепи. Роль матриц выполняют нуклеиновые кислоты. Нуклеиновые кислоты имеются во всех без исключения клетках. Различают две группы нуклеиновых кислот—дезоксирибонуклеиновые кислоты (ДНК) и рибонуклеиновые кислоты (РНК). ДНК содержится главным образом в клеточном ядре, РНК — Э ядре и цитоплазме. [c.122]
Роль матриц выполняют нуклеиновые кислоты. Нуклеиновые кислоты имеются во всех без исключения клетках. Различают две группы нуклеиновых кислот—дезоксирибонуклеиновые кислоты (ДНК) и рибонуклеиновые кислоты (РНК). ДНК содержится главным образом в клеточном ядре, РНК — Э ядре и цитоплазме. [c.122]
Установлено, что первичная аминокислота, синтезируемая бактериями, ассимилирующими молекулярный азот и находящимися в узелках на корнях бобовых, является аспарагиновой кислотой. В организме животного аммиак, необходимый для синтеза глутаминовой кислоты, образуется при дезаминировании аминокислот белков самого организма или белков пищи. О том, в каком виде этот аммиак откладывается в организме, будет сказано ниже. [c.390]
Для нормального синтеза белка в организме человека все незаменимые аминокислоты должны быть доступны одновременно. Если крыс кормить синтетической пищей, содержащей все незаме- [c.825]
Как получение химических соединений и пищевых добавок путем брожения, так и синтез антибиотиков всегда велись в асептических условиях, но некоторые современные процессы (например, образование белка одноклеточными организмами) осуществляют в еще более жестком режиме. Обеспечение таких особых условий —многоплановая задача. Она решается инже-нерами-химиками и микробиологами (подробнее об этом будет рассказано в гл. 10). С другой стороны, использование микроорганизмов при переработке отходов (гл. 6) не требует создания стерильных условий напротив, вообще говоря, чем больше разных микроорганизмов принимает в этом участие, тем лучше. Впрочем, при планировании и создании заводов по переработке отходов инженеры-химики и микробиологи столкнулись с проблемами иного круга. Процесс минерализации органических отбросов, основанный на использовании активного ила, был разработан в 1914 г. С тех пор он был существенно модернизирован, стал более сложным и производительным и используется сегодня во всем мире для переработки стоков. [c.13]
Обеспечение таких особых условий —многоплановая задача. Она решается инже-нерами-химиками и микробиологами (подробнее об этом будет рассказано в гл. 10). С другой стороны, использование микроорганизмов при переработке отходов (гл. 6) не требует создания стерильных условий напротив, вообще говоря, чем больше разных микроорганизмов принимает в этом участие, тем лучше. Впрочем, при планировании и создании заводов по переработке отходов инженеры-химики и микробиологи столкнулись с проблемами иного круга. Процесс минерализации органических отбросов, основанный на использовании активного ила, был разработан в 1914 г. С тех пор он был существенно модернизирован, стал более сложным и производительным и используется сегодня во всем мире для переработки стоков. [c.13]
Про биосинтез белка | Нанотехнологии Nanonewsnet
Новые понятия, которые должен усвоить ребенок: ДНК, белок, генная инженерия, нанотехнологии (манипулирование веществом на уровне атомов и молекул)
Итак, как мы вам обещали, следующей достопримечательностью нашего путешествия будет… наш собственный организм! Ведь он тоже состоит из атомов и молекул, а значит, внутри нас также происходят различные нанопроцессы. Наннос очень переживает, что не все читатели смогут оценить по достоинству эти очень важные процессы наномира (потому что иногда даже взрослые их плохо понимают). А ведь именно эта достопримечательность лежит в основе появления на свет всех всех наноботиков. Так что постарайтесь читать эту главу очень внимательно, и если что-то непонятно, не бойтесь задавать вопросы Нанносу или взрослым.
Наннос очень переживает, что не все читатели смогут оценить по достоинству эти очень важные процессы наномира (потому что иногда даже взрослые их плохо понимают). А ведь именно эта достопримечательность лежит в основе появления на свет всех всех наноботиков. Так что постарайтесь читать эту главу очень внимательно, и если что-то непонятно, не бойтесь задавать вопросы Нанносу или взрослым.
Самый главный нанопроцесс в нашем организме – это биосинтез белкá. Первое, что приходит на ум при слове «белок» – это белок куриного яйца (его научное название – альбумин). Но белки спрятаны не только под яичной скорлупой! Наш собственный организм представляет собой целый завод, на котором постоянно трудится огромное число самых разнообразных белков.
Роль белков в организме чрезвычайно разнообразна. Например, белки-гормоны, участвуют в управлении всеми жизненными процессами организма. Без них человек не мог бы ни расти, ни размножаться. Мы обладаем зрением, благодаря особому глазному белку – родопсину. Мы способны двигаться, потому что наши мышцы сокращаются и расслабляются благодаря белкам миозину и актину. Наши волосы и ногти состоят из белка кератина. Белок гемоглобин разносит кислород от легких ко всем клеткам нашего организма. Без белка пепсина, содержащегося в желудочном соке, мы не смогли бы переваривать пищу, а белок интерферон помогает организму справляться с разными вредными вирусами и защищает нас от болезней и т. д.
Мы способны двигаться, потому что наши мышцы сокращаются и расслабляются благодаря белкам миозину и актину. Наши волосы и ногти состоят из белка кератина. Белок гемоглобин разносит кислород от легких ко всем клеткам нашего организма. Без белка пепсина, содержащегося в желудочном соке, мы не смогли бы переваривать пищу, а белок интерферон помогает организму справляться с разными вредными вирусами и защищает нас от болезней и т. д.
Но если в организме существуют вещества, выполняющие такие разные функции, почему же все они называются белками? А все дело в том, что абсолютно все белки, несмотря на свои различия, состоят из одного и того же «строительного материала» – особых химических веществ – аминокислот.
Внешне молекула белка напоминает нитку, унизанную разноцветными бусинками, где роль бусинок выполняют молекулы аминокислот. Как правило, молекула белка состоит из 300–500 таких «бусинок», а количество существующих в природе аминокислот ограничено – всего 20 видов. Значит, молекула белка может состоять из бусин 20 разных цветов, и, нанизывая бусины на нитку в разных комбинациях, мы получим различные варианты белковых молекул.
Значит, молекула белка может состоять из бусин 20 разных цветов, и, нанизывая бусины на нитку в разных комбинациях, мы получим различные варианты белковых молекул.
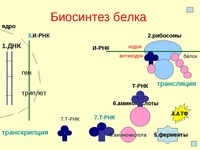
Каким же образом белки образуются в нашем теле? В любом организме существует своеобразная фабрика по производству (синтезу) белков, которая носит название рибосома. Работа рибосомы очень напоминает работу обычной фабрики, например, по производству автомобилей. В обычном мире, как известно, автомобиль строится не абы-как – с бухты-барахты – а строго по определенному чертежу. В наномире при синтезе белковой молекулы происходит абсолютно то же самое.
«Чертежи», необходимые для построения всех белков организма, хранятся в особой «библиотеке», роль которой выполняет молекула ДНК. Каждый «чертеж» в молекуле ДНК, соответствует конкретному белку и называется геном, а вся информация, содержащаяся в ДНК – генетической.
Но «библиотека» – это еще не «сборочный цех». Поэтому прежде, чем построить какой-нибудь белок, необходимо точно «скопировать» его «чертеж» из ДНК и доставить его на место сборки. Такое копирование и перенос осуществляет молекула информационной РНК (и-РНК), на которой специальным образом закодирована вся последовательность аминокислот для каждой белковой молекулы
Такое копирование и перенос осуществляет молекула информационной РНК (и-РНК), на которой специальным образом закодирована вся последовательность аминокислот для каждой белковой молекулы
Рис 1. Схема биосинтеза белка
Итак, молекула и-РНК с записанной на нее информацией направляется к рибосоме. Туда же направляется поток материала, из которого строится белок – молекулы аминокислоты. Аминокислоты попадают в рибосому не самостоятельно, а с помощью подвижных транспортных РНК (т-РНК). Эти молекулы умеют «распознавать» среди всего многообразия аминокислот только «свою» аминокислоту, присоединять её к себе и подтаскивать к рибосоме.
Рибосома медленно ползет по ленте и-РНК и, шаг за шагом, считывает с нее «код» следующей «бусины» в молекуле белка. Считав очередной «код», рибосома ждет, когда к ней «подъедет» т-РНК с необходимой аминокислотой. Если подъехавшая т-РНК «привезла» бусину неподходящего «цвета», ничего не происходит и т-РНК с прицепленной аминокислотой покидает рибосому, а к рибосоме «подъезжает» следующая т-РНК. И только в том случае, если считанный код совпал с подъехавшей «бусиной», аминокислота отделяется от т-РНК и присоединяется к строящейся цепочке белковой молекулы.
И только в том случае, если считанный код совпал с подъехавшей «бусиной», аминокислота отделяется от т-РНК и присоединяется к строящейся цепочке белковой молекулы.
Свободная т-РНК затем выбрасывается из рибосомы в окружающую среду. Здесь она захватывает новую молекулу аминокислоты и весь процесс повторяется снова. Напоминаем, что без этого нанопроцесса жизнь на Земле была бы невозможна, что еще раз подтверждает огромное значение законов наномира в нашей жизни!
Механизм работы рибосомы – это процесс производства конечной «нанопродукции» (белковой молекулы) из первоначального «наносырья» – атомов и молекул, причем процесс не беспорядочный, а по строго заданной программе, или рецепту.
Мы в своем «большом» мире (макромире) тоже производим автомобили, компьютеры, одежду, книги из исходного материала – железа, ткани, древесины и т.д. а методы для производства вещей с наперед заданной структурой называем технологией.
Мы убедились, что эти методы имеют много общего, и отличаются только лишь видом исходного «сырья»: на обычной фабрике – это какое-то вещество в большом объеме, а в наномире сырье – это атомы и молекулы. Поэтому методы манипулирования веществом на атомном и молекулярном уровнях с целью производства конечных продуктов с наперед заданной атомной структурой называется нанотехнологиями. А наномир, по которому мы с вами путешествуем, можно также назвать миром нанотехнологий.
Поэтому методы манипулирования веществом на атомном и молекулярном уровнях с целью производства конечных продуктов с наперед заданной атомной структурой называется нанотехнологиями. А наномир, по которому мы с вами путешествуем, можно также назвать миром нанотехнологий.
Человек часто копирует то, что до него создала природа (Как вы думаете, что появилось раньше – птица или самолет?) После того, как были хорошо изучены основные этапы биосинтеза белка, генные инженеры (ученые, которые занимаются изучением генетических механизмов ДНК) ученые задумались о том, каким образом можно попробовать создать устройства нанометрового размера, которые работали бы аналогично рибосомам в человеческом организме.
Автор: Мария Рыбалкина
Основные закономерности метаболических процессов в организме человека. Часть 2.
Рассматривая обмен веществ в условиях нормального функционирования организма, следует остановиться на безусловно взаимосвязанных, но в то же время достаточно специфичных составляющих метаболизма, а именно на углеводном, белковом, липидном и водно-электролитном обмене.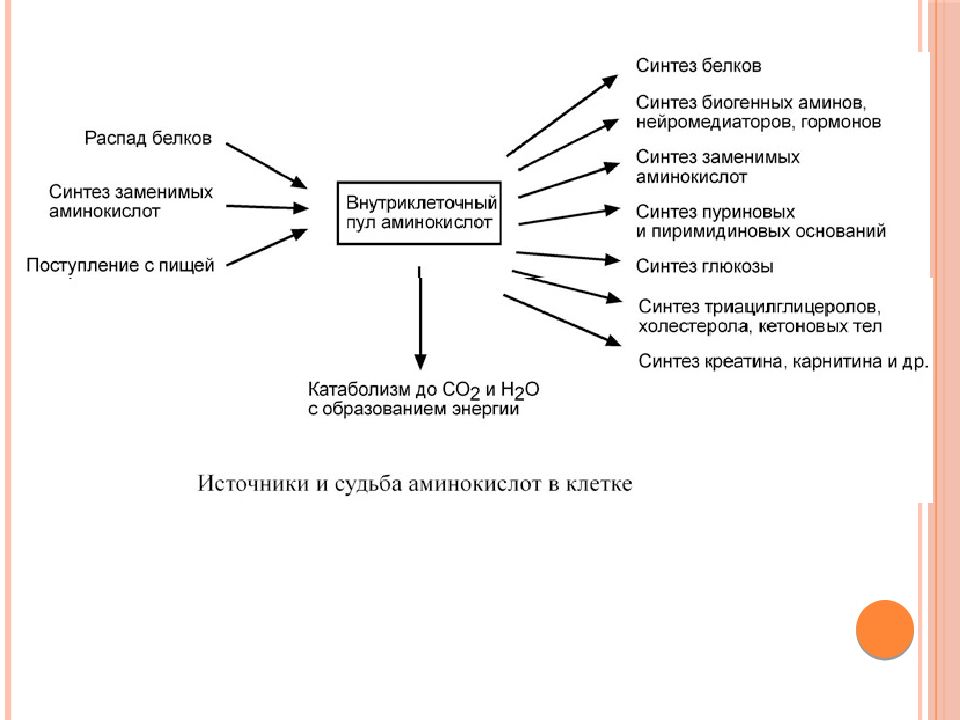
Очевидно, что основная роль углеводов в метаболизме определяется их энергетической функцией. Именно глюкоза крови вследствие наличия простого и быстрого пути гликолитической диссимиляции и последующего окисления в цикле трикарбоновых кислот, а также возможности максимально быстрого извлечения ее из депо гликогена, обеспечивающей экстренную мобилизацию энергетических ресурсов, является наиболее востребованным источником энергии в организме. Использование циркулирующей в плазме глюкозы разными органами неодинаково: мозг задерживает 12% глюкозы, кишечник— 9%, мышцы — 7%, почки — 5%. При этом уровень глюкозы плазмы крови является одной из важнейших гомеостатических констант организма, составляя 3, 3—5, 5 ммоль/л. Как известно снижение уровня глюкозы ниже допустимого передела имеет своим незамедлительным следствием дискоординацию деятельности ЦНС, проявляющуюся соответствующей клинической симптоматикой: головной мозг содержит небольшие резервы углеводов и нуждается в постоянном поступлении глюкозы, поскольку энергетические расходы мозга покрываются исключительно за счет углеводов. Глюкоза в тканях мозга преимущественно окисляется, а небольшая часть ее превращается в молочную кислоту.
Глюкоза в тканях мозга преимущественно окисляется, а небольшая часть ее превращается в молочную кислоту.
Единственной формой углеводов, которая может всасываться в кишечнике, являются моносахара. Они всасываются главным образом в тонкой кишке, током крови переносятся в печень и к тканям. Основная часть поступающей с пищей глюкозы (около 70%) окисляется в тканях до воды и углекислого газа, около 25—28% пищевой глюкозы превращается в жир и только 2—5% ее синтезируется в гликоген. Гликоген печени представляет собой основной резерв углеводов в организме, достигая по своей массе у взрослого человека 150—200 г. Синтез гликогена происходит достаточно быстро, что, наряду с быстрой мобилизацией гликогена и поступлением глюкозы в кровь в процессе гликогенолиза, является одним из механизмов поддержания гликемии в константных пределах. Помимо печени в качестве депо гликогена выступают также мышцы. Однако запас гликогена в мышечной массе по отношению к всему гликогену организма составляет всего 1 — 2%.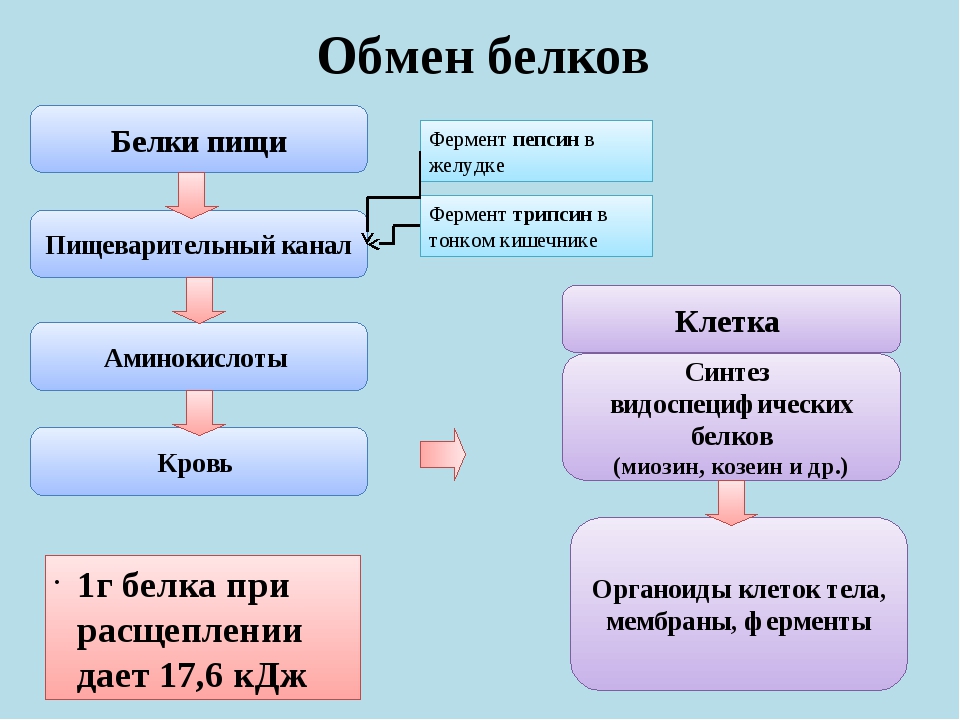 В мышцах под влиянием фермента фосфорилазы, которая активируется в начале мышечного сокращения, происходит усиленное расщепление гликогена, являющегося одним из источников энергии мышечного сокращения. При распаде мышечного гликогена процесс идет до образования пировиноградной и молочной кислот. Этот процесс называют гликолизом. В фазе отдыха из молочной кислоты в мышечной ткани происходит ресинтез гликогена.
В мышцах под влиянием фермента фосфорилазы, которая активируется в начале мышечного сокращения, происходит усиленное расщепление гликогена, являющегося одним из источников энергии мышечного сокращения. При распаде мышечного гликогена процесс идет до образования пировиноградной и молочной кислот. Этот процесс называют гликолизом. В фазе отдыха из молочной кислоты в мышечной ткани происходит ресинтез гликогена.
При полном отсутствии углеводов в пище они образуются в организме из продуктов трансформации жиров и белков. В печени возможно новообразование углеводов как из собственных продуктов их распада (пировиноградной или молочной кислоты), так и из продуктов диссимиляции жиров и белков (кетокислот и аминокислот), что обозначается как глюконеогенез. В результате трансформации аминокислот образуется пировиноградная кислота, при окислении жирных кислот — ацетилкоэнзим А, который может превращаться в пировиноградную кислоту — предшественник глюкозы. Это наиболее важный общий путь биосинтеза углеводов. Между двумя основными источниками энергии — углеводами и жирами — существует тесная физиологическая взаимосвязь. Повышение содержания глюкозы в крови увеличивает биосинтез триглицеридов и уменьшает распад жиров в жировой ткани. Поступление в кровь свободных жирных кислот уменьшается. В случае возникновения гипогликемии процесс синтеза триглицеридов тормозится, ускоряется распад жиров и в кровь в большом количестве поступают свободные жирные кислоты. Гликогенез, гликогенолиз и глюконеогенез являются тесно взаимосвязанными процессами, обеспечивающими оптимальный уровень глюкозы крови сообразно степени функционального напряжения организма.
Между двумя основными источниками энергии — углеводами и жирами — существует тесная физиологическая взаимосвязь. Повышение содержания глюкозы в крови увеличивает биосинтез триглицеридов и уменьшает распад жиров в жировой ткани. Поступление в кровь свободных жирных кислот уменьшается. В случае возникновения гипогликемии процесс синтеза триглицеридов тормозится, ускоряется распад жиров и в кровь в большом количестве поступают свободные жирные кислоты. Гликогенез, гликогенолиз и глюконеогенез являются тесно взаимосвязанными процессами, обеспечивающими оптимальный уровень глюкозы крови сообразно степени функционального напряжения организма.
Центральным звеном регуляции углеводного и других видов обмена и местом формирования сигналов, управляющих уровнем глюкозы, является гипоталамус. Отсюда регулирующие влияния реализуются вегетативными нервами и гуморальным путем, включающим эндокринные железы. Единственным гормоном, снижающим уровень гликемии, является инсулин — гормон, вырабатываемый β-клетками островков Ланхгерганса. Снижение гликемии происходит за счет усиления инсулином синтеза гликогена в печени и мышцах и повышения потребления глюкозы тканями организма. Увеличение уровня глюкозы в крови возникает при действии нескольких гормонов. Это глюкагон, продуцируемый α-клетками островков Ланхгерганса, адреналин — гормон мозгового слоя надпочечников, глюкокортикоиды — гормоны коркового слоя надпочечников, соматотропный гормон гипофиза, тироксин и трийодтиронин — гормоны щитовидной железы. Данные гормоны в связи с однонаправленностью их влияния на углеводный обмен и функциональным антагонизмом по отношению к эффектам инсулина часто объединяют понятием «контринсулярные гормоны».
Снижение гликемии происходит за счет усиления инсулином синтеза гликогена в печени и мышцах и повышения потребления глюкозы тканями организма. Увеличение уровня глюкозы в крови возникает при действии нескольких гормонов. Это глюкагон, продуцируемый α-клетками островков Ланхгерганса, адреналин — гормон мозгового слоя надпочечников, глюкокортикоиды — гормоны коркового слоя надпочечников, соматотропный гормон гипофиза, тироксин и трийодтиронин — гормоны щитовидной железы. Данные гормоны в связи с однонаправленностью их влияния на углеводный обмен и функциональным антагонизмом по отношению к эффектам инсулина часто объединяют понятием «контринсулярные гормоны».
Таким образом биологическая роль углеводов для организма человека определяется прежде всего их энергетической функцией. Обладая энергетической ценностью в 16, 7 кДж (4, 0 ккал) на 1 грамм вещества, углеводы являются основным источником энергии для всех клеток организма, при этом выполняя еще пластическую и опорную функции. Суточная потребность взрослого человека в углеводах составляет около 500 г.
Характерной особенностью белкового обмена является его чрезвычайная разветвленность. Достаточно указать, что в обмене 20 аминокислот, входящих в состав белковых молекул, в организме животных участвуют сотни промежуточных метаболитов, тесно связанных с обменом углеводов и липидов. Число ферментов, катализирующих химические реакции азотистого обмена, также исчисляется сотнями. Собственно белки (протеины и протеиды), высокомолекулярные соединения, построенные из мономеров — аминокислот, занимают ведущее место среди органических элементов организма, составляя более 50 % сухой массы клетки. Как известно, белки в организме выполняют ряд важнейших биологических функций, а именно:
— пластическая (структурная) функция заключается в том, что белки являются главной составной частью всех клеточных и межклеточных структур тканей;
— ферментная (каталитическая, энзимная) функция состоит в обеспечении всех химических реакций, протекающих в ходе обмена веществ в организме (дыхание, пищеварение, выделение), деятельностью ферментов, являющихся по своей структуре белками;
— транспортная функция белков заключается в их способности к соединению с целым рядом метаболитов и переносе последних в связанном состоянии в межтканевой жидкости и плазме крови к области их утилизации;
— защитная функция белков проявляется реализацией иммунного ответа образованием иммуноглобулинов (антител) и системы комплемента при поступлении в организм чужеродного белка, а также способностью к непосредственному связыванию экзогенных токсинов; белки системы гемостаза обеспечивают свертывание крови и остановку кровотечения при повреждении кровеносных сосудов;
— регуляторная функция, направленная на сохранение гомеостаза с поддержанием биологических констатнт организма, реализуется буферными свойствами молекулы протеинов, белковой структурой клеточных рецепторов, активируемых в свою очередь регуляторными полипептидами и гормонами, также имеющими белковую структуру;
— двигательная функция, обеспечивается взаимодействием сократительных белков мышечной ткани актина и миозина;
— энергетическая роль белков состоит в обеспечении организма энергией, образующейся при диссимиляции белковых молекул; при окислении 1 г белка в среднем освобождается энергия, равная 16, 7 кДж (4, 0 ккал).
В организме постоянно происходит распад и синтез белков. Единственным источником синтеза нового белка являются белки пищи. В пищеварительном тракте белки ферментативно расщепляются ферментами до аминокислот и абсорбируются в тонкой кишке. Транспорт их осуществляется двумя путями: через воротную систему печени, ведущую прямо в печень, и по лимфатическим сосудам, сообщающимся с кровью через грудной лимфатический проток. Максимальная концентрация аминокислот в крови достигается через 30 — 50 мин после приёма белковой пищи (углеводы и жиры замедляют всасывание аминокислот). Всасывание L-аминокислот (но не D-изомеров) — активный процесс, требующий затраты энергии. Аминокислоты переносятся через кишечную стенку от слизистой её поверхности в кровь. Перенос через щеточную кайму осуществляется целым рядом переносчиков, многие из которых действуют при участии Na+-зависимых механизмов симпорта, подобно переносу глюкозы.
Из аминокислот и простейших пептидов клетки тканей синтезируют собственный белок, который характерен только для данного организма.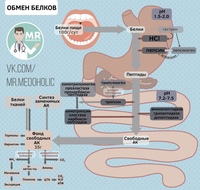 Белки не могут быть заменены другими пищевыми веществами, так как их синтез в организме возможен только из аминокислот. Вместе с тем белок может замещать собой жиры и углеводы, то есть использоваться для синтеза этих соединений. В тканях постоянно протекают процессы распада белка с последующим выделением из организма неиспользованных продуктов белкового обмена и параллельно с этим — синтез белков. Катаболизм большинства аминокислот начинается с отщепления α-аминогруппы результате реакций трансаминирования и дезаминирования. Чаще всего в реакциях трансаминирования участвуют аминокислоты, содержание которых в тканях значительно выше остальных — глутамат, аланин, аспартат и соответствующие им кетокислоты — α—кетоглутарат, пируват и оксалоацетат. Основным донором аминогруппы служит глутамат. Реакции трансаминирования играют большую роль в обмене аминокислот. Поскольку этот процесс обратим, ферменты аминотрансферазы функционируют как в процессах катаболизма, так и биосинтеза аминокислот.
Белки не могут быть заменены другими пищевыми веществами, так как их синтез в организме возможен только из аминокислот. Вместе с тем белок может замещать собой жиры и углеводы, то есть использоваться для синтеза этих соединений. В тканях постоянно протекают процессы распада белка с последующим выделением из организма неиспользованных продуктов белкового обмена и параллельно с этим — синтез белков. Катаболизм большинства аминокислот начинается с отщепления α-аминогруппы результате реакций трансаминирования и дезаминирования. Чаще всего в реакциях трансаминирования участвуют аминокислоты, содержание которых в тканях значительно выше остальных — глутамат, аланин, аспартат и соответствующие им кетокислоты — α—кетоглутарат, пируват и оксалоацетат. Основным донором аминогруппы служит глутамат. Реакции трансаминирования играют большую роль в обмене аминокислот. Поскольку этот процесс обратим, ферменты аминотрансферазы функционируют как в процессах катаболизма, так и биосинтеза аминокислот. Трансаминирование — заключительный этап синтеза заменимых аминокислот из соответствующих α-кетокислот, если они в данный момент необходимы клеткам. В результате происходит перераспределение аминного азота в тканях организма. Трансаминирование — первая стадия дезаминирования большинства аминокислот, то есть начальный этап их катаболизма. Образующиеся при этом кетокислоты окисляются в ЦТК или используются для синтеза глюкозы и кетоновых тел. При трансаминировании общее количество аминокислот в клетке не меняется. В свою очередь дезаминирование аминокислот — реакция отщепления α-аминогруппы от аминокислоты, в результате чего образуется соответствующая α-кетокислота (безазотистый остаток) и выделяется молекула аммиака. Аммиак токсичен для ЦНС, поэтому в организме человека и млекопитающих он превращается в нетоксичное хорошо растворимое соединение — мочевину. В виде мочевины, а также в виде солей аммония аммиак выводится из организма. Безазотистый остаток используется для образования аминокислот в реакциях трансаминирования.
Трансаминирование — заключительный этап синтеза заменимых аминокислот из соответствующих α-кетокислот, если они в данный момент необходимы клеткам. В результате происходит перераспределение аминного азота в тканях организма. Трансаминирование — первая стадия дезаминирования большинства аминокислот, то есть начальный этап их катаболизма. Образующиеся при этом кетокислоты окисляются в ЦТК или используются для синтеза глюкозы и кетоновых тел. При трансаминировании общее количество аминокислот в клетке не меняется. В свою очередь дезаминирование аминокислот — реакция отщепления α-аминогруппы от аминокислоты, в результате чего образуется соответствующая α-кетокислота (безазотистый остаток) и выделяется молекула аммиака. Аммиак токсичен для ЦНС, поэтому в организме человека и млекопитающих он превращается в нетоксичное хорошо растворимое соединение — мочевину. В виде мочевины, а также в виде солей аммония аммиак выводится из организма. Безазотистый остаток используется для образования аминокислот в реакциях трансаминирования.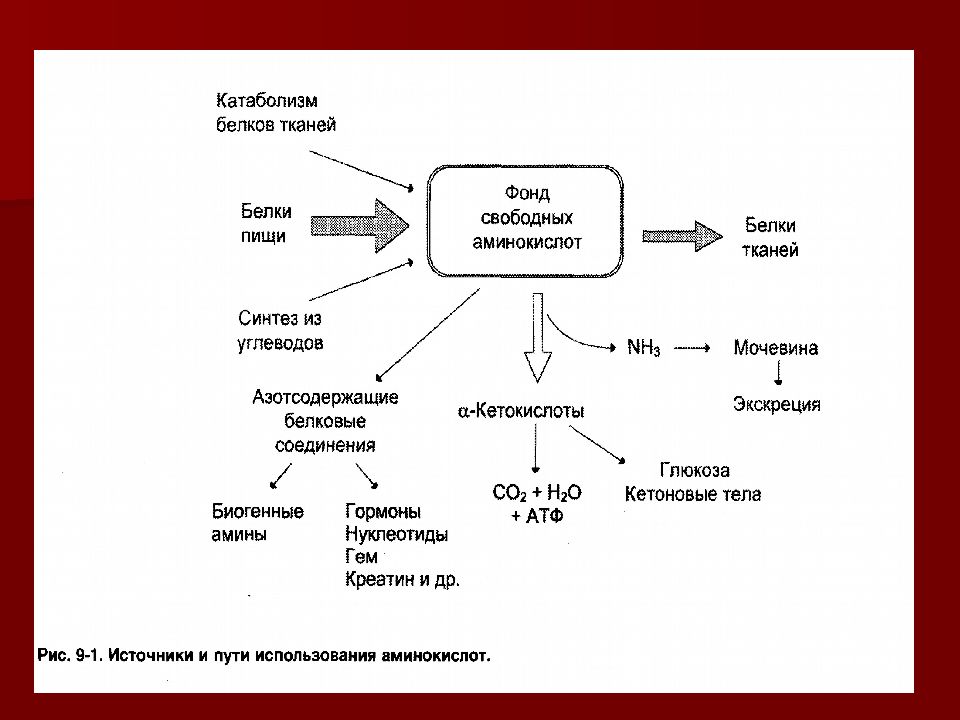
При катаболизме почти все природные аминокислоты сначала передают аминогруппу на а-кетоглутарат в реакции трансаминирования с образованием глутамата и соответствующей кетокислоты. Затем глутамат подвергается прямому окислительному дезаминированию под действием глутаматдегидрогеназы, в результате чего получаются а-кетоглутарат и аммиак. При необходимости синтеза аминокислот и наличии необходимых а-кетокислот обе стадии непрямого дезаминирования протекают в обратном направлении. В результате восстановительного аминирования а-кетоглутарата образуется глутамат, который вступает в трансаминирование с соответствующей а-кетокислотой, что приводит к синтезу новой аминокислоты. В случае использования белков в качестве источника энергии большинство аминокислот окисляются в конечном счёте через цикл лимонной кислоты до углекислого газа и воды. Прежде, чем эти вещества вовлекаются в заключительный этап катаболизма, их углеродный скелет превращается в двухуглеродный фрагмент в форме ацетил-КоА. Именно в этой форме большая часть молекул аминокислот включается в цикл лимонной кислоты.
Именно в этой форме большая часть молекул аминокислот включается в цикл лимонной кислоты.
Белки организма находятся в динамическом состоянии: из-за непрерывного процесса их разрушения и образования происходит обновление белков, скорость которого неодинакова для различных тканей. С наибольшей скоростью обновляются белки печени, слизистой оболочки кишечника, а также других внутренних органов и плазмы крови. Медленнее обновляются белки, входящие в состав клеток мозга, сердца, половых желез и еще медленнее — белки мышц, кожи и особенно опорных тканей (сухожилий, костей и хрящей). Важнейшими азотистыми продуктами распада белков, которые выделяются с мочой и потом, являются мочевина, мочевая кислота и аммиак. Преобладание в организме в данный момент времени синтеза или распада белка отражается понятием азотистого баланса — разностью между количеством азота, содержащегося в пище человека, и его уровнем в выделениях. Азотистым равновесием называют состояние, при котором количество выведенного азота равно количеству поступившего в организм. Азотистое равновесие наблюдается у здорового взрослого человека, если минимальное количество белков в пище соответствует 30-50 г/сут. Оптимальное количество поступления белка с пищей при средней физической нагрузке составляет около 100-120 г/сут. При положительном азотистом балансе количество азота в выделениях организма значительно меньше, чем содержание его в пище, то есть наблюдается задержка азота в организме. Положительный азотистый баланс отмечается у детей в связи с усиленным ростом, у женщин во время беременности, при усиленной спортивной тренировке, приводящей к увеличению мышечной массы, при заживлении обширных ран и при разрешении патологического процесса, связанного с выраженными системными нарушениями. Отрицательный азотистый баланс отмечается тогда, когда количество выделяющегося азота больше содержания его в пище, поступающей в организм. Отрицательный азотистый баланс наблюдается при белковом голодании, лихорадочных состояниях, нарушениях нейроэндокринной регуляции белкового обмена.
Некоторые аминокислоты не могут синтезироваться в организме человека и должны обязательно поступать с пищей в готовом виде. Эти аминокислоты принято называть незаменимыми, или эссенциальными. Экспериментально установлено, что из 20 входящих в состав белков аминокислот 12 синтезируются в организме (заменимые аминокислоты), а 8 не синтезируются (незаменимые аминокислоты) . К незаменимым аминоксилотам относятся: валин, метионин, треонин, лейцин, изолейцин, фенилаланин, триптофан и лизин. Две аминокислоты — аргинин и гистидин — у взрослых образуются в достаточных количествах, однако детям для нормального роста организма необходимо дополнительное поступление этих аминокислот с пищей. Поэтому их называют частично заменимыми. Две другие аминокислоты — тирозин и цистеин — условно заменимые, так как для их синтеза необходимы незаменимые аминокислоты. Тирозин синтезируется из фенилаланина, а для образования цистеина необходим атом серы метионина. Белки, содержащие весь необходимый набор аминокислот, называют биологически полноценными (табл. 1. 1. ). Наиболее высока биологическая ценность белков молока, яиц, рыбы, мяса. Биологически неполноценными называют белки, в составе которых отсутствует хотя бы одна аминокислота, которая не может быть синтезирована в организме. Неполноценными белками являются белки кукурузы, пшеницы, ячменя.
1. 1. ). Наиболее высока биологическая ценность белков молока, яиц, рыбы, мяса. Биологически неполноценными называют белки, в составе которых отсутствует хотя бы одна аминокислота, которая не может быть синтезирована в организме. Неполноценными белками являются белки кукурузы, пшеницы, ячменя.
Таблица 1. 1. Аминокислоты, входящие в состав белков человека.
|
1. Незаменимые
Валин
Лейцин
Изолейцин
Треонин
Метионин
Фенилаланин
Триптофан
Лизин
|
2. Частично заменимые
Гистидин
Аргинин
3. Условно заменимые
Цистеин
Тирозин
|
4. Заменимые
Аланин
Аспарагиновая кислота
Аспарагин
Глутаминовая кислота
Глутамин
Пролин
Глицин
Серин
|
Жиры (липиды) по своей химической структуре представляют собой триглицериды — сложные эфиры глицерина и жирных кислот (табл. 1. 2). Изначально эти соединения были объединены в одну химическую группу по общему признаку растворимости: они не растворяются в воде, но растворяются в органических растворителях (эфир, спирт, бензол). Жиры делят на простые липиды (нейтральные жиры, воски), сложные липиды (фосфолипиды, гликолипиды, сульфолипиды) и стероиды (холестерин). Основная масса липидов представлена в организме человека нейтральными жирами — триглицеридами олеиновой, пальмитиновой, стеариновой, линолевой и линоленовой жирных кислот.
1. 2). Изначально эти соединения были объединены в одну химическую группу по общему признаку растворимости: они не растворяются в воде, но растворяются в органических растворителях (эфир, спирт, бензол). Жиры делят на простые липиды (нейтральные жиры, воски), сложные липиды (фосфолипиды, гликолипиды, сульфолипиды) и стероиды (холестерин). Основная масса липидов представлена в организме человека нейтральными жирами — триглицеридами олеиновой, пальмитиновой, стеариновой, линолевой и линоленовой жирных кислот.
Таблица 1. 2. Классификация липидов организма человека.
|
1. Гликолипиды.
|
Содержат углеводный компонент.
|
|
2. Жиры.
|
Эфиры глицерина и высших жирных кислот. Химическое название — ацилглицерины. Преобладают триацилглицерины.
|
|
3.
|
Свободные жирные кислоты, жирорастворимые витамины, биологически активные вещества липидной природы — простагландины и др.
|
|
4. Стероиды.
|
В основе строения — полициклическая структура циклопентанпергидрофенантрен-стеран.
|
|
А. Стерины (спирты).
|
Наиболее важен холестерин.
|
|
В. Стериды.
|
Эфиры стеринов и высших жирных кислот. Наиболее распространены эфиры холестерина.
|
|
5. Фосфолипипы.
|
Отличительная особенность — остаток фосфорной кислоты в составе молекулы.
|
 Минорные липиды.
Минорные липиды. Жиры растительного и животного происхождения имеют различный состав жирных кислот, определяющий их физические свойства и физиолого-биохимические эффекты.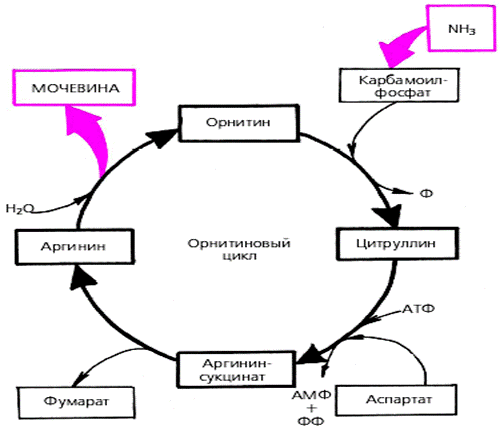 Жирные кислоты подразделяются на два основных класса — насыщенные и ненасыщенные. Насыщенность жира определяется количеством атомов водорода, которое содержит каждая жирная кислота (или, иначе, количеством двойных связей С=С). Жирные кислоты со средней длиной цепи (С8-С14) способны усваиваться в пищеварительном тракте без участия желчных кислот и панкреатической липазы, не депонируются в печени и подвергаются β-окислению. Животные жиры могут содержать насыщенные жирные кислоты с длиной цепи до двадцати и более атомов углерода, они имеют твердую консистенцию и высокую температуру плавления. Как известно высокое потребление насыщенных жирных кислот является важнейшим фактором риска развития диабета, ожирения, атеросклероза. К мононенасыщенным жирным кислотам относятся миристолеиновая и пальмитолеиновая кислоты (жиры рыб и морских млекопитающих), олеиновая (оливковое, сафлоровое, кунжутное, рапсовое масла). Мононенасыщенные жирные кислоты помимо их поступления с пищей в организме синтезируются из насыщенных жирных кислот и частично из углеводов.
Жирные кислоты подразделяются на два основных класса — насыщенные и ненасыщенные. Насыщенность жира определяется количеством атомов водорода, которое содержит каждая жирная кислота (или, иначе, количеством двойных связей С=С). Жирные кислоты со средней длиной цепи (С8-С14) способны усваиваться в пищеварительном тракте без участия желчных кислот и панкреатической липазы, не депонируются в печени и подвергаются β-окислению. Животные жиры могут содержать насыщенные жирные кислоты с длиной цепи до двадцати и более атомов углерода, они имеют твердую консистенцию и высокую температуру плавления. Как известно высокое потребление насыщенных жирных кислот является важнейшим фактором риска развития диабета, ожирения, атеросклероза. К мононенасыщенным жирным кислотам относятся миристолеиновая и пальмитолеиновая кислоты (жиры рыб и морских млекопитающих), олеиновая (оливковое, сафлоровое, кунжутное, рапсовое масла). Мононенасыщенные жирные кислоты помимо их поступления с пищей в организме синтезируются из насыщенных жирных кислот и частично из углеводов.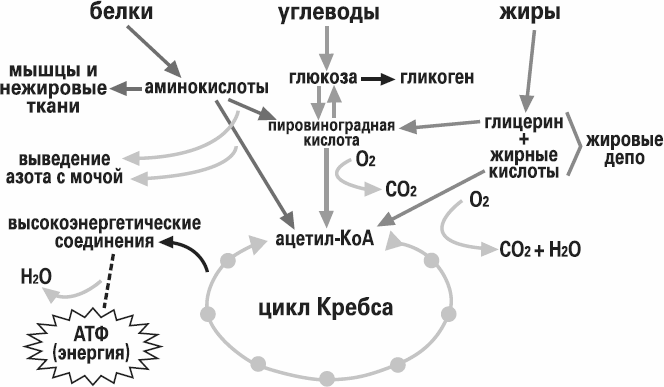 Жирные кислоты с двумя и более двойными связями между углеродными атомами называются полиненасыщенными – ПНЖК. Особое значение для организма человека имеют такие ПНЖК как линолевая, линоленовая, являющиеся структурными элементами клеточных мембран и обеспечивающие нормальное развитие и адаптацию организма человека к неблагоприятным факторам окружающей среды. ПНЖК являются предшественниками образующихся из них биорегуляторов – эйкозаноидов. Двумя основными группами ПНЖК являются кислоты семейств ω-6 и ω-3. Жирные кислоты ω-6 содержатся практически во всех растительных маслах и орехах. ω-3 жирные кислоты также содержатся в ряде масел (льняном, из семян крестоцветных, соевом). Основным пищевым источником ω-3 жирных кислот являются жирные сорта рыб и некоторые морепродукты. Из ПНЖК ω — 6 особое место занимает линолевая кислота, которая является предшественником наиболее физиологически активной кислоты этого семейства — арахидоновой. Арахидоновая кислота является преобладающим представителем ПНЖК в организме человека и служит субстратом для синтеза простагландинов и лейкотриенов.
Жирные кислоты с двумя и более двойными связями между углеродными атомами называются полиненасыщенными – ПНЖК. Особое значение для организма человека имеют такие ПНЖК как линолевая, линоленовая, являющиеся структурными элементами клеточных мембран и обеспечивающие нормальное развитие и адаптацию организма человека к неблагоприятным факторам окружающей среды. ПНЖК являются предшественниками образующихся из них биорегуляторов – эйкозаноидов. Двумя основными группами ПНЖК являются кислоты семейств ω-6 и ω-3. Жирные кислоты ω-6 содержатся практически во всех растительных маслах и орехах. ω-3 жирные кислоты также содержатся в ряде масел (льняном, из семян крестоцветных, соевом). Основным пищевым источником ω-3 жирных кислот являются жирные сорта рыб и некоторые морепродукты. Из ПНЖК ω — 6 особое место занимает линолевая кислота, которая является предшественником наиболее физиологически активной кислоты этого семейства — арахидоновой. Арахидоновая кислота является преобладающим представителем ПНЖК в организме человека и служит субстратом для синтеза простагландинов и лейкотриенов.
Источниками жира в организме являются экзогенный жир, поступающий с пищей, и эндогенный жир, синтезируемый в печени из углеводов. Жир, всасывающийся из кишечника, поступает преимущественно в лимфу и в меньшем количестве — непосредственно в кровь. Большая часть жиров в организме находится в жировой ткани, меньшая часть входит в состав клеточных структур. В жировой ткани жир, находящийся в клетке в виде включений, легко выявляется при микроскопическом и гистохимическом исследованиях. Жировые вакуоли в клетках — это резервный жир, используемый для обеспечения прежде всего энергетических потребностей клетки. Больше всего запасного жира содержится в жировой ткани, а также в некоторых органах, например в печени и мышцах. Количество запасного жира зависит от характера питания, количества пищи, конституциональных особенностей, а также от величины расхода энергии при мышечной деятельности; количество же протоплазматического жира является устойчивым и постоянным. В жировой ткани нейтральный жир депонируется виде триглицеридов. Сложные липиды — фосфолипиды и гликолипиды — входят в состав всех клеток, но в большей степени в состав клеток нервной ткани. Общее количество жира в организме человека колеблется в широких пределах и в среднем составляет 10—20% от массы тела, а в случае патологического ожирения может достигать 50%. Суточная потребность взрослого человека в нейтральном жире составляет 70—80 г. У человека состав и свойства жира относительно постоянны. При употреблении пищи, содержащей даже небольшое количество жира, в теле человека жир все же откладывается в депо. При этом эндогенный жир имеет некоторые видовые особенности, однако видовая специфичность жиров выражена несравнимо меньше, чем видовая специфичность белков.
Сложные липиды — фосфолипиды и гликолипиды — входят в состав всех клеток, но в большей степени в состав клеток нервной ткани. Общее количество жира в организме человека колеблется в широких пределах и в среднем составляет 10—20% от массы тела, а в случае патологического ожирения может достигать 50%. Суточная потребность взрослого человека в нейтральном жире составляет 70—80 г. У человека состав и свойства жира относительно постоянны. При употреблении пищи, содержащей даже небольшое количество жира, в теле человека жир все же откладывается в депо. При этом эндогенный жир имеет некоторые видовые особенности, однако видовая специфичность жиров выражена несравнимо меньше, чем видовая специфичность белков.
Основная биологическая роль жиров — обеспечение пластического и энергетического обмена в организме. Пластическая роль липидов состоит в том, что они входят в состав клеточных мембран, в значительной мере определяя их свойства. Фосфатиды и стерины входят в состав клеточных структур, в частности клеточных мембран, а также ядерного вещества и цитоплазмы. Исключительно важное физиологическое значение имеют стерины, в частности холестерин. Это вещество входит в состав клеточных мембран, является источником образования желчных кислот, а также гормонов коры надпочечников и половых желез, витамина D. Печень является практически единственным органом, поддерживающим уровень фосфолипидов в крови и местом синтеза эндогенного холестерина. В плазме крови холестерин находится в составе липопротеидных комплексов, с помощью которых и осуществляется его транспорт. У взрослых людей 67—70% холестерина плазмы крови находится в составе липопротеидов низкой плотности (ЛПНП), 9—10% — в составе липопротеидов очень низкой плотности (ЛПОНП) и 20—24% — в составе липопротеидов высокой плотности (ЛПВП). Давно доказано, что именно липопротеиды определяют уровень холестерина и динамику его обмена.
Исключительно важное физиологическое значение имеют стерины, в частности холестерин. Это вещество входит в состав клеточных мембран, является источником образования желчных кислот, а также гормонов коры надпочечников и половых желез, витамина D. Печень является практически единственным органом, поддерживающим уровень фосфолипидов в крови и местом синтеза эндогенного холестерина. В плазме крови холестерин находится в составе липопротеидных комплексов, с помощью которых и осуществляется его транспорт. У взрослых людей 67—70% холестерина плазмы крови находится в составе липопротеидов низкой плотности (ЛПНП), 9—10% — в составе липопротеидов очень низкой плотности (ЛПОНП) и 20—24% — в составе липопротеидов высокой плотности (ЛПВП). Давно доказано, что именно липопротеиды определяют уровень холестерина и динамику его обмена.
Энергетическая роль жиров определяется их максимальной среди всех биологических молекул энергоемкостью, более чем в два раза превышающую таковую углеводов или белков. При окислении 1 г жира выделяется 37, 7 кДж (9, 0 ккал) энергии. В отличие от углеводов жиры составляют энергетический резерв организма. Преимущество жира в качестве энергетического резерва заключается в том, что жиры являются более восстановленными веществами по сравнению с углеводами (в молекулах углеводов при каждом углеродном атоме есть кислород — группы -CHOH-; у жира имеются длинные углеводородные радикалы, в которых преобладают группы -Ch3- — в них нет кислорода). От жира можно отнять больше водорода, который затем проходит по цепи митохондриального окисления с образованием АТФ. Еще одним преимуществом жира как энергетического резерва, в отличие от углеводов, является гидрофобность — он не связан с водой. Это обеспечивает компактность жировых запасов — они хранятся в безводной форме, занимая малый объем. В среднем, у человека запас чистых триацилглицеринов составляет примерно 13 кг. Этих запасов могло бы хватить на 40 дней голодания в условиях умеренной физической нагрузки. Для сравнения: общие запасы гликогена в организме — примерно 400 г; при голодании этого количества не хватает даже на одни сутки.
При окислении 1 г жира выделяется 37, 7 кДж (9, 0 ккал) энергии. В отличие от углеводов жиры составляют энергетический резерв организма. Преимущество жира в качестве энергетического резерва заключается в том, что жиры являются более восстановленными веществами по сравнению с углеводами (в молекулах углеводов при каждом углеродном атоме есть кислород — группы -CHOH-; у жира имеются длинные углеводородные радикалы, в которых преобладают группы -Ch3- — в них нет кислорода). От жира можно отнять больше водорода, который затем проходит по цепи митохондриального окисления с образованием АТФ. Еще одним преимуществом жира как энергетического резерва, в отличие от углеводов, является гидрофобность — он не связан с водой. Это обеспечивает компактность жировых запасов — они хранятся в безводной форме, занимая малый объем. В среднем, у человека запас чистых триацилглицеринов составляет примерно 13 кг. Этих запасов могло бы хватить на 40 дней голодания в условиях умеренной физической нагрузки. Для сравнения: общие запасы гликогена в организме — примерно 400 г; при голодании этого количества не хватает даже на одни сутки.
Катаболизм жира включает в себя три этапа: 1) гидролиз жира до глицерина и жирных кислот (липолиз) ; 2) трансформация глицерина с последующим вступлением продуктов в гексозобифосфатный путь, а также окисление жирных кислот до ацетил-КоА; 3) вступление вышеуказанных продуктов в цикл трикарбоновых кислот. Кроме указанных этапов к катаболизму жиров относят также окисление кетоновых тел и перекисное окисление липидов. Обмен полученного в результате липолиза глицерина может осуществляться несколькими путями. Значительная часть образовавшегося при гидролизе липидов глицерина используется для ресинтеза триглицеридов. Второй путь обмена глицерина — включение продукта его окисления в гликолиз или в глюконеогенез. Окисление жирных кислот осуществляется различными путями, наиболее значимым из них является β-окисление. В ходе β-окисления последовательно происходит активация жирной кислоты на мембране митохондрии и ее связывание с молекулой карнитина, прохождение комплекса нв внутреннюю поверхность мембраны митохондрии, внутримитохондриальное окисление жирной кислоты с образованием ацетил-КоА и АТФ.
Одним из продуктов катаболизма жиров, имеющем важное значения для метаболизма в целом являются кетоновые тела. Кетоновые тела — группа органических соединений, являющихся промежуточными продуктами жирового, углеводного и белкового обменов. К кетоновым телам относят β-оксимасляную и ацетоуксусную кислоты и ацетон, имеющие сходное строение и способные к взаимопревращениям. Главным путем синтеза кетоновых тел, происходящего в основном в печени, считается реакция конденсации между двумя молекулами ацетил-КоА, образовавшегося при β-окислении жирных кислот или при окислительном декарбоксилировании пирувата (пировиноградной кислоты) в процессе обмена глюкозы и ряда аминокислот. Данный путь синтеза кетоновых тел более других зависит от характера питания и в большей степени страдает при патологических нарушениях обмена веществ. Из печени кетоновые тела поступают в кровь и с нею во все остальные органы и ткани, где они включаются в универсальный энергообразующий цикл — цикл трикарбоновых кислот, в котором окисляются до углекислоты и воды. Кетоновые тела используются также для синтеза холестерина, высших жирных кислот, фосфолипидов и заменимых аминокислот. При голодании, однообразном безуглеводистом питании и при недостаточной секреции инсулина использование ацетил-КоА в цикле трикарбоновых кислот подавляется, так как все метаболически доступные ресурсы организма превращаются в глюкозу крови. В этих условиях увеличивается синтез кетоновых тел. Следует подчеркнуть важную роль кетоновых тел в поддержании энергетического баланса. Кетоновые тела – поставщики «топлива» для мышц, почек и действуют, возможно, как часть регуляторного механизма с обратной связью, предотвращая чрезвычайную мобилизацию жирных кислот из жировых депо. Печень в этом смысле является исключением, она не использует кетоновые тела в качестве энергетического материала.
Кетоновые тела используются также для синтеза холестерина, высших жирных кислот, фосфолипидов и заменимых аминокислот. При голодании, однообразном безуглеводистом питании и при недостаточной секреции инсулина использование ацетил-КоА в цикле трикарбоновых кислот подавляется, так как все метаболически доступные ресурсы организма превращаются в глюкозу крови. В этих условиях увеличивается синтез кетоновых тел. Следует подчеркнуть важную роль кетоновых тел в поддержании энергетического баланса. Кетоновые тела – поставщики «топлива» для мышц, почек и действуют, возможно, как часть регуляторного механизма с обратной связью, предотвращая чрезвычайную мобилизацию жирных кислот из жировых депо. Печень в этом смысле является исключением, она не использует кетоновые тела в качестве энергетического материала.
Процесс образования, отложения и мобилизации из депо жира регулируется нервной и эндокринной системами, а также тканевыми механизмами и тесно связаны с углеводным обменом. Так, повышение концентрации глюкозы в крови уменьшает распад триглицеридов и активизирует их синтез. Понижение концентрации глюкозы в крови, наоборот, тормозит синтез триглицеридов и усиливает их расщепление. Таким образом, взаимосвязь жирового и углеводного обменов направлена на обеспечение энергетических потребностей организма. При избытке углеводов в пище триглицериды депонируются в жировой ткани, при нехватке углеводов происходит расщепление триглицеридов с образованием неэтерифицнрованных жирных кислот, служащих источником энергии. В обмене жиров одна из важнейших ролей принадлежит печени. Печень — основной орган, в котором происходит образование кетоновых тел (бета-оксимасляная, ацетоуксусная кислоты, ацетон), используемых как альтернативный глюкозе источник энергии.
Понижение концентрации глюкозы в крови, наоборот, тормозит синтез триглицеридов и усиливает их расщепление. Таким образом, взаимосвязь жирового и углеводного обменов направлена на обеспечение энергетических потребностей организма. При избытке углеводов в пище триглицериды депонируются в жировой ткани, при нехватке углеводов происходит расщепление триглицеридов с образованием неэтерифицнрованных жирных кислот, служащих источником энергии. В обмене жиров одна из важнейших ролей принадлежит печени. Печень — основной орган, в котором происходит образование кетоновых тел (бета-оксимасляная, ацетоуксусная кислоты, ацетон), используемых как альтернативный глюкозе источник энергии.
При обильном углеводном питании и отсутствии жиров в пище синтез жира в организме может происходить из углеводов. Источником углерода для синтеза жирных кислот служит ацетил-КоА, образующийся при распаде глюкозы в абсорбтивном периоде. В норме у человека 25—30% углеводов пищи превращается в жиры. Превращение белка в жирные кислоты происходит, вероятнее всего, также через образование углеводов.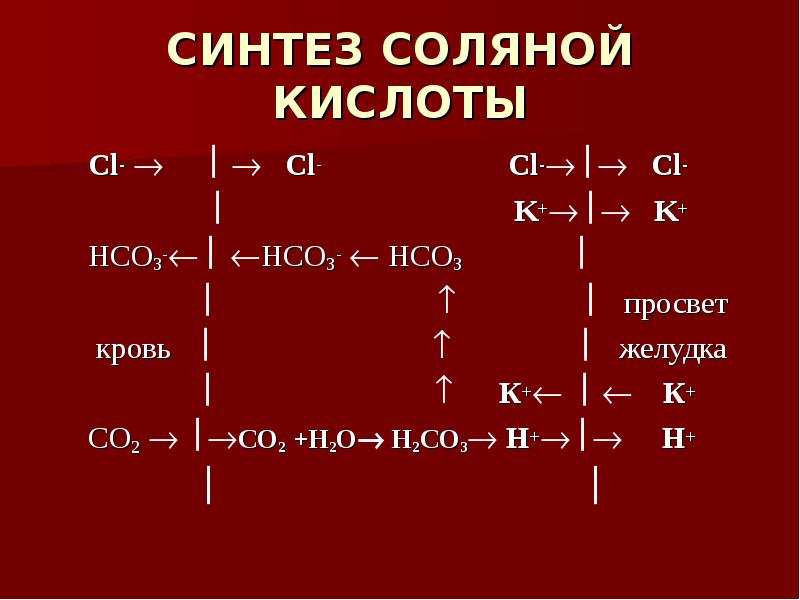 С другой стороны и нейтральные жиры в энергетическом отношении могут быть заменены углеводами. Тем не менее жиры необходимы для нормальной жизнедеятельности. Известно, что длительное исключение жиров из пищевого рациона может явиться причиной возникновения целого ряда тяжелых метаболических нарушений. Отчасти это связано с отсутствием поступления в организм жирорастворимых витаминов (A, D, E, K). Но основная причина метаболических нарушений кроется в возникновении в организме дефицита незаменимых жирных кислот. Некоторые ненасыщенные жирные кислоты (с числом двойных связей более 1), например линолевая, линоленовая и арахидоновая, в организме человека и некоторых животных не образуются из других жирных кислот и поэтому являются незаменимыми. Особенно остро реагирует организм на дефицит незаменимой линолевой кислоты СН3- (СН2) 4 — СН = СН — СН2 — СН = СН — (СН2) 7 — СООН. Возможно это связано с тем, что эта ненасыщенная жирная кислота в организме человека служит предшественником арахидоновой кислоты, которая в свою очередь необходима для синтеза универсальных биорегуляторов — простагландинов.
С другой стороны и нейтральные жиры в энергетическом отношении могут быть заменены углеводами. Тем не менее жиры необходимы для нормальной жизнедеятельности. Известно, что длительное исключение жиров из пищевого рациона может явиться причиной возникновения целого ряда тяжелых метаболических нарушений. Отчасти это связано с отсутствием поступления в организм жирорастворимых витаминов (A, D, E, K). Но основная причина метаболических нарушений кроется в возникновении в организме дефицита незаменимых жирных кислот. Некоторые ненасыщенные жирные кислоты (с числом двойных связей более 1), например линолевая, линоленовая и арахидоновая, в организме человека и некоторых животных не образуются из других жирных кислот и поэтому являются незаменимыми. Особенно остро реагирует организм на дефицит незаменимой линолевой кислоты СН3- (СН2) 4 — СН = СН — СН2 — СН = СН — (СН2) 7 — СООН. Возможно это связано с тем, что эта ненасыщенная жирная кислота в организме человека служит предшественником арахидоновой кислоты, которая в свою очередь необходима для синтеза универсальных биорегуляторов — простагландинов.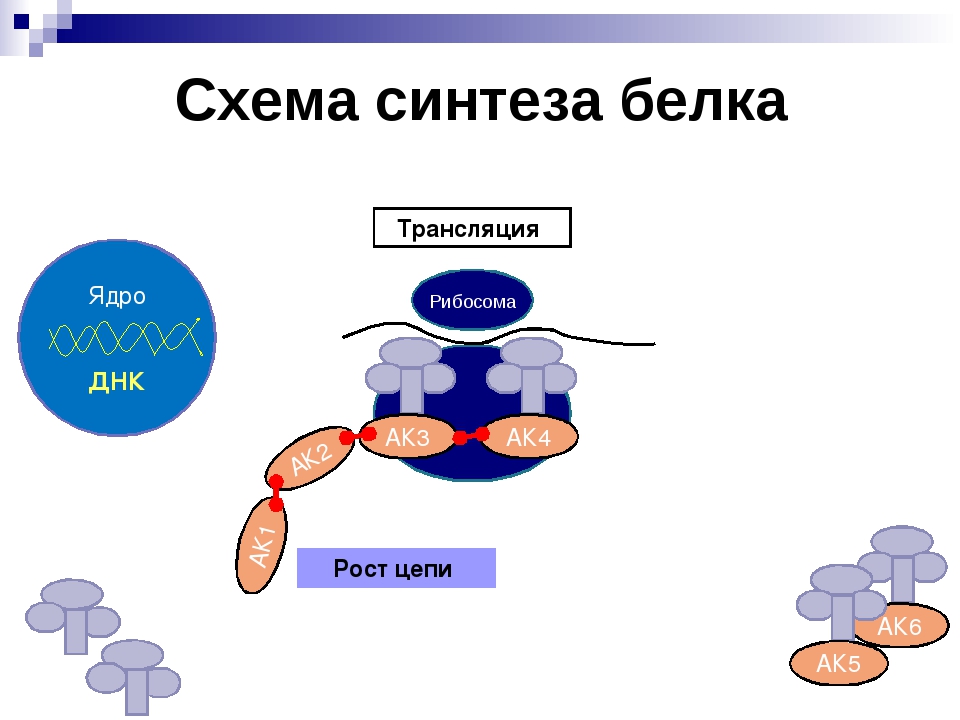 Основными пищевыми источниками полиненасыщенных жирных кислот, в том числе линолевой, являются растительные масла.
Основными пищевыми источниками полиненасыщенных жирных кислот, в том числе линолевой, являются растительные масла.
Как указывалось выше метаболизм жиров контролируется нервной и эндокринной системами. Мобилизация жиров из депо происходит под влиянием гормонов мозгового слоя надпочечников — адреналина и норадреналина. Соматотропный гормон гипофиза также обладает жиромобилизирующим действием. Аналогично действует тироксин — гормон щитовидной железы. Тормозят мобилизацию жира глюкокортикоиды — гормоны коркового слоя надпочечника, вероятно, вследствие того, что они несколько повышают уровень глюкозы в крови. Действие инсулина связано с повышением активности внутриклеточной фосфодиэстеразы, что приводит к снижению концентрации цАМФ и угнетению липолиза. Таким образом, инсулин усиливает синтез жира и уменьшает скорость его мобилизации. Имеются данные, свидетельствующие о возможности прямых нервных влияний на обмен жиров. Симпатические влияния тормозят синтез триглицеридов и усиливают их распад.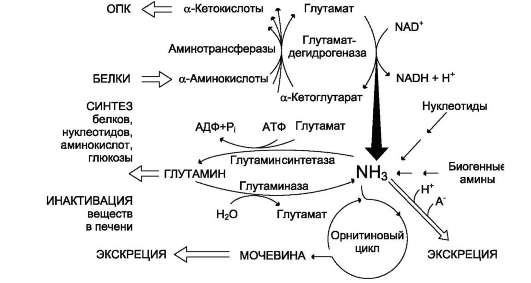 Парасимпатические влияния, напротив, способствуют отложению жира в депо.
Парасимпатические влияния, напротив, способствуют отложению жира в депо.
Статья добавлена 31 мая 2016 г.
Как старение влияет на синтез белка
В последние годы старение как биологический процесс привлекает все большее внимание молекулярных биологов, которые исследуют, что происходит в стареющем организме на молекулярном уровне. Группа исследователей из НИИ ФХБ имени А.Н. Белозерского, Института молекулярной биологии РАН имени В. А. Энгельгардта и Гарвардской медицинской школы изучила, как в ходе старения изменяются транскрипция и трансляция ряда генов, связанных с работой иммунной системы, метаболизмом и защитой от повреждений, а также описала некоторые любопытные возрастные изменения, затрагивающие динамику трансляции.
Старение как биологический феномен привлекает внимание не только биохакеров, стремящихся максимально увеличить продолжительность жизни, но и молекулярных биологов, занимающихся фундаментальной наукой.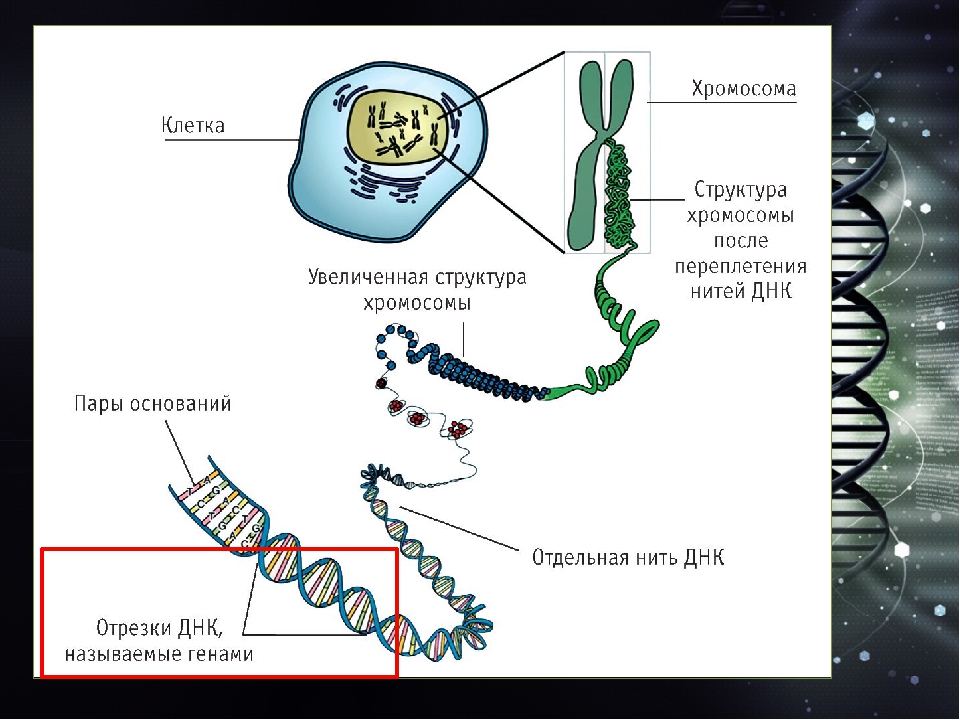 И их старания не проходят даром.
И их старания не проходят даром.
Например, сейчас известно, что в ходе жизни организма в его тканях накапливаются неправильно уложенные, окисленные или иным образом модифицированные белки, которые не только не выполняют свою функцию должным образом, но и могут образовывать нерастворимые скопления, нарушающие функционирование целых тканей [1].
Несколько исследований продемонстрировали, что с возрастом в организме в целом снижается интенсивность синтеза белка, уменьшается концентрация факторов инициации и элонгации трансляции, замедляется аминоацилирование тРНК. Сообщается, что в ходе старения происходит интенсивное метилирование промоторов генов рРНК, а концентрация самих рРНК уменьшается [1].
Вместе с тем биологические виды, отличающиеся необычно высокой для своей таксономической группы продолжительностью жизни, как правило, характеризуются очень стабильным протеомом и высокой устойчивостью к окислению белков. Примечательно, что виды, у которых в целом интенсивность синтеза белка понижена, характеризуются повышенной продолжительностью жизни, кроме того, видам-долгожителям вроде голого землекопа [2] часто присущи высокоточный аппарат трансляции и низкий кругооборот белков [1].
Показано также, что экспериментальный нокаут или нокдаун некоторых компонентов аппарата трансляции у червей-нематод [3] существенно увеличивает продолжительность их жизни [1]. Да и в целом широко распространенное представление о положительном влиянии на продолжительность жизни низкокалорийного питания хорошо вписывается в идею о том, что интенсивный метаболизм, в том числе и активный биосинтез белка, по крайней мере частично лежит в основе старения организма . Но что именно происходит с трансляцией как со сложнейшим молекулярным процессом в ходе старения организма? Трансляция каких генов при старении меняется сильнее всего?
Для ответа на эти вопросы группа ученых из НИИ ФХБ имени А.Н. Белозерского, ИМБ РАН имени В. А. Энгельгардта и Гарвардской медицинской школы досконально изучила паттерны транскрипции и трансляции генов в печени и почке у мышей в возрасте от 1–3 месяцев до 32 месяцев — весьма преклонного для мышей возраста [1]. Анализ профиля транскрипции исследователи проводили с помощью секвенирования РНК, а для изучения трансляции использовали метод, известный как рибосомный профайлинг.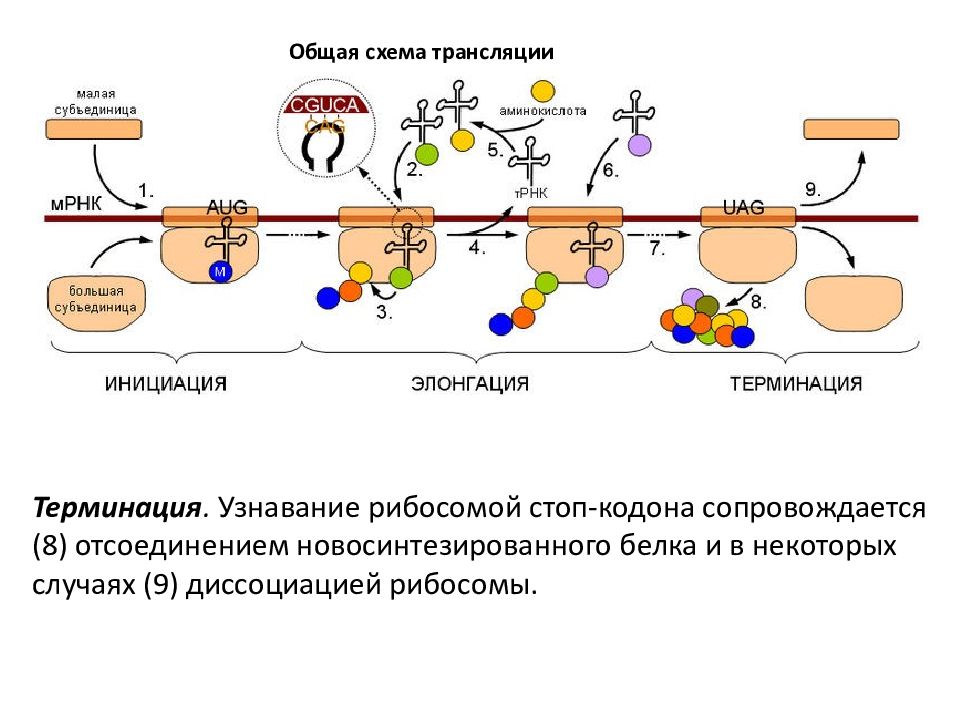 Он заключатся в моментальной остановке рибосом на читаемых ими мРНК с последующим точным определением их положения и количества . В комбинации с секвенированием транскриптома метод позволяет выяснить, на каких мРНК рибосом больше и, следовательно, с каких транскриптов синтезируется больше белка (рис. 1). Включение в исследование мышей нескольких возрастных групп позволило построить зависимость экспрессии определенных генов от возраста. Так, оказалось, что для большинства мышиных генов зависимость уровня экспрессии конкретного гена от времени имеет U-образную выпуклую или вогнутую форму с максимумом в случае одних генов или минимумом в случае других генов в районе 3–11 месяцев, вероятно, отражая переход от ювенильного к взрослому состоянию.
Он заключатся в моментальной остановке рибосом на читаемых ими мРНК с последующим точным определением их положения и количества . В комбинации с секвенированием транскриптома метод позволяет выяснить, на каких мРНК рибосом больше и, следовательно, с каких транскриптов синтезируется больше белка (рис. 1). Включение в исследование мышей нескольких возрастных групп позволило построить зависимость экспрессии определенных генов от возраста. Так, оказалось, что для большинства мышиных генов зависимость уровня экспрессии конкретного гена от времени имеет U-образную выпуклую или вогнутую форму с максимумом в случае одних генов или минимумом в случае других генов в районе 3–11 месяцев, вероятно, отражая переход от ювенильного к взрослому состоянию.
Рисунок 1. Схема исследования, проведенного авторами статьи. Ткани почек и печени, взятые от мышей различных возрастов, были проанализированы с помощью секвенирования РНК и рибосомного профайлинга. Полученные данные позволили оценить, какой вклад вносит трансляция в возрастные изменения экспрессии генов и установить некоторые другие любопытные эффекты, связанные с возрастными изменениями трансляции.
Так, по данным экспериментов, в печени и почках мышей с возрастом заметно повышается экспрессия генов, связанных с воспалением и другими иммунными процессами, что соответствует представлениям о развитии хронического воспаления. Сравнив свои данные с результатами более ранних работ на эту тему, авторы смогли выявить четкую и устойчивую возрастную зависимость уровня экспрессии пяти генов, связанных с воспалением и лизосомной деградацией белков (Ctss, C1qa, C1qb, C1qc и Laptm5), так что их можно рассматривать в качестве потенциальных биомаркеров старения.
В отличие от воспалительных процессов, функции митохондрий с возрастом не усиливаются, а угасают. Постепенное возрастное снижение экспрессии некоторых ядерных генов, кодирующих митохондриальные белки, было обнаружено в клетках почек (в клетках печени тоже наблюдалось, но оказалось менее выраженным). Кроме того, синтез компонентов, ответственных за поддержание окислительно-восстановительного баланса, менялся и в печени, и в почках, что хорошо согласуется с широко распространенной теорией о роли окислительного стресса в процессе старения.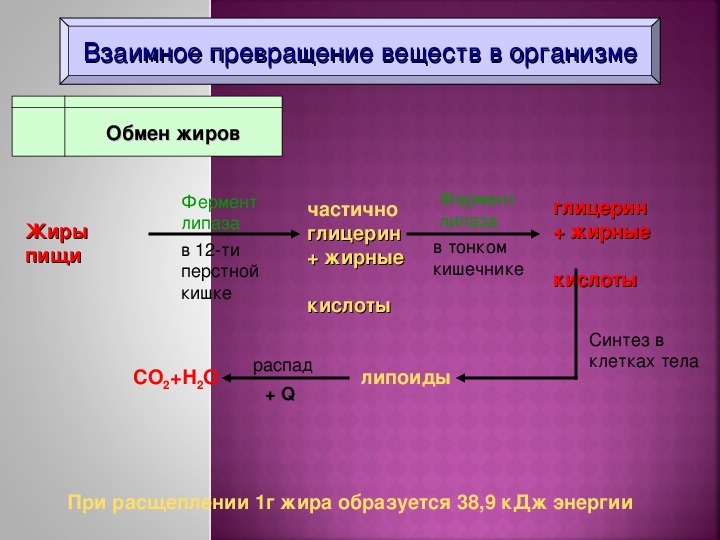 В то же время в обеих тканях удалось выявить возрастное увеличение экспрессии генов, связанных с регуляцией давления в кровеносных сосудах, а также генов, кодирующих предшественники амилоидов. Также с возрастом в почках снижается синтез белков, связанных с ответом на глюкокортикоидные гормоны, биосинтезом кофакторов и метаболизмом липидов. Примечательно возрастное снижение экспрессии в почках двадцати одного гена компонентов пероксисом, что также согласуется с текущим взглядом на роль возрастных изменений в метаболизме липидов и развитии некоторых заболеваний почек.
В то же время в обеих тканях удалось выявить возрастное увеличение экспрессии генов, связанных с регуляцией давления в кровеносных сосудах, а также генов, кодирующих предшественники амилоидов. Также с возрастом в почках снижается синтез белков, связанных с ответом на глюкокортикоидные гормоны, биосинтезом кофакторов и метаболизмом липидов. Примечательно возрастное снижение экспрессии в почках двадцати одного гена компонентов пероксисом, что также согласуется с текущим взглядом на роль возрастных изменений в метаболизме липидов и развитии некоторых заболеваний почек.
В общем и целом, большинство возрастных изменений уровня трансляции коррелирует с изменениями уровня транскрипции как в печени, так и в почках, поэтому, вероятнее всего, ключевые изменения экспрессии генов, связанные со старением, происходят именно на уровне транскрипции. Авторы исследования сумели выявить несколько транскрипционных факторов, которые, предположительно, отвечают за возрастные изменения транскрипции. Бóльшая часть генов-мишеней этих факторов задействована в функционировании иммунной системы и воспалении. Многие из транскрипционных факторов, отвечающих за возрастные изменения профилей транскрипции, оказались общими для печени и почек, но среди них есть и тканеспецифичные факторы. В частности, в почках многие транскрипционные факторы, изменяющие профили транскрипции с возрастом, так или иначе связаны с метаболизмом липидов и функционированием пероксисом.
Бóльшая часть генов-мишеней этих факторов задействована в функционировании иммунной системы и воспалении. Многие из транскрипционных факторов, отвечающих за возрастные изменения профилей транскрипции, оказались общими для печени и почек, но среди них есть и тканеспецифичные факторы. В частности, в почках многие транскрипционные факторы, изменяющие профили транскрипции с возрастом, так или иначе связаны с метаболизмом липидов и функционированием пероксисом.
Авторы работы впервые продемонстрировали, что многие гены, экспрессия которых понижается по мере старения организма, связаны с биосинтезом белка, метаболизмом некодирующих РНК и образованием рибосом. В их числе есть гены рибосомных белков, транскрипционных факторов, а также компонентов, задействованных в биогенезе рРНК и рибосом, процессинге некодирующих РНК и других процессах, имеющих ключевое значение для экспрессии генов и ее регуляции. Стоит отметить, что предыдущие исследования, рассматривавшие преимущественно только транскрипционные возрастные изменения, не могли выявить снижение синтеза этих белков, поскольку оно связано с подавлением трансляции, а не транскрипции.
Для того чтобы данные по трансляции с определенных мРНК можно было сравнивать не только с аналогичными данными для организма другого возраста, но и с данными по трансляции с других транскриптов, авторы работы использовали специальную нормированную метрику — «занятость» рибосом (RO от англ. ribosome occupancy). Для каждого гена RO вычисляется как количество рибосом, «пойманных» на синтезе его белкового продукта, деленное на общее количество транскриптов гена. Иными словами, RO позволяет понять, насколько интенсивно, по сравнению с другими генами, рибосомы взаимодействуют с мРНК данного гена и синтезируют белок по ее матрице (как правило, чем больше рибосом «сидят» на мРНК гена, тем больше белка с него синтезируется).
Анализ данных рибосомного профайлинга, переведенных в термины RO, показал, что с возрастом и в печени, и в почках увеличивается трансляция с мРНК, связанных с воспалением и дифференцировкой клеток различных типов. В то же время в почках с возрастом заметно снижается экспрессия генов, связанных с образованием и функционированием митохондрий, таких как гены компонентов электрон-транспортной цепи [12].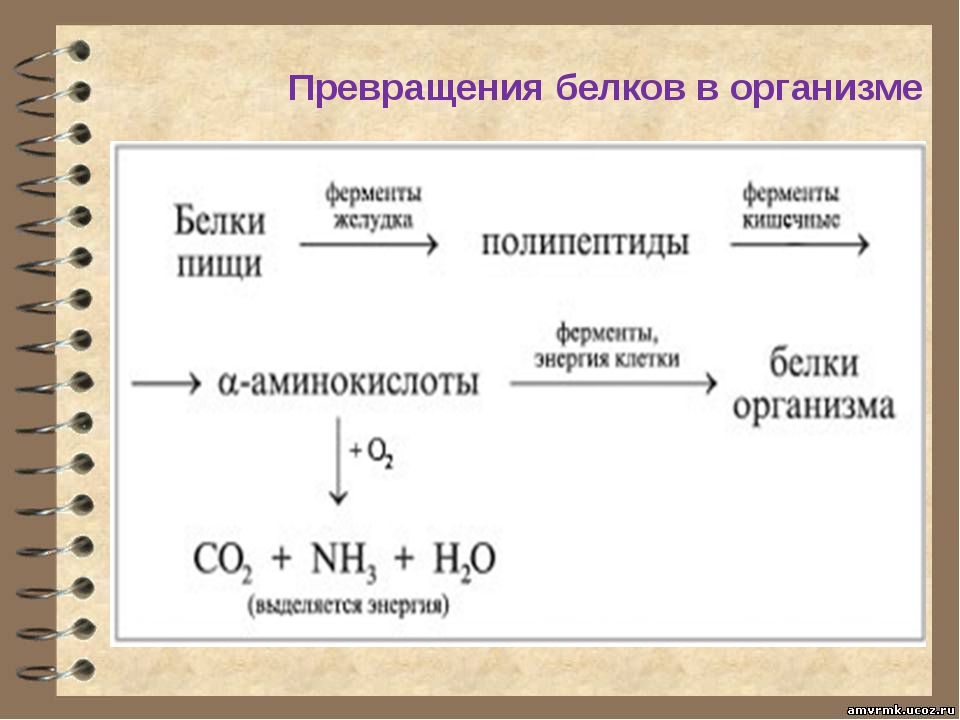 Примечательно, что трансляция многих ядерных генов, кодирующих белки митохондриального назначения, регулируется сигнальным путем mTOR, причем главную роль в этом процессе играет белок 4E-BP1. Этот же белок отвечает за возрастное снижение экспрессии гена интерферонового регуляторного транскрипционного фактора 7 (Irf7). С возрастом показатель RO для этого гена снижается почти в шесть раз. Стоит отметить, что по мере старения в печени увеличивается синтез субъединиц важнейшего регулятора метаболизма железа — ферритина [13], — трансляция которых зависит от особого железозависимого элемента в 5′-нетранслируемых областях их транскриптов. Вероятно, этот эффект связан с хорошо известными возрастными нарушениями метаболизма и запасания железа, которые зачастую являются последствиями возрастного хронического воспаления. Дело в том, что у грызунов по мере старения в тканях организма накапливается железо, а повышение его концентрации приводит к активации экспрессии гена ферритина и других генов, имеющих железозависимый элемент.
Примечательно, что трансляция многих ядерных генов, кодирующих белки митохондриального назначения, регулируется сигнальным путем mTOR, причем главную роль в этом процессе играет белок 4E-BP1. Этот же белок отвечает за возрастное снижение экспрессии гена интерферонового регуляторного транскрипционного фактора 7 (Irf7). С возрастом показатель RO для этого гена снижается почти в шесть раз. Стоит отметить, что по мере старения в печени увеличивается синтез субъединиц важнейшего регулятора метаболизма железа — ферритина [13], — трансляция которых зависит от особого железозависимого элемента в 5′-нетранслируемых областях их транскриптов. Вероятно, этот эффект связан с хорошо известными возрастными нарушениями метаболизма и запасания железа, которые зачастую являются последствиями возрастного хронического воспаления. Дело в том, что у грызунов по мере старения в тканях организма накапливается железо, а повышение его концентрации приводит к активации экспрессии гена ферритина и других генов, имеющих железозависимый элемент.
Сигнальный путь mTOR был упомянут выше неслучайно. Авторы исследования установили, что именно уменьшение активности этого сигнального пути по мере старения отвечает за возрастное снижение трансляции большой группы генов, которые объединяет наличие на 5′-конце особой олигопиримидиновой последовательности (5′-TOP от англ. 5′-terminal oligopyrimidine). Известно, что mTOR на уровне трансляции активирует экспрессию генов, продукты которых сами играют важнейшую роль в трансляции, поэтому снижение активности или ингибирование протеинкиназы mTOR подавляет интенсивность белкового синтеза в целом . Однако сильнее всего ингибирование mTOR сказывается именно на трансляции транскриптов с 5′-TOP. Анализ RO для таких mTOR-зависимых транскриптов показал, что с возрастом их синтез в печени падает сильнее всего по сравнению с остальными генами. В частности, существеннее всего при старении в печени «проседает» экспрессия генов, кодирующих некоторые рибосомные белки и фактор элонгации трансляции Eef2. У всех них в транскриптах имеется 5′-TOP, поэтому трансляция этих мРНК существенно зависит от сигнального пути mTOR. Похожие, хотя и менее выраженные изменения были выявлены в почках. Авторы работы полагают, что синтез рРНК и тРНК также претерпевает существенные возрастные изменения, поскольку он находится под сильным влиянием пути mTOR.
У всех них в транскриптах имеется 5′-TOP, поэтому трансляция этих мРНК существенно зависит от сигнального пути mTOR. Похожие, хотя и менее выраженные изменения были выявлены в почках. Авторы работы полагают, что синтез рРНК и тРНК также претерпевает существенные возрастные изменения, поскольку он находится под сильным влиянием пути mTOR.
Но самый неожиданный и любопытный эффект старения, который выявили авторы работы, состоит в перераспределении рибосом на транскриптах. Оказалось, что с возрастом частота встречаемости рибосом на 5′-концах транскриптов падает и увеличивается на 3′-концах как в печени, так и в почках. Для мышиной печени «точкой невозврата» является возраст 11 месяцев, после которого локализация рибосом становится сильно «перекошенной» в сторону 3′-конца. Особенно выражен эффект смещения рибосом при сравнении образцов тканей от мышей возрастом 11 месяцев и 32 месяца. Наиболее яркий эффект смещения рибосом — вблизи старт- и стоп-кодонов, но его можно пронаблюдать и по всей длине транскрипта.
Один из авторов работы, Сергей Дмитриев, комментирует ее результаты следующим образом: «Наверное, наиболее важным результатом нашей работы с точки зрения понимания механизмов старения является то, что мы показали существенное падение уровня синтеза компонентов трансляционного аппарата в органах стареющих мышей. Интенсивность наработки некоторых рибосомных белков в печени 32-месячных грызунов, например, оказалась в три раза ниже, чем у молодых животных. Причем это снижение происходит именно благодаря регуляции трансляции, поэтому до нас оно не было замечено. То, что белковый синтез у “пожилых” животных идет менее интенсивно, было известно и раньше — но теперь стало понятно, почему это происходит: в клетке просто производится меньше “машин” для биосинтеза белка. Интересный аспект всего этого — что в этой регуляции, по-видимому, замешана протеинкиназа mTOR, которую уже давно “подозревают в связях” со старением. Существуют фармакологические ингибиторы этой киназы (например, рапамицин), которые значимо продлевают жизнь многим модельным организмам.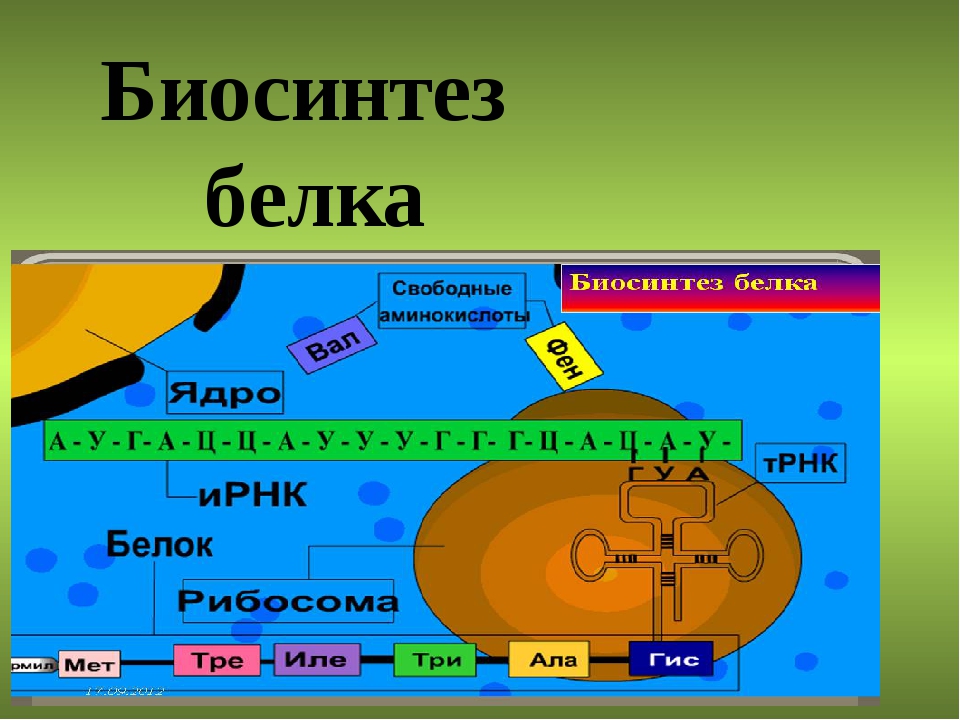 Правда, здесь мы сталкиваемся с парадоксом: с одной стороны, снижение активности белкового синтеза такими лекарствами продлевает жизнь, а с другой — пониженная активность, по нашим данным, характерна как раз для старых животных. Объяснением могло бы быть то, что падение скорости синтеза белка — это не причина старения, а следствие, попытка организма справиться с меняющимися условиями (накоплением в клетке всякого “мусора” и “поломок” — в частности, неправильных белков). Понятно, что когда в квартире не делают уборку, то можно прожить в ней чуточку дольше, если меньше мусорить. Именно это, по-видимому, делает стареющая клетка, и точно так же работают продлевающие жизнь ингибиторы mTOR. К сожалению, эта стратегия лишь отодвигает печальный финал: рано или поздно мусор погребет под собой постояльца. А как провести в клетке “генеральную уборку”, мы пока не придумали».
Правда, здесь мы сталкиваемся с парадоксом: с одной стороны, снижение активности белкового синтеза такими лекарствами продлевает жизнь, а с другой — пониженная активность, по нашим данным, характерна как раз для старых животных. Объяснением могло бы быть то, что падение скорости синтеза белка — это не причина старения, а следствие, попытка организма справиться с меняющимися условиями (накоплением в клетке всякого “мусора” и “поломок” — в частности, неправильных белков). Понятно, что когда в квартире не делают уборку, то можно прожить в ней чуточку дольше, если меньше мусорить. Именно это, по-видимому, делает стареющая клетка, и точно так же работают продлевающие жизнь ингибиторы mTOR. К сожалению, эта стратегия лишь отодвигает печальный финал: рано или поздно мусор погребет под собой постояльца. А как провести в клетке “генеральную уборку”, мы пока не придумали».
- Aleksandra S. Anisimova, Mark B. Meerson, Maxim V. Gerashchenko, Ivan V. Kulakovskiy, Sergey E. Dmitriev, Vadim N.
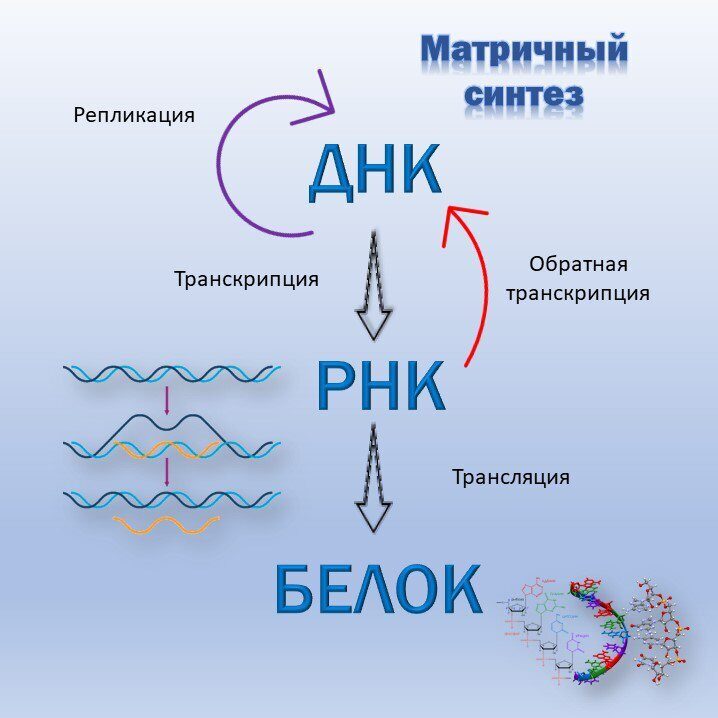 Gladyshev. (2020). Multifaceted deregulation of gene expression and protein synthesis with age. Proc Natl Acad Sci USA. 117, 15581-15590;
Gladyshev. (2020). Multifaceted deregulation of gene expression and protein synthesis with age. Proc Natl Acad Sci USA. 117, 15581-15590;
- Преодолевшие старение. Часть II. Дети подземелья;
- Модельные организмы: нематода;
- 12 методов в картинках: генная инженерия. Часть II: инструменты и техники;
- Старение: остановить нельзя смириться;
- Старение и долголетие: эпигеном раскрывает тайны;
- Почему мы не боимся смерти? Первый шаг к победе над старением;
- Рибосома за работой;
- Чего вы не знаете о рибосоме…;
- Кодирующие некодирующие РНК;
- Длинные некодирующие РНК бактерий;
- Болезни и изменения клеточного метаболизма;
- Jinze Xu, Zhenhua Jia, Mitchell D. Knutson, Christiaan Leeuwenburgh.
 (2012). Impaired Iron Status in Aging Research. IJMS. 13, 2368-2386;
(2012). Impaired Iron Status in Aging Research. IJMS. 13, 2368-2386;
- Аутофагия, протофагия и остальные.
Урок №96. Белки — ХиМуЛя
Белки
в природе
Белки – это молекулы жизни. Каждый живой
организм содержит большое количество различных белковых молекул, при этом
каждому виду присущи особые, свойственные только ему белки. Даже белки,
выполняющие у различных видов одну и ту же функцию, отличаются друг от друга.
Например, у всех позвоночных животных – рыб, птиц, млекопитающих – красные
клетки крови содержат белок гемоглобин, переносящий кислород. Но гемоглобин у
каждого вида животных свой, особенный. Молекула гемоглобина лошади отличается
от соответствующего белка человека в 26 местах, свиньи – в 10 местах, а гориллы
– всего лишь одной аминокислотой.
Функции белков в организме очень
разнообразны. Есть белки – переносчики веществ (молекул, ионов) и электронов;
есть биокатализаторы, ускоряющие реакции в миллиарды раз и отличающиеся
удивительной специфичностью, есть регуляторы различных биологических процессов
в организме – гормоны, например, инсулин, вазопрессин, окситоцин.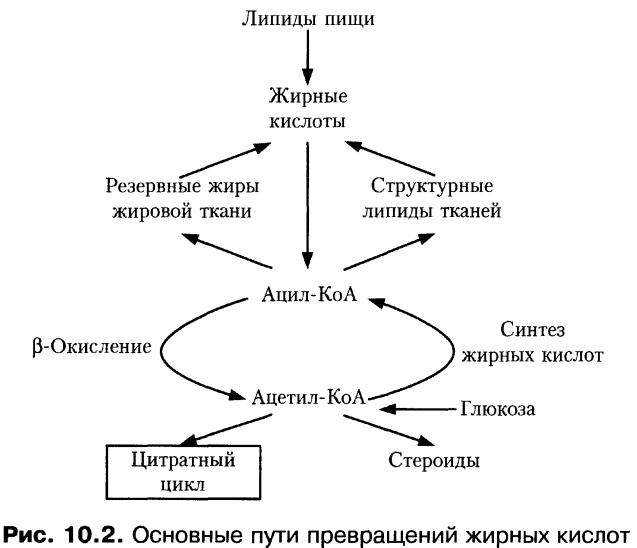 Белки
Белки
защищают организм от инфекции, они способны узнавать и уничтожать чужеродные
объекты: вирусы, бактерии, клетки. Контакты клетки с внешней средой также
выполняют разнообразные белки, умеющие различать форму молекул, регистрировать
изменение температуры, ничтожные примеси веществ, отличать один цвет от другого.
Свойства
белков
Свойства белков весьма разнообразны и
определяются их строением.
1. По растворимости в воде белки делятся
на два класса:
глобулярные белки –
растворяются в воде или образуют коллоидные растворы; фибриллярные белки
– в воде нерастворимы.
2. Денатурация. При нагревании, изменении
кислотности среды происходит разрушение вторичной и третичной структуры белка с
сохранением первичной. Это явление называют денатурацией.
Пример
денатурации
– свертывание яичных белков при варке яиц. Денатурация бывает обратимой
(при употреблении алкоголя,
солёной пищи) и необратимой.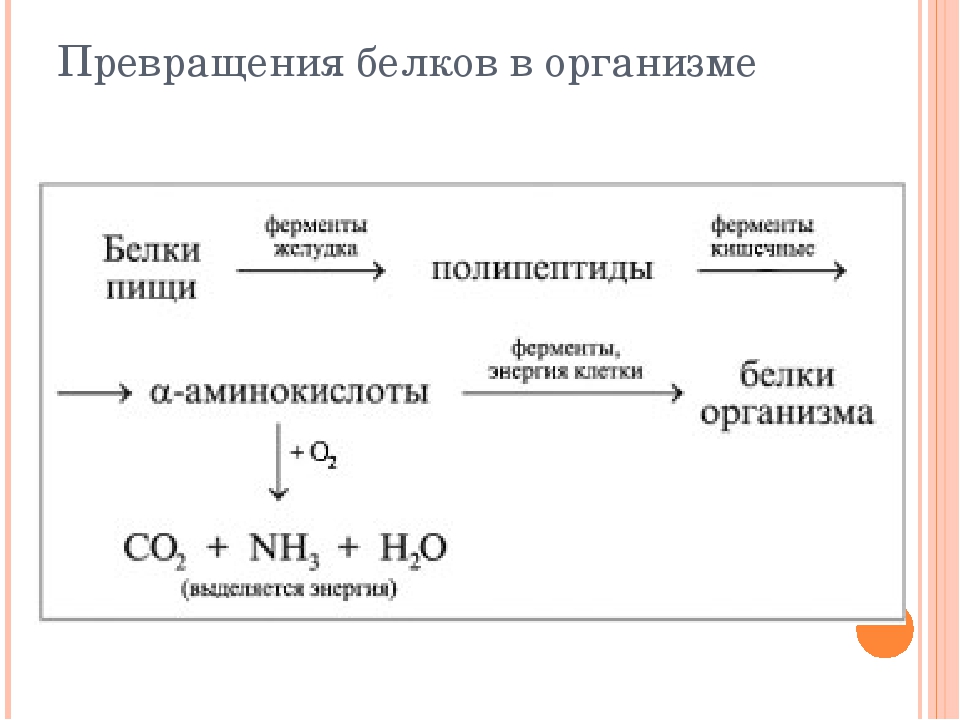 Необратимая денатурация может быть
Необратимая денатурация может быть
вызвана высокими температурами, радиацией,
при отравлении организма солями тяжелых металлов, спиртами, кислотами.
ВИДЕО:
Свертывание белков при нагревании
Осаждение белков солями тяжелых металлов
Осаждение белков спиртом
3. Гидролиз белков – это
необратимое разрушение первичной структуры в кислом или щелочном растворе с
образованием аминокислот. Анализируя продукты гидролиза, можно установить
количественный состав белков.
4.
Для белков известно несколько качественных реакций.
1.
Все
соединения, содержащие пептидную связь, дают фиолетовое окрашивание при
действии на них солей меди (II) в щелочном растворе. Эта реакция
называется биуретовой.
ВИДЕО:
ВИДЕО:
Биуретовая реакция белков
2.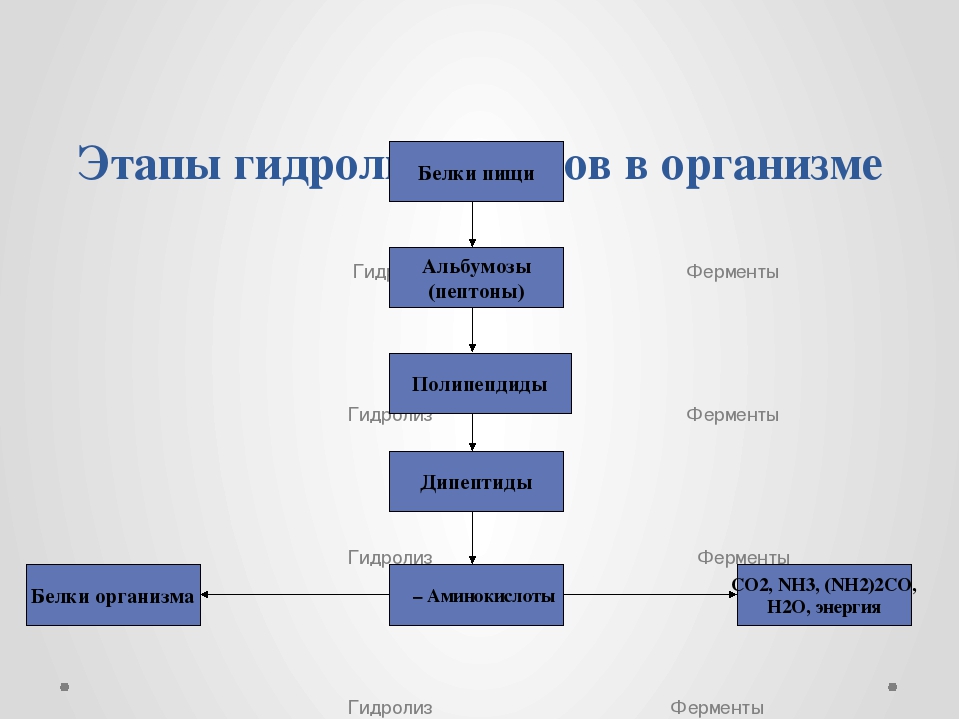
Белки,
содержащие остатки ароматических аминокислот (фенилаланин, тирозин) дают желтое
окрашивание при действии концентрированной азотной кислоты – ксантопротеиновая
реакция.
ВИДЕО:
ВИДЕО:
Качественные реакции на белки: биуретовая и ксантопротеиновая
Ксантопротеиновая реакция белков
Обнаружение серы
5. Амфотерные свойства белков
Очень важным для жизнедеятельности живых
организмов является буферное свойство белков, т.е. способность связывать как
кислоты, так и основания, и поддерживать постоянное значение рН различных систем
живого организма.
Превращение
белков в организме
Животные организмы строят свои белки из
аминокислот тех белков, которые они получают с пищей. Поэтому наряду с жирами и
углеводами белки – обязательный компонент нашей пищи.
Животные и растительные белки в
пищеварительном тракте человека расщепляются на аминокислоты. В процессе
В процессе
переваривания пищи происходит гидролиз белков под влиянием ферментов. В желудке
они расщепляются на более или менее крупные «осколки» – пептиды, которые далее
в кишечнике гидролизуются до аминокислот. Последние всасываются ворсинками
кишечника в кровь и поступают во все ткани и клетки организма. Здесь из
аминокислот под действием ферментов синтезируются белки, свойственные тканям
человеческого тела. Для синтезирования белков необходимо наличие определенных
аминокислот. Но в одних белках, поступающих с пищей, имеются все необходимые
человеку аминокислоты, а в других не все. Организм человека может сам
синтезировать некоторые аминокислоты или заменять их другими. Но 10 аминокислот
он образовать не в состоянии. Их должен непременно
получать с пищей. Эти кислоты называются
незаменимыми (см. Приложения).
Белки, содержащие все необходимые
аминокислоты, называют полноценными. Остальные
белки — неполноценные. Полноценными являются белки
молока, сыра, мяса, рыбы, яиц, бобовых.
Синтезом белков в клетках управляет ДНК. Он осуществляется на поверхности
рибосом с помощью РНК. В организме человека белки почти не откладываются в
запас. Излишки аминокислот в клетках печени превращаются в углеводы — глюкозу и
гликоген или в резервный жир. Поэтому артистам балета слишком больших количеств
белков в пище нужно избегать. Но и намеренное голодание, когда вследствие
больших энерготрат организм, израсходовав запасы углеводов и жира, начинает
тратить резервы белка, очень вредно. Это тратятся белки цитоплазмы клеток.
Судьба аминокислот в организме различна
1. Основная их масса расходуется на
синтез белков, которые идут на увеличение белковой массы организма при его
росте и на обновление белков, распадающихся в процессе жизнедеятельности.
2. Синтез белков идет с поглощением
энергии.
3. Аминокислоты используются в организме
и для синтеза небелковых азотсодержащих соединений, например нуклеиновых
кислот.
4.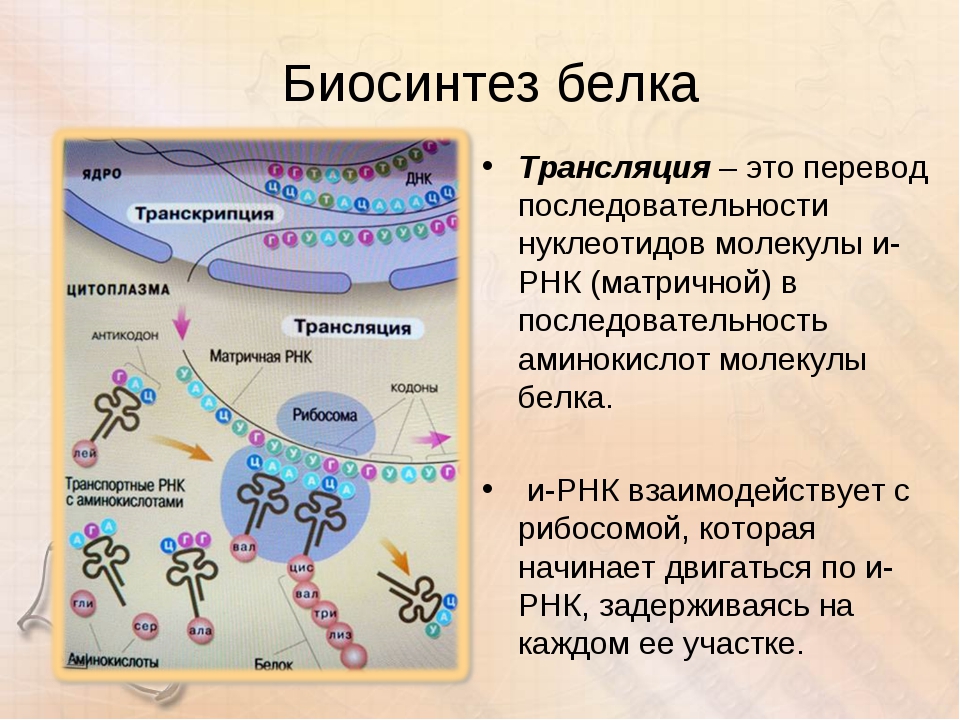 Часть аминокислот подвергается
Часть аминокислот подвергается
постепенному распаду и окислению.
Успехи
в изучении и синтезе белков
Основные сведения о составе и строении
белков были получены при изучении их гидролиза (гидролиз белков – необратимое
разрушение первичной структуры в кислом или щелочном растворе с образованием
аминокислот). Установлено, что в результате гидролиза любого белка получается
смесь α-аминокислот, причем наиболее часто встречаются в составе белков 20 α-аминокислот.
Как же аминокислоты образуют белковую
молекулу? Еще в 80-х годах прошлого века русский ученый-биохимик А.Я. Данилевский на основании своих
опытов впервые высказал гипотезу о пептидной связи между остатками аминокислот
в белковой молекуле. В 1899 году исследованиями белков занялись немецкие
химики-органики Эмиль Фишер и Франц Гофмейстер. Они высказали
предположение, что в белках аминокислоты связаны за счет аминогруппы одной
кислоты и карбоксила другой. При образовании такой связи выделяется молекула
воды. Эта гипотеза была блестяще подтверждена экспериментально в 1907 году и
Эта гипотеза была блестяще подтверждена экспериментально в 1907 году и
получила название “полипептидной теории”.
Фишеру удалось синтетически получить
полипептиды, в молекулы которых входили различные аминокислотные остатки,
соединенные пептидными связями.
Химический синтез широко применяют для
получения пептидов, в т.ч. биологически активных гормонов и их разнообразных
аналогов, используемых для изучения взаимосвязи структуры и биологической
функции, а также пептидов, несущих антигенные детерминанты различных белков и
применяемых для приготовления соответствующих вакцин. Первые химические синтезы
белка в 60-е гг. (инсулина овцы и рибонуклеазы S), осуществленные в растворе с
помощью тех же методов, которые используют при синтезе пептидов, были связаны с
чрезвычайно большими сложностями. В каждом случае требовалось провести сотни
химических реакций и окончательный выход белка был очень низок (менее 0,1%), в
результате чего полученные препараты не удалось очистить. Позже были
синтезированы некоторые химически чистые белки, в частности инсулин человека
(П. Зибер и др.) и нейротоксин II из ядра среднеазиатской кобры (В.Т. Иванов).
Зибер и др.) и нейротоксин II из ядра среднеазиатской кобры (В.Т. Иванов).
Однако до сих пор химический синтез белка представляет весьма сложную проблему
и имеет скорее теоретическое, чем практическое значение. Более перспективны
методы генетической инженерии, которые позволяют наладить промышленное
получение практически важных белков и пептидов.
Упрощенный синтез полипептидов можно
представить так:
Вспомните: связь между остатками аминокислот,
а именно: между группами С = О одной кислоты и N-H другой кислоты – называется
пептидной (амидной), группа атомов –СО─NH ─ называется пептидной (амидной)
группой.
Пептидная или белковая цепь представляет
собой продукт поликонденсации аминокислот. Один из концов цепи, где находится
остаток аминокислоты со свободной аминогруппой, называется N-концом, сама
аминокислота – N-концевой; другой конец цепи с остатком аминокислоты, имеющим
карбоксильную группу, называется С-концом, кислота – С-концевой.
Пептидную цепь всегда записывают,
начиная с N-конца. В названии пептида за основу принимают С-концевую кислоту,
В названии пептида за основу принимают С-концевую кислоту,
остальные аминокислоты указывают как заместители с суффиксом –ил-, перечисляя
их последовательно, начиная с N-конца. Название полученного дипептида: ГЛИЦИЛАЛАНИН
ВИДЕО:
Качественное определение азота в органических соединениях
ПРАКТИКУМ
Карта сайта
Страница не найдена. Возможно, карта сайта Вам поможет.
-
Главная -
Университет-
Об университете -
Структура -
Нормативные документы и процедуры -
Лечебная деятельность -
Международное сотрудничество -
Пресс-центр-
Новости -
Анонсы -
События -
Объявления и поздравления -
Online конференции -
Фотоальбом-
Предупреждение деструктивных проявлений в студенческой среде и влияния агрессивного информационного контента сети интернет -
Онлайн-выставка «Помнить, чтобы не повторить» -
Областная межвузовская конференция «Подвиг народа бессмертен» -
Финал первого Республиканского интеллектуального турнира ScienceQuiz -
Конференция «Актуальные вопросы коморбидности заболеваний в амбулаторной практике: от профилактики до лечения» -
День семьи-2021 -
Диалоговая площадка с председателем Гродненского областного Совета депутатов -
Праздничные городские мероприятия к Дню Победы -
Областной этап конкурса «Королева студенчества-2021″ -
Праздничный концерт к 9 мая 2021 -
IV Республиканский гражданско-патриотический марафон «Вместе – за сильную и процветающую Беларусь!» -
Университетский кубок КВН-2021 -
Музыкальная планета студенчества (завершение Дней ФИУ-2021) -
Молодёжный круглый стол «Мы разные, но мы вместе» -
Дни ФИУ-2021.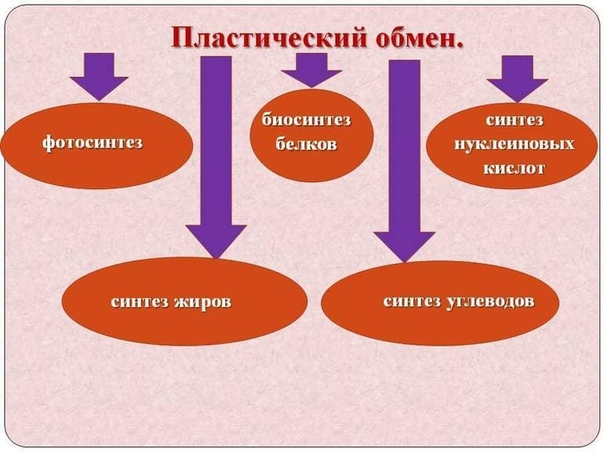 Интеллектуальная игра «Что?Где?Когда?»
Интеллектуальная игра «Что?Где?Когда?»
-
Неделя донорства в ГрГМУ -
Творческая гостиная. Дни ФИУ-2021 -
Открытие XVIII студенческого фестиваля национальных культур -
Передвижная мультимедийная выставка «Партизаны Беларуси» -
Республиканский субботник-2021 -
Семинар «Человек внутри себя» -
Международный конкурс «Здоровый образ жизни глазами разных поколений» -
Вручение нагрудного знака «Жена пограничника» -
Встреча с представителями медуниверситета г. Люблина -
Королева Студенчества ГрГМУ — 2021 -
День открытых дверей-2021 -
Управление личными финансами (встреча с представителями «БПС-Сбербанк») -
Весенний «Мелотрек» -
Праздничный концерт к 8 Марта -
Диалоговая площадка с председателем Гродненского облисполкома -
Расширенное заседание совета университета -
Гродно — Молодежная столица Республики Беларусь-2021 -
Торжественное собрание, приуроченное к Дню защитника Отечества -
Вручение свидетельства действительного члена Белорусской торгово-промышленной палаты -
Новогодний ScienceQuiz -
Финал IV Турнира трех вузов ScienseQuiz -
Областной этап конкурса «Студент года-2020″ -
Семинар дистанционного обучения для сотрудников университетов из Беларуси «Обеспечение качества медицинского образования и образования в области общественного здоровья и здравоохранения» -
Студент года — 2020 -
День Знаний — 2020 -
Церемония награждения лауреатов Премии Правительства в области качества -
Военная присяга -
Выпускной лечебного факультета-2020 -
Выпускной медико-психологического факультета-2020 -
Выпускной педиатрического факультета-2020 -
Выпускной факультета иностранных учащихся-2020 -
Распределение — 2020 -
Стоп коронавирус! -
Навстречу весне — 2020 -
Профориентация — 18-я Международная специализированная выставка «Образование и карьера» -
Спартакиада среди сотрудников «Здоровье-2020″ -
Конференция «Актуальные проблемы медицины» -
Открытие общежития №4 -
Встреча Президента Беларуси со студентами и преподавателями медвузов -
Новогодний утренник в ГрГМУ -
XIX Республиканская студенческая конференция «Язык.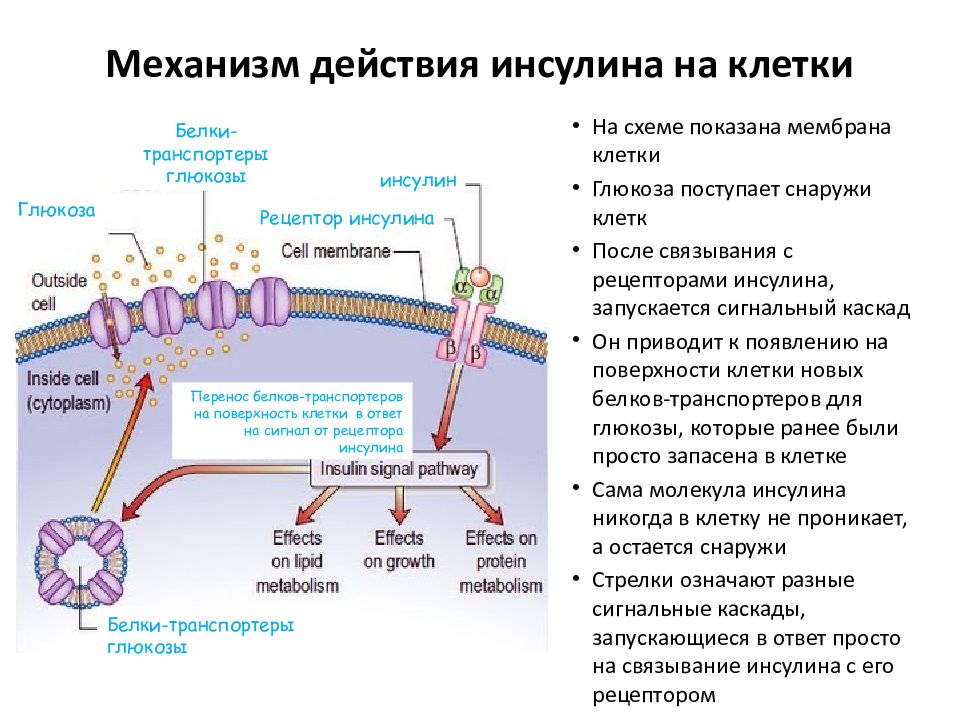 Общество. Медицина»
Общество. Медицина»
-
Alma mater – любовь с первого курса -
Актуальные вопросы коморбидности сердечно-сосудистых и костно-мышечных заболеваний в амбулаторной практике -
Областной этап «Студент года-2019″ -
Финал Science Qiuz -
Конференция «Актуальные проблемы психологии личности и социального взаимодействия» -
Посвящение в студенты ФИУ -
День Матери -
День открытых дверей — 2019 -
Визит в Азербайджанский медицинский университет -
Семинар-тренинг с международным участием «Современные аспекты сестринского образования» -
Осенний легкоатлетический кросс — 2019 -
40 лет педиатрическому факультету -
День Знаний — 2019 -
Посвящение в первокурсники -
Акция к Всемирному дню предотвращения суицида -
Турслет-2019 -
Договор о создании филиала кафедры общей хирургии на базе Брестской областной больницы -
День Независимости -
Конференция «Современные технологии диагностики, терапии и реабилитации в пульмонологии» -
Выпускной медико-диагностического, педиатрического факультетов и факультета иностранных учащихся — 2019 -
Выпускной медико-психологического факультета — 2019 -
Выпускной лечебного факультета — 2019 -
В добрый путь, выпускники! -
Распределение по профилям субординатуры -
Государственные экзамены -
Интеллектуальная игра «Что? Где? Когда?» -
Мистер и Мисс факультета иностранных учащихся-2019 -
День Победы -
IV Республиканская студенческая военно-научная конференция «Этих дней не смолкнет слава» -
Республиканский гражданско-патриотический марафон «Вместе — за сильную и процветающую Беларусь!» -
Литературно-художественный марафон «На хвалях спадчыны маёй» -
День открытых дверей-2019 -
Их имена останутся в наших сердцах -
Областной этап конкурса «Королева Весна — 2019″ -
Королева Весна ГрГМУ — 2019 -
Профориентация «Абитуриент – 2019» (г. Барановичи)
Барановичи)
-
Мероприятие «Карьера начинается с образования!» (г. Лида) -
Итоговое распределение выпускников — 2019 -
«Навстречу весне — 2019″ -
Торжественная церемония, посвященная Дню защитника Отечества -
Торжественное собрание к Дню защитника Отечества — 2019 -
Мистер ГрГМУ — 2019 -
Предварительное распределение выпускников 2019 года -
Митинг-реквием у памятника воинам-интернационалистам -
Профориентация «Образование и карьера» (г.Минск) -
Итоговая коллегия главного управления здравоохранения Гродненского областного исполнительного комитета -
Спартакиада «Здоровье — 2019» -
Итоговая научно-практическая конференция «Актуальные проблемы медицины».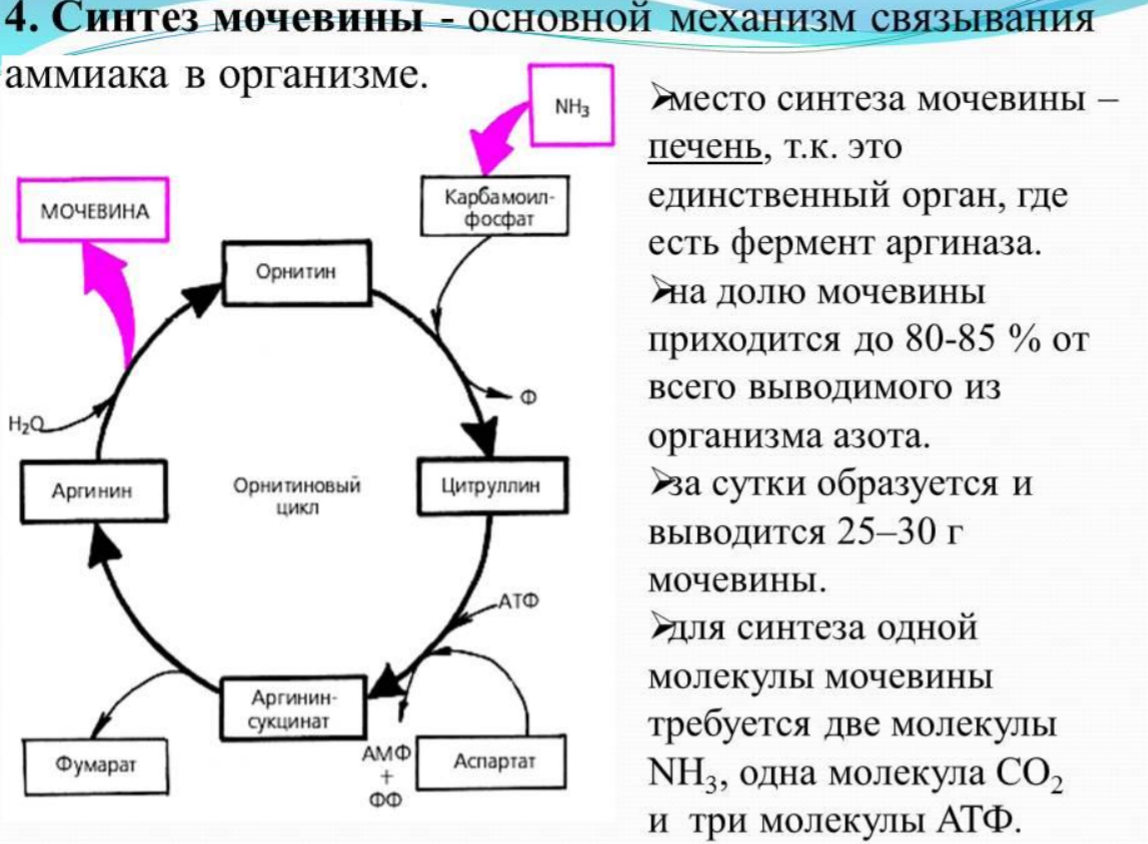
-
Расширенное заседание Совета университета. -
Научно-практическая конференция «Симуляционные технологии обучения в подготовке медицинских работников: актуальность, проблемные вопросы внедрения и перспективы» -
Конкурс первокурсников «Аlma mater – любовь с первого курса» -
XVI съезд хирургов Республики Беларусь -
Итоговая практика -
Конкурс «Студент года-2018» -
Совет университета -
1-й съезд Евразийской Аритмологической Ассоциации (14.09.2018 г.) -
1-й съезд Евразийской Аритмологической Ассоциации (13.09.2018 г.) -
День знаний -
День независимости Республики Беларусь -
Церемония награждения победителей конкурса на соискание Премии СНГ -
День герба и флага Республики Беларусь -
«Стань донором – подари возможность жить» -
VIII Международный межвузовский фестиваль современного танца «Сделай шаг вперед» -
Конкурс грации и артистического мастерства «Королева Весна ГрГМУ – 2018» -
Окончательное распределение выпускников 2018 года -
Митинг-реквием, приуроченный к 75-летию хатынской трагедии -
Областное совещание «Итоги работы терапевтической и кардиологической служб Гродненской области за 2017 год и задачи на 2018 год» -
Конкурсное шоу-представление «Мистер ГрГМУ-2018» -
Предварительное распределение выпускников 2018 года -
Итоговая научно-практическая конференция «Актуальные проблемы медицины» -
II Съезд учёных Республики Беларусь -
Круглый стол факультета иностранных учащихся -
«Молодежь мира: самобытность, солидарность, сотрудничество» -
Заседание выездной сессии Гродненского областного Совета депутатов -
Областной этап республиканского конкурса «Студент года-2017» -
Встреча с председателем РОО «Белая Русь» Александром Михайловичем Радьковым -
Конференция «Актуальные вопросы инфекционной патологии», 27.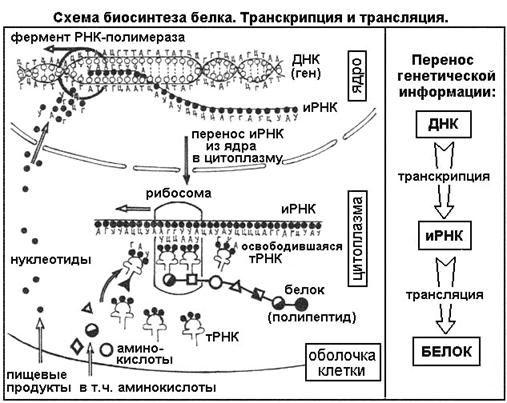 10.2017
10.2017
-
XIX Всемирный фестиваль студентов и молодежи -
Республиканская научно-практическая конференция «II Гродненские аритмологические чтения» -
Областная научно-практическая конференция «V Гродненские гастроэнтерологические чтения» -
Праздник, посвящённый 889-летию города Гродно -
Круглый стол на тему «Место и роль РОО «Белая Русь» в политической системе Республики Беларусь» (22.09.2017) -
ГрГМУ и Университет медицины и фармации (г.Тыргу-Муреш, Румыния) подписали Соглашение о сотрудничестве -
1 сентября — День знаний -
Итоговая практика на кафедре военной и экстремальной медицины -
Квалификационный экзамен у врачей-интернов -
Встреча с Комиссией по присуждению Премии Правительства Республики Беларусь -
Научно-практическая конференция «Амбулаторная терапия и хирургия заболеваний ЛОР-органов и сопряженной патологии других органов и систем» -
День государственного флага и герба -
9 мая -
Республиканская научно-практическая конференция с международным участием «V белорусско-польская дерматологическая конференция: дерматология без границ» -
«Стань донором – подари возможность жить» -
«Круглый стол» Постоянной комиссии Совета Республики Беларусь Национального собрания Республики Беларусь по образованию, науке, культуре и социальному развитию -
Весенний кубок КВН «Юмор–это наука» -
Мисс ГрГМУ-2017 -
Распределение 2017 года -
Общегородской профориентационный день для учащихся гимназий, лицеев и школ -
Праздничный концерт, посвященный Дню 8 марта -
Конкурсное шоу-представление «Мистер ГрГМУ–2017» -
«Масленица-2017» -
Торжественное собрание и паздничный концерт, посвященный Дню защитника Отечества -
Лекция профессора, д. м.н. О.О. Руммо
м.н. О.О. Руммо
-
Итоговая научно-практическая конференция «Актуальные проблемы медицины» -
Меморандум о сотрудничестве между областной организацией Белорусского общества Красного Креста и региональной организацией Красного Креста китайской провинции Хэнань -
Визит делегации МГЭУ им. А.Д. Сахарова БГУ в ГрГМУ -
«Студент года-2016» -
Визит Чрезвычайного и Полномочного Посла Королевства Швеция в Республике Беларусь господина Мартина Оберга в ГрГМУ -
Конкурс первокурсников «Аlma mater – любовь с первого курса» -
День матери в ГрГМУ -
Итоговая практика-2016 -
День знаний -
Визит китайской делегации в ГрГМУ -
Визит иностранной делегации из Вроцлавского медицинского университета (Республика Польша) -
Торжественное мероприятие, посвященное профессиональному празднику – Дню медицинского работника -
Визит ректора ГрГМУ Виктора Александровича Снежицкого в Индию -
Республиканская университетская суббота-2016 -
Республиканская акция «Беларусь против табака» -
Встреча с поэтессой Яниной Бокий -
9 мая — День Победы -
Митинг, посвященный Дню Государственного герба и Государственного флага Республики Беларусь -
Областная межвузовская студенческая научно-практическая конференция «1941 год: трагедия, героизм, память» -
«Цветы Великой Победы» -
Концерт народного ансамбля польской песни и танца «Хабры» -
Суботнiк ў Мураванцы -
«Мисс ГрГМУ-2016» -
Визит академика РАМН, профессора Разумова Александра Николаевича в УО «ГрГМУ» -
Визит иностранной делегации из Медицинского совета Мальдивской Республики -
«Кубок ректора Гродненского государственного медицинского университета по дзюдо» -
«Кубок Дружбы-2016» по мини-футболу среди мужских и женских команд медицинских учреждений образования Республики Беларусь -
Распределение выпускников 2016 года -
Визит Министра обороны Республики Беларусь на военную кафедру ГрГМУ -
Визит Первого секретаря Посольства Израиля Анны Кейнан и директора Израильского культурного центра при Посольстве Израиля Рей Кейнан -
Визит иностранной делегации из провинции Ганьсу Китайской Народной Республики в ГрГМУ -
Состоялось открытие фотовыставки «По следам Библии» -
«Кубок декана» медико-диагностического факультета по скалолазанию -
Мистер ГрГМУ-2016 -
Приём Первого секретаря Посольства Израиля Анны Кейнан в ГрГМУ -
Спартакиада «Здоровье» УО «ГрГМУ» среди сотрудников 2015-2016 учебного года -
Визит Посла Республики Индия в УО «ГрГМУ» -
Торжественное собрание и концерт, посвященный Дню защитника Отечества -
Митинг-реквием, посвященный Дню памяти воинов-интернационалистов -
Итоговое заседание коллегии главного управления идеологической работы, культуры и по делам молодежи Гродненского облисполкома -
Итоговая научно-практическая конференция Гродненского государственного медицинского университета -
Новогодний концерт -
Открытие профессорского консультативного центра -
Концерт-акция «Молодёжь против СПИДа» -
«Студент года-2015» -
Открытые лекции профессора, академика НАН Беларуси Островского Юрия Петровича -
«Аlma mater – любовь с первого курса» -
Открытая лекция Регионального директора ВОЗ госпожи Жужанны Якаб -
«Открытый Кубок по велоориентированию РЦФВиС» -
Совместное заседание Советов университетов г.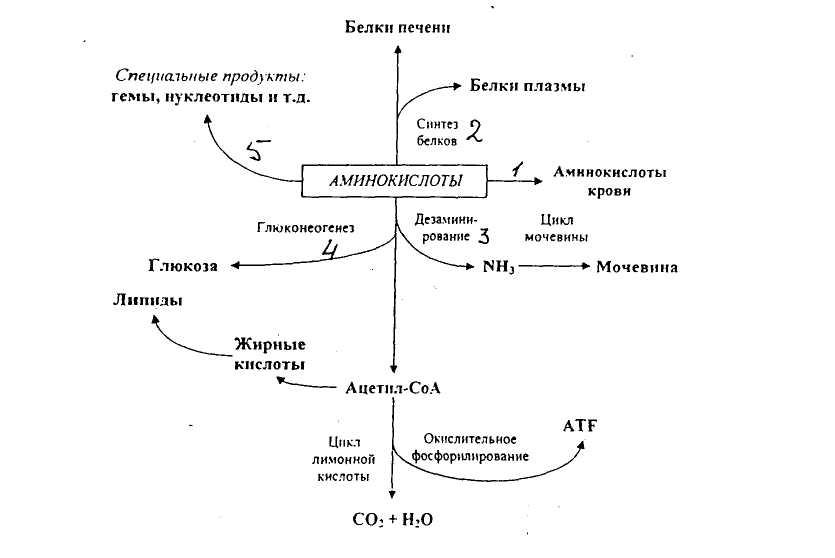 Гродно
Гродно
-
Встреча с Министром здравоохранения Республики Беларусь В.И. Жарко -
День города -
Дебаты «Врач — выбор жизни» -
День города -
Праздничный концерт «Для вас, первокурсники!» -
Акция «Наш год – наш выбор» -
День знаний -
Открытое зачисление абитуриентов в УО «Гродненский государственный медицинский университет» -
Принятие военной присяги студентами ГрГМУ -
День Независимости Республики Беларусь -
Вручение дипломов выпускникам 2015 года -
Республиканская олимпиада студентов по педиатрии -
Открытие памятного знака в честь погибших защитников -
9 мая -
«Вторая белорусско-польская дерматологическая конференция: дерматология без границ» -
Мистер университет -
Мисс универитет -
КВН -
Гродненский государственный медицинский университет -
Чествование наших ветеранов -
1 Мая -
Cовместный субботник
-
-
Наши издания -
Медицинский календарь -
Университет в СМИ -
Видео-презентации
-
-
Общественные объединения -
Комиссия по противодействию коррупции -
Образовательная деятельность
-
-
Абитуриентам -
Студентам -
Выпускникам -
Слайдер -
Последние обновления -
Баннеры -
Иностранному гражданину -
Научная деятельность -
Поиск
Синтез белка Определение и примеры
Синтез белка
n. , Множественное число: синтез белка
, Множественное число: синтез белка
Определение: создание белка.
Синтез белка — это процесс создания белковых молекул. В биологических системах это включает синтез аминокислот, транскрипцию, трансляцию и посттрансляционные события. В синтезе аминокислот есть набор биохимических процессов, которые производят аминокислоты из источников углерода, таких как глюкоза.Не все аминокислоты производятся организмом; другие аминокислоты получают с пищей. В клетках генерируются белки с участием процессов транскрипции и трансляции. Короче говоря, транскрипция — это процесс, с помощью которого матрица мРНК транскрибируется из ДНК. Шаблон используется для следующего шага — перевода. При трансляции аминокислоты связаны друг с другом в определенном порядке, основанном на генетическом коде. После трансляции вновь образованный белок подвергается дальнейшей обработке, такой как протеолиз, посттрансляционная модификация и сворачивание белка.
Определение синтеза белка
Синтез белка — схематическая диаграмма Синтез белка — это создание белков. В биологических системах это осуществляется внутри клетки. У прокариот это происходит в цитоплазме. У эукариот это первоначально происходит в ядре, чтобы создать транскрипт (мРНК) кодирующей области ДНК. Транскрипт покидает ядро и достигает рибосом для трансляции в молекулу белка с определенной последовательностью аминокислот.
В биологических системах это осуществляется внутри клетки. У прокариот это происходит в цитоплазме. У эукариот это первоначально происходит в ядре, чтобы создать транскрипт (мРНК) кодирующей области ДНК. Транскрипт покидает ядро и достигает рибосом для трансляции в молекулу белка с определенной последовательностью аминокислот.
Синтез белка — это создание белков клетками с использованием ДНК, РНК и различных ферментов.Обычно он включает события транскрипции, трансляции и посттрансляционные события, такие как сворачивание белка, модификации и протеолиз.
Этимология
Термин белок произошел от позднегреческого prōteios , prōtos , что означает «первый». Слово синтез произошло от греческого sunthesis , от suntithenai , что означает «соединять». Вариант: биосинтез белка.
Прокариотический синтез в сравнении с синтезом эукариотических белков
Белки являются основным типом биомолекул, которые необходимы всем живым существам для процветания.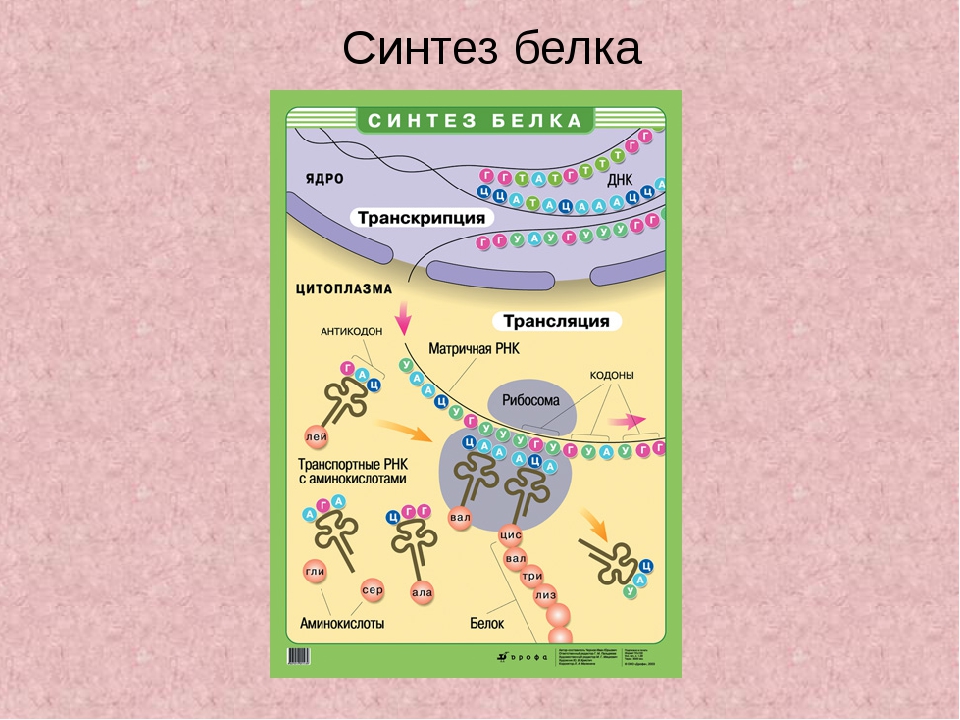 И прокариоты, и эукариоты производят различные белки для различных процессов и функций. Некоторые белки используются для структурных целей, а другие действуют как катализаторы биохимических реакций. Синтезы прокариотических и эукариотических белков имеют явные различия. Например, синтез белка у прокариот происходит в цитоплазме. У эукариот первый этап (транскрипция) происходит в ядре. Когда транскрипт (мРНК) образуется, он попадает в цитоплазму, где расположены рибосомы.Здесь мРНК транслируется в аминокислотную цепь. В таблице ниже показаны различия между синтезом прокариотических и эукариотических белков.
И прокариоты, и эукариоты производят различные белки для различных процессов и функций. Некоторые белки используются для структурных целей, а другие действуют как катализаторы биохимических реакций. Синтезы прокариотических и эукариотических белков имеют явные различия. Например, синтез белка у прокариот происходит в цитоплазме. У эукариот первый этап (транскрипция) происходит в ядре. Когда транскрипт (мРНК) образуется, он попадает в цитоплазму, где расположены рибосомы.Здесь мРНК транслируется в аминокислотную цепь. В таблице ниже показаны различия между синтезом прокариотических и эукариотических белков.
| Синтез прокариотического белка | Синтез эукариотического белка |
|---|---|
| Трансляция происходит еще до транскрипции концов мРНК | Транскрипция происходит с последующей трансляцией |
| За исключением архебактерий, образование мРНК бактерий не включает добавление cap и поли A-хвоста | Образование мРНК включает добавление 5′-кэпа и поли-A-хвоста на 3′-конце транскрипта мРНК |
| Трансляция начинается с кодона AUG | Трансляция начинается через 5 ‘Cap, связывающая мРНК с рибосомной единицей в первом кодоне AUG |
| Инициирующие факторы: PIF-1, PIF-2, PIF-3 | Инициирующие факторы: eIF1-6, eIF4B, eIF4C, eIF4D, eIF4F |
Генетический код
Аминокислотная таблица кодонов РНК.
В биологии кодон относится к тринуклеотидам, которые указаны для конкретной аминокислоты. Например, гуанин-цитозин-цитозин (GCC) кодирует аминокислоту аланин. Коды гуанин-урацил-урацил (GUU) для валина. Урацил-аденин-аденин (UAA) является стоп-кодоном. Кодон мРНК дополняет тринуклеотид (называемый антикодоном) в тРНК.
мРНК, тРНК и рРНК
мРНК, тРНК и рРНК — три основных типа РНК, участвующих в синтезе белка. МРНК (или информационная РНК) несет код для создания белка.У эукариот он образуется внутри ядра и состоит из 5′-кэпа, 5’UTR-области, кодирующей области, 3’UTR-области и поли (A) хвоста. Копия сегмента ДНК для экспрессии гена находится в его кодирующей области. Он начинается со стартового кодона на 5′-конце и стоп-кодона на 3′-конце.
тРНК (или транспортная РНК), как следует из названия, переносит определенную аминокислоту на рибосому, которая будет добавлена к растущей цепи аминокислоты. Он состоит из двух основных сайтов: (1) антикодонное плечо и (2) акцепторный стержень . Плечо антикодона содержит антикодон, который комплементарен парам оснований с кодоном мРНК. Акцепторный стержень — это сайт, к которому прикрепляется определенная аминокислота (в этом случае тРНК с аминокислотой называется аминоацил-тРНК ). Пептидил-тРНК — это тРНК, которая удерживает растущую полипептидную цепь.
Плечо антикодона содержит антикодон, который комплементарен парам оснований с кодоном мРНК. Акцепторный стержень — это сайт, к которому прикрепляется определенная аминокислота (в этом случае тРНК с аминокислотой называется аминоацил-тРНК ). Пептидил-тРНК — это тРНК, которая удерживает растущую полипептидную цепь.
В отличие от первых двух, рРНК (или рибосомная РНК) не несет генетической информации. Скорее, он служит одним из компонентов рибосомы. Рибосома — это цитоплазматическая структура в клетках прокариот и эукариот, которые известны тем, что служат местом синтеза белка.Рибосомы можно использовать для определения прокариота от эукариота. У прокариот есть рибосомы 70S, тогда как у эукариот рибосомы 80S. Однако оба типа состоят из двух субъединиц разного размера. Субъединица большего размера служит рибозимом, который катализирует образование пептидной связи между аминокислотами. рРНК имеет три сайта связывания: сайты A, P и E. Сайт A (аминоацил) — это место стыковки аминоацил-тРНК. Сайт P (пептидил) — это место, где связывается пептидил-тРНК. Сайт E (выход) — это место, где тРНК покидает рибосому.
Сайт P (пептидил) — это место, где связывается пептидил-тРНК. Сайт E (выход) — это место, где тРНК покидает рибосому.
Этапы биосинтеза белка
Транскрипция
Транскрипция — это процесс, с помощью которого матрица мРНК, кодирующая последовательность белка в форме тринуклеотидного кода, транскрибируется из ДНК, чтобы обеспечить матрицу для трансляции с помощью фермент, РНК-полимераза. Таким образом, транскрипция рассматривается как первый этап экспрессии гена. Подобно репликации ДНК, транскрипция происходит в направлении 5 ‘→ 3’. Но в отличие от репликации ДНК, транскрипции не требуется праймер для запуска процесса, и вместо тимина урацил спаривается с аденином.
Этапы транскрипции следующие: (1) инициация, (2) выход промотора, (3) элонгация и (4) терминация. Первый шаг, инициация, — это когда РНК-полимераза с помощью определенных факторов транскрипции связывается с промотором ДНК. Это приводит к открытию (раскручиванию) ДНК в промоторной области, образуя транскрипционный пузырь . Сайт старта транскрипции в пузыре транскрипции связывается с РНК-полимеразой, в частности с РНК-полимеразой , инициирующей NTP , и , продолжающей NTP .Происходит фаза прерывистых циклов синтеза, приводящая к высвобождению коротких транскриптов мРНК (примерно от 2 до 15 нуклеотидов). На следующем этапе РНК-полимераза должна ускользнуть от промотора и вступить в стадию элонгации. Во время элонгации РНК-полимераза пересекает матричную цепь ДНК и пары оснований с нуклеотидами на матричной (некодирующей) цепи. В результате получается транскрипт мРНК, содержащий копию кодирующей цепи ДНК, за исключением тиминов, которые заменены урацилами.Сахарно-фосфатный остов формируется за счет РНК-полимеразы. Последний шаг — прекращение. Во время этой фазы водородные связи спирали РНК-ДНК разрываются. У эукариот транскрипт мРНК проходит дальнейшую обработку. Он проходит полиаденилирования , укупорки и сплайсинга .
Сайт старта транскрипции в пузыре транскрипции связывается с РНК-полимеразой, в частности с РНК-полимеразой , инициирующей NTP , и , продолжающей NTP .Происходит фаза прерывистых циклов синтеза, приводящая к высвобождению коротких транскриптов мРНК (примерно от 2 до 15 нуклеотидов). На следующем этапе РНК-полимераза должна ускользнуть от промотора и вступить в стадию элонгации. Во время элонгации РНК-полимераза пересекает матричную цепь ДНК и пары оснований с нуклеотидами на матричной (некодирующей) цепи. В результате получается транскрипт мРНК, содержащий копию кодирующей цепи ДНК, за исключением тиминов, которые заменены урацилами.Сахарно-фосфатный остов формируется за счет РНК-полимеразы. Последний шаг — прекращение. Во время этой фазы водородные связи спирали РНК-ДНК разрываются. У эукариот транскрипт мРНК проходит дальнейшую обработку. Он проходит полиаденилирования , укупорки и сплайсинга .
Трансляция
Трансляция — это процесс, в котором аминокислоты связываются вместе в определенном порядке в соответствии с правилами, установленными генетическим кодом. Это происходит в цитоплазме, где расположены рибосомы. Он состоит из четырех фаз: (1) активация (аминокислота ковалентно связана с тРНК), (2) инициация (малая субъединица рибосомы связывается с 5′-концом мРНК с помощью факторов инициации), (3 ) элонгация (следующая аминоацил-тРНК в линии связывается с рибосомой вместе с GTP и фактором элонгации) и (4) терминация (сайт A рибосомы обращен к стоп-кодону).
Это происходит в цитоплазме, где расположены рибосомы. Он состоит из четырех фаз: (1) активация (аминокислота ковалентно связана с тРНК), (2) инициация (малая субъединица рибосомы связывается с 5′-концом мРНК с помощью факторов инициации), (3 ) элонгация (следующая аминоацил-тРНК в линии связывается с рибосомой вместе с GTP и фактором элонгации) и (4) терминация (сайт A рибосомы обращен к стоп-кодону).
Пост-трансляция
Следующие события синтеза белка — это события, например.грамм. протеолиз и сворачивание белков. Протеолиз относится к расщеплению белков протеазами. Посредством этого из полипептида удаляются N-концевые, C-концевые или внутренние аминокислотные остатки. Посттрансляционная модификация относится к ферментативному процессингу полипептидной цепи после трансляции и образования пептидной связи. Концы и боковые цепи полипептида можно модифицировать, чтобы гарантировать правильную клеточную локализацию и функцию. Сворачивание белка — это сворачивание полипептидных цепей с принятием вторичных и третичных структур.
См. Также
Ссылки
- Синтез белков. (2019). Получено с веб-сайта Elmhurst.edu: http://chemistry.elmhurst.edu/vchembook/584proteinsyn.html
- Protein Synthesis. (2019). Получено с веб-сайта Estrellamountain.edu: https://www2.estrellamountain.edu/faculty/farabee/biobk/BioBookPROTSYn.html
- Protein Synthesis. (2019). Получено с веб-сайта Nau.edu: http://www2.nau.edu/lrm22/lessons/protein-synthesis/protein-synthesis.htm
© Biology Online.Контент предоставлен и модерируется онлайн-редакторами биологии
Синтез белка | Hepatitis C Trust
Печень — один из важнейших органов для производства белков. Он производит или трансформирует миллионы белковых молекул каждый день. Белки состоят из аминокислот. Некоторые из этих аминокислот уже находятся в организме. Другие, называемые незаменимыми аминокислотами, можно получить только с пищей.
Белки выполняют множество жизненно важных функций. Мы используем их для роста и поддержания тканей тела, таких как мышцы, сердце, почки и стенки кровеносных сосудов.Печень производит сотни различных белков с разными функциями. Некоторые переносят витамины и минералы по телу. Некоторые действуют как катализаторы, ускоряющие метаболические реакции (эти белки называются ферментами). Другие регулируют структуру всех видов деятельности внутри клетки.
Мы используем их для роста и поддержания тканей тела, таких как мышцы, сердце, почки и стенки кровеносных сосудов.Печень производит сотни различных белков с разными функциями. Некоторые переносят витамины и минералы по телу. Некоторые действуют как катализаторы, ускоряющие метаболические реакции (эти белки называются ферментами). Другие регулируют структуру всех видов деятельности внутри клетки.
Альбумин
Альбумин обычно присутствует в крови в больших количествах. Он используется для связывания гормонов, некоторых химических веществ и лекарств. Альбумин также регулирует обмен воды между кровью и тканями.Если концентрация воды в жидкостях организма отличается от концентрации в клетках, существует риск того, что жидкость вырвется из клеток.
В течение нашей жизни печень постоянно контролирует уровень давления в организме, чтобы обеспечить его баланс. Если печень не может синтезировать достаточное количество альбумина, может произойти накопление жидкости в тканях. Обычно это симптом, связанный с декомпенсированным циррозом печени, когда жидкость может скапливаться в лодыжках, ступнях или ногах (отек) или в брюшной полости (асцит).
Обычно это симптом, связанный с декомпенсированным циррозом печени, когда жидкость может скапливаться в лодыжках, ступнях или ногах (отек) или в брюшной полости (асцит).
Утюг
Печень накапливает и выделяет железо по всему телу по мере необходимости. Само по себе железо токсично для клеток организма, поэтому организм связывает его с белком. В печени железо связано с белком ферритином.
Когда заболевание печени связано с увеличением количества ферритина в печени, оно может вызывать повреждение тканей и органов. Это называется гемохроматозом.
Железо также необходимо для образования гемоглобина.Это помогает транспортировать кислород в кровоток. Если железа недостаточно для образования гемоглобина, это может привести к летаргии и анемии.
Конверсия аммиака
Аммиак — токсичное вещество, получаемое в результате метаболизма белков. Печень превращает аммиак в мочевину, которая водорастворима, нетоксична и выводится почками. Дисфункция печени при декомпенсированном циррозе может привести к неспособности преобразовать этот аммиак, который затем накапливается в крови.
Белковая фабрика
Август в нашем календаре PDBe на 2020 год вдохновлен механизмами клеточного производства белка, называемыми рибосомами. Рибосомы — это очень сложные и важные структуры в клетке, которые выполняют жизненно важную роль в синтезе белка.
Белковая фабрика клетки
Каждая клетка нашего тела содержит около 10 миллиардов белков, которые позволяют нам думать, двигаться, есть, играть и делать многое другое. Их эффективное создание — это работа этих макромолекулярных машин, называемых рибосомами, которые обнаружены во всех живых клетках всех видов, от бактерий до людей.
Структура рибосомного комплекса
Глядя на рибосому, кажется, что она представляет собой запутанную мешанину белков и молекул РНК, однако на самом деле она сшита вместе с безупречной точностью.
Две субъединицы рибосомы, собранные вместе с малой и большой субъединицами, показаны серыми и бирюзовыми лентами соответственно (запись PDB 6KE0)
Криоэлектронная микроскопия и рентгеновская кристаллография показали, что рибосома состоит из двух субъединиц: малой и большой субъединиц.Каждая из этих субъединиц образует сложную сеть из нескольких молекул РНК с десятками различных белков. В 2000 году структурные биологи Венкатраман Рамакришнан, Томас А. Стейтц и Ада Э. Йонат разрешили первые кристаллические структуры рибосомы с атомным разрешением. В 2009 году этим трем исследователям была присуждена Нобелевская премия по химии за их исследования структуры и функции рибосомы, что свидетельствует о важности рибосомы.
Синтез белка
Синтез новых белков начинается в ядре, где рибосомы получают команду начать процесс.Участки ДНК (гены), кодирующие определенный белок, копируются на нити информационной РНК (мРНК) в процессе, называемом транскрипцией.
После завершения транскрипции ДНК в мРНК, следующим процессом является трансляция, когда эти мРНК считываются для образования белков. Каждая мРНК определяет порядок, в котором аминокислоты должны быть добавлены к белковой цепи в процессе ее синтеза. Если за основу берется ДНК, то масоны — это рибосомы — они строят белок, используя аминокислоты в качестве «кирпичиков».
Для создания белков две рибосомные субъединицы, малая и большая, собираются вместе, образуя полную рибосому. Он имеет сайты связывания для молекул мРНК и транспортной РНК (тРНК). Большая субъединица находится поверх маленькой субъединицы, а матрица мРНК расположена между ними. После полной сборки рибосома начинает процесс производства белка.
Производство белка
Двигаясь вдоль мРНК, рибосома считывает набор трехнуклеотидных последовательностей на мРНК, называемых кодоном, который кодирует определенную аминокислоту.ТРНК доставляет эти аминокислоты, строительные блоки белка, к рибосоме. Каждая молекула тРНК имеет два разных конца или сайта, один для связывания с определенной аминокислотой, а другой для связывания с соответствующим кодоном мРНК. Во время трансляции эти тРНК переносят аминокислоты на рибосому и присоединяются к своим комплементарным кодонам на мРНК. Впоследствии они преобразуются в правильные аминокислоты в новой белковой цепи.
Собранные аминокислоты сшиваются вместе с помощью молекул рРНК (рибосомной РНК), которые направляют процесс создания новой белковой цепи.Повторяя этот процесс для каждой аминокислоты, весь белок строится в процессе, называемом удлинением. Растущая белковая цепь останавливается только тогда, когда она встречает стоп-кодон на мРНК. Это сигнализирует об окончании полипептидной цепи во время трансляции. Как только аминокислоты сформированы правильно, вновь синтезированная белковая цепь транспортируется либо в цитоплазму, либо в аппарат Гольджи у прокариот или эукариот, соответственно.
Ниже приведено видео с сайта YourGenome, объясняющее этот процесс.
Больше, чем протеиновый завод
Точный и быстрый перевод генетической информации критически важен для производства функциональных белков для жизнеспособности клеток.Скорость производства белка должна быть быстрой и очень точной, чтобы своевременно реагировать на изменения в окружающей среде. Поразительная точность рибосомного оборудования имеет коэффициент ошибки 1 на 1000–10 000 аминокислот. Одна рибосома в эукариотической клетке может добавлять 2 аминокислоты к белковой цепи каждую секунду, однако у прокариот рибосомы могут работать даже быстрее, добавляя к полипептиду около 20 аминокислот каждую секунду. Рибосомы потребляют большое количество энергии для синтеза белков и составляют значительную часть клеточной массы, при этом значительная часть метаболизма клетки направляется на производство рибосомных белков и РНК.
Нацелены на бактериальные рибосомы
Рибосомы встречаются во всех формах жизни и необходимы для синтеза белка, что делает их желательной мишенью для лекарств. Большинство клинически используемых антибиотиков нацелены на рибосомы и ингибируют процесс синтеза белка, вмешиваясь в трансляцию мРНК или блокируя образование пептидных связей.
Бактериальные рибосомы — одна из основных мишеней для антибиотиков. Эти антибиотики не позволяют бактериям синтезировать собственные белки из-за ингибирования их рибосомы, которая в конечном итоге убивает бактерии.Разработка таких антибиотиков стала возможной из-за различий между бактериальными и эукариотическими рибосомами. Они различаются не только по размеру, но и по последовательности и структуре, что позволяет антибиотикам убивать только бактерии, подавляя их рибосомы, не затрагивая человеческие рибосомы.
В PDB доступны структуры многих антибиотиков в комплексе с рибосомами. Эти структуры с разрешением на атомарном уровне позволяют нам лучше понять механизм их действия.
Антибиотики, спасающие жизнь
Антибиотики, такие как неомицин, гентамицин и стрептомицин, относятся к группе аминогликозидов, которые широко используются для лечения тяжелых инфекций брюшной полости и мочевыводящих путей. Они ингибируют малую субъединицу рибосомы, включая тетрациклины, которые блокируют связывание тРНК.
Другой широко назначаемый антибиотик, эритромицин, относится к классу натуральных продуктов. Он оказывает два эффекта на трансляцию: во-первых, предотвращает удлинение полипептидной цепи, а во-вторых, ингибирует образование большой рибосомной субъединицы.
На рисунке ниже показан ряд антибиотиков, которые нацелены на бактериальную рибосому на разных участках большой (голубовато-серый) и малой (желтый) субъединицы рибосомы.
Это изображение взято из статьи «Бактериальная рибосома как мишень для антибиотиков». Nat Rev Microbiol 3, 870–881 (2005). https://doi.org/10.1038/nrmicro1265
Ингибирование эукариотической рибосомы
Некоторые антибиотики, такие как генетицин, также называемый G418, ингибируют стадию элонгации как в прокариотических, так и в эукариотических рибосомах.Рицин, лектин (белок, связывающий углеводы), вырабатываемый семенами клещевины, является сильнодействующим токсином. Всего несколько крупинок очищенного порошка рицина могут убить взрослого человека. Он ингибирует удлинение путем ферментативной модификации рРНК большой рибосомной субъединицы эукариот. Другим известным ингибитором трансляции эукариот является циклогексимид, который обычно используется в лабораториях для подавления синтеза белка.
Противоопухолевые препараты
Биогенез рибосом, процесс создания рибосом, недавно стал эффективной мишенью в терапии рака.Несколько соединений, ингибирующих продукцию или функцию рибосом, преимущественно убивающих раковые клетки, прошли клинические испытания. Недавние исследования показывают, что клетки экспрессируют гетерогенные популяции рибосом и что состав рибосом может играть ключевую роль в онкогенезе, открывая новые терапевтические возможности.
Об изображении
Два произведения искусства, керамическая скульптура (слева) и кусок шелкового батика (справа), были созданы Шином Галаутом и Мари Бишофс, 13-летними учениками Школы Персе и Колледжа Импингтон Вилладж, соответственно.Оба художника черпали вдохновение из комплексов белков и нуклеиновых кислот в базе данных PDB, а их работы основывались на процессе синтеза белка и рибосомах.
Дипти Гупта
Синтез белков · Анатомия и физиология
Синтез белков · Анатомия и физиология
К концу этого раздела вы сможете:
- Объясните, как генетический код, хранящийся в ДНК, определяет белок, который будет образовывать
- Опишите процесс транскрипции
- Опишите процесс перевода
- Обсудить функцию рибосом
Ранее упоминалось, что ДНК обеспечивает «план» структуры и физиологии клетки.Это относится к тому факту, что ДНК содержит информацию, необходимую клетке для построения одного очень важного типа молекулы: белка. Большинство структурных компонентов клетки состоят, по крайней мере частично, из белков, и практически все функции, которые выполняет клетка, выполняются с помощью белков. Один из наиболее важных классов белков — это ферменты, которые помогают ускорить необходимые биохимические реакции, происходящие внутри клетки. Некоторые из этих критических биохимических реакций включают создание более крупных молекул из более мелких компонентов (например, происходит во время репликации ДНК или синтеза микротрубочек) и разрушение более крупных молекул на более мелкие компоненты (например, при сборе химической энергии из молекул питательных веществ).Каким бы ни был клеточный процесс, в нем почти наверняка участвуют белки. Подобно тому, как геном клетки описывает полный набор ДНК, протеом клетки представляет собой полный набор белков. Синтез белка начинается с генов. Ген — это функциональный сегмент ДНК, который обеспечивает генетическую информацию, необходимую для создания белка. Каждый конкретный ген обеспечивает код, необходимый для создания определенного белка. Экспрессия гена , которая преобразует информацию, закодированную в гене, в конечный продукт гена, в конечном итоге определяет структуру и функцию клетки, определяя, какие белки сделаны.
Интерпретация генов работает следующим образом. Напомним, что белки — это полимеры или цепи многих строительных блоков аминокислот. Последовательность оснований в гене (то есть его последовательность нуклеотидов A, T, C, G) транслируется в аминокислотную последовательность. Триплет — это участок из трех оснований ДНК в ряду, который кодирует определенную аминокислоту. Подобно тому, как трехбуквенный код d-o-g сигнализирует об изображении собаки, трехбуквенный базовый код ДНК сигнализирует об использовании определенной аминокислоты.Например, триплет ДНК CAC (цитозин, аденин и цитозин) определяет аминокислоту валин. Следовательно, ген, состоящий из нескольких триплетов в уникальной последовательности, обеспечивает код для построения целого белка с несколькими аминокислотами в правильной последовательности ([ссылка]). Механизм, с помощью которого клетки превращают код ДНК в белковый продукт, представляет собой двухэтапный процесс с молекулой РНК в качестве промежуточного звена.
От ДНК к РНК: транскрипция
ДНК
размещается в ядре, а синтез белка происходит в цитоплазме, поэтому должен быть какой-то промежуточный мессенджер, который покидает ядро и управляет синтезом белка.Этот промежуточный мессенджер представляет собой матричную РНК (мРНК) , одноцепочечную нуклеиновую кислоту, которая несет копию генетического кода одного гена из ядра в цитоплазму, где она используется для производства белков.
Существует несколько различных типов РНК, каждая из которых выполняет свои функции в клетке. Структура РНК похожа на ДНК, за некоторыми небольшими исключениями. Во-первых, в отличие от ДНК, большинство типов РНК, включая мРНК, являются одноцепочечными и не содержат дополнительных цепей.Во-вторых, сахар рибозы в РНК содержит дополнительный атом кислорода по сравнению с ДНК. Наконец, вместо основного тимина РНК содержит основной урацил. Это означает, что аденин всегда будет соединяться с урацилом в процессе синтеза белка.
Экспрессия гена начинается с процесса, называемого транскрипцией , который представляет собой синтез цепи мРНК, комплементарной интересующему гену. Этот процесс называется транскрипцией, потому что мРНК подобна транскрипту или копии кода ДНК гена.Транскрипция начинается примерно так же, как репликация ДНК, когда участок ДНК раскручивается и две нити разделяются, однако только эта небольшая часть ДНК будет разделена. Тройки внутри гена на этом участке молекулы ДНК используются в качестве матрицы для транскрипции комплементарной цепи РНК ([ссылка]). Кодон представляет собой последовательность из трех оснований мРНК, названную так потому, что они непосредственно кодируют аминокислоты. Как и в случае репликации ДНК, транскрипция состоит из трех стадий: инициация, удлинение и завершение.
Этап 1: Инициирование. Область в начале гена, называемая промотором , — конкретная последовательность нуклеотидов — запускает начало транскрипции.
Этап 2: удлинение. Транскрипция начинается, когда РНК-полимераза раскручивает сегмент ДНК. Одна цепь, называемая кодирующей цепью, становится матрицей с генами, которые нужно кодировать. Затем полимераза выравнивает правильную нуклеиновую кислоту (A, C, G или U) с ее комплементарным основанием на кодирующей цепи ДНК. РНК-полимераза — это фермент, который добавляет новые нуклеотиды к растущей цепи РНК. Этот процесс создает цепь мРНК.
Этап 3: Прекращение действия. Когда полимераза достигает конца гена, один из трех конкретных триплетов (UAA, UAG или UGA) кодирует «стоп-сигнал», который запускает ферменты для прекращения транскрипции и высвобождения транскрипта мРНК.
Прежде чем молекула мРНК покинет ядро и приступит к синтезу белка, она модифицируется несколькими способами.По этой причине на этой стадии ее часто называют пре-мРНК. Например, ваша ДНК и, следовательно, комплементарная мРНК, содержит длинные области, называемые некодирующими областями, которые не кодируют аминокислоты. Их функция до сих пор остается загадкой, но процесс, называемый сплайсингом , удаляет эти некодирующие области из транскрипта пре-мРНК ([ссылка]). Сплайсосома — структура, состоящая из различных белков и других молекул — прикрепляется к мРНК и «сплайсирует» или вырезает некодирующие области.Удаленный сегмент транскрипта называется интроном . Остальные экзоны склеены. Экзон представляет собой сегмент РНК, который остается после сплайсинга. Интересно, что некоторые интроны, удаленные из мРНК, не всегда являются некодирующими. Когда различные кодирующие области мРНК сплайсируются, в конечном итоге возникают разные вариации белка с различиями в структуре и функциях. Этот процесс приводит к гораздо большему разнообразию возможных белков и функций белков.Когда транскрипт мРНК готов, он выходит из ядра в цитоплазму.
От РНК к белку: перевод
Подобно переводу книги с одного языка на другой, кодоны цепи мРНК должны быть переведены в аминокислотный алфавит белков. Трансляция — это процесс синтеза цепочки аминокислот, называемой полипептидом . Для перевода требуются два основных вспомогательных средства: во-первых, «транслятор», молекула, которая будет осуществлять трансляцию, и, во-вторых, субстрат, на котором нить мРНК транслируется в новый белок, как «стол переводчика».Оба эти требования выполняются другими типами РНК. Субстратом, на котором происходит трансляция, является рибосома.
Помните, что многие рибосомы клетки связаны с грубым ER и осуществляют синтез белков, предназначенных для аппарата Гольджи. Рибосомная РНК (рРНК) — это тип РНК, которая вместе с белками составляет структуру рибосомы. Рибосомы существуют в цитоплазме в виде двух отдельных компонентов, маленькой и большой субъединицы.Когда молекула мРНК готова к трансляции, две субъединицы объединяются и прикрепляются к мРНК. Рибосома обеспечивает субстрат для трансляции, объединяя и выравнивая молекулу мРНК с молекулярными «трансляторами», которые должны расшифровать ее код.
Другое важное требование для синтеза белка — это молекулы-трансляторы, которые физически «читают» кодоны мРНК. Трансферная РНК (тРНК) — это тип РНК, которая переносит соответствующие соответствующие аминокислоты на рибосому и присоединяет каждую новую аминокислоту к последней, последовательно выстраивая полипептидную цепь.Таким образом, тРНК переносит определенные аминокислоты из цитоплазмы в растущий полипептид. Молекулы тРНК должны быть способны распознавать кодоны на мРНК и сопоставлять их с правильной аминокислотой. ТРНК модифицируется для этой функции. На одном конце его структуры находится сайт связывания определенной аминокислоты. На другом конце находится последовательность оснований, которая соответствует кодону, определяющему его конкретную аминокислоту. Эта последовательность из трех оснований в молекуле тРНК называется антикодоном . Например, тРНК, отвечающая за перемещение аминокислоты глицина, содержит сайт связывания глицина на одном конце.С другой стороны, он содержит антикодон, который дополняет кодон глицина (GGA является кодоном для глицина, и поэтому антикодон тРНК будет читать CCU). Оборудованная своим специфическим грузом и подходящим антикодоном, молекула тРНК может считывать свой распознанный кодон мРНК и переносить соответствующую аминокислоту в растущую цепь ([ссылка]).
Подобно процессам репликации и транскрипции ДНК, трансляция состоит из трех основных стадий: инициации, элонгации и терминации.Инициирование происходит при связывании рибосомы с транскриптом мРНК. Стадия элонгации включает распознавание антикодона тРНК следующим кодоном мРНК в последовательности. Как только последовательности антикодона и кодона связаны (помните, что они являются комплементарными парами оснований), тРНК представляет свой аминокислотный груз, и растущая полипептидная цепь присоединяется к этой следующей аминокислоте. Это прикрепление происходит с помощью различных ферментов и требует энергии. Затем молекула тРНК высвобождает цепь мРНК, цепь мРНК сдвигает один кодон в рибосоме, и следующая подходящая тРНК прибывает с соответствующим ей антикодоном.Этот процесс продолжается до тех пор, пока не будет достигнут последний кодон на мРНК, который обеспечивает сообщение «стоп», сигнализирующее о прекращении трансляции и запускающее высвобождение полного, вновь синтезированного белка. Таким образом, ген в молекуле ДНК транскрибируется в мРНК, которая затем транслируется в белковый продукт ([ссылка]).
Обычно транскрипция мРНК транслируется одновременно несколькими соседними рибосомами. Это увеличивает эффективность синтеза белка.Одна рибосома может транслировать молекулу мРНК примерно за одну минуту; таким образом, несколько рибосом на борту одного транскрипта могут производить в несколько раз больше одного и того же белка за одну минуту. Полирибосома представляет собой цепочку рибосом, транслирующую одну цепь мРНК.
Посмотрите это видео, чтобы узнать о рибосомах. Рибосома связывается с молекулой мРНК, чтобы начать трансляцию своего кода в белок. Что происходит с малыми и большими субъединицами рибосом в конце трансляции?
Обзор главы
ДНК хранит информацию, необходимую для того, чтобы дать клетке команду выполнять все свои функции.Клетки используют генетический код, хранящийся в ДНК, для создания белков, которые в конечном итоге определяют структуру и функцию клетки. Этот генетический код заключается в особой последовательности нуклеотидов, составляющих каждый ген в молекуле ДНК. Чтобы «прочитать» этот код, ячейка должна выполнить два последовательных шага. На первом этапе транскрипции код ДНК преобразуется в код РНК. Молекула информационной РНК, комплементарная конкретному гену, синтезируется в процессе, аналогичном репликации ДНК.Молекула мРНК обеспечивает код для синтеза белка. В процессе трансляции мРНК прикрепляется к рибосоме. Затем молекулы тРНК перемещают соответствующие аминокислоты к рибосоме, одну за другой, кодируемые последовательными триплетными кодонами на мРНК, до тех пор, пока белок не будет полностью синтезирован. По завершении мРНК отделяется от рибосомы, и белок высвобождается. Как правило, несколько рибосом присоединяются к одной молекуле мРНК одновременно, так что из мРНК можно производить несколько белков одновременно.
Вопросы по интерактивной ссылке
Посмотрите это видео, чтобы узнать о рибосомах. Рибосома связывается с молекулой мРНК, чтобы начать трансляцию своего кода в белок. Что происходит с малыми и большими субъединицами рибосом в конце трансляции?
Они разделяются и перемещаются и могут свободно присоединяться к трансляции других сегментов мРНК.
Вопросы для обзора
Что из следующего не является разницей между ДНК и РНК?
- ДНК содержит тимин, тогда как РНК содержит урацил
- ДНК содержит дезоксирибозу, а РНК содержит рибозу
- ДНК содержит чередующиеся сахарофосфатные молекулы, тогда как РНК не содержит сахаров
- РНК одноцепочечная, а ДНК двухцепочечная
Транскрипция и перевод выполняются в ________ и ________, соответственно.
- ,
- ядро; цитоплазма
- ядрышко; ядро
- ядрышко; цитоплазма
- цитоплазма; ядро
Сколько «букв» в последовательности молекулы РНК необходимо, чтобы обеспечить код отдельной аминокислоты?
- 1
- 2
- 3
- 4
Что из следующего не состоит из РНК?
- носители, которые перетасовывают аминокислоты в растущую полипептидную цепь
- рибосома
- молекула-мессенджер, обеспечивающая код для синтеза белка
- интрон
Вопросы о критическом мышлении
Кратко объясните сходство между транскрипцией и репликацией ДНК.
Транскрипция и репликация ДНК включают синтез нуклеиновых кислот. Эти процессы имеют много общих черт, в частности, схожие процессы инициирования, удлинения и завершения. В обоих случаях молекула ДНК должна быть раскручена и разделена, а кодирующая (то есть смысловая) цепь будет использоваться в качестве матрицы. Кроме того, полимеразы служат для добавления нуклеотидов к растущей цепи ДНК или мРНК. Оба процесса получают сигнал о завершении по завершении.
Контрастная транскрипция и перевод.Назовите хотя бы три различия между двумя процессами.
Транскрипция — это на самом деле процесс «копирования», а трансляция — это на самом деле процесс «интерпретации», потому что транскрипция включает копирование сообщения ДНК в очень похожее сообщение РНК, тогда как трансляция включает преобразование сообщения РНК в совершенно другое аминокислотное сообщение. Эти два процесса также различаются по своему расположению: транскрипция происходит в ядре, а трансляция — в цитоплазме. Механизмы, с помощью которых выполняются эти два процесса, также полностью различны: транскрипция использует ферменты полимеразы для создания мРНК, тогда как трансляция использует различные виды РНК для создания белка.
Глоссарий
- антикодон
- последовательная последовательность из трех нуклеотидов в молекуле тРНК, которая комплементарна определенному кодону в молекуле мРНК
- кодон
- последовательная последовательность из трех нуклеотидов на молекуле мРНК, которая соответствует определенной аминокислоте
- экзон
- одна из кодирующих областей молекулы мРНК, остающихся после сплайсинга
- ген
- , обеспечивающая генетическую информацию, необходимую для построения белка
Функциональная длина ДНК
- экспрессия гена
- активная интерпретация информации, закодированной в гене, для производства функционального генного продукта
- интрон
- некодирующих областей транскрипта пре-мРНК, которые могут быть удалены во время сплайсинга
- матричная РНК (мРНК)
- нуклеотидная молекула, которая служит промежуточным звеном в генетическом коде между ДНК и белком
- полипептид
- цепочка аминокислот, связанных пептидными связями
- полирибосома
- одновременная трансляция одного транскрипта мРНК множеством рибосом
- промотор
- область ДНК, которая сигнализирует о начале транскрипции в этом сайте в пределах гена
- протеом
- Полный набор белков, продуцируемых клеткой (определяется экспрессией специфических генов клетки)
- рибосомная РНК (рРНК)
- РНК, составляющая субъединицы рибосомы
- РНК-полимераза
- , который раскручивает ДНК, а затем добавляет новые нуклеотиды к растущей цепи РНК для фазы транскрипции синтеза белка
Фермент
- сплайсосома
- Комплекс ферментов, который служит для расщепления интронов транскрипта пре-мРНК
- сращивание
- процесс модификации транскрипта пре-мРНК путем удаления определенных, обычно некодирующих, областей
- транскрипция
- процесс производства молекулы мРНК, комплементарной определенному гену ДНК
- транспортная РНК (тРНК)
- молекул РНК, которые служат для доставки аминокислот к растущей полипептидной цепи и правильного размещения их в последовательности
- перевод
- процесс получения белка из кода нуклеотидной последовательности транскрипта мРНК
- тройка
- последовательная последовательность из трех нуклеотидов в молекуле ДНК, которая при транскрибировании в кодон мРНК соответствует определенной аминокислоте
Это произведение находится под лицензией Creative Commons Attribution 4.0 Международная лицензия.
Вы также можете бесплатно скачать по адресу http://cnx.org/contents/[email protected]
Атрибуция:
Питание и синтез мышечного белка: описательный обзор
Реферат
Предпосылки
Врачи хиропрактики часто дают пациентам лечебные упражнения и советы по питанию. Роль скелетных мышц в обеспечении здоровья и болезней недооценивается. Создание синергии между потреблением белка и упражнениями способствует синтезу белка и может повлиять на результаты лечения пациентов.
Цель
Изучить литературу, описывающую метаболизм белков и упражнения в связи с практикой хиропрактики.
Метод
Поиск в базах данных PubMed и Web of Science проводился с использованием ключевых терминов «метаболизм белка», «синтез белка», «упражнения», «сыворотка», «соя» и «тренировки с отягощениями» в различных комбинациях. Пределы исключали использование статей, которые не были основаны на людях, относились к младенцам или болезням или были опубликованы до 1988 года.В конечном итоге для анализа было включено 30 работ.
Обсуждение
Количество, тип и время потребления белка играют решающую роль в стимулировании синтеза белка. Внутриклеточный механизм синтеза белка включает множество взаимосвязанных интересных компонентов.
Заключение
Адаптация к упражнениям (синтез белка) может быть улучшена путем контроля типа белка, количества потребляемого белка и времени потребления белка.Врачи хиропрактики могут повлиять на результаты лечения пациентов, используя эмпирические данные о потреблении белка и упражнениях, чтобы максимизировать синтез белка.
Ключевые слова: белок , синтез, метаболизм, упражнения, хиропрактика
Введение
Врачи хиропрактики обычно назначают упражнения и дают рекомендации по питанию пациентов. В отчете «Анализ работы хиропрактики» за 2005 год Национального совета хиропрактиков было обнаружено, что 89% опрошенных хиропрактиков использовали консультации по питанию, терапию или пищевые добавки. 1 Кроме того, более 98% использовали корректирующие или лечебные упражнения в своей практике хиропрактики. Что касается «Процедур по укреплению здоровья и благополучию», 98,3% хиропрактиков инструктировали пациентов по вопросам физической подготовки / физических упражнений, а 97,7% давали инструкции по рекомендациям по питанию / диете. Рецепты упражнений и / или рекомендации по питанию, будь то терапевтические или оздоровительные, даются с ожиданием изменений. Врачи и пациенты ожидают, что физические упражнения приведут к положительной адаптации.Желаемое изменение — это, по сути, анаболическая реакция на синтез белка при минимизации катаболизма белка. Открыто врачи оценивают изменения в силе, выносливости и / или функциях. Явные клинические изменения вызваны, по крайней мере частично, клеточными изменениями. Произойдет ли желаемое изменение или нет, во многом зависит от синергетической взаимосвязи между диетой и упражнениями. В этом обзоре основное внимание уделяется вмешательствам в области питания, которые оптимизируют адаптацию за счет синтеза белка.
Метод
Поиск литературы включал использование баз данных Web of Science и PubMed.При поиске использовались несколько комбинаций следующих ключевых слов: белковый метаболизм, синтез белка, упражнения, сыворотка, соя, и тренировки с отягощениями. Это привело к первоначальному выходу нескольких сотен статей. Когда поиск ограничивался исследованиями с участием людей, за исключением тех, которые касались младенцев или болезней (например, ВИЧ), и статей, опубликованных до 1998 г. (если их содержание не было очень значимым и основополагающим для установления линии доказательств), окончательный результат был тридцать статей по этой теме.
Обсуждение
Основная цель этого обзора — описать белковый метаболизм и упражнения, связанные с практикой хиропрактики. Позвоночник состоит не только из кости. Скелетные мышцы играют ключевую роль в обеспечении позвоночника опорой, движением, проприоцепцией и выносливостью. Когда пациенты занимаются физическими упражнениями, будь то аэробные упражнения или упражнения с отягощениями, мышцы разрушаются и восстанавливают белок в ответ на стимул. Существуют новые эмпирические данные, позволяющие оптимизировать взаимосвязь между стимулом и реакцией.Как указывалось ранее, большинство докторов хиропрактики применяют упражнения и питание на практике. Признание и применение эмпирических данных о том, как тип, время и количество потребления белка влияют на адаптацию, могут помочь врачам и пациентам достичь оптимальных результатов.
Важность скелетных мышц
Функцию скелетных мышц можно недооценить, если приписать ей простую роль «движения тела». Скелетные мышцы являются основным резервуаром аминокислот для других тканей.Фактически, запасы аминокислот в скелетных мышцах — единственное хранилище, способное к большим потерям без ущерба для способности поддерживать жизнь. 2 Когда уровень глюкозы в крови падает, аминокислоты являются основным глюконеогенным субстратом печени. Быстрый оборот клеток пищеварительного тракта требует постоянного поступления аминокислот. Азот, содержащийся в аминокислотах, является жизненно важным компонентом ДНК и других молекул, необходимых для поддержания и репликации клеток. Потребность тканей в аминокислотах становится критической во время стресса, такого как сепсис, рак или травмы.Экспорт аминокислот в мышцы может привести к большой потере белка или саркопении. Саркопения может сильно затруднить выздоровление от будущей болезни или травмы. 3 Это вызывает особую озабоченность у пожилых людей, которые могут не вылечиться от болезни в основном в результате саркопении. Они становятся слишком хрупкими и перестают развиваться. Пожилым людям намного труднее наращивать мышцы; 4 поэтому следует соблюдать осторожность, чтобы сохранить мышечную массу на протяжении всей жизни. Arts обнаружила, что средняя толщина двуглавой мышцы плеча и четырехглавой мышцы бедра у здоровых 90-летних мужчин аналогична таковой у 5-летних детей. 5 Это вызывает беспокойство, если учесть, какой дополнительный вес должен нести взрослый во время повседневной деятельности. В обзоре Вольфа 3 более полно обсуждается недооцененная роль скелетных мышц в здоровье и болезнях. Врачи-хиропрактики должны понимать ценность поддержания мышечной массы на протяжении всей жизни и понимать роль упражнений и питания в синтезе белка.
В этой статье описывается влияние питания и физических упражнений на синтез белка.Упражнения с отягощениями — мощный сигнал, который можно усилить количеством, временем и типом потребляемого белка. Механизм синтеза белка сложен и увлекателен. Более полное понимание этой темы может позволить хиропрактику разработать более эффективные стратегии увеличения или сохранения скелетных мышц на протяжении всей жизни.
Упражнения с отягощениями, питание и синтез белка
Скелетные мышцы постоянно разрушаются и синтезируют белок.Если мышца собирается поддерживать свою массу, чистый уровень белкового баланса должен быть равен нулю. Если мышцы собираются набирать массу, синтез белка должен превышать распад белка. Хотя обычно тренировки с отягощениями воспринимаются как мощный стимул для синтеза белка, важно также понимать, что они являются сигналом к распаду белка. Филлипс и др. 6 показал, что упражнения с отягощениями приводят к увеличению баланса чистого белка в мышцах на 24–48 часов. Как анаболизм белка, так и катаболизм усиливаются после упражнений, но увеличение анаболизма относительно больше, что приводит к положительному балансу чистого мышечного белка.В своем эксперименте Филлипс и др. 6 использовал 8 подходов по 8 концентрических и эксцентрических мышечных движений с 80% от максимального усилия каждого повторения каждого испытуемого. Участникам исследования разрешалось принимать пищу до и после тренировки. Это важно, поскольку более поздние исследования показали отсутствие синтеза белка после упражнений при отсутствии питания. 7 Esmarck et al. 8 2 группы тренировались с отягощениями в течение 12 недель и показали, что в контрольной группе, не получавшей питания в течение 2 часов после тренировки, уменьшилась безжировая масса тела, в то время как у тех, кто потреблял 10 г белка сразу после тренировки, увеличилась безжировая масса.
Как анаболизм, так и катаболизм белков очевидны после тренировки с отягощениями. Тем не менее, если питание отсутствует после тренировки, синтез белка может быть снижен или отсутствовать. Количество потребляемого белка является важным фактором для максимального увеличения скорости и продолжительности синтеза белка. Borsheim et al. 9 показали, что 6 г смешанных аминокислот повышают синтез белка после тренировки. В том же эксперименте 6 г незаменимых аминокислот удвоили синтез белка, что привело исследователей к выводу, что заменимые аминокислоты не требуются для стимулирования синтеза белка.Роль заменимых аминокислот в синтезе белка остается спорной. Cuthbertson et al. 10 показали, что пероральная доза 10 г незаменимых аминокислот максимально стимулировала синтез белка, и выдвинули гипотезу, что это произойдет, если за один прием пищи съесть эквивалент 6 унций мяса, рыбы, яиц или молока. Филлипс и др. 11 заявил, что 25 г качественного источника белка (молочные продукты, мясо и яйца) содержат примерно 10 г незаменимых аминокислот и должны максимально стимулировать синтез белка после тренировки.
Время потребления белка
Время потребления белка имеет решающее значение для увеличения синтеза белка. Потребление протеина сразу после тренировки стимулирует синтез протеина, а ожидание всего через 2 часа после тренировки притупляет реакцию. 8 Rasmussen et al. 7 обнаружили повышенный синтез белка у тех, кто потреблял 6 г незаменимых аминокислот после тренировки, по сравнению с контрольной группой. Белок потребляли через 1 или 3 часа после тренировки.В отличие от Esmark et al., 8 , не было различий в скорости синтеза белка в группах после тренировки через 1 или 3 часа. Типтон и др. 12 обнаружили более выраженный анаболический ответ, когда углеводно-белковый напиток употреблялся перед тренировкой, чем если бы он употреблялся сразу после тренировки. В этом эксперименте обе группы потребляли 6 г незаменимых аминокислот до или после тренировки с отягощениями. Они предположили, что увеличение кровотока и доставки аминокислот к мышцам во время упражнений было причиной наблюдаемого повышенного синтеза белка.
Тип белка
Тип потребляемого белка также может быть важным фактором в стимуляции синтеза белка. Было показано, что добавление соевого и сывороточного протеина увеличивает мышечную массу и силу по сравнению с плацебо. 13 Candow et al. 13 показали это с 6-недельными тренировками с отягощениями у молодых взрослых людей. Энтони и др. 14 обнаружили, что сывороточные и соевые белковые добавки способствуют синтезу белка в скелетных мышцах больше, чем добавки, содержащие только углеводы.Они отметили большую внутриклеточную передачу сигналов (фосфорилирование S6K1 и mTOR) в группе, получавшей сывороточный протеин, по сравнению с группой сои. Внутриклеточная передача сигналов будет обсуждаться более подробно ниже. Энтони и др. 14 использовали аэробное кондиционирование и крыс в своем эксперименте; однако это может не совпадать с тренировкой с отягощениями в человеческой популяции. Wilkinson et al. 15 также продемонстрировал способность как соевого, так и молочного белка увеличивать чистый белковый баланс у людей после тренировок с отягощениями.Они обнаружили более высокую скорость фракционного синтеза белка и больший баланс чистого мышечного белка после приема молока по сравнению с соей. Испытуемые в их эксперименте потребляли 18,2 грамма белка вскоре после тренировки. Morifuji et al. 16 показали, что добавление сывороточного протеина по сравнению с казеином вызывало снижение активности липогенных ферментов печени и повышение активности липогенных ферментов в мышцах. Сывороточный протеин может дать скелетным мышцам преимущество, позволяя им накапливать больше энергии (жира), необходимой для синтеза протеина.Он также может снизить производство жира в печени и помочь в борьбе с ожирением.
Tipton et al. 17 опубликовали наводящее на размышления исследование, сравнивающее способность казеина и сывороточного протеина стимулировать анаболизм скелетных мышц в ответ на упражнения с отягощениями. И казеин, и сыворотка являются молочными белками, но сыворотка остается растворимой в желудке и быстрее выводится в тонкий кишечник. Казеин выходит из желудка медленнее. Они обнаружили, что сывороточный протеин повышает плазменный уровень и уровень внутриклеточного лейцина выше, чем у казеина или в контрольной группе.Сывороточный протеин увеличивал внутриклеточную концентрацию лейцина на 110% через 1 час после употребления. Группа сыворотки также имела более высокие концентрации инсулина в сыворотке после употребления. В группе казеина потребление фенилаланина было на 35% больше, чем в группе, получавшей сыворотку. Поглощение фенилаланина положительно коррелирует с синтезом белка. Это не было долгосрочным исследованием, посвященным наращиванию мышц. Авторы пришли к выводу, что и казеин, и сыворотка стимулируют анаболический ответ в мышцах после тренировки, но неясно, дает ли одно преимущество перед другим.
Типичное измерение синтеза белка обычно включает импорт и экспорт в скелетные мышцы аминокислоты фенилаланина. 18 Скелетные мышцы обладают способностью окислять шесть аминокислот (лейцин, изолейцин, валин, аспартат, аспарагин и глутамат). Импорт этих аминокислот напрямую не коррелирует с их добавлением в структуру белка. Фенилаланин — незаменимая аминокислота, которую мышцы не могут окислить. Считается, что его поглощение и включение в мышцы является точным индикатором синтеза мышечного белка. 18
Внутриклеточная передача сигналов
Результаты Tipton et al. 17 открывают сложную дискуссию относительно внутриклеточной передачи сигналов и синтеза белка. Лейцин, который в больших количествах содержится в сывороточном протеине, является не только строительным блоком в синтезе протеина, но и важным внутриклеточным сигналом, направляющим скелетные мышцы для трансляции протеина. Полный обзор внутриклеточной передачи сигналов выходит за рамки этого обзора; однако Proud 19 предоставляет более глубокий обзор механизмов, участвующих в синтезе клеточного белка.Ниже приводится краткий обзор потенциальных сигналов для увеличения синтеза белка в скелетных мышцах. Три ключевых компонента передачи сигналов включают: энергетический статус мышечной клетки, инсулин и аминокислоту лейцин ().
Ключевые компоненты внутриклеточной передачи сигналов
Изменения скорости синтеза белка начинаются до изменения содержания мРНК. 20 Это означает, что посттранскрипционный механизм играет преобладающую роль в активации синтеза белка. На транскрипцию влияет множество переменных, но энергетический заряд клетки, инсулин и лейцин кажутся очень важными.Во время упражнений энергетический заряд клеток или содержание АТФ (аденозинтрифосфата) снижается. Мишень рапамицина у млекопитающих (mTOR) рассматривается как «главный регулятор» трансляции внутри клетки. 19 Существует 2 типа субъединиц mTOR: mTORC1 и mTORC2. Когда уровни АТФ снижены, активируется АМФ-активированная протеинкиназа (AMPK). AMPK фосфорилирует промежуточную молекулу (TSC2), которая выключает передачу сигналов mTORC1. Bolster et al. 21 показали отрицательную корреляцию между активацией AMPK и фосфорилированием mTOR и других ключевых сигнальных молекул (S6K1 и 4E-BP1).
Снижение клеточного уровня АТФ снижает синтез белка в клетке. В этом есть смысл, поскольку синтез белка требует много энергии. Добавление 1 аминокислоты в процессе трансляции требует разрушения 4 молекул АТФ. 19 Для синтеза белка может потребоваться примерно 485 килокалорий в день у мускулистого молодого мужчины и примерно 120 килокалорий в день у активной пожилой женщины. 3 После хорошего ночного сна без еды синтез белка снижается на 15–30 процентов. 22 Питание клеток после тренировки позволяет им поддерживать высокий энергетический заряд и способствовать непрерывному синтезу белка.
Упражнения с отягощениями изменяют энергетический статус скелетных мышц. Купман и др. 23 документально подтвердили изменения уровней гликогена и липидов в мышцах после тренировки с отягощениями. Один сеанс упражнений с отягощениями снижает уровень мышечного гликогена в волокнах как I, так и II типа. В эксперименте 8 подходов по 10 повторений жима ногами с последующими 8 подходами по 10 повторений разгибаний ног снизили содержание гликогена на 23, 40 и 44% в волокнах типов I, IIa и IIx соответственно.Уровни внутримышечных триглицеридов (IMTG) были снижены на 27% в волокнах типа I, но оставались постоянными в волокнах типов IIa и IIx. Уровни IMTG в волокнах типа I вернулись к исходному уровню через 2 часа отдыха после тренировки. Волокна типа II ответственны за большую часть гипертрофии, наблюдаемой после тренировки с отягощениями. 24 , 25 Они также страдают от большей утечки энергии во время упражнений с отягощениями. Питание и доступность внутриклеточной энергии существенно влияют на метаболизм мышечного белка.
Потребление углеводов и секреция инсулина косвенно влияют на синтез белка в скелетных мышцах. Инсулин активирует путь фосфоинозитол-3-киназы (PI3K), который заставляет белок транспорта глюкозы скелетных мышц (GLUT4) перемещаться в сарколемму и, следовательно, позволяет глюкозе проникать в клетку. 26 Короче говоря, инсулин восполняет запасы глюкозы в клетке. В пути PI3K активируется белок (Akt), который стимулирует mTOR. Akt также фосфорилирует и инактивирует киназу гликогенсинтазы (GSK-3), что позволяет активировать фактор инициации эукариот 2B (eIF2B).И mTOR, и eIF2B стимулируют синтез белка. Тем не менее, повышенный уровень инсулина не увеличивает синтез белка в отсутствие высоких концентраций аминокислот. 27 Biolo et al. 27 выдвинули гипотезу, что предыдущие эксперименты не смогли продемонстрировать повышенный синтез белка в ответ на инсулин, потому что инсулин расщеплял аминокислоты крови на другие типы клеток, создавая гипоаминоацидемию во время испытаний. В присутствии гипераминоацидемии инсулин, по-видимому, способствует синтезу белка, способствуя проникновению аминокислот в клетки и посредством тангенциальной передачи сигналов, возникающей в результате пути PI3K.
Лейцин, вероятно, наиболее влиятельная аминокислота, влияющая на синтез белка в скелетных мышцах. Аминокислоты, особенно лейцин, стимулируют секрецию инсулина β-клетками поджелудочной железы. 28 В печени отсутствуют аминотрансферазы с разветвленной цепью и, следовательно, отсутствует способность значительно окислять аминокислоты с разветвленной цепью, включая лейцин. Потребление аминокислот с разветвленной цепью приводит к повышению уровня лейцина в крови, достигающего периферических тканей, включая скелетные мышцы.Лейцин напрямую стимулирует mTOR, а также косвенно стимулирует mTOR, стимулируя инсулиновый каскад. 22 Типтон и др. 17 открытие, что сывороточный белок значительно увеличивает плазменный и внутримышечный лейцин, может объяснить потенциальный анаболический эффект потребления сыворотки. Потребление сывороточного протеина повышает лейцин; лейцин прямо и косвенно стимулирует синтез белка через mTOR.
Для достижения наилучшего ответа на упражнения врачи-хиропрактики должны учитывать время потребления белка, тип потребляемого белка и количество потребляемого белка — все это играет важную роль в содействии адаптации за счет синтеза белка.
Чрезмерное потребление протеина
Опасения по поводу чрезмерного потребления протеина действительны. Дезаминирование аминокислот приводит к образованию аммиака. Аммиак токсичен, особенно для центральной нервной системы. Основным путем выведения аммиака является образование и выведение мочевины. Можно потреблять белок, превышающий способность организма справляться с ним. Рудман и др. 29 обнаружили, что максимальная скорость выведения мочевины составила 55 мг мочевины N • ч -1 • кг -0.75 . В своем обзоре диетического потребления белка людьми Билсборо и Манн 30 утверждают, что человек весом 80 кг может дезаминировать до 301 грамма белка в день. На ранее указанных уровнях 7 , 9 , 11 , обеспечивающих максимальную скорость синтеза белка, количества, превышающие 300 г в день, были бы совершенно ненужными. Тем не менее, Phillips 11 et al. анализ избыточного потребления протеина не обнаруживает серьезных последствий при потреблении до 3 г протеина на килограмм массы тела.Увеличение количества диетического белка связано с увеличением пиковой костной массы, но не связано с прогрессирующим снижением функции почек. Продолжение исследований относительно избыточного потребления белка оправдано, но только ошибочные рассуждения позволят сделать вывод о потребности в более 300 граммах белка в день.
Заключение
Эмпирические данные подчеркивают важную роль потребления белка в стимулировании синтеза белка после упражнений с отягощениями. Множественные источники протеина способствуют синтезу протеина после тренировки, но только те, которые содержат незаменимые аминокислоты, повышают синтез.Белки молока, по-видимому, более эффективны, чем белки сои. 15 Сывороточный протеин может способствовать важным изменениям клеточной сигнализации благодаря своей способности повышать уровень лейцина в плазме и внутри клетки. 17 Сывороточный протеин также способствует внутримышечному накоплению триглицеридов, подавляя накопление жира в печени. 16 Потребление углеводов может иметь важное значение для облегчения внутриклеточной передачи сигналов через инсулин, а также для повышения и поддержания энергетического статуса клетки.Тем не менее, важно понимать, что значительный синтез белка маловероятен с добавкой, содержащей только углеводы. Потребление протеина до и после тренировки кажется оправданным. 18 Десяти граммов незаменимых аминокислот или двадцати пяти граммов полноценного белка достаточно, чтобы максимально стимулировать синтез белка. 11 Тип, время и количество белка — все это факторы, влияющие на увеличение мышечной массы.
Еще многое предстоит открыть, что поможет в укреплении здоровья и благополучия.Мы начинаем больше понимать роль скелетных мышц в здравоохранении. Признание необходимости создания синергии между упражнениями и диетой имеет решающее значение. Многие хиропрактики дают пациентам советы по питанию и делают упражнения. Признание и применение синергетической взаимосвязи между упражнениями и диетой может помочь в достижении лучшей адаптации к упражнениям. Для врачей-хиропрактиков адаптацию можно увидеть в объективных измерениях силы, выносливости или функциональных способностей пациента.
Биохимия, синтез белков — StatPearls
Введение
Наше понимание каждой из биологических наук улучшается благодаря изучению биохимии и молекулярной биологии. За последние несколько десятилетий успехи в лабораторных методах изучения этих микроскопических наук привели нас к большему пониманию центральной догмы молекулярной биологии — ДНК транскрибирует РНК, которая затем переводится в белок. Понимание синтеза белка имеет первостепенное значение при изучении различных областей медицины, от молекулярных основ генетических заболеваний и разработки антибиотиков до экспрессии рекомбинантных белков в виде лекарств или клинических лабораторных реагентов.Синтез белка как одна из основополагающих концепций в биологии достаточно сложен, и многие полагают, что он возник однажды, давая механизм синтеза белка во всех организмах на планете общего предка. Несмотря на определенное сходство в их механизме, синтез белка в трех основных линиях происхождения (бактерии, археи и эукариоты) разошелся до такой степени, что возникли существенные механистические различия. Эти различия используются в природе, поскольку организмы производят соединения, нацеленные на белковый синтетический аппарат конкурентов, поскольку они соперничают за ограниченные ресурсы.Наука модифицировала многие из этих соединений, которые нацелены на механизмы синтеза белка у патогенных микробов, для использования в клинике в качестве антибиотиков. По мере того, как наше понимание механизмов синтеза белка продолжает расти, вероятно, появятся бесчисленные дополнительные приложения для этих знаний в медицине, исследованиях и промышленности.
Основы
Синтез белка включает сложное взаимодействие многих макромолекул.
-
Рибосомы:
-
- Рибосома эукариот состоит из двух субъединиц: малой субъединицы 40S и большой субъединицы 60S.Вместе эукариотическая рибосома составляет 80S. Существует несколько сайтов, имеющих функциональное значение, но наиболее важными из них являются сайты A (аминоацил), P (пептидил) и E (выход). Рибосома эукариот представляет собой комплекс рибонуклеопротеидов, состоящий из 4 РНК и 80 белков. Многие функции рибосомы, включая катализирование образования пептидных связей, приписываются рибосомной РНК (рРНК), а не рибосомным белкам, которые вместо этого играют основную роль в сборке субъединиц. Рибосомы могут быть либо прикрепленными к мембранам эндоплазматического ретикулума, либо свободными в цитоплазме.[1]
-
Бактериальные рибосомы имеют две субъединицы, 30S и 50S, которые соединяются, образуя частицу 70S. В целом бактериальные рибосомы меньше, чем их эукариотические аналоги, включая меньше рибосомных белков (55) и более короткие рРНК (всего 3). Определенные области рРНК и некоторые рибосомные белки остаются консервативными между бактериями и эукариотами. Другие области рРНК и белков уникальны как для эукариот, так и для бактерий и частично объясняют различия в механизмах синтеза белка, о которых говорилось выше.
-
-
Эукариотические клетки содержат второй тип рибосомы, обнаруженный в митохондрии, который поддерживает систему синтеза белка, отличную от той, что обнаруживается в цитоплазме. Несмотря на их присутствие в эукариотических клетках, происхождение митохондриальной рибосомы можно проследить до бактерий, что согласуется с эндосимбионтной теорией происхождения митохондрий. Во время разработки антибиотиков необходимо проявлять осторожность, чтобы избежать нацеливания на характеристики митохондриальной рибосомы, общей с бактериальными рибосомами.
-
- Информационная РНК (мРНК): мРНК представляет собой другой тип рибонуклеиновой кислоты, которая функционирует, чтобы нести кодирующую часть гена синтеза белка. Он содержит части некодирующих и кодирующих последовательностей. Кодирующая последовательность группирует нуклеотиды в кодоны, которые представляют собой три конкретных нуклеотида, которые соответствуют определенной аминокислоте, указанной генетическим кодом. [2]
- РНК-переносчик (тРНК): тРНК представляют собой адаптеры, соединяющие нуклеотидную последовательность, обнаруженную в мРНК, с аминокислотной последовательностью, обнаруживаемой в растущем белке.РНК переноса предполагают подобную клеверному листу вторичную структуру с аминокислотой, связанной с ее 3 ’концом через сложноэфирную связь и участок из трех нуклеотидов в основании клеверного листа, называемый антикодоном. Три основания пары оснований антикодона с комплементарными последовательностями кодонов в мРНК в процессе синтеза белка. Это взаимодействие пар оснований играет решающую роль в считывании генетического кода с мРНК на белок. Существует 20 различных аминоацил-тРНК синтетаз, по одной для каждой из 20 распространенных аминокислот.Как только аминокислота связывается со своей родственной тРНК, ее называют аминоацильной тРНК, или «заряженной» тРНК. [3]
- Генетический код: Последовательность генетического кода состоит из трех нуклеотидов, изначально кодированных геномом организма, который определяет отдельные аминокислоты, обнаруженные в белках. Существует 20 общих аминокислот, используемых аппаратом синтеза белков, и 64 возможных перестановки последовательностей четырех оснований, используемых для определения 20 аминокислот. Ранние исследования показали, что код был вырожденным, и многие аминокислоты определялись несколькими комбинациями из трех оснований.В общем, когда несколько кодонов определяют одну аминокислоту, вырожденность обнаруживается в третьем или «колеблющемся» положении. [1] Шестьдесят одна из 64 перестановок последовательностей определяет аминокислоты, тогда как три перестановки последовательности служат «стоп-кодонами» для прекращения синтеза белка. Первоначально считалось, что это одно и то же для всех живых организмов, теперь ученые знают, что существует небольшое количество отклонений от универсального кода, обнаруженного в митохондриях и конкретных видах бактерий.
-
-
Генетический код и болезнь человека: Надлежащее считывание генетического кода имеет важное значение для здоровья человека.Мутации, которые изменяют кодирующие белки последовательности, могут влиять на белки по-разному. Влияние мутаций на кодирующую последовательность можно классифицировать как синонимичные или несинонимичные, в зависимости от того, изменяют ли они первичную структуру белка.
-
-
Синонимичные мутации связаны с вырожденностью кода и тем фактом, что изменения в последовательности оснований могут не влиять на то, какую аминокислоту представляет кодон (хотя следует отметить, что некоторые синонимичные мутации могут влиять на пре-мРНК. сплайсинга и таким образом влияют на первичную структуру белка).Синонимичные мутации обычно попадают в третью позицию кодона.
-
Несинонимичные мутации делятся на три разных класса:
-
-
Миссенс-мутации, при которых происходит замена одной аминокислоты на другую.
-
Нонсенс-мутации, которые вводят кодон преждевременной терминации в последовательность мРНК. Эти мутации обычно приводят к усеченному белку.
-
Мутации со сдвигом рамки являются результатом инсерционных или делеционных мутаций, которые сдвигают рамку считывания кодирующей последовательности, так что секвенирование ниже мутационного события больше не кодирует правильную аминокислотную последовательность белка.
-
-
-
- Белковые факторы — процесс синтеза белка требует множества нерибосомных белков, которые временно участвуют во время фаз инициации, удлинения и завершения синтеза белка. Эти факторы названы в честь фазы, в которой они функционируют (например, фактор инициации эукариот 2, eIF2). [2]
Проблемы, вызывающие озабоченность
Белки, которые экспрессирует клетка, являются окончательным проявлением ее фенотипа.Клетки в тканях человеческого тела имеют различную фенотипическую экспрессию, участвующую в определении организации и функции тканей, несмотря на то, что они имеют идентичные геномы из-за различной экспрессии генов в геноме. Хотя дифференциальная регуляция экспрессии генов в основном происходит на уровне транскрипции, регуляция экспрессии генов также может происходить на посттранскрипционном уровне, включая регулируемую трансляцию. Из-за важности экспрессии белка для фенотипических свойств клетки ошибки в клеточном протеоме, проявляющиеся на всех уровнях правильного считывания генетической информации от гена к белку, могут иметь широкие последствия для здоровья.
Клеточная
Эукариотическая клетка разделена на отдельные части, с различными клеточными компартментами, определенными биологическими мембранами. Синтез компонентов трансляционного аппарата начинается с транскрипции мРНК, тРНК и рРНК в ядре РНК-полимеразами II, III и I, соответственно. Трансферные РНК и мРНК, кодирующие рибосомные белки, выходят из ядра, а последние транслируются в цитоплазму. Затем рибосомные белки возвращаются в ядро, где они иерархически собираются на рРНК, транскрибируемых РНК-полимеразой I.Этот процесс сборки определяет отсек ядра, называемый ядрышком. Сборка рибосом — сложный процесс, в котором задействованы сотни дополнительных факторов, которые временно связываются с рибосомными субъединицами во время их созревания. Хотя большинство этапов созревания рибосомных субъединиц происходит внутри ядрышка до того, как субъединицы выходят через ядерные поры, заключительные этапы созревания субъединиц происходят в цитоплазме. Рибосомы, транслирующие большинство клеточных мРНК, делают это как свободные рибосомы в цитоплазме.Напротив, рибосомы, транслирующие мРНК, кодирующие белки, предназначенные для секреции из клетки или резидентные белки эндоплазматического ретикулума, аппарата Гольджи, лизосомы или плазматической мембраны, локализуются на мембране эндоплазматического ретикулума. [4]
Механизм
Вкратце, трансляцию можно разбить на три фазы: инициирование, удлинение и завершение. Инициация заключается в идентификации точного сайта в последовательности нуклеотидов мРНК, с которого начинается трансляция. Этот процесс имеет существенные различия между эукариотами (описанными здесь) и прокариотами.После идентификации стартового сайта для трансляции происходит удлинение по мере того, как рибосома перемещается по группам «считывания» мРНК из трех нуклеотидов, которые определяют каждую аминокислоту, добавленную к растущей полипептидной цепи. Наконец, терминация происходит, когда рибосома встречает один из трех кодонов терминации, и завершенный белок высвобождается из рибосомы.
Трансляция начинается со сборки комплекса инициации 80S на мРНК. Этот процесс включает определение подходящего кодона для инициации трансляции.Кодон AUG определяет аминокислоту метионин, и практически все белки, определенные генетическим кодом, начинаются с метионина. У эукариот AUG, используемый для инициации синтеза белка, обычно является первым AUG ниже кэп-структуры, обнаруживаемым на 5 ’конце мРНК. Белковый комплекс, известный как eIF4F, распознает структуру кэпа. Комплекс eIF4F затем рекрутирует преинициативный комплекс 43S, состоящий из субъединиц 40S вместе с тройным комплексом, образованным тРНК инициатора (Met-тРНК), eIF2 и GTP на 5 ’конце мРНК.Комплекс 40S впоследствии сканирует мРНК до тех пор, пока не встретит первый AUG и формы инициирующего комплекса 48S. В дополнение к eIF4F и eIF2, множество других факторов инициации облегчают образование комплекса инициации 48S. В этот момент рибосомная субъединица 60S присоединяется к инициирующему комплексу 48S, все факторы инициации высвобождаются, и начинается фаза элонгации трансляции. В инициирующем комплексе 80S инициатор Met-тРНК спарен по основанию с инициирующим AUG в рибосомном Р-сайте со следующим кодоном мРНК, расположенным в рибосомном А-сайте.Облегчение повторной инициации трансляции происходит за счет взаимодействия комплекса eIF4F как с 5 ’кэпом, так и с 3’ polyA-хвостом мРНК. [5]
Как и в случае инициации, удлинение требует использования не рибосомных белков, известных как факторы элонгации. Эукариотический EF1A (eEF1A) образует тройные комплексы с аминоацил-тРНК и GTP. Эти тройные комплексы входят в пустой сайт А рибосомы, и если соответствующее взаимодействие кодон-антикодон образуется между поступающей аминоацил-тРНК и кодоном в сайте А, GTP будет гидролизоваться и высвобождаться eEF1A.На этом этапе сайт пептидилтрансферазы рибосомы катализирует образование пептидной связи, поскольку свободная аминогруппа входящей аминоацил-тРНК атакует сложноэфирную связь, связывающую растущий полипептид с тРНК в Р-сайте рибосомы. Образовавшаяся незаряженная тРНК, занимающая сайт P, перемещается в сайт E (выход) и покидает рибосому. Растущая полипептидная цепь, ранее находившаяся в сайте P, теперь удлиняется на одну аминокислоту, поскольку она переносится на аминоацил-тРНК в сайте A. Пептидил-тРНК в сайте A затем перемещается обратно в сайт P с помощью eEF2 и GTP.Сайт A теперь пуст, и весь процесс повторяется снова и снова, когда рибосома движется вниз по мРНК.
Терминация происходит, когда eRF1, фактор высвобождения, структурно аналогичный тРНК, распознает кодоны терминации в мРНК и рекрутирует eRF3 для гидролиза полипептидной цепи из тРНК, занимающей P-сайт. Прекращение трансляции завершается диссоциацией рибосомных субъединиц, которые теперь способны инициировать еще один цикл синтеза белка. Несколько рибосом могут транслировать одну мРНК, одновременно образуя комплексы, известные как полисомы.[5] [6] [7]
Тестирование
Существует множество возможных методов подтверждения того, что конкретный белок синтезируется.
Иммуноокрашивание
Из-за большого количества белков, синтезируемых в типичной клетке, проверка наличия конкретного белка по понятным причинам является сложной задачей. Один из способов подтвердить присутствие определенного белка в клиническом образце — это иммуноокрашивание. Этот метод вводит антитело к интересующему белку, а исключительная специфичность антитела служит для обнаружения белка.
При иммуноокрашивании образец инкубируют с раствором первичных антител. Это антитело может содержать флуоресцентную молекулу на своей тяжелой цепи или фермент (такой как пероксидаза хрена), который будет флуоресцировать в присутствии подходящего субстрата. Выделяемый свет можно визуализировать под микроскопом или экспонировать на светочувствительной пленке в темной комнате для последующего проявления. Иммуноокрашивание может быть прямым, если первичное антитело обладает средствами флуоресцентного обнаружения, или непрямым, когда вторичное антитело, индуцированное против первичного антитела, выявляется с помощью флуоресценции.[8]
Белковый электрофорез
Как и нуклеиновые кислоты, белки можно разделить по размеру и / или заряду с помощью гель-электрофореза. Белки можно запускать в их естественных конфигурациях или денатурировать перед электрофорезом. При денатурирующем электрофорезе детергент, такой как додецилсульфат натрия (SDS), используется для разрушения сил нековалентного связывания в белках. SDS также придает белкам общее отношение заряда к массе, поэтому единственная сила, действующая во время электрофореза в SDS-полиакриламидном геле, — это молекулярное просеивающее действие полиакриламидного геля.Разделенные таким образом белки можно детектировать неспецифично с помощью красителей, таких как кумасси синий, или специфически с использованием антител в процедуре, называемой вестерн-блоттингом или иммуноблоттингом.
Патофизиология
Многие болезни человека возникают в результате изменений в последовательности белка, вызванных мутациями, которые изменяют правильное считывание генетической информации с гена на функциональный белок. Дефекты в аппарате синтеза белка также вызывают небольшое, но растущее число заболеваний человека.Примеры таких патологий приводятся ниже.
Серповидно-клеточная анемия
Гемоглобин человека содержит две альфа- и две бета-цепи для образования гетеротетрамера. При серповидной анемии шестой кодон бета-цепи содержит миссенс-мутацию, в которой глутаминовая кислота, заряженная аминокислота, заменяется валином, нейтральной аминокислотой. Это различие в одной аминокислоте влияет на третичную и четвертичную структуры гемоглобина, так что в определенных условиях оно искажает двояковогнутую форму эритроцитов в серповидную форму.[9]
Мышечная дистрофия Дюшенна
Подобно многим Х-сцепленным заболеваниям, МДД в первую очередь поражает мужчин в раннем возрасте. Клинически он характеризуется мышечной слабостью, псевдогипертрофией икр и симптомом Гауэра у ребенка. Одним из патофизиологических источников этого заболевания является образование преждевременного стоп-кодона в раннем экзоне гена дистрофина, что приводит к усеченному белку дистрофина, который нарушает целостность саркомера и сократительную функцию мышцы.[10]
Анемия Даймонда-Блэкфана
В то время как многие болезни человека возникают в результате мутаций в кодирующих последовательностях генов, влияющих на выработку белка, анемия Даймонда-Блэкфана (DBA) является одним из растущего числа состояний, являющихся следствием дефектов в протеиновый синтетический аппарат. DBA вызывается аутосомно-доминантными мутациями в генах, кодирующих белки 40S или 60S рибосомной субъединицы. Хотя точные механизмы, лежащие в основе патофизиологии DBA, в настоящее время неизвестны, кажется вероятным, что изменения в клеточных протеомах (белковом составе клетки) в результате неоптимального количества рибосом частично влияют на клинические признаки заболевания.Эти клинические особенности включают дефицит продукции красных кровяных телец, небольшой размер и неоднородное количество врожденных аномалий. [11]
Клиническая значимость
Клиническое значение синтеза белка заключается не только в человеческой трансляции, но и в различиях между человеческой и бактериальной трансляцией. Бактериальная рибосома (70S) имеет те же основные компоненты и множество структурно схожих сайтов по сравнению с эукариотической рибосомой (80S). Однако трансляционные различия между людьми и бактериями создают мишени для противомикробных препаратов.Эти различия позволяют определенным антибиотикам избирательно связываться с бактериальными рибосомами в низких концентрациях, избирательно воздействуя на бактерии и либо подавляя рост, либо убивая микроб. Несколько обычно назначаемых антибиотиков нацелены на определенные компоненты бактериальной рибосомы и мРНК. Аминогликозиды нацелены на малую 30S субъединицу рибосомы; в частности, этот класс связывается с сегментом рРНК, активным в A-сайте. Тетрациклины действуют аналогичным образом, конкурируя за сайт A с заряженной аминоацил тРНК.Макролидные антибиотики действуют на большую рибосомную субъединицу 50S. Когда они связываются с рРНК большой субъединицы, это предотвращает образование пептидной связи и способствует раннему изгнанию тРНК в P-сайте. [12] [3]
Клинические проявления различий в синтезе белка также могут быть полезны в диагностике. Электрофорез нативных белков может помочь выявить гемоглобинопатии при обследовании новорожденных. Точно так же электрофорез сывороточного белка может идентифицировать характерные пики М-белка при экспрессии моноклонального белка при множественной миеломе.
Непрерывное обучение / обзорные вопросы
Ссылки
- 1.
- Schmeing TM, Ramakrishnan V. Что недавние структуры рибосом открыли о механизме трансляции. Природа. 29 октября 2009 г .; 461 (7268): 1234-42. [PubMed: 19838167]
- 2.
- Chee NT, Lohse I, Brothers SP. Трансляция мРНК в белок при гипоксии. Молочный рак. 30 марта 2019; 18 (1): 49. [Бесплатная статья PMC: PMC6441220] [PubMed: 30925920]
- 3.
- Хариш А., Каэтано-Аноллес Г.История рибосом раскрывает истоки современного синтеза белка. PLoS One. 2012; 7 (3): e32776. [Бесплатная статья PMC: PMC3299690] [PubMed: 22427882]
- 4.
- Tolsma TO, Hansen JC. Посттрансляционные модификации и динамика хроматина. Очерки Биохимии. 2019 апр 23; 63 (1): 89-96. [PubMed: 31015385]
- 5.
- Simonović M, Steitz TA. Структурный взгляд на механизм образования пептидной связи, катализируемой рибосомами. Biochim Biophys Acta. 2009 сентябрь-октябрь; 1789 (9-10): 612-23.[Бесплатная статья PMC: PMC2783306] [PubMed: 19595805]
- 6.
- Gao H, Zhou Z, Rawat U, Huang C, Bouakaz L, Wang C, Cheng Z, Liu Y, Zavialov A, Gursky R, Sanyal S , Эренберг М., Франк Дж., Сонг Х. RF3 индуцирует рибосомные конформационные изменения, ответственные за диссоциацию факторов высвобождения класса I. Клетка. 1 июня 2007 г .; 129 (5): 929-41. [PubMed: 17540173]
- 7.
- Nissen P, Hansen J, Ban N, Moore PB, Steitz TA. Структурные основы активности рибосом в синтезе пептидных связей.Наука. 2000, 11 августа; 289 (5481): 920-30. [PubMed: 10937990]
- 8.
- Medzihradszky KF, Chalkley RJ. Уроки секвенирования пептидов de novo методом тандемной масс-спектрометрии. Mass Spectrom Rev.2015, январь-февраль; 34 (1): 43-63. [Бесплатная статья PMC: PMC4367481] [PubMed: 25667941]
- 9.
- Azar S, Wong TE. Серповидноклеточная болезнь: краткое обновление. Med Clin North Am. 2017 Март; 101 (2): 375-393. [PubMed: 28189177]
- 10.
- Aartsma-Rus A, den Dunnen JT. Прогнозирование фенотипа для делеций / дупликаций экзонов: руководство пользователя для профессионалов и клиницистов, использующих в качестве примеров мышечную дистрофию Беккера и Дюшенна.Hum Mutat. 2019 Октябрь; 40 (10): 1630-1633. [PubMed: 31356707]
- 11.
- Ellis SR, Gleizes PE. Анемия Даймонда Блэкфана: рибосомные белки выходят из строя. Semin Hematol. 2011 Апрель; 48 (2): 89-96. [PubMed: 21435505]
- 12.
- Витцки А., Толлерсон Р., Ибба М. Трансляционный контроль устойчивости к антибиотикам. Откройте Биол. 2019 26 июля; 9 (7): 1
. [Бесплатная статья PMC: PMC6685928] [PubMed: 31288624]
Какова цель синтеза белка? — Видео и стенограмма урока
Транскрипция и трансляция
ДНК — это окончательный проект клетки, содержащий все инструкции по созданию белков.Эта молекула настолько важна, что клетка хочет делать ее копии для использования в синтезе белка, вместо того, чтобы перемещать настоящую ДНК по клетке. Думайте о ДНК как о главной копии документа, как о свидетельстве о рождении. Чтобы получить другие документы, удостоверяющие личность, вы предъявляете копию свидетельства о рождении, но не настоящее.
Копия ДНК называется мРНК (или информационная РНК), еще одна молекула, которая хранит информацию в клетке. Процесс транскрипции копирует ДНК в мРНК с помощью белка, называемого РНК-полимеразой.После копирования мРНК отправляется в отсек клетки, называемый рибосомой, который выполняет следующий шаг — трансляцию.
Рибосомы — это части клетки, необходимые для синтеза белка. Во время трансляции они считывают мРНК и приказывают другой молекуле, называемой тРНК (или транспортной РНК), получить строительные блоки для белков или аминокислоты . Рибосома связывает аминокислоты в соответствии с инструкциями в мРНК. После того, как все аминокислоты соединены вместе, белок складывается в функциональную форму.
Белки, как мы уже упоминали, отвечают за всю структуру и функции внутри клеток. Белки нужны всем клеткам, а не только нашим клеткам. Итак, теперь давайте рассмотрим несколько ключевых примеров на различных типах организмов.
Синтез белка у животных
Человек — это вид животных, поэтому наши клетки считаются животными клетками. Чтобы понять, насколько важны белки в наших клетках, давайте рассмотрим пример того, что происходит, если возникает проблема с синтезом белка. Белок, называемый Rb, с полным названием белок ретинобластомы, отвечает за то, как часто наши клетки делятся, особенно в клетках сетчатки, важной части наших глаз.Если клетки бесконтрольно делятся, мы получаем рак. У некоторых людей есть мутация или изменение ДНК в инструкциях по созданию Rb. У этих людей нет функционального Rb-белка, поэтому их клетки бесконтрольно разрастаются, и они заболевают ретинобластомой или раком глаза.
Синтез белка в растениях
Растения являются важной частью нашей экосистемы, поскольку все другие организмы зависят от растений в получении энергии. Растения используют процесс, называемый фотосинтезом, для превращения углекислого газа, воды и солнечного света в кислород и сахар, называемый глюкозой.Это происходит в результате серии химических реакций, которые становятся возможными благодаря белкам.
Конкретный белок под названием RuBisCo (рибулозо-1,5-бисфосфаткарбоксилаза / оксигеназа) включает двуокись углерода из воздуха в молекулы внутри клетки, которые в конечном итоге будут производить глюкозу. Без глюкозы люди и все другие организмы, которые не могут производить себе пищу, умрут. Когда растения отмирают, у нас больше не остается кислорода, который также является продуктом фотосинтеза. Ясно, что, несмотря на то, что он не является микроскопическим, RuBisCo необходим для всей жизни на Земле.
Синтез белка в бактериях
Как микроскопические одноклеточные организмы, мы часто пренебрегаем бактериями как важной частью экосистемы. Но, имея более 10 000 видов, они контролируют больше, чем мы думаем. Хотя многие бактерии полезны для нас, например, бактерии в кишечнике, расщепляющие растительный материал, некоторые бактерии являются патогенными или вызывают заболевания. Давайте посмотрим на пример того, как белки помогают бактериям выжить за наш счет.
Бактерия под названием MRSA (или устойчивый к метициллину Staphylococcus aureus ) особенно опасна, и инфекции становятся все более распространенными в больницах, где много больных людей и вводят много антибиотиков.MRSA устойчив ко многим антибиотикам, а это значит, что антибиотики больше не убивают его.
Устойчивость к антибиотикам очень опасна для пациентов, потому что существует меньше вариантов лечения. Бактерии делают это несколькими способами, но один из них — это белок, называемый насосом, который выводит антибиотики из клетки.
Эти белки находятся на мембране или внешнем барьере бактериальной клетки. Они используют энергию и выводят антибиотики из клетки по мере поступления, не позволяя антибиотикам выполнять свою работу.Многие лекарственные препараты для лечения бактериальных инфекций на самом деле сосредоточены на блокировании синтеза белка, например, хлорамфеникол.
Резюме урока
Синтез белка — это процесс, который используют все клетки для производства белков, которые отвечают за всю структуру и функции клетки. Есть два основных шага к синтезу белка. В транскрипции ДНК копируется в мРНК, которая используется в качестве матрицы для инструкций по созданию белка.