ru:about:media:2018:20180108 [Институт химической биологии и фундаментальной медицины]
Оригинал статьи
РИА Новости
от 01.08.2018 г.
Формула молока: грудное кормление защищает от инфекций и рака
МОСКВА, 1 авг — РИА Новости, Альфия Еникеева. Младенец полноценно развивается только при естественном вскармливании, и альтернативы этому пока нет: до сих пор синтезированы лишь несколько важных белков для искусственных смесей. Во Всемирную неделю грудного вскармливания РИА Новости вместе с экспертами выясняет, чем уникально материнское молоко и почему его ничем не заменить.
Найди сотни отличий
«Грудное молоко — это живая ткань, которая постоянно меняет свой состав в зависимости от времени суток, возраста ребенка и его потребностей в воде и пище. Кроме макронутриентов — белков, жиров и углеводов — в нем есть гуморальные (лизоцим, лактоферрин, секреторный иммуноглобулин А) и клеточные компоненты (лейкоциты, макрофаги и нейтрофилы), пребиотики (олигосахариды) и пробиотики (лакто- и бифидобактерии), обеспечивающие защиту младенца от инфекций. Грудное молоко содержит также факторы роста, отвечающие за созревание желудочно-кишечного тракта. Всего — более четырехсот полезных для здоровья и развития младенца компонентов. В искусственных смесях их не более тридцати», — рассказывает доктор медицинских наук Любовь Абольян, главный специалист по организации общественного здоровья Первого МГМУ имени И. М. Сеченова Минздрава России и руководитель Российского научно-практического центра по пропаганде и поддержке грудного вскармливания.
Грудное молоко содержит также факторы роста, отвечающие за созревание желудочно-кишечного тракта. Всего — более четырехсот полезных для здоровья и развития младенца компонентов. В искусственных смесях их не более тридцати», — рассказывает доктор медицинских наук Любовь Абольян, главный специалист по организации общественного здоровья Первого МГМУ имени И. М. Сеченова Минздрава России и руководитель Российского научно-практического центра по пропаганде и поддержке грудного вскармливания.
Если сравнивать молоко разных млекопитающих, то его качественный состав, на первый взгляд, кажется одинаковым — везде одни и те же компоненты, только в разных пропорциях. В молоке животных выше содержание белков, которые необходимы для быстрого роста детенышей, в том числе много трудноперевариваемого казеина. Вроде бы на этом разница заканчивается.
«Корова, коза, любое другое млекопитающее от человека отличаются не только внешне, но и на молекулярном уровне. Названия белков в молоке одинаковые — казеин, лактоферрин, лизоцим, а их структура немного другая.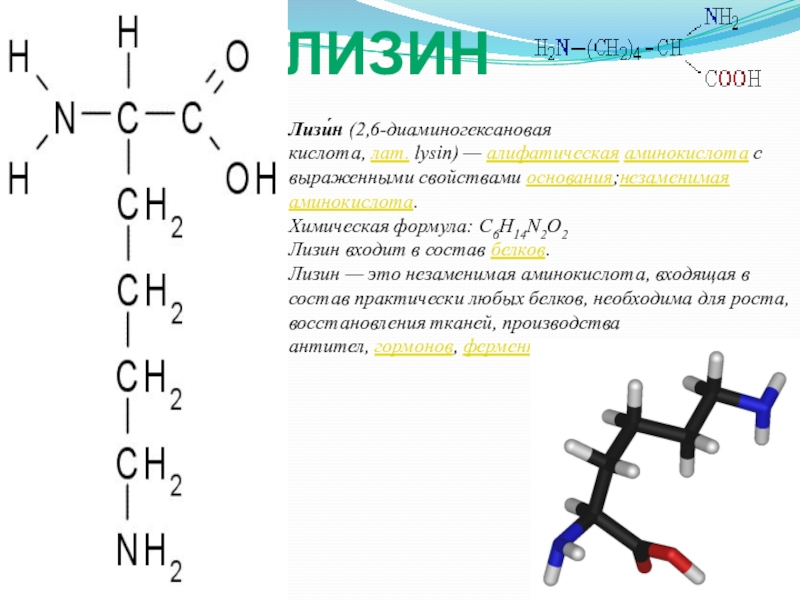 Когда мы используем такие белки при производстве продуктов питания для взрослых или в технических целях, например в качестве консерванта, это никакой роли не играет. Но при изготовлении продуктов для детей данный фактор решающий — именно с ним связаны многие аллергические реакции», — отмечает Алексей Дейкин, заместитель руководителя Центра коллективного пользования Института биологии гена РАН.
Когда мы используем такие белки при производстве продуктов питания для взрослых или в технических целях, например в качестве консерванта, это никакой роли не играет. Но при изготовлении продуктов для детей данный фактор решающий — именно с ним связаны многие аллергические реакции», — отмечает Алексей Дейкин, заместитель руководителя Центра коллективного пользования Института биологии гена РАН.
Смеси маму не заменят
Производители сухих смесей для детского питания стараются приблизить их к женскому молоку. Чтобы адаптировать для младенцев смеси из коровьего и козьего молока, в них снижают почти в два раза общий уровень белка. Благодаря этому азотистый и минеральный обмен грудного ребенка остается в норме, не страдают функции пищеварительного тракта и незрелых почек, сведен к минимуму риск развития ожирения и метаболического синдрома.
«Это обеспечивает безопасность продуктов животноводства и снижает аллергенность. Но когда мы уничтожаем в молочных смесях белки, они перестают выполнять защитную функцию — фактически ребенок лишается защиты со стороны иммунной системы матери. Молоко не способствует развитию кишечника, потому что самых ценных белков там нет», — объясняет Алексей Дейкин.
Молоко не способствует развитию кишечника, потому что самых ценных белков там нет», — объясняет Алексей Дейкин.
Группа российских и белорусских ученых, в которую входил и Алексей Дейкин, в 2008 году впервые в мире вывела генно-модифицированную козу, в чьем молоке содержался человеческий белок лактоферрин — природный антибиотик, убивающий инфекции, вирусы и грибки, но не затрагивающий микрофлору кишечника.
«Идея нашего проекта была в том, что можно добавить в искусственную смесь правильный, целый, полученный биотехнологическим способом белок человека. Да, это только один из множества белков, но он там будет и сможет защитить ребенка, пока не сформировался его собственный иммунитет», — рассказывает ученый.
Трансгенные козы и человеческие белки
Российским и белорусским исследователям тогда удалось создать рекомбинантную ДНК, содержащую кодирующую последовательность гена человека и регуляторные элементы, которые обеспечили работу этой конструкции в молочной железе животного. Затем эту ДНК встроили в ядро яйцеклетки и имплантировали козам.
Затем эту ДНК встроили в ядро яйцеклетки и имплантировали козам.
«Мы провели 113 операций по трансплантации яйцеклеток. Родились 33 козленка, два оказались трансгенными, то есть в их геном эта конструкция встроилась. Теперь у нас около пятидесяти таких животных. Но на этапе создания опытного производства мы вышли из проекта, потому что тут научная часть заканчивается. Дальше дело за инвесторами и промышленностью. Знаете, прошло уже десять лет после того, как мы создали козье молоко с человеческим лактоферрином, а промышленного производства таких смесей до сих пор нет, и мне это душу не греет», — качает головой ученый.
Сейчас в Институте биологии гена РАН при поддержке Российского научного фонда исследователи занимаются новой задачей — заставить животное производить молоко, в котором был бы только человеческий белок.
«У нашей трансгенной козы в молоке как человеческий, так и ее собственный лактоферрин. И их друг от друга очень сложно отделить, из-за чего увеличивается стоимость конечного продукта.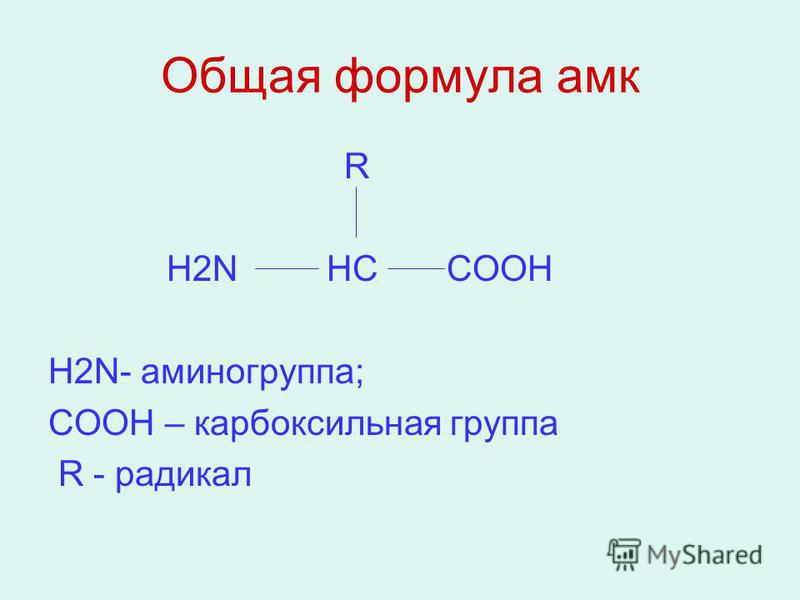 Мы сейчас подошли к созданию технологии, когда в молоке животного будет только человеческий белок. С помощью системы CRISPR/Cas9 мы хотим отключить собственный ген козы, чтобы его продукции в молоке не было. Пока экспериментируем на мышах: у них мы не просто выключили ген, а заменили его. В их молоке есть человеческий белок антитромбин — важный для борьбы с тромбозом сосудов. Следующий этап — кролики», — уточняет Алексей Дейкин.
Мы сейчас подошли к созданию технологии, когда в молоке животного будет только человеческий белок. С помощью системы CRISPR/Cas9 мы хотим отключить собственный ген козы, чтобы его продукции в молоке не было. Пока экспериментируем на мышах: у них мы не просто выключили ген, а заменили его. В их молоке есть человеческий белок антитромбин — важный для борьбы с тромбозом сосудов. Следующий этап — кролики», — уточняет Алексей Дейкин.
Альтернативы нет и не будет
Полностью воспроизвести состав женского грудного молока до сих пор не удалось никому в мире и вряд ли это получится когда-нибудь, отмечают эксперты. В основном научный интерес сфокусирован на получении отдельных белков женского молока, ведь они могут использоваться не только в сухих смесях для питания младенцев, но и при лечении самых разных заболеваний — от рака до ВИЧ. Так, ученые из Института химической биологии и фундаментальной медицины Сибирского отделения РАН разработали нетоксичное лекарство от рака на основе лактаптина — белка, входящего в состав женского молока.
Даже если в ближайшие годы на рынке появятся детские смеси, содержащие человеческие белки, нельзя отказываться от кормления грудью. Смеси — последнее средство, когда грудное вскармливание в силу обстоятельств невозможно. По словам Любови Абольян, кормление грудью — это не только защита ребенка, пока не сформировался его иммунитет, но и профилактика для мамы рака молочных желез и яичников.
Как подсчитали эксперты, ежегодно грудное вскармливание способно спасти около 820 тысяч детских жизней и примерно 20 тысяч материнских, сохранив мировой экономике около трехсот миллиардов долларов. В мае этого года на 71-й сессии Всемирной ассамблеи здравоохранения в Женеве принята резолюция по грудному вскармливанию. Резолюцию внесла Россия, хотя, как пишет The New York Times, США, поддерживающие интересы крупных американских производителей сухих молочных смесей, противились этой инициативе.
Согласно документу, страны — члены Всемирной организации здравоохранения обязаны продвигать идею кормления грудью на государственном уровне и сдерживать рекламу молочных смесей, если в ней говорится о полноценной замене женского молока.
РИА Новости https://ria.ru/science/20180801/1525689856.html
Белки как биополимеры. Химические функции белков. 10-й класс
Цель урока: расширить и углубить знания
учащихся о важнейших органических веществах
клетки – белках.
Задачи урока:
- Образовательные: обобщить знания
учащихся о биологических функциях белков,
строении, структуре, познакомить учащихся с
химическими функциями белков, раскрыть связь
между химическими знаниями и повседневной
жизнью человека, проконтролировать степень
усвоения основных умений и навыков, изученных и
сформированных на предыдущих уроках, а также
уроках биологии. - Воспитательные: продолжить формирование
научного мировоззрения, воспитывать культуру
речи. - Развивающие: развивать познавательный
интерес к предмету, такие процессы как внимание,
логическое мышление, эрудицию; навыки
самостоятельной работы с учебником, умение
анализировать информацию, устанавливать
причинно-следственные связи между составом,
строением, функций и применением веществ;
применять теоретические знания на практике;
навыки составления уравнений реакций
образования пептидов из аминокислот; навыки
работы с химическими веществами; продолжить
развитие химической речи учащихся, таких понятий
как полимер, альфа-аминокислота, денатурация,
реакция поликонденсации, расширить кругозор
учащихся с привлечением дополнительных
источников информации, продолжить формировать
умения и навыки использования знаний и умений в
новых ситуациях.
Тип урока: комбинированный
Оборудование:
- раствор яичного белка, р-ры сульфата меди (II),
хлорида цинка, гидрокисида натрия, этиловый
спирт, пробирки №1– яичный белок, мясной бульон,
молоко, №2 — бульонный кубик, медная проволока,
шерстяная нитка, спиртовка, спички;
дидактический материал (схема “Химический
состав организма”, “Общая формула
альфа-аминокислоты”, структурные формулы
аланина, серина), карточки-задания, компьютер,
телевизор. - На ученических столах: пробирки с веществами (№1
— яичный белок, мясной бульон, молоко, №2 -
бульонный кубик), медная проволока, шерстяная
нитка, спиртовка, спички; задания. - На классной доске: схема “Химический состав
организма”, “Общая формула
альфа-аминокислоты”, структурные формулы
аланина, серина.
Структура урока
I. Организационный момент
Организационный момент
II. Изучение нового материала
III. Закрепление
IV. Заключение
V. Домашнее задание
Ход урока
I. Организационный момент (1 мин)
Приветствие учителя. Проверка готовности
класса к уроку. Психологический настрой класса.
Отмечание отсутствующих.
II. Изучение нового материала (40мин)
1. Подведение к теме урока.
Вступительное слово учителя с элементами
беседы. Заполнение схемы на доске.
Организм состоит из множества веществ. Давайте
их классифицируем. Перед вами список различных
веществ: вода, белки, углеводы, минеральные соли,
жиры, нуклеиновые кислоты. Разделите их на группы
и дайте названия группам.
Заполнение схемы. (ученик у доски)
Откройте учебник параграф 27 стр.227 рис. 36, на
котором показан химический состав организма
человека. Какого вещества в организме больше?
Какого вещества в организме больше?
(воды – 65%).
Из органических веществ, образующих организм,
каких веществ больше? (белков)
Итак, основой живого организма является белки.
Сегодня на уроке мы рассмотрим белки, их состав и
строение, познакомимся с химическими функциями
белков. Для этого нам понадобятся знания из курса
биологии, а также ваш жизненный опыт.
Тема нашего урока: “Белки как биополимеры.
Химические функции белков”. (Слайд 1, Запись
даты и темы урока в тетради)
2. Биологические функции белков.
Беседа, выполнение задания в тетради.
Вспомним биологические функции белков.
Выполните письменно следующее задание. Возьмите
лист с заданиями. (Приложение 1)
Задание №1. (слайд 2)
Перечислены белки и их функции. Сопоставьте
функцию с белком с помощью стрелок. На выполнение
задание 1 минута. Будьте внимательны при
выполнении задания.
- Ферменты – … (каталитическая)
- Миозин – … (двигательная)
- Гемоглобин – … (транспортная)
- Коллаген, кератин – … (структурная)
- Инсулин — … (регуляторная)
- Альбумин -… (Запасающая)
- Тромбин — … (защитная)
- Гликоген — …—————-
Гликоген лишнее вещество в данном перечне, так
как не является белком. Это углевод.
А сейчас вам вопрос от учителя биологии Ирины
Аркадьевны (Слайд 3):
В науке известен следующий факт: влюблённый
студент-химик решил оригинальным способом
воспользоваться информацией о том, что в нашем
организме есть железо. Он решил из железа,
которое содержится в его крови сделать кольцо
для дамы сердца. Небольшими порциями он выпускал
кровь, химическим путём выделял железо. Но такой
романтический способ добычи металла закончился
трагически: он умер от малокровия.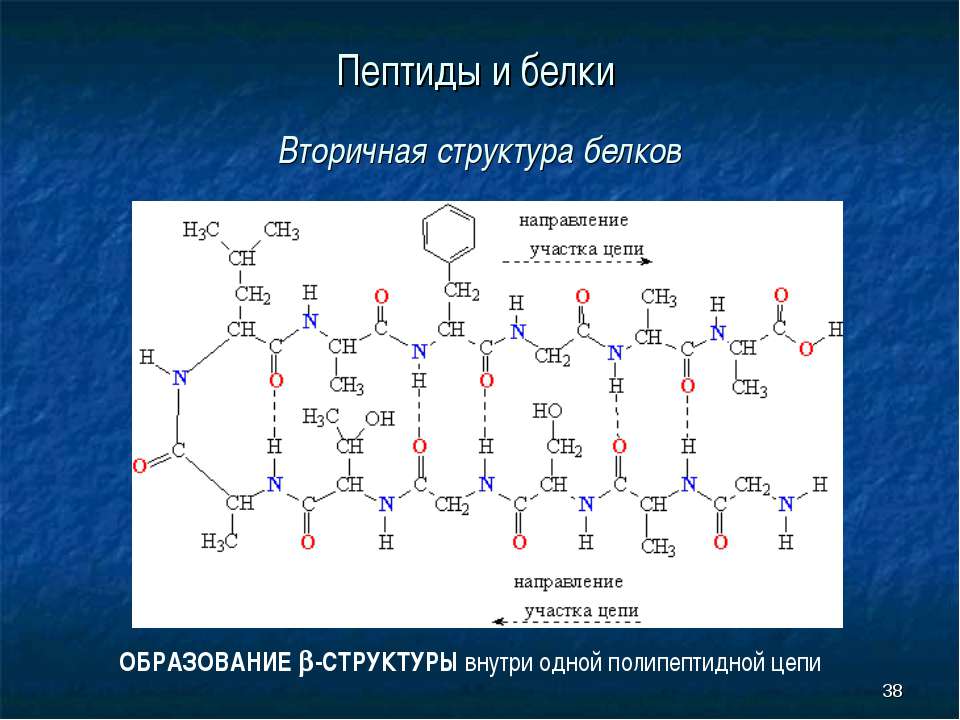 Ведь он не
Ведь он не
знал, что в нашей крови железа содержится
примерно от 3 до 4 грамм. В состав какого белка
входит железо, и какое значение он имеет для
организма? (железо входит в состав
белка-гемоглобина, который участвует в переносе
кислорода).
Посмотрите, насколько разнообразны функции
белков. С белками связаны все жизненные процессы.
Благодаря белкам организм приобрёл возможность
двигаться, усваивать пищу, расти, размножаться,
реагировать на внешние воздействия.
Итак, мы вспомнили некоторые биологические
функции белков. Переходим к следующему вопросу:
белок как химическое вещество.
3. Состав и строение белков.
Беседа с элементами объяснения, выполнение
упражнений.
Рассмотрим состав и строение белков.
Составьте определение белков из предложенных
слов (Слайд 4):
Белок, альфа — аминокислота, биополимер,
мономер. (Белок- это биополимер, мономером
которого является альфа — аминокислота).
— Какие химические элементы входят в состав
белков? (Углерод, водород, кислород, азот, а также
сера, фосфор и другие).
Составьте на доске формулу альфа –
аминокислоты из предложенных частей:
С, NH2, H, COOH, R.
(R – CHNH2 –COOH) (ученик у доски)
— Какие функциональные группы входят в состав
аминокислоты? (аминогруппа, карбоксильная
группа)
— Какими свойствами обладает аминокислота?
(амфотерными)
— Почему аминокислота – амфотерное соединение?
(аминогруппа определяет основные свойства,
карбоксогруппа – кислотные свойства)
— Сколькими аминокислотами образованы белки? (20)
— В результате какой реакции образуются белки?
(поликонденсации)
— Что собой представляет реакция
поликонденсации? (это реакция, в результате
которой образуется полимер, с отщеплением
побочного продукта)
Выполним следующее задание на доске и в
тетради:
Задание№2 (ученик у доски):
Составить уравнение реакции образования
дипептида из глицина, серина. Указать в нём
Указать в нём
пептидные связи.
Образующийся участок представляет собой
дипептидный участок белка-инсулина. Полимерные
цепи белков состоят из десятков тысяч, миллионов
и более остатков аминокислот. Перед вами формулы
некоторых белков (Слайд 5):
- С254Н377О75N65S6
инсулин - С3032Н4876О872N780S6Fe4
– гемоглобин - С1864Н3021О576N468S21 -
лактоглобулин (белок молока)
Как вы думаете, какова молекулярная масса
белков? (Очень большая). Например, молекулярная
масса инсулина – Mr 5727, гемоглобина 66184,
лактоглобулина (белка молока) – 39112.
Белковая цепь настолько длинная, что для
лучшего выполнения функций она упаковывается в
структуры.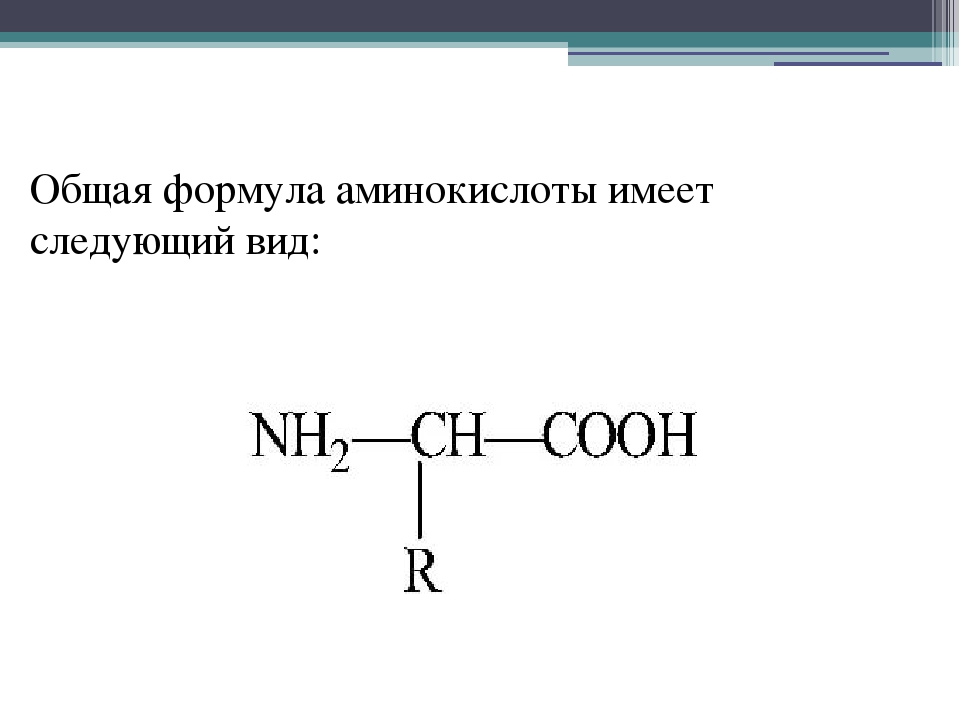
Рассмотрим структуры белков.
— Какие структуры белков вы знаете? (первичную,
вторичную, третичную, четвертичную)
Смоделируем структуры белка из проволоки,
которая лежит у вас на столе. Возьмите её.
— С какой структурой белка её можно сравнить?
(первичной)
— Что представляет собой первичная структура
белка? (чередование аминокислот в полипептидной
цепи)
Намотайте проволоку на ручку. С какой
структурой белка можно сравнить получившуюся
спираль? (вторичной)
— Что изменилось в молекуле? (уменьшился размер,
форма стала другой)
Из данной спирали сделайте комок. Какая
структура белка получилась? (третичная)
— Что представляет собой третичная структура?
(глобулу)
— Повернитесь друг к другу, объедините две
глобулы? Эта какая структура белка?
(четвертичная)
Выполняем следующее задание из карточки.
Задание №3. (Приложение 1)
На карточке представлены рисунки структур
белка. Снизу под цифрами перечислены типы связей,
Снизу под цифрами перечислены типы связей,
определяющих структуры. Определите структуру
белка и тип связей. Подпишите нужную цифру под
рисунком.
Проверим правильность выполнения задания. (Слайд
6)
— Какая структура самая прочная? (первичная)
4. Химические свойства белков.
Объяснение учителя с элементами беседы.
Выполнение демонстрационных и лабораторных
опытов. Запись на доске и в тетради
Начнём изучение свойств белков, связанных с их
структурой. Внимание на экран: вопрос от учителя
технологии Татьяны Леонидовны (Слайд 7):
Любая хозяйка знает, если нужно сварить вкусный
бульон для 1 блюда, мясо кладётся в холодную воду,
а когда вкусное мясо для 2 блюда — в горячее. Есть
ли в этом химический смысл?
— О каком свойстве белков идёт речь? (о
денатурации)
1) Денатурация (под действием нагревания,
химических веществ и т. д.)
д.)
— Что такое денатурация? (процесс утраты
белковой молекулой своей структуры при
изменении внешних факторов).
а) повышение температуры
Что вызывает денатурацию при варке мяса?
(нагревание, повышение температуры)
Итак, мы рассмотрели состав и строение белков.
Переходим к следующему вопросу.
Ответим на вопрос Татьяны Леонидовны.
— Почему для вкусного бульона мясо кладётся в
холодную воду, для вкусного мяса – в горячую?
(Если мясо положить в холодную воду, растворимые
белки переходят в воду и там денатурируются.
Бульон получается вкусным. Если мясо положить в
горячую воду, белки денатурируются сразу в мясе,
поэтому мясо получается вкусным)
– Какие факторы, кроме температуры, вызывают
денатурацию? (изменение температуры, облучением,
действием тяжёлых металлов, кислот, органических
веществ, обезвоживанием и другими воздействиями)
б) действие солей тяжёлых металлов (дем. опыт)
опыт)
Беру раствор яичного белка. Добавляю в один
стакан хлорид цинка, во второй сульфат меди (II).
— Что наблюдаем при этом? (свёртывание белков)
Белки связывают ионы тяжёлых металлов и
обезвреживают их. При отравлении тяжёлыми
металлами пострадавшему дают молоко как
противоядие.
в) действие органических веществ (дем.опыт)
К раствору яичного белка добавляю этиловый
спирт. Наблюдаем осаждение белков.
— Какие структуры белка разрушаются при
денатурации? (вторичная, третичная, четвертичная
с сохранением первичной). Утрачивается
биологическая активность. Белок становится
доступен действию пищеварительных ферментов.
— Какая бывает денатурация? (обратимая и
необратимая). Данная денатурация необратимая.
Может ли структура белка восстановиться? Обратим
ли процесс денатурации? (Да). Ренатурация –
процесс восстановления структуры белка.
Следующие химические свойства помогают
определить белки в растворах.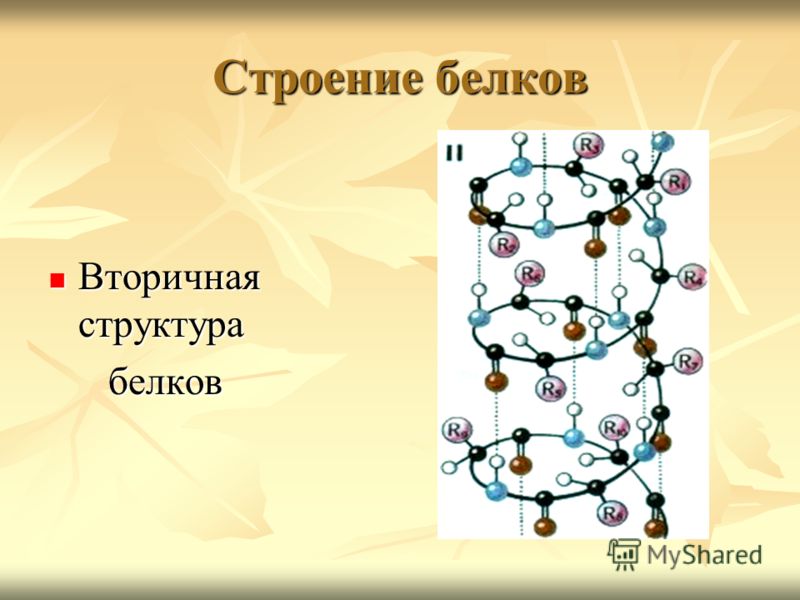
2) Цветные реакции
а) биуретовая (на пептидную связь)
Это универсальная реакция определения белков.
Посмотрим видеоопыт. (Слайд 8)
Заполняем схему в тетради.
Белок + ______________ = ____________ окрашивание
Белок + (щелочь+ CuSO4) = фиолетовое
окрашивание
Проведём экспертизу неизвестных веществ в
пробирках с помощью биуретовой реакции.
Приступайте к выполнению лабораторного опыта по
инструкции. (Слайд 9)
В какой пробирке находится белок? Посмотрите,
какие вещества находились у вас в пробирках (Слайд
10):
1 ряд
- Пробирка №1 – молоко
- Пробирка №2 – бульонный кубик
2 ряд
- Пробирка №1 – мясной бульон
- Пробирка №2 – бульонный кубик
3 ряд
- Пробирка №1 — яичный белок
- Пробирка №2 – бульонный кубик
Я выбрала бульонный кубик для определения
белков. Бульонный кубик белки содержит? (Нет). А в
Бульонный кубик белки содержит? (Нет). А в
его составе указаны растительные жиры и мясо
курицы.
б) ксантопротеиновая (на ароматические кольца) (видеофрагмент,
Слайд 11)
Заполняем схему в тетради:
Белок +__________= ___________окрашивание
белок + конц. HNO3 = жёлтое
окрашивание
При неосторожном обращении с азотной кислотой
от попадания её на кожу остаётся жёлтое пятно.
Это ксантопротеиновая реакция с покровными
тканями.
Эти качественные реакции можно применить в
жизни. А когда и где вам подскажет небольшой
видеофрагмент из фильма Аллы Суриковой “Ищите
женщину” (Слайд 12).
В каких случаях и с какой целью можно применить
их в жизни? (В пищевой промышленности, судебной
медицине для обнаружения белков)
Биуретовая и ксантопротеиновая реакции — это
качественные реакции, реакции, которые позволяют
уверенно судить – белок перед нами или нет.
3) Гидролиз
— Какой процесс называют гидролизом белков?
Вставьте пропущенные слова. (Слайд 13)
Гидролиз – это разрушение … структуры белка
под воздействием …, а также водных растворов
кислот или щелочей. (первичной, ферментов)
— Какие продукты образуются при гидролизе
белков? (аминокислоты)
Как можно изменить уравнение реакции
образования пептида, чтобы превратить её в
реакцию гидролиза? (Написать её наоборот)
Чтобы составить уравнение реакции гидролиза
дипептида глицилаланина добавить воду. Идёт
разрыв пептидной связи. Атом водорода
присоединяется к остатку от аминогруппы,
гидроксогруппа к остатку от карбоксильной
группы.
Общая схема гидролиза белков есть у вас в
учебнике.
Гидролиз – это основа пищеварения.
4) Горение (с образованием углекислого газа,
воды, азота.)
Запах: … .
Как проверить, что изделие из шерсти? (Шерсть –
это белок, надо сжечь кусочек нити и по запаху
определить результат)
Приступайте к выполнению лабораторного опыта
по инструкции. (Слайд 14)
Мы рассмотрели основные химические функции
белков.
III. Закрепление (3 мин.)
Фронтальный опрос.
Задания:
Перед вами на слайде представлены группы слов.
Обобщите несколько слов 1 словом или термином. (Слайд
15-23)
А) третичная, вторичная, первичная,
четвертичная — ? (структуры)
Б) 20, незаменимая – ? (аминокислота)
В) белки, жиры, углеводы -? (органические
вещества)
Г) температура, обратимая, необратимая -?
(денатурация)
Д) пепсин, амилаза, трипсин – ? (ферменты)
Е) Cu(OH)2, фиолетовое окрашивание — ?
(биуретовая реакция)
IV. Заключение (1 мин.)
Заключительное слово учителя.
Сегодня мы познакомились с химией белка. Ни
одно вещество химики не изучали так долго, как
белки, прежде чем удалось разгадать тайну их
строения. Они выполняют большую роль не только в
организме человека, но и в жизни. Не случайно в
переводе с греческого языка белки называют
протеинами, что значит “первый, главный”.
V. Домашнее задание (1 мин.)
Параграф 27
Приготовить сообщения на темы:
А) применение цветных реакций
Б) история изучения белков
В) факторы, вызывающие денатурацию.
Литература
1. Артёменко А.И. Удивительный мир органической
химии. — М.: Дрофа, 2004.
2. Горковенко М.Ю Поурочные разработки по химии
к учебным комплектам О.С.Габриеляна и др. 10 класс.
М. “ВАКО”, 2005.
3. Рябов М.А. Сборник заданий и упражнений по
химии: 10-й кл.: к учебнику О.С.Габриеляна и др.
“Химия. 10 класс”. – М.:Экзамен, 2008.
4. Химия 10 класс. Учеб. для общеобразоват.
учреждений/ О.С.Габриелян, Ф.Н.Маскаев,
С.Ю.Пономарёв, В.И.Теренин. – М.:Дрофа, 2010.
Белки (протеины)
Первичная структура белковых молекул
Под первичной структурой белков понимают последовательность аминокислотных остатков в полипептидной цепи.
Условно считают, что к белкам относятся полипептиды, содержащие в полипептидной цепи более 100 аминокислотных остатков и имеющие молекулярную миссу от 10 000 до нескольких миллионов.
Соединения с меньшим числом аминокислотных остатков называются пептидами, которые делятся на олигопептиды (от 2 до 10 остатков) и полипептиды (от 10 до 100 остатков).
Фрагмент первичной структуры:
Подсчитано, что из 20 различных аминокислот, которые чаще всего встречаются в составе белков, можно построить примерно 1018 белковых молекул.
Вторичная структура
В результате образования внутримолекулярных водородных связей между атомами водорода аминогрупп и атомами кислорода карбонильных групп полипептидные цепи многих белков скручиваются в спираль.
Третичная структура
Третичная структура определяет конфигурацию макромолекулы белка и представляет собой пространственную упаковку спирали, которая стабилизируется связями различного типа между отдельными участками полипептидных цепей.
Такими связями являются:
— дисульфидные -S-S-, возникающие между серусодержащими аминокислотными звеньями;
— сложноэфирные, образующиеся в результате взаимодействия карбоксильной группы в радикалах дикарбоновых кислот и гидроксильной группы в радикалах гидроксиаминокислот;
— солевые мостики между протонированной аминогруппой и ионизированной карбоксильной группой, которые содержатся в радикалах диамино- и дикарбоновых кислот: -NH3—… ООС-.
Четвертичная структура
Для некоторых белков характерно образование ассоциатов из нескольких полипептидных цепей, каждая из которых имеет определенную вторичную и третичную структуру.
Например, белок крови — гемоглобин — сложный белок (протеид), макромолекула которого состоит из четырех полипептидных цепей, соединенных с четырьмя небелковыми образованиями — гемами. В каждом геме содержится один атом железа, который может присоединять одну молекулу кислорода.
Эмпирическая формула гемоглобина:
(C738H1166O208N203S2Fe)4
Физические свойства
Природные белки выполняют в организме различные функции и в соответствии с этим обладают разными свойствами: существуют жидкие и твердые белки, растворимые и нерастворимые в воде.
Белки не имеют температуры плавления и температуры кипения, так как большинство из них при нагревании сворачиваются.
При высокой температуре все белки сгорают.
Физико-химические свойства
Как и аминокислоты, белки являются амфотерными соединениями, так как молекула любого белка содержит на одном конце группу -NH2, а на другом конце — группу -СООН. Большее или меньшее число таких групп содержится также в радикалах диамино- и дикарбоновых кислот. Поэтому свойства белков во многом похожи на свойства аминокислот. Однако белки обладают и специфическими физико-химическими свойствами.
Переваривание белков в организме по своей сути представляет ферментативный гидролиз белковых молекул. В лабораторных условиях и в промышленности проводится кислотный гидролиз. В любом случае в ходе гидролиза белков происходит разрушение пептидных связей. Гидролиз белка имеет ступенчатый характер:
Белки способны осаждаться из растворов под действием таких веществ, как этиловый спирт, ацетон, соли Na+, К+, NH4+ и др. Свойства белков при таком осаждении не меняются, поэтому при добавлении воды белки снова могут переходить в раствор.
При действии таких реагентов, как соли тяжелых металлов (Hg2+, Pb2+, Сu2+ и др.), концентрированные кислоты и щелочи,а также в результате нагревания, облучения УФ- и γ-лучами происходит разрушение (полное или частичное) вторичной и третичной структуры белка и изменение его природных (нативных) свойств.
Пример тепловой денатурации — свертывание яичного белка альбумина при варке яиц.
а) Биуретовая реакция — реакция на пептидные связи
Причина появления окраски — образование комплексных соединений с координационным узлом:
б) Ксантопротеиновая реакция (реакция на радикалы ароматических аминокислот)
Причина появления окраски — образование нитропроизводных ароматических аминокислот, например, фенилаланина.
в) Цистеиновая реакция (реакция на остатки S-содержащих аминокислот)
Причина появления окраски — образование
черного осадка сульфида свинца (II) PbS.
Составлена детальная карта генома нового коронавируса
https://ria.ru/20200410/1569854665.html
Составлена детальная карта генома нового коронавируса
Составлена детальная карта генома нового коронавируса — РИА Новости, 10.04.2020
Составлена детальная карта генома нового коронавируса
Корейские микробиологи составили первую генную карту нового коронавируса. Оказалось, что геном SARS-CoV-2 содержит множество так называемых субгеномных РНК,… РИА Новости, 10.04.2020
2020-04-10T13:21
2020-04-10T13:21
2020-04-10T13:53
наука
южная корея
открытия — риа наука
здоровье
биология
коронавирус covid-19
/html/head/meta[@name=’og:title’]/@content
/html/head/meta[@name=’og:description’]/@content
https://cdnn21.img.ria.ru/images/07e4/03/1b/1569221900_0:56:1440:866_1920x0_80_0_0_fbbfc0ca31ce38e52946048f6e21416a.jpg
МОСКВА, 10 апр — РИА Новости. Корейские микробиологи составили первую генную карту нового коронавируса. Оказалось, что геном SARS-CoV-2 содержит множество так называемых субгеномных РНК, функции которых не до конца понятны. Но многое ученым удалось выяснить. Результаты исследования опубликованы в журнале Cell.Коронавирус SARS-CoV-2 относится к группе РНК-вирусов, довольно сложный геном которых зашифрован в очень длинной молекуле рибонуклеиновой кислоты (РНК). Проникая в клетки-хозяева, вирус реплицирует геномную РНК и создает множество более мелких, называемых субгеномными. Эти субгеномные РНК используются для синтеза различных белков, из которых строятся элементы новых вирусных частиц: шипов, оболочек, мембран. Таким образом, если ученые найдут способ подавить субгеномные РНК, то это нарушит жизненный цикл вируса в организме. Но сперва надо понять, за что отвечает каждая из этих загадочных цепочек.Биологам из Центра исследований РНК Института фундаментальных наук Южной Кореи (IBS) под руководством профессоров Ким Нарри (Kim V. Narry) и Чан Хьешик (Chang Hyeshik) в сотрудничестве с Корейским национальным институтом здоровья (KNIH) удалось детально проанализировать архитектуру генома РНК SARS-CoV-2 и составить его генную карту высокой степени детализации.Исследователи экспериментально подтвердили наличие в геноме коронавируса девяти из десяти ранее известных субгеномных РНК (входящих в структуру вирусных частиц и транслирующихся в конкретные вирусные белки), а также обнаружили десятки неизвестных, образующихся на разных этапах жизненного цикла вируса в результате слияния и разложения. Кроме того, они выяснили, где именно находятся эти гены на геномной РНК.»Это не просто детализация структуры SARS-CoV-2, — приводятся в пресс-релизе IBS слова профессора Ким Нарри. — Мы обнаружили многочисленные новые РНК и множественные неизвестные химические модификации вирусных РНК».Авторы предполагают, что модифицированные в ходе жизненного цикла РНК могут получить новые свойства, отличающие их от немодифицированных, даже если они несут одинаковую генетическую информацию. По мнению исследователей, выявление неизвестных характеристик РНК позволит подобрать ключ для борьбы с новым коронавирусом.»Хотя требуется дальнейшее изучение, уже можно сказать, что подобные молекулярные события могут привести к относительно быстрой эволюции коронавируса. Более того, мы находим множество неизвестных химических модификаций вирусных РНК. Пока неясно, что делают эти модификации, но возможно, что они помогают вирусу избежать атаки со стороны клетки-хозяина», — говорит Ким.Успех корейских микробиологов основан на применении двух дополнительных методов: секвенирования кольцевых молекул ДНК (наноболов) и прямого нанопорового секвенирования РНК.Обычные методы секвенирования требуют пошагового процесса разрезания и преобразования РНК в ДНК перед ее считыванием. Нанопоровое секвенирование позволяет напрямую анализировать всю длинную вирусную РНК без фрагментации, а секвенирование кольцевых молекул ДНК имеет преимущество анализа большого количества последовательностей с высокой точностью. Эти два взаимодополняющих метода оказались очень эффективными для анализа вирусных РНК.»Теперь мы получили генную карту нового коронавируса с высоким разрешением, которая поможет нам найти каждый бит генов на всех РНК SARS-CoV-2 и всех модификациях РНК. Настало время исследовать функции вновь открытых генов и механизм, лежащий в основе слияния вирусных генов. Мы также должны работать над модификациями РНК, чтобы увидеть, какую роль они играют в репликации вируса и иммунном ответе. Мы твердо верим, что наше исследование будет способствовать прогрессу в диагностике и терапии для более эффективной борьбы с вирусом», — отмечает Ким.
https://ria.ru/20200408/1569743831.html
южная корея
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
2020
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
Новости
ru-RU
https://ria.ru/docs/about/copyright.html
https://xn--c1acbl2abdlkab1og.xn--p1ai/
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
https://cdnn21.img.ria.ru/images/07e4/03/1b/1569221900_106:0:1335:922_1920x0_80_0_0_4d9aa5355bb4861b1706a1cd850ca90f.jpg
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
РИА Новости
7 495 645-6601
ФГУП МИА «Россия сегодня»
https://xn--c1acbl2abdlkab1og.xn--p1ai/awards/
южная корея, открытия — риа наука, здоровье, биология, коронавирус covid-19
МОСКВА, 10 апр — РИА Новости. Корейские микробиологи составили первую генную карту нового коронавируса. Оказалось, что геном SARS-CoV-2 содержит множество так называемых субгеномных РНК, функции которых не до конца понятны. Но многое ученым удалось выяснить. Результаты исследования опубликованы в журнале Cell.
Коронавирус SARS-CoV-2 относится к группе РНК-вирусов, довольно сложный геном которых зашифрован в очень длинной молекуле рибонуклеиновой кислоты (РНК). Проникая в клетки-хозяева, вирус реплицирует геномную РНК и создает множество более мелких, называемых субгеномными. Эти субгеномные РНК используются для синтеза различных белков, из которых строятся элементы новых вирусных частиц: шипов, оболочек, мембран. Таким образом, если ученые найдут способ подавить субгеномные РНК, то это нарушит жизненный цикл вируса в организме. Но сперва надо понять, за что отвечает каждая из этих загадочных цепочек.
Биологам из Центра исследований РНК Института фундаментальных наук Южной Кореи (IBS) под руководством профессоров Ким Нарри (Kim V. Narry) и Чан Хьешик (Chang Hyeshik) в сотрудничестве с Корейским национальным институтом здоровья (KNIH) удалось детально проанализировать архитектуру генома РНК SARS-CoV-2 и составить его генную карту высокой степени детализации.
Исследователи экспериментально подтвердили наличие в геноме коронавируса девяти из десяти ранее известных субгеномных РНК (входящих в структуру вирусных частиц и транслирующихся в конкретные вирусные белки), а также обнаружили десятки неизвестных, образующихся на разных этапах жизненного цикла вируса в результате слияния и разложения. Кроме того, они выяснили, где именно находятся эти гены на геномной РНК.
«Это не просто детализация структуры SARS-CoV-2, — приводятся в пресс-релизе IBS слова профессора Ким Нарри. — Мы обнаружили многочисленные новые РНК и множественные неизвестные химические модификации вирусных РНК».
Авторы предполагают, что модифицированные в ходе жизненного цикла РНК могут получить новые свойства, отличающие их от немодифицированных, даже если они несут одинаковую генетическую информацию. По мнению исследователей, выявление неизвестных характеристик РНК позволит подобрать ключ для борьбы с новым коронавирусом.
«Хотя требуется дальнейшее изучение, уже можно сказать, что подобные молекулярные события могут привести к относительно быстрой эволюции коронавируса. Более того, мы находим множество неизвестных химических модификаций вирусных РНК. Пока неясно, что делают эти модификации, но возможно, что они помогают вирусу избежать атаки со стороны клетки-хозяина», — говорит Ким.
Успех корейских микробиологов основан на применении двух дополнительных методов: секвенирования кольцевых молекул ДНК (наноболов) и прямого нанопорового секвенирования РНК.
Обычные методы секвенирования требуют пошагового процесса разрезания и преобразования РНК в ДНК перед ее считыванием. Нанопоровое секвенирование позволяет напрямую анализировать всю длинную вирусную РНК без фрагментации, а секвенирование кольцевых молекул ДНК имеет преимущество анализа большого количества последовательностей с высокой точностью. Эти два взаимодополняющих метода оказались очень эффективными для анализа вирусных РНК.
«Теперь мы получили генную карту нового коронавируса с высоким разрешением, которая поможет нам найти каждый бит генов на всех РНК SARS-CoV-2 и всех модификациях РНК. Настало время исследовать функции вновь открытых генов и механизм, лежащий в основе слияния вирусных генов. Мы также должны работать над модификациями РНК, чтобы увидеть, какую роль они играют в репликации вируса и иммунном ответе. Мы твердо верим, что наше исследование будет способствовать прогрессу в диагностике и терапии для более эффективной борьбы с вирусом», — отмечает Ким.
8 апреля 2020, 13:09НаукаСоздан искусственный вирус, блокирующий коронавирусные инфекции
1.1.3. Химический состав дрожжей — Технология производства дрожжей
Состав
дрожжей
зависит от условий культивирования дрожжей,
состава питательной среды и физиологического состояния клетки. В прессованных
дрожжах содержится 67-75% воды и 25-33% сухого вещества. При этом часть воды
находится в межклеточных пространствах и называется внеклеточной; остальная
вода, находящаяся в цитоплазме дрожжей, называется внутриклеточной. Соотношение
клеточной и внутриклеточной влаги в дрожжах может изменяться в зависимости от
применяемой расы дрожжей,
технологического режима их выращивания и способа ведения технологического
процесса. Так, при выращивании дрожжей в концентрированной среде или с
добавлением осмотически активных веществ, например хлористого натрия, общее
количество влаги в дрожжах снижается в результате уменьшения внутриклеточной
воды, а при обработке дрожжей хлористым натрием (при выделении) общее количество
влаги в дрожжах снижается вследствие внеклеточной воды.
Состав
сухого вещества хлебопекарных дрожжей по элементам следующий (в %): углерод
45-49; водород — 50-70; кислород 30-35; азот 7,1-10,8; фосфор 1,9-5,5; калий
1,4-4,3; магний 0,1-0,7; алюминий 0,002-0,020; сера 0,01-0,05; хлор 0,004-0,100;
железо 0,005-0,012; кремний 0,02-0,20.
Кроме того, в сухом веществе дрожжей содержатся (в %): белки
и другие азотистые вещества — 50; жиры
— 1,6; углеводы
— 40,8; зола — 7,6. Однако этот состав непостоянен и может колебаться в широких
пределах.
Белки
состоят из полипептидов
и аминокислот — простых соединений, имеющих с одной стороны своей молекулы
аминогруппу NH, а с другой — кислотную группу СООН. Самая простая аминокислота —
глицин — имеет следующую формулу: NH-СН3-СООН. Соединяясь между собой,
аминокислоты образуют молекулы простых белков
или протеинов. К ним относятся альбумины, глобулины, гистоны и др. При
присоединении к простому белку небелковой группы образуются сложные белки,
или протеиды. Если небелковая группа состоит из нуклеиновых кислот, от
образовавшийся сложный белок называют нуклеопротеидом, а если к простому белку
присоединяются жиры, то сложный белок называют липопротеидом. Протеиды
осуществляют в клетке ряд сложнейших реакций, которые называют обменом веществ,
— размножение, питание, дыхание, передачу наследственных признаков, регулируют
поступление питательных веществ внутрь клетки и выделение продуктов обмена во
внешнюю среду. Белки
весьма чувствительны к воздействию факторов внешней среды. Например, при
воздействии либо очень высокой, либо очень низкой температуры происходит
свертывание белка или его денатурация, в результате чего клетка отмирает. Такое
же явление наблюдается при действии кислот, щелочей, солей тяжелых металлов,
излучения и др.
Углеводы
состоят
из углерода, кислорода и водорода. Их делят на высшие и низшие. К высшим
углеводам относят полисахариды (крахмал;
гликоген, клетчатка),
а также дисахариды (сахароза, лактоза, мальтоза, галактоза). К низшим углеводам
относят моносахариды (глюкоза, фруктоза, арабиноза, ксилоза и др.). При
разложении полисахаридов, например крахмала,
сначала образуются декстрины, затем дисахариды и моносахариды, а начало синтеза
полисахаридов начинается с моносахаров. Гликоген, или животный крахмал, является
запасным веществом в животном организме и дрожжах, как крахмал
в растениях. Из углеводов
клетка получает энергию.
Жиры
—
запасные вещества клетки. Они являются сложными эфирами трехатомного спирта
(глицерина и органических кислот).
Жировые вещества дрожжей являются важной частью протоплазмы клеток.
Молекулы жировых веществ соединяются в крупные частицы (жировые мицеллы)
палочковидной формы, распределяющиеся между мицеллами протеинов. Иногда они
образуют с последними сложные соединения (липопротеиды) и представляют собой
основной структурный материал клетки. Жиры
превращаются клеткой по мере необходимости в углеводы
и используются затем для получения энергии. Помимо связанных жировых веществ в
протоплазме некоторых дрожжевых грибов имеются и свободные жировые вещества,
обособленные в виде капель, хорошо окрашивающихся специальными
красками.
Зола
дрожжей
составляет около 6,5-12,0% общей массы сухого вещества дрожжей. Состав золы
колеблется в зависимости от условий их культивирования. Зола дрожжей состоит
примерно наполовину из фосфора: большая часть фосфорной кислоты связана в
дрожжах с органическими соединениями. В золе значительно больше калия, чем
натрия, кальция и магния. Содержание серы в хлебопекарных
дрожжах составляет 0,17-0,20%. Минеральные вещества золы дрожжей,
растворяясь в межмицеллярной воде, играют большую роль в обмене веществ клетки.
Наиболее важное значение имеют катионы натрия, калия, кальция, магния, железа,
анионы хлора, фосфора.
Витамины.
В дрожжах
содержится целый ряд витаминов и витаминоподобных веществ. Обмен веществ у
животных и человека, осуществляемый ферментами, протекает при непременном
участии витаминов, тесно связанных с ферментными системами клетки. Так, витамин
B1 содержится в хлебопекарных дрожжах в количестве около 20 мкг на 1 г СВ.
Витамин B1 регулирует деятельность нервной системы человеческого организма,
участвует в обмене белковых веществ и в синтезе жиров, излечивает полиневриты и
различные очень тяжелые нервные заболевания, возникающие при длительном
употреблении пищи, лишенной должного количества этого
витамина.
Витамин
В2 (рибофлавин) содержится в хлебопекарных
дрожжах в количестве около 25-30 мкг на 1 г СВ. Отсутствие рибофлавина в
пище человека приводит к различным поражениям кожного покрова, к расстройствам
зрения.
Витамины
B1 и В2 достаточно устойчивы к высоким температурам, особенно витамин В2,
который может быть отделен от витамина B1 путем шестичасового автоклавирования
при 120 °С; при этом витамин В2 остается без изменения, а витамин B1
разрушается.
Витамин В3 (пантотеновая кислота) в большом количестве содержится в хлебопекарных
дрожжах (15 000-33 000 мг/г СВ). Недостаток его в пищевом рационе животных и
птиц приостанавливает нормальный рост их и нарушает нормальную деятельность
нервной системы и желез внутренней секреции.
Витамин
В5 (РР — никотинамид) является собственно антипеллагрическим фактором; он
содержится в хлебопекарных дрожжах в большом количестве (от 185 до 290 мкг на 1
г СВ).
Витамин
В6 (пиридоксин) содержится в хлебопекарных дрожжах в количестве 1,6-6,5 на 1 г
СВ. Он стимулирует рост животных и микроорганизмов.
Витамин D — антирахитический фактор, регулятор фосфорно-кальциевого
обмена животных и человека. Провитамин D — эргостерин — имеется в огромном
количестве в хлебопекарных
дрожжах — 20 000 мкг на 1 г СВ.
Помимо
перечисленных витаминов хлебопекарные дрожжи
содержат парааминобензойную кислоту в
количестве 8-95 мкг на 1 г CB и фолиевую кислоту 19-35 мкг. Парааминобензойная кислота действует как
активный витамин самостоятельно и в виде составной части фолиевой кислоты. Эти
кислоты входят в состав ферментов,
катализирующих синтез нуклеиновых оснований. Большое значение для
жизнедеятельности дрожжей имеет витамин Вн, или биотин. Сахаромицеты не способны
синтезировать биотин из окружающей среды, поэтому для нормального их развития
биотин должен входить в состав питательной среды, где культивируются дрожжи как
важнейший фактор роста. Содержание этого витамина составляет 0,5-1,8 мкг на 1 г
СВ. Биотин — устойчивое вещество. При
термической обработке, доступе кислорода и воздействии разбавленных кислот и
щелочей биологическая активность его не снижается. Расщепление биотина
происходит лишь при обработке его концентрированными кислотами, щелочами и
раствором перекиси водорода. В дрожжах содержится и другой стимулятор роста
дрожжей — мезоинозит. В хлебопекарных
дрожжах он содержится в количестве 270 мг на 1 г СВ. Состав среды может
способствовать повышению содержания витаминов в дрожжевых клетках. Можно
обогащать хлебопекарные дрожжи витаминами группы В, помещая их в условия брожения
на 1-2 ч в среды, содержащие витамины.
Дрожжи способны поглощать витамин В1, находящийся в бродящей жидкости. В
этом случае общее количество витамина B1 может достигать 2000 мкг на 1 г СВ;
если бродящая жидкость содержит не витамин В1, а его компоненты (пирамидин и
тиазол), дрожжи
способны синтезировать витамин B1; количество его в дрожжах при этом может
достигать 600 мкг на 1 г СВ.
Ферменты.
Все процессы, происходящие в живых организмах при обмене веществ, при росте и
развитии организмов, совершаются с участием биологических катализаторов белковой
природы, ферментов или энзимов. Сущность механизма действия ферментов
заключается в том, что субстрат, на который действует фермент, образует с ним
непрочный продукт фермент
— субстратный комплекс. Промежуточный продукт разлагается с образованием
конечных продуктов и освобождением фермента, который может воздействовать на
новую молекулу субстрата. Считается, что активность фермента зависит не только
от таких факторов, как температура и реакция среды (рН), но и от того, в каком
виде он находится в клетке. Когда фермент находится в свободном состоянии, он
активен, когда же он связан с белками протоплазмы клетки, то активность его
уменьшается или теряется совсем. Синтез ферментов происходит в дрожжевой клетке
непрерывно. По способу образования ферменты делят обычно на конститутивные и
адаптивные. Адаптивными, т. е.
приспособительными, ферментами называют такие, которые образуются в клетке в
результате появления в среде соответствующего субстрата, например сахара.
Фермент мальтаза формируется в клетке при наличии в среде сахара мальтозы. Конститутивные ферменты образуются в
клетке организма независимо от состава среды. Наибольшую активность ферменты
проявляют при определенной температуре, кислотности, а также при отсутствии
тормозящих их действие веществ. Неустойчивость ферментов объясняется их белковой
природой, т. е. они чувствительны, как все белки,
к высоким температурам, кислотности, к солям тяжелых металлов, что вызывает их
денатурацию. Специфичность действия ферментов состоит в том, что один фермент
ускоряет только определенную реакцию, поэтому в микробных клетках действуют
одновременно десятки различных ферментов, не мешая один другому. Например, фермент,
разлагающий сахарозу, не может разлагать белки, жиры или другие вещества.
Отдельные ферменты в живых клетках образуют ферментные системы, состоящие из
10-12 ферментов.
Питание.
В настоящее время известно, что питание дрожжевых клеток состоит из двух фаз:
первая — прохождение веществ через клеточную стенку и цитоплазматическую
мембрану и вторая — сложные биохимические реакции, состоящие из взаимосвязанных
процессов ассимиляции и диссимиляции. Основным барьером, отделяющим внутреннее
содержимое клетки от окружающей среды, является цитоплазматическая мембрана,
основная функция которой заключается в регулировании прохождения в клетку
молекулярных растворов.
По
химическому составу дрожжей видно, что для питания им нужны азот, фосфор, калий,
магний, усвояемые формы углеводов,
микроэлементы и другие вещества. Источниками углерода для дрожжей являются
различные углеводы,
моно- и дисахара, а также спирты,
альдегиды и органические кислоты. При отсутствии аэрации дрожжи
используют обычно лишь сахара. В условиях аэрации при обогащении среды
кислородом, когда усиливается дыхательная функция дрожжей и активируется процесс
накопления биомассы, дрожжи усваивают не только сахара, но и спирты (этиловый
спирт, глицерин, маннит), альдегиды, а также и органические кислоты (молочная,
уксусная, лимонная и яблочная кислоты) и их соли. Доказано, что и аминокислоты
являются для дрожжей источником углерода. Источником азотистого питания для
живых клеток являются растворимые соединения азота (органические и
неорганические). Сложные высокомолекулярные протеины не усваиваются дрожжами,
так как у сахаромицетов не содержится экзоферментов, протеолизирующих сложные
белки среды. Продукты распада белков могут усваиваться дрожжами. Легко
усваиваются аминокислоты, а также амиды и аммонийные соединения. Нитраты не
усваиваются большинством дрожжевых грибов. Аммиак является первоисточником для
синтеза белковых веществ клетки. Аммиачный азот, отщепленный от аммонийных солей
или аминокислот среды и других азотистых соединений, используется дрожжевыми
клетками для синтеза собственных аминокислот. Большую роль в питании дрожжей
играют макроэлементы (калий, натрий, фосфор, магний, кальций) и микроэлементы
(железо, медь, марганец, кобальт, цинк, молибден, никель, кремний, алюминий,
бор).
Аминокислоты — номенклатура, получение, химические свойства. Белки » HimEge.ru
Строение аминокислот
Аминокислоты — гетерофункциональные соединения, которые обязательно содержат две функциональные группы: аминогруппу — NH2 и карбоксильную группу —СООН, связанные с углеводородным радикалом.Общую формулу простейших аминокислот можно записать так:
Так как аминокислоты содержат две различные функциональные группы, которые оказывают влияние друг на друга, характерные реакции отличаются от характерных реакций карбоновых кислот и аминов.
Свойства аминокислот
Аминогруппа — NH2 определяет основные свойства аминокислот, т. к. способна присоединять к себе катион водорода по донорно-акцепторному механизму за счет наличия свободной электронной пары у атома азота.
Группа —СООН (карбоксильная группа) определяет кислотные свойства этих соединений. Следовательно, аминокислоты — это амфотерные органические соединения. Со щелочами они реагируют как кислоты:
С сильными кислотами- как основания-амины:
Кроме того, аминогруппа в аминокислоте вступает во взаимодействие с входящей в ее состав карбоксильной группой, образуя внутреннюю соль:
Ионизация молекул аминокислот зависит от кислотного или щелочного характера среды:
Так как аминокислоты в водных растворах ведут себя как типичные амфотерные соединения, то в живых организмах они играют роль буферных веществ, поддерживающих определенную концентрацию ионов водорода.
Аминокислоты представляют собой бесцветные кристаллические вещества, плавящиеся с разложением при температуре выше 200 °С. Они растворимы в воде и нерастворимы в эфире. В зависимости от радикала R— они могут быть сладкими, горькими или безвкусными.
Аминокислоты подразделяют на природные (обнаруженные в живых организмах) и синтетические. Среди природных аминокислот (около 150) выделяют протеиногенные аминокислоты (около 20), которые входят в состав белков. Они представляют собой L-формы. Примерно половина из этих аминокислот относятся к незаменимым, т. к. они не синтезируются в организме человека. Незаменимыми являются такие кислоты, как валин, лейцин, изолейцин, фенилаланин, лизин, треонин, цистеин, метионин, гистидин, триптофан. В организм человека данные вещества поступают с пищей. Если их количество в пище будет недостаточным, нормальное развитие и функционирование организма человека нарушаются. При отдельных заболеваниях организм не в состоянии синтезировать и некоторые другие аминокислоты. Так, при фенилкетонурии не синтезируется тирозин. Важнейшим свойством аминокислот является способность вступать в молекулярную конденсацию с выделением воды и образованием амидной группировки —NH—СО—, например:
Получаемые в результате такой реакции высокомолекулярные соединения содержат большое число амидных фрагментов и поэтому получили название полимамидов.
К ним, кроме названного выше синтетического волокна капрона, относят, например, и энант, образующийся при поликонденсации аминоэнантовой кислоты. Для получения синтетических волокон пригодны аминокислоты с расположением амино- и карбоксильной групп на концах молекул.
Полиамиды альфа-аминокислот называются пептидами. В зависимости от числа остатков аминокислот различают дипептиды, трипептиды, полипептиды. В таких соединениях группы —NH—СО— называют пептидными.
Изомерия и номенклатура аминокислот
Изомерия аминокислот определяется различным строением углеродной цепи и положением аминогруппы, например:
Широко распространены также названия аминокислот, в которых положение аминогруппы обозначается буквами греческого алфавита: α, β, у и т. д. Так, 2-аминобутановую кислоту можно назвать также α-аминокислотой:
Способы получения аминокислот
В биосинтезе белка в живых организмах участвуют 20 аминокислот.
Более подробно про белки.
Что такое глутамат натрия? – «Еда»
Сергей Белков, химик-технолог, флейворист, блогер:
«Глутамат натрия — это соль
глутаминовой кислоты, одной из аминокислот, из которой построены белки. Она
есть в любом белке без исключения, без нее белка не существует в принципе. Более
того, она является, пожалуй, самой распространенной среди всех двадцати
аминокислот. В некоторых растительных белках ее количество может доходить до
20–40 процентов. В организме она выполняет огромное количество функций, можно
сказать, что организм без нее в принципе не может жить. Но, к счастью, это заменимая
аминокислота: то есть если в пище ее не хватает, то организм ее спокойно
синтезирует, а если в пище избыток — спокойно утилизирует. Отказаться от пищи с
глутаминовой кислотой невозможно — тогда придется полностью отказаться от белка.
Вся шумная история с
глутаматом возникла из-за того, что глутаминовая кислота, помимо того что несет
важные биологические функции, еще и обладает вкусом — вкусом умами. Это один из пяти
базовых вкусов, для которого на языке существуют свои вкусовые рецепторы. Молекулы
глутамата — носители этого вкуса, так же как поваренная соль, хлорид натрия (точнее, ионы натрия в ней), — носитель
соленого вкуса, кислоты в виде ионов водорода — носители кислого вкуса, сахар — сладкого,
а многие
алкалоиды — горького вкуса.
Так сложилось в процессе
эволюции: глутаминовая кислота — это, можно сказать, маркер белка. Если в пище
есть белок — есть, как правило, определенное количество этой аминокислоты, соответственно, распознавание
умами — способ, которым организм находит богатую белком пищу. Именно поэтому
нам этот вкус приятен, что и использует пищевая промышленность.
Но не только
промышленность: большинство методов кулинарной обработки — это фактически
производство глутамата из белка: тушение, жарка, варка. Белок в присутствии кислоты под действием высокой температуры или под действие ферментов, содержащихся в пище или привносимых микроорганизмами, взаимодействует с водой. Эта реакция по сути аналогична той, что происходит с белком в пищеварительном тракте, и неминуемо приводит к распаду его длинной молекулы на отдельные фрагменты или даже отдельные аминокислоты. Они — и прежде всего глутаминовая кислота — как раз и придают пище приятный вкус.
Если говорить о ферментации
— то это и есть производство глутамата: соевые соусы, рыбные соусы, сыры, особенно твердые. Иногда используется и природный источник глутамата, например если мы говорим о кетчупе. Это фактически глутамат из помидоров, вот почему
он такой вкусный. Кулинары нащупали вкус умами случайно, по вкусу, а химия реализовала
эти интуитивные находки в виде вещества, которое можно добавлять в продукты».
(PDF) Все белки имеют основную молекулярную формулу
была определена родственная молекулярная формула. Общее количество
атомов водорода, азота, кислорода и серы было тогда
, разделенное на общее количество атомов углерода. Таким образом были получены коэффициенты
для углерода, водорода, азота, кислорода и
серы. Эти коэффициенты указывают количество
атомов водорода, азота, кислорода и серы в структуре белка
по отношению к количеству атомов углерода.Этот
дает возможность составить базовую формулу для каждого образца белка
. Таблица I представляет собой пример полученной формулы для фермента оксидоредуктазы
.
ТАБЛИЦА I
ПРИМЕР ПОЛУЧЕННОЙ ОСНОВНОЙ ФОРМУЛЫ ДЛЯ ОКСИДОРЕДУКТАЗЫ
ФЕРМЕНТОВ
Ш. РЕЗУЛЬТАТЫ
Полученные коэффициенты позволяют извлечь базовую формулу
для каждого выбранного белка. Наконец, формула базовой единицы
была получена для каждой классификации белков (например,g.,
ферментов) путем определения среднего из коэффициентов. Результаты
показаны в таблице Π.
ТАБЛИЦА №
ОКОНЧАТЕЛЬНАЯ СРЕДНЯЯ ФОРМУЛА ДЛЯ КАЖДОГО ИЗ 9 БЕЛКОВ
КЛАССИФИКАЦИЯ
Белки
Образцы Основная молекулярная формула
Enz
mes
.20002
Oxidoreductasen.
Трансфераза
1030 Cnh2,58nN0,28nO0,29nS0.008n
Гидролаза 1030 Cnh2.56nN0.27nO0.29nS0.007n
Lyase 1050 Cnh2.59nN0.28nO0.30nS0.008n
Изомераза
280 Cnh2.57nN0.27502NO 500 28nO0.30nS0.009n
Запасные белки 380 Cnh2.57nN0.27nO0.30nS0.009n
Гормоны 1020 Cnh2.57nN0.27nO0.296nS0.016n
Сигнальные белки 450 Cnh2.58000nN0.28n 1030 Cnh2.58nN0.27nO0.30nS0.01n
Транспортные белки 1100 Cnh2.59nN0.27nO0.29nS0.008n
Иммуноглобулины
612 Cn h2.57n N0.28n O0.31n S0.007n
Моторные белки
693 Cn h2.62n N0.28n O0.31n S00002n рецептор 9.002n 9.008n
764 Cn h2.56n N0.27n O0.29n S0.01n
Формула среднего значения 10739 Cn h2.58n N0.28n O0.30n S0.01n
Среднее значение коэффициентов, обозначающих углерод,
водород, Определено содержание азота, кислорода и серы
, представленное в Таблице Ш.Затем было вычислено общее среднее значение
средних коэффициентов для каждого атома. Так как
показано в таблице ш. В этой таблице общая корреляция между
и
коэффициентами составила 0,9999.
ТАБЛИЦА Ø
ИЗВЛЕЧЕННЫЕ НАБОРЫ ДАННЫХ НА ОСНОВЕ СРЕДНЕГО
КОЭФФИЦИЕНТОВ C, H, N, O и S
ДЛЯ КАЖДОГО ИЗ 9 БЕЛКОВ
КЛАССИФИКАЦИИ
Коэффициенты
CHNO Оксидоредуктаза 1 1.58 0,27 0,30 0,008
Трансфераза 1 1,58 0,28 0,29 0,008
Гидролаза 1 1,56 0,27 0,29 0,007
Лиаза 1 1,59 0,28 0,30 0,008
Изомераза 1 1,57 0,28 0,30 Хранение 0,018
Лигаза 1 1,58 0,28 0,30 0,009 0,27 0,30 0,009
Гормоны 1 1,57 0,27 0,30 0,016
Сигнальные белки 1 1,58 0,28 0,30 0,011
Структурные белки 1 1,58 0,27 0,30 0,010
Транспортные белки 1 1.59 0,27 0,29 0,008
Иммуноглобулины 1 1,57 0,28 0,31 0,007
Моторные белки 1 1,62 0,28 0,31 0,008
Рецепторные белки 1 1,56 0,27 0,29 0,010
Среднее значение 1 1,58 0,28 0,30 0,010
9000oreducts3 9000oreduct Общая корреляция образцы
Доступ
Код Формула белка Основная молекулярная формула
A5D4R8 C3142H5015N903O907S30 C
n h2.60n N0.29n O0.29n S0.010n
A5D560 C1926h4107N583O548S17 C
n h2.61n N0.30n O0.28n S0.009n
A5D616 C1712h3741N517O481S18 C
0003
73 N H0115 N0481
n h2.57n N0.27n O0.29n S0.007n
A5D685 C1226h2925N347O364S16 Cn h2.57n N0.28n O0.30n S0.013n
A5D686 C879h2426N250O20003. 003n
A5E8U7 C1944h4137N557O560S11 C
n h2.61n N0.29n O0.29n S0.006n
A6SYB5 C1630h3616N448O477S4 C
n h2.60n N0.27n O0.29n S0.002n
A6TA35 C1588h3514N418O474S15 N0.20.59 N0.205
A6TAB8 C1765h3748N518O517S2 C
n h2.56n N0.29n O0.29n S0.001n
A7K3A5 C980h2546N258O297S9 C
n h2.58n C02000 S03000
n h2.58n N07253.00 59n N0.28n O0.30n S0.007n
A7K531 C1741h3722N482O521S12 C
n h2.56n N0.28n O0.30n S0.007n
A7WHY8 C1323h3091N333O379S10 C
n h2.58n N0.25n O0.29n S0.008n
A7ZQY9 C1264h3029N357O3889.00n N0.2.31.60N0 N0.20002.69S8 N0.31
A8A422 C1262h3027N357O387S8 C
n h2.61n N0.28n O0.31n S0.006n
A9HMF9 C2681h5274N802O791S18 C
n h2.5960n C
n h2.59605 N030002 N H2.59605 N03 56n N0.29n O0.30n S0.009n
A9HX35 C1804h3804N516O532S11 C
n h2.55n N0.29n O0.29n S0.006n
A9HXI4 C1629h3597N467O475S8 C
n h2.59n N0.29n O0.29n S0.005n
World Academy of Science, Engineering and Technology 54 2011
Произошла ошибка при настройке вашего пользователя Печенье
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно.Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.
Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie. - Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом.Если часы вашего компьютера показывают дату до 1 января 1970 г.,
браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере. - Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.
Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу.Чтобы предоставить доступ без файлов cookie
потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта.Например, сайт
не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к
остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Новая наука о белке, основанная на полном химическом синтезе — Кент — 2019 — Наука о белке
В этой статье будут описаны примеры, отобранные из научной литературы, чтобы проиллюстрировать ключевые аспекты воздействия, которое полный химический синтез оказал на науку о белке, и дать понимание потенциального будущего вклада, который внесет химический синтез.
Хиральность
Одним из наиболее поразительных аспектов мира природных белков является гомохиральность: все природные белки состоят из 1-аминокислот и ахиральной аминокислоты глицина.20 Химический синтез белков может использоваться для управления хиральностью белков новыми способами, в том числе чтобы облегчить определение структуры белка, создать новые терапевтические ведущие молекулы и улучшить наше понимание молекулярных основ функции белка.
Термодинамическая гипотеза Анфинсена7 предполагала, что полный химический синтез с использованием d-аминокислот и глицина может быть использован для создания зеркального отображения «d-белков»: то есть белковых молекул, складчатые структуры которых являются зеркальным отображением природных белков. Пионерский химический синтез d-белков подтвердил это предположение.21, 22 Доступ к d-белкам, обеспечиваемый полным химическим синтезом, позволяет использовать несколько новых видов науки о белках.
Рентгеновская кристаллография рацемических белков
Есть два важных аспекта определения структуры белковой молекулы с помощью рентгеновской кристаллографии: образование высокоупорядоченных кристаллов белка; и (после получения данных дифракции рентгеновских лучей) получение информации о фазе, необходимой для определения структуры белка.Рацематы белков можно кристаллизовать и использовать для исследований дифракции рентгеновских лучей.22 Такое применение химически синтезированных молекул d-белка может способствовать как образованию кристаллов дифракционного качества, так и фазированию дифракционных данных для структурного раствора23
В качестве примера можно привести антифризный протеин от снежных блох (sfAFP). Хотя было легко получить sfAFP с помощью полного химического синтеза, 24 оказалось, что синтетический белок трудно кристаллизовать.После изучения тысяч условий кристаллизации в течение нескольких месяцев, в конечном итоге были получены кристаллы, и данные синхротронной дифракции рентгеновских лучей были получены с разрешением 0,98 Å. Однако, поскольку в то время не было известно родственного глобулярного белка, структура белка не могла быть решена путем молекулярной замены, и было невозможно получить кристаллы селенометионинсодержащего sfAFP для метода аномальной дисперсии для фазирования.25
В письме на номер Nature в 1989 г.26 Алан Маккей указал, что центросимметричный кристалл рацемата белка значительно упростил бы фазовую проблему, потому что возможные фазы сильно ограничены: недиагональные фазы сокращаются, поэтому все фазы связаны на пи радианы; например, в пространственной группе P1
Опыт программ структурной геномики за последние 15 лет показал, что только около одной трети рекомбинантно экспрессируемых очищенных глобулярных белков может быть кристаллизовано, и что немногим более половины этих белковых кристаллов имеют дифракционное качество27. существует ~ 80% глобулярных белковых молекул для получения кристаллов, пригодных для определения структуры стандартными методами дифракции рентгеновских лучей. Во многих случаях неспособность кристаллизовать молекулу глобулярного белка может быть преодолена с помощью кристаллографии рацемического белка, в которой используется смесь рацемических белков, то есть смесь, содержащая равные количества форм 1-белка и d-белка одной и той же молекулы белка. (Инжир.2). Облегченная кристаллизация смесей рацемических белков была предсказана в 1995 году Вуковицем и Йейтсом на теоретических основаниях28. Это предсказание было подтверждено легкой кристаллизацией рацемата белка sfAFP.25 Кристаллизация рацематов белка в настоящее время используется во все большем числе случаев для выяснения. структуры белковых молекул. На сегодняшний день в нескольких десятках примеров, большинство из которых связано с непокорными белками, сообщалось о более чем 80% успешном получении кристаллов дифракционного качества.29
Кристаллография рацемических белков. Взято из ссылки 29.
Кристаллизация рацемического белка одновременно облегчает кристаллизацию и упрощает фазовую проблему. Ген Rv1738 активируется и транскрибируется в наибольшей степени, когда M. tuberculosis переходит в устойчивое состояние покоя.30 Однако до недавнего времени ни структура, ни функция предсказанной молекулы белка Rv1738 не были известны. Белок Rv1738, экспрессируемый в E.coli не смогла дать кристаллы дифракционного качества в ходе обширных испытаний в течение нескольких лет с использованием полного диапазона изменений молекулы белка, способствующего кристаллизации, а также роботизированного скрининга широкого диапазона условий кристаллизации.31 l-белок и d-белковые формы Rv1738 были получены полным химическим синтезом. Рацемическая смесь l-Rv1738 и d-Rv1738 дала центросимметричные кристаллы, которые дифрагировали до разрешения 1,5 Å, а структура Rv1738 была решена с использованием нового метода фазирования ab initio , допускаемого упрощенными фазами (рис.3). Элементарная ячейка содержала зеркальные гомодимеры, связанные через центр инверсии. L-гомодимер состоял из двух белков l-Rv1738, а d-гомодимер состоял из двух белков d-Rv1738. Поиск на основе структуры выявил сходство между гомодимерной молекулой белка Rv1738 и белками бактериального «фактора, способствующего гибернации», которые отключают синтез рибосомного белка, что позволяет предположить функциональную роль Rv1738 в устойчивом состоянии покоя M. tuberculosis .31
Структура белка Rv1738 из M.tuberculosis определен с помощью кристаллографии рацемических белков. (А) Рацемический кристалл. (B) Рентгенограмма. (C) Зеркальное отображение структур гомодимера Rv1738. Синее изображение — это l-гомодимер, а красное изображение — d-гомодимер. Центр инверсии показан голубой точкой. (D) Карта электронной плотности 2Fo – Fc для боковой цепи Trp. По материалам Ref. 31.
Другой пример полезности кристаллографии рацемических белков включал наномолярное сродство связывания d-белка с VEGF-A, которое было разработано с помощью фагового дисплея с зеркальным отображением (см. Ниже).Чтобы определить, как связывающий d-белок взаимодействует с VEGF-A, был приготовлен раствор, который содержал четыре химически синтезированных белка: энантиомеры белка d-VEGF-A и l-VEGF-A вместе с двумя эквивалентами каждого энантиомера связывающего белка. . Были получены кристаллы рацемического белка и данные синхротронной дифракции были собраны с разрешением 1,6 Å.32 Структура рацемического гетерохирального белкового комплекса была определена путем молекулярного замещения. Краткий отчет об этой работе можно найти здесь: 29
Квази -рацемическая рентгеновская кристаллография белка
Облегченная кристаллизация смесей рацемических белков является свойством зеркального отображения форм энантиомеров белка.Это важное явление также применимо к энантиоморфам белка — близкозеркальным белковым молекулам, которые не являются истинными энантиомерами, но имеют зеркальную форму. Первым примером кристаллографии рацемического белка квази- было определение структуры sfAFP, описанной выше. В этой работе селенсодержащий аналог l-sfAFP (в котором остаток Asn был заменен остатком алкилированного SeCys) был смешан с d-sfAFP, который
, а не , содержат селен для облегчения образования квазирацемических кристаллов дифракционного качества для определения фазы методом аномального диспергирования.1 25 Квазирацемическая кристаллография была объединена с рацемической кристаллографией для определения структуры растительного антимикробного белка snakin-1 с помощью фазирования, вызванного радиационным повреждением.
Важной особенностью квазирацемической кристаллической структуры аналога l-белка с формой d-белка соответствующего нативного белка является то, что путем цифровой инверсии структуры d-белка определяется эталонная структура нативного l-белка. в том же эксперименте.Эта особенность квазирацемической кристаллографии белка была использована с хорошим эффектом при определении кристаллической структуры нового топологического аналога белка крамбина и его сравнении с нативным крамбином [описанным ниже в разделе «Топология» (см. Рис. 5)]. 34
Облегченная кристаллизация смесей квазирацемических белков, состоящих из индивидуальных аналогов l-белка и энантиомера d-белка молекулы нативного белка, может значительно ускорить определение структур аналогового белка методами дифракции рентгеновских лучей.Полезность этого применения квазирацемической кристаллографии была проиллюстрирована при изучении эффектов замены α-аминокислот на β-аминокислоты в небольшой молекуле белка, 35 и при определении структур аналогов диастереомеров боковой цепи белка (см. Диастереомеры белков ниже) 36
Гликопротеины трудно кристаллизовать из-за гибкости сложных гликановых фрагментов. Чтобы исследовать облегченное определение структуры гликопротеина с помощью квазирацемической кристаллографии, хемокин CCL1 был химически синтезирован в форме l-белка, гликозилированного l-белка и d-белка.37 Кристаллы были получены из истинного рацемата {d-CCL1
плюс l-CCL1}, а также из квази -рацемата {d-CCL1
плюс глико-1-CCL1}. Обе кристаллические формы дифрагировали с разрешением 2,15 Å. Кристаллическую структуру CCL1 определяли путем молекулярного замещения с использованием данных дифракции, полученных от истинного рацемата. Кристаллическую структуру гликозилированного CCL1 определяли путем молекулярного замещения с использованием данных дифракции от квазирацемата.38 Несмотря на облегченную кристаллизацию гликопротеина CCL1 в виде квазирацемата, сложный гликановый фрагмент был неупорядочен, и только сахар, непосредственно присоединенный к Asn29, хорошо определялся на полученной карте электронной плотности.
Об изобретательном расширении квазирацемической кристаллографии недавно сообщил Лей Лю (Университет Цинхуа) и его сотрудники.39 Несколько различных разветвленных олигоубиквитиновых конструкций с естественной хиральностью l-белка были совместно кристаллизованы с d-белковой формой мономера убиквитина.Наблюдали облегченное образование кристаллов, давая центросимметричные квазирацемические кристаллы, в которых несколько копий мономера d-убиквитина расположились с образованием квазирацематов с каждым l-убикутином в разветвленных ковалентных олигомерах. Данные рентгеновской дифракции были собраны с хорошим разрешением, и была определена структура каждого из разветвленных олигомерных убиквитинов.39 Этот полезный метод {d-мономер + 1-олигомер} предполагает, что для облегчения кристаллизации можно предусмотреть даже более радикальные квазирацематы. устойчивых белковых молекул, таких как использование слитых белков, связывающих мальтозу (MBP), плюс d-протеиновая форма MBP.
Скрининг библиотек хиральных пептидов, белков и природных продуктов
Природные белки, встречающиеся в природе пептиды и почти все природные продукты вторичных метаболитов с низкой молекулярной массой, встречающиеся в природе, являются хиральными соединениями.40 Может быть желательно создать зеркальные формы этих встречающихся в природе молекул, в случае белков и пептидов, чтобы улучшить стабильность in vivo и снизить иммуногенность, а также для природных продуктов с низкой молекулярной массой, чтобы обеспечить новые ведущие соединения для разработки фармацевтических препаратов.
Белки с зеркальным отображением являются ключевым компонентом оригинального метода «зеркального фагового дисплея» для систематической разработки высокоаффинных лигандов d-пептида для природных белков.41 Форма d-белка природного l-белка получается путем полного химического синтеза, и используется в качестве приманки при выборе связывающих веществ из пептидных библиотек, отображаемых на фаге. После идентификации пептида с высоким сродством синтезируется d-пептид с той же аминокислотной последовательностью.По причинам симметрии 42 d-пептид имеет такое же высокое сродство к нативному l-белку-мишени. Этот мощный метод разработки стабильных, высокоаффинных лигандов d-пептида в настоящее время используется для множества терапевтических белков-мишеней.43-49 Во многих случаях достаточно создать один домен целевого белка в форме зеркального отображения, а не в форме зеркального отражения. чем вся молекула белка, что упрощает задачу синтеза.
Зеркальное изображение белок фаговый дисплей — мощный метод систематической инженерии взаимодействий между левыми и правыми молекулами белка.Чтобы разработать антагонист d-белка ангиогенного белкового гормона фактора роста эндотелия сосудов (VEGF-A) 50, белок d-VEGF-A размером 22 кДа был получен путем полного химического синтеза и использовался для скрининга большого количества варианты белкового каркаса, отображаемые на фаге. Был идентифицирован наномолярный аффинный связывающий d-белок. Данные о рацемической кристаллической структуре гетерохирального комплекса VEGF-A и связующего d-белка (описанного выше) 32 использовали для уточнения свойств этого антагониста d-белка VEGF-A.50
Хиральность — это доминирующий фактор во взаимодействии низкомолекулярных соединений натуральных продуктов с молекулами белков.51 Обычно исследования по открытию лекарственных препаратов в остро конкурентной области фармацевтических разработок на основе натуральных продуктов сильно пересекаются. Обычно одни и те же белки идентифицируются как важные в настоящее время мишени, и часто бывает, что несколько программ разработки лекарств предлагают аналогичные ведущие соединения для дальнейшего уточнения с помощью медицинской химии.По этой причине большое значение будет иметь метод идентификации уникальных соединений свинца. Очень рано было понято, что скрининг библиотек соединений хиральных природных продуктов (NP) против химически синтезированной зеркальной формы белковой мишени с последующим созданием зеркального отображения выбранных NP с помощью обычного органического синтеза может дать уникальные выводы, которые не обнаружены. обычными методами скрининга52 (рис. 4).
Скрининг библиотеки хиральных натуральных продуктов против зеркальных форм фермента может обеспечить новые совпадения.Затем синтез зеркального отражения хитового соединения дает новую молекулу, родственную натуральному продукту, активную против нативного фермента.
Недавно Нобутака Фуджи и др. воплотили эту концепцию в жизнь.53 Они проверили хиральную библиотеку, состоящую из ~ 22 000 соединений природного продукта, против зеркальных форм p53-связывающего домена белка MDM2. Хиральное производное α-токоферола было идентифицировано как ингибитор взаимодействия d-MDM2 / d-p53. Зеркальное отображение идентифицированного производного α-токоферола, полученного обычным органическим синтезом, ингибировало взаимодействие l-MDM2 / l-p53, но не влияло на взаимодействие d-MDM2 / d-p53.53
Диастереомеры белков
Диастереомеры представляют собой белки, которые содержат смесь d-аминокислот и l-аминокислот, 54 или которые имеют перевернутую стереохимию боковой цепи Thr или Ile.55 Эти стереохимические гибриды могут быть полезными зондами молекулярной основы свойств белка.
Термодинамика C-концевого «перекрытия спирали», геометрически ограниченного участка, где альфа-спиральный участок полипептида переходит в более протяженную вторичную структуру, была исследована в молекуле белка убиквитина.56 Полный химический синтез был использован для замены консервативного остатка Gly сразу после альфа-спирали на d- и l-энантиомеры аминокислот Ala и Val, таким образом создавая две пары диастереомеров белка. Измеренные энергетические различия между стабильностью диастереомеров показали, что доминирующим термодинамическим эффектом была необходимость принять левостороннюю конформацию скелета для остатка после альфа-спирали, конформацию, которая может быть принята только Gly из всех протеиногенных аминокислот. или d-аминокислотой.56
Белок ShK токсина яда скорпиона содержит два остатка Ile и четыре остатка Thr. На основании предсказанной относительной стабильности из расчетов молекулярной динамики три диастереомера ShK были химически синтезированы как l-белки, содержащие один остаток со стереохимией перевернутой боковой цепи: [l- allo -Ile 7 ] ShK; [1- алло -Thr 13 ] ShK; и [1- allo -Thr 31 ] ShK.1 36 Скорости сворачивания и относительная стабильность свернутых диастереомеров согласуются с расчетами.Квазирацемическая кристаллография, в которой каждый диастереомер l-ShK был совместно кристаллизован с d-ShK, была использована для определения структуры диастереомеров [l- allo -Ile 7 ] ShK и [l- allo ] -Thr 13 ] ШК. Интересно, что наименее стабильный диастереомер [1- алло -Thr 31 ] ShK спонтанно кристаллизовался сам по себе из квазирацемической смеси с d-ShK.36
Топология
В структурной науке о белках термин «топология» используется для описания вторичных и третичных структур свернутой белковой молекулы.В математическом смысле 57 «топология» отличает линейные полипептидные цепи, обычно встречающиеся в белках, от разветвленных, кольцевых, линейных петель и множества других возможных конфигураций полипептидных цепей. Здесь мы будем использовать термин топология в обоих этих смыслах.
Вторичная структура
Предварительно сформированные компоненты вторичной структуры или фрагменты, индуцирующие вторичную структуру, могут быть введены в пептидную цепь белковой молекулы путем химического синтеза.Последовательность Gly 16 –Gly 17 в обратном повороте в молекуле протеина протеазы ВИЧ-1 была заменена на BTD, непептидный имитатор β-поворота типа II фиксированной геометрии.58 Сложенный BTD-содержащий белок имел полная ферментативная активность, неизменная субстратная специфичность и повышенная термостабильность. Аналогичный подход был использован для изучения домена белка GB1 Staph. Aureus ,59. Совсем недавно лактамы, образованные между боковой цепью аминокислоты и соседней пептидной связью, были использованы для контроля вторичной структуры белка.60
Другой подход к контролю вторичной структуры белков можно назвать «космической инженерией Рамачандрана». Гомодимерная молекула протеина протеазы ВИЧ-1 имеет две идентичные мобильные лоскутные структуры, которые закрываются над связанным субстратом. Ковалентная димерная форма протеазы ВИЧ-1 с полипептидной цепью из 203 аминокислот была получена путем полного химического синтеза, чтобы сделать возможными асимметричные изменения в ковалентных структурах лоскутов.61 Остаток Gly 51 и Gly 51 ‘ были заменены на d-Ala, l-Ala или Aib (α-амино-изомасляная кислота) в одном или обоих створках, что привело к образованию серии молекул аналогового белка, в которых доступное конформационное пространство позвоночника на кончике каждого лоскута было ограничено определенным образом. Симметричная замена Gly 51 и Gly 51 ’ на d-Ala или l-Ala приводила к существенному снижению ферментативной активности. Протеаза ВИЧ-1 [Ala 51 , Aib 51 ’], в которой доступные конформации лоскута были сильно ограничены, имела резко сниженную ферментативную активность.Интересно, что асимметричный аналог [d-Ala 51 , l-Ala 51 ‘] протеаза ВИЧ-1 обладал полной ферментативной активностью, что позволяет предположить, что различная геометрия скелета наблюдалась для Gly 51 и Gly 51′ в исходном состоянии. ферменты функционально значимы.62
Новая ковалентная топология
Synthesis можно использовать для создания уникальных химических вариаций простой линейной топологии от N-конца к C-концу полипептидных цепей в большинстве природных белков.
Ядерные белки cMyc и Max регулируют экспрессию генов, образуя нековалентный гетеродимер, который связывается со специфической нуклеотидной последовательностью в двухцепочечной ДНК.63 Была разработана конструкция гетеродимера из 172 остатков, в которой отдельные домены b / HLHZ cMyc и Max были ковалентно связаны в C-концевой области каждого домена, чтобы получить молекулу белка с беспрецедентной ковалентной структурой: полипептидная цепь изменяет направление в средней точке и имеет два N-концевых остатка, но не имеет C-конца.Целевой ковалентный гетеродимер получали конденсацией четырех синтетических пептидных сегментов с использованием двух различных, взаимно совместимых химических методов лигирования. Этот уникальный химический аналог белка cMyc-Max обладал той же специфичностью связывания, что и нековалентный гетеродимер. Интересно, что КД-спектроскопия синтетической конструкции ковалентного гетеродимерного белка cMyc-Max показала, что он был предварительно свернут в отсутствие лиганда дцДНК, тогда как домены b / HLHZ нативных cMyc и Max образуют только свернутый белок (нековалентный). димер в присутствии дцДНК-мишени.64
Убиквитин — это высококонсервативная белковая молекула, которая в природе присоединяется к другим белкам для изменения их биохимических свойств.65 Присоединение убиквитина к боковой цепи лизина в другой молекуле белка может быть использовано для нацеливания этого белка на деградацию протеасомным механизмом клетки. .66 Ковалентная модификация гистоновых белков убиквитином модифицирует экспрессию генов.67 В обеих этих биохимических системах присоединенный убиквитин может быть дополнительно модифицирован ковалентным присоединением одного или нескольких убиквитинов с образованием убиквитиновых цепей.68 Топологически убиквитинированные белки состоят из разветвленных полипептидных цепей . Несколько исследовательских групп использовали химический синтез белка для создания разветвленных конструкций убиквитин-белок с определенной ковалентной структурой, а затем для исследования их биохимических свойств.69-73 Новые методы химического синтеза олигоубиквитинов предоставили разветвленные белковые конструкции с определенной ковалентной структурой. с массами до 51 кДа, 74 и даже больше мультиубиквитинов путем контролируемой нативной химической лигированной полимеризации.75
В клетке некоторые линейные полипептидные цепи, продуцируемые рибосомной трансляцией, подвергаются посттрансляционной биохимической обработке с образованием белков, которые имеют кольцевых полипептидных цепи : то есть белковые молекулы, в которых N-конец и C-конец полипептидной цепи находятся соединены пептидной связью. Кольцевые белки первоначально были выделены из растений и названы «циклотидами». 76 В исследовательской лаборатории химический синтез является предпочтительным способом создания этих кольцевых белковых молекул для изучения структуры и функции.77 Циркулярные белки были идентифицированы из самых разных природных источников, в том числе из бактерий.78 Группа Боде недавно сообщила об общем химическом синтезе 70-остаточного кольцевого белка бактериоцина As-48.79
Катенаны — это молекулы, в которых две макроциклические структуры взаимопроникают и, таким образом, механически связаны друг с другом. Химический синтез был использован для разработки и создания белковых катенанов s, которые содержат взаимосвязанные макроциклические полипептидные цепи.80
Химический синтез также был использован для получения уникального аналога крамбина, в котором взаимодействие ионной пары между гуанидином боковой цепи остатка Arg10 и α-карбоксилатом полипептидной цепи было заменено ковалентной связью с образованием полипептидной цепи с линейной петлей. ковалентная топология. Сворачивание дало молекулу белка новой топологии, в которой N-конец полипептидной цепи проникает в кольцо макролактама и удерживается на месте дисульфидными связями, напоминая лассо-пептиды.1 34 Структура этого нового топологического аналога белка была определена с помощью квази -рацемической рентгеновской кристаллографии (рис. 5).
Quasi- рацемическая рентгеновская кристаллография: определение структуры взаимопроникающего топологического аналога крамбина с линейной петлей. (A) Кристаллографическая элементарная ячейка. (B) Мультяшное изображение структуры позвоночника. Новая амидная связь показана в виде сфер КФК. C. Электрон карта плотности 2Fo – Fc с подобранной структурой новой амидной связи, показанной в виде стержней. (D) Наложение структуры топологического аналога крамбина (зеленый) и структуры нативного крамбина (голубой), полученного путем инвертирования структуры d-крамбина. Обе белковые структуры были получены в одном эксперименте. По материалам Ref. 34.
Химическая промышленность
Synthesis позволяет использовать весь репертуар органической химии для создания уникальных химических вариантов ковалентной структуры белка.Существует обширная литература по «пептидомиметической химии» 82, и все они могут быть применены к молекулам белков с использованием современных методов химического синтеза. Корреляция изменений ковалентной структуры белковых молекул с эффектами на структуру и свойства свернутых белков обеспечивает новое понимание молекулярных основ функции белка и позволяет создавать новые формы (пере) дизайна белков.
Магистральная инженерия
Ранним примером уникальных химических вариантов белка была «инженерия остова» путем полного химического синтеза молекулы протеина протеазы ВИЧ-1.Две пептидные связи были заменены тиоэфирными связями, что предотвратило образование водородных связей с карбонилами на пептидном субстрате.83 Полученный химически сконструированный фермент имел k cat (число оборотов, когда фермент насыщен субстратом) ниже на ~ В 3000 раз по сравнению с молекулой фермента нативного скелета. Это каталитическое снижение скорости на ~ 5 ккал / моль соответствует энергии двух водородных связей, предполагая, что эти взаимодействия между пептидным субстратом и основной цепью белка являются каталитически релевантными.84 Недавние примеры инженерии белковой основы с помощью химического синтеза включают использование химической конденсации «щелчка» пептидных сегментов с получением белковых молекул, содержащих изостер триазола вместо пептидной связи.85, 86 Химический синтез был использован для проведения систематической C -альфа- и амид-азот-метил-сканирование полипептидной цепи белка для выявления конформаций скелета, которые влияют на структуру и функцию белка.87
Транслокация боковой цепи
В молекулах природного белка боковые цепи аминокислот являются боковыми цепями α-углеродов в основной цепи полипептида.Химический синтез белка можно использовать для сдвига боковой цепи аминокислоты к атому азота пептидной связи. В модельном белке ингибитора трипсина поджелудочной железы крупного рогатого скота (BPTI) остаток Cys38 был заменен пептоидным фрагментом, несущим N α -этанотиоловый фрагмент. Уникальный химический аналог белка [(N α (CH 2 ) 2 SH) Gly 38 )] BPTI имел такую же складчатую структуру и термостабильность, что и нативный BPTI, как показал многомерный ЯМР88. , полусинтез использовали для осуществления серии пептоидных транслокаций боковых цепей отдельных остатков Lys, Glu и His в N-концевой области полипептидной цепи фермента рибонуклеазы A, а также для изучения влияния на ферментативную активность полученных аналогов.89
Медицинская химия
Используя синтетическую химию, принципы медицинской химии могут быть применены к молекулам белков, чтобы оптимизировать их свойства. Для хемокина RANTES это привело к разработке уникального химического аналога белка AOP-Rantes, который связывается с хемокиновым рецептором CCR5 и ингибирует проникновение ВИЧ-1 в клетки периферической крови.90, 91 человеческие терапевтические средства были улучшены за счет включения галоген-модифицированных аминокислот.92 Применение медицинской химии к инсулину в значительной степени облегчается за счет разработки эффективного полного синтеза молекулы белка инсулина через «сложноэфирный инсулин», химический заменитель проинсулина, в котором две полипептидные цепи зрелого инсулина связаны сложноэфирной связью. между боковыми цепями Glu A4 и Thr B30 . Эффективно это заменяет 35 остатков С пептида проинсулина одной ковалентной связью между двумя боковыми цепями. Депсипептидная цепь из 51 остатка сворачивается при физиологическом pH, и полученный сложноэфирный белок инсулина может быть химически преобразован в человеческий инсулин с отличным выходом.93-95 Сложноэфирные депсипротеины основной цепи могут быть получены с помощью химии лигирования KAHA, которая может непосредственно генерировать продукты лигирования сложноэфирной полипептидной цепи.96
Гликопротеиновая инженерия
Делая шаг вперед в науке о синтетических гликопротеинах, Kajihara et al. сообщили об общем химическом синтезе, фолдинге и биологической активности пяти различных гликоформ эритропоэтина (ЭПО). Каждая гликоформа содержала один или несколько двухантенных сиалилолигосахаридов человеческого типа в определенных положениях гликозилирования, и они были получены с высокой степенью чистоты и с определенной химической структурой97 (рис.6).
Гликоформы ЭПО, полученные методом полного химического синтеза. (A) Модульная конвергентная синтетическая схема. В нижнем ряду показаны мультяшные изображения пяти гликоформ. (B) Масс-спектры синтетических гликопротеинов с электрораспылением для прямой инфузии. Наблюдаемые массы синтетических гликопротеинов составляли 20 540 Да, 22 746 Да (три изомерные гликоформы) и 24 952 Да. По материалам Ref. 97.
В более ранней работе был разработан и синтезирован синтетический белок эритропоэза (SEP) — химический «гликопротеиновый миметик» варианта EPO.SEP содержал две тетраразветвленные олиго (этиленоксид-амидные) части, каждая из которых несла четыре отрицательных заряда, которые заменяли сложные сиалогликаны нативного белка. Молекула белка SEP 50 825 Да обладала полной биологической активностью и увеличенной продолжительностью действия in vivo .98, 99
Априорная
конструкция белка
Создание белковых молекул с заданной структурой и новыми функциями — это «святой Грааль» белковой науки.100 Полный химический синтез позволяет разработчику выйти за рамки 20 аминокислот, обычно присутствующих в белках, и обойти парадигму линейной полипептидной цепи, навязанную рибосомной трансляцией. Примером третичной структуры является разработанный трициклический белок «CovCore 3h3». Белок 6280 Да был получен путем полного химического синтеза и содержит кольцевую полипептидную цепь. Белок был заблокирован в том, что в противном случае было бы высокоэнергетической конформацией из-за неестественных ковалентных ограничений.Его структура была определена методом рентгеновской кристаллографии102 (рис. 7).
Рентгеновская структура априорно сконструированной белковой молекулы. Кольцевая полипептидная цепь показана фиолетовым цветом с карикатурным изображением спиральных участков. Ароматическое кольцо, используемое для ковалентного соединения трех спиралей и ограничения складчатой структуры, показано голубым цветом.102
Аналогичный подход был использован для создания тримерного аналога 24 кДа амилоидного агрегата.103
Продвинутые физические техники
Химический синтез обеспечивает исключительно точное введение ядер изотопных зондов в молекулу белка и сайт-специфическое прикрепление других спектроскопических зондов. Это позволило по-новому применить передовые спектроскопические методы для выяснения молекулярных основ структуры и функции белков.
Ядерный магнитный резонанс
Спектроскопия ЯМР
широко используется для определения структуры белков и исследования динамики белков.Назначение резонансов отдельным ядрам и определение молекулярной структуры методами многомерного ЯМР может быть сложной задачей для многодоменных белков, и особенно сложной задачей для интегральных мембранных белков из-за спектрального перекрытия резонансов от гидрофобных аминокислотных остатков в трансмембранных спиралях. 104 Химическое лигирование может использоваться для упрощения этих проблем путем включения сегментов, меченных изотопами ЯМР, и немеченых сегментов в полипептидную цепь белка.105-107 ЯМР-спектроскопия особенно полезна для определения ионизационных свойств функциональных групп в белковых молекулах. Однозначное определение значений функциональности отдельных боковых цепей для p K a важно для понимания ферментативного катализа и в значительной степени облегчается за счет сайт-специфического включения аминокислот, содержащих ядра ЯМР-зонда, в молекулу белка.108
Флуоресценция
Флуоресцентные красители являются полезными датчиками динамики белков и могут использоваться для измерения расстояний внутри белков и белковых комплексов.Химический синтез может включать флуоресцентные красители в определенных участках белковой молекулы для измерения расстояний с помощью Förster Resonance Energy Transfer (FRET) .109 Сайт-специфически меченные флуоресцентные токсинные белки были получены путем полного химического синтеза и использованы для измерений резонансного переноса энергии лантаноидов (LRET). конформационных изменений потенциалзависимого натриевого канала Na V 1.4 в состоянии покоя и в инактивированном состоянии, а также в живых клетках 110 (рис. 8).
Измерение расстояний в натриевом канале Nav1.1 молекула белка. Измерения резонансного переноса энергии лантаноидов (LRET) стали возможными благодаря полному химическому синтезу малых белковых токсинов, сайт-специфично меченных флуорофорным красителем. По материалам Ref. 110.
Ансамблевые измерения усредняют конформации и динамику субпопуляций белков и, таким образом, скрывают поведение отдельных белковых молекул. Флуоресцентные зонды можно использовать для изучения одиночных молекул ферментного катализа. Ковалентную гомодимерную протеазу ВИЧ-1, снабженную связкой ПЭГ-биотин, получали путем полного химического синтеза и закрепляли на предметном стекле, покрытом стрептавидином.Используя пептидный субстрат, меченный одним членом пары красителей FRET на N-конце и другим красителем на C-конце, регистрировали зависимые от времени наблюдения связывания и расщепления субстрата одной молекулы.111 Одномолекулярный FRET также может быть осуществляется с помощью химически синтезированных белковых молекул, содержащих несколько флуоресцентных красителей. Конвергентный общий химический синтез в сочетании с региоспецифической защитой боковой цепи цистеина использовали для сайт-специфического введения трех различных флуорофоров в молекулу белка -синуклеина.Одномолекулярные трехцветные FRET-эксперименты проводили в растворе на свободно диффундирующих молекулах α-синуклеина.112
Инфракрасная спектроскопия
Инфракрасная спектроскопия белков с двумерным преобразованием Фурье с временным разрешением (FTIR) может предоставить информацию о конформационной динамике белков в масштабах от пикосекунд до миллисекунд.113 Чтобы использовать FTIR для исследования роли специфических водородных связей в образовании димеров инсулина, общий химический анализ синтез использовали для получения молекул инсулина, содержащих массивы изотопных зондов 13 C = 18 O в карбонильных фрагментах специфических пептидных связей в двух полипептидных цепях белка.114
Новые вибрационные зонды, доступные для наблюдения через прозрачное окно между 1880 и 2500 см −1 , могут быть введены в определенные места, чтобы действовать как репортеры электростатики и других характеристик физико-химической среды внутри белковой молекулы.115 Эффект Штарка, сдвиг частоты инфракрасного резонанса в ответ на изменения приложенного электрического поля, является чувствительным зондом локальной электростатики внутри белковой молекулы.116 Фрагменты вибрационного зонда, такие как нитрил и тиоцианат, были введены в молекулы ферментного белка на боковых цепях остатков Cys. В пионерских экспериментальных исследованиях нитрильные колебательные зонды с эффектом Штарка использовались для выяснения роли электростатических эффектов в ферментативном катализе.117 Эти и аналогичные исследования эффекта Штарка подтверждают значительный вклад электрических полей в катализатор в молекуле ферментного белка, образующей комплекс с его субстратом118. Недавно был опубликован современный взгляд на роль электростатики в ферментативном катализе.119 Химический синтез был бы более универсальным и контролируемым способом введения фрагментов зонда с вибрационным эффектом Штарка в молекулы белка. Полный химический синтез будет играть важную роль в дальнейших экспериментальных испытаниях роли и величины предварительно ориентированных диполей Н-связи в ферментативном катализе.
Первичная структура белка | Аминокислоты и химический состав — стенограмма видео и урока
Первичная структура белка
Какова первичная структура белка? Белки являются структурными компонентами клеток и выполняют множество функций, например транспортируют материалы, обеспечивают структурную поддержку и катализируют химические реакции.Это макромолекулы, состоящие из мономерных аминокислот. Есть двадцать различных аминокислот, которые могут составлять белки. Порядок расположения определенных аминокислот в белке — это первичная структура белка. Например, часть первичной структуры белка гемоглобина, который переносит кислород у животных, следующая:
val — his — leu — thr — pro — glu
Это означает, что первые шесть аминокислот в гемоглобине — это валин, гистидин, лейцин, треонин, пролин и глутамат.Белки могут состоять из тысяч аминокислот, так что это только начало первичной структуры гемоглобина.
Формирование цепи
Первичная структура белка формируется в процессе синтеза белка. При синтезе белка исходная первичная структура кодируется инструкциями ДНК.ДНК копируется в информационную РНК (мРНК) во время транскрипции. Затем мРНК экспортируется из ядра для выполнения следующего шага — трансляции. Во время трансляции рибосомы прикрепляются к мРНК, которые представляют собой уменьшенные копии отдельных генов ДНК. Рибосомы считывают мРНК группами из трех нуклеотидов, называемыми кодонами. Каждый кодон кодирует одну аминокислоту. Рибосома катализирует образование пептидной связи между указанными аминокислотами посредством синтеза дегидратации, при котором молекула воды теряется.Эта полимеризация формирует полипептидную цепь, которая в конечном итоге сворачивается в трехмерный белок посредством посттрансляционной модификации. Во время посттрансляционной модификации к первичной последовательности белка могут быть добавлены дополнительные модификации. Полипептидная цепь имеет определенную ориентацию, причем один термин называется N-элементом, а другой — C-элементом. Это относится к ориентации аминокислот. Полипептидные цепи всегда синтезируются и считываются от N-члена к C-члену.
Секвенирование первичных структур белка
Существует несколько способов выяснить первичную структуру белка. Если белок можно выделить, существует несколько методов, включая следующие:
- Деградация по Эдману
- Масс-спектрометрия
При деградации по Эдману одна аминокислота удаляется из белка за раз с использованием специального реагента, называемого фенилизотиоцианатом, который не повреждает последовательность. Однако деградация по Эдману может занять много времени и не подходить для нужд высокой производительности.Сегодня многие ученые используют масс-спектрометрию для идентификации белков. Во время масс-спектрометрии белок расщепляется на более мелкие фрагменты, которые затем можно провести с помощью высокоэффективной жидкостной хроматографии для идентификации компонентов. Затем вычислительные алгоритмы могут быть использованы для выяснения исходной последовательности белка.
Если ген белка известен, ген можно секвенировать с помощью технологии секвенирования ДНК. Как только последовательность ДНК известна, генетический код можно использовать для идентификации аминокислот в первичной последовательности белка.
границ | Автоматизированная фрагментация QM / MM Расчет химических сдвигов ЯМР для комплексов белок-лиганд
Введение
Вычислительные методы, основанные на структуре, являются полезными инструментами для анализа способов связывания и сродства для комплексов белок-лиганд (Grinter and Zou, 2014). С развитием рентгеновской кристаллографии и технологии ядерного магнитного резонанса (ЯМР) было определено более 100000 трехмерных белковых структур с высоким разрешением, что полезно для поиска основных соединений и терапевтических целей (Ferreira et al., 2015). По сравнению с экспериментальными методами, вычислительные подходы, такие как молекулярная стыковка, являются быстрыми и эффективными способами открытия лекарств. В программах молекулярного стыковки оценочные функции используются для аппроксимации свободной энергии связи и, следовательно, для ранжирования смоделированных поз-приманок (Sliwoski et al., 2014). Большинство оценочных функций примерно получены из потенциалов, основанных на силовом поле (Morris et al., 1998; Englebienne and Moitessier, 2009), эмпирических (Eldridge et al., 1997; Murray et al., 1998) или основанных на знаниях (Gohlke и другие., 2000; Хуанг и Цзоу, 2006). Однако, основанные на параметризованных функциях, эти методы оценки обычно недостаточно точны, чтобы отличить экспериментальную структуру от пристыкованных структур-ловушек, и иногда рейтинги из разных программных пакетов могут быть несовместимыми (ledź and Caflisch, 2018). Для решения этой проблемы много усилий было направлено на разработку методов стыковки путем введения экспериментальной структурной информации или квантово-механических (КМ) расчетов для оценки исходных и прогнозируемых поз (Mohan et al., 2005; Гринтер и Зоу, 2014; Адени, Солиман, 2017).
Химический сдвиг — один из наиболее эффективных и точных параметров ЯМР в отражении локальной химической среды вокруг атома, которая играет важную роль в определении и уточнении структуры (Zhu et al., 2014; Bratholm and Jensen, 2017). Для расчетов химического сдвига ЯМР используются программы для прогнозирования эмпирического химического сдвига ShiftS (Xu и Case, 2001; Moon and Case, 2007), ShiftX (Neal et al., 2003), ShiftX2 (Han et al., 2011), CamShift (Robustelli et al., 2010), PROSHIFT (Meiler, 2003; Meiler and Baker, 2003), SHIFTCALC (Williamson and Craven, 2009), ProCS (Christensen et al., 2014; Bratholm and Jensen, 2017). ), CheShift (Vila et al., 2009; Garay et al., 2014) и Sparta + (Shen, Bax, 2010). Эти эмпирические методы обладают высокой скоростью вычислений и успешно предсказывают химические сдвиги в основной цепи. Поскольку эмпирические формулы для этих моделей получены на основе экспериментальной или рассчитанной QM базы данных химических сдвигов и высококачественных структур, эти модели не очень подходят для точного предсказания химических сдвигов ЯМР для некоторых сложных систем, таких как комплексы белок-лиганд, нестандартные белковые остатки или неканонические пары оснований в системах нуклеиновых кислот (Swails et al., 2015). Расчеты квантово-механического химического сдвига в принципе способны предсказать химические сдвиги ЯМР для любых сложных систем (Lodewyk et al., 2012; Hartman and Beran, 2014; Merz, 2014). Для расчетов химического сдвига ЯМР белков Cui и Karplus предложили очень эффективный подход QM / MM (Cui and Karplus, 2000), Gao et al. разработали метод молекулярных орбиталей фрагментов (FMO) (Gao et al., 2007, 2010), Экснер с соавторами использовали подход сборщика регулируемой матрицы плотности (ADMA) (Frank et al., 2012; Victora et al., 2014), Тан и Беттенс разработали комбинированный метод фрагментации (CFM) (Tan and Bettens, 2013), а He и его коллеги разработали метод автоматизированной квантовой механики / молекулярной механики фрагментации (AF-QM / MM) (He et al., 2009, 2014; Zhu et al., 2012, 2013, 2014, 2015; Swails et al., 2015; Jin et al., 2016). Эти методы QM на основе фрагментов успешно применялись для расчета химического сдвига ЯМР. белков и нуклеиновых кислот.
Исходя из того, что химические сдвиги или возмущения химического сдвига (CSP) чувствительны к изменениям химической среды, эти параметры вполне подходят для определения структуры (Case, 1998; Cavalli et al., 2007; Шен и Бакс, 2015). Многие методы на основе ЯМР были разработаны для прогнозирования режимов связывания белок-лиганд (Medek et al., 2000; Cioffi et al., 2008; Riedinger et al., 2008; Aguirre et al., 2014). McCoy и Wyss использовали данные протонного CSP, индуцированного эффектом ароматического кольцевого тока в лигандах, чтобы определить положение кольца стыковочных структур (McCoy and Wyss, 2002). Недавно Ten Brink et al. сравнили экспериментальные и смоделированные CSP для проверки конформационных изменений белков и разработали метод стыковки на основе CSP (Ten Brink et al., 2015). Merz et al. разработали оценочные функции на основе CSP для определения положений связывания для комплексов белок-лиганд (Wang et al., 2007; Yu et al., 2017). Однако большинство этих оценочных функций рассчитывают только химический сдвиг протонов в белках. Оценочные функции могут быть более эффективными, если принять во внимание химические сдвиги лиганда 1 H.
В этой работе мы применили подход AF-QM / MM для расчета химического сдвига ЯМР на комплексах связывания белок-лиганд.На основе вычислений методом DFT, химические сдвиги 1 H как для белка, так и для лиганда доступны для определения структуры и улучшения оценочных функций. В рамках подхода AF-QM / MM лиганд также разделяется на более мелкие фрагменты, и, следовательно, это значительно снижает вычислительные затраты для расчета химического сдвига 1 H ЯМР для большого лиганда. В этом исследовании белок неокарзиностатин (NCS) выбран в качестве тестового примера для метода AF-QM / MM из-за его важности в терапии рака.NCS имеет экспериментальные химические сдвиги как для апо, так и для голо форм (Myers et al., 1988; Mohanty et al., 1994; Schaus et al., 2001; Takashima et al., 2005; Wang and Merz, 2010). Кроме того, путем сравнения рассчитанных AF-QM / MM химических сдвигов с данными эксперимента была разработана оценочная функция на основе химического сдвига для ранжирования нативных и предсказанных поз связывания белок-лиганд.
Эта статья организована следующим образом: сначала был проведен тестовый тест с использованием метода AF-QM / MM для расчетов химического сдвига ЯМР комплекса белок-лиганд.Расчетные результаты сравниваются с расчетами химического сдвига ЯМР для крупноразмерной системы. Впоследствии рассчитанные AF-QM / MM химические сдвиги сравниваются с экспериментальными результатами как для апо, так и для голо NCS структур. Далее обсуждается эффективность оценочных функций, основанных на химическом сдвиге, и обычных оценочных функций, основанных на энергии, для ранжирования предсказанных поз связывания белок-лиганд. Наконец, гибридная оценочная функция, которая объединяет вычисленные химические сдвиги ЯМР и энергию связи, применяется для ранжирования экспериментальной структуры и других стыкованных поз связывания.
Вычислительные подходы
Подготовка конструкции
Рентгеновские структуры апо и голо NCS были загружены из банка данных по белкам (идентификаторы PDB: 1NOA и 1NCO соответственно). Данные экспериментального химического сдвига апо-белка и хромофора были получены из предыдущих исследований (Myers et al., 1988; Mohanty et al., 1994). Химические сдвиги эксперимента holo NCS были загружены из банка данных биологического магнитного резонанса (запись BMRB: 5969). Минимизацию структуры рентгеновских структур белков проводили с помощью программы AMBER12 (Case et al., 2012) с силовым полем ff99SB. Структуры апо и голо NCS были сольватированы в усеченном октаэдрическом периодическом ящике молекул воды TIP3P с каждой стороной, по крайней мере, 10 Å от ближайшего атома растворенного вещества (Jorgensen and Jenson, 1998). После того, как вся система была нейтрализована противоионами, были использованы 1000 шагов алгоритма наискорейшего спуска с последующими 4000 шагов метода сопряженных градиентов для удаления неправильных контактов системы. Для получения параметров силового поля лиганда использовали общее силовое поле AMBER (GAFF) (Wang et al., 2004) и модель заряда AM1-связи (AM1-BCC) была использована для лиганда (Jakalian et al., 2002). Молекулярный докинг был выполнен с использованием модуля Glide в программе Schrödinger (Friesner et al., 2004; Halgren et al., 2004). В данном исследовании использовалась функция оценки Glide XP. В этом исследовании структура белка была зафиксирована, когда лиганд был прикреплен к сайту связывания. Поэтому мы не включали гибкость белка во время молекулярного стыковки. Основываясь на оптимизированной экспериментальной структуре с использованием поля молекулярных сил, Glide предсказал 38 стыковочных поз лиганда, среднеквадратичные отклонения которых варьируются от 1.От 5 до 10,5 Å по отношению к исходному положению были выбраны для последующих расчетов химического сдвига на уровне QM.
Метод AF-QM / MM для расчета химического сдвига ЯМР комплекса белок-лиганд
В подходе AF-QM / MM (He et al., 2009, 2014; Zhu et al., 2012, 2013; Swails et al., 2015; Jin et al., 2016) белок апо делится на отдельные остаток, разрезая пептидные связи. Количество фрагментов такое же, как количество остатков в белке.Каждый фрагмент содержит сердцевину (каждую аминокислоту) в белке и буферную область, которая содержит близлежащие остатки, окружающие сердцевину области. И сердцевина, и буферные области обрабатываются квантовой механикой (QM), в то время как остатки за пределами буферной области описываются молекулярной механикой (MM). Детали для определения буферных областей описаны в нашей предыдущей работе (He et al., 2009; Zhu et al., 2012, 2013). Для исследуемого в данной работе голопротеина мы разработали схему фрагментации лиганда и окружающих его белковых остатков.Как показано на рисунке 1, мы также разделили хромофор на три части, разрезав одинарную связь C-O. Фрагмент 1 содержит нафтоатную группу, фрагмент 2 включает ендииновое кольцо, а фрагмент 3 содержит аминосахарную группу, соответственно. Для каждого фрагмента лиганда (взятого в качестве каждой области ядра) остальная часть лиганда и остатки белка, окружающие область ядра (каждый фрагмент лиганда), принимаются в качестве буферной области (см. Рисунок 1). Критерии расстояния для выбора буферной области для каждого фрагмента лиганда такие же, как и для каждого остатка в белке.
Рисунок 1 . Схема фрагментации лиганда в AF-QM / MM. (A) Лиганд разделен на три фрагмента. Каждый фрагмент лиганда принимают за сердцевину. Буферная область содержит оставшуюся часть остатков лиганда и белка в пределах определенного порогового расстояния от центральной области (см. Текст для более подробной информации). Ядро и буферные области рассчитываются на уровне QM, в то время как остальная часть системы описывается вложенными зарядами. (B) Определение каждой области ядра хромофора. Фрагмент 1: нафтоатная группа; фрагмент 2: часть эндиинового кольца; фрагмент 3: аминосахарная группа. Буферная область имеет тот же цвет, что и основная область.
В этом исследовании мы принимаем следующие зависящие от расстояния критерии для включения остатков в буферной области каждой области ядра для лиганда: (1) если тяжелый атом остатка находится на расстоянии менее 3,5 Å от любого атома в области ядра , (2) если расстояние до атома водорода остатка меньше 3.0 Å от любого атома в области ядра. Отсечка обеспечивает достаточный размер буферной области для сходимости расчетов химического сдвига в области ядра (Flaig et al., 2012). Остальные остатки за пределами буферной области описываются включением зарядов для учета электростатического поля за пределами области QM. Заряды белка были получены непосредственно из силового поля ff99SB. Для назначения буферной области для каждого остатка (где каждый остаток определяется как центральная область) лиганд обрабатывают как целую молекулу (нефрагментированную), и определение буферной области следует тем же критериям, что и апо-белок.Оборванные связи закрыты атомами водорода для построения фрагмента с замкнутой оболочкой.
Расчеты QM фрагментов проводились параллельно на уровне B3LYP / 6-31G **. Все расчеты QM проводились с использованием пакета Gaussian 09 (Frisch et al., 2009). Только изотропные экранирования ЯМР атомов центральной области были собраны из расчета QM каждого фрагмента. Химические сдвиги 1 H получены путем обращения к таковому для тетраметилсилана (TMS) на том же вычислительном уровне, который равен 31.66 частей на миллион. Неявная сольватационная модель была применена для аппроксимации эффекта растворителя. Распределение заряда белка поляризует диэлектрический раствор и создает поле реакции, чтобы воздействовать на растворенное вещество до тех пор, пока не будет достигнуто равновесие. Поле реакции, действующее на растворенное вещество, может быть эффективно представлено индуцированными зарядами на поверхности полости. В этой работе поверхностные заряды рассчитываются по модели Пуассона-Больцмана (PB) с использованием программы Delphi (Rocchia et al., 2001). Набор точечных зарядов в среде ММ и на поверхности молекулы, представляющий поле реакции, используется в качестве фоновых зарядов при расчете КМ.Поскольку вычислительные затраты на расчеты химического сдвига QM будут резко увеличены для нескольких конфигураций, если принять во внимание эффект конформационной выборки. В этом исследовании оптимизированная рентгеновская структура с использованием молекулярного силового поля была взята в качестве типичной конфигурации для структуры усреднения ансамбля.
Функции подсчета очков
Чтобы отличить структуру связывания нативный белок-лиганд от поз-приманок, здесь мы предлагаем функцию оценки, основанную на химических сдвигах ЯМР (оценка CS ), которая представляет собой просто среднеквадратичное отклонение (RMSD) вычисленных химических сдвигов с ссылка на экспериментальные значения,
CSscore = ∑i = 1N (δHi-δexpi) N (1)
, где δHi — химический сдвиг –-го атома водорода на лиганде и близлежащих остатках, а δexpi — экспериментальный химический сдвиг соответствующего атома в нативном комплексе (holo NCS). N — число атомов, химические сдвиги которых были выбраны в качестве молекулярного зонда для характеристики структуры связывания NCS-хромофора. В этом исследовании N был установлен на 31 для голо-NCS, 21 из которых являются неамидными протонами на хромофоре, а остальные 10 атомов водорода — с экспериментальными нарушениями химического сдвига (CSP, между связанными и несвязанными комплексами). более 0,5 частей на миллион от остатков в сайте связывания белка (см. рисунок 2 и таблицу S1 дополнительных материалов).
Рисунок 2 . Трехмерная структура переплетного кармана. Неполярные атомы водорода на лиганде и протеиновые протоны, значения CSP которых (между связанными и несвязанными комплексами) больше 0,5 ppm (обозначены желтым цветом), выбираются в оценочных функциях на основе химического сдвига (а именно, CS оценка и CS Gscore ).
Стоит отметить, что в уравнение (1) мы также могли бы добавить химические сдвиги атомов водорода, которые экспериментально не изменяются при связывании лиганда.Ложная поза стыковки может вызвать значительные отклонения в CSP для этих атомов водорода. Однако на остатках в связывающем кармане имеется много таких протонов, которые будут усреднять итоговую оценку, чтобы функция оценки не могла отличить нативную структуру от наборов-ловушек. Протоны с экспериментальными возмущениями более 0,5 ppm более чувствительны к позе связывания, поэтому мы учли эти атомы в функции подсчета очков. Кроме того, хотя химические сдвиги ЯМР протонов амида также очень чувствительны к локальному химическому окружению связывающего кармана, эти атомы были исключены из-за отсутствия экспериментальных данных.
Вторая функция оценки ( CS Gscore ), которую мы предлагаем здесь, представляет собой линейную комбинацию оценки CS и оценки Glide,
CSGscore = CSscore + Оценка αGlide (2)
, где α — весовой коэффициент. В этом исследовании диапазоны CSscore и Glide Score составляют 0,42 ~ 2,90 и -10,96 ~ 10,98 (см. Таблицу S2 дополнительных материалов), соответственно, и поэтому мы выбираем α = (2,90–0,42) / (10,98 — (- 10,96). )) ≈ 0,1. Добавляя оценку Glide к оценке CS , можно избежать нефизической структуры с неблагоприятной энергией связи.Поскольку и оценка CS , и оценка Glide будут меньше по мере приближения стыковочной позы к экспериментальной структуре, исходная стыковочная поза даст самое низкое значение CS Gscore .
Результаты и обсуждение
Сравнительный тест AF-QM / MM на комплексе связывания нативного NCS-хромофора
Сначала мы сравнили рассчитанные химические сдвиги на хромофоре между AF-QM / MM и расчетами систем большого размера. Поскольку голо-NCS содержит более 1500 атомов и был слишком большим для выполнения полных расчетов QM системы, мы альтернативно использовали весь лиганд и его буферную область для расчета QM большого размера.Другие атомы за пределами буферной области считаются фоновыми зарядами, а поверхностные заряды ПБ для всего комплекса также помещаются так, чтобы приблизиться к неявному растворителю. Рассчитанные химические сдвиги для такой модельной системы (около 460 атомов в области QM) приняты в качестве опорных значений. Здесь мы только сравниваем химические сдвиги на лиганде между расчетом AF-QM / MM и расчетом системы большого размера. Как показано на рисунке 3, химические сдвиги 1 H на лиганде, рассчитанные методом AF-QM / MM (где лиганд разделен на три части), хорошо согласуются с расчетом системы большого размера.Средняя ошибка без знака (MUE) между результатами AF-QM / MM и химическими сдвигами из расчета крупноразмерной системы составляет 0,046 ppm, а RMSD между ними составляет 0,051 ppm. Результаты показывают, что подход AF-QM / MM может точно воспроизвести расчет крупногабаритной системы. Кроме того, на уровне DFT общая стоимость вычислений была снижена на 36%, с 5 601 мин (время ЦП) при вычислении системы большого размера до 3585 мин при разделении лиганда на 3 фрагмента. Кроме того, параллельно выполнялись 3 расчета QM / MM на основе фрагментов.Таким образом, время вычислительной стены можно дополнительно сократить примерно на 2/3.
Рисунок 3. (A) Система большого размера содержит лиганд и его буферную область (около 460 атомов) в голо-NCS. (B) Сравнение рассчитанных 1 химических сдвигов H между расчетом крупногабаритной системы и подходом AF-QM / MM. «Полная система» означает расчет крупногабаритной системы.
Затем мы сравниваем рассчитанные химические сдвиги для апо и голо NCS (31 протон в связывающем кармане, как показано на рисунке 2) с экспериментальными значениями.В этом тесте результаты AF-QM / MM хорошо коррелируют с экспериментом (см. Рисунок 4). Для связанного комплекса MUE между расчетным и экспериментальным химическими сдвигами составляет 0,44 м.д., а RMSD составляет 0,57 м.д. Для несвязанного белка и лиганда MUE и RMSD между расчетными и экспериментальными химическими сдвигами составляют 0,45 и 0,62 частей на миллион, соответственно.
Рисунок 4 . Сравнение результатов AF-QM / MM и экспериментальных 1 H химических сдвигов (31 протон в связывающем кармане, как показано на рисунке 2) как для апо, так и для голо NCS.
Прогнозирование возмущений химического сдвига (CSP) между апо и голографическим NCS могло бы дополнительно подтвердить точность подхода AF-QM / MM. Среди атомов водорода в связывающем кармане, h25, h41 лиганда, HD2 Leu45 и HB2 Cys37 значительно зависят от эффекта кольцевого тока, где они близки к ароматическим кольцам в нативной голоструктуре (см. Рисунок 5). . В результате для этих атомов наблюдались большие сдвиги в сильное поле при связывании лиганда. Экспериментальные CSP этих четырех атомов (а именно, h25, h41, Leu45: HD2 и Cys37: HB2) равны -0.93, -0,86, -1,15 и -0,72 частей на миллион, соответственно, тогда как результаты AF-QM / MM для них составляют -0,45, -0,34, -1,25 и -0,71 частей на миллион. Результаты показывают, что большие нарушения химического сдвига между системами апо и голо белок-лиганд могут быть точно предсказаны с помощью подхода AF-QM / MM.
Рисунок 5 . Рентгеновская структура голографической НКС. CSP для протонов h25 (a) и h41 (b) в хромофоре, Leu45: HD2 (c) и Cys37: HB2 (d) в NCS, значительно зависят от кольцевого тока. эффект.
Показатели по шкале CS
Функция подсчета баллов
Оценки за скольжение 38 пристыкованных ложных целей и исходной голографической NCS показаны на рисунке 6. Функция оценки на основе энергии способна отличить экспериментальную структуру от стыкованных поз, структурные RMSD которых больше 4 Å по отношению к исходной позе. . Однако для закрепленных поз, среднеквадратичное отклонение которых составляет от 2 до 4 Å, их оценка за скольжение иногда очень близка к экспериментальной структуре (см. Позу 7 на рисунке 6A).Напротив, по шкале CS легче отличить экспериментальную структуру от наборов приманок, структурные RMSD которых составляют около 2 Å. В основном это связано с тем, что химические сдвиги весьма чувствительны к местной химической среде в месте связывания. При оценке CS протоны в лиганде и выбранные атомы водорода в белковых остатках служат в качестве молекулярных зондов для обнаружения среды связывания. Когда протоны имеют разные тесные контакты между нативными и ложными структурами, такие как взаимодействия с ароматическими кольцами или водородными связями, расчетный химический сдвиг определенных протонов может иметь существенные отклонения между различными режимами связывания.Следовательно, изменение химических сдвигов ЯМР протонов может четко отражать соответствующие связывающие взаимодействия между белком и лигандом.
Рисунок 6 . Оценки Glide (A) и CSscore (B) с различными структурными RMSD хромофора в голографической NCS. Зеленая точка представляет собой оценку позы связывания на рентгеновской структуре голографической NCS. Красные, желтые и фиолетовые точки обозначают стыковочные позы 7, 10 и 18 соответственно.
Сравнение рассчитанных химических сдвигов между нативной позой и позой стыковки может исследовать структурные изменения поз связывания лиганда между ними. На рис. 7 показано, что для положений нафтоатной группы и ендиинового кольца в позе 7 хромофор очень близок к нативной связывающей структуре. Однако группа аминосахаров указывает на другое направление, что приводит к большим отклонениям химических сдвигов для h23 и h25 лиганда (см. Таблицу 1).
Рисунок 7 .Сравнение связывающей структуры позы 7 и естественной связывающей позы. Модель зеленой палочки обозначает естественную позу привязки, а модель красной палочки обозначает структуру привязки Позы 7 из программы стыковки Gilde. Группа аминосахара в хромофоре была смещена в позе стыковки 7 по сравнению с экспериментальной структурой.
Таблица 1 . Сравнение экспериментальных и расчетных химических сдвигов (в ppm).
Слабость CS с оценкой заключается в том, что для ловушек с более крупными структурными RMSD функция оценки на основе химического сдвига может быть не такой эффективной, как у тех, у кого структурные RMSD низкие.В диапазоне высоких структурных среднеквадратичных отклонений некоторые позы лигандов могут быть близки к состоянию апо (меньше взаимодействий с белком), а оценка CS для состояния апо хромофора составляет 0,60. Следовательно, ранжирование этих поз не очень чувствительно к структурным RMSD с использованием CS балла . Примеры случаев для поз 10 и 18 будут обсуждаться в разделе «Улучшение гибридной функции подсчета очков CS Gscore ».
Улучшение гибридной CS
Gscore Функция подсчета очков
На рис. 8 показано, что по шкале CS Gscore можно отличить экспериментальную структуру от ловушек для комплекса белок-лиганд.В этой работе весовой коэффициент α в уравнении (2) должен был быть установлен равным 0,1, чтобы CS набрал баллов и оценки Glide по той же шкале. Как показано на Рисунке 8, для поз, структурные RMSD которых составляют около 2–4 Å, оценка CS по химическим сдвигам ЯМР доминирует над оценочной функцией. Несмотря на то, что оценка Glide для структур-приманок близка к экспериментальной (см. Рисунок 6B), оценка CS может отличить экспериментальную структуру от ложных, в результате чего CS Gscore (комбинация оценок CS и Оценка скольжения) явно оценила позу нативного связывания как наиболее благоприятную структуру.С другой стороны, для поз-приманок, структурные RMSD которых больше 4 Å, функция оценки на основе энергии (оценка скольжения) имеет большое влияние на CS Gscore , что заставляет приманки с большими структурными RMSD больше отклоняться от экспериментальных структура по сравнению с CS оценка баллов (Рисунок 6B).
Рисунок 8 . Рейтинг аборигенов (зеленый) и стыковочных структур (красный: поза 7; желтый: поза 10; фиолетовый: поза 18; и синий: другие закрепленные позы, предсказанные Glide), рассчитанные с помощью CS Gscore .
Поза 10 ранее получала низкую оценку в CS с оценкой , среднеквадратичное значение химического сдвига которого составляет 0,70 ppm между расчетными и экспериментальными данными. Нафтоатная группа лиганда была перевернута в стыковочной структуре, в то время как положения других областей хромофора аналогичны позе нативного связывания (см. Фигуру 9). В нафтоатной группе h33 имеет наибольшее отклонение химического сдвига между нативной структурой связывания и позой 10. В позе нативного связывания h33 находится рядом с ароматическим кольцом, что вызывает химический сдвиг в слабом поле h33.Находясь в позе 10, атом h33 удаляется от индольного кольца Trp39, в результате чего его расчетный химический сдвиг намного ниже, чем у экспериментальной структуры (см. Таблицу 2). Кроме того, на рис. 9а показано, что в позе 10 h41 удаляется от фенильного кольца Phe52, и соответствующее химическое экранирование снижается, что приводит к более высокому химическому сдвигу ЯМР. Кроме того, поскольку положение ароматического кольца нафтоатной группы перемещается в стыковочной структуре, химический сдвиг 1 H на окружающих белковых остатках, таких как Cys37: HB3, также значительно изменился.Между тем, поскольку нафтоатное кольцо сильно влияло на химические сдвиги 1 H из несвязанного в связанное состояния, отклонение нафтоатной группы в Позе 10 привело к тому, что химические сдвиги меньше влияют на связывание лиганда, что приводит к немного более высокому CS . набрать баллов (см. Рисунок 6В, но другие группы близки к родному состоянию). Принимая во внимание физические несвязанные взаимодействия между NCS и хромофором, нафтоатная группа остается глубоко внутри связывающего кармана в нативном состоянии, что вносит наибольший вклад в энергию связывания NCS-хромофора.Следовательно, структурная деформация в позе 10 привела к тому, что энергия взаимодействия между ними стала слабее, и соответствующий показатель скольжения дал более низкий ранг. В гибридной функции оценки CS Gscore рейтинг позы 10 четко отделен от экспериментальной структуры, поскольку он включает как отклонения химического сдвига ЯМР от экспериментальных данных, так и энергию физического взаимодействия между белком и лигандом.
Рис. 9. (a) Связывающие структуры позы 10 (оранжевый) и естественной позы (зеленый).Переворот нафтоатной группы заставил h41 и Cys37: HB3 отойти от области ароматического кольца, что вызовет химический сдвиг в сильном поле. (b) Для позы 10 h33 перемещается от области ароматического кольца, что вызывает химический сдвиг в слабое поле.
Таблица 2 . Сравнение экспериментальных и рассчитанных химических сдвигов для модели нативного связывания и позы 10 (в ppm).
Поза стыковки 18 также является случаем, когда функция энергии более важна, чем оценка CS при ранжировании поз стыковки.На фиг.10 показано, что положение лиганда в позе 18 почти транслируется в направлении от кармана связывания. По мере удаления нафтоатного кольца химическая защита Cys37: HB2, Cys37: HB3 и Leu45: HD2 снижалась по сравнению с нативным состоянием (см. Таблицу 3). Поза лиганда 18 ближе к состоянию апо, что привело к высокому баллу CS и баллу , но энергия взаимодействия между хромофором и NCS будет явно слабее, чем у нативного состояния, а оценка Glide для позы 18 составляет существенно выше, чем нативная поза связывания (см. фиг. 6А).Следовательно, в гибридной функции оценки CS Gscore рейтинг позы 18 явно ниже, чем в исходном состоянии, что правильно отражает то, что рейтинги снижаются по мере того, как структурные RMSD становятся больше.
Рисунок 10 . Структуры позы 18 (фиолетовый) и естественного состояния (зеленый). Хромофор в позе 18 сместился от положения нативного состояния, в результате чего Cys37: HB2, Cys37: HB3 и Leu45: HD2 отошли от ароматических колец хромофора.
Таблица 3 . Сравнение экспериментальных и расчетных химических сдвигов в исходном состоянии и в позе стыковки 18 (в ppm).
Заключение
В данной работе мы применили автоматизированный метод фрагментации для расчета КМ / ММ химических сдвигов ЯМР для комплексов связывания белок-лиганд. В подходе AF-QM / MM химические сдвиги атомного ЯМР были получены путем автоматического разделения белка на фрагменты, ориентированные на остатки. Чтобы снизить вычислительные затраты для лиганда, хромофор, содержащий 81 атом, также был разделен на три меньших фрагмента, чтобы сделать размер QM для расчета лиганда сравнимым с фрагментами белка.Подход AF-QM / MM с неявной сольватационной обработкой является эффективным с точки зрения вычислений и линейным масштабированием с низким предварительным фактором. Более того, этот подход во многом параллелен и может применяться для рутинного расчета ab initio химических сдвигов ЯМР для комплексов белок-лиганд любого размера.
Химические сдвиги 1 H, рассчитанные с помощью подхода AF-QM / MM на уровне DFT, хорошо согласуются с расчетом системы большого размера, где весь лиганд и его буферная область обрабатываются QM, а оставшиеся атомы белки описываются фоновыми зарядами.MUE между AF-QM / MM и расчетом крупногабаритной системы составляет 0,046 ppm. Кроме того, MUE между расчетными и экспериментальными 1 H химическими сдвигами в связывающем кармане апо и голо NCS составляют 0,45 и 0,44 м.д., соответственно. Наши результаты демонстрируют, что подход AF-QM / MM способен воспроизводить расчеты химических сдвигов ЯМР для комплексов белок-лиганд в крупноразмерной системе ab initio , и рассчитанные химические сдвиги хорошо согласуются с экспериментальными результатами.
Результаты оценки CS баллов показывают, что химические сдвиги можно использовать в качестве молекулярных зондов для обнаружения конформации связывания комплекса белок-лиганд. Экспериментальная структура имеет явный лидирующий рейтинг по сравнению с позами связывания ложных целей. Изучая паттерны CSP структур-ловушек, изменения положения лиганда могут быть обнаружены по вариациям химических сдвигов в различных локальных химических условиях.
В этом исследовании мы также предложили гибридную функцию оценки CS Gscore , которая сочетает в себе оценку CS и функцию оценки на основе энергии для оценки скольжения.Гибридная функция подсчета очков CS Gscore может помочь отличить структуру нативного лиганда от поз стыковки-приманки. CS Gscore также может четко отделить оценки структур-ловушек, которые имеют значительно большие структурные значения RMSD и дают относительно низкие оценки CS баллов, от естественной стыковочной позы. CS Gscore включает в себя как экспериментальную информацию о химическом сдвиге ЯМР, так и метод оценки на основе энергии, который может лучше определять структуру сайта связывания комплекса белок-лиганд.Следовательно, подход AF-QM / MM обеспечивает точную и эффективную платформу для предсказания структуры связывания белок-лиганд на основе информации, полученной с помощью ЯМР.
Авторские взносы
XH разработал исследование; XJ, TZ и XH проводили исследования; XJ, TZ, JZ и XH проанализировали данные; и XJ и XH написали статью.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Эта работа была поддержана Национальной программой ключевых исследований и разработок Китая (грант № 2016YFA0501700), Национальным фондом естественных наук Китая (№№ 21673074, 21761132022 и 21433004), Молодежной программой поддержки первоклассных талантов Шанхая, Центром NYU-ECNU по вычислительной химии в Нью-Йоркском университете Шанхая и округе Путуо Шанхая (грант 2014-A-02). Мы благодарим Суперкомпьютерный центр Восточно-Китайского педагогического университета за предоставленное время для вычислений.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https: // www.frontiersin.org/articles/10.3389/fchem.2018.00150/full#supplementary-material
Список литературы
Адении, А.А., Солиман, М.Э.С. (2017). Внедрение QM в расчеты стыковки: это пустая трата вычислительного времени? Drug Discov. Сегодня 22, 1216–1223. DOI: 10.1016 / j.drudis.2017.06.012
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Aguirre, C., ten Brink, T., Cala, O., Guichou, J.-F., and Krimm, I. (2014). Структура белок-лиганд определяется возмущениями химического сдвига протонов в основной и боковой цепи. J. Biomol. ЯМР 60, 147–156. DOI: 10.1007 / s10858-014-9864-9
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Братхольм, Л. А., и Дженсен, Дж. Х. (2017). Уточнение структуры белка с использованием предсказателя химической защиты на основе квантовой механики. Chem. Sci. 8, 2061–2072. DOI: 10.1039 / C6SC04344E
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дело, Д. А. (1998). Использование химических сдвигов и их анизотропии в определении структуры биомолекул. Curr. Opin. Struct. Биол. 8, 624–630. DOI: 10.1016 / S0959-440X (98) 80155-3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Case, D., Darden, T., Cheatham, T. E. III., Simmerling, C., Wang, J., Duke, R., et al. (2012). ЯНТАРЬ 12 . Сан-Франциско, Калифорния: Калифорнийский университет.
Кавалли А., Сальвателла Х., Добсон К. М. и Вендрусколо М. (2007). Определение структуры белков по химическим сдвигам ЯМР. Proc. Nat. Акад.Sci. США 104, 9615–9620. DOI: 10.1073 / pnas.0610313104
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Christensen, A. S., Linnet, T. E., Borg, M., Boomsma, W., Lindorff-Larsen, K., Hamelryck, T., et al. (2014). Проверка и уточнение структуры белков с использованием химических сдвигов амидных протонов, полученных из квантовой механики. PLoS ONE 8: e84123. DOI: 10.1371 / journal.pone.0084123
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чоффи, М., Хантер, К. А., Пакер, М. Дж., Пандья, М. Дж., И Уильямсон, М. П. (2008). Использование количественных изменений химического сдвига 1 H ЯМР для стыковки лиганда с барназой. J. Biomol. ЯМР 43, 11–19. DOI: 10.1007 / s10858-008-9286-7
CrossRef Полный текст | Google Scholar
Цуй, К., и Карплюс, М. (2000). Молекулярные свойства из комбинированных методов QM / MM. 2. химические сдвиги в больших молекулах. J. Phys. Chem. B 104, 3721–3743. DOI: 10.1021 / jp994154g
CrossRef Полный текст | Google Scholar
Элдридж, М.Д., Мюррей, К. В., Аутон, Т. Р., Паолини, Г. В., и Ми, Р. П. (1997). Функции эмпирической оценки: I. Разработка быстрой эмпирической функции оценки для оценки сродства связывания лигандов в рецепторных комплексах. J. Comput. Помощь Мол. Des. 11, 425–445.
PubMed Аннотация | Google Scholar
Englebienne, P., and Moitessier, N. (2009). Стыковка лигандов в гибкие и сольватированные макромолекулы. 5. Основанное на силовом поле прогнозирование аффинности связывания лигандов с белками. J. Chem. Инф. Модель. 49, 2564–2571. DOI: 10.1021 / ci
1k
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Феррейра Л., душ Сантуш Р., Олива Г. и Андрикопуло А. (2015). Молекулярный докинг и стратегии дизайна лекарств на основе структуры. Молекулы 20, 13384–13421. DOI: 10,3390 / молекулы200713384
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Флэйг Д., Бир М. и Оксенфельд К. (2012). Сходимость электронной структуры с размером области QM: пример экранирования QM / MM ЯМР. J. Chem. Теор. Comput. 8, 2260–2271. DOI: 10.1021 / ct300036s
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Франк А., Мёллер Х. М. и Экснер Т. Э. (2012). К квантово-химическому расчету химических сдвигов белков ЯМР. 2. Уровень теории, основы и зависимость модели от растворителей. J. Chem. Теор. Comput. 8, 1480–1492. DOI: 10.1021 / ct200913r
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фриснер Р.А., Бэнкс, Дж. Л., Мерфи, Р. Б., Халгрен, Т. А., Кличич, Дж. Дж., Майнц, Д. Т. и др. (2004). Glide: новый подход к быстрой и точной стыковке и подсчету очков. 1. Методика и оценка точности стыковки. J. Med. Chem. 47, 1739–1749. DOI: 10.1021 / jm0306430
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Frisch, M., Trucks, G., Schlegel, H. B., Scuseria, G., Robb, M., Cheeseman, J., et al. (2009). по Гауссу 09 . Уоллингфорд, Коннектикут: Gaussian Inc.
Google Scholar
Гао, К., Йокодзима, С., Федоров, Д. Г., Китаура, К., Сакураи, М., и Накамура, С. (2010). Расчеты химического сдвига ЯМР на основе фрагментно-молекулярно-орбитального метода для больших молекулярных систем. J. Chem. Теор. Comput. 6, 1428–1444. DOI: 10.1021 / ct100006n
CrossRef Полный текст | Google Scholar
Гао, К., Йокодзима, С., Коно, Т., Исида, Т., Федоров, Д. Г., Китаура, К., и др. (2007). Расчеты химического сдвига ЯМР ab initio белков с использованием фрагментных молекулярных орбиталей с электростатическим окружением. Chem. Phys. Lett. 445, 331–339. DOI: 10.1016 / j.cplett.2007.07.103
CrossRef Полный текст | Google Scholar
Гарай П. Г., Мартин О. А., Шерага Х. А. и Вила Дж. А. (2014). Факторы, влияющие на расчет экранирования 13 C в дисахаридах. J. Comput. Chem. 35, 1854–1864. DOI: 10.1002 / jcc.23697
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гринтер, С., Цзоу, X. (2014). Проблемы, приложения и недавние достижения стыковки белок-лиганд в разработке лекарств на основе структуры. Молекулы 19, 10150–10176. DOI: 10.3390 / молекулы1
150
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Халгрен, Т. А., Мерфи, Р. Б., Фриснер, Р. А., Берд, Х. С., Фрай, Л. Л., Поллард, В. Т. и др. (2004). Glide: новый подход к быстрой и точной стыковке и подсчету очков. 2. Факторы обогащения при проверке базы данных. J. Med. Chem. 47, 1750–1759. DOI: 10.1021 / jm030644s
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хан, Б., Лю Ю., Гинзингер С. В., Вишарт Д. С. (2011). SHIFTX2: значительно улучшенное предсказание химического сдвига белка. J. Biomol. ЯМР 50, 43–57. DOI: 10.1007 / s10858-011-9478-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хартман, Дж. Д., Беран, Г. Дж. О. (2014). Метод электронной структуры на основе фрагментов для расчета химических сдвигов ядерного магнитного резонанса в молекулярных кристаллах. J. Chem. Теор. Comput. 10, 4862–4872. DOI: 10.1021 / ct500749h
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хе Х., Ван Б. и Мерц К. М. младший (2009). Расчеты химического сдвига ЯМР белков на основе метода автоматизированной фрагментации QM / MM. J. Phys. Chem. B 113, 10380–10388. DOI: 10.1021 / jp
2p
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хэ Х., Чжу Т., Ван Х., Лю Дж. И Чжан Дж. З. (2014). Фрагментный квантово-механический расчет белков и его приложения. В соотв. Chem. Res. 47, 2748–2757. DOI: 10.1021 / ar500077t
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хуанг, С.-Й., и Цзоу, X. (2006). Итеративная функция оценки, основанная на знаниях, для прогнозирования взаимодействий белок-лиганд: I. Получение потенциалов взаимодействия. J. Comput. Chem. 27, 1866–1875. DOI: 10.1002 / jcc.20504
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джакалиан А., Джек Д. Б. и Бейли К.И. (2002). Быстрая и эффективная генерация качественных атомных зарядов. Модель AM1-BCC: II. параметризация и проверка. J. Comput. Chem. 23, 1623–1641. DOI: 10.1002 / jcc.10128
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цзинь, X., Чжу, Т., Чжан, Дж. З., и Хэ, X. (2016). Систематическое исследование расчета химического сдвига РНК ЯМР на основе автоматизированного подхода QM / MM фрагментации. RSC Adv. 6, 108590–108602. DOI: 10.1039 / C6RA22518G
CrossRef Полный текст | Google Scholar
Йоргенсен, В.Л. и Дженсон К. (1998). Температурная зависимость воды TIP3P, SPC и TIP4P из моделирования NPT Монте-Карло: поиск температур максимальной плотности. J. Comput. Chem. 19, 1179–1186. DOI: 10.1002 / (SICI) 1096-987X (19980730) 19: 103.0.CO; 2-J
CrossRef Полный текст | Google Scholar
Лодевик, М. В., Зиберт, М. Р., и Тантилло, Д. Дж. (2012). Вычислительное предсказание химических сдвигов 1 H и 13 C: полезный инструмент для естественных продуктов, механистической и синтетической органической химии. Chem. Rev. 112, 1839–1862. DOI: 10.1021 / cr200106v
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Маккой, М.А., и Висс, Д.Ф. (2002). Пространственная локализация сайтов связывания лиганда с поверхностей плотности электронного тока, рассчитанная на основе возмущений химического сдвига ЯМР. J. Am. Chem. Soc. 124, 11758–11763. DOI: 10.1021 / ja026166c
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Медек, А., Хайдук, П. Дж., Мак Дж. И Фесик С. В. (2000). Использование дифференциальных химических сдвигов для определения местоположения сайта связывания и ориентации лигандов, связанных с белками. J. Am. Chem. Soc. 122, 1241–1242. DOI: 10.1021 / ja993921m
CrossRef Полный текст | Google Scholar
Мохан В., Гиббс А.С., Каммингс М.Д., Джагер Э.П. и ДеДжарле Р.Л. (2005). Стыковка: успехи и проблемы. Curr. Drug Metab. 11, 323–333. DOI: 10.2174 / 1381612053382106
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Моханти, С., Зикер, Л. К., и Дробный, Г. П. (1994). Последовательное отнесение 1 H ЯМР комплекса апонеокарзиностатина с бромидом этидия и исследование взаимодействий белок-лекарство в сайте связывания хромофора. Биохимия 33, 10579–10590. DOI: 10.1021 / bi00201a003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Моррис Г. М., Гудселл Д. С., Холлидей Р. С., Хьюи Р., Харт В. Э., Белью Р. К. и др. (1998). Автоматическая стыковка с использованием генетического алгоритма Ламарка и эмпирической функции свободной энергии связи. J. Comput. Chem. 19, 1639–1662. DOI: 10.1002 / (SICI) 1096-987X (19981115) 19: 143.0.CO; 2-B
CrossRef Полный текст | Google Scholar
Мюррей К. В., Аутон Т. Р. и Элдридж М. Д. (1998). Эмпирические оценочные функции. II. Тестирование эмпирической функции оценки для предсказания аффинности связывания лиганд-рецептор и использование байесовской регрессии для улучшения качества модели. J. Comput. Помощь Мол. Des. 12, 503–519.
PubMed Аннотация | Google Scholar
Майерс, А.Г., Протеу П. Дж. И Гендель Т. М. (1988). Стереохимическое отнесение хромофора неокарзиностатина. Структуры аддуктов хромофор-метилтиогликолят неокарзиностатина. J. Am. Chem. Soc. 110, 7212–7214.
Google Scholar
Нил С., Нип А. М., Чжан Х. и Вишарт Д. С. (2003). Быстрый и точный расчет химических сдвигов белка 1 H, 13 C и 15 N. J. Biomol. ЯМР 26, 215–240. DOI: 10.1023 / A: 1023812930288
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ридингер, К., Эндикотт, Дж. А., Кемп, С. Дж., Смит, Л. А., Уотсон, А., Валер, Э. и др. (2008). Анализ изменений химического сдвига показывает способы связывания изоиндолиноновых ингибиторов взаимодействия MDM2-p53. J. Am. Chem. Soc. 130, 16038–16044. DOI: 10.1021 / ja8062088
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Робустелли П., Кольхофф К., Кавалли А. и Вендрусколо М. (2010). Использование химических сдвигов ЯМР в качестве структурных ограничений при моделировании молекулярной динамики белков. Структура 18, 923–933. DOI: 10.1016 / j.str.2010.04.016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Роккиа В., Алексов Э. и Хониг Б. (2001). Расширение применимости нелинейного уравнения Пуассона-Больцмана: множественные диэлектрические проницаемости и многовалентные ионы. J. Phys. Chem. B 105, 6507–6514. DOI: 10.1021 / jp010454y
CrossRef Полный текст | Google Scholar
Шаус, С.Э., Кавальери, Д., и Майерс, А.Г. (2001). Анализ транскрипции генов Saccharomyces cerevisiae, подвергнутых воздействию комплекса белок неокарзиностатин-хромофор, выявляет доказательства повреждения ДНК, потенциального механизма устойчивости и последствий длительного воздействия. Proc. Nat. Акад. Sci. США 98, 11075–11080. DOI: 10.1073 / pnas.1
698
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шен Ю. и Бакс А. (2010). SPARTA +: небольшое улучшение эмпирического предсказания химического сдвига ЯМР с помощью искусственной нейронной сети. J. Biomol. ЯМР 48, 13–22. DOI: 10.1007 / s10858-010-9433-9
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Śledź, P., and Caflisch, A. (2018). Дизайн лекарств на основе структуры белков: от докинга до молекулярной динамики. Curr. Opin. Struct. Биол. 48, 93–102. DOI: 10.1016 / j.sbi.2017.10.010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Swails, J., Zhu, T., He, X., and Case, D. A. (2015). AFNMR: автоматизированный квантово-механический расчет фрагментации химических сдвигов ЯМР для биомолекул. J. Biomol. ЯМР 63, 125–139. DOI: 10.1007 / s10858-015-9970-3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Такашима, Х., Ёсида, Т., Ишино, Т., Хасуда, К., Окубо, Т., и Кобаяши, Ю. (2005). Исследование структуры ЯМР раствора для механизма высвобождения хромофора неокарзиностатина из голопротеина. J. Biol. Chem. 280, 11340–11346. DOI: 10.1074 / jbc.M411579200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тан, Х.-J., И Беттенс, Р. П. (2013). Расчеты химического сдвига Ab initio ЯМР на основе комбинированного метода фрагментации. Phys. Chem. Chem. Phys. 15, 7541–7547. DOI: 10.1039 / c3cp50406a
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тен Бринк, Т., Агирре, К., Экснер, Т. Э. и Кримм, И. (2015). Выполнение стыковки белок-лиганд с моделированием возмущений химического сдвига. J. Chem. Инф. Модель. 55, 275–283. DOI: 10.1021 / ci500446s
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Виктора, А., Мёллер, Х. М., Экснер, Т. Э. (2014). Точное ab initio предсказание химических сдвигов ЯМР нуклеиновых кислот и комплексов нуклеиновых кислот / белков. Nucleic Acids Res. 42, e173 – e173. DOI: 10.1093 / nar / gku1006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вила, Дж. А., Арнаутова, Ю. А., Мартин, О. А., и Шерага, Х. А. (2009). Квантовая механика 13 C α сервер химического сдвига (CheShift) для проверки структуры белка. Proc. Nat. Акад. Sci. США 106, 16972–16977. DOI: 10.1073 / pnas.0
3106
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ван Б. и Мерц К. М. младший (2010). Важность динамики петли в механизмах связывания и высвобождения хромофора неокарзиностатина. Phys. Chem. Chem. Phys. 12, 3443–3449. DOI: 10.1039 / b924951f
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ван Б., Вестерхофф Л. М. и Мерц К.М. (2007). Критическая оценка эффективности функций оценки белок-лиганд на основе возмущений химического сдвига ЯМР. J. Med. Chem. 50, 5128–5134. DOI: 10.1021 / jm070484a
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ван Дж., Вольф Р. М., Колдуэлл Дж. У., Коллман П. А. и Кейс Д. А. (2004). Разработка и тестирование общего янтарного силового поля. J. Comput. Chem. 25, 1157–1174. DOI: 10.1002 / jcc.20035
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сюй, X.-П., И Кейс, Д.А. (2001). Автоматическое предсказание химических сдвигов 15 N, 13 C α , 13 C β и 13 C ‘в белках с использованием базы данных с функциональной плотностью. J. Biomol. ЯМР 21, 321–333. DOI: 10.1023 / A: 1013324104681
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ю. З., Ли П., Мерц К. М. (2017). Использование индуцированных лигандом нарушений химического сдвига белков для определения структур белок-лиганд. Биохимия 56, 2349–2362. DOI: 10.1021 / acs.biochem.7b00170
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжу Т., Хэ Х. и Чжан Дж. З. (2012). Расчет химических сдвигов ЯМР для белков с неявной сольватацией с помощью функциональной теории плотности фрагментов. Phys. Chem. Chem. Phys. 14, 7837–7845. DOI: 10.1039 / C2CP23746F
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Zhu, T., Zhang, J. Z. H., and He, X.(2013). Автоматический расчет химических сдвигов амидных протонов в белках с помощью QM / MM с использованием явной модели растворителя. J. Chem. Теор. Comput. 9, 2104–2114. DOI: 10.1021 / ct300999w
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжу, Т., Чжан, Дж. З. Х., Хе, X. (2014). Коррекция ошибочно упакованных боковых цепей белка в структуре ЯМР на основе ab initio расчетов химического сдвига. Phys. Chem. Chem. Phys. 16, 18163–18169.DOI: 10.1039 / C4CP02553A
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжу, Т., Чжан, Дж. З. Х., Хэ, X (2015). «Квантовый расчет химических сдвигов белкового ЯМР на основе автоматизированного метода фрагментации», в Advance in Structural Bioinformatics , ред. Д. Вей, К. Сюй, Т. Чжао и Х. Дай (Дордрехт: Springer), 827, 49– 70.
Google Scholar
Молекулярные основы химической денатурации белков мочевиной
Реферат
Моделирование молекулярной динамики протеина химотрипсин-ингибитор 2 в 8 М мочевине при 60 ° C было предпринято для исследования молекулярных основ химической денатурации.Белок быстро разворачивался в этих условиях, но при контрольной моделировании в воде при той же температуре он сохранил свою естественную структуру. Общий процесс разворачивания в мочевине был аналогичен наблюдаемому при моделировании термической денатурации выше T m белка при 75 ° C. Первым шагом в развертывании было расширение гидрофобного ядра. Затем керн сольватировали водой, а затем мочевиной. Денатурированные структуры как в мочевине, так и при высокой температуре содержали остаточную нативную спиральную структуру, тогда как β-структура была полностью разрушена.Среднее время пребывания мочевины вокруг гидрофильных групп было в шесть раз больше, чем вокруг гидрофобных остатков, и во всех случаях больше, чем соответствующее время пребывания в воде. Самодиффузия воды снижалась на 40% в 8 М мочевине. Мочевина изменила структуру и динамику воды, тем самым уменьшив гидрофобный эффект и способствуя сольватации гидрофобных групп. Кроме того, из-за ослабления структуры воды мочевиной вода стала свободно конкурировать с внутрибелковыми взаимодействиями. Мочевина также напрямую взаимодействует с полярными остатками и пептидным остовом, тем самым стабилизируя неродные конформации.Это моделирование предполагает, что мочевина денатурирует белки с помощью как прямых, так и косвенных механизмов.
Небольшие органические молекулы в водном растворе могут оказывать сильное влияние на стабильность, структуру и функцию белка. Использование этих растворов для стабилизации или дестабилизации белков в зависимости от сорастворителя является обычным явлением. На самом деле исследования белков проводятся почти исключительно в комплексных растворах. Химическая денатурация с помощью такого агента, как мочевина, является одним из основных способов оценки стабильности белка, влияния мутаций на стабильность и развертывания белка (1).Несмотря на широкое распространение, молекулярная основа способности мочевины денатурировать белки остается неизвестной. Мочевина может оказывать свое действие напрямую, связываясь с белком, или косвенно, изменяя среду растворителя (2–20). Большинство версий модели прямого взаимодействия постулируют, что мочевина связывается и стабилизирует денатурированное состояние (D), тем самым способствуя разворачиванию. Но эта интерпретация не объясняет, как белок преодолевает кинетический барьер для разворачивания. В связи с этим мочевина может связываться с белком и конкурировать с нативными взаимодействиями, тем самым активно участвуя в процессе разворачивания.В качестве альтернативы было предложено, что мочевина действует косвенно, изменяя среду растворителя, тем самым смягчая гидрофобный эффект и облегчая обнажение остатков в гидрофобном ядре. Также возможно, что механизм разворачивания, вызванного мочевиной, зависит от концентрации мочевины. К сожалению, кажется маловероятным, что экспериментальные подходы предоставят молекулярные детали того, как мочевина денатурирует белки, поэтому мы используем моделирование молекулярной динамики (МД) с атомным разрешением для решения этой проблемы.
Использование реалистичных сорастворителей в МД-моделировании белков — редкость. На сегодняшний день опубликовано немного таких исследований: убиквитин в 60% метаноле (21), барназа в 8 М мочевине (22, 23), γ-химотрипсин в гексане (24) и субтилизин в диметилформамиде (25). Первое из этих исследований смогло продемонстрировать конформационное поведение, зависящее от растворителя, приводящее к частично развернутому состоянию убиквитина, согласующемуся с исследованиями ЯМР в тех же условиях растворителя. Исследования барназы в мочевине были направлены на изучение основ химической денатурации, как мы делаем здесь, но, к сожалению, их моделирование было слишком коротким (0.9–2 нс), даже при использовании повышенной температуры (87 ° C), чтобы денатурировать белок. Последние два исследования были разными и вместо этого содержали небольшое количество воды и были почти чистыми органическими растворителями, потому что авторы изучали функцию белков в органических средах. Таким образом, все еще существует потребность в дальнейшем исследовании влияния мочевины на белки, и теперь можно проводить гораздо более длительные симуляции, чтобы просмотреть всю координату реакции разворачивания.
При подготовке к моделированию, описанному здесь, мы рассмотрели влияние мочевины на структуру и динамику воды, особенно в отношении алканов (26) и циклических дипептидов (27).Мочевина мало влияет на структуру воды по отношению к функциям радиального распределения (26). Следовательно, мы использовали количество водородных связей на молекулу воды в качестве метрики, чтобы различать решения и силу водородной связи (28), потому что они могут выявить тонкие различия, не наблюдаемые в других усредненных по ансамблю свойствах. Число водородных связей на воду ниже в гидратной оболочке неполярных молекул, таких как октан, и эти воды ограничены из-за их усилий по максимальному взаимодействию с соседними водами при минимальном взаимодействии с углеводородами (26).
Мочевина приводит к аналогичному снижению взаимодействий вода-вода и прочности водородных связей, а также к локальному упорядочению воды вокруг полярных атомов мочевины, тем самым снижая штраф за воздействие растворителя на неполярные группы по сравнению с чистой водой (27). Интересно, что количество водородных связей на воду не может сходиться к постоянному значению, пока не будет достигнут радиус 6 Å от углеводорода или мочевины. Таким образом, проецирование мочевины на окружающую водную среду приводит к радиусу воздействия 6 Å “.«Сфера влияния примечательна: при такой низкой концентрации, как 4 M, 76% воды находится в контакте, по крайней мере, с одной молекулой мочевины. Таким образом, мочевина, по-видимому, лучше растворяет гидрофобные растворенные вещества за счет возмущения воды и, таким образом, предварительной загрузки растворителя для принятия неполярных групп за счет тонкого нарушения предпочтительной структуры воды и переориентации воды вокруг ее полярных атомов (27). При более высоких концентрациях между мочевиной и растворенным веществом наблюдается более прямое взаимодействие. Таким образом, наши предыдущие модельные исследования предполагают, что мочевина действует как прямо, так и косвенно на модельные соединения, но мы не знаем, применимо ли это к белкам.
Чтобы изучить механизм денатурации белков мочевиной, мы выполнили моделирование ингибитора химотрипсина 2 (CI2) в 8 M мочевине. CI2 был выбран для этого исследования из-за обширного количества доступной информации о его поведении при сворачивании / разворачивании как в теоретических, так и в экспериментальных исследованиях (29). Предыдущие исследования моделирования были сосредоточены на развертывании пути термической денатурации (30–35). Здесь мы описываем моделирование CI2 в 8 M растворе мочевины и в чистой воде (контроль) при 60 ° C [что ниже T m белка (36)], уделяя особое внимание процессу развертывания.Результаты сравниваются с его поведением при разворачивании при высокой температуре (125 ° C) в воде (35).
Методы
МД моделирования были выполнены с использованием encad (37). Были описаны протоколы и потенциальные функции белков (38), воды (39) и мочевины (27). Моделирование началось с кристаллической структуры CI2 [1ypc (40)]. Было описано моделирование 125 ° C в воде (35).
Контрольная траектория CI2 в чистой воде была подготовлена для MD путем сольватации белка с молекулами воды, простирающимися по крайней мере на 8 Å от любого атома белка, с получением 2596 воды.Объем коробки был отрегулирован для воспроизведения экспериментальной плотности 0,983 г / мл при 60 ° C (41). Было выполнено несколько шагов для дальнейшей подготовки системы к MD. Во-первых, только вода была минимизирована для 1001 шага, затем последовали 1001 шаг MD и еще 1001 шаг минимизации. Затем белок подвергали 1001 стадии минимизации. Наконец, вся система была минимизирована на 1001 шаг. Затем было проведено производственное моделирование. Использовалась отсечка без скрепления 8 Å с плавным усечением с силовым смещением, и список без скрепления обновлялся каждые два-пять шагов.Для уменьшения краевых эффектов в микроканоническом ансамбле использовались периодические граничные условия и соглашение о минимальном изображении. Затем моделирование проводилось в течение 20 нс с использованием временного шага 2 фс. Структуры сохранялись каждые 0,2 пс для анализа, в результате получалось 100 000 структур для каждой симуляции.
систем мочевины (8 M; мольная доля 0,186) были сконструированы путем случайной замены молекул воды мочевиной, в результате чего получили 2 103 молекулы воды и 493 молекулы мочевины. Объем коробки был отрегулирован для получения экспериментальной плотности для 60 ° C, 1.103 г / мл (42). Были выполнены три независимых моделирования 8 M мочевины: UR1, UR2 и UR3. Эти системы были подготовлены для MD путем изменения подготовительного протокола, описанного выше для CI2 в чистой воде, для создания независимых траекторий. Внесены следующие изменения: в UR1 выполнено 2001 (вместо 1001) начальные шаги минимизации системы растворителей; в UR2 был выполнен 2001 шаг (вместо 1001) динамики на растворителе; и при моделировании UR3 использовался базовый протокол, описанный выше.Производственное моделирование MD было выполнено, как описано выше.
Результаты
Свойства белка в различных средах с растворителями.
Во время моделирования отслеживали несколько общих свойств CI2 и растворителя. Сравнение среднеквадратичного отклонения Cα (rmsd) для всех траекторий показывает поразительный контраст между CI2 в воде и 8 M мочевине при 60 ° C (рис. 1 a ). Белок начал разворачиваться в мочевине в течение первых нескольких наносекунд, напоминая то, что происходило во время термической денатурации при 125 ° C (рис.1 a ).
Рисунок 1
Общие свойства CI2 в зависимости от условий моделирования. ( a ) Cα среднеквадратичное значение по кристаллической структуре. ( b ) Изменение во времени доступной площади поверхности неполярной боковой цепи для растворителя, рассчитанной с использованием модифицированной версии NACCESS (43).
Неполярная доступная для растворителя площадь поверхности при моделировании мочевины была больше, чем в воде (рис. 1 b ). Экспозиция ключевых остатков (Ile-20, Leu-49 и Ile-57) в гидрофобном ядре CI2 показана на рис.2. В каждом случае гидрофобные остатки ядра сначала сольватировали водой, а затем мочевиной (рис. 2). Доступные для растворителя площади поверхности этих ключевых остатков существенно увеличились в 8 M мочевине, в то время как они оставались погребенными при моделировании воды при 60 ° C (рис. 2). Несколько групп остатков испытали значительное увеличение сольватации в течение первых 2–3 нс в мочевине (рис. 3). Две из этих групп определяют верхний край гидрофобного ядра (остатки 9–11 и 55–58) и образуют ключевые водородные связи в нативном состоянии (N).На рис. 3 показаны снимки нескольких из этих остатков ядра, сольватированных между 0,2 и 8 нс при моделировании UR3. Боковые цепи этих гидрофобных краевых остатков стабильно сольватировались водой, а затем мочевиной до тех пор, пока неполярные внутрибелковые взаимодействия не были полностью экранированы растворителем. Водородная связь основной цепи в этой области между остатками 11 и 57 представляет собой пример прямой конкуренции за стабилизацию нативных взаимодействий растворителем. Эта водородная связь была разорвана в течение ≈1 нс, и вода и, в конечном итоге, молекулы мочевины стали новыми партнерами по водородной связи этих остатков (рис.3).
Рисунок 2
Взаимодействия растворитель-белок в гидрофобном ядре CI2. ( a ) Ленточное представление кристаллической структуры, построенное с использованием MOLSCRIPT (44) и RASTER3D (45). ( b ) Контакты воды (красный) и мочевины (пурпурный) для Ile-20 нанесены на график как функция времени для CI2 в 8 M мочевине при 60 ° C (UR1). Контакты считали, когда тяжелые атомы находились в пределах 4,6 Å или 5,4 Å для алифатических атомов углерода. ( c ) Контакты воды (темно-синий) и мочевины (голубой) для Leu-49.( d ) Контакты воды (темно-зеленый) и мочевины (светло-зеленый) для Ile-57. ( e ) Боковая цепь (с использованием окраски в a ) доступная для растворителя площадь поверхности как функция времени для этих трех гидрофобных остатков ядра в моделировании UR1 и в воде ( f ).
Рисунок 3
Сольватация гидрофобного ядра. Снимки из моделирования UR3, показывающие воду и мочевину в пределах 5,4 Å от остатков 11, 16, 20, 49 и 57 (показаны в виде заполнения пространства).Показано, что вода (зеленая) проникает через верхний край гидрофобной сердцевины за 1 нс. К 2 нс мочевина (окрашенная атомами) попала в гидрофобное ядро. Водородная связь основной цепи остатка 57 и карбонильный атом кислорода остатка 11 также показаны вверху структур. Рисунок был сделан с использованием VMD (46) и визуализирован с помощью RASTER3D (45).
Свойства воды в различных средах.
Самодиффузия воды контролировалась для измерения возможных изменений в динамике растворителя при высокой температуре и в присутствии мочевины.Диффузия воды немного снизилась при добавлении белка с 0,46 до 0,41 Å 2 / пс при 60 ° C, но резко упала в 8 M мочевине до 0,18 и 0,17 Å 2 / пс с и без CI2, соответственно. Для сравнения: экспериментальная константа диффузии воды при этой температуре составляет 0,47 Å 2 / пс (47), что хорошо согласуется со значением из MD (0,46). Время пребывания молекул растворителя вокруг поверхности белка также отслеживали для полярных и неполярных групп. Время пребывания мочевины было одинаково больше, чем у воды.Например, для UR1 среднее время пребывания мочевины составляло 25 пс вокруг полярных остатков и 3,6 пс вокруг неполярных групп, а дополнительные значения для воды составляли 2,8 и 2,6 пс соответственно.
Влияние мочевины на воду проиллюстрировано на рис. 4. Хотя общее количество водородных связей растворителя было одинаковым для CI2 в воде и 8M мочевины, 1818 и 1956, соответственно, произошел сдвиг в сторону более длинных водородных связей в мочевине. Следовательно, произошло снижение количества сильных (≤1.8 Å) водородных связей вода – вода на 136. Эти сдвиги привели к ослаблению структуры воды.
Рисунок 4
Распределение водородных связей для CI2 в воде и в 8 М мочевине. Вероятность разной длины водородной связи в течение 200 пс временного интервала от 0,8 до 1,0 нс перед разворачиванием отображается для взаимодействий вода-вода и вода-мочевина. Значения даны для объемного растворителя (> 3,5 Å для полярных атомов и> 4,5 Å для неполярных атомов) и нормированы на среднее количество молекул в объеме.Критерии водородной связи приведены в легенде к Таблице 1.
Контакты мочевины и воды также отслеживались на предмет основных гидрофобных остатков ядра на протяжении всего моделирования. Как показано на фиг. 2, когда мочевина контактировала с гидрофобным остатком, также присутствовали контакты с водой. Контакты воды и мочевины увеличиваются примерно на 2 нс (рис. 2), что совпадает с переходом через переходное состояние (TS). Как правило, вода опережала мочевину в сольватировании гидрофобного ядра. Количество водородных связей вода – белок увеличивалось на ≈50% на ранних стадиях разворачивания от N к TS (Таблица 1).Напротив, изменение количества водородных связей мочевина-белок было более вариабельным за тот же период времени. В одном случае (UR1) количество водородных связей мочевина-белок мало изменилось от N к TS и, вместо этого, увеличилось позже при разворачивании после распада вторичной структуры. В двух других моделях было большее количество водородных связей мочевина-белок в TS, особенно в UR3. В целом 30–35% групп основной цепи взаимодействовали с мочевиной> 40% времени в ансамблях TS.
Таблица 1
Внутри- и межмолекулярные водородные связи в зависимости от конформационного состояния и среды растворителя
В среднем количество водородных связей вода – белок мало увеличивалось при переходе от TS к D (табл. 1). Этот результат, по крайней мере частично, связан с относительным обогащением мочевиной на поверхности белка; отношение воды к мочевине в первой сольватной оболочке было меньше, чем в массе (3,6 против 4,2, исходя из расстояния контакта тяжелого атома 3 Å).Всего CI2 приобрел 50 водородных связей с растворителем в процессе N → D и потерял ≈25 внутрибелковых водородных связей.
Конформационные свойства, индуцированные растворителем.
Рис. 5 содержит снимки CI2 в различные моменты времени, чтобы проиллюстрировать влияние мочевины на общую укладку белка. Структуры из моделирования разворачивания обладают некоторыми основными характеристиками, такими как полностью сольватированное гидрофобное ядро и степень разрушения α-спирали и β-листа.Гидрофобные контакты между спиралью и β-листом были потеряны, что позволило остаткам, определяющим ядро, Ala-16, Ile-20, Leu-49 и Ile-57 стать сольватированными (рис. 2 и 3), а также Trp 5. (данные не показаны), спектроскопический зонд использовался экспериментально для наблюдения за разворачиванием. Спиральная структура не была полностью потеряна в 8 М мочевине, хотя происходило раскручивание и повторная укладка спирали (рис. 5). Β-структура, напротив, была отменена. В целом конформационные свойства CI2 в 8 M мочевине резко отличались от таковых при контрольном моделировании CI2 в воде при той же температуре, и, вместо этого, они были аналогичны тем, что наблюдались на траектории денатурации 125 ° C (рис.5).
Рисунок 5
Структурные изменения CI2 как функция времени и окружающей среды. Позиции нативной вторичной структуры окрашены в красный цвет (спираль) и голубой цвет (β-тяжи).
Обсуждение
Многие исследования пытались объяснить действие денатурантов, таких как мочевина (2–20, 48). Эти исследования предполагают три возможных механизма: ( i ) прямые взаимодействия мочевины с белком, особенно с D; ( ii ) косвенные эффекты через возмущение среды растворителя, способствующие сольватации гидрофобных остатков; и ( iii ) сочетание прямых и косвенных эффектов.МД моделирования с соответствующими моделями растворителей могут пролить свет на атомные детали химической денатурации мочевиной. Две группы, в частности, использовали такой подход для изучения взаимодействия мочевины с ферментом барназой (22, 23). Оба этих исследования были сосредоточены на описании прямых взаимодействий белок-мочевина с N белка, потому что моделирование было слишком коротким, чтобы наблюдать разворачивание. Кроме того, хотя их результаты в целом совпадают, они не рассматривают возможные косвенные эффекты мочевины посредством анализа растворителя.Здесь мы продолжили эти более ранние исследования, доведя разворачивание до конца и проанализировав не только прямые взаимодействия мочевина-белок, но и свойства растворителя, чтобы получить атомарное описание химической денатурации.
Основываясь на результатах нашего моделирования CI2 в 8 M мочевине, денатурация, катализируемая мочевиной, произошла за счет сочетания прямых и косвенных механизмов. Во-первых, мочевина нарушила структуру и динамику воды, что привело к ослаблению взаимодействия вода-вода и уменьшению диффузии воды.Константы диффузии воды в 8 M мочевине при 60 ° C были аналогичны константам для чистой воды при 25 ° C, что было совершенно неожиданным. Многие из ранних объяснений денатурации, зависимой от мочевины, основывались на хаотропных аргументах: мочевина нарушает структуру воды, так что гидрофобные молекулы легче сольватируются. Наши предыдущие исследования концентраций мочевины ≤4 M согласуются с этой идеей (27).
При высоких концентрациях, таких как описанные здесь, это воздействие на воду усиливается и способствует «хаотропным» свойствам мочевины и способствует более легкой сольватации неполярных остатков.Кроме того, гидрофобный эффект увеличивается с повышением температуры (49, 50), отражая увеличивающееся неравенство между возмущенными водами, вынужденными выстраиваться вокруг неполярных групп (48), и водами, свободными для взаимодействия с объемным растворителем. В 8 M мочевине гидрофобный эффект смягчается снижением динамики воды при 60 ° C. Фактически, среда растворителя лучше способна сольватировать гидрофобные группы, и исключение неполярных боковых цепей из растворителя дает небольшое преимущество; таким образом, косвенные эффекты мочевины действуют для стабилизации не-N белка, включая TS, тем самым ускоряя разворачивание белка.
Вторая часть нашей модели включает прямые взаимодействия между растворителем и CI2, которые четко видны при моделировании. Нарушение структуры воды мочевиной уменьшило когезию воды, сделав ее основным денатурантом на ранних этапах развертывания, тем самым обеспечивая связь между прямым и косвенным механизмами. Количество водородных связей вода-белок увеличилось на ≈50% от N к TS, при меньшем изменении взаимодействия мочевины. Однако мочевина также напрямую взаимодействовала с белком, особенно после разрушения вторичной структуры.Число водородных связей мочевины с основной цепью пептида увеличивалось от TS до D, тогда как водородные связи воды оставались относительно постоянными, но эти числа значительно варьировались. Увеличение «связанной» мочевины при разворачивании согласуется с оценками, основанными на кривых перехода ≈10–15 дополнительных связанных молекул (2) и значениями Махатадзе и Привалова (5) ≈30 для белков, вдвое превышающих размер CI2. Степень взаимодействия основной цепи с мочевиной в TS при моделировании (30–35% остатков) также хорошо согласуется с оценками, основанными на эксперименте (36%) (9).
Растворитель не только участвует в специфических электростатических взаимодействиях с белком, но также проверяет гидрофобные взаимодействия. Например, остатки, составляющие край гидрофобного ядра, взаимодействовали с мочевиной и водой до тех пор, пока не нарушились стабилизирующие гидрофобные и водородные связи, что привело к притоку воды, а затем и мочевины к гидрофобному ядру (рис. 2 и 3). Поскольку белок подвергает воздействию растворителя большее количество атомов основной цепи, мочевина взаимодействует преимущественно с этими атомами и исключает воду в процессе, что согласуется с предыдущими исследованиями калориметрии (51, 52), растворимости (19) и сольватации (11, 53, 54). и недавняя работа Тимашева (10, 11).Относительное обогащение мочевины вокруг белка наблюдалось в предыдущих моделях (4, 22, 23). Более того, расчет коэффициента распределения локально-объемного K p (отношение молярных концентраций сорастворителя около белка к массе) из нашего моделирования дает среднее значение 1,2, что хорошо согласуется с экспериментом (1,1 –1.4; ссылки 7 и 8). Короткое время пребывания мочевины вокруг неполярных остатков (≈4 пс) по сравнению с полярными остатками (25 пс) подтверждает часть нашей модели о прямом взаимодействии.Кроме того, время пребывания мочевины было больше, чем в воде, что согласуется с исследованиями ЯМР (55). Белок содержал остаточную структуру в виде динамической нативной спиральной структуры и кластеров гидрофобных боковых цепей, что согласуется с экспериментом (34).
Выводы
Моделирование CI2 в 8 M мочевине показывает, что мочевина способствует разворачиванию как косвенными, так и прямыми механизмами. Прямые взаимодействия с мочевиной заключаются в связывании водорода с полярными фрагментами белка, особенно с пептидными группами, что приводит к скринингу внутримолекулярных водородных связей.Сольватация гидрофобного ядра происходила за счет притока молекул воды, затем мочевины. Мочевина также способствовала разворачиванию белка косвенным образом, изменяя структуру и динамику воды, что также происходит при введении неполярных групп в воду, тем самым уменьшая гидрофобный эффект и облегчая воздействие на гидрофобные остатки ядра. В целом, воздействие мочевины на воду косвенно способствовало развертыванию, способствуя гидрофобной сольватации, тогда как прямые взаимодействия обеспечивали путь.
Благодарности
Мы благодарим докторов наук. Бен Ли, Дарвин Алонсо и Кейт Лейдиг за полезные обсуждения. Эта работа была поддержана грантом GM50789 Национального института здравоохранения (для V.D.) и учебным грантом GM07750 (для BJB) Национального института здравоохранения.
Сноски
-
↵ * Кому следует направлять корреспонденцию. Электронная почта: daggett {at} u.washington.edu.
-
Этот документ был отправлен напрямую (Трек II) в офис PNAS.
Сокращения
- CI2,
- ингибитор химотрипсина 2;
- MD,
- молекулярная динамика;
- N,
- родное состояние;
- TS,
- переходное состояние;
- D,
- денатурированное состояние
- Получено 8 января 2003 г.
- Copyright © 2003, Национальная академия наук
Химический состав и антигенность протеина Миндаль (Prunus Dulcis) и Macadamia Integolia ) Семена
Сорта миндаля и ореха макадамия были проанализированы на предмет физических характеристик семян, химического состава и белковой антигенности.Также было исследовано влияние условий окружающей среды (географическое положение и год сбора урожая) на состав семян миндаля. Масса отдельных семян миндаля составляла 0,78–1,44 г, длина 18,3–27,8 мм, ширина 9,8–13,8 мм и толщина 6,9–10,2 мм. Образцы орехов макадамия имели массу семян 2,41–3,36 г, диаметр 18,7–20,1 мм и толщину 14,4–15,9 мм. Влага, липиды, белок, зола, растворимые сахара и дубильные вещества варьировались от 2,9 до 5,6%, 50,9-66,7%, 15,7-26%.7%, 2,6-3,5%, 3,0-6,5% и 0,07-0,36% в семенах миндаля и 1,7-2,6%, 60,0-66,2%, 5,6-7,1%, 1,0-1,4%, 5,3-8,7% и 0,03- 0,04% в семенах ореха макадамия соответственно. Кислые аминокислоты (глутаминовая кислота и аспарагиновая кислота) были доминирующими аминокислотами в белках миндаля и ореха макадамия, составляя 24,4-43,3% и 35,1-40,8% от общего количества аминокислот. Белки семян миндаля и ореха макадамия имеют соотношение незаменимых аминокислот к общему количеству аминокислот в диапазоне 25,4-33,8% и 22,5-26,1%, а также являются богатыми источниками аргинина в диапазоне 7.3-11,9% и 9,4-11,5% соответственно. Общее количество свободных аминокислот в семенах миндаля варьировалось от 172,3 до 723,2 мг / 100 г сухого веса, при этом свободный аспарагин составлял 14,9-44,9% от общего количества свободных аминокислот. Моноклональный сэндвич-ELISA и поликлональный ингибирующий ELISA были разработаны для обнаружения и оценки антигенности белков миндаля и орехов макадамии, соответственно. ELISA в сочетании с вестерн-анализом и дот-блоттингом использовали для исследования антигенной стабильности белков миндаля и ореха макадамия при различных обработках (термических и pH) и для количественного определения этих белков в сложных пищевых матрицах.ELISA был способен обнаруживать основной белок миндаля (AMP) и белки ореха макадамии на уровнях от 19,2 ± 1,3 нг / мл и 21,8 ± 0,9 нг / мл, а также был способен обнаруживать белки AMP и ореха макадамии при 10 ppm или ниже в большинство протестированных пищевых матриц.
.