Дефицит магния и калия в организме: симптомы и признаки дефицита
Домашняя аптечка MIRRA
В последние годы в Компании активно проводились исследования, посвященные вопросам возможности применения оздоровительной и косметической продукции для решения общих и косметологических проблем. В этом направлении работали практически все ведущие сотрудники Компании – И. Рудаков (д. м. н.), С. Чубатова (д. б. н.), Е. Аитова (к. б. н.), Л. Кондэ (к. м. н.), А. Голубков (к. х. н.), директор производства В. Тульский, главный технолог Б. Городничев. Также в научных разработках стали принимать участие консультанты и дистрибьюторы некоторых РЦ. С 2010 года было опубликовано множество научных статей, практических руководств и справочников по продукции, в которых рассматривался ряд современных косметологических (космецевтических) концепций и необходимость создания новой продукции, а также идеи о том, что косметические и оздоровительные средства необходимо использовать в комплексе. Результатом проведенных научно-практических работ стала разработка метода информационного тестирования для оценки микронутриентной обеспеченности (МО) организма. Данная информация опубликована в номерах нашего ежемесячника в рубрике «Домашняя аптечка MIRRA».
Результатом проведенных научно-практических работ стала разработка метода информационного тестирования для оценки микронутриентной обеспеченности (МО) организма. Данная информация опубликована в номерах нашего ежемесячника в рубрике «Домашняя аптечка MIRRA».
Эта публикация посвящена дефициту калия и магния.
КАЛИЙ (K)
Калий содержится во многих продуктах питания и усваивается на 90−95 %. В сутки в среднем он поступает в организм с пищей в количестве 1−3 г. В теле человека суммарно присутствует 160−180 г калия. Ниже представлен опросник, содержащий перечень негативных состояний (НС). Их причиной может быть дефицит калия, а точнее, недостаточная микронутриентная обеспеченность. Чтобы понять, существует ли дефицит калия в организме, нужно выделить свойственные состоянию Вашего здоровья НС и в зависимости от их количества оценить МО калием.
- Апатичность, безразличие к происходящему вокруг.
- Склонность к депрессиям.
- Снижение работоспособности.

- Быстрая утомляемость, частая слабость.
- Снижение устойчивости к стрессу.
- Аллергические реакции и болезни.
- Слабость в мышцах.
- Кожный зуд.
- Кожа медленно восстанавливается после порезов и ранок.
- Избыточная сухость кожи.
- Избыточное шелушение и шершавость кожи.
- Сильное выпадение волос.
- Утрата гибкости волосами.
- Волосы стали более тонкими.
- Потускнение волос.
- Слизистые рта и языка воспалены.
- Склонность к диарее.
- Нарушение сердечного ритма, боли в сердце.
- Изменение артериального давления.
- Наклонность к воспалительным процессам с образованием эрозии (гастрит, эрозия шейки матки).
Полученные результаты позволяют судить о степени обеспеченности организма калием. Если НС 6 и менее, это свидетельствует о нормальном уровне МО. При выборе 7−12 НС можно говорить о легком или среднем снижении МО. О дефиците калия в организме могут свидетельствовать НС в количестве 13 и более.
О дефиците калия в организме могут свидетельствовать НС в количестве 13 и более.
Причиной снижения обеспеченности организма калием может стать его недостаточное поступление с пищей. Однако это не единственное объяснение. Так, организм усиленно расходует калий при нервных, психических перегрузках, чрезмерных или хронических стрессах.
Другой важной причиной снижения калиевой МО является его вытеснение в избытке поступающим в организм натрием, прием гормональных препаратов, диуретиков, слабительных средств. Калий быстрее расходуется при нарушении работы выделительных систем (кожи, кишечника, легких, почек).
Таким образом, при выявленном дефиците калия, т. е. снижении МО требуется пересмотреть рацион. При добавлении в него богатых калием продуктов можно восстановить МО. Быстрому восстановлению содержания калия могут способствовать препараты безрецептурного отпуска, содержащие данный микронутриент в форме аспарагината, который хорошо усваивается организмом («Панангин», «Аспаркам»). В их состав также входит аспарагинат магния. Если на организм длительное время воздействуют различные стрессорные факторы, может нарушиться МО не только калием, но и другими веществами, что требует подбора группы средств для ее восстановления.
В их состав также входит аспарагинат магния. Если на организм длительное время воздействуют различные стрессорные факторы, может нарушиться МО не только калием, но и другими веществами, что требует подбора группы средств для ее восстановления.
Если имеет место патология выделительных систем, часто назначается медикаментозное лечение. При нарушении процессов обмена калия на организменном уровне может быть рекомендована регулирующая терапия гомеопатическими средствами. Если МО калием находится в норме, никаких вмешательств проводить не стоит. Если МО сниженная, ее нормализация становится крайне важной, а иногда и жизненно необходимой для организма.
МАГНИЙ (Mg)
В среднем с пищей и водой в сутки в организм поступает 200−400 мг магния, при этом около половины данной дозы усваивается в ЖКТ. Этот микроэлемент является важной составляющей частью каждой клетки, где вместе с кальцием и натрием участвует в обеспечении важнейших внутриклеточных процессов.
Ниже дан перечень негативных состояний (неприятных ощущений, симптомов, заболеваний), которые часто указывают на дефицит магния, или его недостаточную МО. Такое состояние может потребовать коррекции путем нормализации рациона и приема БАД. Посчитайте количество НС, свойственных состоянию Вашего здоровья.
Такое состояние может потребовать коррекции путем нормализации рациона и приема БАД. Посчитайте количество НС, свойственных состоянию Вашего здоровья.
- Повышенная нервная возбудимость.
- Головокружения.
- Тревожность, беспокойство.
- Нарушения сна, бессонница.
- Наклонность к депрессии.
- Повышенная утомляемость, слабость.
- Снижение работоспособности.
- Ухудшение концентрации внимания.
- Снижение устойчивости к стрессу.
- Снижение слуха.
- Иммунодефицитные состояния.
- Выпадение волос.
- Тонкие и ломкие ногти.
- Покалывание и онемение в пальцах рук и ног.
- Подергивания век.
- Судороги и боли в мышцах.
- Колебания артериального давления.
- Нарушения сердечного ритма.
- Боли в сердце.
- Тенденция к диарее или запорам.
Если количество выбранных НС не превышает 6, можно говорить об условно-нормальном уровне МО. На вероятность легкого или среднего снижения МО указывает выбор 7−12 НС. Признаком дефицита магния может стать выделение 13 НС и больше.
На вероятность легкого или среднего снижения МО указывает выбор 7−12 НС. Признаком дефицита магния может стать выделение 13 НС и больше.
Кроме указанных в перечне НС, симптомами дефицита магния (т. е. недостаточной МО) могут стать необъяснимые головные боли, повышенная раздражительность и плаксивость, появление вздорности характера. Из всего комплекса НС очень часто наблюдаются беспокойство, тревожность, снижение усидчивости. На состояние дефицита магния в организме и нарушение его обмена большое влияние оказывает хронический стресс.
Данные факторы не единственные из числа тех, которые могут влиять на нехватку магния. К их числу также можно прибавить воспалительные процессы или дисбактериоз ЖКТ, из-за которых нарушается всасывание полезных веществ. Нарушение обмена магния может стать результатом приема некоторых лекарств. Повышенная потребность в магнии наблюдается в период беременности или выздоровления после длительной болезни. При этом надо исключить вероятность недостаточности магния из-за однообразного рациона, в котором преобладают бедные этим микроэлементом продукты.
Для устранения НС, связанных с дефицитом магния, часто бывает достаточно нормализовать режим питания. Также МО может восстановить прием магний-содержащих препаратов безрецептурного отпуска и БАД. В их состав могут входить магний и калий в форме аспарагинатов («Аспаркам», «Панангин»), или магний и витамин В6, а также другие минерально-витаминные комплексы. Нормальная обеспеченность организма магнием не требует принятия каких-либо мер, но при его недостаточности нормализация данного состояния бывает не только желательной, но и часто необходимой.
И. Рудаков, д. м. н., директор по науке
*При использовании материалов статьи гиперссылка на источник обязательна.
Кальций общий в медицинском центре «Академия здоровья»
Стоимость услуги
90р
Если в организме живого существа недостаточно калия, то начинается рвота, тошнота и мышечная слабость. В некоторых случаях возможно нарушение в работе сердца – сбивается ритм. Для того, чтобы определить концентрацию калия в организме человека, проводится забор крови на биохимический анализ.
Забор материала для лабораторного исследования нужно проводить натощак. При этом, за три дня до предполагаемого забора нужно убрать из потребления жирную пищу, прием антибиотиков (если это целесообразно).
Для нормального количества кальция нужно пить оптимальное для веса количество воды, не употреблять консервированные продукты в большом количестве и заниматься спортом.
Повышенное количество натрия в организме человека может явственно указывать на серьезные недуги. Такое возможно при нефрите, паталогической почечной недостаточности, сахарном диабете и чрезмерному количеству отложения солей. Для выявления количества натрия в организме человека используют анализ мочи. Забор исследуемого материала собирается утром, перед приемом пищи или жидкости, в стерильную посудину. Для точного анализа материал нужно доставить как можно быстрее до лаборатории. Неверные показатели возможны, если посуда была не стерильной или поздно привезли забор.
Неверные показатели возможны, если посуда была не стерильной или поздно привезли забор.
Наиболее распространенным неорганическим элементом для человека является кальций, для диагностического определения объемов которого назначается специализированный биохимический анализ крови. Биологическая роль кальция очень существенна. Так, он отвечает за поддержание нормального сердечного ритма, активно участвует в прямом процессе обмена железа и регулирует ферментную активность, действенно способствует передаче нервных импульсов, оптимальной работе ЦНС, участвует в процедуре сокращения мышц. Помимо того кальций регулирует уровень проницаемости клеточных мембран, деятельно принимает участие в процедуре свертывания крови.
зачем они нужны растениям и как их применять
Виды фосфорных удобрений
Вообще у фосфора несколько функций: он отвечает за рост корневой системы, повышает устойчивость растений к низким температурам, засухам, но главная его задача – стимулировать цветение растений.
Суперфосфат простой
Фосфора 26 %.
Плюсы: Помимо фосфора, содержит азот (6 %), кальций (17 %), серу (10 %) и магний (0,5 %). Прекрасно усваивается растениями.
Минусы: 40 % удобрения состоит из гипса, который бесполезен для растений. То есть, покупая пакет удобрения, вы фактически отдаете половину его стоимости за ненужное вещество.
Когда вносить: осенью под перекопку.
Нельзя смешивать с мочевиной, аммиачной, натриевой и кальциевой селитрой, карбонатом калия, известью.
Полностью безопасное удобрение!
Суперфосфат двойной
Фосфора 40–50 %.
Плюсы: в его составе фосфора в два раза больше, чем в простом суперфосфате. Стоит он чуть дороже обычного, но вносить его нужно в два раза меньше. В итоге получается выгоднее.
Минусов нет!
Когда вносить: осенью или весной под перекопку. Можно вносить в качестве подкормки перед цветением. Но помните: двойной суперфосфат растворяется только на 40 %, поэтому одна только летняя подкормка не даст должного эффекта. Тем более, растения усваивают фосфор постепенно. Подкормки фосфором для однолетних растений не имеют смысла, на урожай он никак не влияет. А вот многолетние удобрить можно – он повысит их морозостойкость. Разбросайте удобрение по поверхности и заделайте в почву граблями (если корневая сисема растений неглубокая) либо перекопайте землю вокруг растения лопатой (если корни глубоко).
Можно вносить в качестве подкормки перед цветением. Но помните: двойной суперфосфат растворяется только на 40 %, поэтому одна только летняя подкормка не даст должного эффекта. Тем более, растения усваивают фосфор постепенно. Подкормки фосфором для однолетних растений не имеют смысла, на урожай он никак не влияет. А вот многолетние удобрить можно – он повысит их морозостойкость. Разбросайте удобрение по поверхности и заделайте в почву граблями (если корневая сисема растений неглубокая) либо перекопайте землю вокруг растения лопатой (если корни глубоко).
Эффективность суперфосфата заметно повышается вкупе с органикой.
Нельзя смешивать с мочевиной, аммиачной, натриевой и кальциевой селитрой, карбонатом калия, известью.
Полностью безопасное удобрение!
Фосфоритная мука
Фосфора 19–30 %.
Плюсы: прекрасное удобрение для кислых почв! Абсолютно безвредно для окружающей среды.
Минусы: фосфор в этом удобрении находится в недоступной для растений форме, высвобождается в почве лишь через несколько месяцев. Поэтому в качестве подкормки не годится.
Поэтому в качестве подкормки не годится.
Но главный минус – это затраты. Килограмм фосфоритной муки стоит на уровне суперфосфата, но вносить ее надо очень много – 30 кг на сотку!
Когда вносить: осенью под перекопку.
Нельзя смешивать с известью, карбонатом калия.
Полностью безопасное удобрение!
В суперфосфате (как простом, так и двойном) фосфор находится в виде фосфорной кислоты. Она практически не передвигается в почве – куда внесли, там и закрепляется. Поэтому эти удобрения надо обязательно заделывать в землю на глубину основной массы корней. Если просто разбросать по поверхности участка – толку никакого не будет.
Виды калийных удобрений
Калий нужен в первую очередь плодам – он повышает количество сахара и витаминов. овощи и фрукты, насыщенные калием, дольше хранятся, растения лучше переносят суровые зимы.
Сульфат калия (сернокислый калий)
Калия 45–53 %.
Плюсы: хорошо растворяется в воде, прекрасно усваивается растениями. Содержит 18 % серы.
Минусы: не для каждой почвы.
Когда вносить: осенью под перекопку. Либо в качестве подкормки для плодовых культур и овощей – в начале плодоношения. Под деревья и кустарники сульфат калия полезно внести в конце августа – начале сентября. Это повысит их зимостойкость.
Нельзя смешивать с мочевиной, аммиачной селитрой, сульфатом аммония, карбонатом калия, известью.
Опасность: подкисляет почву. Использовать только на нейтральных и щелочных участках.
Калимагнезия
Калия 26–28 %, магния – 9–16 %.
Плюсы: растворяется в воде. очень эффективное удобрение на легких почвах, бедных магнием.
Минусов нет!
Когда вносить: осенью под перекопку или весной непосредственно в лунки. Можно в качестве подкормки в начале плодоношения.
Нельзя смешивать с мочевиной, карбонатом калия.
Полностью безопасное удобрение!
Карбонат калия
Калия 52–55 %.
Плюсы: хорошо растворяется в воде, прекрасно усваивается растениями.
Минусы: при высокой влажности воздуха слеживается и даже полностью растворяется. Хранить его нужно только в очень сухих помещениях.
Когда вносить: осенью под перекопку.
Нельзя смешивать вообще ни с чем!
Опасность: сильно подщелачивает грунт. Использовать только на кислых почвах.
Калийная селитра
Калия 46 %.
Плюсы: помимо калия, содержит 13 % азота, который является одним из основных элементов питания растений.
Минусы: одновременно калий и азот доступны растениям только на нейтральных почвах. На кислых азот не работает, а на щелочных мертвым грузом залегает калий.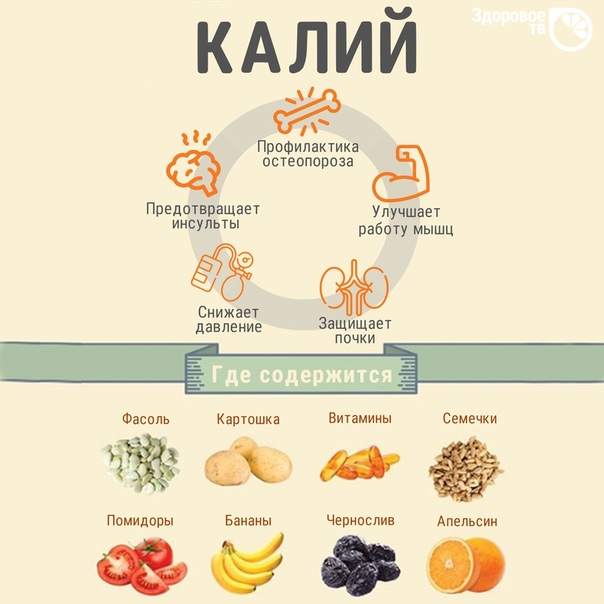
Когда вносить: весной перед посадкой и летом в виде подкормок – в начале плодоношения.
Нельзя смешивать с навозом (может загореться!), сульфатом аммония, карбонатом калия.
Опасность: взрывоопасна при хранении и использовании! Может рвануть, если в нее попадет любое моющее средство или растворитель, а также при высокой температуре в помещении. Хранить ее можно только в прохладном месте и в герметичной таре.
Хлористый калий
Калия 50–60 %.
Плюсы: хорош, если вносить его под свеклу, – она любит соль.
Минусы: в составе вредный для почвы и для растений хлор. Этот элемент на дух не переносят огурцы, перцы, томаты и картофель.
Когда вносить: осенью под перекопку почвы.
Нельзя смешивать с известью, доломитом, мелом, карбонатом калия.
Опасность: содержит хлор.
Виды сезонных удобрений: так ли они хороши?
Сейчас в продаже есть готовые сезонные удобрения, например «осеннее», «Весна-лето», «Лето». Питательные элементы в них сбалансированы в соответствии с потребностями культур в конкретное время года. В весенне-летнем содержится больше азота, в осеннем главные элементы – фосфор и калий.
Питательные элементы в них сбалансированы в соответствии с потребностями культур в конкретное время года. В весенне-летнем содержится больше азота, в осеннем главные элементы – фосфор и калий.
Плюсы: это удобно – не нужно смешивать удобрения, высчитывать количество – в них уже все рассчитано и в инструкции указано, в какой дозе и под какие культуры вносить.
Минус: стоят они в среднем в три раза дороже обычных.
Зола: самое лучшее природное удобрение
Часто золу как удобрение упоминают, когда говорят об органике. На самом же деле это минеральное удобрение. Причем отличное! Зола содержит много калия, фосфора, кальция и практически все нужные растениям микроэлементы: серу, бор, марганец, магний. Стакан золы заменяет 40 г сернокислого калия! При этом она абсолютно безопасна для почвы, растений и людей.
На тяжелых почвах ее лучше вносить под осеннюю перекопку, а на легких – весной. Но и в том и в другом варианте обязательно заделывать в почву на глубину 10 см. Если же оставить ее на поверхности – образуется почвенная корка.
Если же оставить ее на поверхности – образуется почвенная корка.
Если добавить это удобрение в компостную кучу, органика быстрее разложится.
Хранить золу нужно в сухом помещении, поскольку при увлажнении она теряет свою питательную ценность.
Нельзя смешивать с навозом и птичьим пометом – это приводит к потере азота.
Внекорневые подкормки: нужны ли они?
Часто дачникам советуют использовать внекорневые подкормки – в той или иной стадии развития растений. Дескать, они увеличивают урожайность, повышают качество плодов и еще много чего полезного дают. так ли это? Да, так. Но! Эффект у них кратковременный.
Во-первых, концентрацию удобрений для таких подкормок делают меньше – в противном случае можно сжечь листья.
Во-вторых, растения используют их одномоментно и сразу же расходуют, а дальше снова начнут голодать.
Есть и другая проблема. фосфорные удобрения плохо растворяются в воде и могут оставлять налет на листьях. Калийные часто малоэффективны, поскольку ионы калия очень крупные и с трудом проходят внутрь листа.
Калийные часто малоэффективны, поскольку ионы калия очень крупные и с трудом проходят внутрь листа.
Если вы внесли фосфор и калий осенью или весной в рекомендованных количествах, внекорневые подкормки не нужны. Это хлопотно и может быть опасно – чуть переборщите с дозой, растения погибнут. Внекорневые подкормки рядовому дачнику могут
помочь только в одном случае – в прохладную погоду, когда температура почвы понижается и корни не способны добывать себе элементы питания из почвы.
применение, инструкция, состав, полезные советы
Содержание:
- За что отвечает калий
- Как распознать дефицит калия
- Для каких культур и когда использовать
- Способы применения
Плодородная почва в процессе многолетнего использования теряет полезные вещества, а если речь о суспесях и суглинках — получить хороший урожай без удобрений вряд ли удастся. Основные элементы, необходимые всем культурам, — азот, фосфор и калий. Устранить дефицит последнего удается с помощью удобрения сульфата калия. Сернокислый калий (калия сульфат) используют для обогащения почвы в теплицах и под открытым небом, стимулируя правильный рост и активное плодоношение различных садово-огородных культур.
Устранить дефицит последнего удается с помощью удобрения сульфата калия. Сернокислый калий (калия сульфат) используют для обогащения почвы в теплицах и под открытым небом, стимулируя правильный рост и активное плодоношение различных садово-огородных культур.
Удобрение выпускается в виде белого порошка с серым оттенком. Кристаллы калия сульфата (удобрения) имеют кисло-горький вкус, хорошо растворяются в воде. Главным элементом состава является калий (50 %), остальное — сера, магний и кальций. Противники хлора могут не опасаться — в этом удобрении его нет. Поэтому можно смело применять его на грядках, где выращивается картофель и бобовые культуры, которые не любят хлор.
Относительно недавно в роли источника калия выступала древесная зола. Учитывая, что все меньше людей используют в хозяйстве дровяные печи, проще заказать в магазине упаковку готовой подкормки весом 0,5–5 кг. Готовую смесь легче дозировать, подбирая нужную концентрацию.
За что отвечает калий
Калий, как один из трех главных элементов для развития растений, отвечает за их рост и развитие и способствует повышению урожайности. Благодаря калию в клетках растений происходят обменные процессы, культуры приобретают стойкость к вредителям и болезням, зимним морозам. В плодах повышается количество витаминов и веществ, полезных человеку. Поэтому оценивая целесообразность применения разных добавок на огороде, удобрение сульфат калия используют без сомнений, и не только в теплицах и в открытом грунте, но и для выращивания комнатных цветов.
Благодаря калию в клетках растений происходят обменные процессы, культуры приобретают стойкость к вредителям и болезням, зимним морозам. В плодах повышается количество витаминов и веществ, полезных человеку. Поэтому оценивая целесообразность применения разных добавок на огороде, удобрение сульфат калия используют без сомнений, и не только в теплицах и в открытом грунте, но и для выращивания комнатных цветов.
Калий участвует в белковом и углеводном обмене растений, активизирует аммиачный азот, увеличивает водоудерживающую способность и стойкость растений к увяданию. Дефицит калия подавляет в растении перемещение углеводов, синтез белка и фотосинтез. Общие рекомендации сводятся к обогащению почвы, на которой выращиваются культуры из семейства крестоцветных. Очень любят калий перцы, баклажаны и огурцы, не помешает использовать подкормку для клубники, смородины, малины, других кустарников и плодовых деревьев.
Под удобрение сульфатом калия подходят любые почвы — сероземные, песчаные, торфяные, подзолистые и др. Чтобы вещество лучше усваивалось в суглинках, рекомендуется вносить его ближе к корням растений.
Чтобы вещество лучше усваивалось в суглинках, рекомендуется вносить его ближе к корням растений.
Как распознать дефицит калия
Целесообразность применения в виде удобрения калия сульфата не вызывает сомнений, но правильный подход подразумевает поддержание баланса полезных веществ в почве — избыток не менее вреден, чем недостаток. Желательно раз в 2 года проводить анализ состава грунта, чтобы точнее вносить подкормки и избежать переизбытка некоторых веществ.
Распознать нехватку калия можно двумя способами — заметить явные признаки во время развития растений или столкнуться со значительным снижением урожайности. Первый способ предпочтительнее, поскольку позволяет вовремя предпринять меры и не допустить гибели растений и снижения плодоношения.
Основные признаки дефицита калия:
- зеленая часть растения замедляет рост;
- листья желтеют, деформируются и не могут развиваться полноценно;
- листовые пластины покрываются точками или пятнами;
- бутоны засыхают, не раскрывшись;
- растение менее стойко переносит холода;
- урожай хранится плохо;
- вкус плодов ухудшается.
Для каких культур и когда использовать подкормку
Выбор в пользу удобрения сульфата калия обычно не оспаривается, вещество необходимо разным культурам. Есть такие, для которых подкормка жизненно необходима. Это бобовые с их потребностью в увеличенном количестве серы, крестоцветные (редиска, капуста и др.), корнеплоды, плодовые деревья и кустарники, а также растения, которым не нравится хлор (табак, цитрусовые, картофель).
Объем удобрений и целесообразность их применения зависят от состава почвы. Калия сульфат можно использовать комплексно с другими добавками. Такой подход решает сразу несколько задач — насыщает грунт полезными веществами и отпугивает вредителей. Если в точности следовать инструкции и не превышать рекомендованные дозировки, урожай будет полезным и безопасным для человека.
В отношении кислых грунтов рекомендуется вносить удобрение калия сульфат одновременно с известью, чтобы подкормка работала эффективнее. Не стоит совмещать калий с карбамидом и мелом — такое соседство не даст результатов. Сернокислый калий не используют, если почву удобрили компостом или навозом — органика уже содержит достаточное количество этого элемента, поэтому нет нужды в его дополнительном внесении.
Способы применения
Использовать сульфат калия можно как в сухом виде, так и в виде раствора, поливая растения под корень или опрыскивая по листу. В каждом случае полезные элементы хорошо усваиваются культурами, но лучше непосредственно удобрять грунт. Так растение получает необходимые вещества, а почва обогащается.
Основные способы применения сводятся к следующим:
- Весной или в канун осенней перекопки разбрасывают сухую подкормку по участку.
- Во время перекопки сухие кристаллы заделываются в почву на глубину, где будут располагаться корни растений.
- При посадке деревьев в яму добавляют сульфат калия и фосфорную подкормку.
- Летом используют полив раствором сульфата калия под корень, деревья можно удобрять через отверстия, расположенные под углом 45 градусов к корням.
В магазинах агрохимии предлагаются комплексные составы, в которых сульфат калия объединяют с фосфором. Оба вещества отлично работают в тандеме, действуют долго. Узнать больше о роли фосфора в развитии растений можно из других статей, размещенных в блоге.
Значение калия для растений и его содержание в почве
Оглавление
Значение калия для растений
Физиологические функции калия
Калий в почве
Необменно-поглощенный (фиксированный) калий
Обменно-поглощенный калий
Водорастворимый калий
Значение калия для растений
Калий является одним из основных элементов питания, наряду с азотом и фосфором. Функция калия в растениях, как и других необходимых для них элементов, строго специфична.
Первые предположения о необходимости калия растениям выразил Ссср в 1804 г. на основании анализа золы растений, в которой всегда присутствовал калий. Затем Либих сделал вывод о необходимости применения калийных удобрений. Первые экспериментальные данные об абсолютной необходимости калия растениям были получены Сальм-Хорстмар в 1846 г.
В отличие от азота и фосфора калий не входит в состав органических соединений в растениях, а находится в клетках растений в ионной форме в виде растворимых солей в клеточном соке и частично в виде недолговечных комплексов с коллоидами цитоплазмы.
Калия значительно больше в молодых жизнеспособных частях и органах растений, чем в старых. Около 80% калия находится в клеточном соке и может легко вымываться водой (дождями и при поливе). Молодые органы растения содержат калия в 3-5 раз больше, чем старые: его больше в тех органах и тканях, где идут интенсивно процессы обмена веществ и деления клеток. При недостатке калия в питательной среде происходит отток его из старейших органов и тканей в молодые растущие органы, где он подвергается повторному использованию (реутилизации).
| Культуры | Продукция | Содержание элементов питания | ||
|---|---|---|---|---|
| N | P2O5 | K2O | ||
| Пшеница озимая | Зерно | 2.8 | 0.85 | 0.5 |
| Пшеница озимая | Солома | 0.45 | 0.2 | 0.9 |
| Пшеница яровая | Зерно | 3.4 | 0.85 | 0.6 |
| Пшеница яровая | Солома | 0.67 | 0.2 | 0.75 |
| Ячмень | Зерно | 2.1 | 0.85 | 0.55 |
| Ячмень | Солома | 0.5 | 0.2 | 1 |
| Кукуруза | Зерно | 1.91 | 0.57 | 0.37 |
| Кукуруза | Солома | 0.75 | 0.3 | 1.64 |
| Горох | Зерно | 4.5 | 1 | 1.25 |
| Горох | Солома | 1.4 | 0.35 | 0.5 |
| Лен | Семена | 4 | 1.35 | 1 |
| Лен | Солома | 0.62 | 0.42 | 0.97 |
| Подсолнечник | Семена | 2.61 | 1.39 | 0.96 |
| Подсолнечник | Целое растение | 1.56 | 0.76 | 5.25 |
| % на сырую массу | ||||
|---|---|---|---|---|
| Картофель | Клубни | 0.32 | 0.14 | 0.6 |
| Картофель | Ботва | 0.3 | 0.1 | 0.85 |
| Сахарная свекла | Корни | 0.24 | 0.08 | 0.25 |
| Сахарная свекла | Ботва | 0.35 | 0.1 | 0.5 |
| Капуста | Кочаны | 0.33 | 0,09-0,12 | 0,27-0,44 |
| Томаты | Плоды | 0.26 | 0.07 | 0,29-0,36 |
Физиологические функции калия
Физиологические функции калия в растительном организме разнообразны. Он оказывает положительное влияние на физическое состояние коллоидов цитоплазмы, повышает их обводненность, набухаемость и вязкость, что имеет большое значение для нормального обмена веществ в клетках, а также для повышения устойчивости растений к засухе. При недостатке калия и усилении транспирации растения быстрее теряют тургор и увядают.
Калий положительно влияет на интенсивность фотосинтеза, окислительных процессов и образования органических кислот в растениях, участвует в углеводном и азотном обмене. При недостатке калия в растении тормозится синтез белка, в результате нарушается весь азотный обмен.
Недостаток калия особенно сильно проявляется при питании растений аммонийный азотом. Внесение высоких норм аммонийного азота при недостатке калия приводит к накоплению в растениях большого количества не переработанного аммиака, который оказывает вредное действие на растение. При недостатке калия задерживается превращение простых углеводов в более сложные (олиго- и полисахариды).
Калий повышает активность ферментов, участвующих в углеводном обмене, в частности сахаразы и амилазы. Этим объясняется положительное влияние калийных удобрений на накопление крахмала в клубнях картофеля, сахара в сахарной свекле и других корнеплодах. Под влиянием калия повышается морозоустойчивость растений, что связано с большим содержанием сахаров и увеличением осмотического давления в клетках.
При достаточном калийном питании повышается устойчивость растений к различным заболеваниям, например в зерновых хлебов — к мучнистой росе и ржавчине, в овощных культур, картофеля и корнеплодов — к возбудителям гнилей. Значительно улучшает лежкость плодов и овощей. Калий положительно влияет на прочность стеблей и устойчивость растений к полеганию, на выход и качество волокна льна и конопли.
Содержание калия в клетках растения значительно выше, чем других катионов. Внутриклеточная концентрация калия в растениях в 100-1000 раз превышает его концентрацию в почвенном растворе.
Критический период в потреблении калия растениями приходится на первые 15 дней после всходов. Период максимального потребления совпадает с периодом интенсивного прироста биомассы. Поступление калия заканчивается у льна в фазу цветения, у зерновых и зернобобовых — до цветения, молочной спелости. У других культур период поступления калия в растения более растянут, и проходит в течение всего вегетационного периода (картофель, сахарная свекла, капуста).
Относительное содержание элементов минерального питания в основной и побочной продукции различных сельскохозяйственных культур определяется, прежде всего, их видовыми особенностями, но зависит также от сорта и условий выращивания. Содержание азота и фосфора значительно выше в хозяйственно ценной части урожая — зерне, корне и клубнеплодов, чем в соломе и ботве. Калия же больше содержится в соломе и ботве, чем в товарной части урожая.
Калиелюбивые культуры (сахарная и кормовая свекла, картофель, капуста и кукуруза) потребляют этот элемент гораздо больше, чем зерновые и зернобобовые культуры, лен и травы. Также много калия потребляет подсолнечник.
Общий вынос калия с урожаем сельскохозяйственных культур сильно различается. Это обусловлено особенностями химического состава растений, колебаниями в уровне формирования урожая и изменением его структуры.
Недостаток калия вызывает множество нарушений обмена веществ у растений. В результате производительность растения падает, качество продукции снижается, растения начинают чаще поражаться различными болезнями.
Внешние признаки калийного голодания проявляются в побурении краев листовых пластинок — `краевом запале`. Края и кончики листьев приобретают `обожженный` вид на пластинках появляются мелкие ржавые пятна. При недостатке калия клетки растут неравномерно, что вызывает гофрированность, куполообразное закручивание листьев. В картофеле на листьях появляется также характерный бронзовый налет. Особенно часто недостаток калия проявляется при выращивании более требовательных к этому элементу картофеля, корнеплодов, капусты, силосных культур и многолетних трав. Зерновые злаки менее чувствительны к недостатку калия. Но и они при остром дефиците калия плохо кустятся, междоузлия стеблей укорачиваются, а листья, особенно нижние, вянут даже при достаточном количестве влаги в почве.
Чрезмерное калийное питание растений также негативно сказывается на их росте и развитии. Проявляется оно в возникновении между жилками листьев бледных музыкальных пятен, которые со временем буреют, а затем листья опадают. Поэтому оптимально разработанный план калийного питания растений в значительной степени будет влиять на производительность и качество урожая.
Калий в почве
Содержание калия (K2О) в различных почвах колеблется от 0.5 до 3% и зависит от механического состава. Больше калия содержится в глинистой фракции почвы. Поэтому тяжелые глинистые и суглинистые почвы богаче калием (2-3%), чем песчаные и супесчаные (1.5-2%). Очень бедные калием торфянистые почвы (0.03-0.05%). В большинстве суглинистых почв калия содержится 2-2.5%, то есть значительно больше, чем азота и фосфора. Общий (валовый) калий содержится:
- в составе первичных и вторичных минералов (не менее 91%),
- в обменно-поглощенном (0,5-2%) и необменно-поглощенном (до 9%) состояниях,
- в виде солей почвенного раствора (0.05-0.2%),
- в составе пожнивно-корневых остатков, микроорганизмов (до 0.05%).
По степени подвижности и доступности для растений содержащиеся в почве соединения калия можно разделить на следующие основные формы.
Необменно-поглощенный (фиксированный) калий
Bходит в состав крепких алюмосиликатных минералов, главным образом полевого шпата (ортоклаз и др.) И слюды (мусковит, биотит и др.). Калий полевого шпата малодоступен для растений. Однако под воздействием воды и растворенной в ней углекислоты, изменений температуры среды и деятельности почвенных микроорганизмов происходит постепенное разложение этих минералов с образованием растворимых солей калия. Калий мусковита и биотита доступный растениям.
Обменно-поглощенный калий
Поглощенный почвенными коллоидами, составляет 0.8-1.5% общего содержания калия в почве. Ему принадлежит основная роль в питании растений. Хорошая доступность обменного калия для растений обусловлена его способностью при обмене с другими катионами легко переходить в раствор, из которого он усваивается растениями. При усвоении растениями калия из раствора новые порции его переходят из поглощенного состояния в почвенный раствор. По мере использования обменного калия этот процесс все больше замедляется, а остаточный калий все прочнее удерживается в поглощенном состоянии. Содержание обменного калия может служить показателем степени обеспеченности почвы усваиваемым калием. Обычные и мощные черноземы и сероземы богаче обменным калием, чем дерново-подзолистые почвы, особенно песчаные и супесчаные.
Водорастворимый калий
Представлен различными солями, растворимыми в почвенной влаге (нитраты, фосфаты, сульфаты, хлориды, карбонаты), которые непосредственно усваиваются растениями. Содержание его в почве обычно незначительное (около 1/10 от обменного), так как калий из раствора немедленно переходит в поглощенное состояние и потребляется растениями. В некоторых почвах водорастворимый калий (а также калий, внесенный в почву как удобрение) может поглощаться в необменной форме, в результате снижается доступность его для растений. Необменная фиксация калия, как и иона аммония, наиболее сильно выражена на черноземах и серозёмах, особенно при их попеременном увлажнении и высушивании.
Между различными формами калия в почвах существует динамическое равновесие. Количество водорастворимых форм калия может пополняться за счет обменно-поглощенных, уменьшение которых через некоторое время может возмещаться за счет фиксированной формы. Следует иметь в виду, что при внесении водорастворимых калийных удобрений их трансформация может протекать в обратном направлении. Часть калия теряется из корневого слоя за счет инфильтрации (процесса пропитки и просачивания) от 2% в тяжелых и до 5% на легких почвах от внесенного количества удобрений. Также потери могут происходить от водной или ветровой эрозии.
Итак, главным условием для поддержания оптимального баланса питательных веществ в почве, в том числе и калия, является компенсация затрат за счет применения минеральных и органических удобрений.
Российский калий дошел до Индии – Бизнес – Коммерсантъ
Один из крупнейших мировых производителей калия «Уралкалий», который не торопится подписывать по низкой цене контракт с Китаем на поставку своей продукции, договорился с Индией о продажах на 2020 год по $230 за тонну. Хотя такой уровень почти на 18% ниже прошлогоднего, компания считает его более соответствующим текущей конъюнктуре.
Торговая структура «Уралкалия» «Уралкалий Трейдинг» заключила соглашение с крупнейшим индийским импортером минеральных удобрений IPL на поставку хлористого калия в Индию до конца 2020 года. Цена контракта составила $230 за тонну на условиях поставки в порт покупателя (CFR). Предыдущий контракт с IPL был подписан в октябре 2019 года по цене $280 за тонну.
В начале недели аналогичные соглашения подписали совместный трейдер североамериканских производителей хлоркалия Canpotex и белорусская «Белорусская калийная компания» (БКК, трейдер «Беларуськалия»).
Как отмечают в российском производителе, такой уровень цен «в большей степени отражает реальную рыночную ситуацию в отличие от коммерческих условий китайского контракта, которые большинством компаний в индустрии были оценены как неоптимальные».
Традиционно определяющими для калийного рынка являются контракты с Китаем — крупные игроки обычно подписывают сначала их. Однако в этом году крупнейшие игроки рынка по-разному строят свою маркетинговую политику. В начале мая БКК договорилась с Китаем о поставках калия по $220 за тонну, задав ориентир для всего рынка — это на $70 на тонну (31,8%) ниже уровня предыдущего контракта. Объем поставок не раскрывается.
По мнению БКК, такая стоимость нового китайского контракта «создает прочный фундамент для стабилизации, восстановления и последующего поступательного развития мирового калийного рынка». В то же время в «Уралкалии» считают, что согласованная Белоруссией цена не является оптимальной как для контракта, заключаемого на столь длительный период, так и для отрасли в целом. Аналогичное мнение высказывал глава канадской Nutrien Чарльз Магро, отметив, что «цена китайского контракта слишком низкая», и компании «придется решать, что делать со сроками контракта и объемами поставки». «Уралкалий» пока не заключил соглашение с Китаем.
Текущая цена на этом рынке, отмечают в «Уралкалии», не сможет обеспечить инвестиции производителям с учетом, что те несут высокие инвестиционные расходы, связанные с поддержанием существующих производственных мощностей и разработкой новых месторождений.
Как отметил гендиректор «Уралкалий Трейдинг» Александр Терлецкий, ценовые параметры достигнутого с Индией соглашения в целом соответствуют рыночной конъюнктуре. Заключение контракта в условиях экономического кризиса в равной степени отвечает интересам как потребителей, так и производителей, а также позволит максимально сохранить позиции компании на одном из ключевых рынков потребления.
Ольга Мордюшенко
как избежать дефицита жизненно важных микроэлементов — Российская газета
Наше тело нуждается практически во всех химических элементах. Но некоторые мы можем получать только с пищей. И потому неправильное питание приводит порой к серьезным заболеваниям. Ведь модные сегодня диеты, а то и просто привычка питаться на скорую руку консервированными продуктами — все это неизбежно ведет к дефициту многих жизненно важных минералов и веществ. Какие же микроэлементы особенно нужны нам, для чего и как определить их нехватку?
Вот лишь пять элементов из таблицы Менделеева. Нам их требуется совсем немного, но в обязательном порядке:
1 Железо (Fe) — входит в состав гемоглобина крови (60-75% железа в нашем организме содержится в эритроцитах — красных кровяных клетках). Гемоглобин переносит кислород, которым мы дышим, ко всем органам и тканям, поэтому жизнь без него невозможна так же, как и без кислорода. При дефиците железа в организме ухудшается клеточное дыхание, что ведет к дистрофии тканей и органов, потом развивается анемия, а в запущенных случаях — рак крови.
Человек должен получать 15-20 мг железа в день. Оно содержится в твороге и твердом сыре, бобовых, злаках, свекле, печени животных. Лучшему всасыванию железа из пищевых продуктов способствуют лимонная и аскорбиновая кислоты и фруктоза, которые содержатся во фруктах, ягодах, их соках.
Для примера: гречневая крупа, фасоль, горох, шоколад, черника содержат около 4 мг железа на 100 граммов продукта.
2 Медь(Сu) — химический элемент, продлевающий нам молодость, так как он отвечает за эластичность тканей. Преждевременная седина, морщины, обвисание кожи — верные признаки нехватки меди. При нехватке этого элемента в организме наблюдаются: задержка роста (у детей), анемия, дерматозы, депигментация волос, частичное облысение, потеря аппетита, сильное исхудание, понижение уровня гемоглобина, атрофия сердечной мышцы, варикозное расширение вен, причиной которой является все то же нарушение эластичности тканей.
Меди организму человека требуется 1-3 мг в сутки. Ею богаты печень, почки и мясо животных, морская и пресноводная рыба; морепродукты, крупы (перловая, пшеничная, гречневая, овсяная), картофель, укроп, некоторые фрукты и ягоды — черная смородина, малина, клюква, абрикосы, крыжовник, груши, клубника. Кстати, пиво способствует усвоению меди из пищи.
Для примера: 100 граммов печени трески содержит 12 мг меди, какао-порошок — 5 мг, печень говяжья — 4 мг.
3 Цинк (Zn) — элемент, особенно важный для мужчин, так как больше всего его содержится в сперме. Кроме того, он оказывает влияние на активность половых и гонадотропных гормонов гипофиза, участвует в жировом, белковом и витаминном обмене, в процессах кроветворения. Верный признак дефицита цинка — потеря обоняния и вкуса. Дети при недостатке цинка страдают гнойничковыми заболеваниями кожи и слизистых оболочек. У взрослых это проявляется в перевозбуждении нервной системы, быстром утомлении, ослаблением и выпадением волос, утолщением кожи, отеками слизистых оболочек рта и пищевода. Недостаточность цинка также приводит к бесплодию.
Цинк способствует заживлению ран, поэтому его необходимо принимать перед любой хирургической операцией, а также после нее. Кроме того, цинк помогает при лечении катаракты, замедляет разрушение сетчатки. Подобно витамину С, цинк полностью прекращает вирусную инфекцию, если захватить ее достаточно рано.
В среднем нам надо употреблять 10-20 мг цинка ежедневно. А беременным женщинам до 30 мг. Наиболее богаты этим микроэлементом отруби, проросшие зерна пшеницы, хлеб грубого помола.
Для примера: в 100 граммах дрожжей 10 мг цинка, в отварной говядине — 7 мг, в тыквенных семечках 7,4 мг, в какао-порошке — 6,3.
4 Кальций (Сa) — этот элемент не только отвечает за крепость нашего скелета, но и участвует во всех жизненных процессах организма. Нормальная свертываемость крови происходит только в присутствии солей кальция. Кальций играет важную роль в нервно-мышечной возбудимости тканей. При недостатке кальция наблюдаются: тахикардия, аритмия, боли в мышцах, беспричинные рвоты, запоры. Волосы делаются грубыми и выпадают. Ногти становятся ломкими. Кожа утолщается и грубеет. Глазной хрусталик теряет прозрачность. Любое падение может привести к серьезной травме, ведь кости делаются хрупкими.
В среднем человеку надо около 1000 мг кальция в сутки. Но эта величина разнится для людей разного возраста. Много кальция содержится в кунжуте, крапиве, твердом сыре, халве, зелени петрушки, сардинах, капусте.
Для примера: в ста граммах голландского сыра содержится около 1000 мг кальция, а в 100 граммах халвы — 824 мг.
5 Калий (K) — этот элемент отвечает за клеточные оболочки, делая их проницаемыми для прохождения солей. Поэтому он необходим для ясности ума, избавления от шлаков, лечения аллергии. Недостаток калия приводит к замедлению роста организма и нарушению половых функций, вызывает мышечные судороги, перебои в работе сердца.
Ежедневно нам надо до 2000 мг калия. Этот элемент можно пополнить, вписав в меню мясо и субпродукты, черную смородину, овсяную крупу, чернослив, арбуз, кукурузу.
Для примера: один банан среднего размера содержит 450 мг калия, чашка молока — 370 мг, один апельсин — 250 мг.
Кстати
Кроме микроэлементов нам нужны еще и ультрамикроэлементы, которые содержатся в организме человека и вовсе в очень малых количествах. К ним относятся хром, ванадий, селен, бор, никель, олово, серебро, золото и др. Вот как сказывается их недостаток:
Литий. Предполагают, что его недостаток вызывает состояние агрессии, депрессии и, как вторичное явление, пьянство.
Хром и ванадий — их дефицит приводит к заболеванию диабетом, к потере зрения.
Олово — без него человек страдает ранним облысением. А при длительной нехватке еще и развивается глухота.
Бор — его дефицит приводит к остеопорозу, так как этот элемент помогает сохранить в костях употребляемый кальций.
Селен — его нехватка бьет по сердцу. Он отвечает за нормальную работу сердечной мышцы, а также за противоопухолевую активность организма.
Кобальт — спасает от малокровия, так как без него не образуется витамин В12 (излечивающий анемию).
Калий — обзор | Темы ScienceDirect
11.1 Введение
Калий вместе с азотом и фосфором является одним из наиболее важных питательных веществ для растений и третьим элементом в классических химических удобрениях NPK (Velázquez et al., 2016). Он играет важную роль в активации нескольких метаболических процессов, таких как фотосинтез, синтез белков и ферментов, а также в устойчивости к болезням и насекомым. Кроме того, растениям необходим калий для их раннего роста, производства белков и поддержания эффективности использования воды (Теотия и др., 2016). Общее содержание калия в почве превышает 20 000 ppm и подразделяется на различные фракции: недоступный, медленно доступный и легкодоступный (Sharma et al., 2016). Достаточное поступление калия к растениям приводит к плохо развитым корням, образованию мелких семян, медленному росту, снижению урожайности, и растения становятся более восприимчивыми к болезням и вредителям (Meena et al., 2014, 2015). Хотя запасы калия в почве, как правило, велики, большая часть почвенного калия не доступна для усвоения растениями напрямую (Zörb et al., 2014). Таким образом, калийные удобрения используются для удовлетворения потребностей растений. В общей сложности 95% калийных удобрений представляют собой хлористый калий, также называемый хлоридом калия. Используются культуры, которые не переносят хлорид, сульфат калия, нитрат калия и другие соли, не содержащие хлора (Теотия и др., 2016).
Использование химических удобрений небезопасно для окружающей среды, и, кроме того, из-за несбалансированного внесения калийных удобрений дефицит калия становится основным ограничением в производстве сельскохозяйственных культур (Kour et al., 2020d; Ядав и Ядав, 2018б). Кроме того, с внедрением высокоурожайных культур и прогрессирующей интенсификацией сельского хозяйства запасы калия в почве день ото дня истощаются. Таким образом, становится очень важным найти альтернативные стратегии для поддержания уровня калия в почве и обеспечения доступности калия для растений (Supanjani et al., 2006). За последние три десятилетия понимание ризосферных, эндофитных и эпифитных микробов значительно улучшилось, и доступны различные отчеты о микробах, способствующих росту растений (Yadav et al., 2020b, 2020c, 2020d). Микробы, способствующие росту растений (PGPM), способствуют росту растений с помощью различных механизмов, таких как обеспечение доступности для растений различных питательных веществ, таких как калий, фосфор, цинк, они производят различные регуляторы роста растений, защищают растения от патогенов, помогают растения, чтобы выжить в стрессовых условиях (Kour et al., 2020b; Rana et al., 2020a; Yadav, Yadav, 2018a) (рис. 11.1).
Рис. 11.1. Схематическое изображение изоляции, характеристики, идентификации и потенциального применения стрессоустойчивых микробов.
(Адаптировано с разрешения Kour, D., Rana, KL, Yadav, AN, Yadav, N., Kumar, V., Kumar, A., et al., 2019a. Засухоустойчивые микробы, солюбилизирующие фосфор: биоразнообразие и биотехнологические применения для облегчения стресса от засухи у растений. В: Сайед, Р.З., Арора, Н.К., Редди, М.С. (ред.), Ризобактерии, способствующие росту растений для устойчивого управления стрессом: Том 1: Ризобактерии в управлении абиотическим стрессом. Спрингер, Сингапур, стр. 255 -308. DOI: 10.1007 / 978-981-13-6536-2_13).
Сообщается о широком спектре солюбилизирующих калий микробов, а именно: Acidithiobacillus , Agrobacterium , Arthrobacter , Aspergillus , Bacillus , Burkholderia , Enterobacter 900obacter Pantoea Klebsiella , Microbacterium , Myroides , Paenibacillus , Pseudomonas и Stenotrophomonas (Rana et al., 2019b; Verma et al., 2013, 2015а, 2017а). В этой главе рассматривается разнообразие солюбилизации калия из различных мест обитания, механизм солюбилизации и мобилизации калия, а также их потенциальные биотехнологические применения для устойчивого сельского хозяйства и окружающей среды.
Анализ крови на калий: причины низкого и высокого уровней + способы улучшения
Калий — это электролит, который играет ключевую роль в работе нервов и мышц, кровяном давлении и балансе жидкости. Аномальный уровень может быть признаком серьезных проблем со здоровьем.Узнайте о причинах и последствиях низкого и высокого уровня калия. и что вы можете сделать, чтобы их улучшить.
Что такое калий?
Калий в организме человека
Калий — важный минерал и положительный ион, который выполняет множество важных функций в нашем организме.
Калий [1, 2, 3]:
- контролирует кровяное давление
- помогает балансировать жидкости
- необходим для связи нервов и мышц
- помогает хранить питательные вещества, включая глюкозу, внутри клеток
Большинство Калий в организме находится внутри клеток, где он участвует в производстве белков, помогает поддерживать водный баланс, играет роль в делении клеток и ускоряет химические реакции [4, 5].
Наибольшие запасы калия в организме человека находятся в мышцах [1].
Уровень калия в крови, который в медицинской литературе называется сывороточным калием, составляет лишь около 2% от общего содержания калия в организме [1].
Калий всасывается через кишечник и выводится почками с мочой [1, 6].
Тест на калий важен для оценки здоровья почек, сердца и надпочечников.
Нормальный диапазон
Нормальный диапазон составляет около 3.5-5,3 ммоль / л. В разных лабораториях он может незначительно отличаться.
Даже незначительные отклонения от нормы связаны со значительными отрицательными эффектами [7].
На изменение уровня калия в сыворотке влияет несколько факторов. Исследование показало, что уровень калия в сыворотке был самым низким вечером (около 21:00) и самым высоким в полдень (13–15 часов) [8].
Средние уровни калия в сыворотке крови могут варьироваться для людей в разных географических регионах, вероятно, в результате различий в питании [9].Одно исследование показало, что у мужчин в Китае уровень калия в сыворотке ниже, чем в среднем в других частях мира [9].
В Соединенных Штатах концентрация калия в сыворотке крови у афроамериканцев обычно ниже, чем у людей европейского происхождения [10].
Во время беременности повышенный уровень калия является нормальным, поскольку почки выводят меньше калия; Повышение уровня гормона прогестерона может быть ответственным за адаптацию организма к этим изменениям.
Низкий уровень калия
Низкий уровень калия известен как «гипокалиемия» (гипокалиемия = низкий уровень калия, калемия = калий в крови).
Результат ниже нормы не обязательно означает, что у вас есть заболевание, требующее лечения. Ваш врач интерпретирует ваш результат с учетом вашей истории болезни, симптомов и других результатов анализов.
Причины
Указанные ниже причины обычно связаны с низким уровнем калия. Проконсультируйтесь со своим врачом или другим медицинским работником, чтобы поставить точный диагноз.
Факторы, которые могут снизить уровень калия, включают:
- Обезвоживание из-за диареи или рвоты [11, 12]
- Низкое потребление калия, особенно в сочетании с диетой с высоким содержанием натрия [11, 13]
- Алкоголизм [14]
- Дефицит магния [15, 11]
- Хронический стресс или тревога [16, 17]
- Избыток гормонов надпочечников (напр.грамм. Синдром Конна, синдром Кушинга) [18]
- Проблемы с почками [19, 11, 12]
- Солодка в избытке [20, 21]
- Отравление барием [22, 23]
- Редкие генетические нарушения [11]
Уровень калия в сыворотке крови здоровых людей существенно не зависит от содержания калия в рационе или количества калия, хранящегося в организме [3].
В результате недостаточное потребление калия часто не является истинной причиной гипокалиемии. Тем не менее, люди с очень плохим питанием и люди, участвующие в программах тяжелого снижения веса, находятся в опасности развития тяжелой недостаточности калия [24, 25, 7, 26].
Некоторые препараты могут также снижать уровень калия, в том числе:
- Водные таблетки (диуретики) [11, 27]
- Слабительные [11]
- Инсулин [11, 27, 1]
- Глюкокортикоид, часто используемый для подавления аллергических реакций. , воспалительные и аутоиммунные расстройства [28]
- Пенициллин [11, 27]
- Передозировка ацетаминофена (тайленол, панадол, парацетамол) [29, 30]
Симптомы
Симптомы низкого уровня калия включают [11, 27, 31 ]:
- Слабость
- Усталость
- Мышечные судороги
- Запор
- Нерегулярное сердцебиение
Почему низкий уровень калия в сыворотке — плохой?
Низкий уровень калия в сыворотке (гипокалиемия) может вызвать серьезные нарушения функции сердца, мозга и мышц [32].
Повышение уровня калия
Наиболее важно поработать с врачом, чтобы выяснить, что вызывает низкий уровень калия, и лечить любые основные заболевания!
Обсудите со своим врачом дополнительные стратегии, перечисленные ниже. Ни одну из этих стратегий никогда не следует применять вместо того, что рекомендует или предписывает ваш врач!
Чтобы убедиться, что у вас достаточно калия:
- Ешьте больше продуктов, богатых калием, таких как свежие фрукты и свежие овощи.В них содержится больше калия, чем в злаках и мясе [1]. Хорошие источники включают бананы, изюм, чернослив, абрикосы, финики, молочные продукты, клубнику, цитрусовые, арбуз, сою, тыкву, орехи и свеклу.
- Уменьшите потребление соли (хлорида натрия). Соление продуктов и отказ от жидкости приводит к обмену натрия на калий, что снижает содержание калия в пище [1]. Также избегайте продуктов с высоким содержанием натрия, таких как расфасованные продукты, фаст-фуд, копченые и маринованные продукты, мясные консервы, мясные обеды и чипсы.
- Обсудите с врачом использование заменителей соли, содержащих калий [13]. Их нельзя использовать, если у вас есть такое заболевание, как диабет, болезнь сердца или почек, или если они мешают принимать лекарства.
- Уменьшите потребление кофе / кофеина. Кофеин снижает уровень калия в крови [33, 34].
- Проверьте свой магний. Убедитесь, что у вас нет дефицита магния. Дефицит магния может еще больше снизить уровень калия [15, 35, 11]
Исследования показали, что тревога увеличивает уровень гормонов надпочечников, что может снизить уровень калия в крови [17].Избегайте стресса или управляйте им. Хорошие стратегии борьбы со стрессом — это полноценный отдых, физические упражнения, расслабление, медитация и т. Д.
Обсудите с врачом дополнительные калийные добавки. Эта добавка всегда должна предоставляться под наблюдением врача из-за потенциальной токсичности, особенно если у вас заболевание почек [36, 37].
Высокий уровень калия
Высокий уровень калия называется «гиперкалиемией» (гипер = высокий, калемия = калий в крови).
У большинства людей высокий уровень калия можно устранить или отложить.Но у людей с определенными расстройствами или заболеваниями выведение и накопление не работают должным образом. Также некоторые препараты могут повышать уровень калия выше нормы.
Однако результат выше нормы не обязательно означает, что у вас есть заболевание, требующее лечения. Результат может отличаться, например, в зависимости от способа взятия пробы крови. Ваш врач интерпретирует ваш результат с учетом вашей истории болезни, симптомов и других результатов анализов.
Причины
Указанные ниже причины обычно связаны с высоким уровнем калия.Проконсультируйтесь со своим врачом или другим медицинским работником, чтобы поставить точный диагноз.
Высокий уровень калия может быть вызван:
- Заболеванием почек [38, 39]
- Обезвоживанием [38]
- Повреждением и повреждением тканей [38]
- Инфекцией [38]
- Надпочечниковой недостаточностью (например, болезнью Аддисона) [ 40, 41]
- Диабет 1 типа [40]
- Чрезмерное потребление богатых калием фруктов или соков или чрезмерное употребление добавок (почти всегда без других основных проблем) [40]
- Некоторые редкие генетические нарушения [38]
Следующие препараты могут повышать уровень калия:
- Нестероидные противовоспалительные препараты (НПВП), такие как ибупрофен [42]
- Лекарства, снижающие артериальное давление, такие как бета-блокаторы, ингибиторы АПФ и блокаторы рецепторов ангиотензина. [43, 44, 45]
- Калийсберегающие диуретики [39]
- Ингибиторы протонной помпы, снижающие выработку кислоты в желудке, такие как омепразол [46]
Симптомы
Высокий уровень калия вызывает часто без симптомов.В качестве альтернативы, человек может испытать [40, 47]:
- Слабость
- Стук или учащенное сердцебиение
- Онемение конечностей
- Мышечная боль
- Затрудненное дыхание
- Тошнота
Почему высокий уровень калия в сыворотке плох?
Если оставить без внимания, умеренное или сильно повышенное содержание калия в сыворотке (гиперкалиемия) может привести к нарушению сердечного ритма , что может быть фатальным [40]. Таким образом, с чрезвычайно высоким уровнем калия в сыворотке крови следует обращаться как с неотложной ситуацией, и очень важно контролировать уровни лиц, находящихся в группе риска [40].
Пожилые пациенты, пациентов с хронической сердечной недостаточностью, почечной недостаточностью и пациенты с диабетом склонны к развитию гиперкалиемии [48].
Снижение уровня калия
Наиболее важно поработать с врачом, чтобы выяснить, что вызывает высокий уровень калия, и лечить любые основные заболевания.
Убедитесь, что вы хорошо гидратированы [38]. Обезвоживание может увеличить калий в крови.
Избегайте заменителей соли и упакованных продуктов, которые их содержат, поскольку они обычно содержат калий.Если у вас нарушена функция почек, возможно, вам придется сесть на диету с низким содержанием калия.
Специалист по высокому содержанию калия — Хамбл, Техас: Kingwood Kidney Associates: Сертифицированный совет по внутренней медицине
Что мне нужно знать о калии?
Калий — это минерал, который служит в организме электролитом. В этой роли он несет электрические заряды, которые необходимы для работы ваших нервов, мышц и сердца.
Ваше тело строго контролирует количество калия, циркулирующего в кровотоке.Если уровень калия поднимается выше или ниже этого строгого диапазона, у вас развиваются серьезные проблемы со здоровьем. Высокий уровень калия вызывает нерегулярное сердцебиение и может привести к сердечному приступу.
Что вызывает высокий уровень калия?
Основными причинами повышенного содержания калия или гиперкалиемии являются хроническое заболевание почек и почечная недостаточность. Ваши почки отвечают за выведение из организма лишнего калия. Они фильтруют его из кровотока и выводят с мочой.
Когда ваши почки слишком повреждены, чтобы выполнять свою работу, лишний калий остается в вашем кровотоке, где он постепенно накапливается до опасного уровня.
К другим причинам гиперкалиемии относятся такие состояния здоровья, как:
- Надпочечниковая недостаточность
- Застойная сердечная недостаточность
- Неконтролируемый диабет
- Сильное кровотечение
Уровень калия также может стать слишком высоким, если вы принимаете определенные лекарства или обезвоживаетесь. .
Какие симптомы развиваются, если у меня повышенный уровень калия?
В большинстве случаев у вас не будет никаких симптомов, пока уровень калия не станет опасно высоким.Гиперкалиемия обычно развивается медленно, что также затрудняет распознавание симптомов. Однако со временем вы можете заметить мышечную слабость, покалывание или онемение.
При внезапном повышении уровня калия или его очень высоком уровне вы испытываете:
- Тошнота
- Боль в мышцах
- Боль в груди
- Учащенное сердцебиение
- Одышка
- Общая слабость или утомляемость
Тяжелая гиперкалиемия — это опасное для жизни состояние, требующее немедленной неотложной помощи.
Как предотвратить повышенное содержание калия?
Вы можете предотвратить высокий уровень калия, ограничив его количество в своем рационе. В некоторых случаях вам может потребоваться лекарство, связывающее калий, чтобы помочь вашему организму вывести избыток калия, но при заболевании почек вам всегда нужно соблюдать диету с низким содержанием калия.
Kingwood Kidney Associates работает с каждым пациентом, предоставляя им информацию и поддержку, необходимые для успешного соблюдения диеты с низким содержанием калия. Например, вы узнаете о продуктах с высоким и низким содержанием калия и советах по питанию, которые помогут вам придерживаться этого плана.
Не обязательно отказываться от продуктов с высоким содержанием калия, таких как печеный картофель, бананы, молоко, чечевица и куриная грудка. Но вам нужно будет есть меньшими порциями и следить за количеством общего калия, которое вы потребляете в течение дня.
Чтобы узнать больше о диете с высоким содержанием калия и почечной диете, позвоните в Kingwood Kidney Associates или запишитесь на прием через Интернет.
Удобрение 101: Большая тройка — азот, фосфор и калий | TFI
Это возделываемые пахотные земли, которые позволяют человечеству выжить и процветать.Растения обеспечивают пищу, клетчатку, жилье и множество других преимуществ, а удобрения играют ключевую роль в этом процессе. Поскольку ожидается, что к 2050 году численность населения мира превысит 9 миллиардов человек, удобрения будут необходимы больше, чем когда-либо, для повышения урожайности сельскохозяйственных культур, чтобы люди были сыты и здоровы.
Все растущие растения нуждаются в 17 основных элементах, чтобы полностью раскрыть свой генетический потенциал. Из них 17 поглощаются растениями через почву, а остальные три поступают из воздуха и воды.
Поколения почвоведения дали знания о том, как проверять уровни питательных веществ в почве, как растения усваивают их и как лучше всего восполнить эти питательные вещества после сбора урожая.Вот тут-то и пригодятся удобрения.
Азот, фосфор и калий, или NPK, являются основными питательными веществами «большой тройки» в коммерческих удобрениях. Каждое из этих основных питательных веществ играет ключевую роль в питании растений.
Азот считается самым важным питательным веществом, и растения поглощают больше азота, чем любой другой элемент. Азот необходим для того, чтобы растения были здоровыми в процессе их развития и питательными для употребления в пищу после сбора. Это потому, что азот необходим для образования белка, а белок составляет большую часть тканей большинства живых существ.Ниже приведена фотография кукурузы с дефицитом азота.
Вторая из Большой тройки, фосфор, связана со способностью растений использовать и накапливать энергию, включая процесс фотосинтеза. Он также необходим для нормального роста и развития растений. Фосфор в коммерческих удобрениях поступает из фосфоритов. Ниже представлена фотография кукурузы с дефицитом фосфора.
Калий — третье по важности питательное вещество коммерческих удобрений.Он помогает укрепить способность растений противостоять болезням и играет важную роль в повышении урожайности и общего качества. Калий также защищает растение в холодную или сухую погоду, укрепляя его корневую систему и предотвращая увядание. Ниже представлена фотография кукурузы с дефицитом калия.
Большая тройка — азот, фосфор и калий — являются основными питательными веществами современных коммерческих удобрений. Следите за The Voice, поскольку мы продолжим углубленное изучение удобрений в предстоящие недели.
Для получения дополнительной информации о питательных веществах «большой тройки» в коммерческих удобрениях ознакомьтесь с учебными модулями 4R по азоту , фосфору и калию .
хлорид калия и хлорид натрия
Какая самая важная информация, которую я должен знать о хлориде калия и хлориде натрия?
Следуйте всем указаниям на этикетке и упаковке продукта. Расскажите каждому из своих медицинских работников обо всех своих заболеваниях, аллергиях и обо всех лекарствах, которые вы принимаете.
Что такое хлорид калия и хлорид натрия?
Калий — это минерал, который содержится во многих продуктах питания и необходим для некоторых функций вашего тела, особенно для работы вашего сердца.
Хлорид натрия — это химическое название соли. Натрий — это электролит, регулирующий количество воды в организме. Натрий также участвует в нервных импульсах и мышечных сокращениях.
Хлорид калия и хлорид натрия — это комбинированная минеральная добавка, которая может помочь уменьшить усталость, мышечные спазмы или тепловую прострацию, которая может возникнуть, когда вы потеете больше обычного.Этот продукт часто используется для активного отдыха на открытом воздухе при высокой температуре или в помещении, где высокие температуры могут вызвать перегрев.
Неизвестно, эффективны ли хлорид калия и хлорид натрия при лечении какого-либо заболевания. Лекарственное использование этого продукта не было одобрено FDA. Хлорид калия и хлорид натрия не следует использовать вместо лекарств, прописанных вам врачом.
Хлорид калия и хлорид натрия также могут использоваться для целей, не указанных в данном руководстве по продукту.
Что мне следует обсудить с врачом перед приемом хлорида калия и хлорида натрия?
Не используйте этот продукт без консультации врача, если:
- у вас заболевание почек; или
- Вы соблюдаете диету с низким содержанием соли.
Спросите врача, фармацевта или другого поставщика медицинских услуг, безопасно ли для вас использовать этот продукт, если у вас есть:
- высокое кровяное давление;
- порок сердца; или
- Язва или другая проблема в желудке или пищеводе.
Неизвестно, вредит ли хлорид калия и хлорид натрия нерожденному ребенку. Не используйте этот продукт без консультации врача, если вы беременны. Ваши потребности в дозе могут отличаться во время беременности.
Неизвестно, проникают ли хлорид калия и хлорид натрия в грудное молоко или могут нанести вред кормящемуся ребенку. Не используйте этот продукт без консультации врача, если вы кормите ребенка грудью.
Не давайте ребенку какие-либо травяные / лечебные добавки без консультации с врачом.
Как мне принимать хлорид калия и хлорид натрия?
Используйте точно так, как указано на этикетке, или в соответствии с предписаниями врача. Не используйте в больших или меньших количествах или дольше, чем рекомендуется.
Обычная доза хлорида калия и хлорида натрия составляет 1 таблетку от 5 до 10 раз в день.
Запейте этот продукт полным стаканом воды.
Возможно, вам придется скорректировать дозу в зависимости от количества тепла, которому вы подвергаетесь.
Позвоните своему врачу, если состояние, которое вы лечите с помощью хлорида калия и хлорида натрия, не улучшается или ухудшается при использовании этого продукта.
Хранить при комнатной температуре вдали от влаги и тепла.
Что произойдет, если я пропущу дозу?
Пропустите пропущенную дозу, если пришло время для следующей запланированной дозы. Не используйте дополнительно хлорид калия и хлорид натрия для восполнения пропущенной дозы.
Что произойдет, если я передозирую?
Обратитесь за неотложной медицинской помощью или позвоните в справочную службу Poison по телефону 1-800-222-1222.
Чего следует избегать при приеме хлорида калия и хлорида натрия?
Следуйте инструкциям вашего поставщика медицинских услуг относительно любых ограничений на еду, напитки или деятельность.
Каковы возможные побочные эффекты хлорида калия и хлорида натрия?
Получите неотложную медицинскую помощь при наличии признаков аллергической реакции: крапивницы; затрудненное дыхание; отек лица, губ, языка или горла.
Немедленно позвоните своему врачу, если у вас есть:
- симптомы обезвоживания — горячая и сухая кожа, чувство сильной жажды или жара, спутанность сознания, невозможность помочиться; или
- высокий уровень калия в крови (гиперкалиемия) — тошнота, медленное или необычное сердцебиение, мышечная слабость, потеря подвижности.
Хотя не все побочные эффекты известны, хлорид калия и хлорид натрия, вероятно, безопасны для большинства людей при использовании по назначению.
Это не полный список побочных эффектов, которые могут возникнуть. Спросите у своего доктора о побочных эффектах. Вы можете сообщить о побочных эффектах в FDA по телефону 1-800-FDA-1088.
Какие другие препараты будут влиять на хлорид калия и хлорид натрия?
Не принимайте хлорид калия и хлорид натрия без консультации с врачом, если вы принимаете какие-либо из следующих лекарств:
- лекарства для сердца или кровяного давления; или
- Лекарство от простуды или аллергии, содержащее антигистаминные препараты (Бенадрил и другие).
Этот список не полный. Другие препараты могут взаимодействовать с хлоридом калия и хлоридом натрия, включая лекарства, отпускаемые по рецепту и без рецепта, витамины и растительные продукты. В этом руководстве по лекарствам перечислены не все возможные взаимодействия.
Где я могу получить дополнительную информацию?
Ваш фармацевт может предоставить дополнительную информацию о хлориде калия и хлориде натрия.
Помните, храните это и все другие лекарства в недоступном для детей месте, никогда не делитесь своими лекарствами с другими и используйте это лекарство только по назначению.
Были предприняты все усилия для обеспечения точности, актуальности и полноты информации, предоставленной Cerner Multum, Inc. («Multum»), но никаких гарантий на этот счет не дается. Содержащаяся здесь информация о препарате может меняться с течением времени. Информация Multum была собрана для использования практикующими врачами и потребителями в Соединенных Штатах, и поэтому Multum не гарантирует, что использование за пределами США является целесообразным, если специально не указано иное.Информация о лекарственных препаратах Multum не содержит рекомендаций по лекарствам, диагностике пациентов и лечению. Информация о лекарственных препаратах Multum — это информационный ресурс, предназначенный для оказания помощи лицензированным практикующим врачам в уходе за своими пациентами и / или обслуживании потребителей, рассматривающих эту услугу как дополнение к опыту, навыкам, знаниям и суждениям практикующих врачей, а не их замену. Отсутствие предупреждения для данного лекарственного средства или комбинации лекарств никоим образом не должно толковаться как указание на то, что лекарство или комбинация лекарств безопасны, эффективны или подходят для любого данного пациента.Multum не несет никакой ответственности за какие-либо аспекты здравоохранения, управляемые с помощью информации, предоставляемой Multum. Информация, содержащаяся в данном документе, не предназначена для охвата всех возможных способов использования, указаний, мер предосторожности, предупреждений, лекарственных взаимодействий, аллергических реакций или побочных эффектов. Если у вас есть вопросы о лекарствах, которые вы принимаете, проконсультируйтесь с врачом, медсестрой или фармацевтом.
Copyright 1996-2021 Cerner Multum, Inc. Версия: 1.01. Дата редакции: 07.04.2016.
Калийное питание растений и его взаимодействие с другими питательными веществами в гидропонной культуре
\ n
2. Калиевое питание растений и его взаимодействие с другими питательными веществами в гидропонной культуре
\ n \ n
2.1. Питательный раствор для гидропоники обычно используется для растений
\ n
В гидропонной культуре питательные растворы являются единственным источником питания растений. Раствор, содержащий все питательные вещества для растений, должен применяться в правильном соотношении.При выборе удобрений и приготовлении гидропонных питательных растворов следует учитывать следующие факторы:
-
Следует учитывать концентрацию вредных элементов, таких как натрий, хлорид и бор, соленость и качество воды.
-
Значения концентрации необходимых питательных веществ в гидропонном питательном растворе должны быть хорошо отрегулированы.
-
Баланс питательных веществ должен обеспечиваться питательными веществами, которые получают растения.
-
Следует учитывать значение pH гидропонного питательного раствора и исследовать влияние значения pH гидропонного питательного раствора на поглощение питательных веществ растениями.
\ n
В таблице 1 показаны общие диапазоны питательных веществ в гидропонных питательных растворах. В таблице 2 приведены рекомендуемые питательные растворы для различных растений [6].
\ n
| Элемент | Ионная форма, поглощаемая растениями | Общий диапазон (ppm = мг / л) |
|---|---|---|
| Азот | Нитрат (NO 3 — ), аммоний (NH 4 + ) | 100–250 частей на миллион элементарного N |
| Фосфор | Дигидрофосфат (H 2 PO 4 — ), фосфат (PO 4 3-), моногидрофосфат (HPO 4 ) | 30–50 частей на миллион элементарного P |
| Калий | Калий (K + ) | 100–300 частей на миллион |
| Кальций | Кальций (Ca 2+ ) | 80–140 частей на миллион |
| Магний | Магний (Mg 2+ ) | 30–70 страниц в минуту | Сера | Сульфат (SO 4 2-) | 50–120 частей на миллион элементарного S |
| Железо | Ион железа (Fe 2+ ), ион трехвалентного железа (Fe 3+ ) | 1–5 частей на миллион |
| Медь | Медь (Cu 2+ ) | 0 .04–0,2 частей на миллион |
| Марганец | Марганец (Mn 2+ ) | 0,5–1,0 частей на миллион |
| Цинк | Цинк (Zn 2+ ) | 0,3–0,6 частей на миллион |
| Молибден | Молибдат (MoO 4 2-) | 0,04–0,08 частей на миллион |
| Бор | Борная кислота (H 3 BO 3 ), борат (H 2 BO 3 — ) | 0.2–0,5 частей на миллион элементарного B |
| Хлорид | Хлорид (Cl — ) | <75 частей на миллион |
| Натрий | Натрий (Na + ) | <50 ppm ТОКСИЧНО для растений |
Таблица 1.
Общие значения диапазона питательных веществ ионной формы элементов, поглощаемых растениями.
\ n
| Урожай | N | P | K | Ca | Mg |
|---|---|---|---|---|---|
| Концентрация в мг / л (ppm) | |||||
| Помидор | 190 | 40 | 310 | 150 | 45 |
| Огурец | 200 | 40 | 280 | 140 | 40 |
| Перец | 190 | 45 | 285 | 130 | 40 |
| Клубника | 50 | 25 | 150 | 65 | 20 |
| Дыня | 200 | 45 | 285 | 115 | 30 |
| роз | 170 | 45 | 285 | 120 | 40 |
Таблица 2.
Необходимое количество элементов, содержащихся в питательном растворе для различных растений.
\ n
Общее количество солей, растворенных в гидропонном питательном растворе, считается мерой электропроводности (EC) . EC — это параметр, используемый для отслеживания процесса оплодотворения. Данные, относящиеся к ЕС, не отражают минеральное содержание питательного раствора.
\ n
Питательный раствор для гидропоники рециркулирует в закрытых гидропонных системах. Таким образом, элементы (натрий, хлорид, фторид и др.), которые не всасываются в больших количествах растениями, или ионы, выделяемые растением, откладываются в гидропонном питательном растворе. В этом случае электропроводность (ЕС) не может предоставить информацию о содержании питательного раствора.
\ n
Катионообменная емкость (CEC) является краеугольным камнем гидропонного питания. Влияние катиона калия на катионообменную емкость бесспорно. Катионный обмен обеспечивает следующие условия:
-
Катионный обмен является основным резервуаром питательных веществ (макроэлементов) K + (монофосфат калия / сульфат калия, нитрат калия, фосфат калия и фосфат аммония), Ca 2+ и Mg 2+ .
-
Азот (N) необходимо хранить в форме аммония (NH 4 + ).
-
Катионный обмен помогает обеспечить наличие микроэлементов в следовых количествах металлов, таких как Zn 2+ и Mn 2+ в определенном количестве.
-
Катионный обмен обеспечивает устойчивость к изменениям pH , а также поддерживает питательные вещества для растений.
\ n
На рисунке 1 сравниваются катионообменные емкости на поверхности частиц глины и органических материалов с отрицательно заряженными участками, содержащими положительно заряженные ионы.
\ n
Рис. 1.
Внешний вид поверхности частиц глины и органического вещества с отрицательно заряженными участками, на которых находятся положительно заряженные ионы.
\ n
Mo и Mg присутствуют при более высоком pH, чем большинство питательных веществ. С другой стороны, следовые количества металлов, таких как Fe, Zn и Mn, имеют более низкий pH, чем большинство питательных веществ. Идеальное значение pH для многих растений составляет от 5,8 до 7,0. Значения в этом диапазоне являются сбалансированным источником всех питательных веществ [6].
\ n
Гидропонный питательный раствор следует часто проверять.Этот процесс предоставляет информацию о времени замены питательного раствора или времени разбавления пресной водой. Идеальный диапазон pH для гидропонного питательного раствора составляет 5,8–6,3. Для многих растений оптимальный диапазон pH показан на рисунке 2. Значение pH питательных микроэлементов обычно ниже предельного значения. Если уровень pH падает ниже 5,5, увеличивается риск токсичности питательных микроэлементов, а также ухудшение содержания кальция и магния. В закрытой системе гидропоники влияние корней на значение pH гидропонного раствора велико.Это вызывает колебания pH. Серная кислота, фосфорная кислота и азотная кислота используются для увеличения кислотного числа гидропонного питательного раствора. Одним из наиболее важных факторов, влияющих на значение pH питательного раствора , является добавление аммония / нитрата.
\ n
Рис. 2.
Идеальный диапазон pH элементов в гидропонном питательном растворе, используемом для большинства сельскохозяйственных культур.
\ n
Минералы, содержащиеся в сырой воде, и питательные вещества, поступающие в результате удобрения, являются двумя основными факторами, которые обеспечивают гидропонный питательный раствор.Качество сырой воды сильно влияет на выбор удобрений и их концентрацию в гидропонном питательном растворе. По этой причине, прежде чем переходить к составу удобрения для внесения удобрений, следует проверить качество и состав сырой воды. В исходной воде могут присутствовать микроэлементы, такие как бор, марганец, железо и цинк, а также минералы, такие как кальций, магний и сера. Поэтому при приготовлении питательного раствора для гидропоники необходимо также учитывать влияние этих элементов.Кроме того, в сырой воде могут присутствовать нежелательные минералы, такие как натрий, хлорид или фторид. Для гидропонного питательного раствора присутствие этих минералов нежелательно. Чтобы избавиться от такой ситуации, можно предпринять следующие действия:
-
Сырую воду можно разбавить добавлением чистой воды.
-
Сырая вода может быть обессолена.
-
Возможен ионный обмен [6].
\ n
Переворачивание и перекачивание питательного раствора можно выразить как операции по замене резервуара.Эти операции выполняются в определенное время / с определенной частотой. Замена резервуара — один из факторов, которым можно управлять в гидропонных системах. Есть много разных способов сэкономить время при замене резервуара. Одним из них является добавление небольшого количества концентрата питательных веществ к наиболее потребляемым ионам питательных веществ. Как правило, N, K и P составляют содержание добавленной концентрации питательного вещества. В предыдущем исследовании содержание ежедневно добавляемого концентрата питательных веществ составляло 10 частей на миллион для азота и фосфора и 15 частей на миллион для калия [7].
\ n \ n \ n
2.2. Взаимодействие калия с другими питательными веществами в гидропонной культуре
\ n
Добавление исходной воды и нитрата кальция приводит к увеличению содержания ионов Са в некоторых растворах. Чтобы избежать добавления Са, в удобрения добавляют нитрат калия и монокалиевый фосфат. Кроме того, если добавление нитрата калия и монофосфата калия может ограничить значение Са в растворе, составы удобрений регулируются более низким начальным значением Са. Тест на смену резервуара для питательных веществ дает информацию о том, как добавление питательных веществ сравнивается с заменой в резервуаре для нормальных и низкокальциевых составов питательных веществ.Исследователи, проводящие такой тест, смогли сформировать четыре разных питательных раствора. Они подробно описаны в таблицах 3 и 4 [7].
\ n
| Питательные вещества / ион | Исходный нормальный (1) | Исходный низкий уровень кальция (2) | При замене резервуара нормальный (3) | При замене резервуара низкий уровень кальция (4) |
|---|---|---|---|---|
| Азот | 119 | 127 | 129 | 129 |
| Фосфор | 28 | 31 | 26 | 31 |
| Калий | 200 | 233 | 188 | 231 |
| Кальций | 110 | 78 | 116 | 86 |
| Магний | 30 | 33 | 32 | 36 |
| Сера | 97 | 93 | 111 | 111 |
| Натрий | 72 | 72 | 86 | 86 |
| Хлорид | 24 | 24 | 28 | 27 |
| Бор | 0.1 | 0,11 | 0,12 | 0,12 |
| Марганец | 0,04 | 0,07 | 0,04 | 0,05 |
| Медь | 0,08 | 0,08 | 0,07 | 0,08 |
| Цинк | 0,06 | 0,07 901 \ n | 0,06 | 0,08 |
Таблица 3.
Изучение ситуации с заменой резервуара в составе питательных веществ с нормальным и низким содержанием кальция.
\ n
| Питательные вещества / ион | Нормальное изменение (1) | Низкое изменение кальция (2) | Нормально без изменений (3) | Низкое содержание кальция без изменений (4) |
|---|---|---|---|---|
| Азот | 61 | 38 | 41 | 48 |
| Фосфор | 26 | 33 | 75 | 81 |
| Калий | 68 | 116 | 244 | 285 |
| Кальций | 172 | 118 | 95 | 70 |
| Магний | 44 | 52 | 25 | 29 |
| Сера | 197 | 206 | 168 | 167 |
| Натрий | 112 | 115 | 114 | 111 |
| Хлорид | 25 | 24 | 28 | 26 |
| Бор | 0.11 | 0,13 | 0,09 | 0,10 |
| Марганец | <0,01 | <0,01 | <0,01 | < 0,01 |
| Медь | 0,09 | 0,11 | 0,06 | 0,07 |
| Цинк | 0,05 | 0,06 | 0,02 | 0,03 |
Таблица 4.
Питательные растворы, для которых нет эффекта замены резервуара для нормального и низкого содержания кальция в составе питательных веществ.
\ n
Исследователи отметили следующие перспективы в своей работе:
-
В процессе производства была произведена замена резервуара в штатном решении.
-
В обычном растворе замена резервуара не производится. При дальнейшей обработке ежедневно производились добавки KNO 3 и KH 2 PO 4 после замены резервуара.
-
Во время производства была произведена замена резервуара для раствора с низким содержанием кальция.
-
В растворе с низким содержанием кальция замена резервуара не производится.При дальнейшей обработке ежедневно производились добавки KNO 3 и KH 2 PO 4 после замены резервуара [7].
\ n
Чтобы оценить данные, полученные как для растения, так и для раствора, были отмечены следующие результаты:
-
Никаких существенных различий между конечными выходами свежей массы при четырех обработках не наблюдалось.
-
Не было значительных различий в рейтингах ожогов.
-
Наблюдались некоторые различия в оценках ожогового типа при лечении.Предполагается, что это можно сделать менее частой заменой резервуара.
-
Начальная точка условий раствора, подготовленных производителями, должна тщательно контролироваться.
-
Следует изучить взаимосвязь между отдельными условиями в исходной воде и другими параметрами.
-
Наиболее вредными свойствами раствора являются увеличение содержания S и Na к концу процесса. В этом случае необходимо понизить pH.Для этого следует использовать раствор азотной кислоты вместо раствора серной кислоты. Вместо увеличения уровня K за счет добавления KNO 3 , уровень N увеличивается за счет добавления раствора азотной кислоты. Таким образом, накопление S в растворе также снижается.
-
Воздействие исходной воды увеличивает уровень Na в растворе. Уровень Na изменить сложнее. Важное значение имеет своевременное определение повышенного уровня Na.
-
В этом исследовании было замечено, что некоторые элементы, содержащие питательные микроэлементы, такие как Mn, были на более низком уровне.Если интервалы смены резервуаров должны быть увеличены, ежедневные добавки следует выбирать из наиболее часто используемых микроэлементов. При регулярных интервалах замены баков следует применять более высокие начальные значения, чтобы избежать дефектов [7].
\ n
Калий, который присутствует в виде свободных ионов почти во всех питательных растворах, имеет значение pH 2–9 [8]. Как и калий, кальций и магний также имеют широкий диапазон pH. Однако присутствие кальция и магния ограничено из-за присутствия других ионов.Следовательно, если питательный раствор содержит вещества с pH выше 7; Fe 2+ , Mn 2+ , PO 3 −4 , Ca 2+ и Mg 2+ осаждаются в соли. Это означает, что количество питательных веществ, поступающих в растения, ограничено [9].
\ n
Рост, развитие и производство растений основаны на общей концентрации ионов в питательном растворе [10]. Ионы растворенных солей в питательном растворе обладают способностью связываться с питательными растворами.Это свойство вызвано силой, называемой осмотическим давлением (ОП) . Осмотическое давление зависит от количества растворенных веществ [11]. Кроме того, потенциал растворения или осмотический потенциал термины обычно используются в питательных растворах. В питательном растворе растворенные вещества оказывают значительное влияние на водный потенциал. Растворители уменьшают свободную энергию воды за счет разбавления воды [12]. Концентрация соли определяет общее количество солей в растворе. Электропроводность (EC) — это показатель концентрации соли. Таким образом, осмотическое давление питательного раствора косвенно определяется параметром электропроводности (ЕС) . Таким образом, ЭК питательного раствора является хорошим индикатором количества ионов в корневой зоне растений [13].
\ n
Электропроводность или Осмотическое давление — первый исследуемый параметр для концентрации питательного раствора. Регулирование pH и температуры корня также являются другими важными факторами, исследуемыми с точки зрения урожайности и качества [14].Питательные вещества и водопоглотители из питательного раствора постоянно усиливают свою электрическую проводимость (ЕС). Таким образом, в то время как концентрации некоторых ионов уменьшаются, концентрации некоторых ионов также увеличиваются. Такая ситуация возникает как в закрытых, так и в открытых гидропонных системах одновременно. Например, в закрытой гидропонной системе с выращиванием роз питательный раствор в резервуаре контролировался, и результаты показали, что концентрация Fe падала очень быстро, в то время как Ca 2+ , Mg 2+ и Cl — увеличился.Кроме того, нет критических условий в уровнях концентрации K + , Ca 2+ и SO 4 2- [15]. Повторное использование питательных растворов требует регулирования ЕС. Повторное использование питательных растворов было показано в различных исследованиях, которые показали положительные результаты для систем устойчивого сельскохозяйственного производства [16]. В одном из этих исследований Brun et al. [17] снизили ЕС, добавив водный комплекс в дренаж; достигла желаемой EC с использованием систем рециркуляции, содержащих дополнительный питательный раствор.
\ n
Ионы, которые активны на ЭК, — это Ca 2+ , Mg 2+ , K + , Na + , H + , NO 3- , SO 4 Ионы 2-, Cl —, HCO , 3- и OH — [18]. Микроэлементы, такие как Fe, Cu, Zn, Mn, B, Mo и Ni, не оказывают значительного влияния на ЭК, поскольку они с меньшей вероятностью усваиваются растениями, чем макроэлементы [19].
\ n
Питательные растворы содержат по существу шесть питательных веществ вместе с Ca, Mg и S, с предпочтением K, N и P. Ионное соотношение было разработано Штейнером (1961). Эта концепция основана на взаимном соотношении анионов , таких как NO 3 — , H 2 PO 4 — и SO 4 2- и взаимном соотношении катионов , например K + , Ca 2+ и Mg 2+ . Такое соотношение влияет не только на общее количество каждого иона в растворе, но и на количественное соотношение, которое удерживает ионы вместе [10].
\ n
Беспочвенное возделывание дает различные жизнеспособные и контролируемые возможности для повышения качества сельскохозяйственных культур и продукции. Отслеживаются такие параметры, как температура , pH , электропроводность и содержание кислорода в питательном растворе. Важно, чтобы эти параметры проверялись своевременно и точно, чтобы преимущество не превратилось в недостаток [15].
\ n
При повышении температуры питательного раствора увеличивается расход O 2 .Если вентиляции в корне недостаточно, концентрация CO 2 в корне увеличивается [20]. В некоторых овощах были проведены различные исследования по снижению концентрации CO 2 с помощью пероксида калия, который действует как источник кислорода [21].
\ n
Калий является наиболее желательным катионным минералом для растений и составляет 10% от сухого вещества растений. Из-за уменьшения количества удобрений KNO 3 в питательном растворе значительно снизилось содержание сухого вещества в листьях, кронах и корнях.Это замедлило рост и уменьшило количество листьев [22]. Существует ряд исследований, сообщающих, что устьичная проводимость снижается из-за недостатка К. Соответственно, сообщалось, что фиксация CO 2 и экспорт флоэмы также снижаются [23]. Кроме того, при производстве кукурузы и пшеницы наблюдались недостаточные уровни калия для повышения урожайности этих продуктов [24].
\ n
Следует учитывать различные эффекты калиевого питания, принимая во внимание общую концентрацию ионов (ЕС).В компании K Nutrition необходимо исследовать взаимосвязь калия с другими катионами. Среди этих катионов в основном присутствуют Ca, Mg и Na в солевой оросительной воде. В результате увеличения отношения K / Ca улучшается качество хранения. Кроме того, были увеличены вкусовые факторы, такие как содержание сахара и кислоты [25].
\ n
Плохое качество воды может привести к чрезмерной концентрации NaCl в питательном растворе. Следовательно, активность ионов питательных веществ может снизиться, и соотношение Na + : Ca 2+ , Na + : K + , Ca 2+ : Mg 2+ и Cl — : NO3 — может увеличиваться [26].Это может привести к осмотическому и ионному повреждению, дефициту питательных веществ и снижению урожайности и качества растения. Граттан и Грив исследовали, что соленость NaCl на ткани может иметь репрессивный эффект на концентрацию микронутритов и макроэлементов (N, P, K, Ca, Mg и S) [26].
\ n \ n \ n
3. Заключение
\ n
Калий имеет жизненно важное значение для питания растений. В гидропонных системах присутствие калия в питательных растворах положительно влияет на такие процессы, как рост, развитие и сохранение растений.Катион калия выполняет множество задач во многих процессах по сравнению с другими питательными веществами. Эти процессы, влияющие на развитие растений, могут быть перечислены как , катионообменная емкость (CEC) , значение pH , электрическая проводимость (EC) , температура корня , общая ионная концентрация , растворение потенциал (осмотический потенциал) , взаимное соотношение ионов , взаимное соотношение анионов , взаимное соотношение катионов , содержание кислорода и CO 2 концентрация.
\ n
1. Введение
Травы и специи — это семена, фрукты, корни, кора, ягоды, бутоны или растительные вещества, которые десятилетиями использовались в основном для ароматизации, окраски или консервирования пищевых продуктов и приготовления лекарственных средств. . Во всем мире Индия и Пакистан известны своим огромным производством специй и ароматизаторов. Во всем мире признано, что пряности могут иметь широкий спектр физиологических и фармацевтических преимуществ. Многочисленные проблемы со здоровьем, такие как нейродегенеративные заболевания и окислительный стресс, контролируются и восстанавливаются с помощью растительных режимов питания, поскольку они содержат различные ценные химические соединения и антиоксиданты [1].Различные метаболические заболевания и возрастные проблемы, связанные с окислительными механизмами в организме человека, также излечиваются правильным использованием трав. Из-за неопасного воздействия ароматизаторов на людей, они считаются безопасными для употребления в пищу без каких-либо побочных эффектов. На субконтиненте собирают и используют основные специи и травы, а именно. черный перец ( Piper nigrum ), гвоздика ( Syzygium aromaticum) , корица ( Cinnamomum verum ), семена льна ( Linum usitatissimum ), кардамон ( Elettaria cardamomum ), мак ( Papaver somniferum ) , пажитник ( Trigonella foenum-graecum ), тмин ( Cuminum cyminum ), кунжут, фенхель, карамболь, аджвайн, кориандр, куркума, тамаринд, имбирь, лук, чеснок и красный перец чили.Эти специи являются секретом богатства различных лечебных компонентов, которые прямо или косвенно помогают при различных нарушениях здоровья. В этой главе мы сосредоточимся на гвоздике ( Syzygium aromaticum ), которая веками использовалась в качестве пищевого консерванта и в различных лечебных целях, включая ее антимикробные и антиоксидантные свойства. Польза для здоровья, связанная с потреблением Syzygium aromaticum , кратко обсуждается ниже и показана на рисунке 1.
Рисунок 1.
Использование S. aromaticum.
Syzygium aromaticum — это дерево, которое принадлежит к семейству миртовых, происходит из Индонезии и является одной из самых ценных и второй по значимости пряностью в мировой торговле. Гвоздика используется в различных синонимах, а именно. Caryophyllus aromaticus, Caryophyllus silvestris, Eugenia caryophyllus, Jambosa caryophyllus и Myrtus caryophyllus [2]. Дерево Syzygium aromaticum достигает средних размеров и достигает 20 м в высоту.В зависимости от разновидности этого дерева форма его кроны варьируется от цилиндрической до пирамидальной [3]. Продолжительность жизни этого дерева до 100 лет и выше. Дерево предпочитает расти в сильно истощенной почве с достаточной влажностью почвы. Дереву требуется высокая температура воздуха в диапазоне от 25 до 35 ° C с сильным солнечным светом, помимо этого хорошо распределенные осадки и высокая влажность, т.е.> 70%, также являются необходимым условием для их роста [4]. В Индии гвоздичное дерево собирают в глубоких черносуглинистых почвах влажных тропиков и успешно растут на красных почвах средних земель Кералы и на холмистой территории Западных Гатов в Карнатаке и Тамил Наду [5].
Ароматические цветочные бутоны, которые имеют характерный запах этого растения, известны как гвоздика и обычно используются в качестве специи и в нескольких терапевтических целях для лечения различных заболеваний и инфекций, таких как тошнота, рвота, кашель, диарея, диспепсия, метеоризм, вздутие живота. и желудочно-кишечный спазм; ослабить боль; вызывают сокращения матки; и стимулируют нервы [6]. Из-за этого он привлекает к коммерческому выращиванию гвоздики в различных частях мира, таких как Индия, Пакистан, Шри-Ланка, а также в африканских странах.Индия становится вторым по величине потребителем гвоздики после Индонезии [7]. Растение также используется в народной медицине как мочегонное, одонтологическое, желудочное, тонизирующее и приправляющее с ветрогонным и стимулирующим действием [8]. Эфирное масло, полученное из этого ароматического растения, не только служит ароматизатором и ароматизатором, но также используется в качестве диетического антиоксиданта для предотвращения ряда заболеваний, вызываемых свободными радикалами [9, 10]. Гвоздика богата флавоноидами, органическими кислотами, летучими кислотами, фенольными компонентами, такими как эвгенол, терпеноиды, дубильные вещества и галловая кислота, которые имеют большой потенциал для применения в фармацевтике, пище и сельском хозяйстве [11].В Бразилии и других тропических странах он сталкивается с серьезными проблемами со здоровьем, вызванными лихорадкой денге, гвоздику противодействует гвоздика из-за ее ларвицидной активности [12].
1.1 Морфология
Syzygium aromaticum
Гвоздика — ароматное пряное дерево, в основном содержащее листья и почки, которые являются обычной коммерческой частью дерева. Термин «гвоздика» происходит от французских слов «Clove» и «Clou», что означает «гвоздь». Гвоздика — мирт конической формы, среднего размера дерево с прямым стволом, вырастающее до 18-20 м в высоту.Ветви гвоздичного дерева сероватые, толстые, полустоячие. Листья огромные, удлиненно-эллиптические, напротив, обратнояйцевидные, голые, с множеством железок эфирного масла на нижней поверхности. Дерево начало цветение почек примерно через 7 лет и продолжает цвести очень долго или даже дольше после посадки. Цветки маленькие, малинового цвета, распускаются обоеполыми на концах маленьких веточек. На каждом цветоносе от 3 до 4 цветков, длина соцветий составляет от 4 до 5 см.Первоначально цвет бутонов бледно-желтый с глянцевым внешним видом, а при созревании они становятся зелеными или ярко-красными. Их собирают, когда они становятся темно-красными ягодами эллипсоида [12, 13].
1.2 Химические составляющие гвоздики
Фармакологически гвоздика богата фармакофорными фенольными соединениями, такими как флавоноиды (кверцетин), гидроксибензойные кислоты, гидроксифенилпропены, гидроксициновые кислоты, эвгенол и производные галловой кислоты. [11, 12, 13, 14]. Также было обнаружено, что 18–20% эфирного масла присутствует в стебле, бутонах и листе цветка гвоздики с различным химическим составом, имеющим множество применений [15, 16].Эти масла бесцветные или бледно-желтые, с особым ароматом и вкусом [15, 16].
Alma, et al. в 2007 г. [17] извлек и сообщил о наличии эфирного масла из бутонов гвоздики, которое имеет желтый цвет и имеет более высокую плотность, чем вода. Они экстрагировали и охарактеризовали компоненты как эвгенол (87%), хавибетол (19,7%), β -кариофиллен (13%), ацетат эвгенола (8,01%), трисилоксан-1,1,1,5,5,5- гекса-метил-3,3-бис — [(триметилсилил) окси] (1,7%) и др. [18, 19], Marya, et al. 2012 [20] и Kasai, et al. 2016 [21] дополнительно изучили и сообщили о присутствии эвгенола (74,32%), β -кариофиллена (15,94%) и ацетата эвгенола (5,8%) в качестве основных компонентов в бутонах гвоздики. Другой компонент, метил- n -амилкетон, который имеет характерный фруктовый и свежий запах, также был обнаружен в бутонах гвоздики (2001 г.) [22]. Xu, et al. (2016) [23] изучил химический состав эфирного масла почек гвоздики с помощью газовой хроматографии-масс-спектрофометрии (ГХ-МС) и сообщил о присутствии оксида кариофиллена, α -селинена, кадинена, 2-пинена и т. Д.с ранее заявленным маслом. Fankem и др. [24] подтвердили присутствие оксигенированных монотерпенов (89,06%), монотерпенов (0,04%), сесквитерпенов (10,6%) и линейных компонентов (0,03%) в эфирном масле почек гвоздики вместе с эвгенолом.
Эфирное масло, также получаемое из листьев гвоздики, известное как эфирное масло листьев гвоздики. Масло, которое получают из листьев, имеет приятный запах и бледно-желтый цвет. Жировец, и др. и соавторы в 2006 г. [25] сообщили о присутствии 23 других соединений наряду с α -гумуленом, эвгенолом и их ацетатными производными в качестве основных компонентов.Эти данные свидетельствуют о том, что эвгенол, хавибетал, β -кариофиллен, эвгенола ацетат являются основными компонентами гвоздичного дерева. Эти эфирные масла также содержатся в стебле гвоздики [26, 27, 28].
Бао, и др. , 2012 [29], сообщили о нескольких других фенольных компонентах из метенольного экстракта S. aromaticum , а именно. бифлорин, кемпферол, рамноцитрин, мирицетин, галловая кислота и эллаговая кислота (рис. 2). Бао, и др. , 2012 [29] также сообщили о наличии восемнадцати гидролизуемых танинов из водного ацетонового экстракта высушенных цветочных бутонов S.aromaticum , такие как ароматинин A, платикарианин A, бикорнин, сизигинин A, алунуснин A, ругозин C, теллимаграндин II, казуаритин, гетерофиллин D, ругозин D, ругозин F, эупростин A, 1,2-ди- Ogalloyl-3- -дигаллоил-4,6- O — ( S ) -гексагидроксидифеной- β -D-глюкоза, муллианин B, скваррозанин A, казуаринин, сизигинин B, 1,2,3-три- O -галлоил- β -D-глюкоза и 1,2,3,6-тетра- O -галлоил- β -D-глюкоза.
Рисунок 2.
Общие составляющие: (1) эвгенол, (2) β-кариофиллен, (3) ванилин, (4) кратеголовая кислота (5) кемпферол, (6) рамнетин, (7) евгенитин, (8) Евгенин, (9) эллаговая кислота, (10) галловая кислота, (11) бифлорин, (12) мирицетин, (13) кампестерин, (14) стигмастерол, (15) олеаноловая кислота, (16) бикорнин, (17) кверцетин ( 18) Карвакрол, (19) ацетат эвгенола, (20) α-Кариофиллен или α-гумулен, (21) Чавибетол, (22) Кадинен (23) Пинен, (24).
1.3 Фармакологическая активность гвоздичного масла
Эфирное масло или химические соединения, которые были экстрагированы из различных частей гвоздики, обладают разнообразной фармакологической активностью, и в нескольких публикациях сообщалось, что они проявляют противоопухолевое, противомикробное, противодиабетическое, противовоспалительное, антидепрессантное и противоопухолевое действие. противоязвенное, антиоксидантное, противоязвенное, противопротозойное и др.(Рисунок 3).
Рис. 3.
Биологическое использование соединений, экстрагированных из S. aromaticum.
1.4 Противомикробная активность
Масло, выделенное из гвоздики, имеет широкий спектр медицинского применения. Он в основном используется в производстве различных индийских аюрведических и китайских лекарств. Эти масла также используются в производстве антибиотиков из-за их антимикробных свойств. Многочисленные сообщения показывают, что его компонент эвгенол, ингибирующий миграцию бактерий, бактериальную адгезию, образование фимбрий Gram -отрицательных бактерий, таких как Escherichia coli , Salmonella , Pseudomonas aeruginosa и т. Д.и Gram -положительные бактерии, такие как Staphylococcus , Streptococcus , Listeria и др. [30].
Matan et al. (2012) [31] сообщил, что гвоздичное масло проявляет сильную антимикробную активность против Penicillium sp., Aspergillus flavus и Staphylococcus aureus , обнаруженных на сушеной рыбе ( Decapterus maruadsi ). Zengin и Baysal (2014) [32] обнаружили, что они проявляют активность против трех Gram -положительных бактерий, таких как Listeria innocua, Carnobacterium divergens и Staphyloccous aureus , а также подавляют потенциальную активность против четырех Gram -отрицательных бактерий. например, Salmonella typhimurium, Escherichia coli, Serratia liquefaciens и Shewanella putrefacians .Данные убедительно доказали, что эфирное масло гвоздики подавляет рост всех бактерий, будь то грамм — положительный или грамм — отрицательный, в то время как основной компонент эфирного масла оказался неактивным по отношению к Shewanella и Listeria . [33]. Абдулвахаб Каммон, и др. в 2020 году сообщил об антимикробной активности гвоздичного масла против грамм -отрицательных бактерий, выделенных от цыплят E. coli (1 птичьего патогенного E. coli (APEC) и 2 непатогенных E.coli), Salmonella enteritidis и Salmonella spp., , которые были изолированы от кур в Департаменте болезней птицы и рыб [34]. В пищевой промышленности эфирному маслу гвоздики уделяется первостепенное внимание из-за загрязнения L. monocytogenes , особенно для пищевых продуктов, консервированных при низкой температуре, поскольку эти бактерии размножаются при низкой температуре и загрязняют пищу [35]. Singh, et al. (2003) [36] сообщили, что гвоздичное масло было высокоэффективным против L.monocytogenes в пептонной воде и уменьшил популяцию бактерий. Эфирное масло гвоздики (5%) уменьшило количество клеток L. monocytogenes до неопределяемого уровня в курином фарше в течение одного дня воздействия, поэтому его можно использовать для борьбы с L. monocytogenes в курином фарше [37] .
1.5 Антиоксидантная активность
В прошлом механизмы окисления и свободнорадикальные механизмы вызывали большой интерес в живых системах [38]. Поглощение кислорода необходимо для клеточного метаболизма, который приводит к производству активных форм кислорода (АФК).Взаимодействие АФК с молекулами липидов приводит к образованию пероксильных радикалов, которые в дальнейшем взаимодействуют с нуклеиновыми кислотами и белками, что приводит к изменениям и, следовательно, функциональным модификациям [39]. АФК включают свободные радикалы, такие как супероксид-анион-радикалы O 2 — , синглетный кислород, несвободно-радикальный перекись водорода и гидроксильные радикалы, представляют собой различные формы активных форм кислорода [40, 41, 42, 43]. АФК производятся непрерывно во время нормального физиологического процесса и могут легко привести к перекисному окислению липидов, что приводит к накоплению перекисей липидов.АФК отвечает за разрушение важных биомолекул, таких как нуклеиновые кислоты, белки, липиды, а также углеводы. Они также несут ответственность за повреждение ДНК, которое инициирует мутацию [43, 44]. Они предполагают, что, если АФК не удаляются в достаточной степени, они вызывают более 100 заболеваний, таких как болезнь Альцгеймера, сахарный диабет, гипертония [45, 46], рак простаты и толстой кишки, ишемическая болезнь сердца, атеросклероз, рак [47], клеточные травмы и процессы старения [48, 49], поэтому, чтобы избежать вредного воздействия АФК, они могут быть заблокированы антиоксидантными веществами, которые улавливают свободные радикалы и выводят токсины из организма [50].Антиоксиданты — это те соединения, которые могут защищать человеческий организм от воздействия свободных радикалов и АФК и замедлять развитие многих хронических заболеваний путем ингибирования окисления липидов или других молекул путем ингибирования инициирования или распространения окислительных цепных реакций, которые предполагают, что галловая кислота, эвгенол и эвгенилацетат использовались в качестве основных антиоксидантов [51, 52, 53, 54, 55]. Было обнаружено, что антиоксидантная активность эвгенола и эвгенилацетата сопоставима с активностью природного антиоксиданта витамина Е ( α -токоферол).Эвгенол и iso -эвгенол также оказались антиоксидантами и ингибируют перекисное окисление лецитина, индуцированное системой Fe 2+ -H 2 O 2 [56].
Gülçin, et al. [57] сравнивают поглощение радикала DPPH гвоздичного масла с некоторыми искусственными антиоксидантами, такими как α -токоферол, BHT, Trolox и бутилированный гидроксианизол, и показывают, что антиоксидантная активность гвоздичного масла снижается следующим образом: Гвоздичное масло> BHT > α -токоферол> бутилированный гидроксианизол> Trolox.Многочисленные протоколы in vitro , включая DPPH, способность поглощения кислородных радикалов, антиоксидантную способность восстановления железа, 2-дезоксигуанозин, 2,20-азино-бис- (3-этилбензотиазолин-6-сульфоновую кислоту) (ABTS) и ксантиноксидазу, были проверены. использовали для исследования антиоксидантной активности водного экстракта S. aromaticum . После этих исследований они продемонстрировали, что потенциальное антиоксидантное свойство водного экстракта S. aromaticum может быть связано с сильной способностью отдавать водород, улавливанием перекиси водорода, свободных радикалов, супероксидной и хелатирующей способности металлов [58].Таким образом, высокая антиоксидантная активность гвоздичного масла обусловлена присутствием фенольных соединений эвгенола, тимола и эвгенола ацетата [59, 60, 61].
1.6 Противогрибковая активность
Было опубликовано несколько отчетов, которые демонстрируют противогрибковую активность гвоздичного масла, которая, кроме того, обусловлена присутствием фенольного соединения эвгенола и карвакрола [62]. Пинто, и др. в 2009 году [63] сообщил, что гвоздичное масло снижает количество специфического эргостерола клеточных мембран грибов, который лечит распространение грибковой инфекции.Эвгенол также подавляет рост мицелиальных грибов, дрожжей как патогенных грибов человека и грибов, рожденных из пищи [64, 65, 66]. Kumar et al. в 2012 году [67] сообщил, что фенольное соединение карвакрол обладает максимальной противогрибковой активностью против грибков Candida , таких как Candida albicans Candida tropicalis и Candida guilliermondii . Они также проявляли некоторую противогрибковую активность, загрязняющую хлебобулочные изделия грибами, такими как Eurotium spp., Aspergillus spp. и Penicillium spp . Эти результаты подтверждают возможность использования растительных эфирных масел в качестве альтернативы химическим веществам для сохранения хлебобулочных изделий [68]. Виуда-Мартос, и др. [69] также сообщил о противогрибковой активности гвоздичного масла против грибов, вызывающих порчу пищевых продуктов Aspergillus flavus и Aspergillus niger , с использованием метода разбавления агаром, и было обнаружено, что оно проявляет более сильную ингибирующую активность против Aspergillus niger , чем Aspergillus flavus . .Bansod & Rai в 2008 г. [70] дополнительно изучил сравнительную противогрибковую активность различных эфирных масел, включая гвоздичное масло, против Aspergillus fumigates и Aspergillus niger , используя три различных метода: метод дисковой диффузии, метод разбавления бульона и метод разбавления агаром. Их исследования также подтверждают сильную противогрибковую активность гвоздичного масла среди всех других масел. Эстрада-Кано, и др. , 2017 [71] сообщают о сильном противогрибковом ингибирующем действии инкапсулированного гвоздичного масла против Fusarium oxysporum методом оксфордской чашки.На основании экспериментальных наблюдений они пришли к выводу, что голое гвоздичное масло вначале имело наибольшее ингибирующее действие против Fusarium oxysporum . Но через 8 часов эффективность микрокапсулированного гвоздичного масла была максимальной.
1.7 Противораковая активность
Рак является основной проблемой общественного здравоохранения во всем мире, и число пациентов постоянно увеличивается [72]. Это вторая по значимости причина смерти. Когда рак диагностируется на поздней стадии, химиотерапия является наиболее эффективным способом улучшить жизнь пациента и продлить время выживания [73].Недавний обзор литературы показал, что некоторые натуральные продукты играют важную роль в качестве противоопухолевых средств. Установлено, что почти 30-40% лекарств, используемых в мире для лечения рака, получают из растительных источников [74]. Сушеные бутоны гвоздики содержат широкий спектр биоактивных соединений, в том числе эвгенол, β -кариофиллен, гумулен, хавикол, метилсалицилат, α -иланген и эвгенон; флавоноиды евгенин, рамнетин, кемпферол и эвгенитин; тритерпеноиды, такие как олеаноловая кислота, стигмастерол и кампестерин; и несколько сесквитерпенов были проверены как противораковые средства.Хайчжоу Лю, и др. [75] сообщил, что экстракт гвоздики проявляет сильную цитотоксическую активность против нескольких линий раковых клеток человека. Было проведено исследование in vivo противоопухолевой активности экстракта гвоздики с помощью ряда экспериментов на различных раковых клетках, таких как раковые клетки человека, включая клетки рака яичников (SKOV-3), эпителиальные клетки шейки матки (HeLa), клетки рака печени (BEL- 7402), клетки рака толстой кишки (HT-29), клетки рака груди (MCF-7), клетки поджелудочной железы (PANC-1), нормальные эпителиальные клетки толстой кишки (CCD 841 CoN) и нормальные фибробласты легких (IMR-90) по порядку для идентификации биологически активных соединений.Сообщалось, что ингибирующая способность высушенного порошка гвоздики в клетках HT-29 рака толстой кишки человека при использовании анализа МТТ составляла приблизительно 2,2 мг / мл. Антипролиферативные эффекты были усилены примерно в 10 раз при использовании этанольного экстракта порошка гвоздики против панели линий раковых клеток человека, включая клетки рака груди (MCF-7), яичников (SKOV-3), шейки матки (HeLa). , клетки печени (BEL-7402), поджелудочной железы (PANC-1) и толстой кишки (HT-29). После идентификации ингибирующей активности они экстрагировали биоактивное соединение из этилацетатного экстракта гвоздики и обнаружили, что олеаноловая кислота была одним из биоактивных компонентов, обладающих антипролиферативной активностью in vitro, и in vivo, противоопухолевой активностью.Кумар, и др. (2014) [76] сообщил о противоопухолевом биоактивном компоненте различных концентраций воды, этанольного экстракта и эфирного масла гвоздики in vitro с помощью MTT и теста на летальность креветок (BSLT) против клеток рака груди человека MCF-7. Как в MTT, так и в BSLT эфирное гвоздичное масло показало превосходный цитотоксический эффект. В другом исследовании Lesgards, et al. (2014) [77] сообщил, что эфирное масло гвоздики содержит фенилпропаноиды, а терпеноиды проявляют противоопухолевую активность как на клеточной линии, так и на опухолях у животных.Dwivedi, et al. (2011) [78] провели сравнительное исследование противоопухолевого потенциала гвоздичного масла, его этанола и водного экстракта в отношении рака простаты DU-145, рака шейки матки HeLa, рака пищевода TE-13, MDA-MB-231 (ER-ve). и рак груди MCF-7 (ER + ve) вместе с нормальными лимфоцитами периферической крови человека для антипролиферации с использованием анализа MTT. Максимальная цитотоксическая активность и максимальная гибель клеток наблюдались в клетках TF-13 в течение 24 часов при использовании эфирного гвоздичного масла до 300 мкМ л / мл, тогда как минимальная гибель клеток в клетках DU-145, но с той же дозой, не была обнаружена в цитотоксичности. мононуклеарные клетки периферической крови человека.
1.8 Противовоспалительное действие
Гвоздика хорошо известна своим противовоспалительным действием. Традиционное употребление гвоздичного масла в качестве лосьона или употребление гвоздики в чае в повседневной жизни может помочь уменьшить воспаление, вызванное артритом. Предыдущие исследования [79] показали, что их ежедневное употребление может принести большую пользу для здоровья. Susan, et al. пришли к выводу, что разрывы цепей ДНК и воспалительные биомаркеры являются хорошей функциональной мерой биодоступности пищи.Han, et al. [80] недавно предоставил важные доказательства индуцированной эфирным маслом гвоздики (CEO) противовоспалительной активности и активности ремоделирования тканей в дермальных фибробластах человека. CEO в концентрации 0,011% проявляют стойкое антипролиферативное действие на фибробласты кожи человека. Он значительно ингибировал повышенную продукцию нескольких провоспалительных биомаркеров, таких как молекула адгезии сосудистых клеток-1 (VCAM-1), интерферон-индуцируемый α-хемоаттрактант Т-клеток (I-TAC), индуцированный интерфероном γ белок 10 (IP-10). , и монокин, индуцированный γ-интерфероном (МИГ).CEO также значительно ингибировал молекулы белка ремоделирования ткани, а именно коллаген-I, коллаген-III, фактор, стимулирующий колонию макрофагов (M-CSF), и тканевой ингибитор металлопротеиназы-2 (TIMP-2). Кроме того, он значительно модулировал глобальную экспрессию генов и изменял сигнальные пути, важные для воспаления, ремоделирования тканей и сигнальных процессов рака. CEO значительно ингибировал VCAM-1 и коллаген-III на уровнях экспрессии как белка, так и генов. Это исследование также подтверждает противоопухолевые свойства CEO и его основного активного компонента эвгенола.Barboza et al. [81] сообщили, что эвгенол, входящий в состав гвоздики, оказывает благотворное действие на окислительный стресс за счет ингибирования ферментов и окислительных процессов, что связано с профилем противовоспалительного препарата этого соединения. Sugihartini et al. [82] также сообщили, что активность эвгенола, компонента эфирного масла гвоздики в мази на абсорбирующей основе, может быть увеличена с добавлением усилителя. Его активность была лучше, чем у диклофенака натрия в положительном контроле.Однако формулу, содержащую пропиленгликоль, необходимо оценивать на предмет ее противовоспалительной активности в течение более длительного времени, чтобы гарантировать ее эффективность. Banerjee et al. [83] сообщили, что эмульсия гвоздичного масла может заменить местные продукты на химической основе для противовоспалительных и ранозаживляющих применений. Nikoui et al. [84] также сообщили, что введение гвоздичного масла оказывает противовоспалительное и жаропонижающее действие у собак после операции. Leem et al. [85] сообщили, что среди летучих дистиллятных экстрактов 8 лекарственных трав, дистиллятный экстракт гвоздики проявил самую сильную антиоксидантную активность (IC 50 = 8.85 μ мкг / мл) и ингибирующей активности ЦОГ-2 (степень ингибирования составляла 58,15% при концентрации 10 μ мкг / мл), тогда как ингибирующая активность 15-LOX (степень ингибирования составляла 86,15% при концентрации 25 μ г / мл). мл) была второй по величине после Анжелики. Они также измерили антиоксидантную и противовоспалительную активность эвгенола и его производных (метилэвгенола и ацетилэвгенола), эвгенола (IC 50 = 5,99 мкг / мл), которые проявили наивысшую антиоксидантную активность, тогда как метилэвгенол и ацетилэвгенол проявили небольшая активность.В случае ЦОГ-2 эвгенол (85,35%) в концентрации 20 мкМ мкг / мл показал самую сильную ингибирующую активность, тогда как в 15-LOX метилэвгенол (83,29%) в концентрации 20 мкл г / мл показал самую сильную ингибирующую активность, тогда как в 15-LOX метилэвгенол (83,29%) в концентрации 20 мкг / мл г / мл показал самую высокую ингибирующую активность.
1.9 Противодиабетическая активность
Kroda et al. [86] сообщил, что экстракт EtOH гвоздики ( Syzygium aromaticum ) значительно подавлял повышение уровня глюкозы в крови у мышей KK-A (y) с диабетом 2 типа.Результаты показывают, что гвоздика имеет потенциал в качестве функционального пищевого ингредиента для профилактики диабета 2 типа и что 2–4 в основном способствуют его гипогликемическому эффекту за счет активации PPAR-γ. In-vitro оценка показала, что экстракт обладал активностью связывания лиганда рецептора, активируемого пролифератором пероксисом человека (PPAR) -γ, в химерном анализе GAL4-PPAR-γ. Они выделили 8 соединений, из которых дегидродиэвгенол и дегидродиеугенол B обладали мощной активностью связывания лиганда PPAR-γ, а также показали, что они стимулируют дифференцировку преадипоцитов 3 T3-L1 посредством активации PPAR-γ, тогда как основная составляющая олеаноловая кислота в экстракте EtOH имела умеренную активность. .Topal [87] сообщил, что изоэвгенол (2-метокси-4- (проп-1-ен-1-ил) фенол), входящий в состав гвоздичного масла, проявляет превосходные ингибирующие эффекты против некоторых метаболических ферментов, а именно. ферменты ацетилхолинэстеразы (AChE), α -гликозидаза и α -амилаза. Изоэвгенол имеет значения IC 50 , составляющие 411,5, 19,25 и 77,00 нМ для α -амилазы, α -гликозидазы и AChE, соответственно. Значения K i изоэвгенола были обнаружены как 21 ± 9 и 16 ± 3 нМ против α -гликозидазы и AChE, соответственно, тогда как такрин в качестве стандартного ингибитора AChE показал значение IC 50 , равное 20.38 нМ. α -Гликозидазы, обычно называемые блокаторами крахмала, являются антидиабетическими препаратами, которые помогают снизить уровень пищевой глюкозы в крови. Chaudhry et al. [88] сообщил, что экстракт гвоздики обладает эффектом снижения глюкозы у крыс с СТЗ-индуцированным диабетом, и этот эффект зависит от дозы, и доза 750 мг / кг веса тела дала максимальный эффект. Абдулразак и др. [89] оценили влияние добавок гвоздики и ферментированного имбиря на уровень глюкозы в крови, сывороточного инсулина, рецепторов инсулина и лептина при сахарном диабете 2 типа, вызванном диетой с высоким содержанием жиров, у кроликов.Они обнаружили, что значительное (P <0,05) снижение уровня глюкозы в крови было зарегистрировано в группах, получавших добавки, по сравнению с контрольной группой с диабетом. Добавка гвоздики оказалась наиболее эффективной и поддерживающей антигипергликемическую активность, а также значительно снизила эффект на уровень лептина по сравнению с контрольной группой диабетиков. Значительное увеличение уровней инсулина было также обнаружено в группе, получавшей ферментированный имбирь, наряду с более высокими уровнями лептина по сравнению с контрольной группой.Таким образом, в целом исследование показывает, что добавки гвоздики и ферментированного имбиря обладают антидиабетическими свойствами и могут помочь в контроле гиперлептинемии при диабете 2 типа.
1.10 Гербицидная активность
Гвоздичное масло также проявляет гербицидную активность против многих гербицидов. Творкоски (2002) [90] исследовал гербицидное действие различных эфирных масел, включая масло гвоздики и корицы, на отдельные листья одуванчика. Эфирные масла гвоздики и корицы применялись с использованием анализа диска листьев одуванчика и анализа всего растения с травой Джонсона, ягненком обыкновенным и амброзией обыкновенной в теплице.Было замечено, что смерть от выстрела наступала от одного часа до одного дня после нанесения масла. Bainard, et al. (2006) [91] исследовали гербицидную активность гвоздичного масла и его основного составляющего эвгенола на Amaranthus retroflexus (красный свиный водоросль), Chenopodium album (обыкновенные ягнята) и его влияние на целостность мембран клеток листьев и рост проростков. Они заметили, что гвоздичное масло и входящий в его состав эвгенол вызывают снижение целостности клеточной мембраны и ингибирование роста проростков.Evans, et al. (2009) [92] выполнили свои исследования гербицидного действия гвоздичного масла и уксуса на Abutilon theopharst (бархатный лист) и Amaranthus retroflexus (краснокорневой поросенок). В своих исследованиях было обнаружено, что красный свиной корень легче контролировать с помощью обоих продуктов, чем бархатный лист. Park, et al. (2011) [93] дополнительно исследуют гербицидное действие гвоздичного масла на проростки огурцов в светлых и темных условиях. Обработка гвоздичным маслом увеличивала активность супероксиддисмутазы (СОД) и снижала активность каталазы, тогда как активность СОД и каталазы снижалась при обработке паракватом.Они сообщили, что масло гвоздики проявляет гербицидное действие по механизму, отличному от паркетного.
Помимо этого, они также обладают инсектицидной и анестезирующей активностью, и по этому поводу имеется несколько литературных источников. В ходе этих исследований мы обнаружили, что эфирные масла, содержащиеся в гвоздике, полезны и используются для лечения различных заболеваний.
Мы считаем, что финансовые барьеры не должны мешать исследователям публиковать свои выводы. В связи с необходимостью сделать научные исследования более общедоступными и поддержать преимущества открытого доступа, все больше и больше учреждений и спонсоров выделяют ресурсы, чтобы помочь преподавателям и исследователям охватить O pen A ccess P ublishing F ees (ОАПФ) .Кроме того, IntechOpen предоставляет несколько дополнительных вариантов, представленных ниже, все из которых доступны исследователям и могут обеспечить финансирование вашей публикации в открытом доступе.
В IntechOpen большая часть OAPF оплачивается учреждением автора или финансирующим агентством — учреждения (73%) против авторов (23%).
Первый шаг в получении средств для публикации в открытом доступе начинается с вашего учреждения или библиотеки. Издательские стандарты IntechOpen соответствуют большинству программ институционального финансирования.Мы советуем обратиться в ваше учреждение с просьбой о помощи в финансировании вашей публикации в открытом доступе.
Однако по мере того, как открытый доступ становится более широко используемым вариантом публикации для распространения научного и научного контента, в дополнение к учреждениям растет число спонсоров, которые разрешают использовать гранты для покрытия затрат на публикацию открытого доступа или учредили отдельные фонды для той же цели.
Пожалуйста, обратитесь к нашей странице Open Access Funding, чтобы изучить некоторые из этих возможностей финансирования и узнать больше о том, как вы могли бы профинансировать публикацию IntechOpen.Имейте в виду, что этот список не является исчерпывающим, и, хотя мы постоянно обновляем и информируем наших авторов о новых возможностях финансирования, мы рекомендуем вам всегда сначала уточнять у вашего учреждения.
Для авторов, которые не могут получить финансирование от своего учреждения или организаций, финансирующих исследования, но все еще нуждаются в помощи в покрытии расходов на публикацию, IntechOpen предлагает возможность подачи заявления на отказ от прав.
Наша миссия — поддерживать авторов в публикации их исследований и оказывать влияние на научное сообщество.В настоящее время 14% авторов получают полное освобождение и 6% получают частичное освобождение .
Предоставляя поддержку и консультации всем нашим международным авторам, приоритет отказа будет отдаваться тем авторам, которые проживают в странах, которые классифицируются Всемирным банком как страны с низкими доходами. Таким образом, мы можем гарантировать, что проводимая научная работа может оказать влияние на мировое научное сообщество, независимо от того, где проживает Автор.
Процесс подачи заявки начинается после того, как присланная вами рукопись была принята к публикации.Чтобы подать заявку, заполните форму запроса об отказе от прав и отправьте ее своему менеджеру по обслуживанию авторов. Если у вас есть официальное письмо из вашего университета или учреждения, подтверждающее, что средства на вашу публикацию открытого доступа недоступны, приложите и его. Запрос об отказе от прав обычно рассматривается в течение одной недели с даты подачи заявления. Все отделения, которые получают отказы или частичные отказы, будут обозначены как таковые в Интернете.
Не стесняйтесь обращаться к нам по адресу [email protected], если у вас есть какие-либо вопросы о вариантах финансирования или нашей программе отказа.Если вы уже начали процесс и вам нужна дополнительная помощь, обратитесь к своему менеджеру по обслуживанию авторов, который готов помочь вам!
Примечание: все данные, представленные выше, были собраны IntechOpen с 2013 по 2017 год.
В IntechOpen большинство OAPF оплачивает авторское учреждение или финансирующее агентство — учреждения (73%) против авторов (23%) ).
Первый шаг в получении средств для публикации в открытом доступе начинается с вашего учреждения или библиотеки.Издательские стандарты IntechOpen соответствуют большинству программ институционального финансирования. Мы советуем обратиться в ваше учреждение с просьбой о помощи в финансировании вашей публикации в открытом доступе.
Однако по мере того, как открытый доступ становится более широко используемым вариантом публикации для распространения научного и научного контента, в дополнение к учреждениям растет число спонсоров, которые разрешают использовать гранты для покрытия затрат на публикацию открытого доступа или учредили отдельные фонды для той же цели.
Пожалуйста, обратитесь к нашей странице Open Access Funding, чтобы изучить некоторые из этих возможностей финансирования и узнать больше о том, как вы могли бы профинансировать публикацию IntechOpen. Имейте в виду, что этот список не является исчерпывающим, и, хотя мы постоянно обновляем и информируем наших авторов о новых возможностях финансирования, мы рекомендуем вам всегда сначала уточнять у вашего учреждения.
Для авторов, которые не могут получить финансирование от своего учреждения или организаций, финансирующих исследования, но все еще нуждаются в помощи в покрытии расходов на публикацию, IntechOpen предлагает возможность подачи заявления на отказ от прав.
Наша миссия — поддерживать авторов в публикации их исследований и оказывать влияние на научное сообщество. В настоящее время 14% авторов получают полное освобождение и 6% получают частичное освобождение .
Предоставляя поддержку и консультации всем нашим международным авторам, приоритет отказа будет отдаваться тем авторам, которые проживают в странах, которые классифицируются Всемирным банком как страны с низкими доходами. Таким образом, мы можем гарантировать, что проводимая научная работа может оказать влияние на мировое научное сообщество, независимо от того, где проживает Автор.
Процесс подачи заявки начинается после того, как присланная вами рукопись была принята к публикации. Чтобы подать заявку, заполните форму запроса об отказе от прав и отправьте ее своему менеджеру по обслуживанию авторов. Если у вас есть официальное письмо из вашего университета или учреждения, подтверждающее, что средства на вашу публикацию открытого доступа недоступны, приложите и его. Запрос об отказе от прав обычно рассматривается в течение одной недели с даты подачи заявления. Все отделения, которые получают отказы или частичные отказы, будут обозначены как таковые в Интернете.
Не стесняйтесь обращаться к нам по адресу [email protected], если у вас есть какие-либо вопросы о вариантах финансирования или нашей программе отказа. Если вы уже начали процесс и вам нужна дополнительная помощь, обратитесь к своему менеджеру по обслуживанию авторов, который готов помочь вам!
Примечание: все данные, представленные выше, были собраны IntechOpen с 2013 по 2017 год.
Функционирование калия и магния в фотосинтезе, транслокации фотосинтата и фотозащите — Tränkner — 2018 — Physiologia Plantarum
Введение
Почти все виды растений на Земле являются автотрофами и производят сложные углеродные соединения в процессе фотосинтеза, в котором улавливается солнечная энергия.Улавливание солнечной энергии является основой продуктивности, и чистое количество углерода, которое фотосинтетически ассимилируется растительностью в течение 1 года, так называемая чистая первичная продукция, предлагается на уровне 104,9 Пг (Поле 1998) (1 Пг = 10 15 г). Чистая первичная продукция считается доступным количеством энергии, которое может быть передано от растений на другие уровни трофических сетей в экосистемах (Haberl et al. 2007), например человеческое потребление сельскохозяйственных товаров. Предполагается, что сельскохозяйственное производство вырастет на 60–100% в следующие четыре десятилетия (Krausmann et al.2013), подчеркивая важность фотосинтеза и последующих процессов. Для неослабевающего фотосинтеза необходимы оптимальные условия, включающие оптимальное поступление питательных веществ. Калий и магний являются важными минеральными питательными веществами для растений, которые вносят решающий вклад в процесс фотосинтеза и последующий перенос фотоассимилятов на большие расстояния. Среди минеральных питательных веществ растениям обычно требуется азот (N) в наибольшем количестве, за ним следуют калий (K), фосфор (P), кальций (Ca) и магний (Mg).Растения усваивают как минерал Mg, так и K только в катионной форме (как Mg 2+ и K + ). Высокие концентрации K в почвенном растворе ингибируют поглощение Mg и могут вызывать дефицит Mg у растений (Salmon 1963, Heenan and Campbell 1981).
Внутри растений наиболее метаболически активный Mg связывается или включается в клеточные компартменты (Waters 2011), с самыми высокими концентрациями в хлоропластах (Karley and White 2009). Значительная доля Mg в листьях связана в качестве центрального атома в тетрапиррольном кольце молекул хлорофилла a и b , которые являются основными пигментами для фотосинтетического поглощения света (Wilkinson et al.1990). Напротив, K не является частью органических макромолекул и присутствует исключительно в своей ионной форме (K + ) или в слабых комплексах, из которых он легко заменяется (Marschner 2012). В клетках K + присутствует в двух основных пулах, вакуоли и цитозоле. Во втором случае наиболее распространенным катионом является K + . K и Mg взаимно участвуют в ряде физиологических процессов, включая активацию множества ферментов и регуляцию катион-анионного баланса (Marschner 2012).K + служит наиболее важным неорганическим осмотическим соединением в растительных клетках, и, таким образом, достаточное поступление K имеет решающее значение для регуляции тургорных процессов, таких как движение устьиц (Fischer 1968) и удлинение клеток (Mengel and Arneke 1982). K способствует диффузии CO 2 из атмосферы в хлоропласты (Jákli et al., 2017). K и Mg дополнительно участвуют в опосредованном флоэмой транспорте сахарозы и распределении фотоассимилятов в растении.
И K, и Mg играют важную роль в процессах, которые тесно связаны с фотосинтезом и транслокацией фотосинтеза (Рис. 1). До сих пор роль калия и магния в фотосинтезе и связанных процессах не была рассмотрена во всестороннем обзоре. Многие обзоры посвящены перемещению K или Mg внутри растений (Shaul 2002, Gierth and Mäser 2007, Britto and Kronzucker 2008, Szczerba et al. 2009), а также вкладу этих элементов (особенно K) в снятие стресса (Cakmak 2005, Wang et al.2013) или, в несколько более широких подходах, об их статусе в мировом сельском хозяйстве (Römheld and Kirkby 2010, Cakmak 2013, Gransee and Führs 2013, Zörb et al. 2014). Таким образом, наша основная цель состоит в том, чтобы восполнить этот пробел, предоставив всесторонний обзор процессов, на которые влияют, облегчают и регулируют Mg и K в фотосинтезе, транслокации фотоассимилятов и фотозащите.
Локализация анатомических, физиологических и биохимических реакций растений на дефицит калия и магния, связанных с фотосинтезом и транслокацией фотоассимилятов.1) Дефицит калия изменяет анатомию листа и, таким образом, диффузию мезофилла CO 2 . 2) Дефицит калия и магния изменяет ультраструктуру хлоропластов. 3) Mg является центральным атомом пигмента хлорофилла и также необходим для его биосинтеза. 4) И K, и Mg снижают количество и активность Rubisco, а Mg дополнительно действует при активации Rubisco. 5) K и Mg необходимы для транслокации фотоассимилятов источник-сток, так как они выполняют важные функции в апопластной загрузке флоэмы сахарозы.
Сначала мы выделим роль Mg и K в светонезависимых и светозависимых фотосинтетических процессах, включая (1) диффузию CO 2 через пограничный слой листа, устьица и мезофилл, (2) ультраструктуру хлоропластов, (3) синтез хлорофилла и (4) активация и активность Rubisco (рис. 1A – D). Затем мы оценим роль Mg и K в транслокации фотоассимилятов (Fig. 1E). Наконец, мы выделим светозащитные реакции, включая нефотохимическое тушение (NPQ), удаление активных форм кислорода (ROS), фотодыхание, движение хлоропластов и движение листьев.
Участие калия и магния в фотосинтезе
Скорость фотосинтетической ассимиляции CO 2 стабильна в широком диапазоне концентраций K и Mg в листьях. Однако давно признано, что для поддержания фотосинтетической активности сельскохозяйственных культур необходимо адекватное питание K и Mg (например, Cooper et al. 1966, Terry and Ulrich 1973, Peoples and Koch 1979). Ниже определенного порога для листа K скорость ассимиляции резко падает.Концентрация К в тканях, которая имеет решающее значение для фотосинтетического функционирования сельскохозяйственных культур (т. Е. Концентрация К в тканях, при которой нетто-ассимиляция составляет 95% от максимального значения в экспериментальных условиях), составляет от 12,5 до 14,0 мг К · г -1 сухого вещества в масличном рапсе ( Лу и др., 2016a), 13,0 мг г -1 в сое (Singh and Reddy 2017), 22,3 мг г -1 в подсолнечнике (Jákli et al. 2017) и 26,0 мг г -1 в хлопке ( Gerardeaux et al.2010) (критические значения для Singh and Reddy (2017), Jákli et al.(2017) и Gerardeaux et al. (2010) рассчитаны по модельным уравнениям, представленным в соответствующих публикациях, описывающих взаимосвязь между концентрацией калия в листьях и чистой ассимиляцией). О критических концентрациях Mg в листьях для фотосинтеза не сообщалось, но для достижения 90% максимального урожая могут потребоваться концентрации 0,7 мг г -1 сухого вещества листьев (Smith et al. 1985).
Ограничения по CO
2 диффузия
В светонезависимых фотосинтетических реакциях атмосферный CO 2 фиксируется посредством ферментативного карбоксилирования.Скорость фиксации CO 2 во время этого процесса зависит не только от био- и фотохимических процессов, но также от количества CO 2 , доступного для Rubisco в строме хлоропласта. Ряд сопротивлений ограничивает диффузию CO 2 из атмосферы во внутреннее воздушное пространство листа и далее через клеточную стенку, плазматическую мембрану, цитозоль, оболочку хлоропласта и строму (рис. 1A).
Диффузия пограничного слоя
Первое сопротивление движению CO 2 из атмосферы к поверхности листа создает пограничный слой листа.В этом тонком слое, прилегающем к поверхности листа, турбулентный обмен подавлен, и перенос молекул характеризуется диффузией (Schuepp 1993). Толщина пограничного слоя и, следовательно, его проводимость к диффузии CO 2 , определяется энергетическим балансом листа (Woodrow et al. 1990), а не биохимическими параметрами и, следовательно, вряд ли напрямую зависит от статуса питательных веществ в листьях. . Поэтому в исследованиях влияния питательных веществ на обмен CO 2 ему в значительной степени пренебрегали.Однако из-за своих осмотических свойств дефицит калия вызывает потерю тургорного потенциала (Carroll et al. 1994), что может существенно повлиять на наклон листьев растений. Проводимость пограничного слоя зависит как от перехвата излучения, так и от скорости ветра (Martin et al. 1999). Таким образом, изменение среднего наклона листа в условиях дефицита K может существенно повлиять на свойства пограничного слоя и, таким образом, изменить его проводимость на молекулярную диффузию, что повлияет на фотосинтез, транспирацию и эффективность использования воды (Collatz et al.1991, Schuepp 1993, Bridge et al. 2013). Этот эффект, по-видимому, уникален для дефицита K и вряд ли является результатом дефицита Mg, поскольку Mg не способствует осморегуляции. Однако нам не известны какие-либо исследования, напрямую связывающие питание калия или магния с проводимостью пограничного слоя, но следует учитывать его общую концепцию при переносе результатов измерений в листовой камере, где эффекты пограничного слоя в значительной степени устраняются за счет сильного перемешивания воздуха, в поле.
Устьичная проводимость
CO 2 попадает в лист через устьичные поры.Их отверстие опосредуется замыкающими клетками устьиц, которые создают первые активно регулируемое сопротивление движению CO 2 в листья. Важная роль K + в регуляции замыкающих клеток устьиц и эффективности движения устьиц хорошо задокументирована (Fischer 1968, Humble and Raschke 1971). K + — основной неорганический осмолит, управляющий изменениями тургора замыкающих клеток, необходимыми для движения устьиц. Динамическое регулирование устьичной апертуры требует быстрого обмена большого количества K + между апопластом, цитозолем и вакуолями замыкающих клеток устьиц (Roelfsema and Hedrich 2002, Andrés et al.2014). Транспорт K + через плазматическую мембрану замыкающих клеток устьиц опосредуется регулируемыми по напряжению внутренними выпрямляющими каналами K + и сопровождается притоком противоионов (NO 3 — , Cl — ) и синтез малата в цитозоле. Повышенные концентрации ионов уменьшают водный потенциал замыкающих клеток и вызывают приток воды. В результате увеличивается объем замыкающих клеток и открываются устьицы.
Из-за осмотической роли K в регуляции замыкающих клеток неудивительно, что устьичная проводимость часто снижается в условиях дефицита K (например,грамм. Терри и Ульрих 1973, Чжао и др. 2001, Якли и др. 2017). Тем не менее, независимо от важности K + в регуляции замыкающих клеток, изменения устьичной проводимости не ограничивают преимущественно фотосинтез, даже когда общие концентрации K в листьях ниже критического уровня для фотосинтеза (Sen Gupta et al.1989, Zhao et al. др. 2001, Джин и др. 2011). Вклад устьичных ограничений в общее ограничение фотосинтеза у подсолнечника оценивается примерно в 15% (Jákli et al.2017) и не увеличивались даже при сильном дефиците К. Доля устьичных ограничений даже снизилась с 55 и 50% до 36 и 20%, соответственно, у двух сортов К-дефицитного масличного рапса (Lu et al. 2016a).
Имеются данные о том, что скорость, с которой устьица реагируют на экологические или биохимические обратные связи, снижена у люцерны с дефицитом калия (Lindhauer 1983) и оливкового дерева (Erel et al. 2013). Erel et al. (2013) предположили, что функциональная замена K + на Na + может быть ответственной за изменения в скорости движения устьиц у оливок, поскольку транспорт Na + через клеточные мембраны медленный по сравнению с транспортом К + .
Некоторые сообщения указывают на то, что при комбинированном дефиците K и ограничении воды устьичный контроль может быть потерян у выращиваемых перлитом подсолнечника и оливковых растений, что приводит к подавлению закрытия устьиц, вызванного засухой (Fournier et al. 2005, Arquero et al. 2006) , Benlloch-González et al. 2008, 2010). В соответствии с общей концепцией авторы связывают свои результаты не с потерей осмотического контроля устьиц, а с ингибированием сигнального пути ABA из-за чрезмерной продукции этилена в листьях.Эти результаты не подтверждаются выводами Jákli et al. (2017), которые сообщают о снижении показателей устьичной проводимости у подсолнечника, лишенного калия и стрессированного ПЭГ, выращенного в питательном растворе, и Erel et al. (2013), которые не обнаружили какого-либо влияния дефицита калия на устьичную проводимость оливок, выращенных на перлите, подвергшихся воздействию засухи.
Происхождение этого противоречия остается спекулятивным, а эффект взаимодействия дефицита калия и ограничения воды все еще обсуждается.В условиях хорошо обводненной воды все считают, что устьичное функционирование хорошо сохраняется, даже когда концентрация калия в листьях ниже критических значений для фотосинтеза. Jákli et al. (2017) предположили, что снижение устьичной проводимости в условиях дефицита калия является реакцией на низкие скорости ассимиляции CO 2 с целью максимизации эффективности использования воды.
В отличие от K, Mg не выполняет активной функции по контролю устьиц.Следовательно, причинно-следственная связь, предложенная Sun et al. (2001), которые объяснили снижение ассимиляции CO 2 у Mg-дефицитной Pinus radiata сниженной устьичной проводимостью, кажется весьма маловероятным. Снижение чистой ассимиляции, которое обычно наблюдается при дефиците Mg, вызвано, скорее, неустотными факторами. Это предположение подтверждается многочисленными исследованиями, в которых сообщается об отсутствии влияния дефицита Mg на устьичную проводимость или концентрации CO 2 внутри листа (например,Lasa et al. 2000, Tang et al. 2012 г., Ян и др. 2012). Однако, в отличие от K (Lu et al. 2016a, 2016b, Jákli et al. 2017), количественный анализ фотосинтетических ограничений у Mg-дефицитных растений до сих пор не опубликован.
Проводимость мезофилла
После проникновения в лист через устьичную пору ряд сопротивлений дополнительно препятствует движению CO 2 через мезофилл листа и, следовательно, влияет на концентрацию CO 2 в хлоропластах.Эти сопротивления суммируются как (их обратная) «проводимость мезофилла» диффузии CO 2 и суммированы в недавнем обзоре Berghuijs et al. (2016). После диффузии через внутреннее воздушное пространство листа CO 2 растворяется в апопластной воде клеточных стенок. Находясь в жидкой фазе, CO 2 присутствует в виде растворенного CO 2 или HCO 3 — и далее диффундирует через поры клеточной стенки, плазматическую мембрану, цитозоль, оболочку хлоропласта и строма хлоропласта, в которой происходит ферментативная фиксация CO 2 .
Существует общепринятое мнение, что проводимость мезофилла снижается при низком содержании K, что чаще всего связано с анатомическими изменениями листа. Ограничения фотосинтеза, которые происходят из-за ограниченной диффузии CO 2 , связаны скорее с мезофиллом, чем с устьичной проводимостью (например, Terry and Ulrich 1973, Jin et al. 2011, Battie-Laclau et al. 2014). Jákli et al. (2017) сообщили, что доля общего ограничения фотосинтеза, связанная с проводимостью мезофилла, увеличилась с 45 до 64% на бессимптомной площади листьев K-дефицитного подсолнечника по сравнению с K-достаточными растениями, а Lu et al.(2016a) показали увеличение ограничения мезофилла с 28 до 50% в K-неэффективном масличном рапсе при низком предложении K. Однако они не обнаружили значительного влияния дефицита К на ограничения мезофилла у К-эффективных сортов масличного рапса. Battie-Laclau et al. (2014) также не смогли показать влияние дефицита К на ограничения мезофилла в эвкалипте, хотя абсолютные значения проводимости мезофилла снизились, что указывает на видоспецифичную совместную регуляцию между мезофиллом и биохимическими ограничениями фотосинтеза.
Многие переменные влияют на диффузию CO 2 внутри листьев. Газовая диффузия в значительной степени определяется анатомическими свойствами листа, такими как толщина листа и объемное отношение внутреннего воздушного пространства листа к клеткам мезофилла, которые оба влияют на удельную массу листа (SLM). Многочисленные исследования показывают, что дефицит калия значительно увеличивает SLM (O’Toole et al. 1980, Cakmak et al. 1994b, Pettigrew 1999, Gerardeaux et al. 2010, Jin et al. 2011, Lu et al.2016b). SLM листьев с дефицитом калия усиливается за счет уменьшения толщины листа (O’Toole et al. 1980, Battie-Laclau et al. 2014) и уменьшения внутреннего воздушного пространства листа (Zhao et al. 2001, Battie-Laclau et al. 2014) . Более того, K является осмотическим драйвером роста клеток, а дефицит K приводит к уменьшению размера клеток мезофилла (Mengel and Arneke 1982, Claussen et al. 1997). Уменьшение толщины листа, внутреннего воздушного пространства листа и размера клеток уменьшают площадь поверхности хлоропласта, открытую для внутреннего воздушного пространства листа (Giuliani et al.2013) и, следовательно, проводимость мезофилла. Кроме того, Лу и соавт. (2016b) наблюдали большее расстояние между хлоропластами и клеточными стенками в K-дефицитном Brassica napus , увеличивая общую длину пути диффузии жидкой фазы CO 2 и, таким образом, проводимость мезофилла.
Помимо влияния анатомических свойств, увеличение SLM также может быть связано с увеличением неструктурных углеводов в листьях с дефицитом калия (Cakmak et al.1994b, Gerardeaux et al. 2010). Gerardeaux et al. (2010) сообщают, что 15% увеличения SLM в условиях дефицита K было связано с увеличением концентрации растворимого сахара. На основании чисто физических исследований Carroll et al. (2014), Hölttä et al. (2017) предположили, что увеличение концентрации сахара может снизить скорость диффузии жидкой фазы CO 2 . Таким образом, повышенные концентрации сахаров в жидкой фазе мезофилла листа могут способствовать снижению проводимости мезофилла при дефиците калия.
Проводимость мезофилла не только чувствительна к анатомическим особенностям листа и концентрациям растворенных веществ, но может дополнительно зависеть от экспрессии белков, таких как аквапорины, которые способствуют диффузии CO 2 внутри листа. Kanai et al. (2011) продемонстрировали важность адекватного питания калием для функционирования аквапоринов для поглощения и транспортировки воды. Имеются убедительные доказательства того, что транспорт не только воды, но и CO 2 через плазматические и хлоропластные мембраны в значительной степени поддерживается аквапоринами, и сообщается, что снижение экспрессии аквапорина табака NtAQP1 может вызвать 20% -ное снижение проводимость мезофилла (Flexas et al.2006 г., Uehlein et al. 2008 г.). Хотя прямые экспериментальные доказательства все еще отсутствуют, возможно участие аквапориновой активности в общем снижении проводимости мезофилла при дефиците K.
Кроме того, диффузии CO 2 через цитозоль и строму хлоропластов, вероятно, будет способствовать активность угольных ангидраз, группы ферментов, которые катализируют взаимное превращение между CO 2 и HCO 3 — и могут поэтому увеличьте скорость доставки CO 2 в Рубиско (Berghuijs et al.2017). Повышенные концентрации К в листьях усиливали активность углеангидразы в листьях Brassica juncea (Mohammad and Naseem 2006), но общие доказательства связи между активностью карбоангидразы и проводимостью мезофилла в условиях дефицита К пока отсутствуют.
Обобщая влияние дефицита К на проводимость мезофилла, мы заключаем, что снижение фотосинтеза при дефиците К можно в значительной степени отнести к увеличению внутренней устойчивости листа к переносу CO 2 , что приводит к низкой проводимости мезофилла.Повышенные концентрации сахара и SLM, скорее всего, способствуют общему снижению проводимости мезофилла. Уменьшение площади поверхности хлоропластов, подвергающейся воздействию внутреннего воздушного пространства листа, и увеличение длины внутриклеточного пути диффузии жидкой фазы, по-видимому, являются наиболее важными факторами, ограничивающими диффузию мезофилла CO 2 . Вклад различных признаков в общее снижение проводимости мезофилла сильно зависит от вида (Tomás et al.2013) и также может варьироваться между разновидностями одного и того же вида (Lu et al.2016а, 2016б).
В отличие от калия, не существует исследований связи между питанием магнием и проводимостью мезофилла. Следовательно, мы можем только предполагать возможное влияние Mg на диффузию CO 2 через мезофилл листа. Подобно дефициту К, повышенная SLM наблюдается для листьев растений с дефицитом Mg (Cakmak et al.1994b, Li et al.2017), что объясняется накоплением растворимых углеводов в результате нарушенной загрузки флоэмы (например.грамм. Cakmak et al. 1994b, Lavon et al. 1995). Как обсуждалось выше, повышенные концентрации сахаров в жидкой фазе мезофилла листа могут влиять на его проводимость по диффузии CO 2 . Однако, как упоминалось ранее, необходим систематический анализ ограничений для количественной оценки вклада устьичных ограничений и ограничений мезофилла в фотосинтез Mg-дефицитных растений.
Ультраструктура хлоропластов
Ультраструктура хлоропластов образована организацией внутренней мембранной системы.Часть мембран складывается в цилиндрические структуры, которые образуют так называемые граны, в то время как часть мембран остается разложенной, ламеллы стромы. K + и Mg 2+ способствуют хорошо структурированной организации ламелл граны и стромы, поэтому поддерживают целостность хлоропластов, эффективность поглощения света, диффузию Рубиско и, как следствие, ассимиляцию углерода. В условиях дефицита Mg и K наблюдалось повреждение ультраструктуры хлоропластов как таковое: структура ламелл была нерегулярной, рыхлой и частично диспергированной в цитоплазме, а количество гран и ламелл было уменьшено (Hall et al.1972 г., Чжао и др. 2001, Цзя и др. 2008 г.). Напротив, увеличение концентрации Mg привело к появлению многих известных многослойных мембран (Rumak et al. 2010). Укладка граны в основном зависит от сил притяжения Ван-дер-Ваальса и электростатического отталкивания (Puthiyaveetil et al., 2017). Электростатическое отталкивание контролируется поверхностными зарядами мембраны и концентрацией ионов в среде, окружающей тилакоидные мембраны. Экранирование чистых отрицательных поверхностных зарядов K + в качестве одновалентного катиона и более эффективно Mg 2+ в качестве двухвалентного катиона снижает силу электростатического отталкивания.Puthiyaveetil et al. (2017) показали, что при низкой активности K + накопление затруднено, поскольку преобладают силы отталкивания. Напротив, при высоких концентрациях K + и Mg 2+ (> 100 мМ K + и 5 мМ Mg 2+ ) укладка гран была хорошо развита. Кроме того, предполагается, что изменения латеральной микроорганизации LHCII и последующее разделение двух фотосистем (ФС) происходит при низких концентрациях Mg 2+ (Stys 1995, Stoitchkova et al.2006 г.). Однако для укладки граны требуются более высокие концентрации Mg 2+ , чем для сегрегации PSs (Stoitchkova et al. 2006).
Образование Grana увеличивает объем стромы, таким образом, создается больше места для свободной диффузии макромолекул в густо переполненной строме (Anderson et al. 2008). Увеличение объема стромы может уменьшить скученность макромолекул и, таким образом, способствовать диффузии Rubisco и способствовать высокой скорости фиксации углерода (Anderson et al.2008 г.).
Хлорофилл
Хлорофилл a и b — основные пигменты для поглощения световой энергии, и для синтеза обоих пигментов требуется Mg. Обычной реакцией на дефицит Mg является снижение концентрации хлорофилла, что продемонстрировано в многочисленных исследованиях (Mengutay et al. 2013, Faust and Schubert 2016, Tränkner et al. 2016). Однако, если рассматривать хлорофилл a и b по отдельности, ответ неоднороден.В сахарной свекле концентрации хлорофилла a и b снизились из-за дефицита Mg (Hermans et al. 2004). В кофе реакция зависит от сорта, так как не было изменений в соотношении хлорофилла a / b в cv. Катуай (Да Силва и др., 2014), тогда как в cv. Концентрации хлорофилла Acaiá a / b колебались за время эксперимента без четкой корреляции с дефицитом Mg.
Биосинтез хлорофилла начинается с встраивания Mg в протопорфирин IX (рис.1С). Встраивание Mg катализируется Mg-хелатазой, ферментом, состоящим из субъединиц 40, 70 и 140 кДа, обозначенных субъединицами ChlI, ChlD и ChlH соответственно. Для встраивания Mg в протопорфирин IX каталитический цикл Mg-хелатазы проходит две стадии (Masuda 2008). Первым этапом является этап активации, на котором субъединицы I и D образуют комплекс с АТФ и Mg 2+ . Субъединица H связывается с протопорфирином IX и Mg 2+ и предположительно содержит активный сайт для хелатирования.Второй этап включает хелатирование Mg в протопорфирин IX, что требует гидролиза АТФ. Исследования in vitro показали, что хелатирование 1 магния требует гидролиза 15 АТФ, что делает хелатирование магния дорогой реакцией, связанной с гидролизом АТФ (Reid and Hunter 2004). Активность хелатазы Mg чувствительна к концентрации свободного Mg и, таким образом, зависит от суточного цикла. При освещении концентрация свободного Mg в строме хлоропласта может увеличиваться до 6 мМ, обеспечивая приток в путь биосинтеза хлорофилла (Reid and Hunter 2004). Vicia faba растений, выращенных в условиях дефицита Mg, показали снижение количества транскриптов субъединицы H хелатазы Mg даже в тканях, где концентрации Mg не опускались ниже критического уровня (Neuhaus et al. 2013). Следовательно, анализ обилия транскриптов Mg-хелатазы H может служить ранним индикатором дефицита Mg.
При биосинтезе хлорофилла в основном образуется хлорофилл a . Часть хлорофилла a окисляется до хлорофилла b хлорофиллидом a оксигеназой, но хлорофилл b может быть восстановлен обратно до хлорофилла a .Для разложения хлорофилла хлорофилл b сначала должен быть преобразован в хлорофилл a , поскольку производные хлорофилла b не катализируются в последующем пути. Таким образом, соотношение обоих пигментов строго регулируется взаимным превращением хлорофилла a и b (цикл хлорофилла) (Blankenship 2014, Sato et al. 2014). Хлорофилл a и b связаны со стабильностью светособирающего комплекса II (LHCII), а хлорофилл b имеет решающее значение для накопления LHCII (Sato et al.2015). Распад хлорофилла b запускает деградацию LHCII и, таким образом, участвует в регуляции светособирательной способности растений и подавлении фотоповреждений. Он также способствует ремобилизации питательных веществ, поскольку LHCII является белком, который содержится в большом количестве в хлоропласте и служит резервуаром питательных веществ в условиях дефицита (Sato et al. 2015). Таким образом, деградация хлорофилла при дефиците Mg может функционировать для высвобождения Mg и защиты клеток от фотоповреждения, запуская деградацию LHCII.
Аналогичным образом, общая концентрация хлорофилла часто снижается при дефиците калия (Longstreth and Nobel 1980, Bednarz and Oosterhuis 1999, Zhao et al. 2001, Lu et al. 2016b). Было высказано предположение, однако, что дегенерация хлорофилла не является первичным эффектом дефицита К, а скорее связана с чрезмерным продуцированием АФК (Cakmak 2005), на что будет ссылаться далее в тексте.
Активация и активность Rubisco
Магний имеет решающее значение для фиксации CO 2 , поскольку он непосредственно участвует в активации и активности Rubisco.Кроме того, Mg имеет решающее значение для синтеза белка, который влияет на изобилие белка Rubisco и, таким образом, косвенно влияет на фиксацию CO 2 . Было показано, что при дефиците Mg концентрация белка снижается в сахарной свекле (Faust and Schubert, 2016), пшенице (Mengutay et al. 2013) и кукурузе (Mengutay et al. 2013). Более низкие концентрации белка при дефиците Mg могут быть связаны с участием ионов Mg в ассоциации и активности рибосомных субъединиц (Yamamoto et al. 2010). Кроме того, предполагается, что Mg контролирует структурную гибкость многих рибосомных белков (Yamamoto et al.2010). Более 100 ионов магния связаны с большой рибосомной субъединицей (Петров и др., 2012), а часть ионов Mg формирует микрокластер магния, который содержит два тесно связанных иона Mg. Предполагается, что микрокластеры магния обеспечивают структурную целостность большой рибосомной субъединицы и помогают в поддержании компактной нативной структуры сборки (Petrov et al. 2012). Самый распространенный белок на Земле — это Рубиско, и около 30% общего белка в листьях растений (Jensen 2000) и примерно 40% всех стромальных белков составляет Рубиско (Hazra et al.2015). С помощью протеомного анализа Peng et al. (2015) показали, что содержание белка активазы Rubisco и Rubisco (Rca) было снижено при дефиците Mg у Citrus sinensis , что указывает на влияние количества и активности Rubisco, что согласуется с 88% снижением скорости ассимиляции в Mg-дефицитных листьях. . Однако фиксация CO 2 скорее контролируется количеством активного Rubisco, чем общим количеством белка Rubisco (Taylor and Andersson 1996).
Активность
Rubisco обычно измеряется в анализах in vitro, тогда как определение скорости ассимиляции CO 2 in vivo может служить мерой активности Rubisco.Многочисленные исследования показывают снижение активности Rubisco или скорости ассимиляции CO 2 при дефиците Mg у различных видов растений, таких как шпинат (Yuguan et al. 2009), цитрусовые (Tang et al. 2012, Li et al. 2017), кукуруза (Jezek et al. al.2015), подсолнечник (Lasa et al. 2000) и P. radiata (Laing et al. 2000, Sun et al. 2001). Активность Rubisco чувствительна к статусу Mg, что наблюдается по снижению скорости ассимиляции CO 2 у Mg-дефицитной сахарной свеклы и ячменя уже через 7 и 8 дней, соответственно, после начала дефицита Mg в испытаниях питательных растворов (Terry and Ulrich 1974, Tränkner et al.2016). Рубиско катализирует карбоксилирование и / или оксигенацию рибулозо-1,5-бисфосфата (RuBP) и тем самым инициирует путь производства сахара или светодыхательные пути. Для проявления каталитической активности остаток лизина в активном центре фермента должен быть карбамилирован активатором, не являющимся субстратом молекулы CO 2 , а лабильная карбаматная группа впоследствии стабилизируется ионом Mg до связывания субстрата RuBP. (Тейлор и Андерссон 1997, Портис 2003). Однако потеря активатора CO 2 и иона Mg может привести к прочному связыванию субстрата RuBP или других сахарных фосфатов без предварительного карбамилирования, ингибируя его активность (Taylor and Andersson 1997).Ким и Портис (2006) показывают, что низкие концентрации Mg резко увеличивают скорость дезактивации Rubisco и связывают это с декарбамилированием активных центров и более сильным связыванием RuBP с некарбамилированной формой Rubisco. Высвобождение связанного RuBP и других сахарных фосфатов достигается с помощью каталитического шаперона Rca (Kuriata et al. 2014). Rca обладает гидролитической АТФазной активностью, которая необходима для активирующей активности Rubisco. Когда Mg отсутствовал в среде для анализа, ферментативный оборот прекращался в изолированном Rca табака (Hazra et al.2015). Точно так же активность Rca была снижена в шпинате с дефицитом Mg (Yuguan et al. 2009). Hazra et al. (2015) предполагают, что существует второй сайт связывания магния на основе белка, который служит коактиватором, опосредуя контакт между субъединицами. Т.о., необходим и Mg, координированный с ATP и Mg для содействия сборке субъединиц, и полностью активированный комплекс лучше всего представлен Rca • Mg • ATP • Mg (Hazra et al. 2015). Каталитическая активация Rca специфична для Mg, поскольку замена Mg 2+ на Mn 2+ привела к 32-кратному снижению скорости оборота.
Высокие цитозольные концентрации сахарозы в Mg-дефицитных листьях, что является обычным явлением при дефиците Mg в результате снижения скорости экспорта флоэмы (Cakmak et al. 1994a), также должны подавлять экспрессию генов, участвующих в фотосинтезе (Neales и Incoll 1968, Jang and Sheen 1994, Sheen 1994).
Аналогичным образом, дефицит K снижает активность карбоксилирования Rubisco в хлоропластах. Это было показано измерениями in vitro (Weng et al.2007, Ху и др. 2015 г., Захур и др. 2017), а также оценками максимальной скорости карбоксилирования Rubisco, V c, max , на основе комбинированных измерений газообмена листьев и флуоресценции хлорофилла (Jin et al.2011, Battie-Laclau et al.2014, Erel et al. 2015, Якли и др., 2017). Иногда утверждают, что дефицит K ингибирует активацию Rubisco (Oosterhuis et al. 2013), что приводит к снижению скорости карбоксилирования RuBP. В более раннем сообщении по этому вопросу Peoples и Koch (1979) четко заявляют, что в очищенном экстракте Rubisco из K-дефицитной и K-достаточной люцерны специфическая активность RuBPc (= RuBP-карбоксилазы ), выделенной из сильно K-дефицитные и K-достаточные створки существенно не различались, что позволяет предположить, что K не участвует в активности RuBPc ».Напротив, при измерении активности карбоксилирования в неочищенном экстракте авторы обнаружили значительно повышенную активность Rubisco, экстрагированного из листьев с достаточным содержанием калия. Они пришли к выводу, что биосинтез Рубиско заметно подавляется при недостаточном поступлении калия. Это снова было показано на семядолях огурца (Ohya et al. 1986). Как правило, синтез белка оказывается чувствительным не только к Mg, но также и к дефициту K, поскольку K + участвует в рибосомной трансляции (Nissen 2000, Austin and First 2002).Следовательно, концентрация растворимых белков в тканях растений снижается при дефиците К, тогда как концентрация свободных аминокислот увеличивается (Besford 1975, Cakmak et al. 1994a, Li et al. 1997, Faust and Schubert 2016). Как следствие, количество Rubisco снижается в листьях с дефицитом калия. Более низкое содержание Rubisco (на основе площади листа) также было показано в листьях сои с дефицитом K (Wang et al., 2015), но ни общий растворимый белок, ни содержание Rubisco не зависели от статуса K в листьях цитрусовых (Lavon et al.1999). Авторы, однако, сообщают о более высоком общем содержании аминокислот и активности карбоксилирования Рубиско. Тем не менее, влияние K на количественные характеристики Rubisco, по-видимому, зависит от изучаемых видов и соответствующей стадии роста (Osaki et al. 1993).
Помимо количества Rubisco, поставка K может косвенно повлиять на деятельность Rubisco. Оптимальные концентрации хлоропластов K + (т.е. в диапазоне цитозольных концентраций K + около 100 мМ) вызывают более широкий диапазон оптимума pH для функционирования Rubisco, чем при дефиците K (Mengel and Kirkby 2001).Кроме того, подавление активности Rubisco может происходить при низких концентрациях CO 2 в хлоропласте (Galmés et al.2011), что является результатом снижения скорости переноса CO 2 в тканях мезофилла с дефицитом K (Jákli et al., 2017). По сравнению с дефицитом Mg, снижение скорости экспорта флоэмы приводит к высоким концентрациям цитозольной сахарозы (Cakmak et al. 1994a), что может вызывать ингибирование активности Rubisco по обратной связи (Goldschmidt and Huber 1992).
Кроме того, Jin et al.(2011), Battie-Laclau et al. (2014) и Erel et al. (2015) сообщают о снижении максимальной скорости переноса электронов (J max ) через ФС гикори, эвкалипта и оливковых листьев с дефицитом калия. J max связан с регенерацией RuBP, потребляющей АТФ и НАДФН (Farquhar et al. 1980), что подразумевает меньшее количество акцепторных молекул для фиксации CO 2 .
Подводя итог, нет никаких доказательств того, что K + участвует в активации Rubisco, а скорее регулирует активность Rubisco, влияя на его количество, его pH-оптимум и доступность субстрата (CO 2 и, возможно, RuBP).
Участие K и Mg в транслокации фотоассимилятов
Рост и метаболизм растений требуют транслокации углеводов из фотосинтетически активных тканей в приемные органы, такие как корни, цветы и семена. Загрузка сахарозы — основной формы транспорта углеводов в растениях — во флоэму существенно зависит от питания K и Mg (рис. 1E), и нехватка любого из этих элементов может существенно повлиять на эффективность переноса углерода на большие расстояния в пределах растения.Согласно гипотезе напорного потока, первоначально разработанной Мюнхом (1930), движение растворенных веществ во флоэме следует за градиентом гидростатического давления, который возникает из-за различий в концентрациях растворенных веществ. K + является наиболее распространенным катионом во флоэме и вместе с сахарозой и амино-N соединениями является наиболее важным осмотическим соединением сока флоэмы. Следовательно, он способствует скорости транспорта флоэмы (Lalonde et al. 2003), и концентрация K + в соке флоэмы Ricinus communis положительно коррелировала с уровнем внешнего поступления калия (Mengel and Haeder 1977).
При апопластной загрузке флоэмы сахароза должна активно транспортироваться против своего градиента концентрации через плазматические мембраны клеток-компаньонов флоэмы и ситовые элементы транспортерами сахарозы. Необходимая энергия генерируется H + -АТФазами, которые создают градиент протонов через плазматическую мембрану (Michelet and Boutry 1995). Низкие клеточные концентрации Mg могут приводить к диссоциации комплекса Mg • ATP и к снижению активности H + -АТФаз плазматической мембраны (Hanstein et al.2011). Как следствие, трансмембранный протонный градиент не может быть установлен, скорость экспорта флоэмы снижается и сахароза накапливается в ткани листа.
Помимо Mg, K играет решающую роль в загрузке сахарозы. Исследование нокаут-мутантов Arabidopsis akt2 / 3-1 показало, что трансмембранный потенциал, устанавливаемый H + -АТФазами, регулируется внутренним выпрямляющим калиевым каналом AKT2 / 3 (Deeken et al. 2002). Таким образом, AKT2 / 3 способствует основной загрузке и разгрузке сахарозы во флоэму и из нее.Путем множественных шагов посттрансляционной модификации он может быть преобразован в не выпрямляющий канал, который способствует быстрому оттоку K + из комплекса ситовый элемент / сопутствующие клетки (Gajdanowicz et al. 2011, Sandmann et al. 2011). Установленный градиент K + обеспечивает энергию, которая используется другими транспортерами для загрузки флоэмы. Этот механизм, так называемая «калиевая батарея», служит мобильным источником энергии для преодоления местного дефицита АТФ и поддержания эффективности системы транспортировки на большие расстояния (Gajdanowicz et al.2011 г., Дрейер и др. 2017).
Нарушение распределения углерода от источника к поглотителю при дефиците Mg было показано на многих растениях, таких как сахарная свекла (Hermans et al. 2005), кукуруза (Mengutay et al. 2013), фасоль (Cakmak et al. 1994a) и соя ( Ян и др., 2017). Помимо сахарозы, крахмал может накапливаться в хлоропласте при дефиците Mg (Lavon et al. 1995, Ceylan et al. 2016, Yang et al. 2017). Молекулярная регуляция, лежащая в основе накопления углерода при дефиците Mg, все еще неоднозначна.У сахарной свеклы содержание транскриптов специфического симпортера H + / сахарозы BvSUT1 было выше в Mg-дефицитных листьях, но содержание сахарозы было увеличено (Hermans et al. 2005). В V. faba, количество транскриптов изоформ H + -АТФазы плазматической мембраны было снижено, когда растения выращивали в условиях дефицита Mg (Neuhaus et al. 2013). Однако это было не для всех исследованных изоформ. Одна изоформа показала несколько увеличенное количество транскриптов при дефиците Mg, а другая изоформа могла быть обнаружена только при лечении с дефицитом Mg и отсутствовала при контроле поставки Mg (Neuhaus et al.2013).
Подобно Mg, сдвиг в разделении ассимилятов — с повышением концентрации углеводов в первичных листьях и пониженными концентрациями в корнях — а также снижение скорости экспорта сахарозы из флоэмы наблюдались у K-дефицитных Phaseolus vulgaris (Cakmak et al. al. 1994a, 1994b). Аналогичным образом, концентрация сахарозы была увеличена в функциональных листьях хлопчатника с дефицитом калия (Bednarz and Oosterhuis 1999, Hu et al.2015, Zahoor et al.2017).
Эти исследования ясно показывают сильное влияние Mg и K на ассимиляцию и выделение углерода. Однако остается неясным, как связаны статус Mg и K, накопление углеводов и вызванная углеводами обратная связь при фотосинтезе в физиологических и молекулярных аспектах.
K и участие Mg в фотозащите
Как указано в предыдущих абзацах, K и Mg влияют на фотосинтетическую способность, а дефицит одного из двух питательных веществ снижает ассимиляцию CO 2 (рис.2). При таком ограничении фотосинтетической способности поглощенная световая энергия превышает то, что может быть использовано в процессах фотосинтеза, что приводит к усиленному производству АФК, и растения страдают от окислительного стресса (Nishiyama et al. 2006, Takahashi and Badger 2011). АФК ингибируют восстановление фотоповрежденных PSII, преимущественно белка D1 в реакционном центре PSII, вызывая фотоингибирование (Takahashi and Badger 2011, Järvi et al. 2013). На фотоингибирование указывает параметр флуоресценции хлорофилла F v / F m (Niyogi 1999), который описывает максимальную эффективность, при которой свет, поглощенный ФСII, используется для восстановления Q A , первичного акцептора электронов хинона. в PSII (Baker 2008).Дефицит Mg и K вызывает неоднородные ответы у F v / F m . В цитрусовых с дефицитом Mg и сахарной свекле F v / F m было снижено (Hermans et al. 2004, Tang et al. 2012, Yang et al. 2012), тогда как в Mg-дефицитном подсолнечнике и Sulla carnosa , F v / F m не был затронут (Lasa et al. 2000, Farhat et al. 2015). Изменения в F v / F m при дефиците Mg были связаны с повреждением выделяющего кислород комплекса на ФСII или с накоплением восстановленного Q A (Yang et al.2012). Дефицит K вызвал уменьшение F v / F m у двух сортов хлопка (Wang et al. 2012), риса (Jia et al. 2008), оливкового (Erel et al. 2015) и кукурузы (Qu et al.2013), тогда как у ели обыкновенной и подсолнечника снижение F против / F m не наблюдалось (Barnes et al. 1995, Jákli et al. 2017). Два сорта сои, являющиеся толерантными и чувствительными к дефициту K, соответственно, по-разному реагировали на дефицит K: толерантный сорт не показал измененных значений F v / F m , тогда как чувствительный сорт показал значительное снижение F v / F m (Wang et al.2015). В заключение следует отметить, что дефицит Mg и K обычно не вызывает фотоингибирование, которое может зависеть от величины дефицита Mg и K в ткани листа, специфических для сорта реакций и условий окружающей среды и экспериментов.
Влияние дефицита калия и магния на процессы, связанные с фотосинтезом, транслокацией фотоассимилятов и фотозащитой. Первичные эффекты — это прямые анатомические, физиологические или биохимические реакции на дефицит K и / или Mg.Вторичные эффекты являются немедленными последствиями первичных эффектов и в конечном итоге подавляют фотосинтетическую ассимиляцию CO 2 . Третичные эффекты являются результатом ингибирования ассимиляции CO 2 и только косвенно связаны с дефицитом K и / или Mg. Существуют сложные петли обратной связи и взаимодействия. Анатомия листа, транслокация фотоассимилятов, синтез белка, активность Rubisco, ультраструктура хлоропластов, синтез и изобилие хлорофилла напрямую зависят от дефицита Mg и / или K.Измененная анатомия листа препятствует диффузии CO 2 через мезофилл и тем самым снижает концентрацию CO 2 в хлоропластах, что, в свою очередь, ограничивает ассимиляцию CO 2 прямо или косвенно, влияя на активность Rubisco. Нарушение транслокации фотоассимилятов приводит к накоплению сахарозы, которая напрямую влияет на активность Rubisco через ингибирование обратной связи и косвенно влияет на диффузию мезофилла CO 2 . Снижение активности Rubisco в конечном итоге ограничивает ассимиляцию CO 2 .Нарушенная ультраструктура хлоропластов и низкое содержание хлорофилла снижают общую фотосинтетическую способность, снова препятствуя ассимиляции. Как следствие, устьичная проводимость активно регулируется до низких скоростей ассимиляции CO 2 , чтобы повысить эффективность фотосинтетического использования воды. Низкая фотосинтетическая способность вызывает чрезмерное производство АФК и фотоокисление, например, хлорофилл. Как следствие, активируются фотозащитные механизмы.
Удаление АФК
Виды АФК, которые продуцируются в тилакоиде, должны быть немедленно детоксифицированы, прежде чем они диффундируют из места своего образования и повреждают молекулы-мишени (Asada 2006).Детоксикация хлоропласта достигается с помощью антиоксидантов, таких как аскорбатпероксидаза (APX), Fe- и CuZn-супероксиддисмутаза (SOD) и глутатионредуктаза (GR). Взаимодействие между образованием ROS и детоксикацией ROS строго контролируется и необходимо для неограниченной фотохимии. Дефицит Mg и K вызывает обычно повышенный уровень ROS и повышенную активность фермента, детоксифицирующего ROS. При низком содержании калия и магния повышенные концентрации H 2 O 2 или O 2 — были зарегистрированы на многих видах растений, таких как арабидопсис, кукуруза, рис, хлопок и кофе (Shin and Schachtman 2004, Weng и другие.2007, Ху и др. 2016, Да Силва и др. 2017). К-неэффективный сорт хлопка увеличивал концентрации O 2 — и H 2 O 2 в листьях и увеличивал активность SOD и APX, тогда как K-эффективный сорт хлопка увеличивал только APX, но не активность SOD, и ни H 2 O 2 , ни O 2 — концентрации не были затронуты (Wang et al. 2012). Озгюр Узильдай и др. (2017) показали явное влияние статуса Mg на генерацию АФК.Когда Mg отсутствовал в питательном растворе или подавался только с 75 мкМ, концентрация H 2 O 2 в листьях хлопчатника увеличивалась на 29 и 33% соответственно, но когда добавка Mg была увеличена до 150 мкМ, H 2 O 2 концентрация увеличилась на 13%. Точно так же листья ячменя, страдающие тяжелым дефицитом Mg, имели на 55% более высокую концентрацию H 2 O 2 по сравнению с контролем, хотя активность APX, GR и SOD была значительно увеличена.При умеренном дефиците активность APX, GR и SOD все еще увеличивалась, но концентрация H 2 O 2 снижалась до того же уровня, что и в контроле, что указывает на то, что при тяжелом дефиците Mg степень H 2 O 2 Производство превысило производительность по улавливанию (Трэнкнер и др., 2016).
В заключение, дефицит K и Mg вызывает окислительный стресс, на что указывает повышение уровней H 2 O 2 и O 2 —, но в ответ на это усиливается активность антиоксидантного фермента, что выводит токсины из части АФК.Однако не всегда удается снизить АФК до уровней, сопоставимых с уровнями у нестрессированных растений, в зависимости от уровня стресса, вызванного дефицитом калия и магния.
Нефотохимическая закалка
Нефотохимическое рассеяние избыточной поглощенной световой энергии в виде тепла, обычно определяемое как NPQ, расположенное в антенне LHCII, является важным фотозащитным механизмом (Ruban 2016). Рассеяние тепловой энергии как фотозащитный механизм может рассеивать до 75% поглощенной световой энергии (Niyogi 1999).Растения увеличивают рассеяние энергии в виде тепла в ответ на дефицит Mg и K, чтобы защитить PS от повреждения и поддерживать фотосинтетические характеристики. У трех сортов Citrus , подверженных дефициту Mg (Tang et al. 2012, Yang et al. 2012), и двух сортов риса (Jia et al. 2008) и подсолнечника, подверженных дефициту K (Jákli et al. 2017), NPQ был увеличен. Напротив, у S. carnosa , NPQ не был затронут, даже если Mg был исключен из питательного раствора (Farhat et al.2015). Поскольку Mg- и K-дефицитные растения обладают ограниченной способностью преобразовывать световую энергию в химическую энергию, высокая интенсивность света увеличивает образование NPQ, как показано в Mg-дефицитном P. radiata (Laing et al. 2000).
Фотодыхание
Помимо карбоксилирования, Rubisco катализирует оксигенацию RuBP, что приводит к фотодыханию. Фотодыхание считается важным фотозащитным механизмом, который может облегчить чрезмерное восстановление цепи переноса электронов и, таким образом, помогает предотвратить фотоингибирование и чрезмерное образование АФК (Lima Neto et al.2017). У хлопка реакция фотодыхания на дефицит K была специфичной для сорта и привела к увеличению фотодыхания у K-эффективных var. Liaomian18 и контрастное снижение фотодыхания у K-неэффективной вар. NuCOTN99 (Ван и др., 2012). Абсолютные значения фотодыхания также снизились у K-дефицитных листьев люцерны (Peoples, Koch, 1979). Интересно, что фотодыхание относительно увеличилось с 26 до 34% чистой ассимиляции, что указывает на сдвиг в специфичности карбоксилирования / оксигенации Rubisco.Прямое участие K в фотодыхании сомнительно, и изменения при дефиците K, скорее всего, являются результатом неспецифических фотозащитных реакций. Однако фотодыхание не реагировало на интенсивность света выше 400 мкмоль м 2 с -1 плотность потока фотосинтетических фотонов (PPFD) у K-дефицитных оливок, тогда как она увеличивалась с увеличением PPFD у достаточно снабженных растений (Erel et al. 2015 ).
Хлоропласт механизм
Светособирающие комплексы и ФС расположены в хлоропластах клеток мезофилла.Изменения в расположении и структуре хлоропластов влияют на сбор света и фотосинтез. Расположение хлоропластов в зависимости от интенсивности света было предложено Kasahara et al. (2002) в качестве фотозащитного средства, что было признано в обзоре Такахаши и Баджера (2011). При слабой интенсивности света хлоропласты накапливаются у клеточной стенки в вертикальном направлении к свету, чтобы эффективно поглощать слабый свет. При высокой интенсивности света хлоропласты располагаются на клеточных стенках параллельно свету, чтобы минимизировать поглощение избыточного света и максимизировать поглощение CO 2 из межклеточных воздушных пространств (Takahashi and Badger 2011).В листьях B. napus хлоропласты ответили на дефицит калия увеличением своего расстояния от клеточной стенки (Lu et al. 2016b). Обычно хлоропласты выделяются на поверхности клетки для поглощения CO 2 и света. Однако исследование Lu et al. (2016b) фокусируется на диффузионной проводимости CO 2 в мезофилле и не учитывает поглощение света. Тем не менее, он выявил измененную организацию хлоропластов при дефиците K, но необходимы дальнейшие исследования, чтобы описать этот эффект в отношении фотозащиты.То же самое относится и к Mg, где исследования движения хлоропластов в ответ на дефицит Mg полностью отсутствуют в настоящей литературе.
Листовое движение
Парагелиотропное движение листьев — это механизм, с помощью которого предотвращается поглощение избыточного света и, как следствие, избыточное давление возбуждения на реакционные центры. В году у Robinia pseudoacacia парагелиотропное движение листьев привело к эффективному средству снижения стресса из-за сильного света на ФСII (Arena et al.2008 г.). Уровень F v / F m , измеренный ближе к вечеру, был сопоставим с уровнем утром, тогда как в листьях, у которых ось была зафиксирована для предотвращения движения листьев и створок, F v / F m был сокращен ближе к вечеру. Авторы предполагают, что отсутствие восстановления указывает на повреждение PSII, предполагая, что F v / F m является хорошим индикатором фотоингибирования (Arena et al. 2008). Аналогичным образом, у молодых листьев сои изменения угла черешка и средней жилки снижали интенсивность света на поверхности листа и помогали защитить листья от сильного излучения (Jiang et al.2006 г.). Было показано, что парагелиотропизм является стратегией защиты от фотоингибирования у культур фасоли, подверженной водному стрессу (Pastenes et al. 2004) и Vitis california (Gamon and Pearcy 1989). Мы наблюдали сильные гелиотропные движения листьев у подсолнечника, выращенного в условиях дефицита калия (неопубликованные данные). Мы предполагаем, что помимо вышеупомянутого воздействия движения листьев на микроклимат, парагелиотропизм может играть роль в защите листьев подсолнечника от чрезмерного света. Показателем является то, что участки листьев, демонстрирующие симптомы дефицита К, почти затенены от любого падения света.Однако у подсолнечника это явление, по-видимому, является K-специфическим, поскольку Mg-дефицитные растения не проявляли такого поведения. Насколько нам известно, исследований парагелиотропизма при дефиците калия и его вклада в фотозащиту не проводилось. Необходимо провести дальнейшие исследования, чтобы выяснить, является ли парагелиотропное движение листьев активной стратегией фотозащиты или это реакция на снижение тургора в условиях дефицита К. Если это активная стратегия при низкоклеточном статусе K + , движение листьев может быть предпочтительной стратегией, поскольку это управляемый тургором и, следовательно, процесс, требующий K (Oosterhuis et al.2014), следовательно, ионы K + предпочтительно должны быть размещены в местах, где регулируется угол наклона листа.
Сводка
Mg и K прямо или косвенно участвуют во многих физиологических процессах, связанных с фотосинтезом (рис. 2). Снижение скорости чистой ассимиляции углерода наблюдается, когда какое-либо из питательных веществ не может быть поглощено растениями в достаточном количестве, а его концентрация в тканях листа падает ниже критического значения.Первичные эффекты дефицита K, которые могут быть связаны со снижением фотосинтетической способности или ассимиляции CO 2 , включают (1) снижение синтеза белка и, следовательно, количества Rubisco, (2) анатомические изменения в структуре листа, снижающие проводимость мезофилла и, следовательно, концентрация хлоропласта CO 2 и (3) ультраструктура хлоропласта. Хотя K участвует в осморегуляции замыкающих клеток устьиц, дефицит K не препятствует функционированию устьиц и оптимизации эффективности фотосинтетического использования воды.Первичные эффекты дефицита Mg также включают синтез белка и Rubisco, а также ультраструктуру хлоропластов. В отличие от K, Mg непосредственно участвует в синтезе хлорофилла и активации Rubisco, но о влиянии дефицита Mg на анатомию листа или проводимость мезофилла не сообщается. И калий, и магний напрямую влияют на загрузку сахарозы во флоэме, а дефицит этих питательных веществ приводит к накоплению сахарозы в исходных тканях, вызывая дальнейшее подавление активности Rubisco. Фотозащитные механизмы реагируют на дефицит как калия, так и магния, с потенциалом снижения F v / F m и повышением NPQ, активности антиоксидантных ферментов и фотодыхания.Разнообразные эффекты анатомии, физиологии и биохимии растений на фотосинтез, транслокацию фотосинтата и фотозащиту в условиях дефицита K и Mg характеризуются сложными взаимодействиями и межрегулирующими механизмами, и многие вопросы все еще остаются нерешенными.
Вклад авторов
M.