Роль белков в жизнедеятельности организма
Редко можно встретить человека, не слыхавшего о белках. О них упоминается почти во всех работах по питанию, о них же в своих выступлениях говорят диетологи – и медики, и натуропаты.
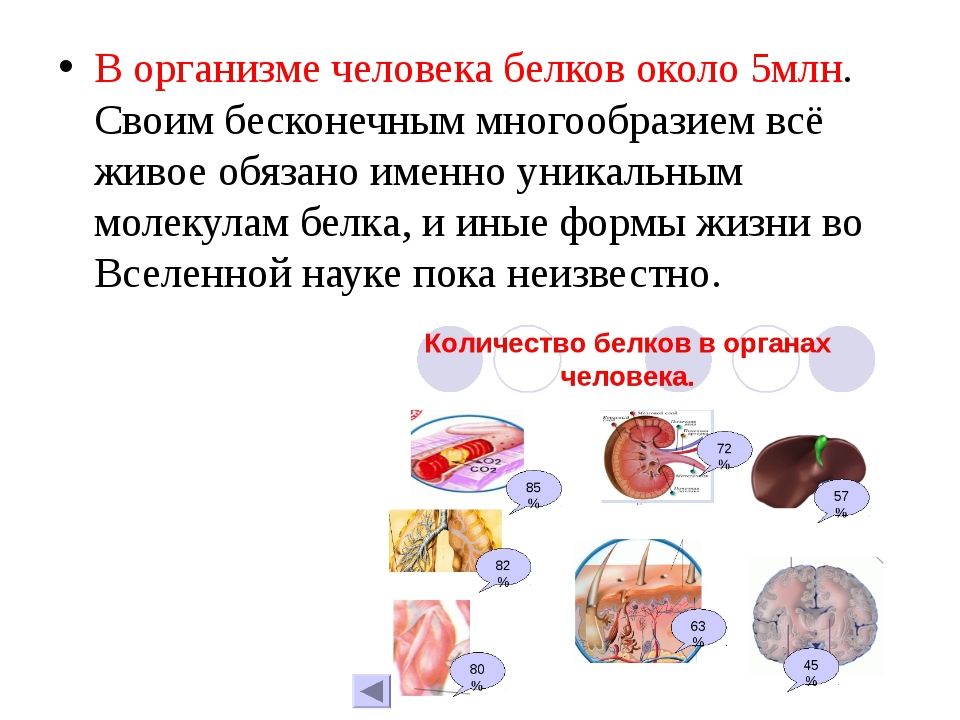
С точки зрения химика, белки – одни из самых сложных компонентов в пище. Значение их чрезвычайно велико, недаром Ф. Энгельс определил нашу биологическую жизнь как «способ существования белковых тел». В клетках человека их содержится в среднем около 20% от общей массы.
Одна из важнейших функций белков – строительная. Все органоиды клетки, мембраны и внеклеточные структуры в своей основе имеют белок. Нет белка – нет и органической жизни на Земле. (По крайней мере в том виде, в каком мы привыкли воспринимать жизнь.)
Белки выполняют и роль катализаторов (ферментов, или энзимов). Почти все химические превращения в живой природе протекают с участием ферментов. Причем каталитическая активность белков весьма специфична. Практически для каждой (!) реакции существуют свои ферменты. Без них реакции идти просто не могут, ведь энзимы ускоряют процессы в десятки и сотни миллионов раз.
Без них реакции идти просто не могут, ведь энзимы ускоряют процессы в десятки и сотни миллионов раз.
Еще одна функция белков – транспортировка необходимых соединений или химических элементов. Гемоглобин, например, переносит кислород, доставляя его в самые удаленные уголки тела, он же транспортирует углекислый газ.
Двигаемся мы также благодаря белкам. Все движения, на которые способны живые организмы – от поворота листьев растений и биения жгутиков простейших до перемещений животных, – все без исключения производятся за счет специального сократительного белка.
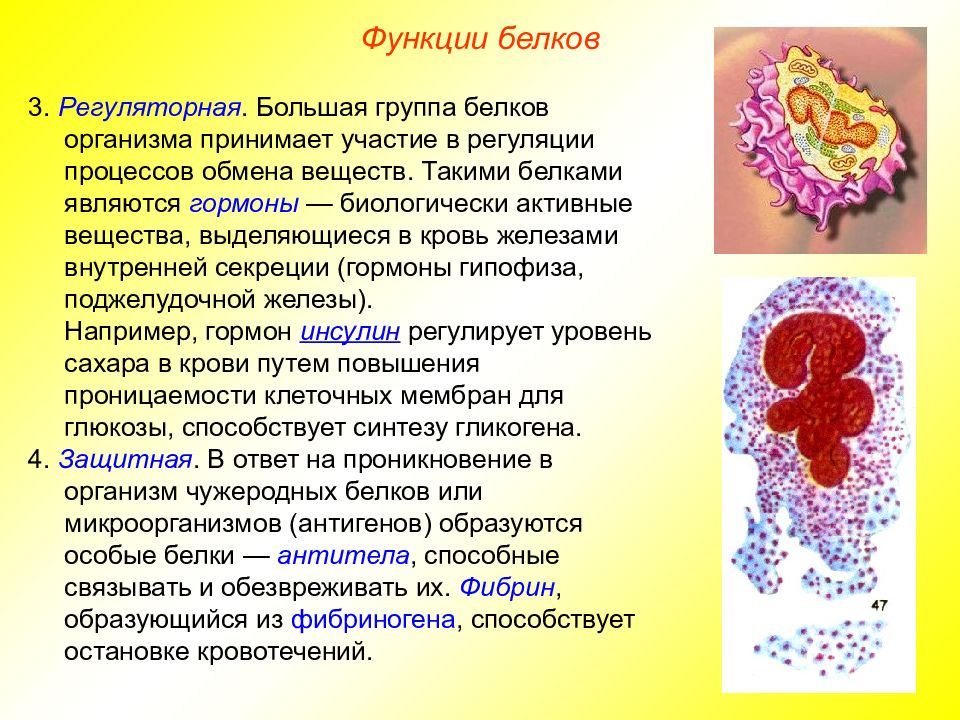
Белки выполняют и защитную функцию. При попадании в организм чужих белков или клеток вырабатываются особые белки – антитела, которые связывают и обеззараживают чужеродные вещества.
И наконец, белки могут служить источником энергии. Но это самое невыгодное «топливо».
Все белки построены из более-менее простых составляющих – аминокислот. Каждая из них наряду с углеродом, водородом и кислородом, входящими в органические соединения, обязательно содержит азот.
Известно около 80 природных аминокислот, но в обычной пище встречаются лишь 22 из них. Из этих элементарных «кирпичиков», стыкуемых в различном порядке, состоит все огромное многообразие белковых молекул. По оценкам ученых, в природе насчитывается около 10 10 –10 12 различных видов белков.
Помимо природных, существуют и синтетические аминокислоты. Из такой искусственной аминокислоты состоит, например, капрон, из которого делают и автомобильные покрышки, и одежду (ходить в которой йоги не советуют).
В природе же аминокислоты производятся живыми организмами. Считается, что 12 аминокислот может синтезировать и человек, поэтому они называются заменимыми. Остальные 10 аминокислот в обычных условиях человеческий организм не производит. Их называют незаменимыми.
Понятно, что незаменимые аминокислоты должны поступать с пищей. В зависимости от их наличия все белки даже подразделяют на «полноценные» (в которых эти аминокислоты присутствуют) и «неполноценные» (где их нет). Однако на практике об этом можно особо не задумываться. При более-менее разнообразном меню мы почти всегда получаем достаточное количество различных аминокислот, к тому же существует кишечная микрофлора, поставляющая массу необходимых соединений, плюс ко всему сам организм в экстремальных условиях или после соответствующей тренировки начинает их синтезировать. Потому-то сам факт «незаменимости» аминокислот некоторые ученые ставят под сомнение.
Однако на практике об этом можно особо не задумываться. При более-менее разнообразном меню мы почти всегда получаем достаточное количество различных аминокислот, к тому же существует кишечная микрофлора, поставляющая массу необходимых соединений, плюс ко всему сам организм в экстремальных условиях или после соответствующей тренировки начинает их синтезировать. Потому-то сам факт «незаменимости» аминокислот некоторые ученые ставят под сомнение.
Серьезные нарушения, вызванные неправильным обменом какой-либо аминокислоты, обычно встречаются только в результате некоторых заболеваний или при злоупотреблении лекарствами, а также при вынужденном недоедании или вынужденном однообразном питании.
Белки содержатся практически во всех натуральных продуктах. При переваривании белки расщепляются на аминокислоты, которые либо используются организмом для синтеза собственных белков, либо окисляются, то есть сжигаются как топливо. При окислении в числе прочих веществ образуется мочевая кислота, которая поступает в кровь и, по идее, должна выводиться почками. Если же организм ослаблен, а мочевой кислоты много (и то и другое – обычный результат злоупотребления мясным), она откладывается в тканях, вызывая подагру.
Если же организм ослаблен, а мочевой кислоты много (и то и другое – обычный результат злоупотребления мясным), она откладывается в тканях, вызывая подагру.
Часто говорят о «норме потребления» белков. Действительно, в каждый период жизни организм, несомненно, нуждается в каком-то определенном их количестве. Но эти потребности зависят от возраста, наследственности, темперамента, нагрузок, климата и множества других причин. Поэтому понятие «норма» здесь совершенно неприменимо.
В раннем детстве, когда потребность в белках наибольшая (за первый год жизни вес тела утраивается), все необходимые вещества ребенок получает с материнским молоком. Нельзя не признать, что это идеальный продукт, отлично обеспечивающий столь интенсивный рост. Между тем на долю белков в грудном молоке приходится лишь 7,4% его общей калорийности.
С возрастом, естественно, потребность в белках снижается. Ткани наращиваются все медленнее и медленнее, и к моменту зрелости на первый план выдвигается уже не строительная функция пищи, а энергетическая. Главным для организма становится компенсация текущих энергозатрат. Еще более отчетливо это проявляется у взрослых, а тем более у пожилых людей.
Главным для организма становится компенсация текущих энергозатрат. Еще более отчетливо это проявляется у взрослых, а тем более у пожилых людей.
Следовательно, доля белка в общей калорийности рациона должна снижаться. Но рассмотрим любопытную таблицу, приводимую Бирхер-Беннером, в которой он демонстрирует распределение калорийности пищи по питательным веществам.
|
Калории
белка, %
|
Калории
жира, %
|
Калории
углеводов, %
|
|
|
В материнском молоке
|
7,4
|
43,9
|
48,7
|
|
В коровьем молоке
|
21,3
|
49,8
|
28,9
|
|
В пище богатого человека
|
19,2
|
29,8
|
51,0
|
|
В пище бедного человека
|
16,7
|
16,3
|
66,9
|
|
В пище, крайне бедной белками
|
8,3
|
38,7
|
52,8
|
То есть получается, что потребление белков с возрастом не уменьшается, а увеличивается! Организм не может принять больше белка, чем ему необходимо, – это уже яд, и избыток обязательно должен быть сожжен. Так и образуются шлаки – конечные продукты белкового обмена: мочевая кислота, мочевина, аммиак, креатинин, креатин и другие. При избытке этих соединений выведение их затрудняется, и они задерживаются в организме, постепенно накапливаясь и нарушая все обменные процессы.
Так и образуются шлаки – конечные продукты белкового обмена: мочевая кислота, мочевина, аммиак, креатинин, креатин и другие. При избытке этих соединений выведение их затрудняется, и они задерживаются в организме, постепенно накапливаясь и нарушая все обменные процессы.
Разумеется, скорость освобождения от шлаков зависит от множества причин: соотношения прихода и расхода энергии, наличия витаминов, макро- и микроэлементов, физической активности, состояния органов и т. п. Но в любом случае белок – самое невыгодное «топливо». Его энергетическая ценность при окислении в организме составляет (по А. А. Покровскому) лишь 70,8 % от полной теплоты сгорания. Для жиров и усвояемых углеводов эти цифры соответственно 96,3 % и 100 %. Это значит, что 1 г белка при простом сжигании дает 5,65 ккал, а при окислении в организме – 4,0 ккал. А куда исчезает остальное? Остальное – шлаки.
Если учесть также, что избыток белка ведет к неоправданной интенсификации обменных процессов (а это способствует преждевременному изнашиванию, то есть старению тканей), то не таким уж парадоксальным кажется вывод Бирхер-Беннера – белок уменьшает ценность пищи. (По данным К. С. Петровского, белки на 30–40 % повышают основной обмен, жиры – на 4–14 %, углеводы – на 4–7 %.)
(По данным К. С. Петровского, белки на 30–40 % повышают основной обмен, жиры – на 4–14 %, углеводы – на 4–7 %.)
Разумеется, какое-то количество белков, и притом разнообразных, необходимо и взрослому человеку. Но даже в «обычной» пище их значительно больше, чем нужно. Иногда действительно не хватает какой-нибудь аминокислоты, но тогда человек инстинктивно набрасывается на нужную еду, и не надо следить за «достаточностью» белка, не надо «питать» организм белком, именно это и приносит вред.
Источник: В. А. Тутельян, А. И. Вялков, А. Н. Разумов, В. И. Михайлов, К. А. Москаленко, А. Г. Одинец, В. Г. Сбежнева, В. Н. Сергеев «Научные основы здорового питания»
Белки и их роль в организме
Голландском химику Герхарду Иоганну Мульдером (1883) приписывают введение в научную терминологию слова белок. Как в растениях, так и животных, по мнению ученого, имеется вещество, выполняет важную функцию. Она является одним из известных компонентов живой материи, и, видимо, без нее жизнь была бы невозможна. Это вещество Мульдер назвал белком или белками (от греч. Proteus — первый).
Это вещество Мульдер назвал белком или белками (от греч. Proteus — первый).
Более двухсотлетняя история химии белка полна непрестанным совершенствованием экспериментальных средств и богатая различные теоретические концепции. Весомый вклад в развитие химии белка сделали как отечественные ученые — А. Я. Данилевский, Н. Д. Зелинский, B. С. Садыков и др.., Так и зарубежные исследователи: Э. Фишер, Т. Курциус, М. Бергман, Ф. сенжери.
Химия белка всегда сочетала идеи и методы биологии, медицины, химии, физики. Белки составляют материальную основу химической деятельности клетки. Они делятся на простые (протеины) и сложные (протеиды). При гидролизе протеинов как продукты расщепления образуются только а-аминокислоты. Протеиды при гидролизе распадаются на белковую часть, состоящая из а-аминокислот, и небелковую, есть другие органические и неорганические соединения. Небелковой части называют простетической группой.
Протеиды обычно классифицируют по характеру простетич-ных групп:
— Гликопротеиды — содержат углеводы как небелковую часть;
— Липопротеиды — содержат липиды;
— Нуклеопротеиды — содержат нуклеиновые кислоты;
— Фосфопротеиды — содержат остаток фосфорной кислоты;
— Металлопротеидов — содержат зализопорфиринвмисни белки (миоглобин, гемоглобин), а также металлоферментов.
Рассмотрим особенности и свойства простых белков — протеинов.
Пептиды и белки — группа соединений, сходных по строению, которые отличаются только по размеру молекулы. Они полиамидами, образованные а-аминокислотами, имеют общий фрагмент Условно считают, что пептиды содержат в молекуле около 100 (молекулярная масса около 1000), а белки — более 100 аминокислотных остатков.
Различают олигопептиды (низкомолекулярные), содержащие в цепи не более 10 аминокислотных остатков, и полипептиды, содержащие от 10 до 100 аминокислотных остатков.
Белки являются основной составной частью мягкой структурной ткани животных и имеют большое значение в биологии.
Последние 100 лет ученые приложили немало усилий, пытаясь определить структуры различных белков, поскольку именно структура белка определяет его свойства.
Новые сведения о строении белковых молекул оказывают большую помощь при разработке средств лечения больных сердечно-сосудистые, наследственные заболевания, рак.
Впервые наличие пептидных групп в молекуле белка доказал профессор Харьковского университета А. Я. Данилевский (1883). Он изучал продукты гидролиза белка и обнаружил, что в результате неполного гидролиза белков образуются аминокислоты и остатки белковой молекулы, причем эти остатки дают биуретовую реакцию: при взаимодействии с щелочным раствором сульфата меди образуются соединения, окрашенные в фиолетовый цвет. При проведении полного гидролиза такая реакция не наблюдается. Поэтому А. Я. Данилевский предположил, что в основе строения белков заключается «биуретовая структура», то есть связь типа-HN-с-между остатками отдельных а-аминокислот с образованием полипеп-тидних цепей.
Связи между отдельными аминокислотами являются сильными. Их называют полипептидными, или «Биуретовая связями» А. Я. Данилевского. Полипептидных связей в макромолекуле много. Порядок чередования аминокислотных остатков в полипеп-тидних фрагментах определяет биологическую функцию и специфичность белка, называют первичной структурой белковой молекулы.
Например, сывороточный альбумин образует цепь, состоящий из более 500 аминокислотных остатков и выполняет двоякую функцию: поддерживает осмотическое (онкотическое) давление в крови и обеспечивает транспорт с кровью биологически активных веществ, например, лекарств. Фредерик сенжери (1953) — автор стратегического плана определения аминокислотной последовательности в белках («блочный» метод) — впервые обнаружил последовательность аминокислотных остатков в простейшем белка — инсулине, за что был удостоен Нобелевской премии 1958 В 1980 г. ему вторично была присуждена Нобелевская премию за работы, связанные с расшифровкой генетического кода. Инсулин состоит из двух пептидных цепей А и В, цепь А содержит 21, а цепь В — 30 аминокислотных остатков.
Оказалось, что в инсулине пептидная связь не является единственным химической связью в строении белка (рис. 6.3). Есть еще три дисульфид-ных связи, вызывающих искривление цепи А.
Действительно, молекула белка не лежит в плоскости, поэтому знание первичной структуры белка не достаточно для полного понимания его строения.
Американские ученые Л. Полинг и Р. Кори доказали, что в некоторых белках (миозин и актомиозин) несколько полипептидных цепей скрученные в спираль и образуют жгуты. При этом диаметр спирали всегда равна 0,54 нм, на отдельном развития размещается 3,6 аминокислотных остатков (рис. 6.4).
Это так называемая а-структура. Отдельные цепи в а-спирали могут быть соединены химическими (дисульфидными) и нехимическими связями. Водородные связи возникают между С = О и NH-группами полипептидных связей отдельных цепей. Конечно дисульфид-ных связей возникает мало, а водородных — много.
Как правило, водородные связи устанавливаются между первым и пятым аминокислотными остатками внутри одной цепи, приводит к скручиванию его в а-спираль. кем. Водородные связи устанавливаются и разрываются легко, без затраты большого количества энергии, и выполняют две важные биологические функции:
— Благодаря своей многочисленности обеспечивают устойчивость молекулы;
— Позволяют образовывать комплексы двух молекул. Условием комплексирования является принцип структурного соответствия (ком-плементарнисть).
Условием комплексирования является принцип структурного соответствия (ком-плементарнисть).
В некоторых белках полипептидные цепи располагаются параллельно друг другу в виде лент, которые могут образовывать структуру сморщенной ленты (рис. 6.5). Это происходит вследствие возникновения водородных связей между пептидными группами разных аминокислотных цепей. Образуется структура ß-сморщенной ленты (ß-фиброин).
За счет спирализации, или заключения в пучки, макромолекула приобретает трехмерности, образуя вторичную структуру белка.
В формировании третичной структуры белка наряду с водородными участвуют другие связи — электростатические (ионные), гидрофобные, дисульфидные.
Было доказано, что в целом для совокупности всех остатков аминокислот наблюдается закономерная локализация гидрофобных групп (углеводородных радикалов) во внутренней области макромолекулы. Гидрофильные (полярные) группы размещаются на поверхности молекул (рис. 6. 6).
6).
Третичную структуру белка называют субъединицей. Несколько субъединиц образуют четвертичное структуру белка.
В Кембриджском университете Д. Кендрю обнаружил третичную структуру белка миоглобина кашалота, который обеспечивает работу мышц с анаэробных условий и состоит из 153 остатков аминокислот и одного железосодержащего гема (Мг = 17000), а М. Пе-рутц установил четвертичное структуру гемоглобина. Этот белок содержит по две пары полипептидных цепей двух типов: две а-спирали (а1 и а2) из 140 остатков и две ß-структуры (ßl и ß 2) из 146 остатков а — аминокислот в каждой. К каждому полипептидной цепи прикреплен железосодержащий гем.
Четвертичная структура обусловлена комбинацией четырех субъединиц третичных структур, содержащихся друг возле друга слабыми связями (в основном силами Ван-дер-Вааль-са). Таким образом, особенностями строения биополимеров, определяющие их свойства, является существование двух различных типов связей (химических и нехимических) и пластичность цепей, обусловленная поворотом звеньев.
Благодаря поворота макромолекула может набирать разнообразной конформации — энергетически неравноценной формы молекул, возникающих при простом повороте без разрыва химической связи.
При изменении конформации макромолекулы могут или скручиваться, образуя глобулы и статические клубки или выпрямляться и состоять в ориентированные структуры-пачки.
Биологическая активность белков тесно связана с высокой организацией структуры. Живые организмы синтезируют белки нужной конформации, которая часто является метастабильной, т.е. из всех возможных структур не стабильной. Под влиянием нагрева, изменение значений рН среды или воздействия химических реагентов белки часто теряют свою биологически активную конформацию, превращаясь в случайные неорганизованные структурные единицы. Такой процесс называется денатурацией.
Белки. Их значение в питании человека реферат по биологии
Содержание 1 Виды белков………… ……………………………………………………………………………………. 3 2 Состав и свойства белков……………………………………………………………………………. 6 3 Функции белков…………………………………………………………………………………………. 9 4 Обмен белков……………………………………………………………………………………………… 11 5 Заключение……………………………………………………………………………………………….. 14 Введение Более 4 млрд лет назад на Земле из маленьких неорганических молекул непостижимым образом возникли белки, ставшие строительными 0 0 1 F 0 0 1 Fбло ками живых организмов. Своим бес конечным разнообразием всё 0 0 1 Fживое обязано именно уникальным молеку лам белка, и иные формы жизни во Вселенной науке пока неизвестны. Белки, или протеины (от греч. «протос» — «первый»), — это 0 0 1 F 0 0 1 Fприрод ные органические соединения, кото рые обеспечивают все 0 0 1 Fжизненные процессы любого организма.
……………………………………………………………………………………. 3 2 Состав и свойства белков……………………………………………………………………………. 6 3 Функции белков…………………………………………………………………………………………. 9 4 Обмен белков……………………………………………………………………………………………… 11 5 Заключение……………………………………………………………………………………………….. 14 Введение Более 4 млрд лет назад на Земле из маленьких неорганических молекул непостижимым образом возникли белки, ставшие строительными 0 0 1 F 0 0 1 Fбло ками живых организмов. Своим бес конечным разнообразием всё 0 0 1 Fживое обязано именно уникальным молеку лам белка, и иные формы жизни во Вселенной науке пока неизвестны. Белки, или протеины (от греч. «протос» — «первый»), — это 0 0 1 F 0 0 1 Fприрод ные органические соединения, кото рые обеспечивают все 0 0 1 Fжизненные процессы любого организма. Из бел ков построены хрусталик 0 0 1 Fглаза и па утина, панцирь черепахи и ядовитые вещества грибов… С помощью белков мы перевариваем пищу и боремся с болезнями. Благодаря 0 0 1 Fособым белкам по ночам светятся светлячки, а в глу бинах океана мерцают 0 0 1 Fтаинствен ным светом медузы. Белковых молекул в живой клетке во много раз больше, чем всех 0 0 1 Fдругих (кроме воды, разумеется!). Учёные вы яснили, что у большинства 0 0 1 F 0 0 1 Fорганиз мов белки составляют более полови ны их сухой массы. 0 0 1 FВпервые белок был выделен (в ви де клейковины) в 1728 г. итальянцем Якопо Бартоломео Беккари (1682— 1766) из пшеничной муки. 0 0 1 F 0 0 1 FЭто собы тие принято считать рождением хи мии белка. С тех пор почти за три столетия из природных источников получены тысячи различных белков и исследованы их свойства. плохо. Плохое усвоение растительного белка вызвано несколькими причинами: • толстые оболочки клеток растительных белков, часто не поддающиеся действию пищеварительных соков; • наличие ингибиторов пищеварительных ферментов в некоторых растениях, например, в бобовых; • трудности расщепления растительных белков до аминокислот.
Из бел ков построены хрусталик 0 0 1 Fглаза и па утина, панцирь черепахи и ядовитые вещества грибов… С помощью белков мы перевариваем пищу и боремся с болезнями. Благодаря 0 0 1 Fособым белкам по ночам светятся светлячки, а в глу бинах океана мерцают 0 0 1 Fтаинствен ным светом медузы. Белковых молекул в живой клетке во много раз больше, чем всех 0 0 1 Fдругих (кроме воды, разумеется!). Учёные вы яснили, что у большинства 0 0 1 F 0 0 1 Fорганиз мов белки составляют более полови ны их сухой массы. 0 0 1 FВпервые белок был выделен (в ви де клейковины) в 1728 г. итальянцем Якопо Бартоломео Беккари (1682— 1766) из пшеничной муки. 0 0 1 F 0 0 1 FЭто собы тие принято считать рождением хи мии белка. С тех пор почти за три столетия из природных источников получены тысячи различных белков и исследованы их свойства. плохо. Плохое усвоение растительного белка вызвано несколькими причинами: • толстые оболочки клеток растительных белков, часто не поддающиеся действию пищеварительных соков; • наличие ингибиторов пищеварительных ферментов в некоторых растениях, например, в бобовых; • трудности расщепления растительных белков до аминокислот. Рыбный белок. Установлено, что изолят рыбного белка еще значительно медленнее, чем казеин расщепляется до аминокислот. Расщепление изолята до пептидов не прекращалось даже через 3 часа с момента введения белка. Состав и свойства белков Белки — высокомолекулярные органические вещества, характерными особенностями которых является их строго определенный элементарный 0 0 1 Fсо став. Таблица 2. Состав белка. Наименование элемента 0 01 FСодержание элемен та (в %) Углерод Водород Азот Кислород Сера Зола 50-55 6,5-7,3 15-18 21-24 0-2,4 0-0,5 Особенно характерен для белков 15-18% уровень содержания азота. На заре белковой химии, когда не умели еще определять ни молекулярную массу белков, ни их химический состав, ни тем более структуру белковой молекулы, этот показатель играл большую роль при решении вопроса о 0 0 1 Fпринадлежно сти высокомолекулярного вещества к классу белков. Естественно, что сейчас данные об элементарном составе белков утратили свое былое значение для их характеристики. Белки вступают во взаимодействие с самыми различными веществами.
Рыбный белок. Установлено, что изолят рыбного белка еще значительно медленнее, чем казеин расщепляется до аминокислот. Расщепление изолята до пептидов не прекращалось даже через 3 часа с момента введения белка. Состав и свойства белков Белки — высокомолекулярные органические вещества, характерными особенностями которых является их строго определенный элементарный 0 0 1 Fсо став. Таблица 2. Состав белка. Наименование элемента 0 01 FСодержание элемен та (в %) Углерод Водород Азот Кислород Сера Зола 50-55 6,5-7,3 15-18 21-24 0-2,4 0-0,5 Особенно характерен для белков 15-18% уровень содержания азота. На заре белковой химии, когда не умели еще определять ни молекулярную массу белков, ни их химический состав, ни тем более структуру белковой молекулы, этот показатель играл большую роль при решении вопроса о 0 0 1 Fпринадлежно сти высокомолекулярного вещества к классу белков. Естественно, что сейчас данные об элементарном составе белков утратили свое былое значение для их характеристики. Белки вступают во взаимодействие с самыми различными веществами. Объединяясь друг с другом или нуклеиновыми кислотами, полисахаридами и липидами, они образуют рибосомы, митохондрии, 0 0 1 Fлизосомы, мембраны эн доплазматической сети и другие субклеточные 0 0 1 Fстрктуры, в которых благо даря пространственной организации белков и 0 0 1 Fсвойственной ряду из них фер ментативной активности осуществляются многообразные процессы обмена веществ. Поэтому именно белки играют выдающуюся роль в явлениях жизни. По своей химической природе белки 0 0 1 Fявляются гетерополимерами протеино генных аминокислот. Их молекулы 0 0 1 Fимеют вид длинных цепей, которые состо ят из аминокислот, соединенных пептидными связями. В самых маленьких полипептидных цепях белков содержится около 50 аминокислотных остатков. В самых больших — около 1500. В настоящее время первичная структура белка выявлена примерно у 2 тысяч белков. У инсулина, рибонуклеазы, лизоцима и гормона роста она 0 0 1 Fпод тверждена путем химического синтеза. Число аминокислотных остатков, входящих в молекулы отдельных белков, весьма различно: в инсулине 51, в миоглобине — около 140.
Объединяясь друг с другом или нуклеиновыми кислотами, полисахаридами и липидами, они образуют рибосомы, митохондрии, 0 0 1 Fлизосомы, мембраны эн доплазматической сети и другие субклеточные 0 0 1 Fстрктуры, в которых благо даря пространственной организации белков и 0 0 1 Fсвойственной ряду из них фер ментативной активности осуществляются многообразные процессы обмена веществ. Поэтому именно белки играют выдающуюся роль в явлениях жизни. По своей химической природе белки 0 0 1 Fявляются гетерополимерами протеино генных аминокислот. Их молекулы 0 0 1 Fимеют вид длинных цепей, которые состо ят из аминокислот, соединенных пептидными связями. В самых маленьких полипептидных цепях белков содержится около 50 аминокислотных остатков. В самых больших — около 1500. В настоящее время первичная структура белка выявлена примерно у 2 тысяч белков. У инсулина, рибонуклеазы, лизоцима и гормона роста она 0 0 1 Fпод тверждена путем химического синтеза. Число аминокислотных остатков, входящих в молекулы отдельных белков, весьма различно: в инсулине 51, в миоглобине — около 140. Поэтому и относительная молекулярная масса белков колеблется в очень широких пределах — от 10 тысяч до многих миллионов На основе определения относительной молекулярной массы и элементарного анализа установлена эмпирическая формула белковой молекулы — гемоглобина крови (C738h2166O208S2Fe)4 Меньшая молекулярная масса может быть у простейших ферментов и некоторых гормонов белковой природы. Например, молекулярная масса гормона инсулина около 6500, а белка вируса гриппа — 320 000 000. Вещества белковой природы (состоящие из остатков 0 0 1 Fаминокислот, соединенных между собой пептидной связью), имею щие 0 0 1 Fотносительно меньшую молекулярную массу и меньшую сте пень пространственной организации макромолекулы, называются полипептидами. Провести резкую границу между белками и полипептидами трудно. В большинстве случаев белки отличаются от других природных полимеров 0 0 1 F(каучука, крахмала, целлюлозы), тем, что чистый инди видуальный белок содержит только молекулы одинакового строения и массы.
Поэтому и относительная молекулярная масса белков колеблется в очень широких пределах — от 10 тысяч до многих миллионов На основе определения относительной молекулярной массы и элементарного анализа установлена эмпирическая формула белковой молекулы — гемоглобина крови (C738h2166O208S2Fe)4 Меньшая молекулярная масса может быть у простейших ферментов и некоторых гормонов белковой природы. Например, молекулярная масса гормона инсулина около 6500, а белка вируса гриппа — 320 000 000. Вещества белковой природы (состоящие из остатков 0 0 1 Fаминокислот, соединенных между собой пептидной связью), имею щие 0 0 1 Fотносительно меньшую молекулярную массу и меньшую сте пень пространственной организации макромолекулы, называются полипептидами. Провести резкую границу между белками и полипептидами трудно. В большинстве случаев белки отличаются от других природных полимеров 0 0 1 F(каучука, крахмала, целлюлозы), тем, что чистый инди видуальный белок содержит только молекулы одинакового строения и массы.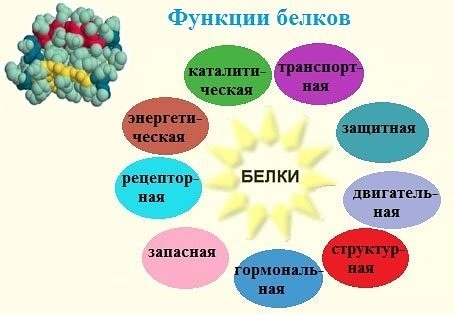 Исключением является, например, желатина, в составе которой входят макромолекулы с молекулярной массой 12 000— 70000. 0 0 1 FСтроением белков объясняются их весьма разнообразные свой ства. Они имеют разную растворимость: некоторые растворяются в воде, другие — в разбавленных растворах нейтральных солей, а некоторые совсем не обладают свойством растворимости (например, белки покровных тканей). При растворении белков в воде образуется своеобразная молекулярно- Сократительную функцию выполняют белки, в результате взаимодействиях которых происходит передвижение в пространстве, сокращение и расслабление сердца, движение других внутренних органов. Структурная функция белков заключается в том, что они составляют основу строения клетки; некоторые из них (коллаген соединительной ткани, кератин волос, ногтей и кожи, эластин сосудистой стенки и др.) выполняют почти исключительно структурную функцию. В комплексе с липидами (преимущественно фосфолипидами) белки участвуют в построении мембран клеток и внутриклеточных образований.
Исключением является, например, желатина, в составе которой входят макромолекулы с молекулярной массой 12 000— 70000. 0 0 1 FСтроением белков объясняются их весьма разнообразные свой ства. Они имеют разную растворимость: некоторые растворяются в воде, другие — в разбавленных растворах нейтральных солей, а некоторые совсем не обладают свойством растворимости (например, белки покровных тканей). При растворении белков в воде образуется своеобразная молекулярно- Сократительную функцию выполняют белки, в результате взаимодействиях которых происходит передвижение в пространстве, сокращение и расслабление сердца, движение других внутренних органов. Структурная функция белков заключается в том, что они составляют основу строения клетки; некоторые из них (коллаген соединительной ткани, кератин волос, ногтей и кожи, эластин сосудистой стенки и др.) выполняют почти исключительно структурную функцию. В комплексе с липидами (преимущественно фосфолипидами) белки участвуют в построении мембран клеток и внутриклеточных образований. Гормональную функцию выполняют белки-регуляторы обмена веществ. Они относятся к гормонам, которые образуются в железах внутренней секреции, некоторых органах и тканях организма. Питательная функция осуществляется белками, которые являются резервными, или питательными. Белки яйца обеспечивают рост и развитие плода, белки молока служат источником питания для новорожденного. Перечисленные функции белков являются наиболее важными и специфичными, но ими не ограничивается значение белков для жизнедеятельности организма. Обмен белков После расщепления белков в пищеварительном тракте образовавшиеся аминокислоты всасываются в кровь. В кровь всасывается также незначительное количество полипептидов — соединений, состоящих из нескольких аминокислот. Из аминокислот клетки нашего тела синтезируют белок, причем белок, который образуется в клетках человеческого организма, отличается от потребленного белка и характерен для человеческого организма. Образование нового белка в организме человека и животных идет беспрерывно, так как в течении всей жизни взамен отмирающих клеток крови, кожи, слизистой оболочки, кишечника и т.
Гормональную функцию выполняют белки-регуляторы обмена веществ. Они относятся к гормонам, которые образуются в железах внутренней секреции, некоторых органах и тканях организма. Питательная функция осуществляется белками, которые являются резервными, или питательными. Белки яйца обеспечивают рост и развитие плода, белки молока служат источником питания для новорожденного. Перечисленные функции белков являются наиболее важными и специфичными, но ими не ограничивается значение белков для жизнедеятельности организма. Обмен белков После расщепления белков в пищеварительном тракте образовавшиеся аминокислоты всасываются в кровь. В кровь всасывается также незначительное количество полипептидов — соединений, состоящих из нескольких аминокислот. Из аминокислот клетки нашего тела синтезируют белок, причем белок, который образуется в клетках человеческого организма, отличается от потребленного белка и характерен для человеческого организма. Образование нового белка в организме человека и животных идет беспрерывно, так как в течении всей жизни взамен отмирающих клеток крови, кожи, слизистой оболочки, кишечника и т. д. создаются новые, молодые клетки. Для того чтобы клетки организма синтезировали белок, необходимо, чтобы белки поступали с пищей в пищеварительный канал, где они подвергаются расщиплению на аминокислоты, и уже из всосавшихся аминокислот будет образован белок. Если же, минуя пищеварительный тракт, ввести белок непосредственно в кровь, то он не только не может быть использован человеческим организмом, он вызывает ряд серьезных осложнений. На такое введение белка организм отвечает резким повышением температуры и некоторыми другими явлениями. При повторном введении белка через 15-20 дней может наступить даже смерть при параличе дыхания, резком нарушение сердечной деятельности и общих судорогах. Белки не могут быть заменены какими-либо другими пищевыми веществами, так как синтез белка в организме возможен только из аминокислот. Для того чтобы в организме мог произойти синтез присущего ему белка, необходимо поступление всех или наиболее важных аминокислот. Из известных аминокислот не все имеют одинаковую ценность для организма.
д. создаются новые, молодые клетки. Для того чтобы клетки организма синтезировали белок, необходимо, чтобы белки поступали с пищей в пищеварительный канал, где они подвергаются расщиплению на аминокислоты, и уже из всосавшихся аминокислот будет образован белок. Если же, минуя пищеварительный тракт, ввести белок непосредственно в кровь, то он не только не может быть использован человеческим организмом, он вызывает ряд серьезных осложнений. На такое введение белка организм отвечает резким повышением температуры и некоторыми другими явлениями. При повторном введении белка через 15-20 дней может наступить даже смерть при параличе дыхания, резком нарушение сердечной деятельности и общих судорогах. Белки не могут быть заменены какими-либо другими пищевыми веществами, так как синтез белка в организме возможен только из аминокислот. Для того чтобы в организме мог произойти синтез присущего ему белка, необходимо поступление всех или наиболее важных аминокислот. Из известных аминокислот не все имеют одинаковую ценность для организма. Среди них есть аминокислоты, которые могут быть заменены другими или синтезированными в организме из других аминокислот; наряду с этим есть и незаменимые аминокислоты, при отсутствии которых или даже одной из них белковый обмен в организме нарушается. Белки не всегда содержат все аминокислоты: в одних белках содержится большее количество необходимых организму аминокислот, в других — незначительное. Разные белки содержат различные аминокислоты и в разных соотношениях. Белки, в состав которых входят все необходимые организму аминокислоты, называются полноценными; белки, не содержащие всех необходимых аминокислот, являются неполноценными белками. Для человека важно поступление полноценных белков, так как из них организм может свободно синтезировать свои специфические белки. Однако полноценный белок может быть заменен двумя или тремя неполноценными белками, которые, дополняя друг друга, дают в сумме все необходимые аминокислоты. Следовательно, для нормальной жизнедеятельности организма необходимо, чтобы в пище содержались полноценные белки или набор неполноценных белков, по аминокислотному содержанию равноценных полноценным белкам.
Среди них есть аминокислоты, которые могут быть заменены другими или синтезированными в организме из других аминокислот; наряду с этим есть и незаменимые аминокислоты, при отсутствии которых или даже одной из них белковый обмен в организме нарушается. Белки не всегда содержат все аминокислоты: в одних белках содержится большее количество необходимых организму аминокислот, в других — незначительное. Разные белки содержат различные аминокислоты и в разных соотношениях. Белки, в состав которых входят все необходимые организму аминокислоты, называются полноценными; белки, не содержащие всех необходимых аминокислот, являются неполноценными белками. Для человека важно поступление полноценных белков, так как из них организм может свободно синтезировать свои специфические белки. Однако полноценный белок может быть заменен двумя или тремя неполноценными белками, которые, дополняя друг друга, дают в сумме все необходимые аминокислоты. Следовательно, для нормальной жизнедеятельности организма необходимо, чтобы в пище содержались полноценные белки или набор неполноценных белков, по аминокислотному содержанию равноценных полноценным белкам. Поступление полноценных белков с пищей крайне важно для растущего организма, так как в организме ребенка не только происходит восстановление отмирающих клеток, как у взрослых, но и в большом количестве создаются новые клетки. Обычная смешанная пища содержит разнообразные белки, которые в сумме обеспечивают потребность организма в аминокислотах. Важна не только биологическая ценность поступающих с пищей белков, но и их количество. При недостаточном количестве белков нормальный рост организма приостанавливается или задерживается, так как потребности в белке не покрываются из-за его недостаточного поступления. Белки поступают в организм человека и животных с различными пищевыми продуктами, в которых содержание белка колеблется в широких пределах. Приведем таблицу, дающую представление о содержании белка в некоторых продуктах питания.
Поступление полноценных белков с пищей крайне важно для растущего организма, так как в организме ребенка не только происходит восстановление отмирающих клеток, как у взрослых, но и в большом количестве создаются новые клетки. Обычная смешанная пища содержит разнообразные белки, которые в сумме обеспечивают потребность организма в аминокислотах. Важна не только биологическая ценность поступающих с пищей белков, но и их количество. При недостаточном количестве белков нормальный рост организма приостанавливается или задерживается, так как потребности в белке не покрываются из-за его недостаточного поступления. Белки поступают в организм человека и животных с различными пищевыми продуктами, в которых содержание белка колеблется в широких пределах. Приведем таблицу, дающую представление о содержании белка в некоторых продуктах питания.
Press-room — IBCh RAS
Объявления →
- science news Barnase*Barstar-guided two-step targeting approach for drug delivery to tumor cells in vivo
November 19To reduce side effects in the process of oncotherapy, it seems promising to use two-step targeting delivery of active agents, or pre-targeting: at the first stage, a non-toxic targeting module (also including antibody or non-immunoglobulin scaffolds) is selectively delivered to a cell of a certain molecular profile, and at the second stage, a cytotoxic agent capable of specifically interacting with the first module is administrated into the organism.

- science news Antigen-specific stimulation and expansion of CAR-T cells using membrane vesicles as target cell surrogates
November 11Development of CAR-T therapy led to immediate success in the treatment of B cell leukemia. Manufacturing of therapy-competent functional CAR-T cells needs robust protocols for ex vivo/in vitro expansion of modified T-cells. In this work, a team of scientists from the IBCh RAS in collaboration with a group of scientists from Dmitry Rogachev National Medical Research Center and colleagues from the Faculty of Biology of Moscow State University have created a new technology for the expansion of CAR T cells using artificial vesicles carrying surface tumor antigens. This approach will allow in the future to obtain CAR T-cells with improved functional properties and to minimize the level of premature «exhaustion» of the CAR T-cell population.
- science news Development of single-domain inhibitory antibodies targeting the ErbB3 receptor for cancer therapy
November 11The human ErbB3 receptor is an important pharmacological target in the treatment of various types of cancer.
 A variety of anti-ErbB3 monoclonal antibodies are currently in development and are classic immunoglobulins. However, the search for new sources of antibodies or nanoantibodies consisting only of the heavy chain is being conducted more and more actively. Thus, in this work, a team of scientists from the Laboratory of Biocatalysis of the IBCh RAS, together with colleagues from the Laboratory of Renewable Energy Sources of the Academic University discovered a group of new single-domain llama antibodies targeting the extracellular domain of ErbB3 using the phage display method. It was found that the single-domain antibodies are not only highly affine for various receptor epitopes, but also have an inhibitory effect on the growth of tumor cells expressing ErbB3.
A variety of anti-ErbB3 monoclonal antibodies are currently in development and are classic immunoglobulins. However, the search for new sources of antibodies or nanoantibodies consisting only of the heavy chain is being conducted more and more actively. Thus, in this work, a team of scientists from the Laboratory of Biocatalysis of the IBCh RAS, together with colleagues from the Laboratory of Renewable Energy Sources of the Academic University discovered a group of new single-domain llama antibodies targeting the extracellular domain of ErbB3 using the phage display method. It was found that the single-domain antibodies are not only highly affine for various receptor epitopes, but also have an inhibitory effect on the growth of tumor cells expressing ErbB3. - science news Engineered Removal of PD-1 From the Surface of CD19 CAR-T Cells Results in Increased Activation and Diminished Survival
November 8CAR-T cell therapy is the most advanced way to treat therapy resistant hematologic cancers, in particular B cell lymphomas and leukemias.
 T cells equipped ex vivo with chimeric receptor recognize target tumor cells and kill them. CAR-T cells that recognize CD19 marker of B cells (CD19 CAR-T) are considered the gold standard of CAR-T therapy and are approved by FDA. But in some cases, CD19 CAR-T cell therapy fails due to immune suppressive microenvironment.
T cells equipped ex vivo with chimeric receptor recognize target tumor cells and kill them. CAR-T cells that recognize CD19 marker of B cells (CD19 CAR-T) are considered the gold standard of CAR-T therapy and are approved by FDA. But in some cases, CD19 CAR-T cell therapy fails due to immune suppressive microenvironment. - science news DARPin_9-29-Targeted Gold Nanorods Selectively Suppress HER2-Positive Tumor Growth in Mice
November 8Breast cancer is one of the most common cancer among women. According to the WHO, in 2020, over 2.2 million cases of this disease were registered worldwide. High level of HER2, a tyrosine kinase receptor, is associated with a more aggressive clinical behavior and poor prognosis for breast cancer patients.
- science news First crystal structure of bacterial oligopeptidase B in an intermediate state: the roles of the hinge region modification and spermine
October 27Oligopeptidase B (OpB) is a two-domain serine peptidase with trypsin-like substrate specificity.
 OpB belongs to the prolyl oligopeptidase (POP) family and are found only in bacteria and protozoa. It is known that OpB are pathogenesis factors of protozoan infections and protect bacterial cells from a number of antimicrobial peptides. Nevertheless, they are the least studied representatives of POP, especially bacterial OpB, for which there was a complete lack of structural information. The researches from IBC RAS together with those of NRC Kurchatov Institute and the IBOC of the National Academy of Sciences of Belarus managed to obtain crystal structures of bacterial OpB from Serratia proteomaculans (PSP) with a modified hinge region.
OpB belongs to the prolyl oligopeptidase (POP) family and are found only in bacteria and protozoa. It is known that OpB are pathogenesis factors of protozoan infections and protect bacterial cells from a number of antimicrobial peptides. Nevertheless, they are the least studied representatives of POP, especially bacterial OpB, for which there was a complete lack of structural information. The researches from IBC RAS together with those of NRC Kurchatov Institute and the IBOC of the National Academy of Sciences of Belarus managed to obtain crystal structures of bacterial OpB from Serratia proteomaculans (PSP) with a modified hinge region. - science news Mambalgin-2 inhibits growth, migration, and invasion of metastatic melanoma cells by targeting the channels containing an asic1a subunit whose up-regulation correlates with poor survival prognosis
October 16Melanoma is aggressive cancer characterized by acidification of extracellular environment. Scientists from the Laboratory of bioengineering of neuromodulators and neuroreceptors IBCh RAS together with colleages from NN Blokhin NMRCO showed for the first time that extracellular media acidification increases proliferation, migration, and invasion of patient-derived metastatic melanoma cells and up-regulates cell-surface expression of acid sensitive channels containing the ASIC1a, α-ENaC, and γ-ENaC subunits.
 No influence of media acidification on these processes was found in normal keratinocytes.
No influence of media acidification on these processes was found in normal keratinocytes. - science news Pseudomonas phage MD8: genetic mosaicism and challenges of taxonomic classification of lambdoid bacteriophages
October 8Fundamental questions of the evolution of viral genomes are the most important topic of virological research. As a result of the joint work of virologists from the Laboratory of Molecular Bioengineering of the IBCh RAS and the Limnological Institute of the RAS, a group of bacteriophages of the dangerous pathogen Pseudomonas was identified, the genomes of these bacterial viruses were studied, and it was shown that their formation was greatly influenced by multiple horizontal transfers, which led to pronounced genetic mosaicism. Scientists also put forward hypotheses about the origin of the new group and proposed basic principles for the taxonomic classification of lambdoid phages.
- science news The secreted protein disulfide isomerase Ag1, lost by ancestors of poorly regenerating vertebrates, is required for Xenopus laevis tail regeneration
October 6As is known, unlike cold-blooded vertebrates, warm-blooded vertebrates are not able to regenerate such complex structures as a limb or tail.
 Earlier, researchers from the Laboratory of Molecular Bases of Embryogenesis IBCH RAS proposed a hypothesis about the relationship between the weakening of regenerative abilities in warm-blooded animals and the loss of some genes that regulate regeneration in cold-blooded animals. In support of this hypothesis, we showed that there are indeed genes essential for the regeneration among the found genes lost by warm-blooded vertebrates, particularly the gene for the secreted disulfide isomerase Ag1. Strong activation of this gene on 1 and 2 days post-amputation of the tail in a model object, the frog Xenopus laevis tadpoles, indicated its essential role at the beginning of regeneration processes. It was shown that knockdown of ag1 reduces the ability to regenerate the amputated tail. At the same time, this ability can be restored either by overexpression of ag1 or by the addition of its recombinant protein to the tadpoles.
Earlier, researchers from the Laboratory of Molecular Bases of Embryogenesis IBCH RAS proposed a hypothesis about the relationship between the weakening of regenerative abilities in warm-blooded animals and the loss of some genes that regulate regeneration in cold-blooded animals. In support of this hypothesis, we showed that there are indeed genes essential for the regeneration among the found genes lost by warm-blooded vertebrates, particularly the gene for the secreted disulfide isomerase Ag1. Strong activation of this gene on 1 and 2 days post-amputation of the tail in a model object, the frog Xenopus laevis tadpoles, indicated its essential role at the beginning of regeneration processes. It was shown that knockdown of ag1 reduces the ability to regenerate the amputated tail. At the same time, this ability can be restored either by overexpression of ag1 or by the addition of its recombinant protein to the tadpoles. - science news Mechanism of coelenterazine chromophore photoinactivation from Beroe abissycola photoprotein is proposed
August 27Scientists from IBCh RAS with colleagues from Photobiology Lab (IBP SB RAS) elucidated the structures of Beroe abissycola photoprotein’s chromophore photoinactivation products and proposed a mechanism of the photoinactivation process.
 The similarity of chemical transformations of photoprotein and GFP-like fluorescent protein chromophores was demonstrated for the first time. The results are published in Organic Letters. The project was funded by the RScF grant № 17-14-01169p.
The similarity of chemical transformations of photoprotein and GFP-like fluorescent protein chromophores was demonstrated for the first time. The results are published in Organic Letters. The project was funded by the RScF grant № 17-14-01169p.
- science news Lipids 2021 Conference
October 11–13 (This event is over)Lipids 2021 Conference, October 11-13, 2021, IBCH RAS, Moscow
- ys-ad Gene therapy 2.0: AAV beyond monogenic gene correction
June 11–23 (This event is over)Founding and managing partner of 4BIO Capital — Dmitry Kuzmin — will make a presentation in conference room 22 June 2022 at 15:00
- science news «Molecular Brain» seminar dedicated to Eugene Grishin’s 75th anniversary
April 27 (This event is over)The open seminar of the Department of Molecular Neurobiology will take place on April 27 at 11:00 in the Great Assembly Hall.
 The program includes presentations by colleagues, students, friends and associates of Eugene Grishin. We cordially invite everyone interested in modern work in the field of toxins, ion channels and neurobiology! Live broadcast in Zoom.
The program includes presentations by colleagues, students, friends and associates of Eugene Grishin. We cordially invite everyone interested in modern work in the field of toxins, ion channels and neurobiology! Live broadcast in Zoom. - conferences International School «Molecular mechanisms of neurodegenerative diseases»
November 26, 2020 (This event is over)Moscow Institute of Physics and Technology (MIPT) invites you to take part in the International school for young scientists «Molecular mechanisms of neurodegenerative diseases», which will be held on November 26, 2020 online.
- science news Lecture by Director-General of the ICGEB Lawrence Banks «Human Papillomaviruses: From Infectious Entry to Malignancy»
January 27, 2020 (This event is over)ICGEB Director-General Group Leader Lawrence Banks will deliver a lecture entitled «Human Papillomaviruses: From Infectious Entry to Malignancy».
Date and time: Mon 27 January 2020 14:00. Location: Small conference hall at 3rd floor BON IBCh.
Location: Small conference hall at 3rd floor BON IBCh. - science news LIGHTS ON: Molecular Imaging of disease dynamics in vivo
September 27 October 11, 2019 (This event is over)Abhijit De PhD
Scientific Officer ‘F’ and Principal Investigator
Head, Molecular Functional Imaging Lab
Advanced Centre of Training Research and Education in Cancer, Tata Memorial Centre, Kharghar, Navi Mumbai, India. - science news Seminar «Molecular Brain»: Anton Maximov
October 8, 2019 (This event is over)The seminar will be held on the 8th of October at 3 pm in the Small lecture hall (3rd floor, BON, IBCh). Everyone is welcome!
- conferences II Joint Life Sciences Forum: VI Russian Congress on Biochemistry and IX Russian Symposium «Proteins and Peptides»
October 1–6, 2019 (This event is over)Dear Colleagues! We are pleasure to invite you to participate the VI Russian Congress on Biochemistry, which will be held in Sochi, Russia (Dagomys Hotel) on October 1-6, 2019.

- science news Lecture by Prof. Yibo Wang «Drug Discovery Targeting Transmembrane Protein-Protein Interactions»
August 26, 2019 (This event is over)Prof. Yibo Wang from the Changchun Institute of Applied Chemistry will deliver a lecture entitled «Drug Discovery Targeting Transmembrane Protein-Protein Interactions». Date and time: Mon 26 August 2019 11:30. Location: Conference hall at 5th floor BON IBCh.
- conferences 12th INTERNATIONAL CONFERENCE “BIOCATALYSIS.FUNDAMENTALS & APPLICATIONS” “BIOCATALYSIS-2019”
June 24–28, 2019 (This event is over)Dear colleagues,
The Lomonosov Moscow State University and RAS institutes, including IBCH RAS, is planning to convene a traditional biannual 12 th International Conference «BIOCATALYSIS-2019» in June, 24–28, 2019. Conference will be convened on board a ship cruising via the route St. Petersburg – Valaam – Kizhi – St. Petersburg.
More info is available at http://bc2019.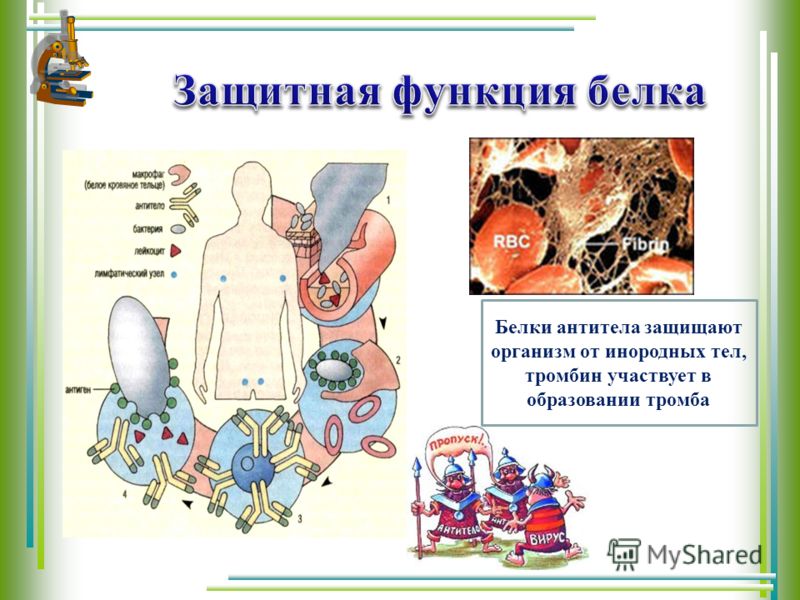 org/.
org/.
Белки острой фазы и маркеры воспаления
Воспаление – это основная защитная реакция организма на внедрение чужеродного агента, введение антигена или физическое повреждение тканей.
Острофазные белки – это белки коагуляции, транспортные белки, которые также выполняют функции медиаторов иммунной системы. Они синтезируются в печени, их концентрация существенно изменяется и зависит от стадии, течения заболевания и массивности повреждения, что и определяет ценность этих тестов для клинической лабораторной диагностики.
В Клинико-диагностической лаборатории производится определение следующих острофазных белков и маркеров воспаления:
- С-реактивный белок (СРБ) – один из самых чувствительных и ранних индикаторов воспаления. Часто используется для диагностики и мониторинга различных воспалительных процессов, дифференциальной диагностики между бактериальной и вирусной инфекциями, обнаружения послеоперационных осложнений, мониторинга эффективности лечебных мероприятий.

- Гаптоглобин – увеличение содержания его в крови отмечается при острых воспалительных процессах, опухолях, нефротическом синдроме.
- Церрулоплазмин – возрастание его уровня наблюдается при острых и хронических инфекциях, циррозе печени, гепатитах, инфаркте миокарда, системных заболеваниях, злокачественных новообразованиях (в 1,5-2 раза).
- Определяются иммуноглобулины классов А, М и G (Ig A, Ig M, Ig G).
Диагностика ревматизма
ASO (антистрептолизин) – является лабораторным критерием ревматизма.Повышение свидетельствует о сенсибилизации организма к стрептококковым антигенам. В период реконвалесценции показатель снижается по сравнению с острым периодом, поэтому может использоваться для наблюдения за динамикой течения, оценки степени активности ревматического процесса.
Ревматоидный фактор – определяется у больных ревматоидным артритом. Также обнаруживается у людей с иными видами аутоиммунной и хронической воспалительной патологией.
% PDF-1.4
%
16 0 объект
>
эндобдж
xref
16 127
0000000016 00000 н.
0000003699 00000 н.
0000004051 00000 н.
0000004231 00000 п.
0000004514 00000 н.
0000004652 00000 п.
0000004751 00000 п.
0000004889 00000 н.
0000005034 00000 н.
0000005180 00000 н.
0000005318 00000 н.
0000005628 00000 н.
0000005939 00000 н.
0000006251 00000 н.
0000006568 00000 н.
0000007039 00000 п.
0000007924 00000 н.
0000008076 00000 н.
0000008398 00000 п.
0000009325 00000 н.
0000010284 00000 п.
0000010383 00000 п.
0000011475 00000 п.
0000011620 00000 п.
0000011767 00000 п.
0000012085 00000 п.
0000012402 00000 п.
0000013835 00000 п.
0000015003 00000 п.
0000016218 00000 п.
0000027900 00000 н.
0000032056 00000 п.
0000033129 00000 п.
0000033312 00000 п.
0000033457 00000 п.
0000033601 00000 п.
0000033796 00000 п.
0000033918 00000 п.
0000042219 00000 п.
0000042413 00000 п.
0000042853 00000 п.
0000043045 00000 п.
0000043206 00000 п.
0000043376 00000 п.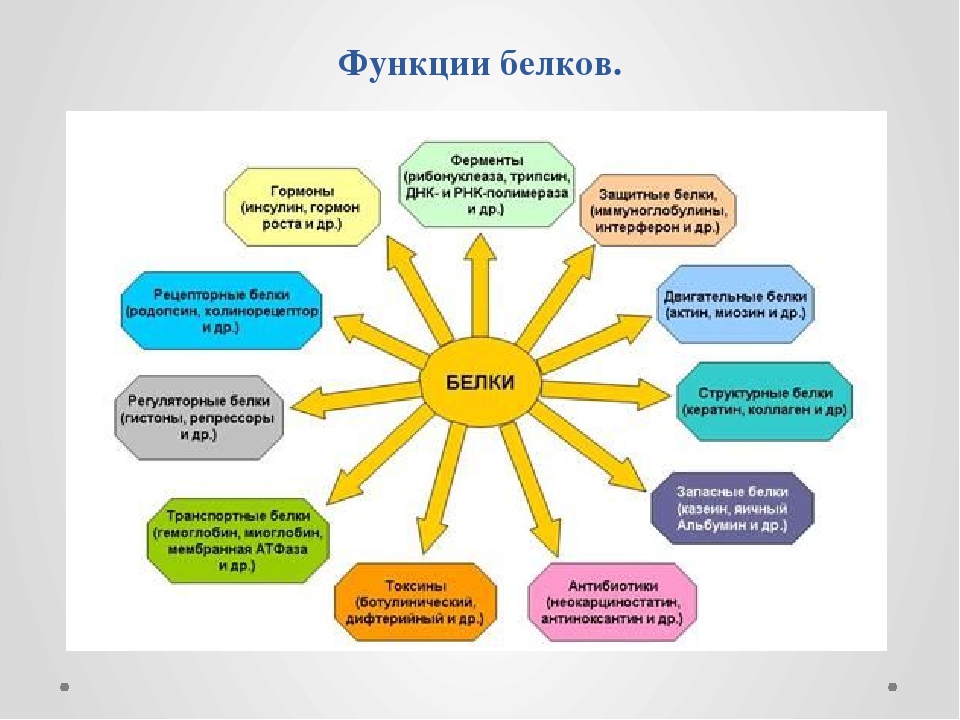
0000043495 00000 п.
0000051100 00000 п.
0000051298 00000 п.
0000051628 00000 п.
0000051826 00000 п.
0000051931 00000 п.
0000054474 00000 п.
0000054653 00000 п.
0000054672 00000 п.
0000054865 00000 п.
0000055178 00000 п.
0000055582 00000 п.
0000055719 00000 п.
0000072092 00000 п.
0000072292 00000 п.
0000072818 00000 п.
0000073018 00000 п.
0000073146 00000 п.
0000073308 00000 п.
0000073880 00000 п.
0000074178 00000 п.
0000075850 00000 п.
0000076053 00000 п.
0000078687 00000 п.
0000078878 00000 п.
0000078912 00000 п.
0000079106 00000 п.
0000079418 00000 п.
0000079572 00000 п.
0000079723 00000 п.
0000080031 00000 н.
0000080053 00000 п.
0000080172 00000 п.
0000080281 00000 п.
0000087924 00000 п.
0000088116 00000 п.
0000088194 00000 п.
0000088386 00000 п.
0000088512 00000 п.
0000088579 00000 п.
0000088746 00000 п.
0000089076 00000 н.
0000089103 00000 п.
0000089235 00000 п.
0000089350 00000 п.
0000094886 00000 п.
0000095080 00000 п.
0000095201 00000 п.
0000095396 00000 п.
0000095537 00000 п.
0000095659 00000 п.
0000095774 00000 п.
0000099669 00000 н.
0000099870 00000 н.
0000100046 00000 н.
0000100248 00000 н.
0000100390 00000 н.
0000100496 00000 н.
0000105433 00000 п.
0000105618 00000 п.
0000105638 00000 п.
0000105839 00000 н.
0000106156 00000 п.
00001
00000 н.
00001
00000 н.
0000191426 00000 н.
0000191609 00000 н.
0000191729 00000 н.
0000191804 00000 н.
0000191923 00000 н.
0000202836 00000 н.
0000203046 00000 н.
0000203370 00000 н.
0000203579 00000 п.
0000203677 00000 н.
0000203783 00000 н.
0000205261 00000 н.
0000205442 00000 н.
0000205462 00000 н.
0000205662 00000 н.
0000205979 00000 н.
0000206160 00000 н.
0000002836 00000 н.
трейлер
] / Назад 254641 >>
startxref
0
%% EOF
142 0 объект
> поток
hb«`f`) a`g`P7cb @
Функции белков в организме — Human Nutrition
Белки являются «рабочими лошадками» тела и участвуют во многих функциях организма.Белки бывают всех размеров и форм, и каждый из них специально структурирован для своей конкретной функции.
Структура и движение
Рисунок 7.9 Структура коллагена
Тройная спираль коллагена от Невита Дилмена / CC BY-SA 3.0
В организме человека было обнаружено более сотни различных структурных белков, но наиболее распространенным является коллаген, который составляет около 6 процентов от общей массы тела. Коллаген составляет 30 процентов костной ткани и включает большое количество сухожилий, связок, хрящей, кожи и мышц.Коллаген — это прочный волокнистый белок, состоящий в основном из глицина и пролина. Внутри его четвертичной структуры три пептидных нити скручиваются друг с другом, как веревка, а затем эти коллагеновые нити перекрываются друг с другом. Эта высокоупорядоченная структура даже прочнее, чем стальные волокна того же размера. Коллаген делает кости крепкими, но гибкими. Волокна коллагена в дерме кожи придают ей структуру, а сопутствующие фибриллы белка эластина делают ее гибкой. Зажмите кожу на руке и отпустите; белки коллагена и эластина в коже позволяют ей вернуться к своей первоначальной форме. Гладкомышечные клетки, которые выделяют белки коллагена и эластина, окружают кровеносные сосуды, придавая им структуру и способность растягиваться назад после того, как через них прокачивается кровь. Еще один сильный волокнистый белок — кератин, из которого состоят кожа, волосы и ногти. Плотно упакованные коллагеновые фибриллы в сухожилиях и связках обеспечивают синхронные механические движения костей и мышц и способность этих тканей возвращаться назад после завершения движения.
Гладкомышечные клетки, которые выделяют белки коллагена и эластина, окружают кровеносные сосуды, придавая им структуру и способность растягиваться назад после того, как через них прокачивается кровь. Еще один сильный волокнистый белок — кератин, из которого состоят кожа, волосы и ногти. Плотно упакованные коллагеновые фибриллы в сухожилиях и связках обеспечивают синхронные механические движения костей и мышц и способность этих тканей возвращаться назад после завершения движения.
Ферменты
Хотя белки в наибольшем количестве содержатся в соединительных тканях, таких как кости, их наиболее необычная функция — это ферменты.Ферменты — это белки, которые проводят определенные химические реакции. Задача фермента — обеспечить место для химической реакции и снизить количество энергии и время, необходимое для того, чтобы эта химическая реакция произошла (это известно как «катализ»). В среднем каждую секунду в клетках происходит более ста химических реакций, и для большинства из них требуются ферменты. Одна только печень содержит более тысячи ферментных систем. Ферменты специфичны и будут использовать только определенные субстраты, которые подходят их активному сайту, подобно тому, как замок может быть открыт только с помощью определенного ключа.Почти каждая химическая реакция требует определенного фермента. К счастью, фермент может снова и снова выполнять свою роль катализатора, хотя в конечном итоге он разрушается и восстанавливается. Все функции организма, включая расщепление питательных веществ в желудке и тонком кишечнике, преобразование питательных веществ в молекулы, которые клетка может использовать, и построение всех макромолекул, включая сам белок, включают ферменты (см. Рисунок 7.10 «Роль ферментов в переваривании углеводов») .
Одна только печень содержит более тысячи ферментных систем. Ферменты специфичны и будут использовать только определенные субстраты, которые подходят их активному сайту, подобно тому, как замок может быть открыт только с помощью определенного ключа.Почти каждая химическая реакция требует определенного фермента. К счастью, фермент может снова и снова выполнять свою роль катализатора, хотя в конечном итоге он разрушается и восстанавливается. Все функции организма, включая расщепление питательных веществ в желудке и тонком кишечнике, преобразование питательных веществ в молекулы, которые клетка может использовать, и построение всех макромолекул, включая сам белок, включают ферменты (см. Рисунок 7.10 «Роль ферментов в переваривании углеводов») .
Рисунок 7.Роль 10 ферментов в переваривании углеводов
Изображение Эллисон Калабрезе / CC BY 4.0
Гормоны
Белки отвечают за синтез гормонов. Гормоны — это химические сообщения, производимые железами внутренней секреции. Когда эндокринная железа стимулируется, она выделяет гормон. Затем гормон транспортируется с кровью к своей клетке-мишени, где он передает сообщение, чтобы инициировать определенную реакцию или клеточный процесс. Например, после еды уровень глюкозы в крови повышается.В ответ на повышение уровня глюкозы в крови поджелудочная железа выделяет гормон инсулин. Инсулин сообщает клеткам организма, что глюкоза доступна и может забирать ее из крови и хранить или использовать для производства энергии или создания макромолекул. Основная функция гормонов — включать и выключать ферменты, поэтому некоторые белки могут даже регулировать действие других белков. Хотя не все гормоны состоят из белков, многие из них таковы.
Когда эндокринная железа стимулируется, она выделяет гормон. Затем гормон транспортируется с кровью к своей клетке-мишени, где он передает сообщение, чтобы инициировать определенную реакцию или клеточный процесс. Например, после еды уровень глюкозы в крови повышается.В ответ на повышение уровня глюкозы в крови поджелудочная железа выделяет гормон инсулин. Инсулин сообщает клеткам организма, что глюкоза доступна и может забирать ее из крови и хранить или использовать для производства энергии или создания макромолекул. Основная функция гормонов — включать и выключать ферменты, поэтому некоторые белки могут даже регулировать действие других белков. Хотя не все гормоны состоят из белков, многие из них таковы.
Жидкостный и кислотно-щелочной баланс
Правильное потребление белка позволяет основным биологическим процессам организма поддерживать статус-кво в изменяющейся окружающей среде.Баланс жидкости относится к поддержанию распределения воды в организме. Если слишком много воды в крови внезапно попадает в ткань, это приводит к отеку и, возможно, к гибели клеток. Вода всегда течет из области с высокой концентрацией в область с низкой концентрацией. В результате вода перемещается в области с более высокими концентрациями других растворенных веществ, таких как белки и глюкоза. Чтобы вода равномерно распределялась между кровью и клетками, белки постоянно циркулируют в крови в высоких концентрациях.Самый распространенный белок в крови — это белок в форме бабочки, известный как альбумин. Присутствие альбумина в крови делает концентрацию белка в крови похожей на таковую в клетках. Таким образом, обмен жидкости между кровью и клетками не является экстремальным, а скорее сводится к минимуму, чтобы сохранить статус-кво.
Вода всегда течет из области с высокой концентрацией в область с низкой концентрацией. В результате вода перемещается в области с более высокими концентрациями других растворенных веществ, таких как белки и глюкоза. Чтобы вода равномерно распределялась между кровью и клетками, белки постоянно циркулируют в крови в высоких концентрациях.Самый распространенный белок в крови — это белок в форме бабочки, известный как альбумин. Присутствие альбумина в крови делает концентрацию белка в крови похожей на таковую в клетках. Таким образом, обмен жидкости между кровью и клетками не является экстремальным, а скорее сводится к минимуму, чтобы сохранить статус-кво.
Рисунок 7.11 Белковый альбумин
PDB 1o9x EBI Джавахара Сваминатана и сотрудников MSD из Европейского института биоинформатики / Public Domain Белок в форме бабочки, альбумин, выполняет множество функций в организме, включая поддержание жидкостного и кислотно-щелочного баланса и транспортировку молекул.
Белок также важен для поддержания правильного баланса pH (мера того, насколько кислым или основным является вещество) в крови. PH крови поддерживается между 7,35 и 7,45, что является слегка щелочным. Даже небольшое изменение pH крови может повлиять на функции организма. Напомним, что кислая среда может вызвать денатурацию белка, что останавливает функционирование белков. В организме есть несколько систем, которые удерживают pH крови в пределах нормы, чтобы этого не происходило. Один из них — это циркулирующий альбумин.Альбумин имеет слабую кислотность и, поскольку он отрицательно заряжен, уравновешивает множество положительно заряженных молекул, таких как протоны (H +), кальций, калий и магний, которые также циркулируют в крови. Альбумин действует как буфер против резких изменений концентраций этих молекул, тем самым уравновешивая pH крови и поддерживая статус-кво. Белок гемоглобин также участвует в кислотно-щелочном балансе, связывая и высвобождая протоны.
PH крови поддерживается между 7,35 и 7,45, что является слегка щелочным. Даже небольшое изменение pH крови может повлиять на функции организма. Напомним, что кислая среда может вызвать денатурацию белка, что останавливает функционирование белков. В организме есть несколько систем, которые удерживают pH крови в пределах нормы, чтобы этого не происходило. Один из них — это циркулирующий альбумин.Альбумин имеет слабую кислотность и, поскольку он отрицательно заряжен, уравновешивает множество положительно заряженных молекул, таких как протоны (H +), кальций, калий и магний, которые также циркулируют в крови. Альбумин действует как буфер против резких изменений концентраций этих молекул, тем самым уравновешивая pH крови и поддерживая статус-кво. Белок гемоглобин также участвует в кислотно-щелочном балансе, связывая и высвобождая протоны.
Транспорт
Альбумин и гемоглобин также играют роль в молекулярном транспорте.Альбумин химически связывается с гормонами, жирными кислотами, некоторыми витаминами, необходимыми минералами и лекарствами и переносит их по кровеносной системе. Каждый эритроцит содержит миллионы молекул гемоглобина, которые связывают кислород в легких и транспортируют его ко всем тканям организма. Плазматическая мембрана клетки обычно не проницаема для больших полярных молекул, поэтому для доставки необходимых питательных веществ и молекул в клетку в клеточной мембране существует множество транспортных белков. Некоторые из этих белков являются каналами, которые позволяют определенным молекулам входить и выходить из клеток.Другие действуют как такси с односторонним движением и требуют энергии для работы.
Каждый эритроцит содержит миллионы молекул гемоглобина, которые связывают кислород в легких и транспортируют его ко всем тканям организма. Плазматическая мембрана клетки обычно не проницаема для больших полярных молекул, поэтому для доставки необходимых питательных веществ и молекул в клетку в клеточной мембране существует множество транспортных белков. Некоторые из этих белков являются каналами, которые позволяют определенным молекулам входить и выходить из клеток.Другие действуют как такси с односторонним движением и требуют энергии для работы.
Защита
Рисунок 7.12 Белки антител
Абаговомаб (моноклональное антитело) по Blake C / CC BY-SA 3.0
Рисунок 7.13 Антигены
Цепи антител от Fred the Oyster / Public Domain
Белок антитела состоит из двух тяжелых цепей и двух легких цепей. Вариабельная область, которая отличается от одного антитела к другому, позволяет антителу распознавать соответствующий ему антиген.
Ранее мы обсуждали, что прочные волокна коллагена в коже обеспечивают ей структуру и поддержку. Плотная сеть коллагеновых волокон кожи также служит преградой для вредных веществ. Функции атаки и разрушения иммунной системы зависят от ферментов и антител, которые также являются белками. Фермент лизоцим выделяется со слюной и атакует стенки бактерий, вызывая их разрыв. Определенные белки, циркулирующие в крови, могут быть направлены на создание молекулярного ножа, который пронзает клеточные мембраны чужеродных захватчиков. Антитела, выделяемые лейкоцитами, исследуют всю систему кровообращения в поисках вредных бактерий и вирусов, которые можно окружить и уничтожить.Антитела также запускают другие факторы иммунной системы для поиска и уничтожения нежелательных злоумышленников.
Плотная сеть коллагеновых волокон кожи также служит преградой для вредных веществ. Функции атаки и разрушения иммунной системы зависят от ферментов и антител, которые также являются белками. Фермент лизоцим выделяется со слюной и атакует стенки бактерий, вызывая их разрыв. Определенные белки, циркулирующие в крови, могут быть направлены на создание молекулярного ножа, который пронзает клеточные мембраны чужеродных захватчиков. Антитела, выделяемые лейкоцитами, исследуют всю систему кровообращения в поисках вредных бактерий и вирусов, которые можно окружить и уничтожить.Антитела также запускают другие факторы иммунной системы для поиска и уничтожения нежелательных злоумышленников.
Заживление ран и регенерация тканей
Белки участвуют во всех аспектах заживления ран, процесса, который проходит в трех фазах: воспалительной, пролиферативной и ремоделирующей. Например, если вы шили и укололи палец иглой, ваша плоть покраснела бы и воспалилась. Через несколько секунд кровотечение прекратится.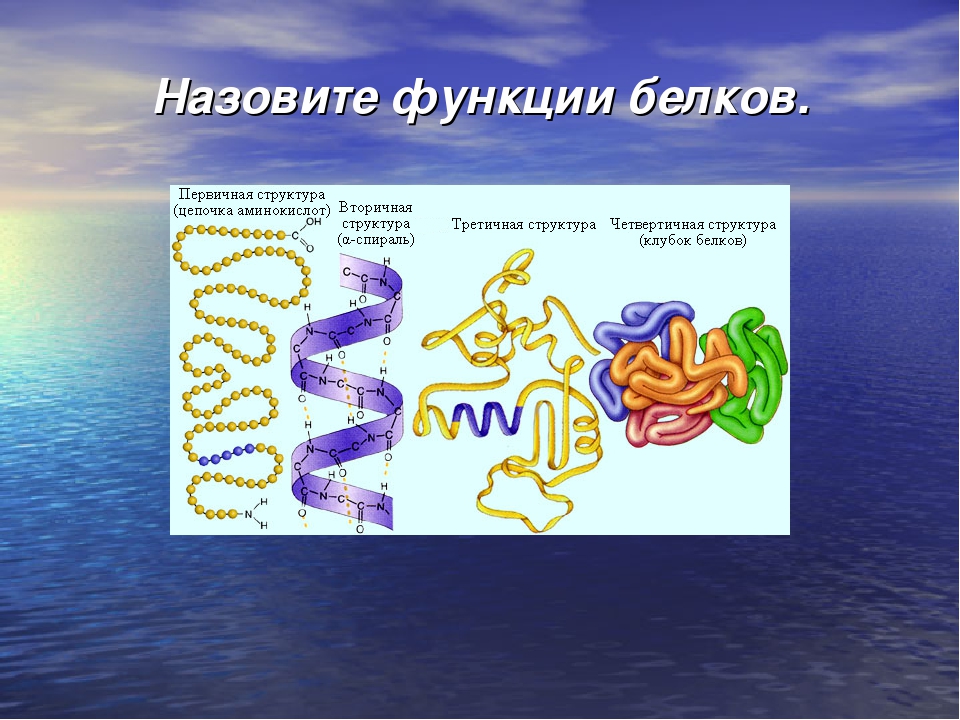 Процесс заживления начинается с белков, таких как брадикинин, которые расширяют кровеносные сосуды в месте повреждения.Дополнительный белок, называемый фибрином, помогает защитить тромбоциты, которые образуют сгусток, чтобы остановить кровотечение. Затем, в фазе пролиферации, клетки перемещаются и восстанавливают поврежденную ткань, устанавливая новые волокна коллагена. Волокна коллагена помогают сблизить края раны. В фазе ремоделирования откладывается больше коллагена, образуя рубец. Рубцовая ткань только на 80 процентов функциональна, чем нормальная неповрежденная ткань. Если в диете недостаточно белка, процесс заживления ран заметно замедляется.
Процесс заживления начинается с белков, таких как брадикинин, которые расширяют кровеносные сосуды в месте повреждения.Дополнительный белок, называемый фибрином, помогает защитить тромбоциты, которые образуют сгусток, чтобы остановить кровотечение. Затем, в фазе пролиферации, клетки перемещаются и восстанавливают поврежденную ткань, устанавливая новые волокна коллагена. Волокна коллагена помогают сблизить края раны. В фазе ремоделирования откладывается больше коллагена, образуя рубец. Рубцовая ткань только на 80 процентов функциональна, чем нормальная неповрежденная ткань. Если в диете недостаточно белка, процесс заживления ран заметно замедляется.
В то время как заживление ран происходит только после получения травмы, в организме продолжается другой процесс, называемый регенерацией тканей. Основное различие между заживлением ран и регенерацией тканей заключается в процессе регенерации точной структурной и функциональной копии утраченной ткани. Таким образом, старая умирающая ткань заменяется не рубцовой тканью, а совершенно новой, полностью функциональной тканью. Некоторые клетки (например, клетки кожи, волос, ногтей и кишечника) имеют очень высокую скорость регенерации, в то время как другие (например, клетки сердечной мышцы и нервные клетки) не регенерируют на каких-либо заметных уровнях.Регенерация тканей — это создание новых клеток (деление клеток), для чего требуется множество различных белков, включая ферменты, синтезирующие РНК и белки, транспортные белки, гормоны и коллаген. В волосяном фолликуле клетки делятся, и волосы растут в длину. Рост волос в среднем составляет 1 сантиметр в месяц, а ногтей — около 1 сантиметра каждые сто дней. Клетки, выстилающие кишечник, восстанавливаются каждые три-пять дней. Неадекватные белковые диеты ухудшают регенерацию тканей, вызывая множество проблем со здоровьем, включая нарушение переваривания и усвоения питательных веществ и, что наиболее заметно, роста волос и ногтей.
Некоторые клетки (например, клетки кожи, волос, ногтей и кишечника) имеют очень высокую скорость регенерации, в то время как другие (например, клетки сердечной мышцы и нервные клетки) не регенерируют на каких-либо заметных уровнях.Регенерация тканей — это создание новых клеток (деление клеток), для чего требуется множество различных белков, включая ферменты, синтезирующие РНК и белки, транспортные белки, гормоны и коллаген. В волосяном фолликуле клетки делятся, и волосы растут в длину. Рост волос в среднем составляет 1 сантиметр в месяц, а ногтей — около 1 сантиметра каждые сто дней. Клетки, выстилающие кишечник, восстанавливаются каждые три-пять дней. Неадекватные белковые диеты ухудшают регенерацию тканей, вызывая множество проблем со здоровьем, включая нарушение переваривания и усвоения питательных веществ и, что наиболее заметно, роста волос и ногтей.
Производство энергии
Некоторые аминокислоты в белках можно разобрать и использовать для производства энергии (Рисунок 7. 14 «Аминокислоты, используемые для получения энергии»). Только около 10 процентов пищевых белков катаболизируются каждый день для производства клеточной энергии. Печень способна расщеплять аминокислоты до углеродного скелета, которые затем могут быть включены в цикл лимонной кислоты. Это похоже на то, как глюкоза используется для производства АТФ. Если диета человека не содержит достаточного количества углеводов и жиров, его организм будет использовать больше аминокислот для производства энергии, что ставит под угрозу синтез новых белков и разрушает мышечные белки.Кроме того, если в рационе человека содержится больше белка, чем необходимо организму, лишние аминокислоты расщепляются и превращаются в жир.
14 «Аминокислоты, используемые для получения энергии»). Только около 10 процентов пищевых белков катаболизируются каждый день для производства клеточной энергии. Печень способна расщеплять аминокислоты до углеродного скелета, которые затем могут быть включены в цикл лимонной кислоты. Это похоже на то, как глюкоза используется для производства АТФ. Если диета человека не содержит достаточного количества углеводов и жиров, его организм будет использовать больше аминокислот для производства энергии, что ставит под угрозу синтез новых белков и разрушает мышечные белки.Кроме того, если в рационе человека содержится больше белка, чем необходимо организму, лишние аминокислоты расщепляются и превращаются в жир.
Рисунок 7.14 Аминокислоты, используемые для получения энергии
Изображение Эллисон Калабрезе / CC BY 4.0
Как работают G-белки
Как работают G-белки
Во время летнего похода по Каскадам вы прогуливаетесь, как вдруг
за поворотом вы видите огромного медведя гризли. Ваше тело переходит в состояние красной тревоги.
Ваше тело переходит в состояние красной тревоги.
Адреналин мгновенно накачивается по всей вашей кровеносной системе, что побуждает вас
печень и мышцы для высвобождения глюкозы, производящей энергию.Ваше сердце бьется
быстро, ваши легкие дышат быстрее, и вы готовы бежать так же быстро, как вы
когда-либо за всю свою жизнь.
В данный момент вам может не быть интересно, как все это работает внутри
ваши клетки, но это был вопрос для ученых, так как гормоны были первыми
изолированные. В начале этого века ученые знали, что гормон,
таких как адреналин, будет сигнализировать ответ в клетке — «высвободить глюкозу в
кровь «. Но как именно это сообщение было передано через клеточную стенку
было непонятно — тогда думали, сработало в один оборот, как замок и
ключ.
Но работа Мартина Родбелла, получившая Нобелевскую премию, доказала, что есть много
больше происходит — необходимо перебросить ряд переключателей химическим способом, чтобы
сообщение, которое нужно передать. Ключ к этим химическим переключателям — G
белки.
И если с этим процессом что-то не так, например, оставить переключатель во включенном состоянии или
«выключено» слишком долго, есть проблемы. Ученые теперь знают, что холера меняет
белки G, оставляя переключатель «включенным» слишком долго, что препятствует нормальному
всасывание соли и воды в кишечнике — что приводит к обезвоживанию или даже
смерть.
Считается, что диабет и алкоголизм частично вызваны G-белком.
неисправности, и ученые также проследили форму рака гипофиза до
нарушенные G-белки. Пока исследования G-протеина еще не привели к чуду
вылечить, открытие может привести к возможному лечению.
Сама система представляет собой сложную серию реакций. Когда вы видите медведя в
в лесу надпочечники выделяют адреналин. Гормональные скачки повсюду
ваше тело, но только клетки, которым нужен сигнал, например сердечная мышца,
есть рецепторы, которые могут его обнаружить.Молекула гормона «щекочет одну из
рецептор за пределами клеточной стенки, а внутри стены, другой конец хихикает, »
— объясняет Константин Лондос, коллега Родбелла из NIH.
«Хихикающая» сторона рецептора меняет форму, превращаясь в
непреодолимая стыковка G-белка. Как только G
сам белок претерпевает изменение. Он держался за молекулу под названием
ВВП. Теперь он высвобождает это и захватывает родственную молекулу, называемую GTP.
Но это еще не конец трансформации.С GTP в руках G
белок теперь работает с другими мембранными процессами, такими как ферменты и ионные
каналы. Эти ферменты теперь могут создавать новую молекулу-мессенджер, называемую циклической.
AMP. Далее следует «ферментативный каскад», в котором циклический АМФ вызывает
белковая реакция. Если ваша печень и мышцы реагируют на взгляд
гризли, ферменты высвобождают глюкозу из клеток, давая вам
энергия убежать.
Роль, которую G-белки играют в процессе коммуникации, происходит мгновенно.В качестве
вы поворачиваетесь и бежите, все, что вы можете надеяться, это то, что G-белки гризли не
работает так же, как и ваш собственный. — Том Гриффин
Вернуться на начальную страницу Родбелла
Отправьте письмо редактору на columns@u. washington.edu.
.
Количественный анализ белков Р тельца дрожжей показывает принципы состава и специфичности
[Примечание редакции: далее следует план авторов по внесению исправлений.]
Основные опасения рецензентов можно резюмировать следующим образом:
1.Новизна работы не подчеркнута. Ясно, что он более всеобъемлющий, чем предыдущие работы этих авторов, но требует более перспективного и дополнительного тестирования.
Что касается новизны, мы считаем, что наша работа добавляет важные новые знания о Р-телах и биомолекулярных конденсатах в целом. Во-первых, наша работа — это единственный отчет об абсолютных или относительных концентрациях основных белков в любом конденсате. Такие концентрации являются важными знаниями при рассмотрении природы и биохимических функций тел фосфора (и, в более широком смысле, других конденсатов).Например, модели конденсатов как отсеков хранения подвергаются сомнению нашими данными на рисунке 3, показывающими, что только небольшая часть большинства белков присутствует в P-тельцах. Кроме того, модели конденсатов как участков высокой биохимической активности должны учитывать тот факт, что большинство неосновных компонентов сконцентрированы только <10 раз относительно окружающей цитоплазмы и что большинство молекул все еще находится в цитоплазме. Таким образом, конденсатные функции с большей вероятностью возникают из основных компонентов или из совокупностей непрофильных компонентов (например,грамм. в каскаде реакций). Это важные концепции для данной области. Во-вторых, наши данные показывают, что лишь небольшая часть многих белков в P-телескопах имеет высокую концентрацию в структуре. Эта информация является ключевой для понимания строения конденсатов. Это говорит против большинства преобладающих моделей состава конденсата, которые рассматривают отсеки как трудноразрешимо сложные, с сотнями компонентов, все из которых неявно имеют одинаковый вес с точки зрения функциональной значимости. Наши данные показывают, что компартменты намного проще и предоставляют первые пути к надежному биомиметическому восстановлению.
Кроме того, модели конденсатов как участков высокой биохимической активности должны учитывать тот факт, что большинство неосновных компонентов сконцентрированы только <10 раз относительно окружающей цитоплазмы и что большинство молекул все еще находится в цитоплазме. Таким образом, конденсатные функции с большей вероятностью возникают из основных компонентов или из совокупностей непрофильных компонентов (например,грамм. в каскаде реакций). Это важные концепции для данной области. Во-вторых, наши данные показывают, что лишь небольшая часть многих белков в P-телескопах имеет высокую концентрацию в структуре. Эта информация является ключевой для понимания строения конденсатов. Это говорит против большинства преобладающих моделей состава конденсата, которые рассматривают отсеки как трудноразрешимо сложные, с сотнями компонентов, все из которых неявно имеют одинаковый вес с точки зрения функциональной значимости. Наши данные показывают, что компартменты намного проще и предоставляют первые пути к надежному биомиметическому восстановлению.
Наконец, хотя нам предстоит еще поработать, чтобы понять корреляции на Рисунке 7, как эмпирический предиктор степени, в которой полноразмерные белки могут быть сконцентрированы в конденсате, они весьма ценны для данной области.
2. Авторам необходимо рассмотреть другие модели для сборки, а не только клиент / скаффолд. Расхождения между данными и их моделью не устранены. Есть несколько подробных экспериментов, которые могли бы сделать больше, чтобы прояснить туманные выводы (см. Ниже).Например, вместо мечения одним флуорофором они могут лучше оценить состав и вариацию с помощью попарного анализа двух флуорофоров.
3. Могут ли они предоставить какие-либо дополнительные данные или идеи, чтобы пролить свет на несоответствие, которое они указывают между их моделью и результатами для DCP2?
Обеспокоенность по поводу модели каркаса / клиента, по-видимому, частично возникает из-за неправильного представления рецензентов о самой модели и о нашем намерении представить корреляции валентностей на рисунке 7. Оглядываясь назад, мы понимаем, как возникли эти неправильные представления, и полагаем, что обширный пересмотр текста в сочетании с дополнительными данными должен привести к гораздо более четкому представлению и более точной модели. Короче говоря, мы не намеревались изобразить модель, в которой валентность взаимодействий является единственным параметром, важным для определения состава конденсата, и где РНК не важна. Скорее, как предполагают составители обзора, мы полагаем, что A) РНК является ключевым каркасом для P-телец, B) важно сродство связывания белков друг с другом и с РНК и C) важна валентность взаимодействий.Мы также ошибочно отождествили «основные» белки с «каркасными» белками, а «неосновные» — с «клиентом» в предложении в начале обсуждения, цитируемом вторым рецензентом, вместо того, чтобы описать их как коррелированные, но не идентичные, что было наше намерение до конца обсуждения. Это вызвало ряд проблем, которые можно решить в пересмотренном введении и обсуждении.
Оглядываясь назад, мы понимаем, как возникли эти неправильные представления, и полагаем, что обширный пересмотр текста в сочетании с дополнительными данными должен привести к гораздо более четкому представлению и более точной модели. Короче говоря, мы не намеревались изобразить модель, в которой валентность взаимодействий является единственным параметром, важным для определения состава конденсата, и где РНК не важна. Скорее, как предполагают составители обзора, мы полагаем, что A) РНК является ключевым каркасом для P-телец, B) важно сродство связывания белков друг с другом и с РНК и C) важна валентность взаимодействий.Мы также ошибочно отождествили «основные» белки с «каркасными» белками, а «неосновные» — с «клиентом» в предложении в начале обсуждения, цитируемом вторым рецензентом, вместо того, чтобы описать их как коррелированные, но не идентичные, что было наше намерение до конца обсуждения. Это вызвало ряд проблем, которые можно решить в пересмотренном введении и обсуждении.
Соответственно, мы не предполагали, что данные на Рисунке 7 представляют тест или валидацию основанной на валентности модели каркаса / клиента или результата, вытекающего из предыдущих данных в рукописи. Скорее, мы задумали это просто как эмпирическую корреляцию, которая потенциально может быть полезна при прогнозировании других конденсатов, где количественные данные визуализации получить нелегко. Очевидно, это не было правильно, и в нашей презентации модель была основана исключительно на валентности, что не входило в наши намерения. Это можно исправить в доработке. Тем не менее, мы согласны с авторами обзора в том, что для нас важно понять, почему мутанты Dcp2 не попадают в ту же корреляцию, что и различные полноразмерные белки, и разработали дополнительный набор мутантов, чтобы лучше это понять.
Скорее, мы задумали это просто как эмпирическую корреляцию, которая потенциально может быть полезна при прогнозировании других конденсатов, где количественные данные визуализации получить нелегко. Очевидно, это не было правильно, и в нашей презентации модель была основана исключительно на валентности, что не входило в наши намерения. Это можно исправить в доработке. Тем не менее, мы согласны с авторами обзора в том, что для нас важно понять, почему мутанты Dcp2 не попадают в ту же корреляцию, что и различные полноразмерные белки, и разработали дополнительный набор мутантов, чтобы лучше это понять.
Второй рецензент также обращает внимание на то, что стехиометрия ~ 1: 1 многих основных белков предполагает, что они могут формировать стереотипную сборку, которая связывает РНК, а затем рекрутирует другие белки, что приводит к несколько иной модели состава. У нас есть несколько мыслей по этому поводу. Во-первых, мы согласны с тем, что относительная стехиометрия поразительна. Фактически, в более ранней версии рукописи это обсуждалось довольно подробно, как и в комментариях рецензента. Тем не менее, мы в конечном итоге удалили это обсуждение, потому что A) относительная стехиометрия отличается у штаммов дикого типа, лишенных глюкозы, и B) широкий диапазон концентраций P-тел каждого компонента и отсутствие многокомпонентных корреляций в отдельных дрожжевых клетках сделали это сложно претендовать на конкретную сборку. Тем не менее, мы согласны с авторами обзора в том, что наше понимание состава тела P было бы значительно улучшено путем сбора данных с двумя цветами, где мы могли бы количественно определить абсолютные концентрации 2 белков одновременно в отдельных клетках.Хотя непрактично собирать такие данные обо всех возможных парах белков P-тел, мы создадим штаммы для изучения ключевых корреляций внутри основной группы и между стержневыми и неосновными белками. Эти данные позволят нам различать разные модели состава тела Р.
Тем не менее, мы в конечном итоге удалили это обсуждение, потому что A) относительная стехиометрия отличается у штаммов дикого типа, лишенных глюкозы, и B) широкий диапазон концентраций P-тел каждого компонента и отсутствие многокомпонентных корреляций в отдельных дрожжевых клетках сделали это сложно претендовать на конкретную сборку. Тем не менее, мы согласны с авторами обзора в том, что наше понимание состава тела P было бы значительно улучшено путем сбора данных с двумя цветами, где мы могли бы количественно определить абсолютные концентрации 2 белков одновременно в отдельных клетках.Хотя непрактично собирать такие данные обо всех возможных парах белков P-тел, мы создадим штаммы для изучения ключевых корреляций внутри основной группы и между стержневыми и неосновными белками. Эти данные позволят нам различать разные модели состава тела Р.
[Примечание редакции: авторы отправили на повторное рассмотрение после принятия решения после экспертной оценки. Далее следует письмо о решении после первого раунда проверки. ]
]
Рецензенты считают, что они не могут поддержать повторное представление этой работы.Причины этого лучше всего резюмирует один из рецензентов, который утверждает:
«Я не сторонник повторного представления. Если я что-то не понял неправильно, авторы не планируют решать проблемы, связанные со значимостью и достоверностью их модели.
Авторы заявляют, что: «Как предполагают обозреватели, мы считаем, что A) РНК является ключевым каркасом для P-телец, B) важно сродство связывания белков друг с другом и с РНК и C) важна валентность взаимодействий» .
Но не предлагайте никаких экспериментов / данных, подтверждающих эти точки.
Приносим извинения за то, что в нашем первоначальном ответе не было достаточно четко заявлено, что мы намеревались серьезно пересмотреть нашу модель, лучше проиллюстрировать ее значение и проверить ее дальше. Все это мы сделали в ревизии.
В качестве краткого резюме здесь мы больше не представляем работу как тестирование модели строительных лесов / клиента для сборки конденсата.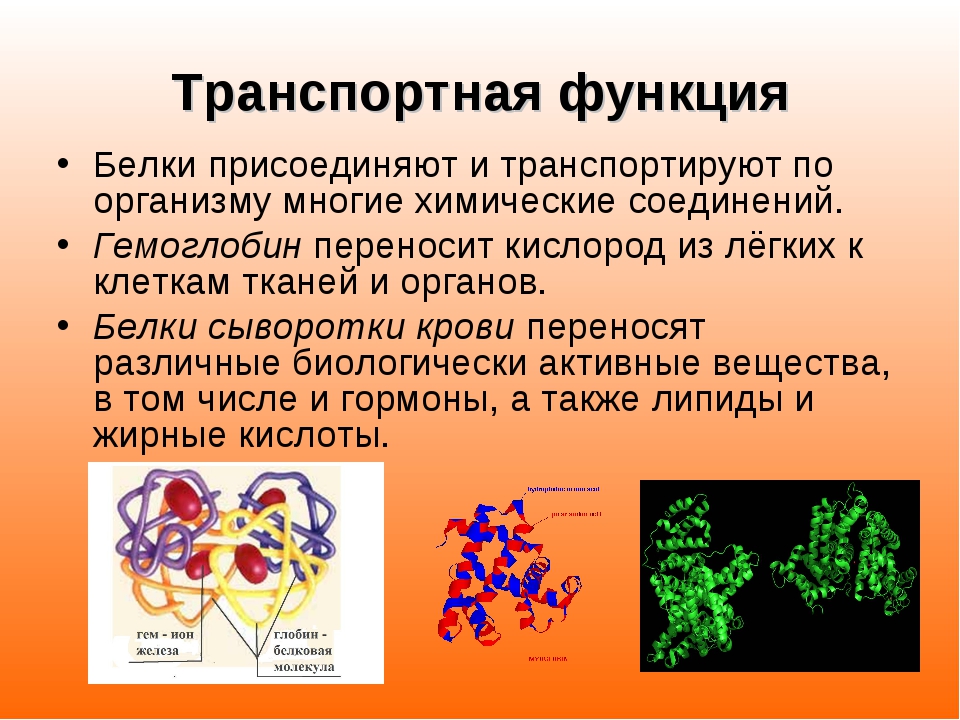 Скорее, теперь мы предлагаем гораздо более детальный взгляд на сборку P-телец, подчеркивая роль высокой связности (связанной с валентностью, но отличной от нее) между белками и РНК в производстве конденсата, но также обсуждая важность аффинности связывания и активности процессы.Мы также обсуждаем несколько других моделей и объясняем, как они несовместимы с данными наших и других лабораторий как в этой статье, так и в литературе. Как было предложено рецензентами и обсуждено с редактором в апреле, теперь мы получили двухцветные данные (изучение корреляции между двумя компонентами Р-тельцов) и данные, исследующие роль высокоаффинных связывающих элементов в управлении рекрутированием Р-тельцов, которые оба оказались полезными при сравнении разных моделей.
Скорее, теперь мы предлагаем гораздо более детальный взгляд на сборку P-телец, подчеркивая роль высокой связности (связанной с валентностью, но отличной от нее) между белками и РНК в производстве конденсата, но также обсуждая важность аффинности связывания и активности процессы.Мы также обсуждаем несколько других моделей и объясняем, как они несовместимы с данными наших и других лабораторий как в этой статье, так и в литературе. Как было предложено рецензентами и обсуждено с редактором в апреле, теперь мы получили двухцветные данные (изучение корреляции между двумя компонентами Р-тельцов) и данные, исследующие роль высокоаффинных связывающих элементов в управлении рекрутированием Р-тельцов, которые оба оказались полезными при сравнении разных моделей.
Наконец, теперь мы гораздо более подробно описываем новизну наших открытий, которые включают, среди 5 пунктов, подробно описанных в сопроводительном документе: A) открытие, что, несмотря на большое количество молекул, присутствующих в P-телах, только небольшое количество (7) сильно сконцентрированы там с большими коэффициентами разделения и B) открытие, что межмолекулярная связь играет ключевую роль в управлении концентрациями молекул в P-телах.
Они также заявляют, что: «Мы не намеревались, чтобы данные на Рисунке 7 представляли тест или валидацию основанной на валентности модели каркаса / клиента или результата, вытекающего из предыдущих данных в рукописи. Скорее, мы намеревались это просто как эмпирическая корреляция, которая потенциально может быть полезна при прогнозировании других конденсатов, где количественные данные визуализации получить нелегко ».
Это не похоже на заголовок статьи, обещающей «композиционные принципы».Их предложение изобразить два компонента одновременно, не добавит ничего, кроме подтверждения того, что они уже показали?
Мы удалили количественные корреляции между валентностью и физическими характеристиками P-тел (исходный рисунок 7), что было проблематично по ряду причин. Наши двухцветные эксперименты предоставили важные проверки потенциальных моделей образования конденсата. Кроме того, как указано выше, мы значительно пересмотрели нашу модель.
Мы полагаем, что в рукописи изложены несколько принципов, касающихся состава P-тел.Они описаны в пересмотренном разделе «Обсуждение» статьи, который полностью переписан. К ним относятся: A) концепция (подтвержденная доказательствами) о том, что биологические конденсаты, которые кажутся довольно сложными при качественном анализе их компонентов, на самом деле могут иметь гораздо более простую первичную сложность и организацию при количественном рассмотрении; Б) идея о том, что различия между высококонцентрированными и слабо концентрированными белками Р-тельцов, вероятно, тесно связаны с их паттернами межмолекулярной связи внутри Р-тельцов; C) идея о том, что взаимодействия, которые производят конденсаты, распределяются по их высоко валентным компонентам, они действуют с разной степенью кооперативности, способствуя формированию более крупной сборки; и D) предсказание того, что термодинамика образования конденсата и состав результирующей структуры должны быть связаны, на основе моделей связности в сети взаимодействия; я.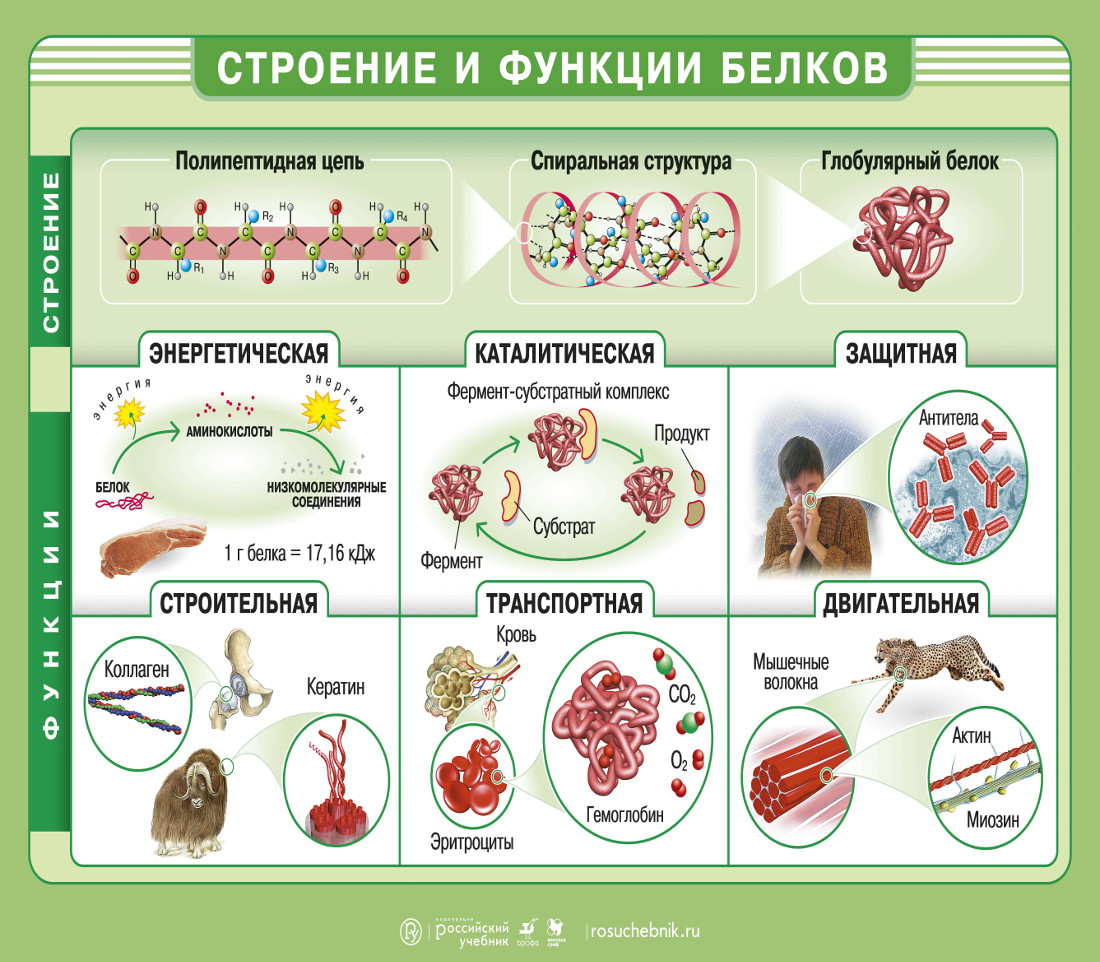 е. Удаление сильно связанной молекулы должно влиять как на концентрации других факторов, необходимых для образования конденсата, так и на относительные концентрации компонентов в конденсате.
е. Удаление сильно связанной молекулы должно влиять как на концентрации других факторов, необходимых для образования конденсата, так и на относительные концентрации компонентов в конденсате.
Наконец, утверждение о том, что «наши данные показывают, что лишь небольшая часть многих белков в Р-тельцах сильно сконцентрирована в структуре. Эта информация является ключевой для концептуализации строения конденсатов. Это говорит против большинства преобладающих моделей состава конденсата, которые Отобразить отсеки как непреодолимо сложные, с сотнями компонентов, все неявно взвешенные в равной степени с точки зрения функциональной значимости.Наши данные предполагают, что компартменты намного проще и предоставляют первые пути к надежному биомиметическому восстановлению «.
Биомиметическое восстановление гранул РНК уже началось, и уже было показано, что нескольких компонентов может быть достаточно для имитации структуры конденсата (Feric et al., 2016, Putnam et al.
, 2019). Утверждение также, кажется, подразумевает, что знание концентрации каждого фактора поможет идентифицировать такие ключевые компоненты, концепция, которая остается непроверенной ».
Хотя работа, процитированная Feric и Putnam, элегантна, обе используют только два компонента в ядрышках и гранулах P соответственно.Таким образом, степень, в которой они захватывают свойства клеточных структур, неизвестна. Более того, насколько нам известно, не было установлено, что компоненты, используемые в этих реконструкциях, в количественном отношении являются доминирующими компонентами клеточных структур или что другие компоненты одинаково сконцентрированы. Таким образом, опять же, степень, в которой эти воссозданные структуры воспроизводят биологические структуры, является неопределенной. На наш взгляд, важным достижением в биохимическом восстановлении конденсатов, которые очень похожи на клеточные конденсаты, является знание доминирующего (т.е. наиболее концентрированные) компоненты и их динамические свойства in vivo. Обладая этой информацией, можно комбинировать соответствующие молекулы in vitro при их общих клеточных концентрациях и узнать, образуют ли они конденсаты соответствующих концентраций компонентов (как относительных, так и абсолютных) и динамики. Если они соответствуют этим критериям для нескольких компонентов, то можно быть уверенным в том, что биохимия действительно является биомиметической. Без этой предварительной информации человек делает обоснованное предположение о том, какие белки следует комбинировать биохимически и насколько хорошо воссозданная структура имитирует клеточную структуру.
Обладая этой информацией, можно комбинировать соответствующие молекулы in vitro при их общих клеточных концентрациях и узнать, образуют ли они конденсаты соответствующих концентраций компонентов (как относительных, так и абсолютных) и динамики. Если они соответствуют этим критериям для нескольких компонентов, то можно быть уверенным в том, что биохимия действительно является биомиметической. Без этой предварительной информации человек делает обоснованное предположение о том, какие белки следует комбинировать биохимически и насколько хорошо воссозданная структура имитирует клеточную структуру.
Резюме рецензента: В этой рукописи сообщается о количественном анализе состава тела P в дрожжах. Авторы характеризуют концентрацию, обогащение относительно цитоплазматического пула и динамику 31 зарегистрированного белка P-тельца, меченного GFP. Они идентифицируют 19 белков, которые были достаточно сконцентрированы в P-тельцах в тестируемых условиях, и делят их на два класса: «основная группа», состоящая из 7 белков с высоким обогащением и низкой динамикой, а остальные 12 белков с низким обогащением и высокой динамикой.
Авторы утверждают, что эти наблюдения поддерживают модель каркаса / клиента для сборки конденсата, где количество партнеров по взаимодействию для данного белка (валентность) может быть использовано для прогнозирования его обогащения конденсатами / P тел.
Мы тщательно пересмотрели текст и больше не представляем работу как тестирование модели строительных лесов / клиента для сборки конденсата. Фактически, на основе наших данных мы теперь представляем новый, более детальный взгляд на номенклатуру строительных лесов / клиентов, который лучше отражает поведение природных конденсатов и будет более полезным для полевых работ.Мы также удалили анализы, описывающие количественные отношения между обогащением тел P и валентностью взаимодействий. Однако мы утверждаем, что существует значимая качественная взаимосвязь между связностью в сети взаимодействия P-тел и молекулярным поведением, с множеством оговорок, которые теперь подробно описаны в разделе «Обсуждение». Подробнее о каждом из этих вопросов мы расскажем ниже.
Основные опасения рецензентов можно резюмировать следующим образом:
1) Новизна работы не подчеркнута.Ясно, что он более всеобъемлющий, чем предыдущие работы этих авторов, но требует более перспективного и дополнительного тестирования.
Мы тщательно пересмотрели обсуждение, чтобы выделить новые принципы, которые, по нашему мнению, вытекают из наших данных. Во-первых, наша работа — это первый всесторонний количественный анализ состава и динамики любого природного биомолекулярного конденсата. Во-вторых, это количественное определение показало, что, несмотря на большое количество молекул, присутствующих в P-телах, только небольшое количество (7) сильно сконцентрировано там с большими коэффициентами разделения.Это значительно упрощает процесс формирования, регуляции и функции P-телец. В-третьих, количественный анализ показал, что только небольшая часть большинства белков присутствует в P-тельцах, что имеет важное значение для функций конденсатов.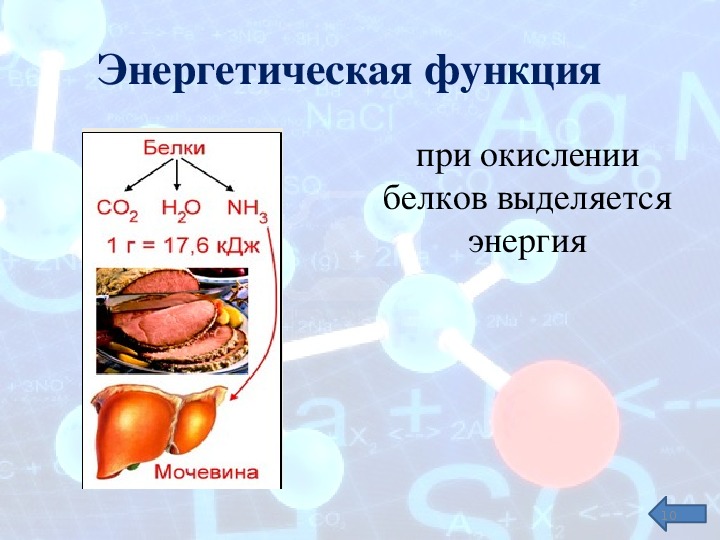 В-четвертых, большинство (6 из 7) высококонцентрированных белков / комплексов сильно связаны в сети взаимодействия P-тельцов, и ни один из слабо концентрированных белков сильно связан. Эти корреляции предполагают, что связность играет важную роль в управлении составом структур.Однако важно отметить, что другие факторы, включая сродство связывания и активные процессы, также вносят вклад в молекулярное поведение, и теперь мы объясним такие сложности в нашем обсуждении. Наконец, основываясь на наших данных и других данных в литературе, мы пересмотрели понятие каркасов и клиентов в биомолекулярных конденсатах. Мы утверждаем, что эти термины не следует использовать для классификации молекул на бинарные группы, как мы делали раньше, а скорее как качественные дескрипторы степени, в которой молекула способствует образованию и составу конденсата.Таким образом, молекула должна быть описана как более подобная каркасу или более подобная клиенту, в зависимости от того, имеет ли она больший или меньший эффект, соответственно.
В-четвертых, большинство (6 из 7) высококонцентрированных белков / комплексов сильно связаны в сети взаимодействия P-тельцов, и ни один из слабо концентрированных белков сильно связан. Эти корреляции предполагают, что связность играет важную роль в управлении составом структур.Однако важно отметить, что другие факторы, включая сродство связывания и активные процессы, также вносят вклад в молекулярное поведение, и теперь мы объясним такие сложности в нашем обсуждении. Наконец, основываясь на наших данных и других данных в литературе, мы пересмотрели понятие каркасов и клиентов в биомолекулярных конденсатах. Мы утверждаем, что эти термины не следует использовать для классификации молекул на бинарные группы, как мы делали раньше, а скорее как качественные дескрипторы степени, в которой молекула способствует образованию и составу конденсата.Таким образом, молекула должна быть описана как более подобная каркасу или более подобная клиенту, в зависимости от того, имеет ли она больший или меньший эффект, соответственно. Такое использование терминов лучше отражает экспериментальные данные, но при этом сохраняет идею о том, что одни молекулы вносят больший вклад, чем другие, в образование и состав конденсата.
Такое использование терминов лучше отражает экспериментальные данные, но при этом сохраняет идею о том, что одни молекулы вносят больший вклад, чем другие, в образование и состав конденсата.
2) Авторам необходимо рассмотреть другие модели для сборки, а не только клиент / скаффолд. Расхождения между данными и их моделью не устранены.Есть несколько подробных экспериментов, которые могли бы сделать больше, чтобы прояснить туманные выводы (см. Ниже). Например, вместо мечения одним флуорофором они могут лучше оценить состав и вариацию с помощью попарного анализа двух флуорофоров.
Теперь мы даем гораздо более детальное представление о сборке Р телец, подчеркивая роль сильно связанных молекул (как белков, так и РНК) в производстве конденсата, но также обсуждая важность сродства связывания и других параметров.По просьбе обозревателей мы также выполнили двухцветную визуализацию, чтобы изучить корреляцию между обогащением между различными молекулярными парами (новый рисунок 3). Эти данные выявили значительную корреляцию (коэффициент Пирсона 0,6-0,7) между обогащением Dcp2 и Edc3, Pat1 и Xrn1. Корреляция Dcp2-Xrn1 особенно интересна, поскольку неизвестно, как эти два белка связываются друг с другом напрямую, что указывает на корреляции, опосредованные косвенной связностью в конденсате (вероятно, через РНК или другие компоненты ядра P-тела).Однако корреляции не столь сильны, чтобы предполагать, что стехиометрически определенный комплекс лежит в основе образования Р-телец (модель, предложенная одним из рецензентов). Более того, как мы приводим ниже, существующие данные говорят против модели, в которой РНК является единственной подобной каркасу молекулой в P-тельцах (вторая модель, предложенная в обзоре), поскольку ряд белков также генетически показали, что они играют важную роль в образование конденсата. Эти вопросы — кооперативность в рекрутинге, возможность стехиометрического комплекса, роли взаимодействий РНК-РНК и РНК-белок и другие факторы — теперь довольно подробно рассматриваются в разделе «Обсуждение» рукописи.
Эти данные выявили значительную корреляцию (коэффициент Пирсона 0,6-0,7) между обогащением Dcp2 и Edc3, Pat1 и Xrn1. Корреляция Dcp2-Xrn1 особенно интересна, поскольку неизвестно, как эти два белка связываются друг с другом напрямую, что указывает на корреляции, опосредованные косвенной связностью в конденсате (вероятно, через РНК или другие компоненты ядра P-тела).Однако корреляции не столь сильны, чтобы предполагать, что стехиометрически определенный комплекс лежит в основе образования Р-телец (модель, предложенная одним из рецензентов). Более того, как мы приводим ниже, существующие данные говорят против модели, в которой РНК является единственной подобной каркасу молекулой в P-тельцах (вторая модель, предложенная в обзоре), поскольку ряд белков также генетически показали, что они играют важную роль в образование конденсата. Эти вопросы — кооперативность в рекрутинге, возможность стехиометрического комплекса, роли взаимодействий РНК-РНК и РНК-белок и другие факторы — теперь довольно подробно рассматриваются в разделе «Обсуждение» рукописи. Мы надеемся, что этот более подробный взгляд на образование конденсата понравится рецензентам.
Мы надеемся, что этот более подробный взгляд на образование конденсата понравится рецензентам.
3) Могут ли они предоставить какие-либо дополнительные данные или идеи, чтобы пролить свет на несоответствие, которое они указывают между их моделью и результатами для DCP2?
Мы решили эту проблему двумя способами. Во-первых, мы создали дополнительные мутанты Dcp2, чтобы лучше понять, как его связывание с Edc3 контролирует его обогащение P-тельцами (новый рисунок 6). Это приводит к важному выводу, что когда сродство между двумя конденсатными белками низкое, увеличение сродства может увеличить обогащение, но когда сродство уже высокое, его дальнейшее увеличение не увеличивает обогащение.Таким образом, настройка обогащения, вероятно, происходит за счет изменений в режиме низкого / умеренного сродства. Вдобавок, как описано выше, новое обсуждение дает более детальное представление о найме, поскольку происходит из комбинации возможности подключения, аффинности и активных процессов. При совместном рассмотрении они объясняют поведение наших мутантов Dcp2. По сути, не все регионы Dcp2 в равной степени способствуют обогащению; удаление элемента связывания с высоким сродством (либо к РНК, либо к Edc3) имеет гораздо более выраженный эффект на обогащение, чем удаление элемента с низким сродством.
При совместном рассмотрении они объясняют поведение наших мутантов Dcp2. По сути, не все регионы Dcp2 в равной степени способствуют обогащению; удаление элемента связывания с высоким сродством (либо к РНК, либо к Edc3) имеет гораздо более выраженный эффект на обогащение, чем удаление элемента с низким сродством.
Рецензент № 1:
[…] Для дальнейшего улучшения новинки предлагаем дополнительные вопросы, на которые можно было бы ответить:
1) Основным достижением этого исследования является точное измерение концентрации GFP-слитых белков, которое осуществляется путем сравнения интенсивности клеточного GFP со стандартными кривыми. Все выводы зависят от этих стандартов и вычитания фона. Несмотря на то, что лаборатория Розена ранее опубликовала статью, в которой использовался аналогичный подход (Banani et al., 2016), подробности этих калибровок и вычитания фона следует указывать в дополнительных материалах.
Мы добавили подробное описание получения изображения (показано на рисунке 1 — рисунок в приложении 2) и процедур анализа в раздел «Материалы и методы».
2) Чтобы оценить концентрацию белка в Р-тельцах, авторы сделали фундаментальное предположение: все Р-тельца имеют одинаковый и постоянный состав. Однако на Рисунке 1 авторы показали, что основные компоненты Р-телец, которые также являются наиболее распространенными белками, имеют сильную (4-5-кратную) изменчивость по концентрации и коэффициенту распределения.Неясно, как эта изменчивость влияет на относительные количественные характеристики, представленные в этой рукописи (рис. 3). Контрольный эксперимент, который мог бы ответить на этот вопрос, заключался бы в том, чтобы пометить несколько компонентов Р-телец спектрально различными флуоресцентными белками (2 или 3 одновременно) и убедиться, что ожидаемые относительные соотношения повторяются.
Отметим, что при измерении абсолютных концентраций различных видов мы не делали никаких предположений о том, имеют ли P-тела постоянный или переменный состав.Предполагая, что GFP-тегирование не изменяет концентрации данного белка (что подтверждают наши данные на рисунке 1 — рисунок в приложении 3), наши данные охватывают диапазон значений концентрации, взятых для каждого компонента в популяции клеток. Эти данные показывают, что составы имеют существенную изменчивость, как отмечает рецензент.
Эти данные показывают, что составы имеют существенную изменчивость, как отмечает рецензент.
При интерпретации данных становится важным вопрос относительной изменчивости. Здесь мы благодарим рецензента за предложение многоцветных экспериментов. На новом рисунке 3 мы использовали двухцветную визуализацию для одновременной количественной оценки концентраций трех пар белков в отдельных клетках: Dcp2 / Edc3, Dcp2 / Pat1 и Dcp2 / Xrn1.Мы обнаружили, что во всех случаях концентрации тел P в парах значительно коррелируют (R Пирсона 0,6–0,7). Таким образом, концентрации этих компонентов колеблются вместе (хотя и с некоторой остающейся вариабельностью, поскольку значения R не равны 1). Учитывая высокую взаимосвязь взаимодействий между белками Р-тельцов, мы полагаем, что многие компоненты, вероятно, демонстрируют сходные корреляции, но более обширный анализ этого момента выходит за рамки этого и без того длительного исследования.
Могут ли быть в популяции клеток несколько типов P-телец с разными композиционными профилями — это сложный вопрос.Мы не видим убедительных доказательств мультимодального распределения в профилях концентрации, что говорит против такой возможности. Но опять же, для всестороннего рассмотрения этой возможности потребуется гораздо больше данных, чтобы оценить, может ли распределение концентраций лучше соответствовать одной или нескольким популяциям.
3) В том же направлении неясно, экспрессировались ли белки, меченные GFP, которые использовались для количественной оценки компонентов Р-телец в фоне dcp1∆, из эндогенного локуса (Таблица 1: YRP1936, yRP2254, yRP2237, yRP2246, yRP2230, yRP1840, yRP1736, yRP2269, yRP1844, yRP1916, yRP1842).Авторы должны указать это, поскольку это будет иметь значительные последствия для количественной оценки, если будет смешанная популяция меченых и немаркированных белков.
Все меченные GFP и mCherry белки, использованные для создания рисунков 1-4, экспрессировались из своего эндогенного локуса. Об этом говорится в основном тексте и в Материалах и методах.
4) На рисунке 2 авторы обнаружили отрицательную корреляцию между скоростью восстановления фракции и концентрацией P-тел.Однако молекулярный механизм этой корреляции не обсуждается. Авторам следует расширить обсуждение и связать его с валентностью взаимодействия.
Как было предложено рецензентом, в Обсуждении мы теперь связываем связность в сети (количество различных молекулярных типов, с которыми взаимодействует данный белок, что связано с валентностью взаимодействия, количество различных молекул, с которыми взаимодействует данная молекула) с Поведение FRAP. Мы заявляем: «За исключением Xrn1 (см. Ниже), все основные белки обладают высокой валентностью взаимодействия (количество взаимодействующих молекул) и высокой связностью с другими белками Р-тельцов и РНК (≥ 4 непосредственно взаимодействующих молекул).[…] Эти особенности обычно должны обеспечивать более низкую концентрацию тел P и более быстрый и полный обмен ».
5) На рисунках 4 и 5 авторы измерили коэффициенты разделения нескольких мутантов Dcp2. Было бы важно продемонстрировать, что эти мутации существенно не влияют на стабильность и экспрессию белка, с помощью вестерн-блоттинга этих мутантов.
На рис. 5 — приложение к рисунку 1B и 1C и на рисунке 7 — приложение к рисунку 1A и 1B, теперь мы показываем вестерн-блоттинг почти всех мутантов Dcp2, исследованных под микроскопом.Эти данные показывают, что белки, сравниваемые на одном рисунке, экспрессируются примерно на одном уровне. Мутанты, не исследованные с помощью вестерн-блоттинга, были исследованы с помощью флуоресцентной визуализации для сравнения общего уровня экспрессии в проанализированных клетках (фигура 6 — рисунок в приложении 1). Мутанты Dcp2, сравниваемые на рисунке 6, действительно экспрессируются на несколько разных уровнях, так что Dcp2C∆5H, который слабо делится на P-тельца, экспрессируется на ~ 30% выше, чем Dcp2C∆5H-h2, который делит намного сильнее (рисунок 6 и рисунок 6 — приложение к рисунку 1).Однако направление этой разницы фактически усиливает наш вывод о том, что нижнее разделение Dcp2C∆5H происходит не из-за пониженной экспрессии.
6) На рисунке 5 авторы предполагают, что декапирующая активность Dcp2 влияет на коэффициент разделения и скорость восстановления. Однако, как авторы упоминают в тексте, неясно, связано ли это с увеличением цитоплазматической мРНК или прямым результатом каталитической активности Dcp2. Эта проблема может быть решена путем экспрессии вариантов Dcp2 в клетках дикого типа и индукции Р-телец с голоданием.
Благодарим рецензента за это предложение. Мы выполнили этот эксперимент, и результаты суммированы на новом рисунке 7. Вкратце, в условиях голодания динамика и распределение количества Р-телец одинаковы для двух мутантов, что позволяет предположить, что эти параметры в основном отвечают на мРНК. уровни при нарушении катализа. Однако общая доля материала в P-тельцах у каталитического мутанта во время голодания все же выше. Таким образом, этот параметр P-телец, по-видимому, является ответом на потерю каталитической активности.Из этих данных мы заключаем, что как увеличение уровней мРНК, так и потеря каталитической активности объясняют поведение мутанта WD.
7) На рис. 7 авторы обнаружили корреляцию между разделением Р-телец и валентностью взаимодействия белков. Это открытие было бы более убедительным, если бы авторы могли продемонстрировать, что когда валентность взаимодействия белка увеличивается, этот белок становится более концентрированным в Р-телец. Например, это можно сделать путем слияния клиентского белка с доменом каркасного белка, такого как Dcp2C ∆5H.Слитый белок должен обладать большей валентностью взаимодействия, и можно ожидать, что слитый белок будет больше концентрироваться в Р-тельцах. Этот эксперимент также усилит вывод, сделанный на Рисунке 6.
Мы удалили рисунок 7 из статьи и больше не обсуждаем количественные отношения между валентностью и разделением. Проблема с вызовом одной только валентности заключается в том, что она игнорирует сродство (проблема, о которой мы упоминали в предыдущем тексте, но не особо подчеркивали), а сродство играет важную роль в разделении Dcp2.В исправленной рукописи мы рассмотрели это двумя способами. Во-первых, мы прямо заявляем, что данные здесь и в литературе показывают, что связывание РНК N-концевым доменом и связывание Edc3 с помощью HLM1 происходит с высокой аффинностью и сильно способствует разделению Dcp2, в то время как C-концевые элементы HLM имеют меньшее сродство и вносят меньший вклад (рис. 5). Во-вторых, мы провели эксперименты, аналогичные предложенным рецензентом. Мы добавили высокоаффинный Edc3-связывающий мотив, HLM1, как к Dcp2 ∆5H, который разделяет слабо, так и к Dcp2 дикого типа, который сильно разделяет.Мы обнаружили, что добавление HLM1 только увеличивает разделение первого белка (рис. 6D). Из этих данных мы заключаем, что когда сродство к Edc3 низкое (Dcp2 ∆5H), добавление дополнительного сайта связывания с высоким сродством может увеличивать разделение Dcp2. Но когда аффинность уже высока (дикий тип Dcp2), дальнейшее ее увеличение не имеет никакого эффекта. Этот результат, вероятно, является общим при рассмотрении рекрутирования белков в конденсаты.
Рецензент № 2:
[…]
Эта модель была получена группой Розена в предыдущем исследовании, в котором изучались свойства разделения фаз in vitro искусственного набора белков с различным количеством низкоаффинных связывающих мотивов (валентность).Модель предсказывает, что «каркасы» (белки, необходимые для сборки конденсата) демонстрируют высокое обогащение и высокую валентность, тогда как «клиенты» демонстрируют низкое обогащение и низкую валентность. В соответствии с моделью авторы отмечают грубую корреляцию между валентностью и обогащением среди исследованных белков тела 19 P (но см. Ниже). Чтобы проверить модель напрямую, они удалили связывающие мотивы в одном предсказанном «высокомалентном» белке Р-телец (Dcp2). Удивительно, но они не обнаружили сильной корреляции между валентностью и обогащением (многие мутанты Dcp2 не лежат в корреляциях на Рисунке 7.Нам непонятно, как разрешить это несоответствие). Они также обнаружили, что некоторые известные клиенты (белки, не необходимые для сборки P-телец) демонстрируют сильное обогащение, вопреки модели. Несмотря на эти несоответствия, авторы продолжают утверждать, что их результаты подтверждают модель каркаса / клиента.
Мы тщательно пересмотрели текст и наш анализ наших данных, чтобы отреагировать на эту критику. Наиболее важно то, что мы удалили рисунок 7 и устранили все обсуждения, связанные с количественной корреляцией между валентностью и концентрацией P-тел.Как утверждает рецензент, одна только валентность не может объяснить наши данные о мутантах Dcp2. Теперь мы представляем гораздо более детальное представление о том, как, по-видимому, определяются концентрация и динамические свойства компонентов P-тела. Мы считаем, что связность взаимодействий (количество типов молекул, с которыми контактирует конкретная молекула) играет важную роль, поскольку большинство (6 из 7) высококонцентрированных белков также сильно связаны (≥4 партнеров по взаимодействию), и все слабоконцентрированные белки имеют низкую связность (≤2 партнеров по взаимодействию).Кроме того, было показано, что большинство (те же 6 из 7) сильно связанных молекул вносят вклад в сборку тел P, и ни одна из молекул с низкой связностью не делает этого. Тем не менее, связность — не единственный фактор, и сродство связывания и активные процессы также могут быть очень важны. В этом свете мы теперь обсуждаем, как взаимодействия N-концевого домена Dcp2 с РНК и элемента HLM1 с Edc3 имеют более высокое сродство, чем другие взаимодействия Dcp2, и, таким образом, играют более важную роль в определении концентрации белка в P-тельцах. (объясняя данные об удалении).Точно так же Xrn1 связывает РНК с высоким сродством. Таким образом, хотя его связность невысока, он сильно сконцентрирован в P-телах и демонстрирует умеренную динамику. Что касается активных процессов, гидролиз АТФ с помощью Dcp2 также вносит вклад в динамику белка. Таким образом, связь важна, но не вся история в определении поведения молекул конденсата. Эти вопросы обсуждаются в новом Обсуждении.
Благодаря этим соображениям мы пришли к другому взгляду на то, как следует использовать понятия «эшафот» и «клиент».Вместо того, чтобы классифицировать молекулы как каркас или клиент, мы теперь считаем, что эти термины следует использовать как дескрипторы. Молекула более подобна каркасу, если она играет большую роль в сборке Р-телец, и более подобна клиенту, если играет меньшую роль. Таким образом, мы можем объяснить тот факт, что разные молекулы по-разному влияют на делецию. Например. РНК, по-видимому, является наиболее подобным каркасу компонентом P-телец, поскольку ее удаление РНКазой разрушает конденсаты (что согласуется с тем фактом, что все 19 компонентов P-телец, количественно определяемые здесь, связывают РНК), в то время как Edc3 и Pat1 менее подобны каркасу, поскольку их делеционные штаммы сохраняют некоторые Р-тельца, хотя и меньшего размера, чем штаммы дикого типа.Напротив, все 12 изученных нами неосновных белков являются клиентоподобными, поскольку ни один из них не продемонстрировал значительного уменьшения сборки P-телец при удалении. Мы отмечаем, что связность (и, вероятно, центральность сети), по-видимому, играет роль в определении того, является ли молекула более похожей на каркас или более похожей на клиента, поскольку все наиболее связанные молекулы подобны каркасу (и более центральны в теле P. сеть взаимодействия) и молекулы с небольшим количеством подключений все похожи на клиентов (и более периферийны в сети).Эти вопросы обсуждаются в новом Обсуждении в новом разделе под названием «Общие принципы строительных лесов и клиентов в природных конденсатах».
Очень удивительно, что авторы не рассматривают другие модели, которые могли бы лучше соответствовать их данным. Одна возможность состоит в том, что РНК функционирует как истинный каркас для Р-тельцов и что белки рекрутируются в Р-тельца в силу их сродства к РНК или другим белкам, которые связывают РНК. В этом отношении интересно, что компоненты мультибелковых комплексов обнаруживаются в примерно стехиометрических количествах в P-тельцах, что позволяет предположить, что комплексы сохраняются в конденсатах.В этой альтернативной модели высокого сродства к РНК одного белка в комплексе было бы достаточно для обогащения всего комплекса, даже если ни одна из других субъединиц в комплексе не связывается с другими компонентами P-тельца.
Насколько мы понимаем этот комментарий, рецензент предполагает, что Р-тельца образуются посредством взаимодействий между мРНК, а затем эти мРНК рекрутируют основные белки Р-тельца за счет связывания РНК с высоким сродством. Однако несколько наблюдений в литературе опровергают эту модель.Например, как мы указываем в рукописи, формирование Р-телец явно требует белок-белковых взаимодействий, поскольку генетически показано, что Dcp2, Edc3, Dhh2, комплекс Lsm1-7 и Pat1 способствуют образованию Р-телец (например, Decker et al. , 2007; Sheth, Parker, 2006; Hondele et al., 2019; Rao, Parker, 2017). Более того, механизмы, с помощью которых белки способствуют образованию P-телец, могут быть напрямую связаны со специфическими белок-белковыми взаимодействиями, такими как димеризация Edc3 (Ling et al., 2008).Кроме того, некоторые белки нуждаются в других компонентах Р-телец для их повторного превращения в Р-тела. Напр., Dcp1 требует, чтобы Dcp2 рекрутировался в P-тела, а комплекс Lsm1-7 требует Pat1 (Teixeira and Parker, 2007). Таким образом, образование Р-телец и привлечение основных компонентов Р-телец к Р-тельцам требует взаимодействия белков.
Мы согласны с рецензентом в том, что РНК способствует образованию Р-телец. Явные данные свидетельствуют о том, что для образования Р-телец необходим пул нетранслирующих мРНК (Teixeira et al., 2005). Одна очевидная роль РНК заключается в обеспечении сайтов связывания для взаимодействующих белков, тем самым обеспечивая сборку Р-телец. В этой роли мы согласны с тем, что РНК функционирует подобно каркасу. Способствует ли РНК также формированию Р-телец посредством межмолекулярных взаимодействий РНК-РНК, еще предстоит установить. Тем не менее, модель, в которой РНК является единственным каркасом, который просто связывает белковые компоненты P-телец, маловероятна. Чтобы прояснить этот вопрос в рукописи, мы добавили этот аргумент в новое Обсуждение.
Более того, как описано выше, мы теперь представляем гораздо более подробный и детализированный вид сборки P-тела, который не зависит от простой бинарной классификации молекул как каркасов, так и клиентов. В пересмотренном Обсуждении также рассматривается вторая модель, предложенная здесь рецензентом, согласно которой высококонцентрированные белки образуют стехиометрический комплекс, который затем собирается на РНК с образованием более крупной структуры. Стехиометрический комплекс будет демонстрировать чрезвычайно высокую кооперативность при рекрутировании в P-тела, поскольку все элементы будут входить или выходить вместе.Хотя новый рисунок 3 действительно показывает корреляции между концентрациями Dcp2 в теле P с Edc3, Pat1 и Xrn1, корреляции недостаточно высоки (значения R Пирсона 0,6-0,7), чтобы предполагать стехиометрически определенный набор, даже если белки имеют примерно равные значения. средние концентрации в конденсате. Кроме того, концентрации P-телец Pat1 и комплекса Lsm1-7 снижаются примерно в два раза в штаммах дикого типа в условиях голодания по глюкозе по сравнению со штаммами dcp1Δ , что снова говорит против дискретной сборки.Эти аргументы представлены в новом абзаце Обсуждения.
Авторы также утверждают, что их анализ показывает, что относительно небольшое количество ядерных белков (7) составляет большую часть содержания белка в P-тельцах. Однако, поскольку другие компоненты P-тела, возможно, еще не обнаружены, это утверждение может быть только приближенным в настоящее время. Более того, даже если это правда, значение этой гипотезы неясно — белки, присутствующие в низких концентрациях в P-тельцах, могут играть важную роль.
Мы удалили это утверждение из текста.
В заключение, хотя в этом обзоре документируются концентрации и динамика значительного количества белков P-тела, оно, по-видимому, не дает существенного понимания «правил», которые регулируют состав P-тела. Модель, предложенная авторами, не согласуется с данными, и другие более правдоподобные модели не рассматриваются.
Как описано в нашем ответе на первый сводный комментарий в верхней части этого обзора, мы чувствуем, что наша работа выявила ряд новых и важных принципов, касающихся формирования и состава P-тел.В новом тексте представлено гораздо более подробное описание сборки P-тела, которое теперь согласуется со всеми нашими данными, и рассматривает множество различных механистических проблем и возможностей.
Рецензент № 3:
[…]
— Предел новизны. Уже было известно, что эти 9 белков являются основными белками P-тельца, которые также играют некоторую роль в функции P-тельца. Эти более ранние исследования также уже показали, что некоторые из этих белков содержат несколько доменов взаимодействия, через которые взаимодействуют компоненты P-тельца.
Как описано в ответе на Сводный пункт 1 в верхней части этого письма, мы считаем, что наша работа является новой во многих отношениях, включая то, что она является первым всеобъемлющим количественным анализом любого биомолекулярного конденсата. С помощью этого количественного анализа мы выявили два различных класса молекул на основе концентрации в структурах, чего нельзя было вывести из предыдущей работы, а также важную идею о том, что почти для всех компонентов только небольшая часть молекул присутствует в P тела.Анализ этих данных в свете известных молекулярных взаимодействий также выявил корреляции между физическим поведением компонентов P-тел и их молекулярной связностью, аффинностью связывания и активностью гидролиза АТФ. Опять же, это не могло быть выведено из предыдущих данных, поскольку они по своей сути зависят от нашего количественного анализа. Если рецензент сможет указать нам на соответствующую литературу, в которой документируется, что основные белки Р-телец, которые мы идентифицируем как высокообогащенные, имеют более высокие коэффициенты разделения, чем другие идентифицированные компоненты Р-телец, мы были бы счастливы соответствующим образом отредактировать нашу рукопись.
— Пределы количественного анализа. Конфокальный анализ имеет четкие пределы пространственного разрешения, а количественные измерения основаны на интенсивности флуоресценции, а не на измерениях отдельных молекул. Поскольку все компоненты помечены одним и тем же флуорофором (GFP), нельзя использовать парный анализ или анализ более высокого порядка для определения различий между гранулами, занятости белков, корреляции между белками, чтобы, например, проверить, все ли компоненты находятся в грануле одновременно. , и с аналогичной концентрацией.
Как подробно описано выше, мы выполнили двухцветную визуализацию нескольких пар белков P-телец и обнаружили существенную кооперативность между их обогащением. Это привело нас к более подробному рассмотрению сборки P body, которая описана в практически полностью переписанном разделе «Обсуждение» рукописи.
— Неясная актуальность анализа DCP2. Анализ DCP2, похоже, не сильно улучшает выводы статьи. Вместо того, чтобы показывать, что этот белок поливалентен, как было показано в прошлом путем разделения доменов других белков «каркаса», было бы информативно, если бы авторы предоставили, действительно ли «клиентские» белки имеют меньше доменов взаимодействия.
Наш анализ Dcp2 проанализировал механизмы, с помощью которых он обогащается внутри P тел. Хотя понятие многовалентности было продемонстрировано и для других белков, это рассмотрение предоставляет полезную информацию о Dcp2. Кроме того, в исследованиях Dcp2 мы обнаружили два ранее неизвестных свойства, которые, вероятно, имеют отношение к компонентам других конденсатов. Во-первых, мы обнаружили, что два фрагмента белка, которые только слабо разделяются на P-тельца по отдельности, при слиянии вместе сильно и совместно разделяются.Это происходит, даже если фрагменты связываются с разными компонентами P-тел, и, таким образом, не то же самое, что простая авидность. Когда мы приступили к работе, этот результат был для нас далеко не очевиден. Во-вторых, в новых данных, представленных на новом рисунке 6, мы обнаруживаем, что добавление элемента, связывающего Edc3 с высоким сродством к белку Dcp2, только увеличивает разделение на P тельца, когда Dcp2 имеет низкое сродство к Edc3. Когда сродство уже высокое, добавление дополнительных элементов привязки не имеет никакого эффекта. Таким образом, разделение компонента конденсата может быть насыщенным, вероятно, из-за ограниченных концентраций партнеров по связыванию.Опять же, это не был очевидный результат, когда мы начали эти эксперименты.
Что касается белков с поведением, подобным клиентскому (см. Ответ на пункт 1 сводки для нашего нового способа использования этого термина), как подробно описано выше, мы показываем в таблице 3 и на рисунке 2 — добавлении к рисунку 3, что все молекулы, которые слабо сконцентрированы в P Тельца имеют низкую связь с другими белками Р-телец и, как известно, не вносят значительный вклад в сборку Р-телец. Итак, действительно, поведение, подобное клиенту, коррелирует с ограниченным взаимодействием с другими компонентами тела Р.Корреляция не всегда верна в противоположном направлении. То есть существует небольшое количество белков, которые имеют лишь несколько связей с другими компонентами P-телец, но при этом сильно сконцентрированы в конденсате (Xrn1, Dcp1 и Pby1). Эти белки, как известно, обладают высоким сродством к другому высококонцентрированному компоненту P-тельца, который их рекрутирует. Эти вопросы теперь подробно рассматриваются в нашем обновленном Обсуждении.
[Примечание редакции: до принятия были предложены дальнейшие исправления, как описано ниже.]
Редакций:
Рецензенты сочли, что рукопись и работа вносят вклад в эту область, но необходимы некоторые исправления, чтобы учесть оговорки при интерпретации данных. Например, рецензент 2 считает, что термин «кооперативность» надлежащим образом не подтверждается данными и что мРНК не рассматривается как компонент P-телец.
Мы изменили наше описание данных на рисунке 5F, источника этого утверждения, на «синергетический», в котором говорится, что эффект объединения двух компонентов вместе приводит к большему эффекту, чем сумма компонентов по отдельности.В случае Dcp2 конструкция Dcp2 300Δh2 имеет ПК, равное 2,5, а конструкция Dcp2C Δ5H имеет ПК, равное 3,5, в то время как конструкция Dcp2Δh2 Δ5H, которая объединяет Dcp2 300Δh2 и Dcp2C Δ5H, имеет ПК 31, что> 2,5 *. 3.5. Таким образом, система соответствует определению синергии. Теперь отметим, что этот эффект может быть результатом кооперативного связывания с молекулами в теле P.
В рукописи РНК упоминается как важный компонент P-тельца. Мы считаем, что этого достаточно, особенно с учетом того, что статья сосредоточена на описании белков Р-тел, а не РНК.
Reviewer 3 считает, что функциональные свойства недостаточно изучены, особенно с учетом специфичности (почему исключены некоторые мРНК и RBP?).
Наше исследование не пытается рассмотреть функцию P-тел. Это сложный вопрос, который потребует гораздо больше и других данных, чем мы реально можем поместить в рукопись. Наша работа действительно обращается к специфичности рекрутирования белков в значительной степени посредством идеи синергетического рекрутирования Dcp2 в P тельца.То есть, резидентные белки часто содержат несколько слабых элементов рекрутирования, и, следовательно, белки, которые содержат мало таких элементов, рекрутируются плохо, если вообще рекрутируются. Рекрутирование РНК может следовать схожим принципам, и теперь мы прямо упоминаем эту идею.
Рецензент № 2:
В этой в значительной степени отредактированной рукописи авторы сообщают о всесторонней количественной оценке составляющих белков биологического конденсата, обедненных Dcp1 Р-телец дрожжей. Благодаря этому они идентифицировали семь белков «ядра» P-тельцов, которые существенно разделены, имеют высокую концентрацию и медленную подвижность при обогащении P-тельцами (рис. 1 и 2).Концентрация этих семи белков в тельцах P положительно коррелирует, что согласуется с предыдущими данными, показывающими, что эти белки взаимодействуют биохимически (рис. 3). Сообщенные параметры обогащения кажутся похожими, когда обедненные Dcp1 P-тельца сравниваются с P-тельцами дикого типа в условиях глюкозного голодания, однако Dcp1 и его партнер по связыванию Pby1 отсутствуют, а деградация РНК является дефектной в модели P-тельцов, используемой на протяжении всего анализа. Таким образом, хотя на уровне их биофизического анализа P-тела, обедненные Dcp1, кажутся похожими на P-тела в физиологических условиях, функционально эти «P-тела» различны.Это снижает актуальность данного исследования.
Автор обзора прав в том, что мы изучали Р-тельца в 2 различных условиях, только одно из которых является диким типом. Отметим, что это не полностью отличается от большинства исследований в этой области. Например, большинство анализов стрессовых гранул индуцируют в них мышьяк, токсин, который редко встречается в естественной биологии. Многие другие исследования связаны с избыточной экспрессией отдельных компонентов, что также является нефизиологической ситуацией.Тем не менее, такой анализ имеет существенное значение для понимания принципов, по которым собирается конденсат, и мы чувствуем, что наше исследование проводится в том же ключе. В то время как Dcp1 — / — P тельца и P тельца дикого типа, лишенные глюкозы, могут иметь разные функции, что поразительно, так это то, что их составы тесно связаны. Кроме того, сохраняются ключевые паттерны состава — небольшое количество высококонцентрированных белков и большее количество слабоконцентрированных белков — что является важной особенностью этих клеточных структур, которая, насколько нам известно, не была исследована в других местах (даже 3 недавно опубликованных Cell в статьях о стрессовых гранулах указаны только коэффициенты распределения, в которых отсутствует информация относительно абсолютных концентраций, указанных здесь).
На рис. 4 авторы наблюдают, что концентрации корового белка в Р-теле в значительной степени не зависят от стрессовых условий, однако авторы рассматривали только белковые компоненты, в то время как регулируемым субъектом стрессовых Р-телец являются мРНК, а не белки.
Мы заявляем, что наши данные не касаются изменений в содержании мРНК во время голодания: «… и они не говорят о секвестрации / хранении РНК…». Мы также отмечаем, что основной смысл рисунка 4 состоит в том, чтобы показать, что для большинства белков Р тельца существенно не истощают количество молекул из цитоплазмы, не сравнивая Dcp1 — / — и Р тельца дикого типа, лишенные глюкозы.Если обозреватель заинтересован, недавние результаты, полученные на клетках млекопитающих, предполагают, что состав мРНК P-телец, по-видимому, в значительной степени регулируется скоростью их трансляции, возможно, с дополнительным входом от длины поли (A) хвоста (Matheny et al., 2019) .
Затем авторы анализируют локализацию P-тельца DCP2 в двойном нуль-мутанте Dcp1, Dcp2, который все еще образует некоторые P-тела (хотя на рис. S1A показано только присутствие небольших структур, подобных «P-тельцам», но без количественного сравнения с контрольными P-тельцами, в дикого типа при голодании по глюкозе и истощении Dcp1).
Мы не совсем уверены, о чем здесь спрашивает рецензент, поскольку мы количественно оценили PC и динамику для различных повторно экспрессируемых мутантов Dcp2. Теперь мы количественно оценили распределение по размерам точки Edc3 в двойном нулевом мутанте Dcp1, Dcp2, показанном на рисунке S5A (рисунок S1A не имеет отношения к этим экспериментам; мы предполагаем опечатку от автора обзора), и сообщаем, что на рисунке 5 — дополнение к рисунку 1А.
В исследованиях делеций авт. Обнаруживают, что различные домены Dcp2 вносят вклад в разделение P тел и демонстрируют некоторую избыточность между доменом h2 и др. Спиральными доменами в CTP.Авторы приходят к выводу, что элементы в Dcp2 действуют «кооперативно». Однако их данные показывают аддитивность или, в лучшем случае, синергию, но не кооперативность. Чтобы использовать термин кооперативность, авторам необходимо продемонстрировать, что начальное событие связывания увеличивает сродство связывания последующих событий связывания.
Как указано выше, мы изменили наше описание данных на рисунке 5F, источника этого утверждения, на «синергетический», в котором говорится, что эффект объединения двух компонентов вместе приводит к большему эффекту, чем сумма компонентов по отдельности.В случае Dcp2 конструкция Dcp2 300Δh2 имеет ПК, равное 2,5, а конструкция Dcp2C Δ5H имеет ПК, равное 3,5, в то время как конструкция Dcp2Δh2 Δ5H, которая объединяет Dcp2 300Δh2 и Dcp2C Δ5H, имеет ПК 31, что> 2,5 *. 3.5. Таким образом, система соответствует определению синергии. Теперь отметим, что этот эффект может быть результатом кооперативного связывания с молекулами в теле P.
Авт. Используют исследование структуры-функции DCP2, чтобы доказать, что связность, аффинность и каталитическая активность являются тремя основными факторами для рекрутирования DCP2 P телец и размера и количества P телец.Однако необходимо провести более конкретные эксперименты, чтобы укрепить этот аргумент или обобщить открытие для других компонентов P-телец или других типов RNP-телец: 1) проведение исследования делеции домена / сайта связывания на Edc3 или других коровых белках; 2) добавление валентного или высокоаффинного сайта связывания к клиентоподобному белку, чтобы превратить его в каркас (достаточность). 3) Тестирование модели с белками скаффолда стрессовых гранул.
Мы согласны с тем, что наши аргументы будут подкреплены дополнительными экспериментами, которые запланированы для дополнительных проектов и публикаций в будущем.
В целом, это исследование технически выполнено хорошо, но довольно ограничено по своему содержанию или новизне. Истинное функциональное считывание отсутствует, поскольку экспериментальная установка не связана с функцией P-тельца (например, уровнем РНК в различных гранулах и активностью по удалению колпачков). Рукопись можно легко сжать до четырех рисунков, каждая из которых представляет собой одну точку для характеристики конкретного типа конденсата: Рисунок 1 и 2: обогащение определенных факторов в P-телах, Рисунки 3 и 4 соотношение концентраций между компонентами и в различных условиях. и рисунки 5 и 6, структурный анализ одного компонента, предполагающий частичную избыточность между CTD-спиралями, и рисунок 7 с исследованиями, касающимися связывания РНК.
Мы согласны с тем, что мы не пытались рассматривать функцию тела P в этой работе. Мы считаем, что такой анализ потребует отдельной публикации. Учитывая отсутствие ограничений по цифрам в eLife , мы не сжимали данные в меньшее количество цифр, так как мы опасаемся, что это затруднит чтение и понимание статьи (например, рисунок 4 не предназначен для сравнения различных условий, но скорее, чтобы подчеркнуть, что большинство молекул не присутствует в P-тельцах для большинства видов.Отсюда его название: «Р-тельца не сильно секвестрируют свои резидентные белки».
Рецензент № 3:
В этой отредактированной рукописи Xing et al. проанализировать обогащение и динамические свойства белков тельца ~ 20 P в дрожжах. Описательная часть исследования достаточно обширна (31 белок включен в первоначальный обзор, 19, численность которых была определена количественно) и предполагает существование двух групп: белки, сильно обогащенные P-тельцами, некоторые из которых также необходимы для сборки P-телец — и менее обогащенные белки — меньшее количество которых влияет на сборку P-телец.Исходя из этого, авторы предполагают, что P-тельца «биохимически проще, чем предполагает протеомика». В лучшем случае это предположение, а в худшем — грубое упрощение, поскольку изобилие не обязательно является предиктором функции — особенно для ферментов.
Для ясности мы изменили вывод на «композиционно проще, чем предполагает протеомика». Это должно устранить основную озабоченность рецензента. Однако мы отмечаем, что наши данные на рисунке 4 противоречат утверждению рецензента о том, что численность не предсказывает функцию.Этот рисунок показывает, что для почти всех белков, обогащенных Р-тельцами, подавляющее большинство молекулярных видов составляют , а не в организме, а скорее присутствуют в цитоплазме. Для всех молекул, кроме шести верхних, менее 20% находится в теле, даже при консервативной поправке на ненаблюдаемые субдифракционные Р-тела (без этой поправки значение падает до 8%). Таким образом, трудно утверждать, что количество P-тельца имеет большое значение для общей активности этого вида в клетке.Есть способы обойти эту проблему, включая, например, огромное увеличение удельной активности (активности на молекулу) внутри P-тельца по сравнению с цитоплазмой или совместную концентрацию нескольких молекул в каскаде. Но это требует дополнительных предположений, и, по нашему мнению, точка зрения первого порядка состоит в том, что изобилие действительно играет важную роль в формировании биохимических функций, возникающих из конденсатов, и, поскольку только небольшое количество молекул является высококонцентрированным, компартменты фактически являются , вероятно, будет биохимически проще, чем можно было бы представить из исследования протеомики, которое не определяет концентрации и, таким образом, по существу взвешивает все компоненты одинаково.
Во второй части исследования они авторы исследуют домены в DCP2, необходимые для локализации P тела, и идентифицируют множественные домены, которые, по-видимому, функционируют частично избыточно. Они пришли к выводу, что совокупность синергетических взаимодействий управляет сборкой P-телец, к этому выводу пришли несколько других исследований (рассмотренные в Lu Na and Slavoff, 2018), включая анализы in vitro (Shutz et al., 2017).
В целом, у нас осталось исследование, которое будет полезным справочным материалом для будущих исследований тела P, но оно не соответствует обещанию «принципов состава и специфичности».В частности, вопрос специфичности (почему не все РНК-связывающие белки локализуются в P-тельцах?) Напрямую не рассматривается. Авторы проводят исследования структуры-функции только одного белка Р-тельца и никогда не рассматривают, что исключает другие РНК-связывающие белки из Р-тельцов.
Как указано выше, мы считаем, что наша работа действительно затрагивает специфичность рекрутирования белков в значительной степени посредством идеи синергетического рекрутирования Dcp2 в P тельца. То есть, резидентные белки часто содержат несколько слабых элементов рекрутирования, и, следовательно, белки, которые содержат мало таких элементов, рекрутируются плохо, если вообще рекрутируются.Рекрутирование РНК может следовать схожим принципам, и теперь мы прямо упоминаем эту идею.
Наша работа также предполагает, что другие связывающие РНК белки действительно накапливаются в Р-тельцах, но их уровень просто отражает количество доступных сайтов связывания в мРНК Р-тельцов. В частности, хотя РНК-связывающий белок MS2 обычно не накапливается в Р-тельцах, добавление множества сайтов связывания для этого белка в мРНК, резидентную в Р-тельцах, может привести к накоплению MS2 в Р-тельцах.Во-вторых, мы действительно наблюдаем другие РНК-связывающие белки в Р-тельцах, просто они менее эффективно делятся на Р-тельца, чем основные компоненты (рис. 1).
Отметим, что исключение молекул из конденсата (PC <1) будет очень сложно наблюдать и значимо количественно определять в клетках, учитывая небольшой размер структур. Более того, более вероятно, что большинство молекул просто не будет задействовано (PC ~ 1), чем будет открыто исключено; мы не знаем о молекулах, исключенных из P-тел.В общем, физические механизмы, которые позволяют исключить образование конденсатов in vivo, вообще не поняты и выходят далеко за рамки настоящего исследования.
Мы также отмечаем, что, насколько нам известно, это первый раз, когда синергизм был количественно продемонстрирован для рекрутирования белка в конденсат in vivo. Упомянутая выше работа Sprangers интересна, но проводится исключительно in vitro. В клетках, чтобы заявить о синергии, необходимо удалить белок, а затем количественно оценить коэффициенты разделения для различных фрагментов при повторном введении индивидуально и вместе, как мы это сделали.Нам неизвестно, чтобы такой уровень количественного анализа проводился где-либо еще.
Дополнительные комментарии:
1) Авторы не последовательны в описании CTD. Во-первых, они предполагают, что «N-концевой домен, HLM1 и C-концевой домен все необходимы для эффективного разделения и поддержания характерной медленной динамики Dcp2». Однако позже они говорят, что «удаление CTD не влияет на ПК с полноразмерным Dcp2». Представленные данные предполагают, что C-концевой домен не требуется (300-конец), но может компенсировать отсутствие HLM1.
Благодарим рецензента за обнаружение этого несоответствия. Мы изменили первое утверждение на «Поскольку N-концевой домен и HLM1 необходимы для эффективного разделения и поддержания характерной медленной динамики Dcp2, а C-концевой домен может компенсировать отсутствие HLM1, мы заключаем, что элементы, управляющие разделением и динамика распределяется по белку ».
2) Неповрежденный C-концевой домен никогда не тестируется; вместо этого авторы использовали CTD с 5 мутированными доменами HLM.Хотя авторы подразумевают, что эта версия CTD имеет более слабую связь с телами P, данные не показаны, и из текста неясно, почему была использована эта версия CTD.
У нас были эти данные, и теперь мы показываем их на рисунке 5 — рисунок в приложении 1С.
3) Для данных на фиг. 7 авторы используют конструкции DCP2 с пониженным связыванием с Edc3, чтобы исследовать роль связывания РНК и активности декапирования в ассоциации P-телец. Однако неясно, почему связывание РНК изучали с использованием мутанта Dcp2 300, а декапирование изучали на мутанте Dcp2Δh2.
Это были по сути исторические особенности работы, связанные с тем, когда мы получали различные конструкции. Мы не считаем, что различия здесь влияют на наши выводы.
4) Цифра, сравнивающая концентрацию тельцов P / коэффициент распределения и скорости обмена для различных мутантов DCP2 (аналогично рис. 2A и B), может быть более интуитивным способом сравнения всех различных мутантов DCP2, использованных в этом исследовании.
Мы считаем, что, хотя в некоторых отношениях такой сюжет может помочь, в других он отвлекает внимание, и не включает его.Принципиальный недостаток состоит в том, что на рисунке 2 мы специально подчеркиваем корреляции между концентрацией тел P и обменными курсами. Но точки на рисунках 5 и 6 отличаются, и они сосредоточены на понимании различных аспектов рекрутирования Dcp2.
5) Исследования структурной функции DCP2 идентифицируют домены в DCP2, которые способствуют локализации P-телец. Достаточно ли этих доменов, чтобы направить белок, не являющийся Р-телом, в Р-тельца, не решается. Без таких «достаточных» экспериментов остается неясным, можно ли обобщить результаты для DCP2 для других белков.Например, они предполагают, что Xrn1 стабильно рекрутируется из-за его высокой аффинности связывания РНК, однако эта гипотеза не объясняет, почему другие высокоаффинные связывающие РНК белки исключаются из P тельцов.
В этом комментарии рассматриваются два момента. Во-первых, могут ли домены внутри Dcp2, которые способствуют локализации P-телец, действовать доминирующим образом. По сути, мы продемонстрировали этот феномен с GFP как чужеродным белком. Связанный с этим вопрос, но один выходит за рамки этой рукописи: может ли нацеливающий домен P-тельца Dcp2 перекрывать свойства других связывающих РНК белков, которые либо исключают их из P-тельцов, либо нацеливают эти связывающие РНК белки на другие конденсаты.Мы согласны с тем, что это будет интересный вопрос, и рассмотрим его в дальнейшей работе.
Второй вопрос, который поднимает рецензент, заключается в том, что мы не понимаем, почему другие связывающие РНК белки, которые могут связывать РНК с высоким сродством, не сильно разделяются на Р-тельца. Два наблюдения предполагают, что другие связывающие РНК белки действительно накапливаются в Р-тельцах, но их уровень просто отражает количество доступных сайтов связывания в мРНК Р-тельцов. В частности, хотя РНК-связывающий белок MS2 обычно не накапливается в Р-тельцах, добавление множества сайтов связывания для этого белка в мРНК, резидентную в Р-тельцах, может привести к накоплению MS2 в Р-тельцах.Во-вторых, мы действительно наблюдаем другие РНК-связывающие белки в Р-тельцах, просто они менее эффективно делятся на Р-тельца, чем основные компоненты (рис. 1).
6) В конце обсуждения авторы вводят концепцию авидности, когда множественные взаимодействия приводят к сильному связыванию. Возможно, будет полезно познакомить вас с этой концепцией раньше.
Теперь мы представили идею жадности в разделе результатов на рисунке 5.
https://doi.org/10.7554 / eLife.56525.sa2
Глава 3. Белки и аминокислоты
Глава 3. Белки и аминокислоты
1. БЕЛКИ
2. ПИЩЕВАРЕНИЕ БЕЛКОВ
И МЕТАБОЛИЗМ
3. ОБЩИЕ ТРЕБОВАНИЯ К БЕЛКАМ
4. АМИНОКИСЛОТЫ
5. КОЛИЧЕСТВО
ТРЕБОВАНИЯ К АМИНОКИСЛОТЕ
6. ДОБАВКА
ДИЕТЫ С АМИНОКИСЛОТАМИ
7. ССЫЛКИ
J. E. Halver
Вашингтонский университет
Сиэтл, Вашингтон
1.1 Классификация
1.2 Структура
1.3 Свойства
1.4 Химическое определение
Белки представляют собой сложные органические соединения, состоящие из многих аминокислот, связанных вместе пептидными связями и поперечно связанных между цепями сульфгидрильными связями, водородными связями и силами Ван-дер-Ваальса. Химический состав белков больше, чем у любой другой группы биологически активных соединений. Белки в различных клетках животных и растений придают этим тканям их биологическую специфичность.
1.1 Классификация
Белки можно разделить на:
(а) Простые белки. При гидролизе они дают только аминокислоты и иногда небольшие углеводные соединения. Примеры: альбумины, глобулины, глютелины, альбуминоиды, гистоны и протамины.
(б) Конъюгированные белки. Это простые белки в сочетании с некоторыми небелковыми веществами в организме. Примеры: нуклеопротеины, гликопротеины, фосфопротеины, гемоглобины и лецитопротеины.
(c) Производные белки. Это белки, полученные из простых или конъюгированных белков физическими или химическими способами. Примеры: денатурированные белки и пептиды.
1,2 Конструкция
Потенциальная конфигурация белковых молекул настолько сложна, что многие типы белковых молекул могут быть сконструированы и обнаружены в биологических материалах с различными физическими характеристиками. Глобулярные белки обнаруживаются в крови и тканевых жидкостях в аморфной глобулярной форме с очень тонкими или отсутствующими мембранами.Коллагеновые белки находятся в соединительной ткани, такой как кожа или клеточные мембраны. Волокнистые белки содержатся в волосах, мышцах и соединительной ткани. Кристаллические белки представлены хрусталиком глаза и подобными тканями. Ферменты — это белки с определенными химическими функциями, которые опосредуют большинство физиологических процессов жизни. Несколько небольших полипептидов действуют как гормоны в тканевых системах, контролируя различные химические или физиологические процессы. Мышечный белок состоит из нескольких форм полипептидов, которые позволяют мышцам сокращаться и расслабляться при физических движениях.
1.3 Недвижимость
Белки также можно охарактеризовать по их химическим реакциям. Большинство белков растворимы в воде, спирте, разбавленной основе или в различных концентрациях солевых растворов. Белки имеют характерную спиралевидную структуру, которая определяется последовательностью аминокислот в первичной полипептидной цепи и стереоконфигурацией радикальных групп, присоединенных к альфа-углероду каждой аминокислоты. Белки термолабильны, проявляя различную степень лабильности в зависимости от типа белка, раствора и температурного профиля.Белки могут быть обратимыми или необратимыми, денатурированными при нагревании, концентрации соли, замораживании, ультразвуковом воздействии или старении. Белки подвергаются характерному связыванию с другими белками в так называемой пластеиновой реакции и соединяются со свободными альдегидными и гидроксильными группами углеводов с образованием соединений типа Майяра.
1.4 Химическое определение
Содержание азота в большинстве белков, обнаруженных в тканях животных, орехов и зерна, составляет около 16 процентов; поэтому содержание белка обычно выражается как содержание азота × 6.25.
Проглоченные белки сначала расщепляются на более мелкие фрагменты пепсином в желудке или трипсином или химотрипсином из поджелудочной железы. Эти пептиды затем дополнительно восстанавливаются под действием карбоксипептидазы, которая гидролизует одну аминокислоту за раз, начиная со свободного карбоксильного конца молекулы, или с помощью аминопептидазы, которая отщепляет одну аминокислоту за раз, начиная со свободного амино-конца полипептида. цепь. Свободные аминокислоты, высвобождаемые в пищеварительную систему, затем всасываются через стенки желудочно-кишечного тракта в кровоток, где они затем повторно синтезируются в новые тканевые белки или катаболизируются для получения энергии или фрагментов для дальнейшего тканевого метаболизма.
Валовая потребность в белке была определена для нескольких видов рыб (см. Таблицу 1). Имитация цельного яичного протеина в тестовых диетах содержит избыток незаменимых аминокислот. Эти диеты поддерживались приблизительно изокалорийными за счет корректировки общего белка и усвояемых углеводных компонентов до фиксированного количества, поскольку лечение белковыми диетами варьировалось в испытанных диапазонах. Испытания на кормлении мальков, сеголетков и годовалых рыб показали, что общие потребности в белке наиболее высоки у начальных кормовых мальков и что они уменьшаются по мере увеличения размера рыбы.Чтобы расти с максимальной скоростью, мальки должны иметь диету, в которой почти половина легкоусвояемых ингредиентов состоит из сбалансированного белка; через 6-8 недель это требование снижается примерно до 40 процентов рациона лосося и форели и примерно до 35 процентов рациона годовалых лососевых, выращенных при стандартной температуре окружающей среды (SET). См. Рисунки 1 и 2. Общие потребности в белке молоди сома, по-видимому, меньше, чем у лососевых. Первоначально кормление мальков требует, чтобы около 50 процентов усвояемых компонентов рациона составлял белок, и потребность в них уменьшается с увеличением размера.Некоторые испытания кормления лососем показали прямую связь между изменениями потребности в белке молоди рыбы и изменениями температуры воды. Лосось чавычи в воде с температурой 7 ° C требует около 40 процентов цельного яичного белка для максимального роста; той же рыбе в воде с температурой 15 ° C требуется около 50% белка. Лосось, форель и сом могут использовать больше белка, чем требуется для максимального роста, благодаря эффективному удалению азотистых отходов в виде растворимых соединений аммиака через ткань жабр непосредственно в водную среду.Эта система удаления азота более эффективна, чем система, доступная для птиц и млекопитающих. Птица и млекопитающие потребляют энергию для синтеза мочевины, мочевой кислоты или других соединений азота, которые выводятся через ткань почек и выводятся с мочой. Перевариваемые углеводы и жиры сохранят избыток белка в рационе до тех пор, пока удовлетворяются потребности в белке для максимального роста (рисунки 1 и 2).
Таблица 1 — Расчетная потребность в белке с пищей для некоторых видов рыб 1/
|
Виды |
Уровень сырого протеина в рационе для оптимального роста (г / кг) |
|
Форель радужная ( Salmo gairdneri ) |
400-460 |
|
Карп ( Cyprinus carpio ) |
380 |
|
Чавыча ( Oncorhynchus tshawytscha ) |
400 |
|
Угорь ( Ангилья японский ) |
445 |
|
Камбала ( Pleuronectes platessa ) |
500 |
|
Золотистый лещ ( Chrysophrys aurata ) |
400 |
|
Белый амур ( Ctenopharyngodon idella ) |
410-430 |
|
Brycon sp. |
356 |
|
Морской лещ ( Chrysophrys major ) |
550 |
|
Желтохвост ( Seriola quinqueradiata ) |
550 |
1/ По материалам C.B. Cowey, 1978
Рис. 1. Потребность в белке чавычи при 47 ° F. Верхняя кривая: исходный индивидуальный средний вес рыбы, 1.5г. Нижняя кривая: исходная индивидуальная средняя масса рыбы 5,6 г.
Рис. 2. Потребность в белке чавычи при температуре 58 ° F. Верхняя кривая: исходный индивидуальный средний вес рыбы 2,6 г. Нижняя кривая: исходная индивидуальная средняя масса рыбы 5,8 г.
(Оба рисунка адаптированы из: DeLong, D.C., J.E. Halver and E.T. Mertz, 1958, J.Nutr ., 65: 589-99)
Обычно рыбе нужно давать диету, содержащую дифференцированный уровень высококачественного белка и энергии, а также адекватный баланс незаменимых жирных кислот, витаминов и минералов в течение длительного периода времени.Из полученной кривой доза / ответ потребность в белке обычно получают по графику Альмквиста. Считается, что эти различия в очевидной потребности в белке связаны с различиями в методах культивирования и составе рациона.
Относительно высокие уровни пищевого белка, необходимые для максимального роста некоторых рыб, таких как белый амур, Ctenopharyngodon idella, и Brycon spp. удивительны, поскольку эти рыбы всеядны. Brycon spp.выращиваются на нежелательных фруктах и другом растительном материале с низким содержанием белка, и в этих условиях, по-видимому, существенный вклад в потребление ими белка вносит естественная пищевая цепь.
Потребность в белке эвриталинных рыб, таких как радужная форель, Salmo gairdneri, и кижуч, Oncorhynchus кисач, , выращенных в воде с соленостью 20 ppt, примерно такая же, как потребность в пресной воде. Нет данных о потребности этих видов в белке в морской воде с полной концентрацией.(35 п.
4.1 Essential
и заменимые аминокислоты
4.2 Незаменимые
Аминокислоты и качество протеина
Аминокислоты являются строительными блоками белков; около 23 аминокислот были выделены из природных белков. Десять из них незаменимы для рыб. Животное не способно синтезировать незаменимые аминокислоты и поэтому должно получать их с пищей.
4.1 Незаменимые и заменимые аминокислоты
Корм для лосося, форели и канального сома, лишенный аргинина, гистидина, изолейцина, лейцина, лизина, метионина, фенилаланина, треонина, триптофана или валина, не рос (рис.3). Те же самые рыбы, которых кормили рационами, лишенными других L-аминокислот, росли так же, как и рыбы, получавшие все 18 протестированных аминокислот (рис. 4). Азотный компонент в тестируемых диетах состоял из 18 L-аминокислот по образцу цельного яичного белка. Вся тестируемая рыба быстро выздоравливала, когда в рационе была заменена недостающая аминокислота. Наклон кривой роста в группе восстановления был идентичен таковому у рыб, получавших полный тестовый рацион с аминокислотами.
Заменяемые аминокислоты были протестированы: аланин, аспарагиновая кислота, цистин, глутаминовая кислота, глицин, пролин, серин и тирозин.Было обнаружено, что эти аминокислоты не являются необходимыми для роста лосося, форели и канального сома.
Для количественных исследований потребности в 10 незаменимых аминокислотах использовалась смесь казеина и желатина с добавлением кристаллических L-аминокислот. Тестируемая диета содержала 40 процентов цельного яичного белка для азотного компонента. Эксперименты, проведенные с карпом и угрем, показали аналогичное отсутствие роста, когда в рационе отсутствовала незаменимая аминокислота.
Рис. 3. Рост рыб с дефицитом аргинина. Группа с дефицитом была разделена через шесть недель на диете с дефицитом, и недостающая аминокислота была заменена в одной из двух частей.
Рис. 4. Рост рыб с дефицитом цистина.
(Оба рисунка взяты из: DeLong, D.C., J.E. Halver and E.T. Mertz, 1958, J.Nutr., 65: 589-99)
4.2 Основные аминокислоты и качество белка
Если известны потребности рыбы в незаменимых аминокислотах, должно быть возможно удовлетворить эти потребности в системах культивирования различными способами за счет различных пищевых белков или комбинаций пищевых белков.
Фенилаланин избавлен от тирозина. Неизвестно, что он химически модифицирован или становится недоступным из-за суровых условий, которым обычно подвергаются кормовые белки во время обработки. Измерение фенилаланина в белках несложно, поэтому обеспечение и оценка фенилаланина в белках в практических диетах не представляет особых трудностей.
Лизин — основная аминокислота. В дополнение к -аминокислотной группе, обычно связанной пептидной связью, он также содержит вторую -аминогруппу.Эта альфа-аминогруппа должна быть свободной и реакционной, иначе лизин, хотя и поддается химическому измерению, не будет доступен биологически. Во время обработки белков корма α-аминогруппа лизина может реагировать с небелковыми молекулами, присутствующими в корме, с образованием дополнительных соединений, которые делают лизин биологически недоступным.
Цистин избавлен от метионина. Однако измерить содержание метионина в кормовых белках непросто, поскольку аминокислота подвержена окислению во время обработки.После обработки метионин может присутствовать как таковой, или в виде сульфоксида, или в виде сульфона. Сульфоксид может образовываться из метионина во время кислотного гидролиза кормового белка перед измерением его кислотного состава, не содержащего кислоты. Кислотный гидролиз белков перед анализом нарушает исходное равновесие между двумя соединениями, так что состав гидролизата больше не отражает состав белка. При определении содержания метионина в чистых белках окисление аминокислоты до метионинсульфона обычно является количественным.В случае кормовых белков, однако, это не покажет, сколько метионина или сульфоксида метионина присутствовало в белке до его окисления и гидролиза.
Сульфоксид метионина может иметь некоторую биологическую ценность для рыб, которые могут иметь некоторую способность обратного преобразования его в метионин и, таким образом, частично восполнять часть метионина, окисленного во время обработки.
Недавно появились сообщения о методах измерения метионина в белках с использованием йодоплатинатного реагента до и после восстановления трихлоридом титана, чтобы получить значения как для метионина, так и для сульфоксида в исходном белке.Также был описан способ измерения метионина конкретно по расщеплению цианогенбромида. Оба метода еще предстоит оценить независимо. Микробиологический анализ метионина в белках кормов является ценным инструментом, хотя существует опасность того, что оксиды метионина могут различаться по своей активности в отношении микроорганизмов и искажать значения.
Количественные потребности лососевых в десяти незаменимых аминокислотах определялись путем кормления линейными приращениями одной аминокислоты за раз в тестируемой диете, содержащей аминокислотный профиль, идентичный цельному яичному белку, за исключением тестируемой аминокислоты.Повторяющиеся группы рыб подвергались диетическому лечению до тех пор, пока не появлялись большие различия в росте исследуемых партий. График реакции роста Альмквиста показывает уровень аминокислот, необходимый для максимального роста в этих конкретных условиях испытания. Рационы были разработаны таким образом, чтобы содержать белок на уровне или немного ниже оптимальной потребности в белке для данного вида и условий испытаний, чтобы гарантировать максимальное использование ограничивающей аминокислоты. Сравнение требований к десяти незаменимым аминокислотам между видами показано в таблице 2.
Недавним нововведением стало использование в тестовых диетах белков, относительно дефицитных по данной незаменимой аминокислоте. Таким образом, комбинации рыбной муки и зеина использовались в тестовых диетах для определения потребности радужной форели в аргинине. Рационы, содержащие различные относительные количества казеина и желатина, показали, что увеличение уровня связанного с белком аргинина с 11 до 17 г / кг привело к значительному увеличению роста канального сома.
Таблица 2 Потребность семи животных в аминокислотах 1/
|
Аминокислота |
Молодь угря |
Мальки карпа |
Канальный сом |
Молодь чавычи |
цыпленок |
Молодой поросенок |
Крыса |
|
Аргинин |
3.9 (1,7 / 42) |
4,3 (1,65 / 38,5) |
6,0 (2,4 / 40) |
6,1 (1,1 / 18) |
1,5 (0,2 / 13) |
1,0 (0,2 / 19) |
|
|
Гистидин |
1,9 (0,8 / 42) |
1,8 (0,7 / 40) |
1,7 (0,3 / 18) |
1.5 (0,2 / 13) |
2,1 (0,4 / 19) |
||
|
Изолейцин |
3,6 (1,5 / 42) |
2,6 (1,0 / 38,5) |
2,2 (0,9 / 41) |
4,4 (0,8 / 18) |
4,6 (0,6 / 13) |
3,9 (0,5 / 13) |
|
|
лейцин |
4.1 (1,7 / 42) |
3,9 (1,5 / 38,5) |
3,9 (1,6 / 41) |
6,7 (1,2 / 18) |
4,6 (0,6 / 13) |
4,5 (0,9 / 19) |
|
|
Лизин |
4,8 (2,0 / 42) |
5,1 (1,23 / 24,0) |
5,0 (2,0 / 40) |
6.1 (1.1 / 18) |
4,7 (0,65 / 13) |
5,4 (1,0 / 19) |
|
|
метионин 2/ |
4,5 (2,1 / 42) 3/ |
3,1 (1,2 / 38,5) |
2,3 (0,56 / 24,0) |
4,0 (1,6 / 40) 3/ |
4.4 (0,8 / 18) |
3,0 (0,6 / 20) |
3,0 (0,6 / 20) |
|
Фенилаланин 4/ |
5,1 (2,1 / 41) 5/ |
7,2 (1,3 / 18) |
3.6 (0,45 / 13) |
5,3 (0,9 / 17) |
|||
|
Треонин |
3,6 (1,5 / 42) |
2,2 (0,9 / 40) |
3,3 (0,6 / 18) |
3,0 (0,4 / 13) |
3,1 (0,2 / 19) |
||
|
Триптофан |
1,0 (0,4 / 42) |
0.5 (0,2 / 40) |
1,1 (0,2 / 18) |
0,8 (0,2 / 25) |
1,0 (0,2 / 19) |
||
|
Валин |
3,6 (1,5 / 42) |
3,2 (1,3 / 40) |
4,4 (0,8 / 18) |
3,1 (0,4 / 13) |
3,1 (0,4 / 13) |
1/ Выражается в процентах от диетического белка.В скобках числители — это потребности в процентах от сухого рациона, а знаменатели — это процент общего содержания белка в рационе.
2/ При отсутствии цистина
3/ Метионин плюс цистин
4/ При отсутствии tyro sine
5/ Фенилаланин плюс тирозин
(По материалам: Национальный исследовательский совет, 1977 г.)
Потребность радужной форели в аргинине была определена по стандартной кривой доза / реакция (рост), а также путем измерения уровней свободного аргинина в тканях (крови и мышцах) в группах форели, получавших возрастающее количество аргинина в рационе.После того, как диетическая потребность форели в аргинине была удовлетворена, любое дальнейшее увеличение потребления аргинина привело к увеличению концентрации свободного аргинина в крови и мышцах. Было получено хорошее согласие между двумя методами.
Данные, представленные в таблице 2, позволяют предположить, что между видами рыб существуют реальные различия в их потребностях в определенных аминокислотах. Это приводит к трудностям при составлении белкового компонента практического рациона для тех видов, потребности которых в аминокислотах еще не известны.Возможное решение — использовать для каждой аминокислоты наивысший уровень, необходимый для любого из тех видов, по которым имеются данные. Необходимость дополнительных количественных данных о потребностях рыб в аминокислотах, особенно тех, которые действительно или потенциально могут использоваться в качестве сельскохозяйственных животных, очевидна.
Одним из решений использования белков, относительно дефицитных по одной или нескольким аминокислотам, является добавление в белок соответствующих количеств аминокислоты, необходимых в практических диетах. Рыба, по-видимому, использует свободные аминокислоты с разной степенью эффективности.
Молодой карп, Cyprinus carpio, оказался неспособным расти на диетах, в которых белковый компонент (казеин, желатин) был заменен смесью аминокислот, аналогичных по общему составу. Гидролизат трипсина казеина также оказался неэффективным. Однако, если диета, содержащая свободные аминокислоты в качестве белкового компонента, тщательно нейтрализуется NaOH до pH 6,5-6,7, то некоторый рост молоди карпа все же происходит. Этот рост был заметно ниже, чем при сопоставимой казеиновой диете в тех же условиях.
Канальный сом также не может использовать свободные аминокислоты в качестве добавок к дефицитным белкам. Когда соевый шрот был заменен изоназотом на муку менхадена, рост и эффективность корма канального сома были значительно снижены. Добавление свободного метионина, цистина или лизина, наиболее ограничивающих аминокислот, к этим заменителям сои не привело к увеличению веса.
Повышение уровня аргинина в рационе сома с 11 до 17 г / кг путем изонитрогенной замены желатина на казеин значительно увеличивало набор веса, но добавление свободного аргинина, цистина, триптофана или метионина к казеину мало влияло на рост или преобразование пищи.
Лососевые могут использовать свободные аминокислоты для роста. Было показано, что зеин-желатиновая диета с добавлением лизина и тритофана заметно превосходит зеин-желатиновую диету для радужной форели, когда в качестве критериев использовались прибавка в весе и использование белка.
Несколько исследователей продемонстрировали возможность дополнения белков с дефицитом аминокислот ограничивающими аминокислотами в рационах лососевых. Казеин с добавлением шести аминокислот давал коэффициенты конверсии корма для атлантического лосося, аналогичные тем, которые были получены при использовании изолированного рыбного белка в качестве источника пищевого белка.Соевый шрот с добавлением пяти или более аминокислот (включая метионин и лизин) был лучшим источником белка для радужной форели по сравнению с соевым шротом. Однако однократное добавление метионина и лизина не привело к повышению ценности соевого шрота. Эти результаты позволяют предположить, что аминокислотный спектр выделенного рыбьего белка, который они использовали, может приблизительно соответствовать потребности в аминокислотах радужной форели. Пищевая ценность изолята соевого белка может быть увеличена путем добавления в него первой ограничивающей аминокислоты; я.е., метионин.
Рационы, содержащие в качестве белкового компонента рыбную муку, мясокостную муку, а также дрожжевую и соевую муку, можно улучшить путем одновременного добавления цистина (10 г / кг) и триптофана (5 г / кг). Рыбную муку можно полностью заменить без снижения конверсии корма в рационах для радужной форели смесью из субпродуктов домашней птицы и перьевой муки вместе с 17 г лизина HCL / кг, 4,8 г DL-метионина / кг и 1,44 г DL. -триптофан / кг.
Коуи, К.Б. и Дж. Р. Сардженты, 1972 Кормление рыб. Adv.Mar.Biol., 10: 383-492
Cowey, C.B., 1979 Потребности рыб в белках и аминокислотах. В Технология кормления и кормления рыб, под редакцией Дж. Э. Халвера и К. Тьюса. Материалы Всемирного симпозиума, спонсируемого EIFAC / FAO, ICES и IUNS, Гамбург, 20-23 июня 1978 г. Schr . Bundesforschungsanst . Fisch ., Hamb ., (14/15) vol. 1: 3-16
Мерц, Э.Т., 1972 г. Потребности в белке и аминокислотах. В Питание рыб, под редакцией Дж. Э. Халвера. Нью-Йорк, Academic Press, стр. 106-43.
Национальный исследовательский совет, Подкомитет по тепловодным рыбам 1977 года, Потребности теплопроводных рыб в питательных веществах. Вашингтон, округ Колумбия, Национальная академия наук (потребности домашних животных в питательных веществах) 78 стр.
Роль белка в организме »МАЛЕСТРОМ
Белок очень важен для вашего здоровья.Белки состоят из аминокислот, и все они соединяются в цепочки, почти как нить бусинок. Тысячи белков в организме состоят только из 20 вариантов аминокислот. Эти белки продолжают выполнять различные жизненно важные задачи в организме.
Прочтите подробное описание этих важнейших функций.
Рост и обслуживание
Вашему организму необходим белок для роста и поддержания различных тканей. Белки в организме находятся в постоянном состоянии обмена, поэтому потребление белка важно.Обычно организм расщепляет только то количество белка, которое используется; однако иногда он расщепляет больше белка, чем может произвести, что затем увеличивает потребность организма. Потребности человека в белке будут зависеть от его здоровья и уровня активности.
Биохимические реакции
Ферменты — это белки, которые помогают в биохимических реакциях, происходящих внутри вашего тела. Структура этих ферментов позволяет им соединяться с субстратами, которые затем действуют как каталитические реакции, которые жизненно важны для вашего метаболизма.Ферменты также выполняют другие функции, такие как пищеварение, свертывание крови, сокращение мышц и выработку энергии. Следовательно, недостаток этих ферментов может быть вредным.
Химические посланники
Некоторые белки образуют гормоны, которые действуют как посредники между клетками, тканями и органами. Гормоны можно разделить на три категории: белки и пептиды, стероиды и амины. Аминокислотные цепи образуют белок и пептиды, которые составляют большую часть гормонов организма.
Обеспечение структуры
Некоторые белки более волокнистые; они придают тканям и клеткам большую структуру. Кроме того, они образуют соединительный каркас для некоторых структур внутри тела. Они также содержат некоторые белки, которые способствуют укреплению и эластичности, такие как кератин, коллаген и эластин.
pH Balance
Белок может помочь регулировать концентрацию кислот и щелочей в жидкостях вашего организма. Этот баланс измеряется с помощью шкалы pH.Постоянный pH имеет решающее значение, поскольку небольшое изменение может быть чрезвычайно пагубным, а иногда и смертельным. Некоторые белки помогают поддерживать этот баланс.
Fluid Balance
Белки помогают регулировать другие процессы в организме, что помогает поддерживать баланс жидкости в организме. Они притягивают и удерживают воду; если вы не потребляете достаточное количество белка, эти белки уменьшаются, что затем приводит к накоплению жидкости, что приводит к белковой недостаточности, что плохо для вас.
укрепляет иммунную систему
Белки помогают создавать антитела, которые борются с инфекциями.Они защищают организм от бактерий и вирусов. Без них бактерии и вирусы могли бы размножаться и в конечном итоге поражать организм. После того, как они были приготовлены, ваше тело запоминает рецепт, а это означает, что он может быстрее отреагировать, если вы снова заразитесь — это основная предпосылка, лежащая в основе вакцин.
Транспортировка питательных веществ
Некоторые белки переносят вещества по всему телу, через кровоток в клетки и из них. Питательные вещества, такие как витамины, минералы, сахар, кислород и холестерин, переносятся белками.Например, гемоглобин — это белок, который переносит кислород по телу. Белки также удерживают питательные вещества. Например, ферритин — это белок, хранящий железо.
Обеспечение энергией
Белки помогают снабжать организм энергией. Белки и углеводы содержат около четырех калорий на грамм, а жиры — девять калорий на грамм. Однако, поскольку белки служат многим другим целям, ваше тело не хочет использовать их для получения энергии; вместо этого он предпочитает хранить и использовать жиры и углеводы, когда происходит увеличение веса.С другой стороны, в случае недоедания или голодания организм начинает лишать мышцы, чтобы использовать запасенные в них аминокислоты для создания энергии.
Важность белка
Теперь должно быть очевидно, что белок важен. Есть много разных источников белка, которые вы можете включить в свой рацион. Например, есть животные белки из мяса, рыбы, молочных продуктов и яиц. Или, если вы растительный, то есть варианты, такие как соя, тофу или сейтан.Существуют также добавки и протеиновые порошки, которые вы можете купить, чтобы получать протеин с собой. Becomeio предлагает ряд различных протеиновых добавок, которые подходят всем — независимо от того, растительные вы или нет. У них также есть информативный блог, в котором можно найти ответы на все ваши вопросы, включая обзоры протеинового порошка, советы по тренировкам и улучшение усвоения протеина.
Заключение
Белок выполняет множество различных ролей в организме, все из которых одинаково важны, от построения и восстановления тканей до метаболических функций и функций организма.Они также обеспечивают структурную основу, поддерживают баланс и улучшают иммунную систему, поэтому очень важно следить за потреблением белка. В совокупности функции, которые различные белки обеспечивают в организме, делают его одним из наиболее важных питательных веществ для вашего здоровья и благополучия.
Белки и масса тела | Молочное питание
Вернуться к белку
г.Харви Андерсон, доктор философии
Профессор диетологии и физиологии; Директор программы по безопасности пищевых продуктов, питанию и нормативным вопросам, Департамент диетологии, медицинский факультет, Университет Торонто
Роль белка в регулировании долгосрочного энергетического баланса и поддержании здоровой массы тела остается неясной.Тем не менее, диеты с высоким содержанием белка остаются популярными из-за очевидных преимуществ, таких как повышенное чувство сытости. Появляется все больше свидетельств того, что источник белка очень важен в этом отношении.
Роль молочных продуктов
Было показано, что молочные продукты и / или молочные компоненты подавляют кратковременное потребление пищи, повышают субъективное чувство сытости и стимулируют механизмы, которые, как известно, сигнализируют о насыщении и сытости. 1 Кальций также может играть роль в регулировании массы тела. 2 Однако несколько исследований показывают, что добавки кальция имеют менее значительный эффект, чем молочный кальций, что позволяет предположить, что в молоке могут действовать другие компоненты, кроме кальция. 2 Молочные белки содержат намного больше аминокислот с разветвленной цепью (BCAA), чем мясные или растительные белки, особенно лейцин, что является предполагаемым преимуществом для регулирования приема пищи и поддержания безжировой массы тела. 3
Преимущества высококачественного белка
Исследования показывают четыре линии доказательств, подтверждающих роль белков в регулировании приема пищи и поддержании здоровой массы тела. 3 Кратко:
- Белок подавляет потребление пищи больше, чем жиры или углеводы
- Белок способствует насыщению и задерживает возвращение чувства голода по сравнению с жирами и углеводами
- Диеты с ограничением энергии с высоким содержанием белка поддерживают поддержание безжировой массы тела, тем самым способствуя потере веса, прежде всего, за счет жировой ткани
- Переваривание белков приводит к стимуляции многих физиологических и метаболических реакций, которые, как известно, участвуют в регуляции приема пищи.
Разнообразные исследования показывают, что источник белка является важным фактором. 3,4 Например, исследования на крысах показывают, что подавление приема пищи в следующий час кормления больше после кормления через желудочный зонд с сывороткой по сравнению с яичным альбумином и соевым белком. 5 Два основных белка в молоке, казеин и сыворотка, каждый вносит свой вклад в чувство насыщения. 4 Казеин, который переваривается медленнее, способствует долгосрочному насыщению, в то время как сыворотка способствует более краткосрочному насыщению. 4 Кроме того, пептиды и другие биоактивные компоненты, содержащиеся в молочных продуктах, по-видимому, обладают рядом преимуществ, включая модуляцию артериального давления, воспалительные процессы и контроль уровня глюкозы в крови. 4 К сожалению, большинство канадцев не могут удовлетворить свою суточную потребность в молочных продуктах. 6
Основные моменты
- Растущее количество данных подтверждает роль молочных продуктов в регулировании массы тела.
- Добавки кальция, по-видимому, не обладают такой же пользой, как молочный кальций, что позволяет предположить, что другие компоненты в молоке могут быть фактором.
- Белки в молоке, включая казеин и сыворотку, улучшают чувство сытости, регулируют прием пищи и способствуют поддержанию мышечной массы.
- Пептиды и другие биоактивные компоненты в молочных продуктах обладают дополнительными преимуществами, включая модуляцию артериального давления, воспаления и уровня глюкозы в крови.
Список литературы
- Азиз А. и Андерсон Г. Влияние молочных компонентов на потребление пищи и сытость: механизмы действия и последствия для разработки функциональных продуктов питания.На Саареле М. (ред.): «Функциональные молочные продукты». Кембридж, Великобритания: Woodhead Publishing Ltd, т. 2, 2007.
- Major GC et al. Последние достижения в исследованиях ожирения, связанного с кальцием. Обзоры ожирения 2008; 9: 428-445.
- Андерсон Г. Х. и Мур С. Е.. Диетические белки в регулировании приема пищи и массы тела у людей. J Nutr 2004; 134: 974S-979S.
- Луховый Б.Л. и соавт. Сывороточные протеины в регуляции приема пищи и сытости. J Am Coll Nutr 2007; 26 (6): 704S-712S.
- Морган Г. Роль холецистокининовых рецепторов в подавлении потребления пищи у крыс, вызванном гидролизатом белка. В: Департамент диетологии , Торонто, Онтарио: Университет Торонто; 1998: 143.
- Статистическое управление Канады. Питание: результаты Обследования состояния здоровья населения Канады, 2004 г. Обзор пищевых привычек канадцев. Оттава, Канада: Отдел статистики здравоохранения, 2006: 47.
Ключевые слова:
белок
.